tumor-penetrating irgd peptide inhibits metastasis
TRANSCRIPT

Small Molecule Therapeutics
Tumor-Penetrating iRGD Peptide InhibitsMetastasisKazuki N. Sugahara1,2, Gary B. Braun1,3, Tatiana Hurtado de Mendoza1,VenkataRamanaKotamraju1, Randall P. French4, AndrewM. Lowy4,Tambet Teesalu1,5, andErkki Ruoslahti1,3
Abstract
Tumor-specific tissue-penetrating peptides deliver drugs intoextravascular tumor tissue by increasing tumor vascular permeabil-ity through interactionwith neuropilin (NRP). Here, we report thata prototypic tumor-penetrating peptide iRGD (amino acidsequence:CRGDKGPDC)potently inhibits spontaneousmetastasisinmice. The antimetastatic effectwasmediatedby theNRP-bindingRXXK peptide motif (CendR motif), and not by the integrin-binding RGD motif. iRGD inhibited migration of tumor cells and
caused chemorepulsion in vitro in a CendR- andNRP-1–dependentmanner. The peptide induced dramatic collapse of cellular process-es and partial cell detachment, resulting in the repellent activity.These effects were prominently displayed when the cells wereseeded on fibronectin, suggesting a role of CendR in functionalregulation of integrins. The antimetastatic activity of iRGD mayprovide a significant additional benefit when this peptide is usedfor drug delivery to tumors.MolCancer Ther; 14(1);120–8.�2014AACR.
IntroductionTumor blood vessels are structurally defective (1). They often
lack pericytes, and their basement membrane is abnormallyloose. The irregular structure leads to leakiness and predisposestumors to metastasis. The leakiness of tumor vasculature alsoresults in high interstitial fluid pressure, which prevents drugpenetration into tumor tissue (2). Poor drug penetrationreduces the potential for antitumor efficacy and results inacquired drug resistance. Factors that increase vascular perme-ability, such as VEGF, bradykinin, and nitric oxide, improvedrug distribution into tumor tissue, but can also promotemetastasis by enhancing tumor cell access to and from bloodvessels (2–4). miR105, a miRNA that disrupts vascular integrityand increases vascular permeability facilitates metastasis for-mation (5). Forced expression of miR105 in nonmetastatictumor cells leads to enhanced vascular permeability and metas-tasis, whereas inhibition of miR105 in highly metastatic tumorcells suppresses metastasis.
Neuropilin-1 (NRP-1) and neuropilin-2 (NRP-2) are some ofthe key molecules that regulate vascular permeability (6). Tissue-penetrating peptides bind toNRPs through the consensus RXXR/Kamino acid sequence, increasing vascular permeability and trans-port of molecules through tissue (7). The RXXR/K sequencemotifis not active unless it occupies a C-terminal position in a peptide,therefore referred to as the C-end Rule (CendR) motif (8). Nota-bly, many natural ligands for NRPs, such as VEGF and class 3semaphorins, carry a CendR motif, and share various biologicactivities with tissue-penetrating peptides (6, 7).
iRGD (cyclic CRGDK/RGPD/EC; the CRGDKGPDC form wasused in this study) is a prototypic tumor-specific tissue-penetrat-ing peptide, which delivers drugs deep into extravascular tumortissue (9). Intravenously injected iRGD first targets av integrinsspecifically expressed on tumor vasculature (10). iRGD is thenproteolytically processed into CRGDK/R, exposing an activeCendR motif at the C-terminus. The interaction of the CendRmotif with NRPs initiates an active bulk transport system throughthe tumor tissue, allowing drugs conjugated to iRGD, and evenfree drugs coadministered with iRGD, to extravasate and spreadwithin the tumor tissue (11). Thus, iRGDprovides a simpleway toenhance the therapeutic index of various anticancer drugs. How-ever, the effect of iRGDon thepermeability of tumor bloodvesselshas led to a hypothetical concern that iRGD may promotemetastasis through antidromic tumor cell dissemination into thecirculation (12). While our earlier studies have shown that iRGDdoes not promote seeding of nonmetastatic tumor cells (11), theeffects of iRGD on metastatic tumors remain elusive. Here, wehave examined the iRGD effects on spontaneous metastasisformation in mice bearing aggressive cancers.
Materials and MethodsPeptides, cells, tumor models, and in vivo treatment studies
Peptides were synthesized as described previously (8, 9, 11).PC-3 human prostate cancer cells were purchased from andauthenticatedbyATCC.GFP-PC-3 cellswerepreparedby infecting
1Cancer Research Center, Sanford-Burnham Medical Research Insti-tute, La Jolla, California. 2Department of Surgery, Columbia UniversityCollege of Physicians and Surgeons, New York, New York. 3Center forNanomedicine andDepartment of Cell, Molecular andDevelopmentalBiology, University of California Santa Barbara, Santa Barbara, Cali-fornia. 4Division of Surgical Oncology and Moores Cancer Center,University of California, San Diego, La Jolla, California. 5Centre ofExcellence for Translational Medicine, University of Tartu, Tartu,Estonia.
Note: Supplementary data for this article are available at Molecular CancerTherapeutics Online (http://mct.aacrjournals.org/).
Corresponding Author: Kazuki N. Sugahara, Sanford-Burnham MedicalResearch Institute, 10901 North Torrey Pines Road, La Jolla, CA 92037. Phone:858-646-3100, ext. 3517; Fax: 858-795-5353; E-mail:[email protected]
doi: 10.1158/1535-7163.MCT-14-0366
�2014 American Association for Cancer Research.
MolecularCancerTherapeutics
Mol Cancer Ther; 14(1) January 2015120
on January 18, 2015. © 2015 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst November 12, 2014; DOI: 10.1158/1535-7163.MCT-14-0366

the PC-3 cells with GFP lentiviruses (9, 13). LM-PmC cells wereprepared by stable mCherry transduction in LM-P cells, whichwere derived from liver metastasis of pancreatic ductal adenocar-cinoma (PDAC) in KrasG12D/þ;LSL-Trp53R172H/þ;Pdx-1-Cre (KPC)mice and authenticated as described earlier (14). Both cell lineswere cultured in DMEM supplemented with 10% FBS and pen-icillin/streptomycin, and used for no longer than 6months beforebeing replaced. Tumor mouse models were created by orthotopicinjections of 1million GFP-PC-3 or LM-PmC cells into nudemice2 weeks (GFP-PC-3) or 1 week (LM-PmC) before the initiation ofthe treatment study. The mice were intravenously treated everyother day with 4 mmol/kg of peptides or vehicle (PBS) alone. After21 days (GFP-PC-3) or 14 days (LM-PmC) of treatment, the micewere dissected under deep anesthesia, imaged underUV lightwithan Illumatool Bright Light System LT-9900 (Lightools Research),and perfused through the heart with PBS containing 1% bovineserum albumin (BSA) before harvesting tissues. All animal exper-imentation was performed according to procedures approved bythe Animal Research Committee at Sanford-Burnham MedicalResearch Institute (La Jolla, CA).
Flow cytometryThe experiments were performed as described previously (9).
The primary antibodies were rabbit anti-human NRP-1 b1b2prepared in-house as by immunizing rabbits with a humanNRP-1 b1b2 protein, goat anti-human NRP-2 (R&D Systems),mouse anti-human avb3 (LM609) (EMDMillipore), mouse anti-humanavb5 (P1F6) (EMDMillipore), rat anti-mouseav (RMV-7;eBioscience), rat anti-mousea5 (MHR5; SouthernBiotech),mouseanti-human a5b1 (JBS5; Thermo Scientific), mouse anti-humanb1 (TS2/16) (eBioscience),mouse anti-human active b1 (HUTS-4;EMDMillipore), rat anti-mouseb1 (HMb1-1; eBioscience), and ratanti-mouse active b1 (9EG7; BD Biosciences). The primary anti-bodies were detected with corresponding secondary antibodiesconjugated to Alexa488, 594, or 647 (Molecular Probes). The cellswere analyzed with an LSR Fortessa System (BD Biosciences), andthe data were analyzed with a Flowjo software.
