on the developmental pathway to nonanadromy in sockeye salmon, oncorhynchus nerka
TRANSCRIPT

On the developmental pathway to nonanadromy in sockeye salmon, Oncorhynchus nerka
CHRIS J. FOOTE School of' Fisheries WH-10, University of Washington, Seattle, WA 981 85, U.S.A.
IAN MAYER~ West Vancouver Laboratory, Department of Fisheries and Oceans, 4160 Marine Drive, West Vancouver, BC V7V lN6,
Canada
AND
CHRIS C. WOOD, W. CRAIG CLARKE, AND J. BLACKBURN Pat+ Biological Station, Department of Fisheries and Oceans, Nanaimo, BC V9R 5K6, Canada
Received June 14, 1993 Accepted January 10, 1994
FOOTE, C.J., MAYER, I., WOOD, C.C., CRAIG, W., and BLACKBURN, J. 1994. On the developmental pathway to nonanadromy in sockeye salmon, Oncorhynchus nerka. Can. J. Zool. 72: 397 -405 .
The evolution of nonanadromous forms of salmonids has been linked to adaptive similarities between migration to the sea and migration to lakes, and to inhibition of smoltification by rapid development rates and early maturation. We examined the seasonal cycle of seawater adaptability in a landlocked (10 000 years) population of kokanee, Oncorhynchus nerka, to determine if the typical anadromous cycle persists in the absence of recent gene flow from sockeye salmon, as would be expected if components of smoltification are beneficial to a lacustrine existence. We also monitored the effects of maturation on seawater adaptability, to determine if they had an inhibitory effect on smoltification, as would be expected if a developmental conflict was responsible for the evolution of nonanadromous forms. Maturation was measured through the growth of gonadal tissue and the increase in circulating testosterone and I 1 -ketotestosterone. Landlocked kokanee showed the seawater adaptability cycle typical of sockeye salmon, and maturation had no inhibitory effect on seawater adaptability until near, or at, full maturity. We conclude that at least some components of smoltification must be beneficial to a life in lakes to account for the continued maintenance of the cycle. Further, rapid development and maturation on their own do not appear to account for the evolution of nonanadromous populations of 0 . nerka.
FOOTE, C.J., MAYER, I., WOOD, C.C., CRAIG, W., et BLACKBURN, J. 1994. On the developmental pathway to nonanadromy in sockeye salmon, Oncorhynchus nerka. Can. J. Zool. 72 : 397-405.
L'kvolution des formes anadromes de salmonidks est peut-Etre attribuable d'une part a des similaritks entre l'adaptation a la migration a la mer et l'adaptation a la migration vers les lacs et, d'autre part, a l'klimination du stade saumonneau entrainke par un dkveloppement rapide et une maturation prkcoce. Nous avons ktudik le cycle saisonnier de l'adapatibilitk a l'eau de mer chez une population non anadrome (en eau douce depuis 10 000 ans) de Kokanis, Oncorhynchus nerka, afin de dkterminer si le cycle anadrome typique persiste mEme s'il n'y a pas eu de flux rkcent de gttnes de Saumons rouges anadromes, phknomttne auquel on doit s'attendre si la tranformation en saumonneau comporte des avantages pour les poissons qui vivent en eau douce. Nous avons kgalement mesurk les effets de la maturation sur l'adaptabilitk a l'eau de mer afin de dkterminer si ces effets inhibent la transformation en saumonneau, comme on devrait s'y attendre si c'est un problkme de dkveloppement qui est responsable de l'existence des formes non anadromes. La maturation est kvaluke en fonction de la croissance des tissus gonadiques et de l'augmentation de la testostkrone et de la ckto-1 1 testostkrone en circulation. Les saumons d'eau douce ont manifest6 le cycle d'adaptabilitk 21 l'eau de mer typique des saumons anadromes et la maturation n'a eu d'effet inhibiteur sur l'adaptabilitk a l'eau de mer qu'a la toute fin. 11 semble donc qu'au moins certaines composantes de la transformation en saumonneau comportent des avantages pour la vie en eau douce puisque le cycle persiste toujours. Le dkveloppement rapide et la maturation ne semblent pas pouvoir expliquer a eux seuls l'kvolution des populations non anadromes d'O. nerka.
[Traduit par la Rkdaction]
Introduction Salmonid species may comprise solely anadromous individ-
uals, nonanadromous individuals, or some combination of both (Thorpe 1987). Nonanadromous individuals may be members of polymorphic anadromous-nonanadromous populations (e.g., Jones and King 1952; Krogius 1981; Saunders et al. 1982; Aida et al. 1984; Jonsson 1985; Langdon and Thorpe 1985; Dellefors and Faremo 1988; Hutchings and Myers 1988; Taylor 1989; Lundqvist et al. 1989; Jordan and Youngson 1992) or they may form their own discrete populations in either the presence or absence of the anadromous form (e.g., Maekawa 1977; Hindar and Jonsson 1982; Berg 1985; Skulason et al. 1989; Foote et al. 1989).
'Present address: Department of Zoology, University of Stockholm, S- 106 9 1 Stockholm, Sweden.
There is debate on how nonanadromous populations arose from anadromous populations. Undoubtedly, geological events following the retreat of the last glaciation played a major role, physically "locking" fish into freshwater environments (Ricker 1940; Nelson 1968; Berg 1985; Thorpe 1987). Also, it is clear that anadromous forms often give rise to nonanadromous individuals within populations (see references above). These nonanadromous individuals are typically males that adopt an alternative mating tactic, "sneaking" in on the spawning of anadromous pairs (e.g., Hutchings and Myers 1988; Jordan and Youngson 1992). However, the physiological basis on which nonanadromous individuals and populations arose, and continue to arise, is not clear. In the case of Atlantic salmon, Salmo sa la r , Berg (1985) pointed out the clear association between strictly nonanadromous populations and a lake-reared existence. He speculated that many of the traits associated with
Printed in Canada 1 Imprime au Canada
Can
. J. Z
ool.
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
Fish
erie
s an
d O
cean
s on
08/
08/1
3Fo
r pe
rson
al u
se o
nly.

398 CAN. J. ZOOL. \ 'OL. 72. 1994
smoltification may also be beneficial to freshwater rearing in lakes, thus making permanent lake residence a relatively minor evolutionary step away from anadromy. In contrast, Thorpe (1986, 1987) has pointed to a developmental conflict between smoltification and maturation as the basis for the evolution of nonanadromous populations: that is, traits selected for in a freshwater environment are incompatible with those selected for in a marine environment. Factors that led to rapid development rates in fresh water, whether environmental and (or) genetic, were thought to stimulate early maturation and inhibit the smoltification process, thus leading to the existence of nonanadromous individuals.
Anadromous and nonanadromous individuals are com- mon in sockeye salmon, Oncorhynchus nerka. Nonanad- romous individuals may occur within polymorphic populations, of which a proportion, usually males, remain in fresh water throughout their lives (Ricker 1938; Smirnov 1959; Krogius 1981), or they may occur as genetically distinct populations from sockeye salmon, where they are commonly referred to as kokanee (Foote et al. 1989; Wood and Foote 1990; Taylor and Foote 1991). Kokanee may occur allopatrically or sympatrically with sockeye salmon and in some cases they spawn together in the same place and at the same time (Hanson and Smith 1967; Nelson 1968; McCart 1970; Foote and Larkin 1988).
Kokanee have arisen from sockeye salmon on numerous occasions (Ricker 1940; Nelson 1968; Foote et al. 1989) but, as is the case in Atlantic salmon, the mechanism by which one form has arisen from the other remains unclear. Ricker (1938, 1940) showed that nonanadromous sockeye salmon (termed "residuals" or "residents") were the fastest growing individuals, supporting Thorpe's (1987) linkage between increased development rate and nonanadromy. However, Foote et al. (1 992) found that kokanee sympatric with sockeye salmon maintained the seasonal cycle of seawater adapt- ability necessary for migration to the sea, suggesting some benefits to be found in the continued maintenance of components of smoltification in a freshwater existence, and thus providing indirect support for Berg's view of the evolution of nonanadromy. Further, this smoltification process was not inhibited by maturation. However, Foote et al. (1992) were unable to rule out the possibility that components of smoltification in kokanee were maintained simply as the result of persistent gene flow, or their recent divergence, from sympatric sockeye salmon. Also, they did not measure the concentrations of circulating androgens in maturing kokanee, the agents thought to be responsible for the inhibition of smoltification (e.g., Miwa and Inui 1985; Lundqvist et al. 1989).
