trends of karyotype evolution in neotropical oedionychina (coleoptera: chrysomelidae: alticinae)
TRANSCRIPT

Hereditas 119: 263-283 (1993)
Trends of karyotype evolution in neotropical Oedionychina ( Coleoptera: Chrysomelidae: Alticinae) NIILO VIRKKI’ and JORGE A. SANTIAGO-BLAY’
Crop Protection Department, Agricultural Experiment Station, University of Puerto Rico,
Department of Entomological Sciences, University of California, Berkeley, CA 94720-0001, U.S.A Rio Piedras 00928-1360 Puerto Rico
VIRKKI, N. and SANTIAGO-BLAY, J. A. 1993. Trends of karyotype evolution in neotropical Oedionychina (Coleoptera: Chrysomelidae: Alticinae). - Hereditas 119: 263-283. Lund, Sweden. ISSN 001 8-0661. Received March 22, 1993. Accepted August 26, 1993
Although the cytology of the Oedionychina flea beetles is unorthodox in many ways, their karyotypes are conservative in chromosome number and sex chromosome system: typically 2n = 22(X, y); 10 + X + y. Differences between karyotypes are expressed mainly in arm length ratios. This study describes instances where this conservatism is broken and new karyotypes have emerged or seem to be emerging. The cases include distance sex multivalents up to 7x + Y (Asphaera spp. and Aspicela scutata), a full series of autosomal centric fusions: 5 + X + y (Alagoasa equesrris), up to 17 B chromosomes (“Al. transparenre” and AI. oblecta), and erosion of achiasmate arms ( A / . januaria).
Niilo Virkki, Crop Protection Department, Agricultural Experiment Station, Uniuersiry of Puerto Rico, Rio Piedras 00928-1360 Puerto Rico Jorge A. Santiago-Blay, Department of Entomological Sciences, University of California, Berkeley, CA 94720-0001, U S A
The chromosome number and meioformula [2n = 22(X, y) and 10 + X + y, respectively] of neotropical Oedionychina are conservative (PETIT- PIERRE et al. 1988). The interspecific karyotype differences concern usually the length of the arms, especially of the free (non-chiasmate) arms of the bivalents, and the arms of Y chromosome (Virkki and Santiago-Blay, unpubl.).
We recently published a hypothetical cladogram of Oedionychina, suggesting some recent trends in their karyotype evolution (VIRKKI et al. 1991). We consider them to be relatively recent because they are few in number and possibly derived from the prevailing 10 + X + y of Oedionychina. They in- clude a full series of autosomal centric fusions in one species of Alagoasa, tentatively identified as AI. equestris, and a full series of autosomal centric fissions in Walterianella venusta, as well as distance sex multivalents (meioformulae 10 + nx + Y ; n from 1 to 7) in Aspicela scutata and in several species of Asphaera. Except for the chromosomal idiosyncrasies in Walterianella venusta, Asphaera clerica, and Asph. daniela (VIRKKI 1963, 1964, 1967, 1968), the cases herein reported have not been documented. We have studied the chromo- somes of all neotropical Oedionychina in our pos- session and now document these and most deviant
karyotypes of the putative subtribe ( BECHYNE and SPRINCLOVA D E BECHYNE 1966).
Materials and methods The material (Table 1) was fixed in the field (“sa- fari fixation”), as follows. The abdomens were either opened with sharp-pointed scissors and the beetles immersed in freshly mixed acetic acid: ethanol (1:3) or the fixative was injected into the abdomen. Thereafter, the beetles were completely immersed in the fixative. Next day, they were transferred to 70 YO ethanol, and, weeks later, in the laboratory, they were re-transferred to fresh 70 % ethanol and stored up to 28 years at 0°C. Excised testes were refixed shortly in Kahle-Smith ( 1 part 100 YO acetic acid: 3 parts formalin: 7.5 parts 95 YO ethanol) to reduce the cytoplasm’s brit- tleness, and squashed on albuminized slides. This method works well for the large spermatocytes of Oedionychina but cannot be recommended for species with smaller spermatocytes ( = most of Coleoptera). For proper fixation, it is vital that beetles are completely immersed in an abundant quantity of fixative.

264 N. VlRKKl AND J. A. SANTIAGO-BLAY Hereditas 119 (1993)
Table 1. Oedionychina with atypical karyotypes
Species No. of beetles Date Locality
Alagoasa eyuestris Fabricius
A/. januaria Bechyne AI. ohlecta Baly “AI. transparenre” Asphaera abbreuiara Fabricius Asph. ubendrothi Harold Asph. clerica (Erichson) I
Asph. daniela Bechyne’ Asph. limilata Harold’ Asph. pauperala Harold’
Asph. refle.xico1li.s Bechyne’ Asph. semifulua Jacoby Asphaera. sp. Ashp. near weyrauchi Bechyne Aspicela scurara Latreille
Omophoira personara llliger
9.11.68 10.11.68 13.11.68 2.1X.80-24.111.81 24.X11.65 9.11.68 16.1V.68 9.1V.68 6.VI.64 28.V1.64 5.XII.65 2.X11.65 5.X11.65 30.X1.65 16.1V.68 14.IX.80 I2.XII.65 I I JV.68
2.IX.80-23.111.8 I
Tripui, Minas Gerais, Brazil Saramenha, Minas Gerais, Brazil Soares, Minas Gerais, Brazil Rio Claro, Srio Paulo, Brazil Coroico, La Paz, Bolivia Tripui, Minas Gerais, Brazil La Mesa, Panama Santo Doming0 de 10s Colorados, Pichincha, Ecuador Serro-do-Navio, Amapa, Brazil Maracay, Aragua, Venezuela Pucallpa, Oreto, Peru Divisoria, Oreto, Peru Pucallpa, Oreto, Peru Tingo Maria, Huanuco, Peru El Valle, Panama, Panama Rio Claro, S5o Paulo, Brazil Limatambo, Cuzco, Peru Silante, Pichincha, km 63 road Quito to Esmeraldas,
Rio Claro, Srio Paulo, Brazil Ecuador
I “Omophoira clerica Erichson“. J . Bechynt det. ( V I R K K I 1967, 1970) ’ In V I R K K I (1968, 1970) ’ Listed in SMITH and V I R K K I (1978); PETITPIERRE et al. (1988)
Nomenclature and presentation of results follow VIRKKI et al. (1992). Most generic names of the present material begin with the letter “A.”. To distinguish the genera unequivocally in the text, we have used longer abbreviations, namely: Al. for Alagoasa, Asph. for Asphaera, and Aspi. for As- picela. As before, in a general presentation, the sex chromosomes are written X and Y, but a lower- case letter is used where we want to emphasize a smaller size of sex chromosomefs) in comparison to the orientation partner( s). Meiograms were made for all species in this study. For comparison, we have also included meiograms of Asphaem spp. already published (VIRKKI 1967, 1968, 1970) or
al. 1988). As condensation of the bivalents and, especially, of the univalent sex chromosomes pro- ceeds from PM I to late M I (phases used for meiograms), chromosomal size comparisons be- tween meiograms must be made with great caution. This is true also for those meiograms where the sex chromosomes and the autosomes had to be taken from different cells. The magnification of all rneiograms is 1334 x .
Most observations and photographies were made under phase contrast optics using Zeiss Pho- tomicroscopes I and I1 with Kodak Plus-X Pan 35 mm film. Some preparations were Ag- or C- banded, as described in VIRKKI (1983).
listed (SMITH and V l R K K l 1978; PETITPIERRE et
Identifications were made by comparing our specimens with identified material, frequently types, at the Bowditch Collection of Coleoptera of the Museum of Comparative Zoology, Harvard University (Cambridge, Massachusetts, USA). Owing to the difficulties of species-level taxonomy of neotropical alticines, most identifications are tentative. Since deforestation or development have shrouded areas where collections of the species mentioned here already extinct.