In vitro peptide internalization assayAs described elsewhere (8, 9), tumor cells were grown on
collagen type I–coated coverslips (BD Biosciences) overnight infully supplemented DMEM, incubated with 10 mmol/L of fluo-rescein-conjugated iRGD (FAM-iRGD) or FAM-labeled iNGRpeptide (cyclic CRNGRGPDC) in the presence of 10 mg/mL ofanti-NRP-1 b1b2or control IgG for 4 hours. The cells werewashedwith warm PBS, fixed in 4% paraformaldehyde (PFA), stainedwith 4,6-diamidino-2-phenylindole (DAPI; Molecular Probes),and observed under a Fluoview 500 confocal microscope (Olym-pus America).
In vivo peptide homing assayAs described previously (9), 100 mg of peptide in PBS were
intravenously injected into tumor mice and allowed to circulatefor 60 minutes. The mice were perfused through the heart withPBS containing 1%BSA, and tissues were harvested and processedfor immunofluorescence. The tissue sections were examined by aFluoview 500 confocal microscope.
Transwell migration assayCell migration was analyzed using 24-well Transwell cham-
bers containing polycarbonate membranes with 8 mm pores
(Corning Inc.; ref. 15). Both sides of the membranes werecoated with 50 mg/mL of collagen type-I (BD Biosciences) tofacilitate initial cell attachment to the membranes. GFP-PC-3 (4� 104 cells) or LM-PmC (2 � 105 cells) cells in DMEM contain-ing 0.1% BSA were added to the upper compartment. The lowercompartment was filled with 600 mL of DMEM containing 0.1%BSA. Peptides at a final concentration of 10 mmol/L or PBS wereadded to both the upper and lower compartments, or in somecases, only to the lower compartment. In some experiments, thecells were treated with 10 mg/mL of anti-NRP-1 b1b2 or controlIgG (Abcam) for 30 minutes before seeding and throughout theassay. After incubation in a CO2 incubator at 37�C for 24 hours,the cells on the upper side of the membranes were gently wipedoff, and the membranes were fixed in methanol and stainedwith hematoxylin and eosin. The membranes were mounted onglass slides, and imaged under a light microscope. The totalnumber of cells that migrated to the lower side of the mem-branes was determined by counting cell numbers under lowmagnification (�100).
Chemorepulsion assaySilver nanoparticles (AgNPs; diameter, 70 nm) were prepared
as described previously (16). In brief, AgNPs synthesized bypolyvinylpyrolidone/ethylene glycol reduction method were pre-cipitated with acetone, and redissolved in water. The AgNPs werethen coated with NeutrAvidin (NA)–5-kDa polyethylene glycol(PEG)-Orthopyridyl disulfide (OPSS; NA, Thermo Scientific; N-Hydroxysuccinimide-PEG-OPSS, JenKem Technology USA),backfilled with lipoic-3-kDa PEG-amine (Nanocs), and labeledwith amine-reactive CF555-succinimidyl ester dye (Biotium Inc.).The NA-AgNPs were then coated with biotinylated peptides orwith free biotin (Sigma-Aldrich). iRGD-AgNPs, CRGDC-AgNPs,andAgNPswithout any peptides were blotted along the peripheryof glass-bottom wells by applying 10 mL drops of nanoparticlesolutions on the glass surface and drying the drops in a laminarairflow chamber for 5 minutes. The wells were briefly rinsed withPBS three times, and 2� 105 LM-PmC cells in full culture DMEMwere seeded using a cloning cylinder in the center of the wells inclose proximitywith the AgNPs, or in some experiments, on topofthe AgNP-covered areas. The cells were allowed to attach to theglass surface in a CO2 incubator for 3 hours before the cylinderwas removed, and further cultured in DMEMwith 1% BSA for 24hours. The wells were then subjected to live cell imaging by takingtime-lapse images every 15minutes for 48 hours with an InvertedIX81Wide Field and FluorescenceMicroscope equippedwithCO2
and Temperature Controlled Time Lapse System (OlympusAmerica).
Cell attachment assayThe assays were performed following a protocol described
elsewhere (17). Cells resuspended in DMEM containing 1% BSAwere treated with peptides for 30 minutes at 37�C under mildrotation. The cells were seeded in 96-well plates coated withhuman fibronectin (R&D Systems) or bovine collagen type-I(BioPioneer) at 4 � 105 cells/mL (LM-PmC) or 2 � 105 cells/mL (GFP-PC-3), and allowed to attach to thewells for 30minutesat 37�C in a CO2 incubator in the presence of the peptides. Insome cases, the cells were treated with the peptides in thepresence of 10 mg/mL of anti-NRP-1 b1b2 or control IgG, ortreated solely with anti-human b1 (EMD Millipore) or anti-mouse b1 (eBioscience) integrin subunit antibodies or control
iRGD Inhibits Metastasis and Repels Tumor Cells
www.aacrjournals.org Mol Cancer Ther; 14(1) January 2015 121
on January 18, 2015. © 2015 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst November 12, 2014; DOI: 10.1158/1535-7163.MCT-14-0366
IgG (Abcam). After the incubation, the cells were treated with0.25% crystalline trypsin for 2 minutes, and the reaction wasstopped by adding 0.5 mg/mL soybean inhibitor. The wells weregently washed with warm DMEM to remove detached cells, andthe cells that remained attached to the wells were cultured in fullDMEM containing 0.5 mg/mL of MTT. Two hours later, the cellswere lysed with a buffer containing dimethylsulfoxide andmethanol at 1:1 volume ratio and absorbance at 595 nm wasread with a microplate reader.
In vitro cell retraction assayCells were cultured on fibronectin-coated coverslips for 1 hour
at 37�C in a CO2 incubator, and peptides were added at a finalconcentration of 10 mmol/L. The cells were cultured for another30 minutes at 37�C in a CO2 incubator, fixed in 4% PFA, stainedwith a rabbit anti-phospho-paxillin pTyr118 antibody (PierceBiotechnology) and DAPI, and imaged under a Fluoview 500confocal microscope. The anti-phospho-paxillin pTyr118 wasdetected with an Alexa488 donkey anti-rabbit antibody (Molec-ular Probes).
Statistical analysisData were analyzed by the two-tailed Student t test, one-way
ANOVA followed by a suitable post hoc test or a Pearson test. Theresults are summarized in Supplementary Table S1.
