vibrational communication during spawning behavior in the himé salmon (landlocked red...
TRANSCRIPT

J Comp Physiol A (1991) 168:417-428 dkxltnal of
Sensory, Neural,
Pt~y=kdogy
�9 Springer-Verlag 1991
Vibrational communication during spawning behavior in the him salmon (landlocked red salmon, Oncorhynchus nerka) Masahiko Satou 1, *, Atsushi Shiraishi 1, Toshiya Matsushima 1, and Naoto Okumoto 2
Zoological Institute, Faculty of Science, University of Tokyo, Tokyo 113, Japan 2 National Research Institute of Aquaculture, Nikko Branch, Nikko 2482 3, Japan
Accepted January 2, 1991
Summary. Sexual behaviors of the salmon are composed of a stimulus-reaction chain, which ensures synchronous spawning between the sexes and successful fertilization. To characterize the signals involved in such a stimulus- reaction chain, the body vibration and electromyograph- ic activity of the trunk muscles during spawning were simultaneously recorded from freely behaving male and female pairs of him~ salmon (landlocked red salmon, Oncorhynehus nerka) and were analyzed in combination with a videographic analysis of behavior sequences. The results showed that the him~ salmon have an elaborate communication system in which characteristic vibration- al signals are exchanged. These are produced by body vibration due to trunk muscle activity related to spawn- ing and are transmitted between the sexes with an ac- curate timing through the stimulus-reaction chain. They act as timing cues to synchronize gamete release and are thought to be shared among a wide variety of fishes. It was hypothesized that the lateral line sense is involved in the detection of these vibrational signals. Furthermore, based on the sequence matrix analysis as well as on information theory, intersexual behavioral sequences during spawning were analyzed statistically. The results showed that statistically significant interactions occur between the sexes and statistically significant amounts of information are transmitted through the interactions, supporting the results from recording experiments men- tioned above. Characters of the signalling system and possible origins of the vibrational signals are also dis- cussed.
Key words: Salmon - Sexual behavior - Spawning - Vibrational communication - Body vibration - Elec- tromyographic activity - Lateral line sense
Abbreviations: E M G electromyogram; L P e musculus lateralis profundus, epaxial portion; L P h m. lateralis profundus, hypaxial portion; L S m. lateralis superficialis; P D protractor dorsalis; S D standard deviation
* Present address: Department of Information Science, Graduate School of Integrated Science, Yokohama City University, 22-2 Seto, Kanazawa-ku, Yokohama 236, Japan
Offprint requests to: Dr. M. Satou
Introduction
For many animals it is important that mating occurs at a particular, optimal moment. This is especially so in species, such as most fishes, that practice external fer- tilization, where there is a considerable risk that gametes are dispersed before they can meet.
After coming back to their home streams from a long- ranging open sea migration (or migration within the lake in the case of the landlocked form), Pacific salmon mate during brief periods at the very last stage of their lives and die shortly after completing reproductive activity (Childerhose and Trim 1979). Mating occurs in fast-flow- ing shallow streams, and the gametes rapidly lose their fertilizability after having been released into the water (Nakano 1969). These physiological and environmental constraints may cause a specific communication system to be evolved that can accurately synchronize the gamete release and thus lead to successful fertilization.
In accordance with this supposition, synchronous spawning is achieved in the salmon through a stimulus- reaction chain of behaviors between the sexes (Jones and King 1949; Tautz and Groot 1975; Satou and Ueda 1982; Satou 1987). This suggests that signals as to the timing of gamete release are exchanged during spawning behavior. Since body vibration is seen during spawning, vibrational signals are likely involved (Satou and Ueda 1982; Uematsu and Yamamori 1982; Satou 1987; Satou et al. 1987). Our recent behavioral experiments have provided evidence supporting this view, in which we used a 'female' model shaken by a motorcontrolled vibrator, and showed that both vibrational and visual cues are necessary for eliciting spawning in the male (Satou et al. 1987).
However, experimental evidence showing that the vibrational signals are actually exchanged and are used as the timing signals in the natural behavioral sequence is still limited. The properties of the vibrational signals (e.g., frequency characteristics, magnitude) and type of communication system (e.g., 'cooperative' or 'manipula- tive'?; see Dawkins and Krebs 1978) are also unclear. Whether or not significant amounts of information are

418 M. Satou et al.: Vibrational communication in salmon
transmitted through the male-female interactions has not been examined quantitatively. We aimed at clarifying these points, using a combination of physiological, ex- perimental behavioral, and statistical methods.
Materials and methods
Animals. Him6 salmon (landlocked red salmon, Oncorhynchus ner- ka, known as 'kokanee' in North America) were used for the present study; they were captured during the spawning migration (Septem- ber-October, 1988) at the mouth of a river ( 'home' river) flowing into Lake Chuzenji (Nikko, Japan). They were stocked in large flow-through ponds until they reached full maturity. A total of 16 animals, weighing 390-590 g each, were selected for the experi- ments.
Aquarium. Experiments were performed using an aquarium 142 cm long, 45 cm high, and 42 cm wide. It had gravel on its bottom, and the water from the 'home' river was introduced to it using a pump. The water flowed along the longer axis at a rate of 5-6 cm/s.
Recordin9 of body vibration and electromyoqraphic (EMG ) activi- ties. The body vibration and the related electromyographic (EMG) activities of trunk muscles were simultaneously recorded from freely behaving male and female pairs. Vibration of the body was mea- sured by using a piezoelectric acceleration transducer (MT 3T, Nihon Kohden) attached to the upper part of the flank at a level 2.3-4.8 cm rostral to the dorsal fin.
Surgery for attaching the acceleration transducer and for im- planting the EMG electrodes was performed under 0.03 % tricain methanesulfonate (MS-222) anesthesia. The acceleration trans- ducer was sewn on the body skin using surgical threads. EMG activities were recorded by using bipolar copper wire electrodes (diameter: 200 lam; bared tip length: 1 mm) from the following 4 trunk muscles at the level 1.0-7.5 cm rostral to the dorsal fin [musculus lateralis profundus (epaxial portion, LPe; and hypaxial portion, LPh); m. lateralis superficialis, LS; protractor dorsalis, PD] (cf. Uematsu and Yamamori 1982; Matsushima et al. 1986, 1989). EMG activities from 2 of these 4 muscles were simultaneous- ly recorded.
After amplification through AC amplifiers (AB-632J, Nihon Kohden), the signals from the acceleration transducers and the EMGs were stored with the aid of a magnetic tape recorder (XR-50, TEAC) and analyzed later. Low- and high-cutoff frequencies of the amplifiers were selected so that the overall frequency response of the recording system was fiat between 1.6 Hz and 1 kHz for signals from the acceleration transducers, while fiat between 53.5 Hz and 1 kHz for EMGs. A multichannel Pt-Hg sliding contact (a combination of two 8-channel and one 16-channel sliding contacts) was spe- cially designed for relaying these signals, while preventing the lead wires from being intertwined as well as from being twisted.
After recovery from anesthesia, a pair of animals were in- troduced into the aquarium. They behaved quite normally, as the intact pair did, and all spawned 1.5-8 h after introduction. The spawning behavior was recorded on a videotape recorder (NV-8950, National). Time correspondence between the behavioral events and the recorded signals was accomplished by using a video timer (VTG-55, FOR.A), timing signals from which were fed into the magnetic tape recorder. To confirm the time correspondence, the output signals from the acceleration transducers and the inte- grated EMGs were superimposed on the video signals through a Videogram (VG-40, FOR.A).