were made, many may be rare or
Results Distance sex multivalents and identification of their sex chromosomes in species of Asphaera and in Aspicela scutata
A coleopteran distance sex multivalent was first described in Asphaera daniela ( 10 + X + 6y) by VIRKKI (1968). Later on, the same meioformula was reported for Asph. t-album by VIDAL (1984), as well as meioformulae 10 + X + 2y (Fig. 78, in SMITH and VIRKKI 1978) and 10+X+4y in Asph. pauperata and Asph. reJexico[Iis, respec- tively, by VIRKKI (1970). The chromosome list of PETITPIERRE et al. (1988) adds four further cases: 10 + X + 3y and 9 + X + 7y in two undetermined
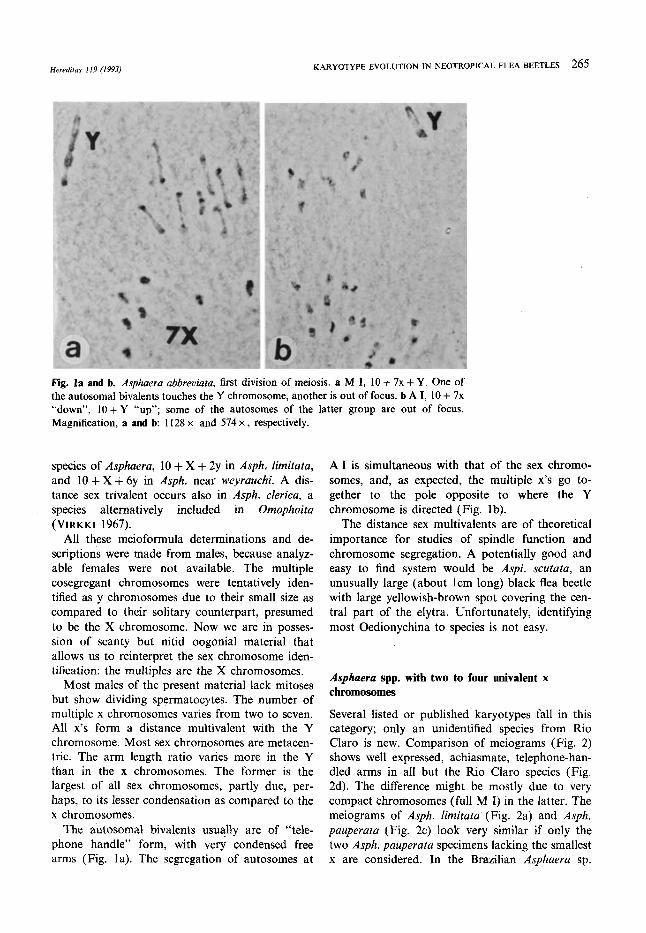
Hereditas 119 (1993) KARYOTYPE EVOLUTION IN NEOTROPICAL FLEA BEETLES 265
Fig. la and b. Asphaera ubbreviuru, first division of meiosis. a M I, 10 + 7x + Y. One of the autosomal bivalents touches the Y chromosome, another is out of focus. b A I, 10 + 7x “down”, lO+Y “up”; some of the autosomes of the latter group are out of focus. Magnification, a and b 1128 x and 574 x , respectively.
species of Asphaera, 10 + X + 2y in Asph. limitata, and 10 + X + 6y in Asph. near weyruuchi. A dis- tance sex trivalent occurs also in Asph. clerica, a species alternatively included in Omophoitu (VIRKKI 1967).
All these meioformula determinations and de- scriptions were made from males, because analyz- able females were not available. The multiple cosegregant chromosomes were tentatively iden- tified as y chromosomes due to their small size as compared to their solitary counterpart, presumed to be the X chromosome. Now we are in posses- sion of scanty but nitid oogonial material that allows us to reinterpret the sex chromosome iden- tification: the multiples are the X chromosomes.
Most males of the present material lack mitoses but show dividing spermatocytes. The number of multiple x chromosomes varies from two to seven. All x’s form a distance multivalent with the Y chromosome. Most sex chromosomes are metacen- tric. The arm length ratio varies more in the Y than in the x chromosomes. The former is the largest of all sex chromosomes, partly due, per- haps, to its lesser condensation as compared to the x chromosomes.
The autosomal bivalents usually are of “tele- phone handle” form, with very condensed free arms (Fig. la). The segregation of autosomes at
A I is simultaneous with that of the sex chromo- somes, and, as expected, the multiple x’s go to- gether to the pole opposite to where the Y chromosome is directed (Fig. lb).
The distance sex multivalents are of theoretical importance for studies of spindle function and chromosome segregation. A potentially good and easy to find system would be Aspi. scutata, an unusually large (about lcm long) black flea beetle with large yellowish-brown spot covering the cen- tral part of the elytra. Unfortunately, identifying most Oedionychina to species is not easy.
Asphaera spp. with two to four univalent x chromosomes
Several listed or published karyotypes fall in this category; only an unidentified species from Rio Claro is new. Comparison of meiograms (Fig. 2) shows well expressed, achiasmate, telephone-han- dled arms in all but the Rio Claro species (Fig. 2d). The difference might be mostly due to very compact chromosomes (full M I) in the latter. The meiograms of Asph. limitatu (Fig. 2a) and Asph. pauperutu (Fig. 2c) look very similar if only the two Asph. pauperata specimens lacking the smallest x are considered. In the Brazilian Asphaera sp.

266 N VIRKKI AND J. A. SANTIAGO-BLAY Hereditas 119 (1993)
Fig. 2a-e. Meiograms of four Asphaera spp. M I (a, c to e); PM I (b). a Asph. limifata from Pucallpa, Perk 10 + 2x + Y. b Asph. clerica from Sero-do-Navio, Brazil 7+2x+Y, with one autosomal ring bivalent. e Asph. pauperafa from Divisoria, Peru. 10 + 3x + Y and 10 + 2x + Y (the smallest x is not present in all specimens). d Asphaera. sp. from Rio Claro, Brazil. 10 + 3x + Y. e Asph. rejlexicol- lis from Tingo Maria, Peru. 10 + B + 4x + Y .
(Fig. 2d) and the Peruvian Asph. reJexicollis (Fig. 2e), each of the x chromosomes is well defined by size and centromere position. The x chromosomes differ most between the species in number, relative size, and arm length ratio. All Y chromosomes are roughly J-shaped, although the arm length ratio seems to differ. The Asph. pauperata meiogram
showing 3x (Fig. 2c) approximates that of As- phaera sp. (Figs. 2d, 3a) in number although not in arm length ratio of the sex chromosomes. The only acrocentric x was found in Asph. clerica (Fig. 2b), which also shows, like Asph. octopunctata and “Asph. pauperata” (SMITH and VIRKKI 1978; PE- TITPIERRE et al. 1988), one or two ring bivalents,
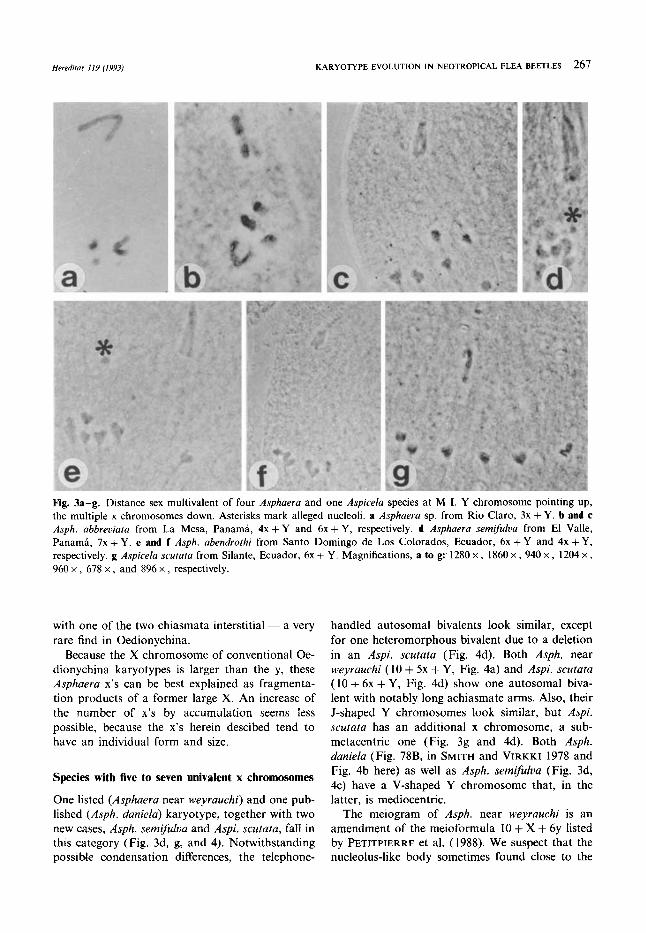
Herediras 119 (1993) KARYOTYPE EVOLUTION I N NEOTROPICAL FLEA BEETLES 267
Fig. 3a-g. Distance sex multivalent of four Asphaera and one Aspieela species at M I. Y chromosome pointing up, the multiple x chromosomes down. Asterisks mark alleged nucleoli. a Asphaera sp. from Rio Claro, 3x + Y. b and c Asph. abbreviata from La Mesa, Panama, 4x + Y and 6x + Y, respectively. d Asphaera semifulva from El Valle, Panama, 7x + Y. e and f Asph. abendrothi from Santo Doming0 de Los Colorados, Ecuador, 6x + Y and 4x + Y, respectively. g Aspieela seutata from Silante, Ecuador, 6x + Y. Magnifications, a to g: 1280 x , 1860 x , 940 x , 1204 x , 960 x , 678 x , and 896 x , respectively.
with one of the two chiasmata interstitial - a very rare find in Oedionychina.
Because the X chromosome of conventional Oe- dionychina karyotypes is larger than the y, these Asphaera x’s can be best explained as fragmenta- tion products of a former large X. An increase of the number of x’s by accumulation seems less possible, because the x’s herein descibed tend to have an individual form and size.