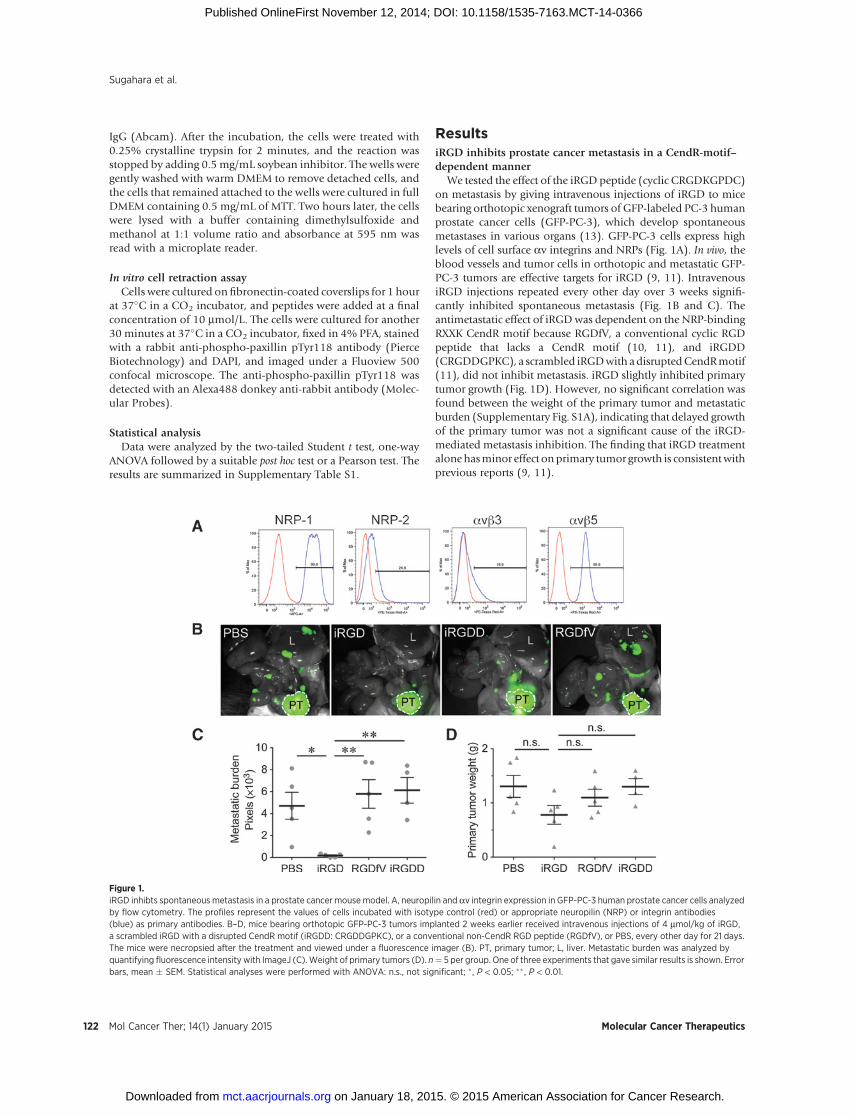
ResultsiRGD inhibits prostate cancer metastasis in a CendR-motif–dependent manner
We tested the effect of the iRGD peptide (cyclic CRGDKGPDC)on metastasis by giving intravenous injections of iRGD to micebearing orthotopic xenograft tumors of GFP-labeled PC-3 humanprostate cancer cells (GFP-PC-3), which develop spontaneousmetastases in various organs (13). GFP-PC-3 cells express highlevels of cell surface av integrins and NRPs (Fig. 1A). In vivo, theblood vessels and tumor cells in orthotopic and metastatic GFP-PC-3 tumors are effective targets for iRGD (9, 11). IntravenousiRGD injections repeated every other day over 3 weeks signifi-cantly inhibited spontaneous metastasis (Fig. 1B and C). Theantimetastatic effect of iRGDwas dependent on the NRP-bindingRXXK CendR motif because RGDfV, a conventional cyclic RGDpeptide that lacks a CendR motif (10, 11), and iRGDD(CRGDDGPKC), a scrambled iRGDwith adisruptedCendRmotif(11), did not inhibit metastasis. iRGD slightly inhibited primarytumor growth (Fig. 1D). However, no significant correlation wasfound between the weight of the primary tumor and metastaticburden (Supplementary Fig. S1A), indicating that delayed growthof the primary tumor was not a significant cause of the iRGD-mediated metastasis inhibition. The finding that iRGD treatmentalonehasminor effect onprimary tumor growth is consistentwithprevious reports (9, 11).
Figure 1.iRGD inhibts spontaneous metastasis in a prostate cancer mousemodel. A, neuropilin and av integrin expression in GFP-PC-3 human prostate cancer cells analyzedby flow cytometry. The profiles represent the values of cells incubated with isotype control (red) or appropriate neuropilin (NRP) or integrin antibodies(blue) as primary antibodies. B–D, mice bearing orthotopic GFP-PC-3 tumors implanted 2 weeks earlier received intravenous injections of 4 mmol/kg of iRGD,a scrambled iRGD with a disrupted CendR motif (iRGDD: CRGDDGPKC), or a conventional non-CendR RGD peptide (RGDfV), or PBS, every other day for 21 days.The mice were necropsied after the treatment and viewed under a fluorescence imager (B). PT, primary tumor; L, liver. Metastatic burden was analyzed byquantifying fluorescence intensity with ImageJ (C).Weight of primary tumors (D). n¼ 5 per group. One of three experiments that gave similar results is shown. Errorbars, mean � SEM. Statistical analyses were performed with ANOVA: n.s., not significant; � , P < 0.05; �� , P < 0.01.
Sugahara et al.
Mol Cancer Ther; 14(1) January 2015 Molecular Cancer Therapeutics122
on January 18, 2015. © 2015 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst November 12, 2014; DOI: 10.1158/1535-7163.MCT-14-0366
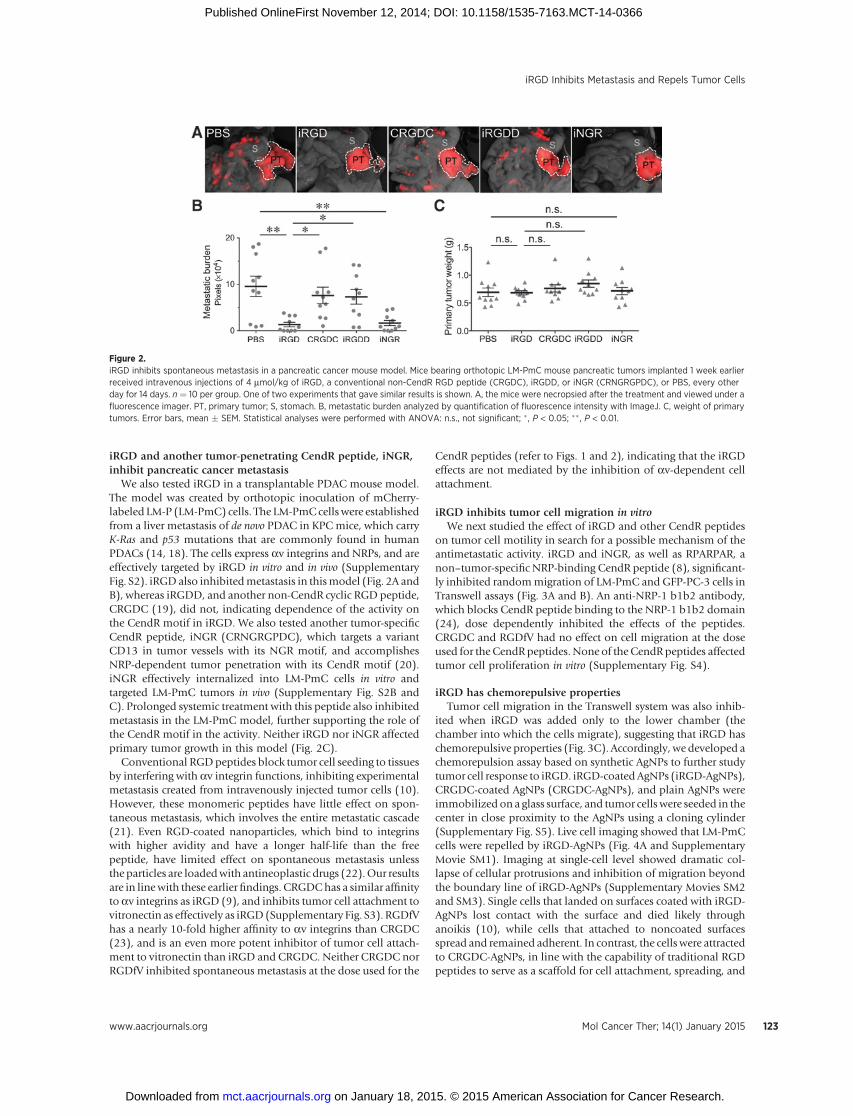
iRGD and another tumor-penetrating CendR peptide, iNGR,inhibit pancreatic cancer metastasis
We also tested iRGD in a transplantable PDAC mouse model.The model was created by orthotopic inoculation of mCherry-labeled LM-P (LM-PmC) cells. The LM-PmC cells were establishedfrom a liver metastasis of de novo PDAC in KPC mice, which carryK-Ras and p53 mutations that are commonly found in humanPDACs (14, 18). The cells express av integrins and NRPs, and areeffectively targeted by iRGD in vitro and in vivo (SupplementaryFig. S2). iRGDalso inhibitedmetastasis in thismodel (Fig. 2A andB), whereas iRGDD, and another non-CendR cyclic RGD peptide,CRGDC (19), did not, indicating dependence of the activity onthe CendR motif in iRGD. We also tested another tumor-specificCendR peptide, iNGR (CRNGRGPDC), which targets a variantCD13 in tumor vessels with its NGR motif, and accomplishesNRP-dependent tumor penetration with its CendR motif (20).iNGR effectively internalized into LM-PmC cells in vitro andtargeted LM-PmC tumors in vivo (Supplementary Fig. S2B andC). Prolonged systemic treatment with this peptide also inhibitedmetastasis in the LM-PmC model, further supporting the role ofthe CendR motif in the activity. Neither iRGD nor iNGR affectedprimary tumor growth in this model (Fig. 2C).