In this paper, we address the possible mechanisms for the evolution of nonanadromy in 0. nerka. If components of smoltification are beneficial to a lacustrine existence we may expect to find the typical anadromous seasonal cycle of seawater adaptability maintained in kokanee populations long isolated physically from the sea (and hence from sockeye salmon). Further, if a developmental conflict between smoltification and maturation has led to the existence of nonanadromous populations, then we may expect to find lowered seawater tolerance in association with gonad growth and increasing concentrations of circu- lating plasma androgens, as has been the case for all other salmonids examined.
Methods Broodstock collection and rearing environment
On September 12, 1989, we collected kokanee from Meadow Creek spawning channel, Kootenay Lake, using a small seine. Kootenay Lake is on the headwaters of the Columbia River and has been physically isolated from the sea by a waterfall at its outlet for approximately 10 000 years (Northcote 1973). We created 12 inde- pendent full-sib families. The fertilized eggs were placed in individual I-L jars filled with creek water, packed in coolers in an ice-water bath, and immediately transported by truck and plane to the Rosewall Creek hatchery on Vancouver Island, where they were incubated at approximately 8S°C until January 2, 1990. At this point, equal allotments of the 12 families were combined to create a mixed-family population from which later samples were taken. The pooled fish were raised in three 400-L tanks at initial densities of approximately 200 fish per tank. In mid-December 1990 we combined fish from these three tanks and placed them in a single 1800-L tank with an original density of 400 fish. All subsequent samples were taken from this tank.
All tanks were maintained on a simulated natural-light schedule at a nearly constant temperature of 8.5"C. Food of appropriate size (Biodiet Starter Mix, Whitecrest Mills Hatchery, and Whitecrest Mills Salmon Developer) was dispensed by automatic feeders every half hour during daylight, at a rate of 3.8-1 .O% body mass per fish each day, decreasing progressively over time.
In the spring of each year we compared the seawater adaptability of Kootenay Lake kokanee with sockeye salmon from other lake systems to obtain a direct measure of their performance relative to their allopatric anadromous counterpart. The sockeye salmon populations used spanned a large geographic range in British Columbia. The specific sockeye salmon population used varied with availability from year to year. We compared underyearlings of the Kootenay Lake kokanee with those of sockeye salmon from Babine Lake (Skeena River drainage) and Takla Lake (upper Fraser River). We compared yearlings of the Kootenay kokanee with those of sympatric sockeye salmon and kokanee from Shuswap Lake (Fraser River drainage, B.C.). The Takla and Babine sockeye salmon sampled were from the combined progeny of 3 and 4 full-sib families each, respectively. The parents were spawned on August 14, 1989, incubated at Rosewall Creek hatchery at 8S°C, placed in one of two 400-L rearing tanks on December 6, 1989, and raised at a density of 300 fry per tank. The Shuswap sockeye salmon and kokanee were the progeny of parents raised in the hatchery. The parents were the siblings of those families used in our previous examination of seawater adaptability in sympatric sockeye salmon and kokanee (Foote et al. 1992). Five full-sib families of each group were created on October 16, 1989, incubated at 8S°C, and transferred to the pond on February 7, 1990. Progeny were combined by cross-type, and raised at a density of 170 fry per 400-L tank until mid-December 1990, at which point they were combined with other Shuswap crosses and raised at a density of 520 fish in a 1800-L tank. Seawater adaptability tests
The seawater adaptability of Kootenay Lake kokanee was measured by 24-h seawater challenge tests carried out at the Pacific Biological Station on 11 occasions from 4 to 21 months after swim-up (see Blackburn and Clarke 1987 for challenge test details). At least 4 d before each test, 20-45 fish were transported from Rosewall Creek hatchery to the Pacific Biological Station, where they were maintained without food in a tank(s) of dechlorinated Nanaimo city water (8.5-9.0°C). After 4 days, the fresh water was displaced by natural seawater (29-30%) at a controlled temperature of 8.5-9.0°C. After 24 h in seawater each fish was sacrificed in anaesthetic (500 ppm 2-phenoxyethanol) and a blood sample was taken from the caudal artery by means of a capillary tube. This sample was analyzed for plasma sodium concentration using a Turner (Case Instrument) flame photometer. Fish that have acquired the hypoosmoregulatory capacity for living in seawater typically show plasma sodium levels lower than 170 mmol/L after seawater challenge, whereas fish that
Can
. J. Z
ool.
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
Fish
erie
s an
d O
cean
s on
08/
08/1
3Fo
r pe
rson
al u
se o
nly.
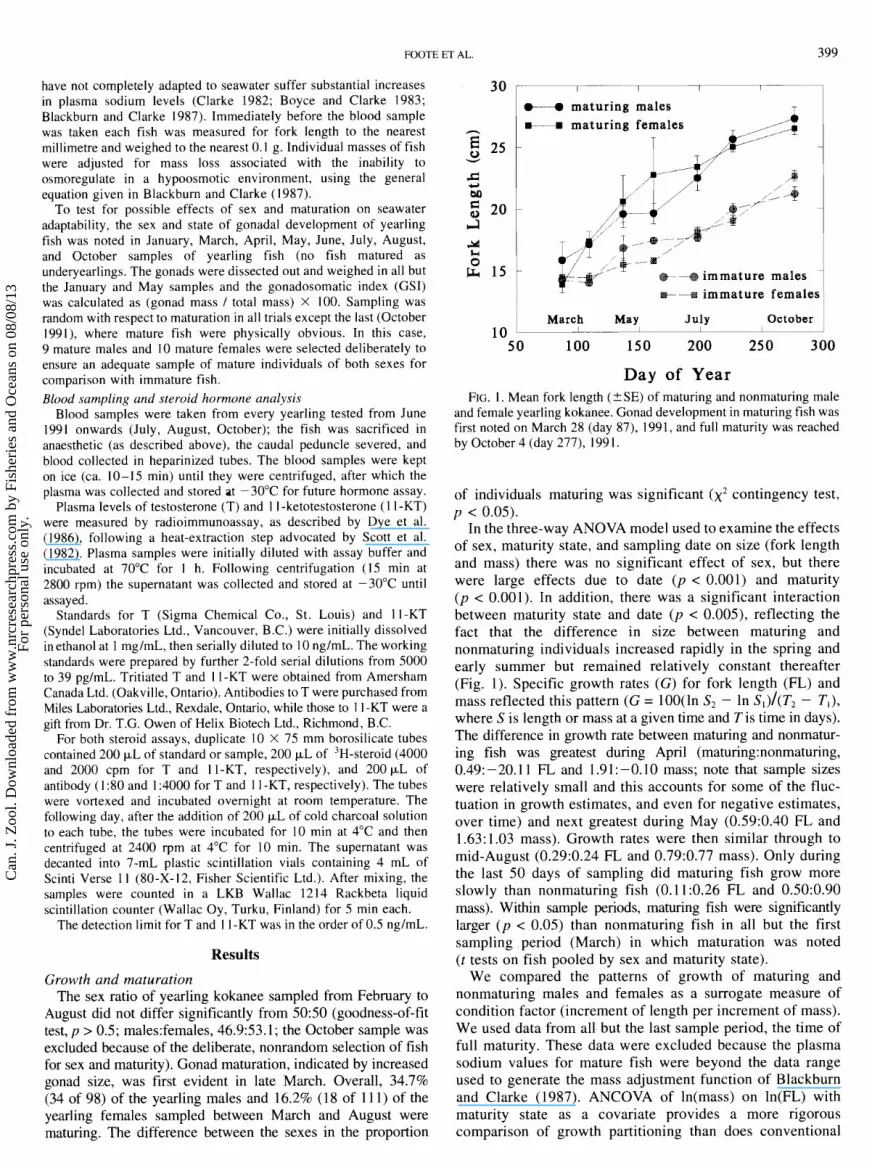
FOOTE ET AL. 399
have not completely adapted to seawater suffer substantial increases 30 r-- T - 7
in plasma sodium levels (Clarke 1982; Boyce and Clarke 1983; Blackburn and Clarke 1987). Immediately before the blood sample was taken each fish was measured for fork length to the nearest millimetre and weighed to the nearest 0.1 g. Individual masses of fish were adjusted for mass loss associated with the inability to osmoregulate in a hypoosmotic environment, using the general equation given in Blackburn and Clarke (1987).