Species with five to seven univalent x chromosomes
One listed (Asphaera near weyrauchi) and one pub- lished (Asph. daniela) karyotype, together with two new cases, Asph. sem$ulva and Aspi. scutata, fall in this category (Fig. 3d, g, and 4). Notwithstanding possible condensation differences, the telephone-
handled autosomal bivalents look similar, except for one heteromorphous bivalent due to a deletion in an Aspi. scututa (Fig. 4d). Both Asph. near weyrauchi ( 10 + 5x + Y, Fig. 4a) and Aspi. scutata (10 + 6x + Y, Fig. 4d) show one autosomal biva- lent with notably long achiasmate arms. Also, their J-shaped Y chromosomes look similar, but Aspi. scurata has an additional x chromosome, a sub- metacentric one (Fig. 3g and 4d). Both Asph. daniela (Fig. 78B, in SMITH and VIRKKI 1978 and Fig. 4b here) as well as Asph. semifulva (Fig. 3d, 4c) have a V-shaped Y chromosome that, in the latter, is mediocentric.
The meiogram of Asph. near weyrauchi is an amendment of the meioformula 10 + X + 6y listed by PETITPIERRE et al. (1988). We suspect that the nucleolus-like body sometimes found close to the
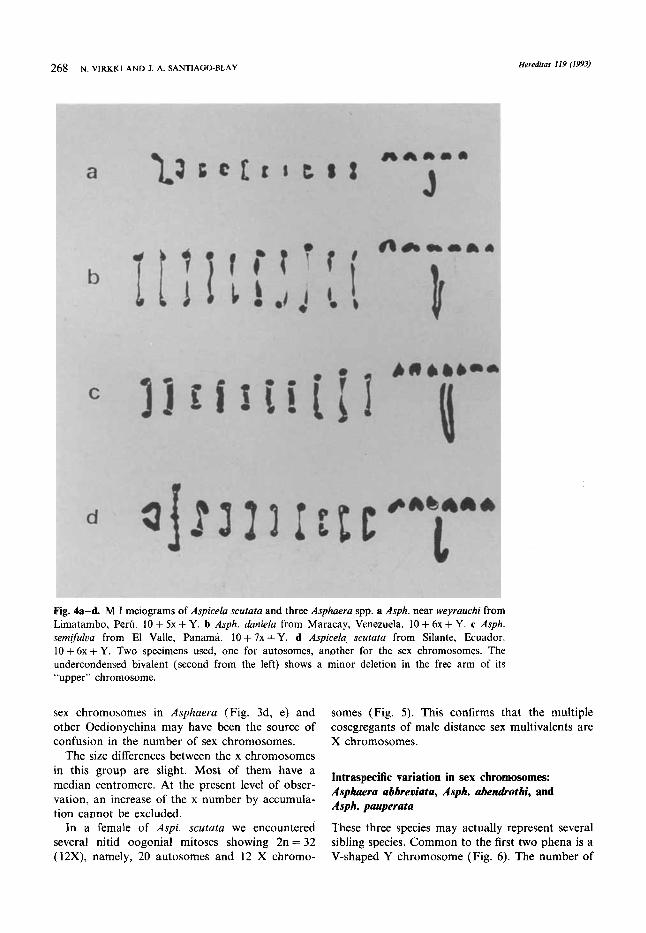
268 N. VIRKKI AND I. A. SANTIAGO-BLAY Herediros 119 (1993)
Fig. 4a-d. M I meiograms of Aspicela scutafa and three Asphaera spp. a Asph. near weyrauchi from Limatambo, Peru. 10 + 5x + Y. b Asph. duniela from Maracay, Venezuela. 10 + 6x + Y. c Asph. semifuluu from El Valle, Panama. 10+7x+Y. d Aspicela scufata from Silante, Ecuador. 10 + 6x + Y. Two specimens used, one for autosomes, another for the sex chromosomes. The undercondensed bivalent (second from the left) shows a minor deletion in the free arm of its “upper” chromosome.
sex chromosomes in Asphaera (Fig. 3d, e) and other Oedionychina may have been the source of confusion in the number of sex chromosomes.
The size differences between the x chromosomes in this group are slight. Most of them have a median centromere. At the present level of obser- vation, an increase of the x number by accumuta- tion cannot be excluded.
In a female of Aspi. scutata we encountered several nitid oogonial mitoses showing 2n = 32 (12X), namely, 20 autosomes and 12 X chromo-
somes (Fig. 5). This confirms that the multiple cosegregants of male distance sex multivalents are X chromosomes.
Intraspecific variation in sex chromosomes: Asphaera abbreviaru, Asph. abendrothi, and Asph. pauperata
These three species may actually represent several sibling species. Common to the first two phena is a V-shaped Y chromosome (Fig. 6). The number of
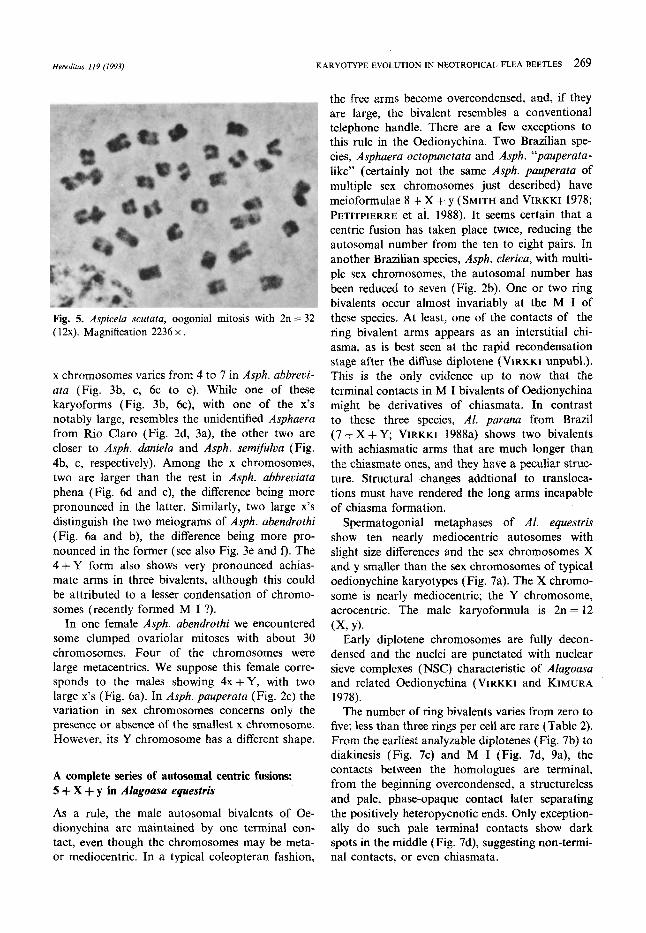
Hereditas 119 (1993) KARYOTYPE EVOLUTION IN NEOTROPICAL FLEA BEETLES 269
Fig. 5. Aspicela scutata, oogonial mitosis with 2n = 32 (12x). Magnification 2236 x .
x chromosomes varies from 4 to 7 in Asph. abbrevi- ata (Fig. 3b, c, 6c to e). While one of these karyoforms (Fig. 3b, 6c), with one of the x’s notably large, resembles the unidentified Asphaera from Rio Claro (Fig. 2d, 3a), the other two are closer to Asph. daniela and Asph. semifulua (Fig. 4b, c, respectively). Among the x chromosomes, two are larger than the rest in Asph. abbreuiata phena (Fig. 6d and e), the difference being more pronounced in the latter. Similarly, two large x’s distinguish the two meiograms of Asph. abendrothi (Fig. 6a and b), the difference being more pro- nounced in the former (see also Fig. 3e and f ) . The 4 + Y form also shows very pronounced achias- mate arms in three bivalents, although this could be attributed to a lesser condensation of chromo- somes (recently formed M I ?).
In one female Asph. abendrothi we encountered some clumped ovariolar mitoses with about 30 chromosomes. Four of the chromosomes were large metacentrics. We suppose this female corre- sponds to the males showing 4x + Y, with two large x’s (Fig. 6a). In Asph. pauperata (Fig. 2c) the variation in sex chromosomes concerns only the presence or absence of the smallest x chromosome. However, its Y chromosome has a different shape.
A complete series of autosomal centric fusions: 5 + X + y in Alagoasa equestris
As a rule, the male autosomal bivalents of Oe- dionychina are maintained by one terminal con- tact, even though the chromosomes may be meta- or mediocentric. In a typical coleopteran fashion,
the free arms become overcondensed, and, if they are large, the bivalent resembles a conventional telephone handle. There are a few exceptions to this rule in the Oedionychina. Two Brazilian spe- cies, Asphaera octopunctata and Asph. “pauperata- like” (certainly not the same Asph. pauperata of multiple sex chromosomes just described) have meioformulae 8 + X + y (SMITH and VIRKKI 1978; PETITPIERRE et ai. 1988). It seems certain that a centric fusion has taken place twice, reducing the autosomal number from the ten to eight pairs. In another Brazilian species, Asph. clerica, with multi- ple sex chromosomes, the autosomal number has been reduced to seven (Fig. 2b). One or two ring bivalents occur almost invariably at the M I of these species. At least, one of the contacts of the ring bivalent arms appears as an interstitial chi- asma, as is best seen at the rapid recondensation stage after the diffuse diplotene (VIRKKI unpubl.). This is the only evidence up to now that the terminal contacts in M I bivalents of Oedionychina might be derivatives of chiasmata. In contrast to these three species, Al. parana from Brazil (7 + X + Y; VIRKKI 1988a) shows two bivalents with achiasmatic arms that are much longer than the chiasmate ones, and they have a peculiar struc- ture. Structural changes addtional to transloca- tions must have rendered the long arms incapable of chiasma formation.