Conventional RGDpeptides block tumor cell seeding to tissuesby interfering with av integrin functions, inhibiting experimentalmetastasis created from intravenously injected tumor cells (10).However, these monomeric peptides have little effect on spon-taneous metastasis, which involves the entire metastatic cascade(21). Even RGD-coated nanoparticles, which bind to integrinswith higher avidity and have a longer half-life than the freepeptide, have limited effect on spontaneous metastasis unlessthe particles are loadedwith antineoplastic drugs (22).Our resultsare in linewith these earlierfindings. CRGDChas a similar affinityto av integrins as iRGD (9), and inhibits tumor cell attachment tovitronectin as effectively as iRGD (Supplementary Fig. S3). RGDfVhas a nearly 10-fold higher affinity to av integrins than CRGDC(23), and is an even more potent inhibitor of tumor cell attach-ment to vitronectin than iRGD and CRGDC. Neither CRGDC norRGDfV inhibited spontaneous metastasis at the dose used for the
CendR peptides (refer to Figs. 1 and 2), indicating that the iRGDeffects are not mediated by the inhibition of av-dependent cellattachment.
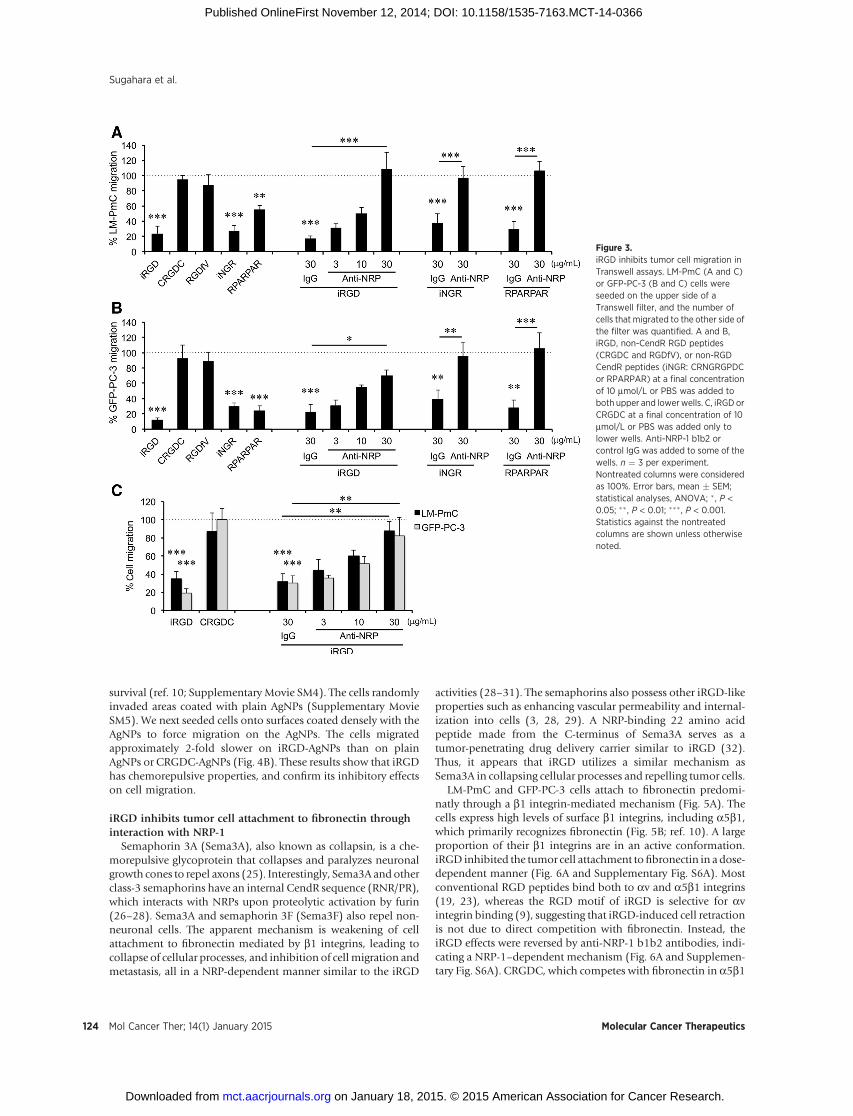
iRGD inhibits tumor cell migration in vitroWe next studied the effect of iRGD and other CendR peptides
on tumor cell motility in search for a possible mechanism of theantimetastatic activity. iRGD and iNGR, as well as RPARPAR, anon–tumor-specific NRP-binding CendR peptide (8), significant-ly inhibited randommigration of LM-PmC and GFP-PC-3 cells inTranswell assays (Fig. 3A and B). An anti-NRP-1 b1b2 antibody,which blocks CendR peptide binding to the NRP-1 b1b2 domain(24), dose dependently inhibited the effects of the peptides.CRGDC and RGDfV had no effect on cell migration at the doseused for the CendRpeptides. None of the CendRpeptides affectedtumor cell proliferation in vitro (Supplementary Fig. S4).
iRGD has chemorepulsive propertiesTumor cell migration in the Transwell system was also inhib-
ited when iRGD was added only to the lower chamber (thechamber into which the cells migrate), suggesting that iRGD haschemorepulsive properties (Fig. 3C). Accordingly, we developed achemorepulsion assay based on synthetic AgNPs to further studytumor cell response to iRGD. iRGD-coatedAgNPs (iRGD-AgNPs),CRGDC-coated AgNPs (CRGDC-AgNPs), and plain AgNPs wereimmobilized on a glass surface, and tumor cells were seeded in thecenter in close proximity to the AgNPs using a cloning cylinder(Supplementary Fig. S5). Live cell imaging showed that LM-PmCcells were repelled by iRGD-AgNPs (Fig. 4A and SupplementaryMovie SM1). Imaging at single-cell level showed dramatic col-lapse of cellular protrusions and inhibition of migration beyondthe boundary line of iRGD-AgNPs (Supplementary Movies SM2and SM3). Single cells that landed on surfaces coated with iRGD-AgNPs lost contact with the surface and died likely throughanoikis (10), while cells that attached to noncoated surfacesspread and remained adherent. In contrast, the cells were attractedto CRGDC-AgNPs, in line with the capability of traditional RGDpeptides to serve as a scaffold for cell attachment, spreading, and
Figure 2.iRGD inhibits spontaneous metastasis in a pancreatic cancer mouse model. Mice bearing orthotopic LM-PmC mouse pancreatic tumors implanted 1 week earlierreceived intravenous injections of 4 mmol/kg of iRGD, a conventional non-CendR RGD peptide (CRGDC), iRGDD, or iNGR (CRNGRGPDC), or PBS, every otherday for 14 days. n ¼ 10 per group. One of two experiments that gave similar results is shown. A, the mice were necropsied after the treatment and viewed under afluorescence imager. PT, primary tumor; S, stomach. B, metastatic burden analyzed by quantification of fluorescence intensity with ImageJ. C, weight of primarytumors. Error bars, mean � SEM. Statistical analyses were performed with ANOVA: n.s., not significant; � , P < 0.05; �� , P < 0.01.