Statistics for intersexual behavioral sequences. To analyze statistical- ly the behavioral sequences between the male and female, the num- ber of occurrences of behaviors in one sex that followed the preced- ing behaviors in the opposite sex was counted from videotape
recordings in combination with the body vibration and EMG re- cordings. These data were collected and presented in contingency tables as sequence matrices, where the rows and columns of the tables were preceding behaviors in one sex and following behaviors in opposite sex, respectively.
Since these contingency tables turned out to include cells, whose expected values were 5 or less and the percentages of these cells were 20% or more, they were reduced by combining categories in rows and columns to make the percentages of cells with small expected values (5 or less) become less than 20% or to make a 2 x 2 contin- gency table. Z 2 statistics or Fisher's exact probability test (Colgan 1978; Kasuya and Fujita 1984) were applied to check the indepen- dency of behavioral sequences.
Analysis based on information theory (Hazlett and Bossert 1965; Dingle 1969) was also applied to examine statistically the amount of information involved in the preceding and following behaviors as well as that transmitted during intersexual interac- tions. From the same contingency tables as described above, the amounts of information, H(S), H(R), H(S,R) and T(S,R), were calculated. H(S) determines the amount of information in the av- erage distribution of preceding behaviors. It is given by H(S) = - EPilog Pi, where Pi is the probability of occurrence of
i
the behaviour i preceded. The logarithm was taken to the base 2, thus H(S) [and also H(R), H(S,R), T(S,R)] is expressed in 'bits'. H(R) determines the amount of information in the average distribu- tion of the behaviors followed. It is given by H(R) = - Z Pj log Pj,
J where Pj is the probability of occurrence of the behavior j followed. H(S,R) determines the amount of information in the average distri- bution of the dyad, a set of preceding and following behaviors. It is given by H(S,R) = - .Z Pij log Pij, where Pij is the joint proba-
i, j
bility of the behavior i being followed by the behavior j. T(S,R) determines the amount of information transmitted during the in- teractions. It is given by T(S,R) = H(S)+ H(R) -H(S ,R) .
Hmax(S), Hmax(R), and Hmax(S,R) determine the upper limit to the information present and thus represent the value of H(S), H(R), H(S,R), respectively, if the behaviors occurred with equal frequency. They are given by Hmax(S)= logm, Hmax(R)=logn, Hmax(S,R) = logmn, where m and n are the number of categories of preceding and following behaviors, respectively, i.e., the number of rows and columns in a m x n contingency table. Whether or not H(S), H(R), and H(S,R) statistically differed from Hmax(S), Hmax(R), and Hmax(S,R) was tested by Miller's method (Colgan 1978; Kasuya and Fujita 1984). Whether or not T(S,R) statistically differed from zero was tested also by Miller's method.
Behavioral terminoloqy and spawnin9 sequence. The sexual behavior of the him6 salmon consists of courting by the male and nest building by the female, as in other salmonid fishes. The following behaviors can be distinguished from aquarium observations (Satou and Ueda 1982; Satou et al. 1984; Satou 1987; Matsushima et al. 1989): 'Probing,' in which the female checks gravel on the bottom using her fins; 'digging,' in which the female digs the nest by a series of body flexures with strong up- and downstrokes while in a hor- izontal position; 'crouching,' in which the female crouches deep in the nest. The female usually performs these behaviors while point- ing upstream. After 'probing,' 'crouching,' or 'digging,' the female swims downstream and turns upstream again ('turning'). 'Slow swimming' is a behavior in which the female swims slowly in the bottom layer without touching the gravel. During the time she is building the nest, the male stays downstream of her and waits for a chance to court or spawn while swimming slowly ('waiting'). 'Quivering' is a courtship behavior in which the waiting male, after darting towards the female from behind ('approaching'), quivers his body. The male sometimes slowly follows the female ('following'). 'Prespawning act' is a female behavior that usually occurs following 'crouching' and resembles the female 'spawning act' mentioned below except that she does not release eggs. 'Spawning act' in the male involves, after swimming up to the female from behind

M. Satou et al. : Vibrational communication in salmon 419
('spawning approach'), widely opening his mouth next to her and releasing milt. 'Spawning act' in the female occurs following the 'prespawning act' and is quite similar to the male 'spawning act' except that the female releases eggs instead of milt. Shortly after the 'spawning act' the female covers the eggs released in the nest by a series of body flexures with up- and downstrokes while in a hor- izontal position ('covering'). While 'covering,' the body flexures are less vigorous than in the case of 'digging.' Besides these behaviors mentioned above which we tentatively classify as 'sexual', both the male and female occasionally perform 'searching' within the aquar- ium, 'escape attempt' from the aquarium, and 'air snapping', which we tentatively classify as 'nonsexual' behaviors. Aggressive behav- iors, such as 'biting' and 'butting,' are also observed between the sexes.
Three successive stages (stages 1-3) can be distinguished during reproductive activity in the female; stages 1 and 2 are the early and late phases, respectively, before spawning, while stage 3 is after spawning. In stage 1 the female repeats 'digging' and/or ' turning' following 'probing.' In stage 2 the female repeats 'crouching' and/or ' turning' following 'probing' in addition to the behaviors performed in stage 1. The male repeats 'approaching' and 'quivering' towards the probing or crouching female following 'waiting' both in stages 1 and 2. At the end of stage 2 the sexual behaviors culminate in spawning within the nest by both sexes through a chain of stimuli
and reactions during interactions between the male and female. In this behavioral chain, the female 'probing' or 'crouching' is followed by 'quivering' in the male, which in turn is followed by the 'prespawning act' in the female. The female also per- forms the 'prespawning act' spontaneously with no change in the preceding male behaviors or with no male in the aquarium. The female 'prespawning act' is succeeded by the 'spawning ap- proach' and 'spawning act' in the male, which in turn are follo- wed by the 'spawning act' in the female. Thus, a synchronous spawning by both sexes is achieved. In stage 3, the female repeats 'covering' and/or ' turning' following 'probing.' The male also repeats 'approaching' and 'quivering' following 'waiting' in sta- ge 3, although the frequency of the male sexual behaviors is con- siderably reduced, and the frequency of nonsexual behaviors such as 'searching' and 'escape attempt' increases (Satou and Ueda 1982; Satou et al. 1984; Satou 1987). Figure 1 schemati- cally illustrates the behavioral sequences during the 3 successive stages.
Body vibration is seen during 'quivering' and 'spawning act' in the male, while it is seen during 'prespawning act' and 'spawning act' in the female. However, to obtain further information, such as concerning the exact temporal character and frequency content, analysis using physiological as well as videographic methods is necessary.
IS,tage 2]
r
I Stage l]
wait
wait u~
quiver~~ probe
/'1 \
t u r n
P /
spa~ a p ~ prespa~n
I S t a g e 31
J d
walt
/
search
\
Fig. 1. Schematic illustration of behavioral sequences during re- the male, non-stippled ones the female. See text for further explana- productive activity in the him6 salmon. Stippled animals indicate tion

420 M. Satou et al. : Vibrational communication in salmon
Results
Body vibration and related electromyographic (EMG) activities
A total of 10 spawning sequences from 8 males and 8 females were analyzed. The same chain of male-female behaviors as seen in the intact pairs was observed. Body vibration detected as outputs from the acceleration trans- ducer occurred during 4 behaviors, i.e., the 'prespawning act' and 'spawning act' in the female and the 'quivering' and 'spawning act' in the male, confirming the previous behavioral observations. The timing, frequency, and duration of the body vibration were all characteristic of each behavior. Characteristic EMG activity of trunk muscles accompanied the body vibration. Besides these behaviors, 'digging' and 'covering' in the female were also accompanied by a low-frequency (4-7 Hz) body vibration which corresponded to the cycle of body flexures.