Spermatogonial metaphases of Al. equestris show ten nearly mediocentric autosomes with slight size differences and the sex chromosomes X and y smaller than the sex chromosomes of typical oedionychine karyotypes (Fig. 7a). The X chromo- some is nearly mediocentric; the Y chromosome, acrocentric. The male karyoformula is 2n = 12
Early diplotene chromosomes are fully decon- densed and the nuclei are punctated with nuclear sieve complexes (NSC) characteristic of Alagoasa and related Oedionychina (VIRKKI and KIMURA 1978).
The number of ring bivalents varies from zero to five; less than three rings per cell are rare (Table 2). From the earliest analyzable diplotenes (Fig. 7b) to diakinesis (Fig. 7c) and M I (Fig. 7d, 9a), the contacts between the homologues are terminal, from the beginning overcondensed, a structureless and pale, phase-opaque contact later separating the positively heteropycnotic ends. Only exception- ally do such pale terminal contacts show dark spots in the middle (Fig. 7d), suggesting non-termi- nal contacts, or even chiasmata.
(X, Y>.

270 N. VIRKKI AND J. A. SANTIAGO-BLAY
Fig. 6a-e. M I meiograms showing variation in the X chromosome number of two Asphaera spp., Asph. abendrothi from Santo Doming0 de 10s Colorados, Ecuador, and Asph. abbreviata from La Mesa, Panami. a Asph. abendrothi. 10 + 4x + Y. Two cells of the same specimen used, one for autosomes, another for the sex chromosomes. b Asph. abendrothi, another specimen. 10 + 6x + Y. Three cells were used, one for autosomes, and one each for the 6x and Y. c Asph. abbreviata. 10+4x+Y. d Asph. abbreviata. 10+6x+Y. Two specimens were used, one for autosomes, another for the sex chromosomes. e Asph. abbreviata. 10 + 7x + Y; still another specimen.
Hereditas 119 (1993)

Hereditas 119 (1993) KARYOTYPE EVOLUTION IN NEOTROPICAL FLEA BEETLES 27 1
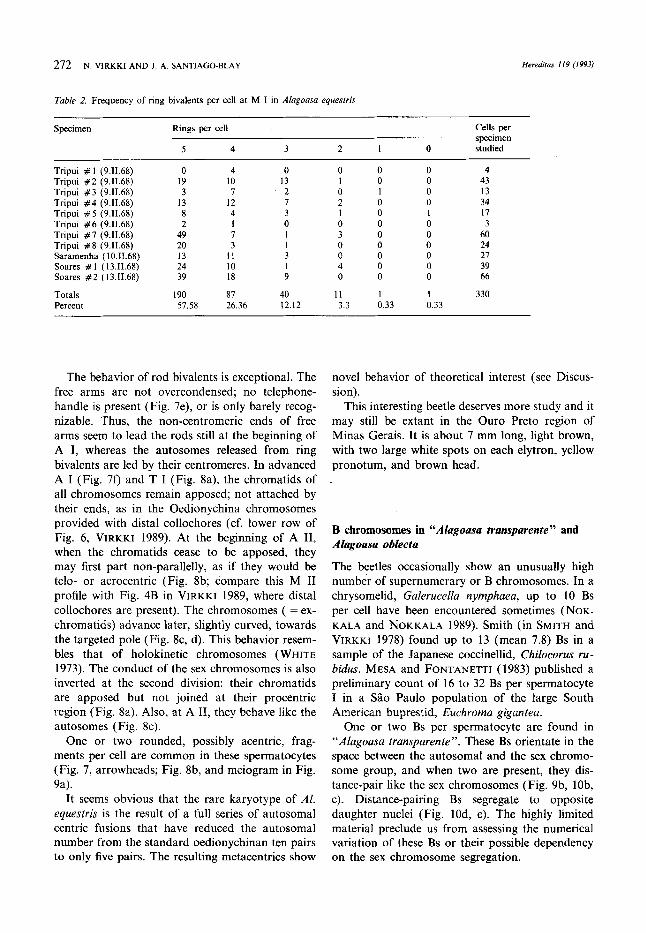
Fig. 7a-f. Alagoasa equesfris (Tripui, Brazil) chromosomes in spermatogenesis. Arrowheads point to possible achiasmatic fragments. a Spermatogonial mitosis, 2n = 12(X, yj. b Diplotene. The two ring bivalents show terminal contact of slightly heteropycnotic ends. c Diakinesis, 5 + X + y; all autosomal bivalents look rings maintained by terminal contacts. d M I, 5 + X + y, extended terminal contacts of the five ring bivalents. Asterisk highlights the leftmost bivalent in higher magnification to show dark spots in the middle of the extended terminal contacts. e Beginning of A I. Note that the free arms of the unichiasmate rod bivalent are not much bent nor overcondensed. f Late A I, apposition of chromatids in all chromosomes. Magnifications, a to E 1344 x , 294 x , 693 x , 670 x (insert, 1678 x j, 770 x , and 608 x , respectively.
Heredifas 119 (1993) 272 N. VIRKKI AND J. A. SANTIAGO-BLAY
Table 2. Frequency of ring bivalents per cell at M I in Alagoasa equeslris
Specimen Rings per cell Cells per specimen
5 4 3 2 I 0 studied
Tripui # 1 (9.11.68) Tripui # 2 (9.11.68) Tripui # 3 (9.11.68) Tripui # 4 (9.11.68) Tripui # 5 (9.11.68) Tripui # 6 (9.11.68) Tripui # 7 (9.11.68) Tripui #8 (9.11.68) Saramenha (10.11.68) Soares # I (13.11.68) Soares # 2 (13.11.68)
Totals Percent
0 4 0 19 10 13 3 7 2
13 12 7 8 4 3 2 1 0
49 7 I 20 3 I 13 11 3 24 10 1 39 18 9
190 87 40 57.58 26.36 12.12
0 0 0 1 0 0 0 1 0 2 0 0 1 0 1 0 0 0 3 0 0 0 0 0 0 0 0 4 0 0 0 0 0
11 1 1 3.3 0.33 0.33
4 43 13 34 17 3
60 24 27 39 66
330
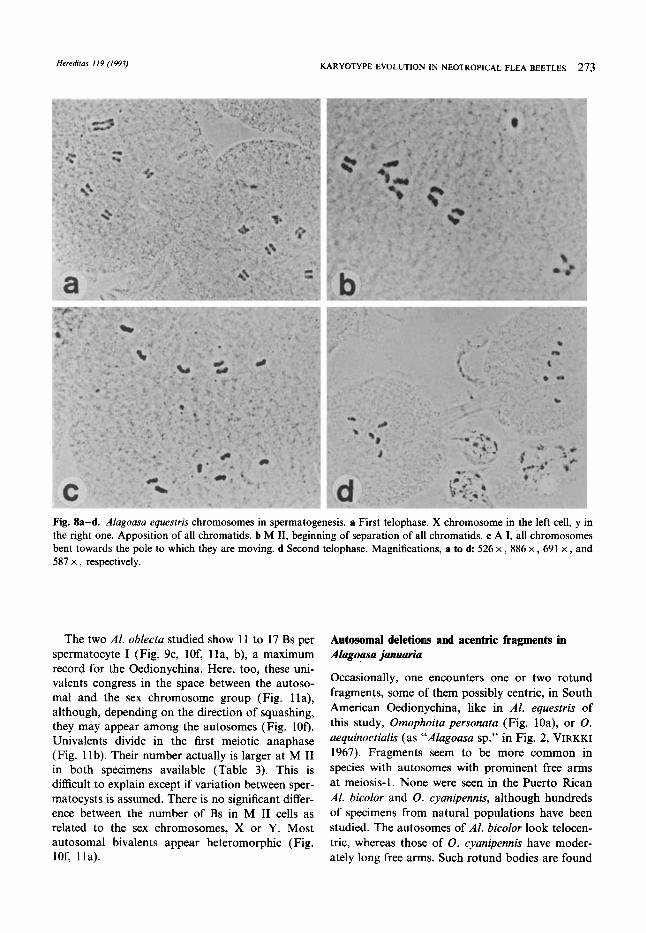
The behavior of rod bivalents is exceptional. The free arms are not overcondensed; no telephone- handle is present (Fig. 7e), or is only barely recog- nizable. Thus, the non-centromeric ends of free arms seem to lead the rods still at the beginning of A I, whereas the autosomes released from ring bivalents are led by their centromeres. In advanced A I (Fig. 7f) and T I (Fig. 8a), the chromatids of all chromosomes remain apposed; not attached by their ends, as in the Oedionychina chromosomes provided with distal collochores (cf. lower row of Fig. 6, VlRKKI 1989). At the beginning of A 11, when the chromatids cease to be apposed, they may first part non-parallelly, as if they would be telo- or acrocentric (Fig. 8b; compare this M I1 profile with Fig. 4B in VIRKKI 1989, where distal collochores are present). The chromosomes ( = ex- chromatids) advance later, slightly curved, towards the targeted pole (Fig. 8c, d). This behavior resem- bles that of holokinetic chromosomes (WHITE 1973). The conduct of the sex chromosomes is also inverted at the second division: their chromatids are apposed but not joined at their procentric region (Fig. 8a). Also, at A 11, they behave like the autosomes (Fig. 8c).