iRGD Inhibits Metastasis and Repels Tumor Cells
www.aacrjournals.org Mol Cancer Ther; 14(1) January 2015 123
on January 18, 2015. © 2015 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst November 12, 2014; DOI: 10.1158/1535-7163.MCT-14-0366
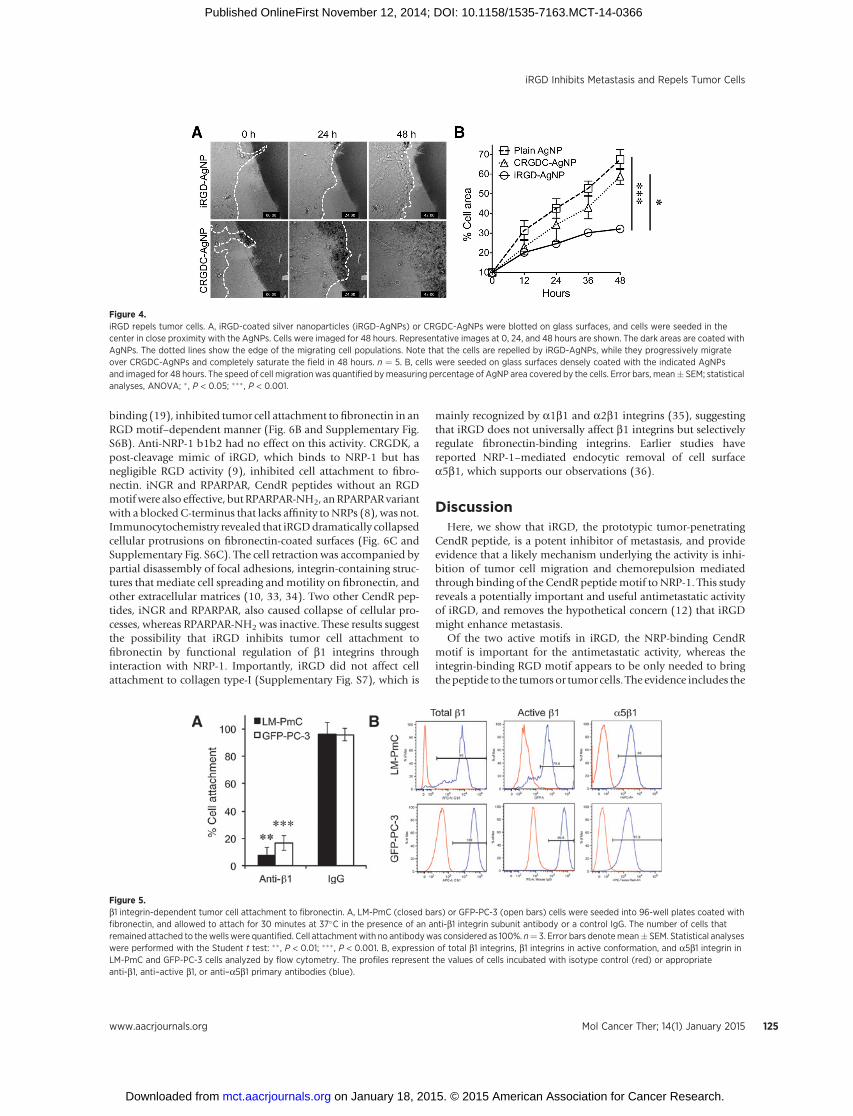
survival (ref. 10; Supplementary Movie SM4). The cells randomlyinvaded areas coated with plain AgNPs (Supplementary MovieSM5). We next seeded cells onto surfaces coated densely with theAgNPs to force migration on the AgNPs. The cells migratedapproximately 2-fold slower on iRGD-AgNPs than on plainAgNPs or CRGDC-AgNPs (Fig. 4B). These results show that iRGDhas chemorepulsive properties, and confirm its inhibitory effectson cell migration.
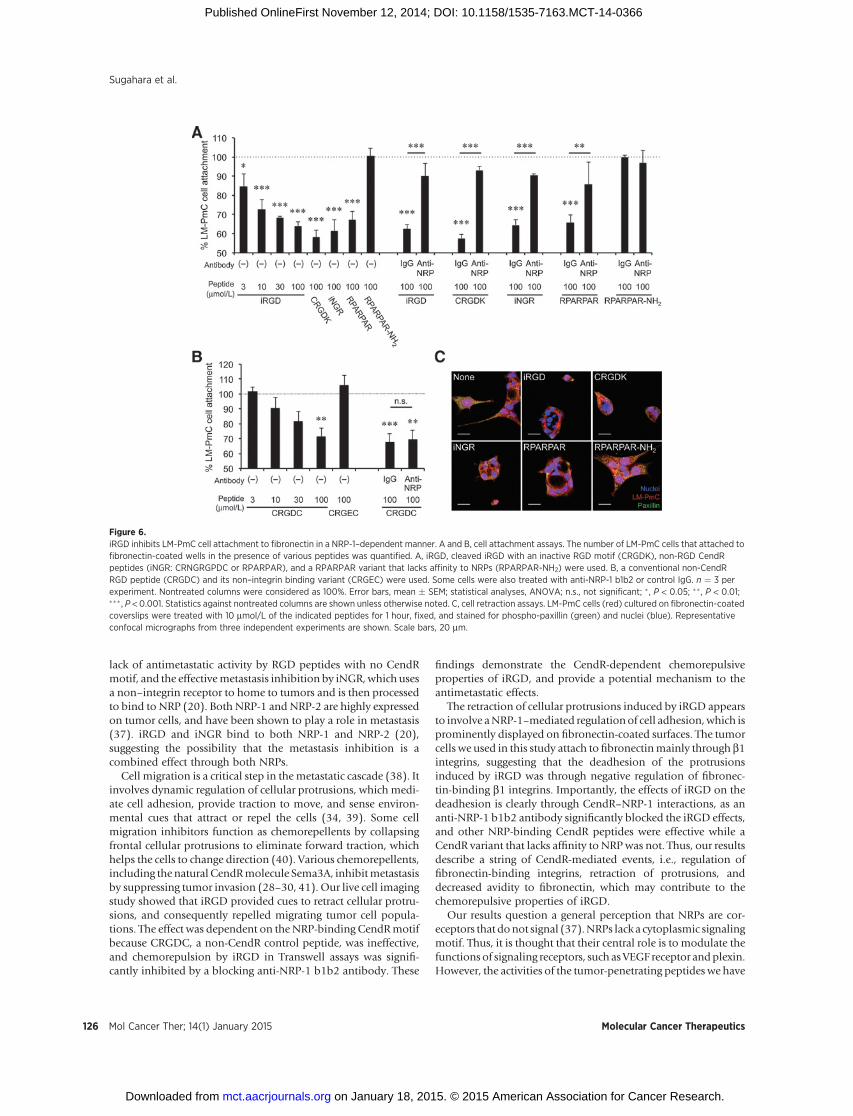
iRGD inhibits tumor cell attachment to fibronectin throughinteraction with NRP-1
Semaphorin 3A (Sema3A), also known as collapsin, is a che-morepulsive glycoprotein that collapses and paralyzes neuronalgrowth cones to repel axons (25). Interestingly, Sema3A andotherclass-3 semaphorins have an internal CendR sequence (RNR/PR),which interacts with NRPs upon proteolytic activation by furin(26–28). Sema3A and semaphorin 3F (Sema3F) also repel non-neuronal cells. The apparent mechanism is weakening of cellattachment to fibronectin mediated by b1 integrins, leading tocollapse of cellular processes, and inhibition of cellmigration andmetastasis, all in a NRP-dependent manner similar to the iRGD
activities (28–31). The semaphorins also possess other iRGD-likeproperties such as enhancing vascular permeability and internal-ization into cells (3, 28, 29). A NRP-binding 22 amino acidpeptide made from the C-terminus of Sema3A serves as atumor-penetrating drug delivery carrier similar to iRGD (32).Thus, it appears that iRGD utilizes a similar mechanism asSema3A in collapsing cellular processes and repelling tumor cells.