The female performed the 'prespawning act' either spontaneously (Fig. 2) or following the male 'quivering' (Fig. 3). The body vibration during the spontaneously occurring 'prespawning act' varied in intensity. Figure 2 shows 2 examples of the records of body vibration and related EMG activities during the weak (Fig. 2A) and strong (Fig. 2B) 'prespawning act' performed by the same female. In Fig. 2A the female performed two weak 'prespawning acts' of short duration successively, which were 0.36 and 1.00 s in duration (time during which the female opened her mouth), respectively, and were 0.17 G in magnitude (peak-to-peak value • 0.5) in both cases. Weak discharges from muscle LS (Fig. 2Ab), but not from muscle LPe (Fig. 2Ac), accompanied each cycle of the body vibration. In Fig. 2B the female performed a strong 'prespawning act' which lasted for 4.42 s. The magnitude of body vibration increased gradually and reached the plateau (about 0.5 G) 1.6 s after the onset of the 'prespawning act' (Fig. 2Ba). Rather regular rhyth- mic discharges from muscle LS, but not from muscle LPe, also accompanied each cycle of the body vibration
(Fig. 2Bb, c). Spontaneous 'prespawning act' with body vibration of a barely detectable magnitude was also ob- served occasionally (not illustrated).
Figure 3 shows a typical example of the records during the spawning sequence. In Fig. 3C the sequence of female (upper) and male (lower) behaviors analyzed from the videotape recordings (Fig. 3A) and from the EMG pat- terns (Fig. 3B) is illustrated schematically. In this case, the female 'crouching' preceded the male 'quivering' by 1.06 s, while the male 'quivering' preceded the female 'prespawning act' by 0.27 s. The female 'prespawning act' preceded the male 'spawning approach' by 1.30 s and the male 'spawning act' by 2.39 s, while the male 'spawning act' preceded the female 'spawning act' by 1.09 s. The time when the female started her 'spawning act' was judged from the time the strong arrhythmic EMG activi- ties started (Fig. 3Bb, c, right; see also below). It should be noted that even through close inspection of the behav- ior on videotape recordings the exact starting time of the female 'spawning act' could not be determined since the
female 'spawning act' resembled the preceding female 'prespawning act' to the naked eye (cf. Fig. 3Aa, b, c). Thus, the only and most reliable way to know the exact moment when the female started her 'spawning act' was to examine the EMG patterns. The duration of each behavior was 3.48 s (female 'prespawning act'), 4.02 s (female 'spawning act'), 1.21 s (male 'quivering'), and 3.91 s (male 'spawning act') in the case in Fig. 3. The range, mean, and standard deviation (SD) of the inter- vals between the male-female behaviors as well as their durations obtained from a number of measurements are shown in Table 1.
These measurements and the facts that there was no case in which the female 'spawning act' preceded the female 'prespawning act,' the female 'spawning act' preceded the male 'spawning act,' or the male 'spawning act' preceded the female 'prespawning act' suggest that behaviors in one sex occur in a characteristic sequence, and each behavior in one sex precedes and follows behav- iors in the opposite sex according to a characteristic time relationship.
A a I
c I m
Prespawning Act
B
Prespawning Act
Fig. 2A, B. Body vibration and electromyographic activity during the weak (A) and strong (B) 'prespawning act' which occurred spontaneously. Records A and B were obtained from the same female. (Aa, Ba) Body vibration detected by the acceleration transducer. (Ab, e, Bb, e) Electromyograms from right LS (Ab, Bb) and right LPe (Ac, Be). Horizontal calibration: 1 s; vertical calibration: 0.5 G (Aa, Ba), 0.5 mV (Ab, c, Bb, c)

M. Satou et al. : Vibrational communication in salmon 421
B a b ~ c d
F e m a l e ' ~ r ' t
v - ~ , , , \%~,~.,~, ' / I r ' TYro ' l ,1 ~L:lJ~.i!.k.. I v ' | I " , I L t L [ I I , I I I ' : , ' I i
c . . . . . . . \ I Mate
"~' "~'":% t ~,~ t~ , ~ ~ , ~ , , , , i i ' h ' i ~ ' ' ~
e
f
C ...Crouching Prespawning Act
Spawning Act Quivering
Spawning Approach Spawning Act (Male)
(FemaLe)
Fig. 3A-C. Body vibration, electromyographic activity, and sequence of spawning behavior. (A) Photographs taken from video- tape recordings showing behaviors during spawning sequence. Aa--d were respectively taken at the times indicated by arrows with letters a d in Ba. The female is in front of the male. Note that the male approached towards the female from behind (Aa), and lay closely side by side with her (Ab, c). The white cloud in Ad is the released milt. Superimposed traces on the background are the outputs from the acceleration transducers and the integrated electromyograms
(EMGs). (B) Body vibration detected by acceleration transducers (Ba, d) and EMGs from trunk muscles (Bb, e, e, t). The spike-like deflections with large amplitude in trace Ba during the female 'spawning act' were artifacts caused by mechanical shocks imposed on the acceleration transducer. EMGs from right LS (Bb, e); right LPe (Be, f). Horizontal calibration: 1 s; vertical calibration: 2.5 G (Ba), 2 mV (Bb, c), 1.0 G (Bd), 1 mV (Be, f). (C) Schematic illustra- tion of spawning sequence. Same time scale as in B. Different animals from Fig. 2
Table 1. Intervals between male and female behaviors and their durations
Range Mean SD n (s) (s) (s)
Interval
'prespawning act' 0.38-2.60 1.47 0.82 10 to d' 'spawning act' 'spawning act' 0.75-3.53 1.68 0.77 9 to ~ 'spawning act'
Duration 2 'prespawning act' 1.27-6.13 3.21 1.43 9
'spawning act' 2.83-5.08 4.02 0.73 9 'spawning act' 3.92-5.98 4.53 0.65 9
d' 'quivering' 0.395.92 1.06 0.60 31
A s t rong b o d y vibrat ion was detected dur ing the female 'p respawning act ' (Fig. 3Ba, left), the male 'quivering ' (Fig. 3Bd, left), and the male ' spawning act ' (Fig. 3Bd, right; see also Fig. 5), in agreement with previous behavioral observations, while a sudden de- crease and later resumpt ion o f b o d y vibrat ion was ob- served dur ing the female ' spawning act ' (Fig. 3Ba, right), a character which has no t been noticed previously. These were c o m m o n results f rom 10 spawning sequences exam- ined, a l though the female occasional ly per formed the 'p respawning act ' (1 case ou t o f 10 'p respawning acts ' which cont inued until the female ' spawning act ' ) with body vibrat ion o f a barely detectable magni tude.