One or two rounded, possibly acentric, frag- ments per cell are common in these spermatocytes (Fig. 7, arrowheads; Fig. 8b, and meiogram in Fig. 9a).
It seems obvious that the rare karyotype of A1 equestris is the result of a full series of autosomal centric fusions that have reduced the autosomal number from the standard oedionychinan ten pairs to only five pairs. The resulting metacentrics show
novel behavior of theoretical interest (see Discus- sion).
This interesting beetle deserves more study and it may still be extant in the Our0 Preto region of Minas Gerais. It is about 7 mm long, light brown, with two large white spots on each elytron, yellow pronotum, and brown head.
B chromosomes in “Alugousu tvanspavente” and Alagoasa oblecta
The beetles occasionally show an unusually high number of supernumerary or B chromosomes. In a chrysomelid, Galerucella nymphaea, up to 10 Bs per cell have been encountered sometimes ( NOK- KALA and NOKKALA 1989). Smith (in SMITH and VIRKKI 1978) found up to 13 (mean 7.8) Bs in a sample of the Japanese coccinellid, Chilocorus ru- bidus. MESA and FONTANETTI (1983) published a preliminary count of 16 to 32 Bs per spermatocyte I in a SBo Paulo population of the large South American buprestid, Euchvoma gigantea.
One or two Bs per spermatocyte are found in “Alagousu trunsparente”. These Bs orientate in the space between the autosomal and the sex chromo- some group, and when two are present, they dis- tance-pair like the sex chromosomes (Fig. 9b, lob, c). Distance-pairing Bs segregate to opposite daughter nuclei (Fig. 10d, e). The highly limited material preclude us from assessing the numerical variation of these Bs or their possible dependency on the sex chromosome segregation.
Hereditas 119 (1993) KARYOTYPE EVOLUTION IN NEOTROPICAL FLEA BEETLES 273
Fig. 8a-d. AIugousu equesfris chromosomes in spermatogenesis. a First telophase. X chromosome in the left cell, y in the right one. Apposition of all chromatids. b M 11, beginning of separation of all chromatids. c A I, all chromosomes bent towards the pole to which they are moving. d Second telophase. Magnifications, a to d 526 x , 886 x , 691 x , and 587 x , respectively.
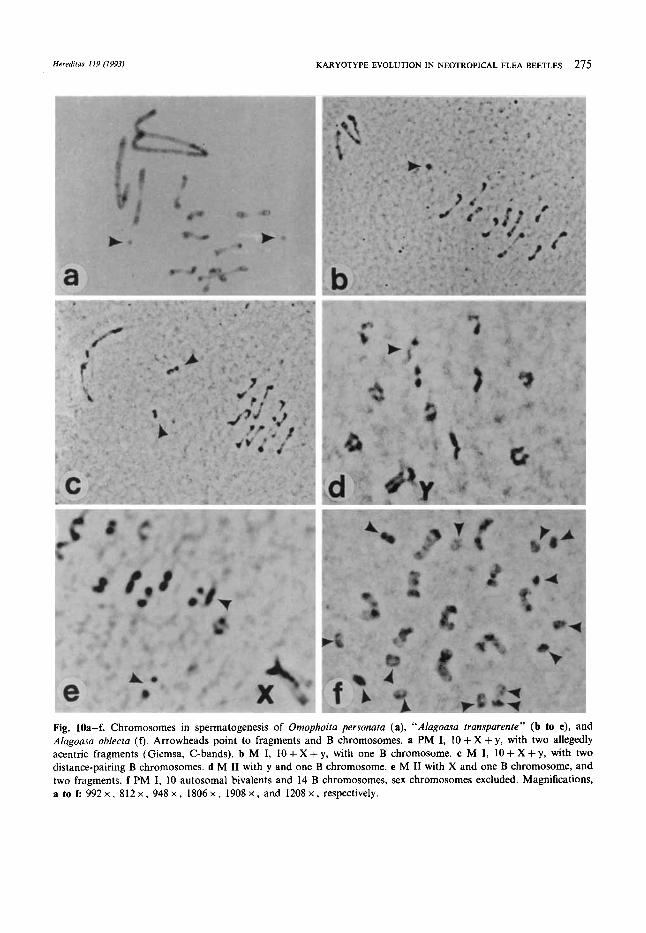
The two Al. oblecta studied show 11 to 17 Bs per spermatocyte I (Fig. 9c, lOf, l l a , b), a maximum record for the Oedionychina. Here, too, these uni- valents congress in the space between the autoso- ma1 and the sex chromosome group (Fig. lla), although, depending on the direction of squashing, they may appear among the autosomes (Fig. 100. Univalents divide in the first meiotic anaphase (Fig. 1 lb). Their number actually is larger at M I1 in both specimens available (Table 3). This is difficult to explain except if variation between sper- matocysts is assumed. There is no significant differ- ence between the number of Bs in M I1 cells as related to the sex chromosomes, X or Y. Most autosomal bivalents appear heteromorphic (Fig. lOf, lla).
Autosomal deletions and acentric fragments in Alagoasa januaria
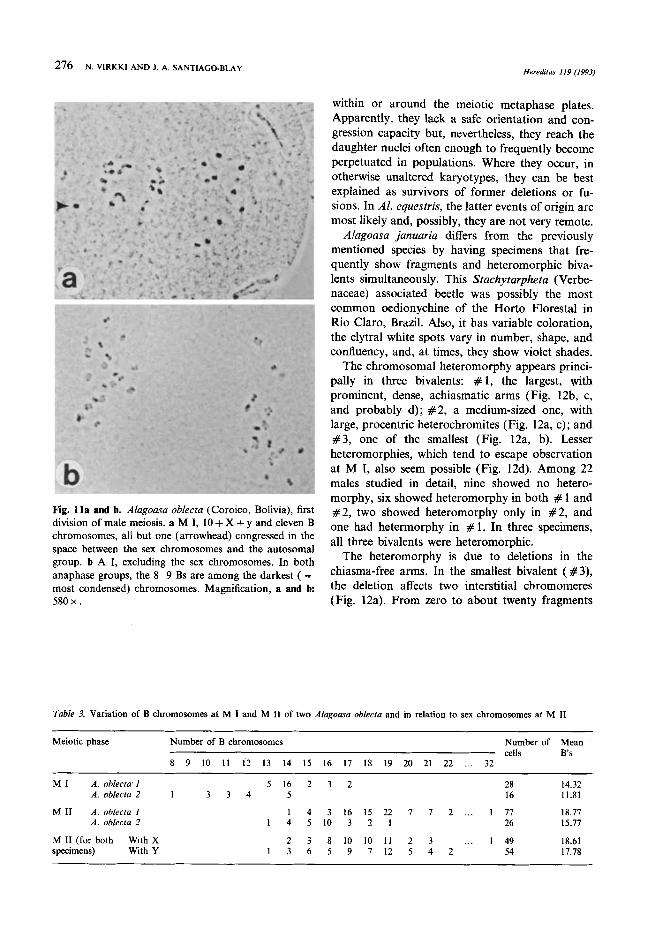
Occasionally, one encounters one or two rotund fragments, some of them possibly centric, in South American Oedionychina, like in AI. equestris of this study, Omophoita personata (Fig. lOa), or 0. aequinoctialis (as “Alagoasa sp.” in Fig. 2, VIRKKI 1967). Fragments seem to be more common in species with autosomes with prominent free arms at meiosis-1. None were seen in the Puerto Rican AI. bicolor and 0. cyanipennis, although hundreds of specimens from natural populations have been studied. The autosomes of Al. bicolor look telocen- tric, whereas those of 0. cyanipennis have moder- ately long free arms. Such rotund bodies are found

214 N. VIRKKI AND J. A. SANTIAGO-BLAY Hereditas 119 (1993)
Fig. 9a-d Meiograms in four Alagoasu spp. with altered karyotypes. M I (a to c); PM I (d). a AI. equestris from Soares, Brazil. 5 + 1B + X + y. b “ A l . transparente” from Tripui, Brazil. 10 + 2B + X + y. Autosomes, Bs, and sex chromosomes, respectively, from three different cells of the same individuum. c Al . oblecta from Coroico, Bolivia. 10 + 15B + X + y. d AI. januuvia from Rio Claro, Brazil. 10 + X + y. This meiogram shows only one hetermorphous bivalent (no. 2, first from the left), and no fragments.