To test for possible effects of sex and maturation on seawater adaptability, the sex and state of gonadal development of yearling fish was noted in January, March, April, May, June, July, August, and October samples of yearling fish (no fish matured as underyearlings. The gonads were dissected out and weighed in all but the January and May samples and the gonadosomatic index (GSI) was calculated as (gonad mass / total mass) X 100. Sampling was random with respecito maturation in all trials except the last (October 1991), where mature fish were physically obvious. In this case, 9 mature males and 10 mature females were selected deliberately to ensure an adequate sample of mature individuals of both sexes for comparison with immature fish. Blood sampling and steroid hormone analysis
Blood samples were taken from every yearling tested from June 1991 onwards (July, August, October); the fish was sacrificed in anaesthetic (as described above), the caudal peduncle severed, and blood collected in heparinized tubes. The blood samples were kept on ice (ca. 10-15 min) until they were centrifuged, after which the plasma was collected and stored at -30°C for future hormone assay.
Plasma levels of testosterone (T) and 1 1 -ketotestosterone (1 1 -KT) were measured by radioimmunoassay, as described by Dye et al. (1986), following a heat-extraction step advocated by Scott et al. (1982). Plasma samples were initially diluted with assay buffer and incubated at 70°C for I h. Following centrifugation (15 min at 2800 rpm) the supernatant was collected and stored at -30°C until assayed.
Standards for T (Sigma Chemical Co., St. Louis) and 11-KT (Syndel Laboratories Ltd., Vancouver, B.C.) were initially dissolved in ethanol at 1 mg/mL, then serially diluted to 10 ng/mL. The working standards were prepared by further 2-fold serial dilutions from 5000 to 39 pg/mL. Tritiated T and 1 1 -KT were obtained from Amersham Canada Ltd. (Oakville, Ontario). Antibodies to T were purchased from Miles Laboratories Ltd., Rexdale, Ontario, while those to 1 1 -KT were a gift from Dr. T.G. Owen of Helix Biotech Ltd., Richmond, B.C.
For both steroid assays, duplicate 10 X 75 mm borosilicate tubes contained 200 pL of standard or sample, 200 pL of 3~-steroid (4000 and 2000 cpm for T and 1 1-KT, respectively), and 200 pL of antibody (1 :80 and 1 :4000 for T and 1 1 -KT, respectively). The tubes were vortexed and incubated overnight at room temperature. The following day, after the addition of 200 pL of cold charcoal solution to each tube, the tubes were incubated for 10 min at 4°C and then centrifuged at 2400 rpm at 4°C for 10 min. The supernatant was decanted into 7-mL plastic scintillation vials containing 4 mL of Scinti Verse 1 I (80-X-12, Fisher Scientific Ltd.). After mixing, the samples were counted in a LKB Wallac 1214 Rackbeta liquid scintillation counter (Wallac Oy, Turku, Finland) for 5 min each.
The detection limit for T and 1 1 -KT was in the order of 0.5 ng/mL.
Results
Growth and maturation The sex ratio of yearling kokanee sampled from February to
August did not differ significantly from 50:50 (goodness-of-fit test, p > 0.5; malesfemales, 46.9:53.1; the October sample was excluded because of the deliberate, nonrandom selection of fish for sex and maturity). Gonad maturation, indicated by increased gonad size, was first evident in late March. Overall, 34.7% (34 of 98) of the yearling males and 16.2% (18 of 11 1) of the yearling females sampled between March and August were maturing. The difference between the sexes in the proportion
M maturing males T
c---. maturing females
T
@+ +B immature males I E+ + immature females
March May July October 10 _L---- I I I
50 100 150 200 250 300
Day of Year FIG. 1. Mean fork length (+SE) of maturing and nonmaturing male
and female yearling kokanee. Gonad development in maturing fish was first noted on March 28 (day 87), 1991, and full maturity was reached by October 4 (day 277), 199 1.
of individuals maturing was significant (x2 contingency test, p < 0.05).
In the three-way ANOVA model used to examine the effects of sex, maturity state, and sampling date on size (fork length and mass) there was no significant effect of sex, but there were large effects due to date (p < 0.001) and maturity (p < 0.001). In addition, there was a significant interaction between maturity state and date (p < 0.005), reflecting the fact that the difference in size between maturing and nonmaturing individuals increased rapidly in the spring and early summer but remained relatively constant thereafter (Fig. I). Specific growth rates (G) for fork length (FL) and mass reflected this pattern (G = 100(ln S2 - In s,)/(T, - T,), where S is length or mass at a given time and T is time in days). The difference in growth rate between maturing and nonmatur- ing fish was greatest during April (maturing:nonmaturing, 0.49: -20.1 1 FL and 1.9 1 : -0.10 mass; note that sample sizes were relatively small and this accounts for some of the fluc- tuation in growth estimates, and even for negative estimates, over time) and next greatest during May (0.59:0.40 FL and 1.63:1.03 mass). Growth rates were then similar through to mid-August (0.29:0.24 FL and 0.79:0.77 mass). Only during the last 50 days of sampling did maturing fish grow more slowly than nonmaturing fish (0.1 1:0.26 FL and 0.50:0.90 mass). Within sample periods, maturing fish were significantly larger ( p < 0.05) than nonmaturing fish in all but the first sampling period (March) in which maturation was noted (t tests on fish pooled by sex and maturity state).
We compared the patterns of growth of maturing and nonmaturing males and females as a surrogate measure of condition factor (increment of length per increment of mass). We used data from all but the last sample period, the time of full maturity. These data were excluded because the plasma sodium values for mature fish were beyond the data range used to generate the mass adjustment function of Blackburn and Clarke (1987). ANCOVA of ln(mass) on ln(FL) with maturity state as a covariate provides a more rigorous comparison of growth partitioning than does conventional
Can
. J. Z
ool.
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
Fish
erie
s an
d O
cean
s on
08/
08/1
3Fo
r pe
rson
al u
se o
nly.
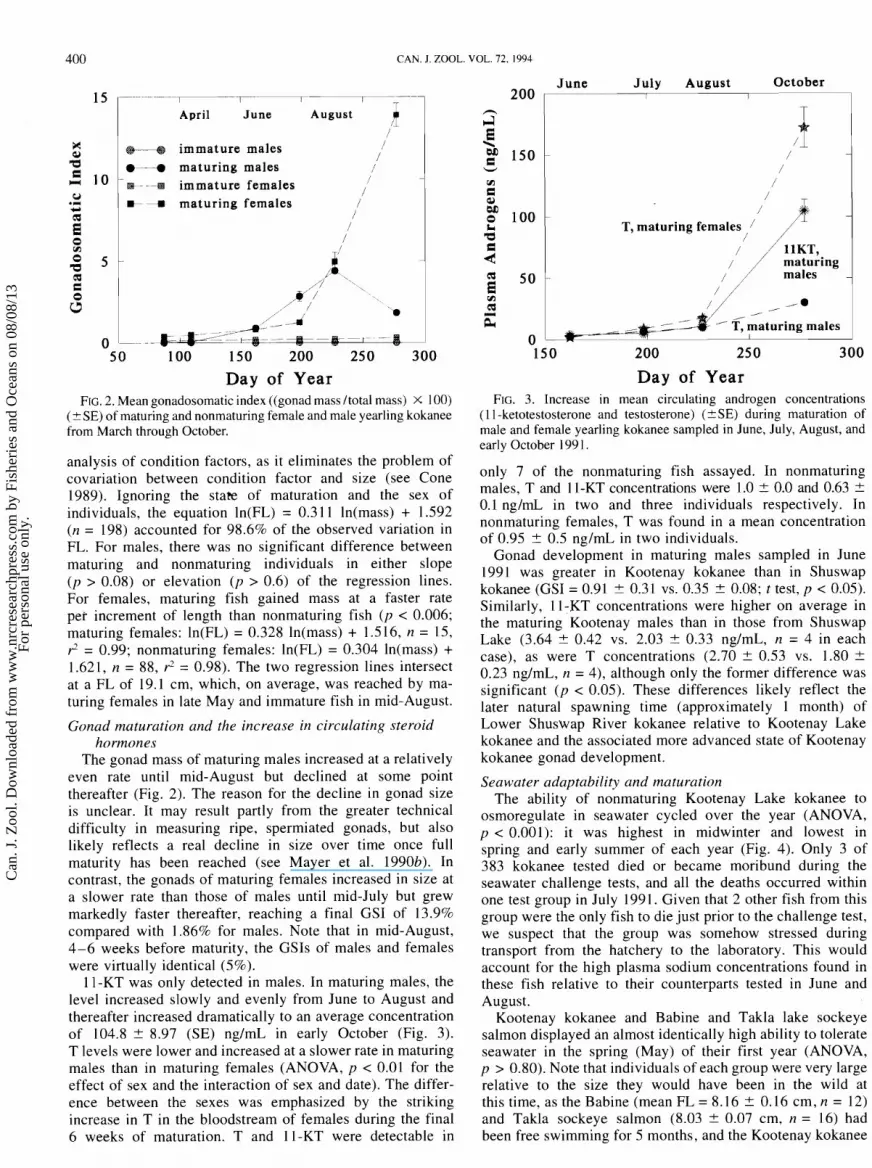
400 CAN. J. ZOOL. VOL. 72, 1994
June July August October 15 , -..-..--.--7-----i---v -----T---l 200 1 1 I
April June August /
immature males / /
a ---a maturing males / m - - -w immature females /
/ E- maturing females /
0 . - .. -
50 100 150 200 250 300
Day of Year FIG. 2. Mean gonadosomatic index ((gonad mass /total mass) X 100)
(? SE) of maturing and nonmaturing female and male yearling kokanee from March through October.