In the case in Fig. 3, the magni tude o f b o d y vibrat ion was 0.42 G during the male 'quivering, ' 0.57 G during the male ' spawning act , ' and 2.05 G during the female 'pre-

422
Table 2. Magnitude of body accelerations
Range Mean SD n (G) (G) (G)
'prespawning act' 0.00-3.22 1.10 0.94 10 5' 'spawning act' 0.57-5.60 3.85 1.00 26* 3' 'quivering' 0.42-4.29 2.67 0.97 31
* Data from male 'spawning act' which was not followed by female 'spawning act' are included
Table 3. Frequency characteristics of body vibration. Significant differences (P < 0.01) were found between peak frequencies (PFs) of female 'spawning act' and male 'spawning act', female 'spawning act' and male 'quivering', or male 'spawning act' and male 'quiver- ing' (Mann-Whitney's U-test, two-tailed)
Range Number of peaks n (Hz)
'prespawning act' 2-32 2-3 5
Range of PF Mean of PF SD n (Hz) (Hz) (Hz)
'spawning act' 5.0- 7.4 6.3 0.8 5 3 ̀'spawning act' 9.9-18.9 14.3 2.7 7* 3` 'quivering' 12.4-21.0 17.5 2.0 31
* Data from male 'spawning act' which was not followed by female 'spawning act' are excluded; in two other cases double peaks (10.9 Hz, 13.1 Hz; Fig. 4d and 11.8 Hz, 14.2 Hz) were observed
spawning act'. Table 2 summarizes the body accelera- tions during these behaviors obtained from a number of measurements. It should be noted that these values seem to be underestimated, since the acceleration transducer was attached so that its axis was rather oblique to the axis of body vibration. Since the body accelerations exceeded 103-104 times greater than the acceleration threshold of the mechanoreceptive lateral line of the him6 salmon (see Discussion), we can assume that the vibrational signals are picked up by the lateral line organ and thus are transmitted to the recipient animal. In several cases, it was observed that the body of the recipient animal was passively vibrated during the male 'quivering,' male 'spawning act,' and female 'prespawning act' (not illus- trated). This also supports the view mentioned above that the vibrational signals are actually transmitted be- tween the sexes during these behaviors.
The frequency characteristics of the body vibrations were examined by calculating their power spectra by the FFT (fast Fourier transform) method with the aid of a computer (7T 18, Sanei). Figure 4 shows typical examples of the results. Figure 4a, c, d, and e correspond respec- tively to 4 records in Fig. 3Ba and d mentioned above. Figure 4b shows another example of the power spectrum during the 'prespawning act' performed by a different female. Table 3 summarizes the frequency characteristics of body vibration obtained from a number of measure- ments. The body vibration during the female 'prespawn- ing act' extended over a broad frequency range (2-32 Hz)
M. Satou et al. : Vibrational communication in salmon
1.0
0.5 �84
o 0 0_
1.o d~ > .t--
1~ 0.5
0
1.0
0.5
0 0
a
c
10 20 50
Frequency 4O
HZ
L-
O
0.5
0
d v
1.0
0.5
e
o o ~o 20 3o 40 Hz Frequency
Fig. 4a-e. Power spectra of body acceleration. Female 'prespawning act' (a, b); female 'spawning act' (c); male 'spawning act' (d); male 'quivering' (e). Calculation for power spectra was performed during periods between arrowheads in inset records. (a), (c)-(e) same re- cords as in Fig. 3Ba, d. Horizontal calibration: 1 s; vertical calibra- tion: 1 G (a, c-e), 0.5 G (b)

M. Satou et al. : Vibrational communication in salmon 423
B a b c
Female l l,, J I
c - - . . . . . . . . . . . . . I
M a l e , ,. :, I!A/, ' , - - ~ ^ s ' / ~ b : J ! , i ; , '~ : ; � 8 4 1 8 4
' i'! i' t f1!! k' e - - ,~,1} !]l[l ltiit!tj!tit,! t r ii,
C Prespawning Act (Female)
i1!
Spawning Approach ,, Spawning Act (MaLe)
~ . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
Fig. 5A-C. Body vibration, electromyographic activity, and behav- ioral sequence in the case that the male spawned normally, but the female failed to spawn. (A) Photographs of behaviors taken from the videotape recordings. Aa--e were respectively taken at the times indicated by arrows with letters a - c in Ba. The male is in front of the female. Note that the male approached towards the female from behind (Aa) but could not lay closely side by side with her due to slight shortness of the lead wires (Ah, e). The photograph Ah was taken at the moment when the male positioned nearest to the female. Superimposed traces on the background are the outputs from the acceleration transducers and the integrated electromyo- grams (EMGs). (B) Body vibration detected by acceleration trans- ducers (Ba, d) and EMGs from trunk muscles (Bb, e, e, f). EMGs from right LS (Bb, f); right LPe (Be, e). Note that the female performed the 'prespawning act' spontaneously, during which a small LS activity with less regular rhythmicity continued (Bb), and
: , !J, (
i,,, L J, ,
the accompanying body vibration (0.25 G in magnitude) with a broad frequency range (4~25 Hz) occurred intermittently (Ba). The EMG activity of LPe largely remained silent (Be), although a very small activity (Be, asterisk) could be recognized which seemed to be a reflection of the EMG activity for the female 'spawning act' not having developed into the full-sized one. During the male 'spawning act' a large, rhythmic EMG activity in both LS and LPe continued (Be, f), accompanied by body vibration with a large amplitude (5.60 G) and a regular rhythmicity (peak frequency: 15.8 Hz) (Bd). Hor- izontal calibration : 1 s; vertical calibration: 0.5 G (Ba), 0.5 mV (Bb, e, e, t), 2.5 G (Bd). (C) Schematic illustration of behavioral sequence. Dotted line indicates the period during which the milt was released. Milt release started 0.9 s after the beginning of the male 'spawning act' and lasted for about 2 s. Same time scale as in B. Different animals from Figs. 2 and 3
with 2-3 b road peaks, a pat tern conspicuously different f rom that o f other vibrational behaviors (Fig. 4; Table 3). This range agreed well with the effective frequency range o f a vibrat ing ' female ' model for eliciting the male ' spawning act ' (unpublished results). Statistically signifi- cant differences ( P < 0.01; Mann-W hi tn ey ' s U-test, two- tailed) were found a m o n g peak frequencies o f the female ' spawning act , ' male ' spawning act , ' and male 'quivering ' (Table 3). These results suggest tha t each vibrat ional behavior has a characterist ic frequency.
A rhythmic E M G activity o f t runk muscles (LS, LPe, LPh, PD), o f which each peak cor responded to each cycle o f the b o d y vibrat ion, was consistently observed (Figs.
2, 3, 5). The E M G activity on one side al ternated with that in the opposi te side, while it was synchronized a m o n g muscles on each side, as observed previously (Matsushima et al. 1986, 1989). E M G rhythmici ty dur ing the male 'quiver ing ' (Fig. 3Be, f, left) and the male ' spawning act ' (Fig. 3Be, f, r ight; Fig. 5Be, f) was very regular, which cor responded to the na r row range spectra (Fig. 4d, e; Table 3). E M G rhythmici ty dur ing the female 'p respawning act ' (Fig. 3Bb, left; Fig. 5Bb) was less regular, which cor responded to the b road range spectra (Fig. 4a, b; Table 3). In muscle LS, such a rhythmic E M G activity was observed in all ' p respawning acts ' examined (N = 7), while in muscles LPe, LPh, or P D it

424 M. Satou et al. : Vibrational communication in salmon
was often very small or completely silent (Fig. 3Bb, c, left; Fig. 5Bb, c). E M G activity during the female 'spawning act ' (Fig. 3Bb, c, right) started as a sudden arrhythmic activity with large amplitude. This corre- sponded to the sudden decrease in the body vibration (Fig. 3Ba, right; see Discussion). In the later part of the E M G activity of LS, slow rhythmicity was observed (Fig. 3Bb, right), which corresponded to the low frequency peak in the spectrum (Fig. 4c; Table 3).
The milt release started approximately 1 s after the beginning of the rhythmic E M G activity for the male 'spawning act ' and lasted for about 2 s or more (see Fig. 5C), while the egg release started within 1 s of the begin- ning of the arrhythmic E M G activity for the female 'spawning act ' and lasted for a few s. Thus, the gametes from both sexes can meet shortly after their release.