Herediras 119 (1993) KARYOTYPE EVOLUTION IN NEOTROPICAL FLEA BEETLES 275
Fig. 10a-f. Chromosomes in spermatogenesis of Omophoira personata (a), “Alagoasa transparente” (b to e), and Alagoasa oblecta (9. Arrowheads point to fragments and B chromosomes. a PM I, 10 + X + y, with two allegedly acentric fragments (Giemsa, C-bands). b M I, 10 + X + y, with one B chromosome. c M I , 10 + X + y, with two distance-pairing B chromosomes. d M I1 with y and one B chromosome. e M I1 with X and one B chromosome, and two fragments. f PM I, 10 autosomal bivalents and 14 B chromosomes, sex chromosomes excluded. Magnifications, a to fi 992 x , 812 x , 948 x , 1806 x , 1908 x , and 1208 x , respectively.
276 N. VIRKKI AND J. A. SANTIAGO-BLAY Herediras 119 (1993)
Fig. l l a and b. Alugousu oblecta (Coroico, Bolivia), first division of male meiosis. a M I, 10 + X + y and eleven B chromosomes, all but one (arrowhead) congressed in the space between the sex chromosomes and the autosomal group. b A I, excluding the sex chromosomes. In both anaphase groups, the 8-9 Bs are among the darkest ( = most condensed) chromosomes. Magnification, a and b 580 x .
within or around the meiotic metaphase plates. Apparently, they lack a safe orientation and con- gression capacity but, nevertheless, they reach the daughter nuclei often enough to frequently become perpetuated in populations. Where they occur, in otherwise unaltered karyotypes, they can be best explained as survivors of former deletions or fu- sions. In Al. equestris, the latter events of origin are most likely and, possibly, they are not very remote.
Alagoasa januaria differs from the previously mentioned species by having specimens that fre- quently show fragments and heteromorphic biva- lents simultaneously. This Stuchytarpheta (Verbe- naceae) associated beetle was possibly the most common oedionychine of the Horto Florestal in Rio Claro, Brazil. Also, it has variable coloration, the elytral white spots vary in number, shape, and contluency, and, at times, they show violet shades.
The chromosomal heteromorphy appears princi- pally in three bivalents: #1, the largest, with prominent, dense, achiasmatic arms (Fig. 12b, c, and probably d); #2, a medium-sized one, with large, procentric heterochromites (Fig. 12a, c); and #3, one of the smallest (Fig. 12a, b). Lesser heteromorphies, which tend to escape observation at M I, also seem possible (Fig. 12d). Among 22 males studied in detail, nine showed no hetero- morphy, six showed heteromorphy in both # 1 and #2, two showed heteromorphy only in #2, and one had hetermorphy in # 1. In three specimens, all three bivalents were heteromorphic.
The heteromorphy is due to deletions in the chiasma-free arms. In the smallest bivalent ( # 3), the deletion affects two interstitial chromomeres (Fig. 12a). From zero to about twenty fragments
Tabie 3. Variation of B chromosomes at M I and M 11 of two Ahgoma oblecta and in relation to sex chromosomes at M I1
Meiotic phase Number of B chromosomes Number of Mean cells B’s
8 9 10 I I 12 13 14 15 16 17 18 19 20 21 22 _.. 32
M I A . obiecra- 1 5 1 6 2 3 2 28 14.32 A . oblecra 2 1 3 3 4 5 16 11.81
M I1 A . oblecta I A . oblectu 2
M I1 (for both With X specimens) With Y
1 4 3 16 15 22 7 7 2 ... 1 4 5 1 0 3 2 I
2 3 8 1 0 1 0 1 1 2 3 ... 1 3 6 5 9 7 1 2 5 4 2
1 77 18.77 26 15.77
1 49 18.61 54 17.78
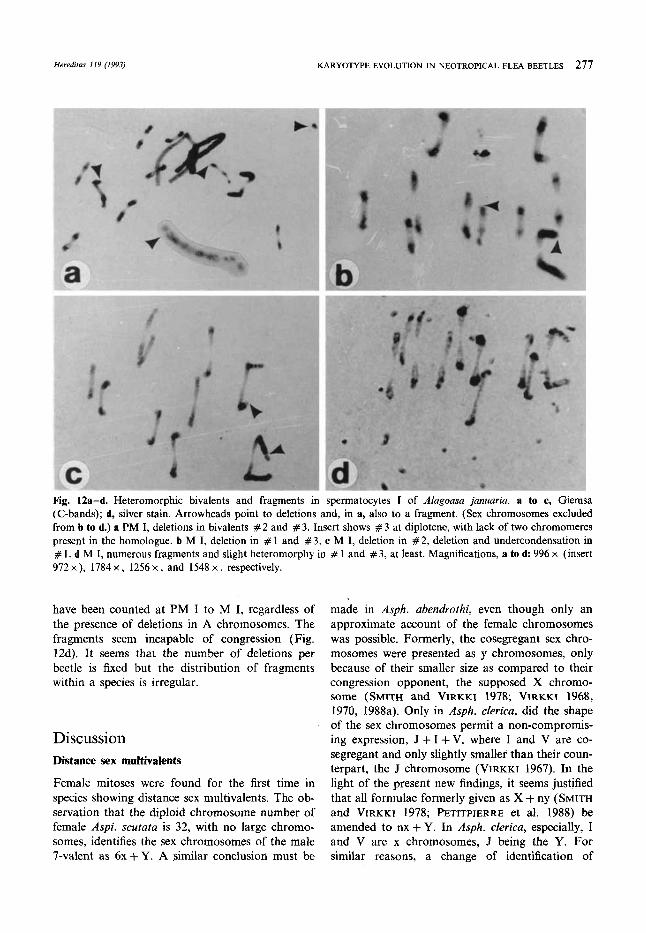
Heredifas 119 (1993) KARYOTYPE EVOLUTION IN NEOTROPICAL FLEA BEETLES 277
Fig. 12a-d. Heteromorphic bivalents and fragments in spermatocytes I of AIagoasa januaria. a to c, Giemsa (C-bands); d, silver stain. Arrowheads point to deletions and, in a, also to a fragment. (Sex chromosomes excluded from b to d.) a PM I, deletions in bivalents # 2 and # 3. Insert shows # 3 at diplotene, with lack of two chromomeres present in the homologue. b M I, deletion in # 1 and # 3. c M I, deletion in # 2, deletion and undercondensation in # 1. d M I, numerous fragments and slight heteromorphy in # 1 and #3 , at least. Magnifications, a to d: 996 x (insert 972 x ), 1784 x , 1256 x , and 1548 x , respectively.
have been counted at PM I to M I, regardless of the presence of deletions in A chromosomes. The fragments seem incapable of congression (Fig. 12d). It seems that the number of deletions per beetle is fixed but the distribution of fragments within a species is irregular.
Discussion Distance sex multivalents
Female mitoses were found for the first time in species showing distance sex multivalents. The ob- servation that the diploid chromosome number of female Aspi. scutata is 32, with no large chromo- somes, identifies the sex chromosomes of the male 7-valent as 6x + Y. A similar conclusion must be
made in Asph. abendrothi, even though only an approximate account of the female chromosomes was possible. Formerly, the cosegregant sex chro- mosomes were presented as y chromosomes, only because of their smaller size as compared to their congression opponent, the supposed X chromo- some (SMITH and VIRKKI 1978; VIRKKI 1968, 1970, 1988a). Only in Asph. clerica, did the shape of the sex chromosomes permit a non-compromis- ing expression, J + I + V, where I and V are co- segregant and only slightly smaller than their coun- terpart, the J chromosome (VIRKKI 1967). In the light of the present new findings, it seems justified that all formulae formerly given as X + ny (SMITH and VIRKKI 1978; PETITPIERRE et al. 1988) be amended to nx + Y. In Asph. clerica, especially, I and V are x chromosomes, J being the Y. For similar reasons, a change of identification of

278 N. VIRKKI AND J. A. SANTIAGO-BLAY Hereditas 119 (1993)
Disonychina sex chromosomes has been already made (VIRKKI 1988b).
Although the lack of sufficient mitoses does not allow us to compose significant karyograms, com- parison of dividing spermatocytes convinced us of two general trends. First, the size of multiple X chromosomes tends to be the smaller as their num- ber increases (in addition to the present illustra- tion, see also Fig. 78 in SMITH and VIRKKI 1978). Second, the X chromosomes of an alleged species (or species within it) are not always equal in num- ber, size, and shape. Thus, it seems possible that the multiple X chromosomes arise through disinte- gration of a former large X into pieces, with doubtful accumulation of such pieces.