analysis of condition factors, as it eliminates the problem of covariation between condition factor and size (see Cone 1989). Ignoring the state of maturation and the sex of individuals, the equation ln(FL) = 0.31 1 ln(mass) + 1.592 (n = 198) accounted for 98.6% of the observed variation in FL. For males, there was no significant difference between maturing and nonmaturing individuals in either slope (p > 0.08) or elevation (p > 0.6) of the regression lines. For females, maturing fish gained mass at a faster rate per increment of length than nonmaturing fish (p < 0.006; maturing females: ln(FL) = 0.328 ln(mass) + 1.5 16, n = 15, r2 = 0.99; nonmaturing females: ln(FL) = 0.304 ln(mass) + 1.621, n = 88, r2 = 0.98). The two regression lines intersect at a FL of 19.1 cm, which, on average, was reached by ma- turing females in late May and immature fish in mid-August.
Gonad maturation and the increase in circulating steroid hormones
The gonad mass of maturing males increased at a relatively even rate until mid-August but declined at some point thereafter (Fig. 2). The reason for the decline in gonad size is unclear. It may result partly from the greater technical difficulty in measuring ripe, spermiated gonads, but also likely reflects a real decline in size over time once full maturity has been reached (see Mayer et al. 1990b). In contrast, the gonads of maturing females increased in size at a slower rate than those of males until mid-July but grew markedly faster thereafter, reaching a final GSI of 13.9% compared with 1.86% for males. Note that in mid-August, 4-6 weeks before maturity, the GSIs of males and females were virtually identical (5%).
11-KT was only detected in males. In maturing males, the level increased slowly and evenly from June to August and thereafter increased dramatically to an average concentration of 104.8 + 8.97 (SE) ng/mL in early October (Fig. 3). T levels were lower and increased at a slower rate in maturing males than in maturing females (ANOVA, p < 0.01 for the effect of sex and the interaction of sex and date). The differ- ence between the sexes was emphasized by the striking increase in T in the bloodstream of females during the final 6 weeks of maturation. T and 11-KT were detectable in
5 blD 0 100 1 LC T, maturing females a 1 c 4
50 + - / z i n g 1
E V1 Q ' ,a iZ /
' ' / - / - ' T, maturing males
0 150 200 250 300
Day of Year FIG. 3. Increase in mean circulating androgen concentrations
(1 1 -ketotestosterone and testosterone) (2SE) during maturation of male and female yearling kokanee sampled in June, July, August, and early October 199 1.
only 7 of the nonmaturing fish assayed. In nonmaturing males, T and 11-KT concentrations were 1.0 + 0.0 and 0.63 + 0.1 ng/rnL in two and three individuals respectively. In nonmaturing females, T was found in a mean concentration of 0.95 + 0.5 ng/mL in two individuals.
Gonad development in maturing males sampled in June 1991 was greater in Kootenay kokanee than in Shuswap kokanee (GSI = 0.91 + 0.31 vs. 0.35 + 0.08; t test, p < 0.05). Similarly, 11-KT concentrations were higher on average in the maturing Kootenay males than in those from Shuswap Lake (3.64 + 0.42 vs. 2.03 + 0.33 ng/mL, n = 4 in each case), as were T concentrations (2.70 + 0.53 vs. 1.80 + 0.23 ng/mL, n = 4), although only the former difference was significant ( p < 0.05). These differences likely reflect the later natural spawning time (approximately 1 month) of Lower Shuswap River kokanee relative to Kootenay Lake kokanee and the associated more advanced state of Kootenay kokanee gonad development.
Seawater adaptability and maturation The ability of nonmaturing Kootenay Lake kokanee to
osmoregulate in seawater cycled over the year (ANOVA, p < 0.001): it was highest in midwinter and lowest in spring and early summer of each year (Fig. 4). Only 3 of 383 kokanee tested died or became moribund during the seawater challenge tests, and all the deaths occurred within one test group in July 1991. Given that 2 other fish from this group were the only fish to die just prior to the challenge test, we suspect that the group was somehow stressed during transport from the hatchery to the laboratory. This would account for the high plasma sodium concentrations found in these fish relative to their counterparts tested in June and August.
Kootenay kokanee and Babine and Takla lake sockeye salmon displayed an almost identically high ability to tolerate seawater in the spring (May) of their first year (ANOVA, p > 0.80). Note that individuals of each group were very large relative to the size they would have been in the wild at this time, as the Babine (mean FL = 8.16 + 0.16 cm, n = 12) and Takla sockeye salmon (8.03 + 0.07 cm, n = 16) had been free swimming for 5 months, and the Kootenay kokanee
Can
. J. Z
ool.
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
Fish
erie
s an
d O
cean
s on
08/
08/1
3Fo
r pe
rson
al u
se o
nly.
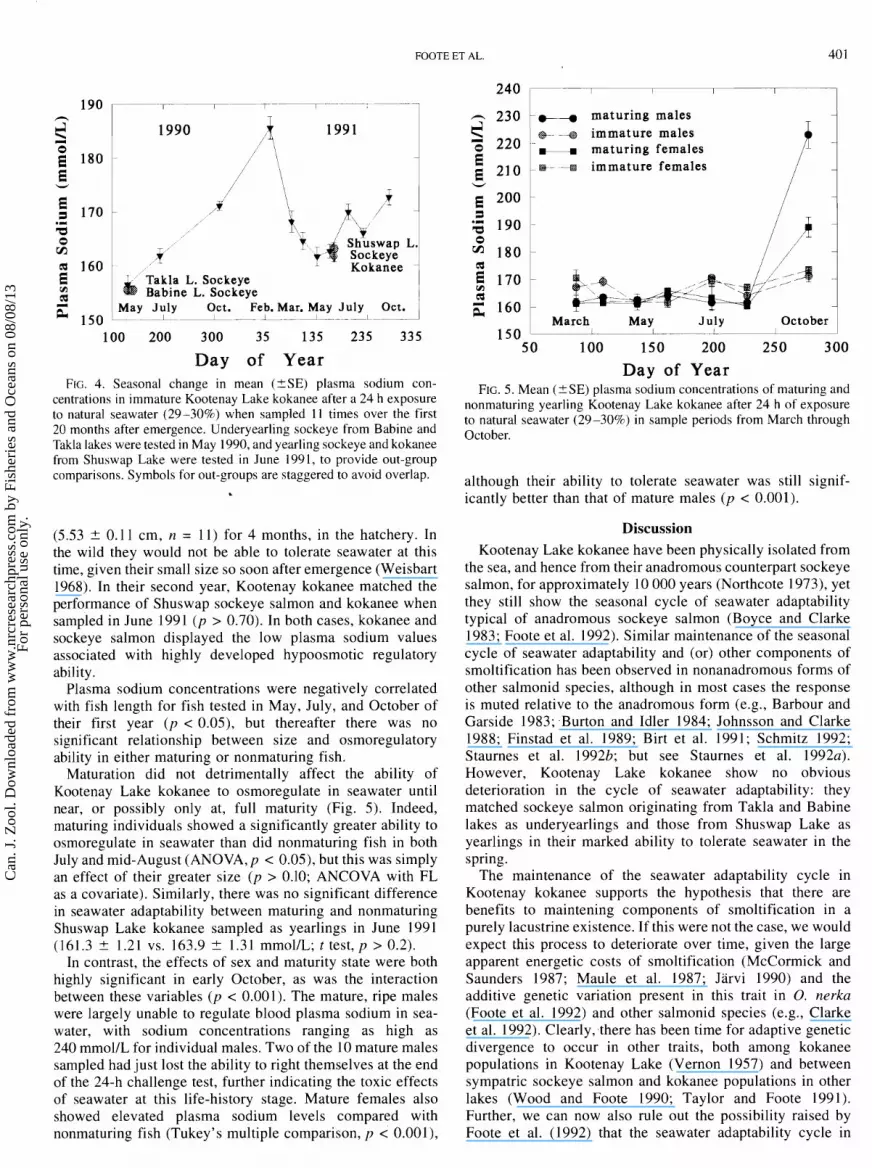
FOOTE ET AL. 40 1
-r.- -I-
1 9 0 1 9 9 0
1991 0 maturing females E 180 E E- -+ immature females w
/ ,$
/
%
b Takla L. Sockeye Babine L. Sockeye
May July Oct. Feb. Mar. May July Oct. 1 L- L 2 - - A .--I 1
100 200 300 35 135 235 335
Day of Year FIG. 4. Seasonal change in mean (+SE) plasma sodium con-
centrations in immature Kootenay Lake kokanee after a 24 h exposure to natural seawater (29-30%) when sampled 11 times over the first 20 months after emergence. Underyearling sockeye from Babine and Takla lakes were tested in May 1990, and yearling sockeye and kokanee from Shuswap Lake were tested in June 1991, to provide out-group comparisons. Symbols for out-groups are staggered to avoid overlap.