LM-PmC and GFP-PC-3 cells attach to fibronectin predomi-natly through a b1 integrin-mediated mechanism (Fig. 5A). Thecells express high levels of surface b1 integrins, including a5b1,which primarily recognizes fibronectin (Fig. 5B; ref. 10). A largeproportion of their b1 integrins are in an active conformation.iRGD inhibited the tumor cell attachment tofibronectin in adose-dependent manner (Fig. 6A and Supplementary Fig. S6A). Mostconventional RGD peptides bind both to av and a5b1 integrins(19, 23), whereas the RGD motif of iRGD is selective for avintegrin binding (9), suggesting that iRGD-induced cell retractionis not due to direct competition with fibronectin. Instead, theiRGD effects were reversed by anti-NRP-1 b1b2 antibodies, indi-cating a NRP-1–dependent mechanism (Fig. 6A and Supplemen-tary Fig. S6A). CRGDC, which competes with fibronectin in a5b1
Figure 3.iRGD inhibits tumor cell migration inTranswell assays. LM-PmC (A and C)or GFP-PC-3 (B and C) cells wereseeded on the upper side of aTranswell filter, and the number ofcells that migrated to the other side ofthe filter was quantified. A and B,iRGD, non-CendR RGD peptides(CRGDC and RGDfV), or non-RGDCendR peptides (iNGR: CRNGRGPDCor RPARPAR) at a final concentrationof 10 mmol/L or PBS was added toboth upper and lowerwells. C, iRGD orCRGDC at a final concentration of 10mmol/L or PBS was added only tolower wells. Anti-NRP-1 b1b2 orcontrol IgG was added to some of thewells. n ¼ 3 per experiment.Nontreated columns were consideredas 100%. Error bars, mean � SEM;statistical analyses, ANOVA; �, P <0.05; �� , P < 0.01; ��� , P < 0.001.Statistics against the nontreatedcolumns are shown unless otherwisenoted.
Sugahara et al.
Mol Cancer Ther; 14(1) January 2015 Molecular Cancer Therapeutics124
on January 18, 2015. © 2015 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst November 12, 2014; DOI: 10.1158/1535-7163.MCT-14-0366
binding (19), inhibited tumor cell attachment to fibronectin in anRGD motif–dependent manner (Fig. 6B and Supplementary Fig.S6B). Anti-NRP-1 b1b2 had no effect on this activity. CRGDK, apost-cleavage mimic of iRGD, which binds to NRP-1 but hasnegligible RGD activity (9), inhibited cell attachment to fibro-nectin. iNGR and RPARPAR, CendR peptides without an RGDmotifwere also effective, butRPARPAR-NH2, anRPARPARvariantwith a blocked C-terminus that lacks affinity toNRPs (8), was not.Immunocytochemistry revealed that iRGDdramatically collapsedcellular protrusions on fibronectin-coated surfaces (Fig. 6C andSupplementary Fig. S6C). The cell retraction was accompanied bypartial disassembly of focal adhesions, integrin-containing struc-tures that mediate cell spreading andmotility on fibronectin, andother extracellular matrices (10, 33, 34). Two other CendR pep-tides, iNGR and RPARPAR, also caused collapse of cellular pro-cesses, whereas RPARPAR-NH2 was inactive. These results suggestthe possibility that iRGD inhibits tumor cell attachment tofibronectin by functional regulation of b1 integrins throughinteraction with NRP-1. Importantly, iRGD did not affect cellattachment to collagen type-I (Supplementary Fig. S7), which is
mainly recognized by a1b1 and a2b1 integrins (35), suggestingthat iRGD does not universally affect b1 integrins but selectivelyregulate fibronectin-binding integrins. Earlier studies havereported NRP-1–mediated endocytic removal of cell surfacea5b1, which supports our observations (36).
DiscussionHere, we show that iRGD, the prototypic tumor-penetrating
CendR peptide, is a potent inhibitor of metastasis, and provideevidence that a likely mechanism underlying the activity is inhi-bition of tumor cell migration and chemorepulsion mediatedthrough binding of the CendR peptidemotif to NRP-1. This studyreveals a potentially important and useful antimetastatic activityof iRGD, and removes the hypothetical concern (12) that iRGDmight enhance metastasis.
Of the two active motifs in iRGD, the NRP-binding CendRmotif is important for the antimetastatic activity, whereas theintegrin-binding RGD motif appears to be only needed to bringthepeptide to the tumors or tumor cells. The evidence includes the
Figure 4.iRGD repels tumor cells. A, iRGD-coated silver nanoparticles (iRGD-AgNPs) or CRGDC-AgNPs were blotted on glass surfaces, and cells were seeded in thecenter in close proximity with the AgNPs. Cells were imaged for 48 hours. Representative images at 0, 24, and 48 hours are shown. The dark areas are coated withAgNPs. The dotted lines show the edge of the migrating cell populations. Note that the cells are repelled by iRGD-AgNPs, while they progressively migrateover CRGDC-AgNPs and completely saturate the field in 48 hours. n ¼ 5. B, cells were seeded on glass surfaces densely coated with the indicated AgNPsand imaged for 48 hours. The speed of cell migration was quantified bymeasuring percentage of AgNP area covered by the cells. Error bars, mean� SEM; statisticalanalyses, ANOVA; � , P < 0.05; ��� , P < 0.001.
Figure 5.b1 integrin-dependent tumor cell attachment to fibronectin. A, LM-PmC (closed bars) or GFP-PC-3 (open bars) cells were seeded into 96-well plates coated withfibronectin, and allowed to attach for 30 minutes at 37�C in the presence of an anti-b1 integrin subunit antibody or a control IgG. The number of cells thatremained attached to thewellswere quantified. Cell attachmentwith no antibodywas considered as 100%. n¼ 3. Error bars denotemean� SEM. Statistical analyseswere performed with the Student t test: �� , P < 0.01; ��� , P < 0.001. B, expression of total b1 integrins, b1 integrins in active conformation, and a5b1 integrin inLM-PmC and GFP-PC-3 cells analyzed by flow cytometry. The profiles represent the values of cells incubated with isotype control (red) or appropriateanti-b1, anti–active b1, or anti–a5b1 primary antibodies (blue).
iRGD Inhibits Metastasis and Repels Tumor Cells
www.aacrjournals.org Mol Cancer Ther; 14(1) January 2015 125
on January 18, 2015. © 2015 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst November 12, 2014; DOI: 10.1158/1535-7163.MCT-14-0366
lack of antimetastatic activity by RGD peptides with no CendRmotif, and the effectivemetastasis inhibition by iNGR, which usesa non–integrin receptor to home to tumors and is then processedto bind to NRP (20). Both NRP-1 and NRP-2 are highly expressedon tumor cells, and have been shown to play a role in metastasis(37). iRGD and iNGR bind to both NRP-1 and NRP-2 (20),suggesting the possibility that the metastasis inhibition is acombined effect through both NRPs.
Cell migration is a critical step in themetastatic cascade (38). Itinvolves dynamic regulation of cellular protrusions, which medi-ate cell adhesion, provide traction to move, and sense environ-mental cues that attract or repel the cells (34, 39). Some cellmigration inhibitors function as chemorepellents by collapsingfrontal cellular protrusions to eliminate forward traction, whichhelps the cells to change direction (40). Various chemorepellents,including the natural CendRmolecule Sema3A, inhibitmetastasisby suppressing tumor invasion (28–30, 41). Our live cell imagingstudy showed that iRGD provided cues to retract cellular protru-sions, and consequently repelled migrating tumor cell popula-tions. The effect was dependent on the NRP-binding CendRmotifbecause CRGDC, a non-CendR control peptide, was ineffective,and chemorepulsion by iRGD in Transwell assays was signifi-cantly inhibited by a blocking anti-NRP-1 b1b2 antibody. These
findings demonstrate the CendR-dependent chemorepulsiveproperties of iRGD, and provide a potential mechanism to theantimetastatic effects.
The retraction of cellular protrusions induced by iRGD appearsto involve aNRP-1–mediated regulationof cell adhesion,which isprominently displayed on fibronectin-coated surfaces. The tumorcells we used in this study attach to fibronectinmainly through b1integrins, suggesting that the deadhesion of the protrusionsinduced by iRGD was through negative regulation of fibronec-tin-binding b1 integrins. Importantly, the effects of iRGD on thedeadhesion is clearly through CendR–NRP-1 interactions, as ananti-NRP-1 b1b2 antibody significantly blocked the iRGD effects,and other NRP-binding CendR peptides were effective while aCendR variant that lacks affinity to NRPwas not. Thus, our resultsdescribe a string of CendR-mediated events, i.e., regulation offibronectin-binding integrins, retraction of protrusions, anddecreased avidity to fibronectin, which may contribute to thechemorepulsive properties of iRGD.