In all 10 spawning sequences examined, the female 'spawning act ' and accompanying egg release occurred when the male approached the female f rom behind (Fig. 3Aa) and lay, while vibrating his body, closely side by side with her (Fig. 3Ab, c). However, it did not occur if the male did not move to a position close to the female, or if he did not vibrate his body at an appropriate time. One case observed is worth noting, in which the male could not lie closely side by side with the female due to slight shortness of the lead wires (Fig. 5Ab, c). In this case, the female did not perform her 'spawning act ' and accompanying egg release, in spite of the fact that she performed a normal 'prespawning act ' ('false spawning
act, ' Fig. 5Aa-c, Ba-c, C) and the male also performed a normal 'spawning act ' with a large amount of milt release (Fig. 5Ab, c, Bd-f, C) following the female 'pre- spawning act'. The female repeated such a 'false spawn- ing act ' 9 times, before she finally performed a ' true spawning act ' with egg release, which occurred shortly after the length of the lead wires were readjusted so that the male could lie closely next to the female (not illus- trated). We observed 4 other such 'false spawning acts' in 2 other females. In two of them, the male-female distance was too large, while in the other two the timing of the male body vibration was too late.
Statistical analysis for &tersexual interactions
The results f rom experiments recording the body vibra- tion and E M G activities suggested that each behavior in one sex precedes and follows behaviors in the opposite sex in a characteristic way, and information as to the timing of gamete release is transmitted between the sexes during the male-female interactions. This was confirmed by statistically analyzing the behavioral sequences.
The same 10 spawning sequences as those analyzed in the recording experiments were examined. Data from a total o f 104 min of observation (2-32 min of observation per spawning sequence) during the late phase of stage 2 were collected. The number of occurrences of male be- haviors that followed preceding female behaviors was counted and displayed at the top of each cell in Table 4,
Table 4. Sequence matrix of female-to-male behavioral sequences. Each row corresponds to the preceding female behaviors, and each column corresponds to the subsequent male behaviors. Values
at the top in each cell are the number of occurrences, those at the middle are expected values, and those at the bottom are Z 2 values
Female behaviors Male behaviors
'waiting' 'following' 'approaching' 'quivering' 'spawning 'spawning Marginal totals approach' act'
'slow swimming' 9 0 10 10 0 0 29 11.14 1.69 9.88 3.93 1.27 1.10 0.41 1.69 0.00 9.40 1.27 1.10
'turning' 156 40 7 9 0 0 212 81.47 12.34 72.21 28.70 9.26 8.02 68.19"** 61.96"** 58.89*** 13.52"** 9.26** 8.02**
'digging' 10 0 1 0 0 0 11 4.23 0.64 3.75 1.49 0.48 0.42 7.88 0.64 2.01 1.49 0.48 0.42
'probing' 47 0 118 42 0 0 207 79.55 12.05 70.51 28.02 9.04 7.83 13.32"** 12.05"** 31.99"** 6.97** 9.04** 7.83**
'crouching' 36 0 92 29 1 0 158 60.72 9.20 53.82 21.39 6.90 5.98 10.06"* 9.20** 27.09*** 2.71 5.05* 5.98*
'prespawning act' 2 0 6 3 29 26 66 25.36 3.84 22.48 8.99 2.88 2.50 21.52"** 3.84 12.08"** 3.94* 236.69 221.11
'spawning act' 4 0 0 0 0 0 4 1.54 0.23 1.36 0.54 0.18 0.15 3.95 0.23 1.36 0.54 0.18 0.15
Marginal totals 264 40 234 93 30 26 687
* Significant at P<O.05, ** significant at P<O.O1, *** significant at P<O.O01 0r 2 test)
M. Satou et al. : Vibrational communication in salmon
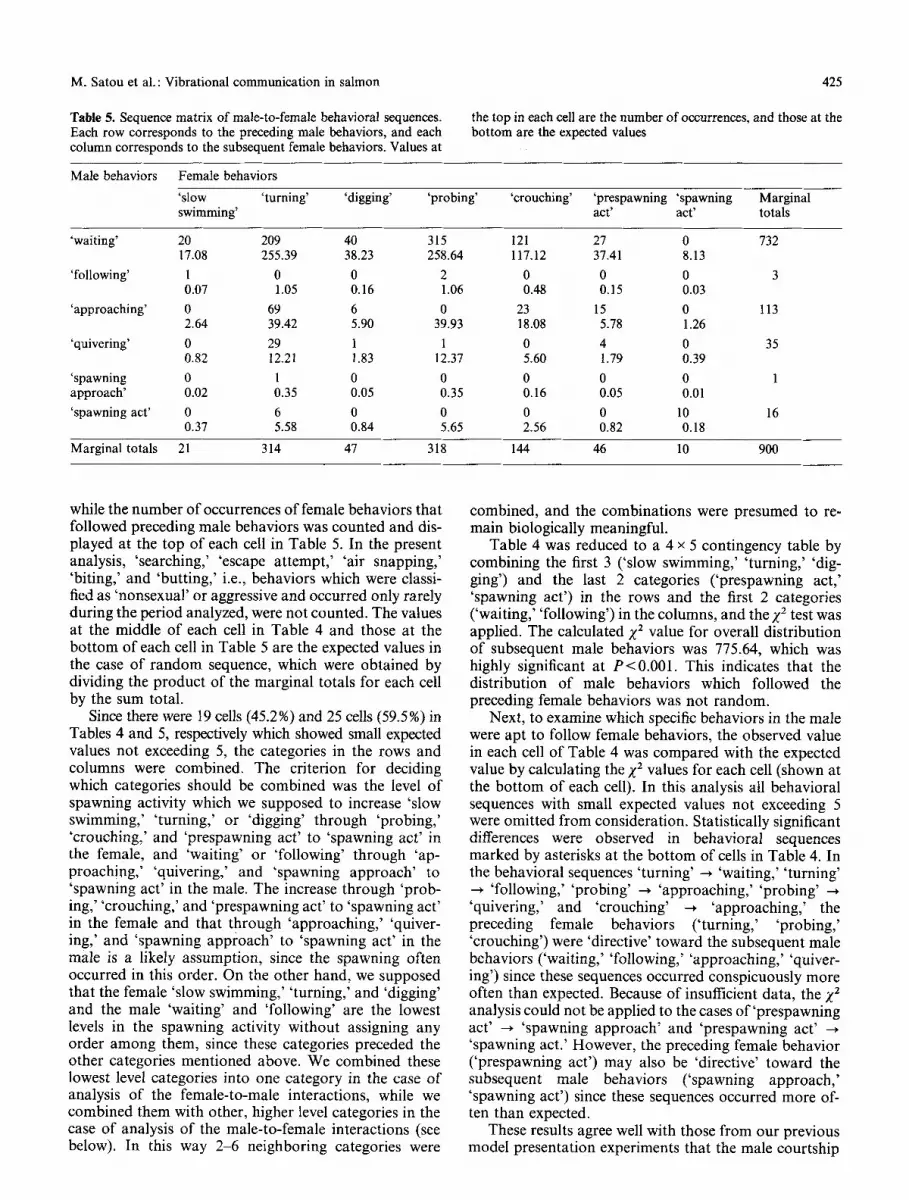
Table 5. Sequence matrix of male-to-female behavioral sequences. Each row corresponds to the preceding male behaviors, and each column corresponds to the subsequent female behaviors. Values at
425
the top in each cell are the number of occurrences, and those at the bottom are the expected values
Male behaviors Female behaviors
'slow 'turning' 'digging' 'probing' 'crouching' 'prespawning 'spawning Marginal swimming' act' act' totals
'waiting' 20 209 40 315 121 27 0 732 17.08 255.39 38.23 258.64 117.12 37.41 8.13
'following' 1 0 0 2 0 0 0 3 0.07 1.05 0.16 1.06 0.48 0.15 0.03
'approaching' 0 69 6 0 23 15 0 113 2.64 39.42 5.90 39.93 18.08 5.78 1.26
'quivering' 0 29 1 1 0 4 0 35 0.82 12.21 1.83 12.37 5.60 1.79 0.39
'spawning 0 1 0 0 0 0 0 1 approach' 0.02 0.35 0.05 0.35 0.16 0.05 0.01
'spawning act' 0 6 0 0 0 0 10 16 0.37 5.58 0.84 5.65 2.56 0.82 0.18
Marginal totals 21 314 47 318 144 46 10 900
while the number of occurrences of female behaviors that followed preceding male behaviors was counted and dis- played at the top of each cell in Table 5. In the present analysis, 'searching,' 'escape attempt,' 'air snapping,' 'biting,' and 'butting,' i.e., behaviors which were classi- fied as 'nonsexual' or aggressive and occurred only rarely during the period analyzed, were not counted. The values at the middle of each cell in Table 4 and those at the bottom of each cell in Table 5 are the expected values in the case of random sequence, which were obtained by dividing the product of the marginal totals for each cell by the sum total.