(5.53 -+ 0.1 1 cm, n = 11) for 4 months, in the hatchery. In the wild they would not be able to tolerate seawater at this time, given their small size so soon after emergence (Weisbart 1968). In their second year, Kootenay kokanee matched the performance of Shuswap sockeye salmon and kokanee when sampled in June 1991 (p > 0.70). In both cases, kokanee and sockeye salmon displayed the low plasma sodium values associated with highly developed hypoosmotic regulatory ability.
Plasma sodium concentrations were negatively correlated with fish length for fish tested in May, July, and October of their first year (p < 0.05), but thereafter there was no significant relationship between size and osmoregulatory ability in either maturing or nonmaturing fish.
Maturation did not detrimentally affect the ability of Kootenay Lake kokanee to osmoregulate in seawater until near, or possibly only at, full maturity (Fig. 5). Indeed, maturing individuals showed a significantly greater ability to osmoregulate in seawater than did nonmaturing fish in both July and mid-August (ANOVA, p < 0.05), but this was simply an effect of their greater size (p > 0.10; ANCOVA with FL as a covariate). Similarly, there was no significant difference in seawater adaptability between maturing and nonmaturing Shuswap Lake kokanee sampled as yearlings in June 1991 (161.3 -+ 1.21 vs. 163.9 + 1.31 mmol/L; t test, p > 0.2).
In contrast, the effects of sex and maturity state were both highly significant in early October, as was the interaction between these variables (p < 0.001). The mature, ripe males were largely unable to regulate blood plasma sodium in sea- water, with sodium concentrations ranging as high as 240 mmol/L for individual males. Two of the 10 mature males sampled had just lost the ability to right themselves at the end of the 24-h challenge test, further indicating the toxic effects of seawater at this life-history stage. Mature females also showed elevated plasma sodium levels compared with nonmaturing fish (Tukey's multiple comparison, p < 0.001),
March May July October 1 150 ' L _ _L- I 1
50 100 150 200 250 300
Day of Year FIG. 5. Mean (?SE) plasma sodium concentrations of maturing and
nonmaturing yearling Kootenay Lake kokanee after 24 h of exposure to natural seawater (29-30%) in sample periods from March through October.
although their ability to tolerate seawater was still signif- icantly better than that of mature males (p < 0.001).
Discussion Kootenay Lake kokanee have been physically isolated from
the sea, and hence from their anadromous counterpart sockeye salmon, for approximately 10 000 years (Northcote 1973), yet they still show the seasonal cycle of seawater adaptability typical of anadromous sockeye salmon (Boyce and Clarke 1983; Foote et al. 1992). Similar maintenance of the seasonal cycle of seawater adaptability and (or) other components of smoltification has been observed in nonanadromous forms of other salmonid species, although in most cases the response is muted relative to the anadromous form (e.g., Barbour and Garside 1983; .Burton and Idler 1984; Johnsson and Clarke 1988; Finstad et al. 1989; Birt et al. 1991; Schmitz 1992; Staurnes et al. 1992b; but see Staurnes et al. 1992~) . However, Kootenay Lake kokanee show no obvious deterioration in the cycle of seawater adaptability: they matched sockeye salmon originating from Takla and Babine lakes as underyearlings and those from Shuswap Lake as yearlings in their marked ability to tolerate seawater in the spring.
The maintenance of the seawater adaptability cycle in Kootenay kokanee supports the hypothesis that there are benefits to maintening components of smoltification in a purely lacustrine existence. If this were not the case, we would expect this process to deteriorate over time, given the large apparent energetic costs of smoltification (McCormick and Saunders 1987; Maule et al. 1987; Jarvi 1990) and the additive genetic variation present in this trait in 0. nerka (Foote et al. 1992) and other salmonid species (e.g., Clarke et al. 1992). Clearly, there has been time for adaptive genetic divergence to occur in other traits, both among kokanee populations in Kootenay Lake (Vernon 1957) and between sympatric sockeye salmon and kokanee populations in other lakes (Wood and Foote 1990; Taylor and Foote 199 1). Further, we can now also rule out the possibility raised by Foote et al. (1992) that the seawater adaptability cycle in
Can
. J. Z
ool.
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
Fish
erie
s an
d O
cean
s on
08/
08/1
3Fo
r pe
rson
al u
se o
nly.

402 CAN. J. ZOOL. VOL. 72, 1994
kokanee is maintained through continued gene flow, or recent divergence, from sockeye salmon as neither is possible in isolated Kootenay Lake. Rather, with slight variation (see Foote et al. 1992), it appears that the cycle is remarkably similar in 0. nerka populations between forms and across drainages, whether the forms occur in sympatry or allopatry.
The smoltification process likely facilitated the transition from anadromous sockeye salmon to kokanee following the former's invasion of new lake habitats after the retreat of the last Pleistocene glaciation (10 000 - 15 000 years before present in British Columbia), in the manner Berg (1985) described for nonanadromous lacustrine Atlantic salmon. The ease of this transition from one form to the other is highlighted by the successful introduction of sockeye salmon into a landlocked lake in Japan, where they persisted as kokanee, and by their subsequent movement and reconversion to anadromous sockeye salmon more than 15 generations later (Kaeriyama et al. 1992; see also Foerster 1947; Scott 1984).
Potential benefits of maintening components of smolti- fication in a freshwater existence were reviewed by Hoar (1976, 1988), Berg (1985), and Foote et al. (1992). These include the positive effects of increased growth hormone (see also McCormick et al. 1991) and thyroxin production, increased deposition of purines in the scales (silvering), and finally the benefits o'f reduced aggression on schooling behaviour in open waters. It is probable that these traits benefited sockeye salmon that remained in lakes, as the circumstantial evidence suggests that components of smoltification continue to benefit long-standing kokanee populations today (see also Leatherland et al. 1974).
If components of smoltification are beneficial to an open freshwater existence, why does the ability of kokanee (and sockeye salmon) to tolerate seawater deteriorate in the fall through to midwinter before increasing again? We suspect that the answer lies in the limited energy available in the natural environment in the fall and winter to maintain characters associated with smoltification (and growth in general). The fish are likely preprogrammed to limit energy input to these processes during seasonal times of reduced primary production, simply in order to maintain stasis (see Johnsson et al. 1993). Our results indicate that photoperiod is a primary cue entraining such a process, as temperature fluctuated only minimally over the year in the hatchery (20.5"C).
The onset of maturity is thought to inhibit the smoltification process in salmonids in both males (e.g., McCormick and Naiman 1985; Langdon and Thorpe 1985; Thorpe 1986,1987; Lundqvist et al. 1990; Foote et al. 1991) and females (Sutterlin and MacLean 1984). Exactly how this inhibition occurs remains unknown, but the release of androgens associated with maturation has been clearly implicated. For example, androgen adminstration has been shown to inhibit seawater adaptability in masu salmon, 0. masou (Ikuta et al. 1987), amago salmon, 0. rhodurus (Miwa and Inui 1985), and Atlantic salmon (Lundqvist et al. 1989). Similarly, castration of mature parr has been shown to stop the inhibition (Aida et al. 1984), although this is not always the case (Lundqvist et al. 1989, 1990).