The origin of the new centromeres constitutes a problem in this cytoevolutionary scenario. Auto- some numbers remain unaltered (except for Asph. clerica (VIRKKI 1967), a couple of other Asphaera spp. (PETITPIERRE et al. 1988), and B chromosome “donors” are seldom present. Two feasible possibil- ities remain: centric fissions or the fragmentation and activation of potential “centrophilic” chro- momeres hidden in the arms of the original large X chromosomes. If the formation of new centromeres occurs by centric fissions, then we face the same problem most beetle cytologists already know from autosomes: the supposed fission products are meta- centric. Because their arms are often unequal, and because an interarm pairing has never been re- ported, isochromosome formation, as a first step following fission (WU 1992), seems unlikely. Some immediate addition of chromatin to fission telocen- trics does occur, according to in situ hybridization studies (SCHUBERT et al. 1992), but a more exten- sive accretion of chromatin is necessary to explain this neo-metacentry. The favored hypothesis among beetle cytologists is that centric fissions are soon followed by pencentric inversions. Studies to follow the change of DNA-amounts in these cases are necessary but still lacking. Postfission restoration of metacentry allows new centric fissions to occur and, thus, further increase the X chromosome number.
Fission X chromosomes formed by accretion of a second arm may be inert and dispensable, or the original large X may have had genetically inert segments. Variation in the number of the fission Xs in a species would thus be comprehensible. We cannot exclude, however, the possibility of sibling species, each one with a different set of X chromo- somes. Even in that case, the repeated rearrange- ment of X chromosomes seems to be a rapid evolutionary process.
The distance sex multivalents are now known to occur in two closely related genera, Asphaera and Aspicela. Most likely the trend of splitting X chro- mosomes started in a common ancestor of these genera, and it may be present in some other, still cytologically unknown, related genera ( SEENO and WILCOX 1982, p. 140). The fact that Asphaera still includes conventional 10 + X + y karyotypes (PE- TITPIERRE et al. 1988) warrants a taxonomic revi- sion of the genus.
Former speculation (SMITH and VIRKKI 1978, p. 153) that Asphaera might be a primitive Oe- dionychina with a tendency of fusion of Disonychina-type X univalents, and a failure of pairing of a Disonychina-type Xy bivalent, is un- likely because their repeated fusion would increase only one of the sex chromosomes. Also exopheno- typically, Asphaera is closer to the rest of the Oedionychina than to the Disonychina. We con- sider Asphaera and Aspicela to be relatively apo- morphic oedionychine genera.
Like distance sex bivalents (VIRKKI 1972, 1990), the distance sex multivalents arise on the site of the clump, staying there when the autosomal bivalents leave to-congress at the equatorial plate. Certainly, there are casual contacts of sex multiples during the tightest clump stage but such contacts hardly have anything to do with the final orientation of the sex chromosomes (contra VIRKKI 1968). The multiple Xs somehow “know” to co-orientate to- wards the same pole, opposite to the pole to which the Y chromosome becomes orientated. Whether proper orientation of multiple sex chromosomes is related to competition of centromeres of different assembly capacity for the microtubule precursor available in each hemisphere of the “sex spindle,” as originally speculated (VIRKKI 1967), or do the nx vs. Y centromeres recognize and insert to microtubules polarized to opposite poles (Eu-
MCDONALD 1989), is a matter still unknown. TENEUER and MCINTOSH 1981; MCINTOSH and
Effect of centric fusion on meiotic behavior of autosomes
The fusion autosomes of Al. equestris look very similar since, apparently, the repeated fusions have occurred in a rapid succession, and no additional rearrangements such as pericentric inversions, have occurred yet. It is interesting to compare such a karyotype and its meiotic behavior with the con- ventional 10 + X + y karyotypes of Oedionychina.

Hereditas 119 (1993) KARYOTYPE EVOLUTION I N NEOTROPICAL FLEA BEETLES 279
Another unusual feature of Al. equestris is the relatively small size of its sex chromosomes. We know a few similar 10 + X + y karyotypes of Brazilian Alagoasa spp. (Virkki and Santiago-Blay, unpubl.), possibly close relatives of AI. equestris. Their autosomal bivalents have notable free arms, as indeed do most bivalents of Oedionychina. Thus, there is good reason to assume that the pre-fusion autosomes of the Al. equestris ancestor have been typical metacentrics that associate in the “telephone handle” fashion. Centric fusion of such metacentrics is possible. If the fusing metacentrics are diphasic, as in the coccinellid genus Chilocorus (see p. 281), these fusions may produce two new metacentrics, viz.: one euchromatic and indispens- able; the other inert and dispensable. The inert and dispensable become B chromosomes, whose ulti- mate fate depends much on their resistance to chromosomal erosion and/or on drive mechanisms that may favor them (SMITH and VIRKKI 1978; JONES and REES 1982). We have indeed discovered two such fragments or chromosomes of poor congression capability in our two AI. equestris specimens. It seems possible, although not yet statistically supported, that, contrary to Ch. stigma, the fusion Bs are rapidly eliminated from Al. equestris karyotypes.
Although one arm-to-arm association suffices to maintain the fusion bivalents, both arm pairs ap- pear associated in most Al. equestris bivalents. This pattern of persistence of the association of the euchromatic arms of the allegedly diphasic pre-fu- sion autosomes, is just as SMITH (1966, 1970) found in the fusion metacentrics of Chilocorus (Coccinellidae) and Pissodes (Curculionidae). Thus, there is little need to assume an extreme chiasma interference across the centromere in the fusion metacentrics. The double association seems to occur between precondensed distal segments, with no indication of chiasmata, except perhaps, the dark spots sometimes seen in the middle of extended terminal contacts of M I rings. Even in such extreme cases, the presence of a chiasma is not excluded as a possibility (JONES 1987).
Calculated from Table 3, 209 (12.7 %) of the observed 1650 autosomal bivalents had a single end-to-end association. This would correspond to a notable univalent frequency in conventional 10 + X + y karyotypes. Actually, pairing of the Oedionychina autosomes seldom fails, and in those cases where it does, the chromosomes tend to face one another at M I, showing, most probably, distance pairing after precocious cession (see Fig.
17b in VIRKKI 1972). Therefore, it seems that the obligatory association of euchromatic arms, inher- ited from the 10 + X + y condition, has started to relax in the fusion karyotype of Al. equestris. We found no structural causes for this failure in our preparations. It is worth noticing, however, that we may be witnessing an incipient microevolution- ary case towards single-arm association bivalents, leading back to the telephone-handle form and, perhaps, to a true diphasism. Before the latter condition is reached, all important genes of the free arms must have been transferred to the pairing arms. Further research on Al. equestris cytogenet- ics and that of its taxonomic allies promise to elucidate the formation of the typical unichiasmate metacentric bivalents in male Coleoptera.
No heterozygous trivalents formed by fused and unfused arms were seen in Al. equestris, although we assume that a rapid succession of fusions have taken place. Probably, the fusion chromosomes have been strongly favored by selection. Small, isolated demes, typical of many Oedionychina (VIRKKI 1980; VIRKKI and ZAMBRANA 1980), have bottlenecked a rapid fixation of the present karyotype later spread by inbreeding (CHARLES- WORTH 1985). Lack of chain multivalents further indicates that the fusions have not occurred at random but each time between two predetermined autosomes. This is just what SMITH (1966; SMITH and VIRKKI 1978; VIRKKI 1984) encountered in the C . stigma karyocline. This predetermination obviously is related to linkage disequilibrium rela- tions (CHARLESWORTH I985), although structural and “somatogrammic” ( interphasic chromosomal neighborhoods, WAGENAAR 1969) relations may have helped in the rapid succession of the rear- rangements.
Asphaera is the only other oedionychine genus where both lowered autosomal number and ring bivalents co-occur. One undetermined species of Asphaera has 9 + 7x + Y , “Asph. pauperata-like” as well as Asph. octopunctata have 8 + X + y, and the lowest autosomal number was encountered in Asph. (Omophoita) clerica: 7 + 2x + Y (all these multiple sex chromosome data modified or cor- rected from SMITH and VIRKKI 1978, and PETT- TIERRE et al. 1988). In the last three species, one or two bivalents appear in a ring form, with one terminal contact, the other one is a subterminal chiasma. Also, in a deletion rod bivalent of Aspi. scutata (this study), a subterminal chiasma was found. Thus, some rearrangements can modify the otherwise very conservative extreme distal localiza-

Heredifas 119 (1993) 280 N. VIRKKI AND J. A. SANTIAGO-BLAY
tion of the chiasma. All these cases in Asphaera are presumably older and have accumulated more postfusion rearrangements than AI. equestris.