Our results question a general perception that NRPs are cor-eceptors that donot signal (37).NRPs lack a cytoplasmic signalingmotif. Thus, it is thought that their central role is to modulate thefunctions of signaling receptors, such as VEGF receptor andplexin.However, the activities of the tumor-penetrating peptides we have
Figure 6.iRGD inhibits LM-PmC cell attachment to fibronectin in a NRP-1–dependent manner. A and B, cell attachment assays. The number of LM-PmC cells that attached tofibronectin-coated wells in the presence of various peptides was quantified. A, iRGD, cleaved iRGD with an inactive RGD motif (CRGDK), non-RGD CendRpeptides (iNGR: CRNGRGPDC or RPARPAR), and a RPARPAR variant that lacks affinity to NRPs (RPARPAR-NH2) were used. B, a conventional non-CendRRGD peptide (CRGDC) and its non–integrin binding variant (CRGEC) were used. Some cells were also treated with anti-NRP-1 b1b2 or control IgG. n ¼ 3 perexperiment. Nontreated columns were considered as 100%. Error bars, mean � SEM; statistical analyses, ANOVA; n.s., not significant; � , P < 0.05; �� , P < 0.01;��� , P < 0.001. Statistics against nontreated columns are shown unless otherwise noted. C, cell retraction assays. LM-PmC cells (red) cultured on fibronectin-coatedcoverslips were treated with 10 mmol/L of the indicated peptides for 1 hour, fixed, and stained for phospho-paxillin (green) and nuclei (blue). Representativeconfocal micrographs from three independent experiments are shown. Scale bars, 20 mm.
Sugahara et al.
Mol Cancer Ther; 14(1) January 2015 Molecular Cancer Therapeutics126
on January 18, 2015. © 2015 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst November 12, 2014; DOI: 10.1158/1535-7163.MCT-14-0366

used in this study suggest independent signaling by NRPs. Theeffects of our peptides resemble those of Sema3A with an impor-tant difference: Semaphorins are large proteins that have activitiesother than NRP binding, whereas the small size of our peptidesand the fact that their only common denominator is the presenceof a CendR motif, strongly suggest that NRP binding alone isresponsible for the antimetastatic and cell-repelling activities.Supporting this possibility, recent studies demonstrate that NRPsalone are capable ofmediating various effects, such as endocytosisand phosphorylation of signaling adaptor molecules (6). Tumor-penetrating peptides may be ideal tools for dissecting intrinsicNRP functions.
The dose and administration schedules we used to establish theantimetastatic effects of iRGD (and iNGR) were identical to thosewe have previously used to promote drug delivery into tumors(11). Thus, our results suggest that tumor-penetrating CendRpeptides will simultaneously achieve metastasis inhibition whenused as an adjuvant therapy to promote tumor-specific drugdelivery. The antimetastatic effects of iRGD could be useful inpreventing initialmetastasis, in suppressing additionalmetastasiswhen metastasis has already taken place, and in overcoming theprometastatic side effects of antiangiogenic therapies (41, 42).
Disclosure of Potential Conflicts of InterestK.N. Sugahara, V.R. Kotamraju, T. Teesalu, and E. Ruoslahti have ownership
interest (including patents) in CendR Therapeutics Inc. No potential conflicts ofinterest were disclosed by the other authors.
DisclaimerThe views and opinions of authors expressed on OER websites do not
necessarily state or reflect those of the U.S. Government, and they may not beused for advertising or product endorsement purposes.
Authors' ContributionsConception and design: K.N. Sugahara, T. Teesalu, E. RuoslahtiDevelopment of methodology: K.N. Sugahara, G.B. Braun, T. Teesalu,E. RuoslahtiAcquisition of data (provided animals, acquired and managed patients,provided facilities, etc.): K.N. Sugahara, G.B. Braun, T.H. de Mendoza, A.M.Lowy, T. Teesalu, E. RuoslahtiAnalysis and interpretation of data (e.g., statistical analysis, biostatistics,computational analysis): K.N. Sugahara, G.B. Braun, T. Teesalu, E. RuoslahtiWriting, review, and/or revision of themanuscript:K.N. Sugahara, G.B. Braun,T.H. de Mendoza, T.H. de Mendoza, A.M. Lowy, T. Teesalu, E. RuoslahtiAdministrative, technical, or material support (i.e., reporting or organizingdata, constructing databases): K.N. Sugahara, T. Teesalu, E. RuoslahtiStudy supervision: K.N. Sugahara, E. RuoslahtiOther (synthesis of peptides and contributed to the chemistry aspects of thestudy): V.R. KotamrajuOther [derived and supplied cell lines used in some of the experiments(mouse and human CAF cell lines; mouse and human PDAC cell lines)]: R.P.French
Grant SupportThis work was supported by grants R01CA167174 (to K.N. Sugahara) and
R01CA152327 (to E. Ruoslahti) from the National Cancer Institute of NIH,Career Development Award from American Association of Cancer Research/Pancreatic Cancer Action Network (to K.N. Sugahara), and Blasker ScienceAward from the San Diego Foundation (to K.N. Sugahara). G.B. Braun wassupported by CA121949 NIH T32 Fellowship and T. Teesalu was supported byKG110704 Susan Komen for Cure Foundation Career Development Award,European Research Council Starting Grant (GliomaDDS), and Wellcome TrustInternational Fellowship (WT095077MA).
The costs of publication of this articlewere defrayed inpart by the payment ofpage charges. This article must therefore be hereby marked advertisement inaccordance with 18 U.S.C. Section 1734 solely to indicate this fact.
Received April 28, 2014; revised September 22, 2014; accepted October 29,2014; published OnlineFirst November 12, 2014.
References1. McDonald DM, Baluk P. Significance of blood vessel leakiness in cancer.
Cancer Res 2002;62:5381–5.
2. Heldin CH, Rubin K, Pietras K, €Ostman A. High interstitial fluid pressure –An obstacle in cancer therapy. Nat Rev Cancer 2004;4:806–3.
3. Prabhakar U, Maeda H, Jain RK, Sevick-Muraca EM, Zamboni W, Farokh-zad OC, et al. Challenges and key considerations of the enhanced perme-ability and retention effect for nanomedicine drug delivery in oncology.Cancer Res 2013;73:2412–7.
4. Fukumura D, Kashiwagi S, Jain RK. The role of nitric oxide in tumourprogression. Nat Rev Cancer 2006;6:521–34.
5. Zhou W, Fong MY, Min Y, Somlo G, Liu L, Palomares MR, et al. Cancer-secreted miR105 destroys endothelial barriers to promote metastasis.Cancer Cell 2014;25:501–15.
6. Prud'homme GJ, Glinka Y. Neuropilins are multifunctional coreceptorsinvolved in tumor initiation, growth, metastasis and immunity. Oncotar-get 2012;3:921–39.
7. Teesalu T, Sugahara KN, Ruoslahti E. Tumor-penetrating peptides. FrontOncol 2013;3:216.
8. Teesalu T, Sugahara KN, Kotamraju VR, Ruoslahti E. C-end Rule: Peptideswith C-terminal arginine cause neuropilin-1 dependent internalization,vascular leakage and tissue penetration. Proc Natl Acad Sci U S A2009;106:16157–62.
9. Sugahara KN, Teesalu T, Karmali PP, Kotamraju VR, Agemy L, Girard OM,et al. Tissue-penetrating delivery of compounds and nanoparticles intotumors. Cancer Cell 2009;16:510–20.
10. Desgrosellier JS, Cheresh DA. Integrins in cancer: biological implicationsand therapeutic opportunities. Nat Rev Cancer 2010;10:9–22.