In this study, the commencement of gonad maturation in yearling kokanee was first detected in March, 6 months prior to their natural spawning period. The progression of maturity was associated with increased concentrations 'of plasma
androgens in males and females. 11-KT and T were the prominent androgens in males and females, respectively, as is typical of salmonids (Scott et al. 1980a, 1980b; Dye et al. 1986; Mayer et al. 1990b). Androgen levels in kokanee raised in the hatchery were comparable to those measured in wild kokanee from Kootenay Lake at maturity (Liley et al. 1993), but somewhat lower than those measured in sockeye salmon during their final migration to the spawning grounds (Truscott et al. 1986). However, we took only a single sample during the final maturation period, and absolute and relative steroid levels can increase or decrease during this time (e.g., Truscott et al. 1986; Liley et al. 1993).
The process of maturation was not associated with any decrease in seawater adaptability in either males or females during the typical smolting period of 0. nerka from April through June (see Burgner, 1987). Indeed, maturing males and females showed plasma sodium concentrations similar to or lower than nonmaturing individuals and typical of anad- romous smolts during this period (<I65 mmol/L; see Clarke 1982; Lundqvist et al. 1990; Foote et al. 1992), even though they showed significant somatic and gonadal growth and elevated androgen levels. Thereafter, maturing fish maintained their strong ability to osmoregulate in seawater until at least mid-August. However, by early October, mature males and females showed significant loss of hypoosmoregulatory ability in comparison with immature fish, with seawater all but lethal to males after 24 h exposure.
The lack of an inhibitory effect of maturation on seawater adaptability during the smoltification period in kokanee is in marked contrast to that observed in other salmonid species (see references above and Thorpe 1987 for review). Further, this appears to be a species-specific trait in 0. nerka. Foote et al. (1992) found similar results in Shuswap Lake kokanee, as we did again here, and in hybrids between sockeye salmon and kokanee. Combined, these results do not support the hypothesis proposed by Thorpe (1986, 1987) that a devel- opmental conflict between maturation and smoltification was responsible for the origination of nonanadromous 0. nerka, as no apparent physiological conflict exists in this species during the time of smoltification. However, we cannot rule out the possibility that maturation affects smolt migratory behaviour as in other salmonids (e.g., Dellefors and Faremo 1988). To date, we have twice been unsuccessful in demon- strating any consistent seasonal trend or any overall difference between sockeye salmon and kokanee in migratory behaviour (C.J. Foote and C.C. Wood, unpublished data).
The absence of a developmental conflict between maturation and smoltification in 0. nerka does not negate the likely role of such a process in the evolution of nonanadromy in other salmonids. Rather, it prompts us to ask, when and where such a conflict would develop. Given that kokanee living in lakes show many of the traits associated with a marine existence, perhaps it is not surprising that maturation does not negatively affect their ability to osmoregulate during the time kokanee are occupying open waters. We would not expect to find an inhibitory effect of maturation on the ability to osmoregulate in anadromous salmonids until at or just before entrance to fresh water (e.g., Boykov et al. 1988; Hirano et al. 1990), even though the process of maturation would have commenced many months before.
The above argument suggests that there are physiological differences associated with maturation in an open water
Can
. J. Z
ool.
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
Fish
erie
s an
d O
cean
s on
08/
08/1
3Fo
r pe
rson
al u
se o
nly.

FOOTE ET AL. 403
marine/lacustrine versus a riverine existence. Precocious demonstrated ease of transition between the two life-history males in riverine species typically show highly developed parr marks (e.g., Dellefors and Faremo 1988; Taylor 1989; Foote et al. 1991) and relatively high condition factors relative to immature fish (e.g., Rowe and Thorpe 1990; Berglund et al. 1992). Both traits are directly affected by the addition of androgens (Miwa and Inui 1985; Lundqvist et al. 1989; Berglund et al. 1992). In contrast, maturing kokanee remained silver until shortly before full maturity (all were silver in mid-August, just 4-6 weeks prior to attaining full maturity) and did not show any increase in condition factor during maturation. Rather, maturing males maintained the same relationship between increment of length versus increment of mass as did nonmaturing fish, and thus maintained the same fusiform body shape. Hence, while precocious riverine forms maintained traits adaptive for their environment (relatively deep bodies and parr marks) and in contrast with those necessary for smoltification (relatively slender bodies and no parr marks), maturing kokanee maintained traits adaptive for an open water environment (see Swain and Holtby 1989), traits that are in general accord with smoltification and seaward migration.
We know that the lack of morphological differentiation in maturing kokanee during the time of species-typical smoltification is not the reshlt of the absence of increasing levels of circulating androgens. Therefore, we suggest that the absence of differentiation is more likely associated with a
types.
Acknowledgements W e are grateful to G. Johnston, I. Baker, and R. Traber
for their excellent care of the fish throughout this study. W e thank D. Rutherford for technical help, N.R. Liley and H. Olskn for their help in making the crosses, and H. Andrusack and R.J. Hammond of the B.C. Ministry of Environment for permission to work at Meadow Creek. The comments of reviewers improved the manuscript. Much of this work was done while C.J.F. was a postdoctorate fellow at the Department of Biology, McGill University, and the support of members of the Department is gratefully acknowledged.
Aida, K., Kato, T., and Awaji, M. 1984. Effects of castration on the smoltification of precocious male masu salmon Oncorhynchus rnasou. Bull. Jpn. Soc. Sci. Fish. 50: 565 -57 1.
Barbour, S. E., and Garside, E. T. 1983. Some physiologic distinctions between freshwater and diadromous forms of the Atlantic salmon, Salrno salar L. Can. J. Zool. 61: 1 165 -1 170.
Berg, O.K. 1985. The formation of nonanadromous populations of Atlantic salmon, Salrno salur L., in Europe. J. Fish Biol. 27: 805 -8 15.
Berglund, I., Mayer, I., and Borg, B. 1992. Effects of sexual maturation, castration, and androgen implants on growth in one- and two-year-old Darr in a Baltic Atlantic salmon (Salrno salar L.)
reduced binding capacity of the skin and possibly other target stock. J. Fish Bioj. 40: 281-292. organs compared with that observed in maturing riverine Birt, T.P., Green, J.M., and Davidson, W.S. 1991. Contrasts in salmonids. For example, Pottinger (1988) showed that the development and smolting of genetically distinct sympatric testosterone binding capacity of skin nuclear extract was anadromous and nonanadromous Atlantic salmon, Sulrno salar.
- A - consistently higher in mature versus immature brown trout, Salrno trutta. The increased skin binding capacity appears to be directly linked with increased dermal and epidermal thickening (Pottinger and Pickering 1985). Given that we observed no external change in maturing fish until late August, just 3-4 weeks before full maturity was reached, it is possible that the maturing kokanee simply delay the onset of increased binding capacity relative to riverine salmonids. Further, given that the marked decrease in seawater adaptability in mature fish appears to coincide directly with morphological change associated with maturity, it is tempting to speculate that this delayed change, which occurs in association with the movement of kokanee into the rivers to spawn (Liley et al. 1993), is the proximate reason for both the loss of seawater adaptability and the difference between kokanee and other salmonids in the onset of this inhibition. The greater loss of hypoosmoregulatory ability in mature males versus mature females adds some support to this argument, as the males undergo the greater physical change, in both skin thickness and body depth (C.J. Foote, G.S. Brown, and C.C. Wood, unpublished data). In addition, there is growing evidence that it is the structural effects of the androgens and not the androgens themselves that conflict with smoltification (Lundqvist et al. 1990; Mayer et al. 1990a)
Berg (1985) argued that the presence of lakes greatly facilitated the origination of landlocked populations of Atlantic salmon because many of the characters necessary for seaward migration were also likely beneficial to a nonanadromous existence. Our results suggest that these arguments also apply to kokanee, and may explain their widespread distribution (see Nelson 1968) and the
Can. J. Zool. 69: 2075 -2084. Blackburn, J., and Clarke, W.C. 1987. Revised procedure for the
24 hour seawater challenge test to measure seawater adaptability of juvenile salmonids. Can. Tech. Rep. Fish. Aquat. Sci. No. 15 15.