Since there were 19 cells (45.2%) and 25 cells (59.5%) in Tables 4 and 5, respectively which showed small expected values not exceeding 5, the categories in the rows and columns were combined. The criterion for deciding which categories should be combined was the level of spawning activity which we supposed to increase 'slow swimming,' 'turning,' or 'digging' through 'probing,' 'crouching,' and 'prespawning act' to 'spawning act' in the female, and 'waiting' or 'following' through 'ap- proaching,' 'quivering,' and 'spawning approach' to 'spawning act' in the male. The increase through 'prob- ing,' 'crouching,' and 'prespawning act' to 'spawning act' in the female and that through 'approaching,' 'quiver- ing,' and 'spawning approach' to 'spawning act' in the male is a likely assumption, since the spawning often occurred in this order. On the other hand, we supposed that the female 'slow swimming,' 'turning,' and 'digging' and the male 'waiting' and 'following' are the lowest levels in the spawning activity without assigning any order among them, since these categories preceded the other categories mentioned above. We combined these lowest level categories into one category in the case of analysis of the female-to-male interactions, while we combined them with other, higher level categories in the case of analysis of the male-to-female interactions (see below). In this way 2-6 neighboring categories were
combined, and the combinations were presumed to re- main biologically meaningful.
Table 4 was reduced to a 4 x 5 contingency table by combining the first 3 ('slow swimming,' 'turning,' 'dig- ging') and the last 2 categories ('prespawning act,' 'spawning act') in the rows and the first 2 categories ('waiting,' 'following') in the columns, and the Z 2 test was applied. The calculated Z 2 value for overall distribution of subsequent male behaviors was 775.64, which was highly significant at P<0.001. This indicates that the distribution of male behaviors which followed the preceding female behaviors was not random.
Next, to examine which specific behaviors in the male were apt to follow female behaviors, the observed value in each cell of Table 4 was compared with the expected value by calculating the Z 2 values for each cell (shown at the bottom of each cell). In this analysis all behavioral sequences with small expected values not exceeding 5 were omitted from consideration. Statistically significant differences were observed in behavioral sequences marked by asterisks at the bottom of cells in Table 4. In the behavioral sequences 'turning' ~ 'waiting,' 'turning'
'following,' 'probing' ~ 'approaching,' 'probing' 'quivering,' and 'crouching' -~ 'approaching,' the preceding female behaviors ('turning,' 'probing,' 'crouching') were 'directive' toward the subsequent male behaviors ('waiting,' 'following,' 'approaching,' 'quiver- ing') since these sequences occurred conspicuously more often than expected. Because of insufficient data, the Z 2 analysis could not be applied to the cases of'prespawning act' -~ 'spawning approach' and 'prespawning act' --. 'spawning act.' However, the preceding female behavior ('prespawning act') may also be 'directive' toward the subsequent male behaviors ('spawning approach,' 'spawning act') since these sequences occurred more of- ten than expected.
These results agree well with those from our previous model presentation experiments that the male courtship

426 M. Satou et al.: Vibrational communication in salmon
behaviors ('approaching,' 'quivering') were elicited by a model which mimicked the female performing 'probing' or 'crouching' (Takeuchi et al. 1987) and that the male spawning behaviors ('spawning approach,' 'spawning act') were elicited by a vibrating model which mimicked the female performing the 'prespawning act' (Satou et al. 1987; unpublished results). Present statistical results showing that the male 'waiting' and 'following' were apt to follow the female 'turning' (Table 4) also suggest that these male behaviors are elicited by the female 'turning.' As to the other behavioral sequences which showed sta- tistically significant differences, the preceding female be- haviors were 'inhibitive' toward the subsequent male behaviors, since these sequences occurred conspicuously less often than expected (Table 4). Here, the terms 'direc- tive' and 'inhibitive' were used in a purely statistical sense (Hazlett and Bossert 1965; Dingle 1969).
Table 5 was reduced to a 2 x 2 contingency table by combining the first 5 categories ('waiting,' 'following,' 'approaching,' 'quivering,' 'spawning approach') in the rows and the first 6 categories ('slow swimming,' 'turn- ing,' 'digging,' 'probing,' 'crouching,' 'prespawning act') in the columns, and Fisher's exact probability test was applied. The calculated probability was 8.76x 10 -2~ This indicates that the distribution of female behaviors which followed the preceding male behaviors was not random. Because of insufficient data, we did not statistic- ally examine which specific behaviors in the female were apt to follow the male behaviors. However, from com- parison between the observed and expected values the male 'spawning act' may be 'directive' toward the female 'spawning act' (Table 5).
Furthermore, analysis of behavioral sequences was performed based on the information theory (Hazlett and Bossert 1965; Dingle 1969). After reducing Table 4 to a 4 x 5 contingency table by combining the first 3 ('slow swimming,' 'turning,' 'digging') and the last 2 categories ('prespawning act,' 'spawning act') in the rows and the first 2 categories ('waiting,' 'following') in the columns, amounts of information, H(S), Hmax(S), H(R), Hmax(R), H(S,R), Hmax(S,R), and T(S,R), were cal- culated (Table 6). Miller's Z 2 test revealed that H(S), H(R), and H(S,R), differed significantly (P < 0.001) from Hmax(S), Hmax(R), and Hmax(S,R), respectively. This
Table 6. Amounts of information calculated
Amounts of information (bit)
H(S) 1.88" H (S) 2.00 H(R) 1.82"* H,~,,(R) 2.32 H(S, R) 3.10"** Hm,,,(S, R) 4.32 T(S, R) 0.60 +
*, ** *** Differed significantly at P< 0.001 from H x(S ), Hm~x(R), and FI=~x(S, R), respectively; + Differed significantly at P< 0.001 from zero. Tested by Miller's method (Colgan 1978; Kasuya and Fujita 1984)
indicates that the female behaviors preceded, the male behaviors followed, and the female-to-male behavioral sequences did not occur randomly, confirming the results of the sequence matrix analysis mentioned above. Sim- ilarly, T(S,R) differed significantly (P < 0.001) from zero. This indicates that statistically significant amounts of information were transmitted during female-to-male in- teractions. The efficiency of information transmission, TE = T(S,R)/H(S) was 0.32, indicating that relatively ef- fective transmission of the information occurred. Be- cause of insufficient data, we did not apply similar statis- tical tests to the case of male-to-female interactions.