11. Sugahara KN, Teesalu T, Karmali PP, Kotamraju VR, Agemy L, GreenwaldDR, et al. Coadministration of a tumor-penetrating peptide enhances theefficacy of cancer drugs. Science 2010;328:1031–5.
12. Feron O. Tumor-penetrating peptides: a shift from magic bullets to magicguns. Sci Transl Med 2010;2:34ps26.
13. Yang M, Jiang P, Sun FX, Hasegawa S, Baranov E, Chishima T, et al. Afluorescent orthotopic bone metastasis model of human prostate cancer.Cancer Res 1999;59:781–6.
14. Tseng WW, Winer D, Kenkel JA, Choi O, Shain AH, Pollack JR, et al.Development of an orthotopic model of invasive pancreatic cancer in animmunocompetent murine host. Clin Cancer Res 2010;16:3684–95.
15. Sugahara KN,Murai T, NishinakamuraH, KawashimaH, SayaH,MiyasakaM. Hyaluronan oligosaccharides induce CD44 cleavage and promotecell migration in CD44-expressing tumor cells. J Biol Chem 2004;279:4541–50.
16. BraunGB, Friman T, PangHB, PallaoroA, deMendoza TH,Willmore AMA,et al. Etchable plasmonic nanoparticle probes for cell biology. Nat Mater2014;13:904–11.
17. Ruoslahti E, Hayman EG, Pierschbacher MD, Engvall E. Fibronectin:purification, immunochemical properties, and biological activities. Meth-ods Enzymol 1982;82:803–31.
18. Olive KP, Tuveson DA. The use of targeted mouse models for preclinicaltesting of novel cancer therapeutics. Clin Cancer Res 2006;12:5277–87.
19. Koivunen E, Gay DA, Ruoslahti E. Selection of peptides binding to thealpha 5 beta 1 integrin from phage display library. J Biol Chem1993;268:20205–10.
20. Alberici L, Roth L, Sugahara KN, Agemy L, Kotamraju VR, Teesalu T,et al. De novo design of a tumor-penetrating peptide. Cancer Res2013;73:804–12.
21. Saiki I, Murata J, Iida J, Sakurai T, Nishi N, Matsuno K, et al. Antimetastaticeffects of synthetic polypeptides containing repeated structures of the celladhesive Arg-Gly-Asp (RGD) and Tyr-Ile-Gly-Ser-Arg (YIGSR) sequences.Br J Cancer 1989;60:722–8.
www.aacrjournals.org Mol Cancer Ther; 14(1) January 2015 127
iRGD Inhibits Metastasis and Repels Tumor Cells
on January 18, 2015. © 2015 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst November 12, 2014; DOI: 10.1158/1535-7163.MCT-14-0366

22. Murphy EA, Majeti BK, Barnes LA, Makale M, Weis SM, Lutu-Fuga K, et al.Nanoparticle-mediated drug delivery to tumor vasculature suppressesmetastasis. Proc Natl Acad Sci U S A 2009;105:9343–8.
23. Ye Y, Xu B, Nikiforovich GV, Bloch S, Achilefu S. Exploring new near-infrared fluorescent disulfide-based cyclic RGD peptide analogs for poten-tial integrin-targeted optical imaging. Bioorg Med Chem Lett 2011;21:2116–20.
24. Zanuy D, Kotla R, Nussinov R, Teesalu T, Sugahara KN, Alem�an C, et al.Sequence dependence of C-end rule peptides in binding and activation ofneuropilin-1 receptor. J Struct Biol 2013;182:78–86.
25. Luo Y, Raible D, Raper JA. Collapsin: a protein in brain that induces thecollapse and paralysis of neuronal growth cones. Cell 1993;75:217–27.
26. He Z, Tessier-Lavigne M. Neuropilin is a receptor of the axonal chemor-epellent Semaphorin III. Cell 1997;90:739–51.
27. Parker MW, Linkugel AD, Vander Kooi AW. Effect of C-terminal sequenceon competitive semaphorin binding to neuropilin-1. J Mol Biol2013;425:4405–14.
28. Neufeld G, Kessler O. The semaphorins: versatile regulators of tumourprogression and tumour angiogenesis. Nat Rev Cancer 2008;8:632–45.
29. Kruger RP, Aurandt J, Guan KL. Semaphorins command cells to move. NatRev Mol Cell Biol 2005;6:789–800.
30. Bielenberg DR, Hida Y, Shimizu A, Kaipainen A, Kreuter M, Kim CC, et al.Semaphorin 3F, a chemorepulsant for endothelial cells, induces a poorlyvascularized, encapsulated, nonmetastatic tumor phenotype. J Clin Invest2004;114:1260–71.
31. Serini G, Valdembri D, Zanivan S, Morterra G, Burkhardt C, Caccavari F,et al. Class 3 semaphorins control vascular morphogenesis by inhibitingintegrin function. Nature 2003;424:391–7.
32. Shin TH, Sung ES, Kim YJ, Kim KS, Kim SH, Kim SK, et al. Enhancement oftumor penetration of monoclonal antibody by fusion of a neuropilin-targeting peptide improves the antitumor efficacy. Mol Cancer Ther2014;13:651–61.
33. Woo S, Gomez TM. Rac1 and RhoA promote neurite outgrowth throughformation and stabilization of growth cone point contacts. J Neurosci2006;26:1418–28.
34. Geiger B, Spatz JP, Bershadsky AD. Environmental sensing through focaladhesions. Nat Rev Mol Cell Biol 2009;10:21–33.
35. Prockop DJ, Kivirikko KI. Collagens: molecular biology, diseases, andpotentials for therapy. Annu Rev Biochem 1995;64:403–34.
36. Valdembri D, Caswell PT, Anderson KI, Schwarz JP, K€onig I, Astanina E,et al. Neuropilin-1/GIPC1 signaling regulates a5b1 integrin traffic andfunction in endothelial cells. PLoS Biol 2009;7:e1000025.
37. Bagri A, Tessier-Lavigne M, Watts RJ. Neuropilins in tumor biology. ClinCancer Res 2009;15:1860–4.
38. Nguyen DX, Bos PD, Massagu�e J. Metastasis: from dissemination to organ-specific colonization. Nat Rev Cancer 2009;9:274–84.
39. Friedl P, Wolf K. Tumour-cell invasion and migration: diversity andescape mechanism. Nat Rev Cancer 2003;3:362–74.
40. Luo Y, Raper JA. Inhibitory factors controlling growth cone motility andguidance. Curr Opin Neurobiol 1994;4:648–54.
41. Maione F, Capano S, Regano D, Zentilin L, Giacca M, Casanovas O, et al.Semaphorin 3A overcomes cancer hypoxia and metastatic disseminationinduced by antiangiogenic treatment in mice. J Clin Invest 2012;122:1832–48.
42. Carmeliet P, Jain RK. Principles and mechanisms of vesel normalization forcancer and other angiogenic diseases. Nat RevDrugDiscov 2011;10:417–27.
Mol Cancer Ther; 14(1) January 2015 Molecular Cancer Therapeutics128
Sugahara et al.
on January 18, 2015. © 2015 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst November 12, 2014; DOI: 10.1158/1535-7163.MCT-14-0366

2015;14:120-128. Published OnlineFirst November 12, 2014.Mol Cancer Ther Kazuki N. Sugahara, Gary B. Braun, Tatiana Hurtado de Mendoza, et al. Tumor-Penetrating iRGD Peptide Inhibits Metastasis
Updated version
10.1158/1535-7163.MCT-14-0366doi:
Access the most recent version of this article at:
Material
Supplementary
http://mct.aacrjournals.org/content/suppl/2014/11/13/1535-7163.MCT-14-0366.DC1.html
Access the most recent supplemental material at:
Cited Articles
http://mct.aacrjournals.org/content/14/1/120.full.html#ref-list-1
This article cites by 42 articles, 14 of which you can access for free at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications Department at
Permissions
To request permission to re-use all or part of this article, contact the AACR Publications Department at
on January 18, 2015. © 2015 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst November 12, 2014; DOI: 10.1158/1535-7163.MCT-14-0366