Boyce, N.P., and Clarke, W.C. 1983. Eubothriurn salvelini (Cestoda: Pseudophyllidea) impairs seawater adaption of migrant sockeye salmon yearlings (Oncorhynchus nerka) from Babine Lake, British Columbia. Can: J. Fish. Aquat. Sci. 40: 821-824.
Boykov, P.Ya., ~ a k s i m o v i c h ; ~ . ~ . , Natochin, Yu.V., Polyakov, V.N., and Plyusnin, V.V. 1988. Osmoregulation and electrolytes in the blood serum of spawning pink salmon, Oncorhynchus gorbuscha, during migration from the sea into the river and when transferred back into sea water. J. Ichthyol. (Engl. Transl. Vopr. Ikhtiol.), 5: 1-6.
Burgner , R. L. 1987. Factors influencing age and growth of juvenile sockeye salmon (Oncorhynchus nerka) in lakes. Can. Spec. Publ. Fish. Aquat. Sci. No. 96. pp. 129-142.
Burton, M.P., and Idler, D.R. 1984. Can Newfoundland landlocked salmon, Salrno salar L., adapt to sea water? J. Fish Biol. 24: 59-64.
Clarke, W.C. 1982. Evaluation of the seawater challenge test as an index of marine survival. Aquaculture, 28: 177-183.
Clarke, W.C., Withler, R.E., and Shelbourn, J.E. 1992. Genetic control of juvenile life history pattern in chinook salmon (Oncorhynchus tshawytscha). Can. J. Fish. Aquat. Sci. 49: 2300-2306.
Cone, R.S. 1989. The need to reconsider the use of condition indices in fishery science. Trans. Am. Fish. Soc. 118: 5 10-5 14.
Dellefors, C., and Faremo, U. 1988. Early sexual maturation in males of wild sea trout, Salrno trutta L., inhibits smoltification. J. Fish Biol. 33: 741-749.
Dye, H.M., Sumpter, J.P., Fagerlund, U.H.M., and Donaldson, E.M. 1986. Changes in reproductive parameters during spawning migration of pink salmon, Oncorhynchus gorbuscha (Walbaum). J. Fish Biol. 29: 167-176.
Can
. J. Z
ool.
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
Fish
erie
s an
d O
cean
s on
08/
08/1
3Fo
r pe
rson
al u
se o
nly.

404 CAN. J. ZOOL. VOL. 72, 1994
Finstad, B., Nilssen, K.J., and Arnesen, A.M. 1989. Seasonal changes in sea-water tolerance of Arctic charr (Salvelinus alpinus). J. Comp. Physiol. B, 159: 371-378.
Foerster, R.E. 1947. Experiment to develop sea-run from land-locked sockeye salmon (Oncorhynchus nerka kennerlyi). J. Fish. Res. Board Can. 7: 83-93.
Foote, C.J., and Larkin, P.A. 1988. The role of male choice in the assortative mating of sockeye salmon and kokanee, the anadromous and non-anadromous forms of Oncorhynchus nerka. Behaviour, 106: 43-62.
Foote, C.J., Wood, C.C., and Withler, R.W. 1989. Biochemical genetic comparison of sockeye salmon and kokanee, the anadromous and nonanadromous forms of Oncorhynchus nerka. Can. J. Fish. Aquat. Sci. 46: 149-158.
Foote, C.J., Clarke, W.C., and Blackburn, J. 1991. Inhibition of smolting in precocious male chinook salmon, Oncorhynchus tshawytscha. Can J. Zool. 69: 1848 -1 852.
Foote, C.J., Wood, C.C., Clarke, W.C., and Blackburn, J. 1992. Circannual cycle of seawater adaptability in Oncorhynchus nerka: genetic differences between sympatric sockeye salmon and kokanee. Can. J. Fish. Aquat. Sci. 49: 99-109.
Hanson, A. J., and Smith, H. D. 1967. Mate selection in a population of sockeye salmon (Oncorhynchus nerka) of mixed age groups. J. Fish. Res. Board Can. 24: 1955-1977.
Hindar, K., and Jonsson, B. 1982. Habitat and food segregation of dwarf and normal Arctic charr (Salvelinus alpinus) from Vangsvatnet Lake, western Norway. Can. J. Fish. Aquat. Sci. 39: 1030-1045.
Hirano, T., Ogasawara, T., Hasegawa, S., Iwata, M., and Nagahama, Y. 1990. Changes in plasma hormone levels during loss of hypoosmoregulatory capacity in mature chum salmon (Oncorhynchus keta) kept in freshwater. Gen. Comp. Endocrinol. 78: 254-262.
Hoar, W.S. 1976. Smolt transformation: evolution, behaviour and physiology. J. Fish. Res. Board Can. 33: 1233-1252.
Hoar, W.S. 1988. The physiology of smolting salmonids. In Fish physiology. Volume 11B. Edited by W.S. Hoar and D.J. Randall. Academic Press, New York.
Hutchings, J.A., and Myers, R. A. 1988. Mating success of alternate maturation phenotypes in male Atlantic salmon, Salmo salar. Oecologia, 75: 169-1 74.
Ikuta, K., Aida, K., Okumoto, N., and Hanyu, I. 1987. Effects of sex steroids on the smoltification of masu salmon, Oncorhynchus masou. Gen. Comp. Endocrinol. 65: 99-1 10.
Jarvi, T. 1990. Cumulative acute physiological stress in Atlantic salmon smolts: the effect of osmotic imbalance and the presence of predators. Aquaculture, 89: 337-350.
Johnsson, J., and Clarke, W.C. 1988. Development of seawater adaption in juvenile steelhead trout (Salmo gairdneri) and domesticated rainbow trout (Salmo gairdneri)-effects of size, temperature and photoperiod. Aquaculture, 71: 247-263.
Johnsson, J.I., Clarke, W.C. and Withler, R.E. 1993. Hybridization with domesticated rainbow trout (Oncorhynchus mykiss) reduces seasonal variation in growth of steelhead trout (0 . mykiss). Can. J. Fish. Aquat. Sci. 50: 480-487.
Jones, J.W., and King, G.M. 1952. The spawning of male salmon parr (Salmo salar Linn. Juv.). Proc. Zool. Soc. Lond. 122: 61 5-61 9.
Jonsson, B. 1985. Life history variation of freshwater resident and searun migrant brown trout in Norway. Trans. Am. Fish. Soc. 114: 182-194.
Jordan, W.C., and Youngson, A.F. 1992. The use of genetic marking to access the reproductive success of mature male Atlantic salmon parr (Salmo salur L.) under natural spawning conditions. J. Fish Biol. 41: 61 3-61 8.
Kaeriyama, M., Urawa, S., and Susuki, T. 1992. Anadromous sockeye salmon (Oncorhynchus nerka) derived from nonanadromous kokanees: life history in Lake Toro. Sci. Rep. Hokkaido Salmon Hatchery, 46: 157-174.
Krogius, F. V. 198 1. The role of resident fish in the reproduction of anadromous sockeye salmon, Oncorhynchus nerka. J. Ichthyol. (Engl. Transl. Vopr. Ikhtiol.), 21: 14-21.
Langdon, J.S., and Thorpe, J.E. 1985. The ontogeny of smoltification: developmental patterns of gill Na+/K+-ATPase, SDH, and chloride cells in juvenile Atlantic salmon, Salmo salar L. Aquaculture, 45: 83-95.
Leatherland, J.F., McKeown, B.A., and John, T.M. 1974. Circadian rhythm of plasma prolactin, growth hormone, glucose and free fatty acid in juvenile kokanee salmon, Oncorhynchus nerka. Comp. Biochem. Physiol. A, 47: 821-828.
Liley, N.R., Olsen, H.K., Foote, C.J., and Van Der Kraak, G. 1993. . Endocrine changes associated with spawning behaviour in male kokanee salmon (Oncorhynchus nerka) and the effects of anosmia. Horm. Behav. 27: 470-487.
Lundqvist, H., Borg, B., and Berglund, I. 1989. Androgens impair seawater adaptability in smolting Baltic salmon (Salmo salar). Can. J. Zool. 67: 1733-1 736.
Lundqvist, H., Berglund, I., Mayer, I., and Borg, B. 1990. Seawater adaptability in Baltic salmon, Salmo salar, immature smolt and mature male parr: lack of effect of springtime castration. Can. J. Zool. 68: 2 18 1-2 1 84.