Discussion
The present results have shown that the spawning behav- iors of the him6 salmon are composed of an accurately timed sequence of characteristic vibrational behaviors with characteristic rhythmic trunk muscle activity. Body vibration detected during the male 'quivering,' male 'spawning act,' female 'prespawning act,' and female 'spawning act' was described in both time and frequency domains. The observation that body vibration in one sex was followed by a certain behavior in the other (Fig. 3; Table 1) and the fact that the effective frequency range of a vibrating 'female' model for eliciting the male 'spawning act' was close to that of body vibration during the female 'prespawning act' in natural behavior (Fig. 4; Table 3; unpublished results) suggest that vibrational signals are actually exchanged and are used as timing cues to synchronize gamete release. The observation that the body of the recipient animal was sometimes vibrated passively also supports this view. Statistical analysis of the male-female interactions during spawning also showed that significant interactions occurred and signifi- cant amounts of information were transmitted through the interactions (Tables 4-6). The finding that the female did not vibrate her body during the early,phase of the 'spawning act' (Fig. 3Ba, right) should be stressed, since this was not noticed in previous visual observations and has revealed an important property of the signalling system (see below).
The observations that the female 'spawning act' and accompanying egg release occurred only if the male per- formed his 'spawning act' in a close position and at an appropriate time suggest that the female carries out her 'spawning act' if she has received vibrational signals which are strong enough and correctly timed. However, the involvement of other sensory cues, such as visual ones, for eliciting the female 'spawning act' should not be overlooked, since we previously showed the impor- tance of visual cues for eliciting the male 'spawning act' (Satou et al. 1987). The idea that the visual cues are important for eliciting the male 'spawning act' was also supported by the present observation that the female occasionally carried out her 'prespawning act' with body vibration of a barely detectable magnitude. Presumably, besides the vibrational cues, the postural cues during the female 'prespawning act' may also be involved in eliciting the male 'spawning act.'

M. Satou et al. : Vibrational communicat ion in salmon 427
The present recording experiments revealed that the mean magnitude of body vibration during the male 'quivering,' male 'spawning act,' or female 'prespawning act' ranged from 1.10 to 3.85 G (Table 2). The accelera- tion of the 'near field-local flow' of the water particles caused by a sinusoidally vibrating 'sphere' decreases ap- proximately in proportion to the inverse cube of the radial distance from the vibrating source (Harris and Van Bergeijk 1962; Kalmijn 1988). In contrast to the case of a vibrating sphere, it decreases approximately in proportion to the inverse square of the radial distance in the case of sinusoidally vibrating 'cylinder' with infinite length, as is given by A = a2/r 2 • A 1 • COS 0 • sin to t, where A : acceleration, to: angular frequency, a: radius of vibrating cylinder, r: radial distance, AI: amplitude of axial source acceleration, 0: angle of radiation, t: time (unpublished results). If we suppose that a = 2 cm, r - 3 cm, A~=1.10-3.85 G, 0=0, a maximal value of A, Ama~= 0.49-1.71 G is obtained. This value seems to be a rough estimate that the recipient animal actually re- ceives the vibrational signals at its body surface. It much exceeds the threshold (an order of 10 -4 G) of the lateral line detectors in the him6 salmon (unpublished results) as well as the known threshold (an order of 10 - 5 - 10 -3 G ~) in other fishes (Harris and van Bergeijk 1962; Bleck- mann 1980; Bleckmann and Topp 1981; Sand 1981; Coombs and Janssen 1989).
These considerations suggest that the lateral line sense is one likely candidate involved in the detection of vibra- tional signals during intersexual interactions in the him6 salmon (Satou 1987). In agreement with this hypothesis, we have recently shown that the cobalt ion, which blocks the mechanoelectrical transduction in the lateral line detectors (Sand 1975; Karlsen and Sand 1987), sup- presses the spawning in the the male him6 salmon which was elicited by a vibrating 'female' model (Satou et al., in press). However, since we observed that one body of an interacting pair was sometimes vibrated passively, there remains a possibility that the tactile receptor and/or inner ear (besides the mechanoreceptive lateral line) are also involved in detecting the vibrating neighbor.
Smith (1965) distinguished between the 'message' and the 'meaning' involved in the signals. The message is what the signal encodes about the signaller's state, while the meaning is the information extracted from the signals by the receiver. In the communication system of the him6 salmon, signals from the female 'prespawning act' are likely to contain a message as to her readiness for egg release, while those from the male 'spawning act' contain a message as to the start of milt release. The finding that the frequency of body vibration was characteristic of each vibrational behavior (Fig. 4; Table 3) suggests that
t In previous reports the threshold had been expressed as 'displace- ment ' at a particular frequency. Here, this was given as 'accelera- t ion' by calculating according to the formula: A = (2nf)2x D, where A: acceleration, f : frequency, D: displacement. However, it should be noted that this calculation was done for the purpose of comparison, and we do not intend to argue that the lateral line detectors of the him6 salmon are designed to detect 'acceleration' rather than 'displacement' (or 'velocity'). This is yet to be deter- mined.
differences in the messages involved in the signals are reflected in alterations in the vibrational frequency.
On the other hand, since little body vibration was produced during the female 'spawning act' except for its later phase (i.e., a phase after egg release) (Fig. 3Ba, right), it is likely that the female does not send any signals with messages as to the time of egg release. Observations that the male sometimes released milt without leading to egg release in the female (Fig. 5) support this view. Thus, the female seems to 'eavesdrop' on signals from the male as to the time of milt release without sending back any return signals to the male. This may be an example of 'manipulation' of the signaller by the receiver (Dawkins and Krebs 1978), since the receiver (female) unilaterally gets the benefits (information as to the timing of sperm release) from the signaller (male).
In considering the origin and evolution of vibrational signals, the remarkable differences in the EMG activity patterns (rhythmic vs. arrhythmic) between the male and female 'spawning act' (which are 'isomorphic' behaviors indistinguishable from each other by the naked eye) are interesting. A function of the trunk muscle activity during spawning is undoubtedly to squeeze out the gametes by increasing the internal body pressure (Uematsu and Yamamori 1982; Matsushima et al. 1989). The most effective way to do so is to contract all trunk muscles simultaneously, which produces an arrhythmic EMG activity pattern, as was seen during the female 'spawning act' (Fig. 3Bb, c, right). This inevitably results in stiffening of the body and hence disturbs the produc- tion of the body vibration. This was actually seen during the female 'spawning act' (Fig. 3Ba, right).
In contrast, the EMG activity during the male 'spawn- ing act' showed a regular, rhythmic pattern with a large amplitude (Fig. 3Be, f, right; Fig. 5Be, f), which caused a strong body vibration (Fig. 3Bd, right; Fig. 5Bd). Such a pattern is thought to have dual functions: to produce the body vibration (signal production) and to increase the internal body pressure (gamete release). Since inter- mittent muscular activity alternating on each side of the body is the most effective way to produce the body vibration but a less effective way to increase the internal body pressure, it may be hypothesized that the vibration- al signals from the male 'spawning act' stem from the muscular activity for milt release and have evolved to- wards increased signalling efficiency. Body vibration during the female 'prespawning act' may have a similar origin related to egg release. On the other hand, since the female 'spawning act,' in contrast to the male 'spawning act,' does not have any signalling function concerning the timing of gamete release (see above), the observed ar- rhythmic EMG pattern during the female 'spawning act,' which is most effective for egg release but is ineffective in producing vibrational signals, may have been pre- served during the course of evolution of the signalling system.
Due to the short transmission distances of vibrational signals, they are used for close-range 'private' communi- cation in a variety of social interactions of animals (Markl 1983). The best-known example is the courtship vibration of drosophilid flies (Bennet-Clark 1971). Since

428 M. Satou et al. : Vibrational communication in salmon
a n u m b e r o f pa i rs o f s a l m o n s imul t aneous ly spawn in a reas where m a n y nests are c rowded close together , and the spawning genera l ly occurs at twi l ight when vis ion is reduced, the ' nea r field' v ib ra t iona l s ignals ac t ing as nonv i sua l c lose- range signals a n d hence free o f inter- ference ( ' j a m m i n g ' ) f rom ne ighbor ing pa i rs m a y well be sui ted for such s i tuat ions .
M a n y fish species wi th a g rea t d ivers i ty o f r ep roduc - tive styles exhibi t b o d y v ib r a t i on dur ing spawning (Breder a n d R o s e n 1966; Keen leys ide 1979). This sug- gests t ha t v ib r a t i ona l s ignals are very bas ic a m o n g m a n y fishes and are used as t iming signals for gamete release, as p rev ious ly po in t ed ou t by Keen leys ide (1979). To test this hypo thes i s fur ther , exper imen t s f rom a c o m p a r a t i v e view p o i n t a n d us ing phys io log ica l as well as exper imen- tal behav io ra l me thods , such as those used in the p resen t s tudy, a re requi red .
Acknowledgements. We thank Dr. A. D. Munro for critically read- ing an early version of the manuscript, and Dr. T. Maruyama, the Director of National Research Institute of Aquaculture, Nikko Branch, and the staff of the Lake Chuzenji Fishery Association for kindly providing facilities for the experiments. We also thank Dr. H.-A. Takeuchi for valuable discussion in preparing the manuscript and for help in the initial stage of the recording experiments. Sup- ported by grants from the Ministry of Education, Science and Culture of Japan, the Ministry of Agriculture, Forestry and Fish- eries of Japan, and the Mitsubishi Foundation.
References
Bennet-Clark HC (1973) Acoustics of insect song. Nature 234:255-259
Bleckmann H (1980) Reaction time and stimulus frequency in prey localization in the surface-feeding fish Aplocheilus lineatus. J Comp Physiol 140:163-172
Bleckmann H, Topp G (1981) Surface wave sensitivity of the lateral line organs of the topminnow Aplocheilus lineatus. Naturwissen- schaften 68 : 624-625
Breder CM Jr, Rosen DE (1966) Modes of reproduction in fishes. The Natural History Press, Garden City
Childerhose RJ, Trim M (1979) Pacific salmon. Douglas and Mclntyre Ltd, Vancouver
Colgan PW (ed) 1978 Quantitative ethology. Academic Press, New York London
Coombs S, Janssen J (1989) Peripheral processing by the lateral line system of the mottled sculpin (Cottus bairdi). In : Coombs S, G6rner P, M/inz H (eds) The mechanosensory lateral line: Neurobiology and evolution. Springer, Berlin Heidelberg New York, pp 299-319
Dawkins R, Krebs JR (1978) Animal signals: information or mani- pulation? In: Krebs JR, Davies NB (eds) Behavioural ecology. Blackwell Scientific Publications, Oxford, pp 282-309
Dingle H (1969) A statistical and information analysis of aggres- sive communication in the mantis shrimp Gonodactylus bredini. Anim Behav 17: 567-581
Harris GG, van Bergeijk WA (1962) Evidence that the lateral-line organ responds to near-field displacements of sound sources in water. J Acoust Soc Am 34:1831-1941
Hazlett BA, Bossert WH (1965) A statistical analysis of the aggres- sive communication systems of some hermit crabs. Anim Behav 13 : 357-373
Jones JW, King GM (1949) Experimental observations on the spawning behaviour of the Atlantic salmon (Salmo salar Linn.). Proc Zool Soc Lond 119:33-48
Kalmijn AJ (1988) Hydrodynamic and acoustic field detection. In: Atema J, Fay RR, Popper AN, Tavolga WN (eds) Sensory biology of aquatic animals. Springer, Berlin Heidelberg New York, pp 83-130
Karlsen HE, Sand O (1987) Selective and reversible blocking of the lateral line in freshwater fish. J Exp Biol 133:249-262
Kasuya E, Fujita K (1984) Statistics for ethology. Tokai Univ Press, Tokyo (in Japanese)
Keenleyside MHA (1979) Diversity and adaptation in fish behav- iour. Springer, Berlin Heidelberg New York
Markl H (1983) Vibrational communication. In: Huber F, Markl H (eds) Neuroethology and behavioral physiology. Springer, Ber- lin Heidelberg New York, pp 332-353
Matsushima T, Kitamura S, Takei K, Satou M, Okumoto N, Ueda K (1986) An electromyographic study of sexual behavior in him6 salmon (landlocked red salmon, Oncorhynchus nerka). Zool Sci 3 : 563-567
Matsushima T, Takei K, Kitamura S, Kusunoki M, Satou M, Okumoto N, Ueda K (1989) Rhythmic electromyographic ac- tivities of trunk muscles characterize the sexual behavior in the him~ salmon (landlocked sockeye salmon, Oneorhynchus nerka). J Comp Physiol A 165:293-314
Nakano E (1969) Fishes. In: Metz CB, Monroy A (eds) Fertiliza- tion; comparative morphology, biochemistry, and immunology, vol 2. Academic Press, New York, pp 295-324
Sand O (1975) Effects of different ionic environments on the mecha- nosensitivity of lateral line organs in the mudpuppy. J Comp Physiol 102:27-42
Sand O (1981) The lateral line and sound reception. In: Tavolga WN, Popper AN, Fay RR (eds) Hearing and sound communi- cation in fishes. Springer, Berlin Heidelberg New York, pp 459-478
Satou M (1987) A neuroethological study of reproductive behavior in the salmon. Proc 3rd Int Symp Reprod Physiol Fish, pp 154-159
Satou M, Ueda K (1982) Brain mechanisms of salmon sexual behavior. In: Onishi E, Hidaka T (eds) Mechanisms of sexual behavior. Sangyo Tosho, Tokyo, pp 5-19 (in Japanese)
Satou M, Oka Y, Kusunoki M, Matsushima T, Kato M, Fujita I, Ueda K (1984) Telencephalic and preoptic areas integrate sexual behavior in him6 salmon (landlocked red salmon, Oncorhynchus nerka) : results of electrical brain stimulation experiments. Phys- iol Behav 33:441-447
Satou M, Takeuchi H, Takei K, Hasegawa T, Okumoto N, Ueda K (1987) Involvement of vibrational and visual cues in eliciting spawning behaviour in male him6 salmon (landlocked red sal- mon, Oncorhynchus nerka). Anim Behav 35:1556-1558
Satou M, Takeuchi H-A, Kitamura S, Kudo Y, Nishii J, Tanabe M (1991) Involvement of lateral line sense in intersexual vibra- tional communication during spawning behavior in the him6 salmon (landlocked red salmon, Oncorhynchus nerka). Neurosci Res Suppl 12 (in press)
Smith WJ (1965) Message, meaning and context in ethology. Am Naturalist 99: 405-409
Takeuchi H, Takei K, Satou M, Matsushima T, Okumoto N, Ueda K (1987) Visual cues as key stimuli for courtship behav- iour in male him6 salmon (landlocked red salmon, Oncorhyn- chus nerka). Anim Behav 35:936-939
Tautz AF, Groot C (1975) Spawning behavior of chum salmon (Oncorhynchus keta) and rainbow trout (Salmo gairdneri). J Fish Res Bd Can 32:633-642
Uematsu K, Yamamori K (1982) Body vibration as a timing cue for spawning in chum salmon. Comp Biochem Physiol 72A: 591-594