Maekawa, K. 1977. Studies on the variability of the landlocked miyabe charr, Salvelinus malma miyabei Oshima. 1. Development and early life history. Jpn. J. Ecol. 27: 91-102.
Maule, A.G., Schreck, C.B., and Kaattari, S. L. 1987. Changes in the immune system of coho salmon (Oncorhynchus kisutch) during the parr-to-smolt transformation and after the implantation of cortisol. Can. J. Fish. Aquat. Sci. 44: 161-166.
Mayer, I., Berglund, I., Rydevik, M., Borg, B., and Scultz, R. 1990a. Plasma levels of five androgens and 17a-hydroxy- 20P-dihydroprogesterone in immature and mature male Baltic salmon (Salmo salar) parr, and the effects of castration and androgen replacement in mature parr. Can. J. Zool. 68: 263-267.
Mayer, I., Lundqvist, H., Berglund, I., Schmitz, M., Scultz, R., and Borg, B. 1990b. Seasonal endocrine changes in Baltic salmon, Salmo salar, immature parr and mature male pan. 1. Plasma levels of five androgens, 17a-hydroxy-20P-dihydroxyprogesterone, and 17P-estradiol. Can. J. Zool. 68: 1360-1365.
McCart, P. 1970. A polymorphic population of Oncorhynchus nerka in Babine Lake, British Columbia. Ph.D. thesis, University of British Columbia, Vancouver.
McCormick, S.D., and Naiman, R.J. 1985. Hypoosmoregulation in an anadromous teleost: influence of sex and maturation. J. Exp. Zool. 234: 193-198.
McCormick, S.D., and Saunders, R.L. 1987. Preparatory physiological adaptations to marine life: osmoregulation, growth and metabolism. Am. Fish. Soc. Symp. 1: 21 1-229.
McCormick, S.D., Sakamoto, T., Hasegawa, S., and Hirano, T. 199 1. Osmoregulatory actions of insulin-like growth factor-1 in rainbow trout (Oncorhynchus mykiss). J. Endocrinol. 130: 87-92.
Miwa, S., and Inui, Y. 1985. Inhibitory effects of 17-methyltestosterone and oestradiol-17 on smoltification of sterilized amago salmon Oncorhynchus rhodurus. Aquaculture, 45: 383.
Nelson, J.S. 1968. Distribution and nomenclature of North American kokanee, Oncorhynchus nerka. J. Fish. Res. Board Can. 25: 409-414.
Northcote, T.G. 1973. Some impacts of man on Kootenay Lake and its salmonids. Great Lakes Fishery Commission Tech. Rep. No. 25.
Pottinger, T.G. 1988. Seasonal variation in specific plasma- and target-tissue binding of androgens, relative to plasma steroid levels, in the brown trout, Salmo trutta L. Gen. Comp. Endocrinol. 70: 334-344.
Pottinger, T.G., and Pickering, A.D. 1985. Changes in skin structure associated with elevated androgen levels in maturing male brown trout, Salmo trutta L. J. Fish. Biol. 26: 745-753.
Ricker, W.E. 1938. "Residual" and kokanee salmon in Cultus Lake. J. Fish. Res. Board Can. 4: 192-21 8.
Ricker, W.E. 1940. On the origin of kokanee, a freshwater type of sockeye salmon. Trans. R. Soc. Can. 34: 121-135.
Can
. J. Z
ool.
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
Fish
erie
s an
d O
cean
s on
08/
08/1
3Fo
r pe
rson
al u
se o
nly.

FOOTE ET AL. 405
Rowe, D.K., and Thorpe, J.E. 1990. Differences in growth between maturing and non-maturing male Atlantic salmon, Salmo salar L., parr. J. Fish Biol. 36: 643-658.
Saunders, R.L., Henderson, E.B., and Glebe, B.D. 1982. Precocious sexual maturation and smoltification in male Atlantic salmon (Salmo salar). Aquaculture, 28: 2 1 1-229.
Schmitz, M. 1992. Annual variations in rheotactic behaviour and seawater adaptability on landlocked Arctic char (Salvelinus alpinus). Can. J. Fish. Aquat. Sci. 49: 448-452.
Scott, A.P., Bye, V.J., and Baynes, S. M. 1980a. Seasonal variations in sex steroids of female rainbow trout (Salmo gairdneri Richardson). J. Fish Biol. 17: 587-592.
Scott, A.P., Bye, V.J., Baynes, S.M., and Springate, J.C. 1980h. Seasonal variations in plasma concentrations of 1 1 -ketotestosterone and testosterone in male rainbow trout (Salmo gairdneri Richardson). J. Fish Biol. 17: 495-505.
Scott, A.P., Sheldrick, E.L., and Flint, A.P.F. 1982. Measurement of 17a,20P-dihydroxy-4-pregen-3-one in plasma of trout (Salmo gairdneri, Richardson): seasonal changes in response to salmon pituitary extract. Gen. Comp. Endocrinol. 46: 444-45 1.
Scott, D. 1984. Origin of New Zealand sockeye salmon, Onco- rhynchus nerka. J. R. Soc. N.Z. 14: 245 -249.
Skulason, S., Noakes, D.L.G., and Snorrason, S.S. 1989. Ontogeny of trophic morphology in four sympatric morphs of Arctic charr Salvelinus alpinus in Thingvallavatn, Iceland. Biol. J. Linn. Soc. 38: 28 1-20 1.
Smirnov, A.I. 1959. Differences in the biology and reproduction and the development of residual ck dwarf sockeye and anadromous sockeye (Oncorhynchus nerka) Walbaum. Nauchn. Dokl. Vyssh. Shk., Biol. Nauki No. 3. pp. 59-65. [Fish. Res. Board Can. Translation No. 266.1
Staurnes, M., Lysfjord, G., and Berg, O.K. 1992a. Parr-smolt transformation of a nonanadromous population of Atlantic salmon (Salmo salar) in Norway. Can. J. Zool. 70: 197-199.
Staurnes, M., Sigholt, T., Lysfjord, G., and Gulseth, D.A. 1992h. Difference in the seawater tolerance of anadromous and landlocked
populations of Arctic char (Salvelinus alpinus). Can. J. Fish. Aquat. Sci. 49: 443-447.
Sutterlin, A.M., and MacLean, D. 1984. Age at first maturity and the early expression of oocyte recruitment processes in two forms of Atlantic salmon (Salmo salar) and their hybrids. Can. J. Fish. Aquat. Sci. 41: 1 139-1 149.
Swain, D.P., and Holtby, L.B. 1989. Differences in morphology and behavior between juvenile coho salmon (0ncorhynchu.s kisutch) rearing in a lake and in its tributary stream. Can. J. Fish. Aquat. Sci. 46: 1406-1414.
Taylor, E.B. 1989. Precocial male maturation in laboratory reared populations of chinook salmon, Oncorhynchus tshawytscha. Can. J. Zool. 67: 1665-1669.
Taylor, E.B., and Foote, C.J. 1991. Critical swimming velocities of juvenile sockeye salmon and kokanee, the anadromous and non-anadromous forms of Oncorhynchus nerka. J. Fish Biol. 38: 407- 4 1 9.
Thorpe, J.E. 1986. Age at first maturity in Atlantic salmon, Salmo salar: freshwater period influences and conflicts with smolting. Can. Spec. Publ. Fish. Aquat. Sci. No. 89. pp. 7-14.
Thorpe, J.E. 1987. Smolting versus residency: developmental conflict in salmonids. Am. Fish. Soc. Symp. 1: 244-252.
Truscott, B., Idler, D.R., So, Y.P., and Walsh, J.M. 1986. Ma- turational steroids and gonadotropin in upstream migratory sockeye salmon. Gen. Comp. Endocrinol. 62: 99-1 10.
Vernon, E.H. 1957. Morphometric comparison of three races of kokanee (Oncorhynchus nerka) within a large British Columbia lake. J. Fish. Res. Board Can. 14: 573-598.
Weisbart, M. 1968. Osmotic and ionic regulation of embryos, alevins, and fry of the five species of Pacific salmon. Can. J. Zool. 46: 385 -397.
Wood, C.C., and Foote, C.J. 1990. Genetic differences in the early development and growth of sympatric sockeye salmon and kokanee (Oncorhynchus nerka) and their hybrids. Can. J. Fish. Aquat. Sci. 47: 2250-2260.
Can
. J. Z
ool.
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
Fish
erie
s an
d O
cean
s on
08/
08/1
3Fo
r pe
rson
al u
se o
nly.