Because proximal collochores (VIRKKI 1989) tend to co-occur with procentric heterochromatin, one would expect distal collochores co-occurring with the overcondensed distal ends seen in the fusion autosomes. At A I, the V-form metacentrics should thus advance towards the poles with their telomeres together, and the chromatidal cen- tromeres slightly apart (Fig. 6 in VIRKKI 1989). This is not the case in Al. equestris, where both the autosomal and sex chromosome chromatids re- main apposed although separate. This arrangement survives the congression and orientation to M 11, and when the anaphase movements begin, it sug- gests a switch to a diffuse centromere structure in all chromosomes. Deviant centromeric structure and function, consisting of the lack of procentric heterochromatin (ENNIS 1974) and a tendency to assemble nucleolar material in male meiosis (SMITH 1969), were also found in the euchromatic fusion monophasics of several species of Chilo- corus.
Are such changes the result of centric fusions? We do not know the karyotype immediately pre- ceding in an evolutionary sequence and we cannot be sure if, how, and when some of these features were present. There are 10 + X + y karyotypes in Oedionychina with uncertain distal localization of collochore (Virkki and Santiago-Blay, unpubl.), as well as karyotypes where individual autosomes show a proximal collochore while most show distal ones (Fig. 4 in VIRKKI 1989).
The switch from a localized to diffuse (or polycentric) centromere has been studied both in Lepidoptera (GASSNER and KLEMETSON 1974; HOLM and RASMUSSEN 1980) and Trichoptera (WOLF et al. 1992). Among the Oedionychina herein reported, the fusion karyotype is unique in showing such a switch. Alagoasa equestris differs from the lepidopterans and trichopterans because its centromere turns diffuse in the second meiotic division; it is localized in the first one. The situa- tion in Alagoasa equestris reminds of the hemipter- ans studied by NOKKALA (1985) and NOKKALA and NOKKALA (1985). These authors attribute the switch from monokinetism to holokinetism to a change in the exposition pattern of the kinetic DNA sequences of axial core: the kinetic DNA is exposed only at the centromeres in M I, but all along the chromosome in M I1 and in the mitoses. Ultrastructural studies would help understanding
how the collochore and centromere changes may relate to autosomal fusions.
Origin of fragments and B chromosomes
The Chilocorus karyocline studied by SMITH ( 1966) offers a plausible, albeit characteristically Coleopteran, mechanism for the origin of the B chromosomes. In a transect from Florida via Nova Scotia to Alberta, C. stigma loses about half of its chromatin, due to successive fusion of diphasic metacentrics in such a way that two monophasic metacentrics arise: one euchromatic, the other, a heterochromatic B chromosome (SMITH and VIRKKI 1978; VIRKKI 1984). Loss of the inert arms may occur also by deletion, as can be inferred from the karyotypic differences between C. kuwanae and its close taxonomic allies. Therefore, the non-chias- mate arms of diphasics deserve to be considered as a potential source for fragments and B chromo- somes in Coleoptera. In the case of C. stigma-type rearrangements, the number of autosomes de- creases, as in Al. equestris herein reported, whereas it remains unchanged in the C. kuwanae-type suc- cessive deletions.
Our material precluded a colchicine reaction needed to discover the diphasism (SMITH 1965). On the other hand, all dispensable achiasmate arms do not show the differential colchicine reac- tion reliably, as found by SMITH (1970, 1973) in Pissodes weevils. Apparently, the diphasism can evolve in degrees and in different ways, depending on heterochromatinization of old chromatin as well as on accretion of a new one.
The acentric fragments found in Al. januaria apparently have derived from at least three achias- matic autosomal arms by erosion of dispensable segments. The survival expectancy of such frag- ments cannot be large, but it is possible that new ones are frequently formed. Sometimes, many of them can be expected to be included by chance in the gamete(s), resulting in new individuals with ususually high number of fragments. Another pos- sibility is that there has been, within such an individual, a major quirk of erosion.
It is particularly interesting that the B chromo- somes of “Alagoasa transparente” and Al. oblecta congress in the space between the “sex spindle” and the autosomal spindle-portion at M I. Their placement in that space could be due to their engagement with the PM I spindle later than the regular autosomal bivalents, although earlier than the sex chromosomes (VIRKKI 1972, 1990). Also,

Hereditas 119 (1993) KARYOTYPE EVOLUTION IN NEOTROPICAL FLEA B E E ~ E S 281
when two Bs are present in “ A . transparente”, they distance-pair like the sex chromosomes. Similarly, in the cases of precessive autosomes sporadically encountered in many Oedionychina M I, they tend to lie adjacent to the sex spindle. There is little reason to assume that they have remained trapped inside of the sex spindle. On the contrary, they may stay very close to other autosomes, although facing the sex chromosomes (Fig. 17b in VIRKKI 1972). It seems that the sex spindle can induce distance pairing in chromosomes in its vicinity, at least if they are syntelically orientated. The B chromo- somes of Al. oblecta, although loosely congressed in the same space, are supposedly amphiorientated, and form no distance bivalents or multivalents. The fragments of Al. januaria show no affinity towards the sex spindle.
It is difficult to explain the origin of the I to 2 Bs in “Al. transparente”. A guess is that they repre- sent what is left of a heterochromatic autosomal arm once separated by a centric fission, which provided the centromere. The frequent hetero- morphy in autosomal bivalents of Al. oblecta sug- gests multiple and more recent origin of Bs by the same means. For reasons hard to explain in terms of selection, their centromeres must have switched to an amphitelic orientation, because the Bs divide at A I. Whether a postreduction follows, remains unknown.
Diphasic metacentrics: the favored “autosomal phenotype” of Coleoptera: Polyphaga?
Using cytological and hybridization criteria, Smith demonstrated that the overcondensed, achiasmate arms of C. stigma are dispensable and perishable, whereas the chiasmate arms survive the transloca- tions and can still be identified in the successive fusion karyoforms of increasing monophasedness (see Smith’s summary in SMITH and VIRKKI 1978, p. 161-173). ENNIS (1974, 1976) showed that the condensed telephone-handle arms of Chilocorus cannot be C-banded. We call this type of dispens- able arms “cryptically heterochromatic” (SMITH and VIRKKI, 1978; cf. also CRAIG and BICKMORE 1993).
In the present material, Al. equestris suggests a C. stigma-like fusion history, with autosomal num- ber lowered to one half, autosomal bivalents bibrachially bichiasmate, euchromatic and mono- phasic, altered centromeric structure as well as function, and presence of B chromosomes. Alagoasa januaria also suggests a C. kuwanae-type
evolution, where the dispensable arms are eroded away, and the chromosome number remains unal- tered.
The question now arises whether such arm dispensability, partial or total, might be more common in beetle chromosomes. Metacentric auto- somes forming rod bivalents with subterminal or “terminal” localization of the only chiasma are very common in male meiosis of beetles belonging in the Suborder Polyphaga (SMITH and VIRKKI 1978). Such a chiasma distribution limits the ge- netic crossing-over to one extreme of, as we as- sume, one and the same arm (MANNA and SMITH 1959; VIRKKI and REYES-CASTILLO 1972). The overcondensation of the achiasmate arms (straight rod of telephone-handle shape) may not be the consequence of a lacking chiasma and, indeed, it is not in the fusion rod bivalents of Al. equestris. On the contrary, structural changes in the achiasmate arms could be the reason that impedes chiasma formation. This is to be assumed especially in cases like the two largest bivalents of AZ. parana (Fig. 5D in VIRKKI 1988a), with grossly longer achias- matic arms as compared to the chiasmatic ones.
Recent studies show that such structural changes might be due to formation and controlled disper- sion of repetitive DNA (DOOLIITLE 1985). More- over, if one agrees with DOVER (1980) in assuming that in the chromosomes there is a continuous formation and elimination of repetitive sequences, with the sheltering of more important chromosome regions from this turnover, then we can visualize the following model for repeated structural evolu- tion of polyphagan autosomes as observed at MI of males:
1. Telocentric autosomes either turn to metacentry by means of pericentric inversion, or accrete a second arm by accumulating repetitive DNA.
2. Metacentric autosomes that initially may form bibrachially bichiasmate rings, increasingly accu- mulate repetitive DNA in one of the arms, which turns overcondensed and achiasmate.
3. Most major genes are localized in the chiasmate arm, but their recombination by chiasmata is lim- ited to the distal 25 %, or less, of the arm length. Repetitive DNA arising in this arm is transposed to the achiasmate arm.
The end result would be a Smith’s diphasic, metacentric chromosome with one arm genetically inert and dispensable, slower in replication and reaction to colchicine than a chiasmate arm, but

Hereditas 119 (1993) 282 N. VIRKKI AND J. A. SANTIAGO-BLAY
most Akigousu, most Wulierianelk~ Aspicelu scutata, Oinophoira, Walrerianella Alagoasa equestris
most Asphaera some Asphaera venusta
repeated centric fissions in X chromosomes followed by pencentric inversions? 10+nx+Y (n = 1 to 7)
centric fissions even distribution of nuclear pores in spermatocyte I
10 pairs of autosomes
Fig. 13. Hypothetical cladogram of some genera in the Oedionychina based on cytological character of the male meiosis. (Modified from VIRKKI et al. 1991, with publisher’s errors corrected, by permission of Psyche’s editor.) spz/b = spermatozoa per bundle.
not necessarily showing all characteristics of con- stitutive heterochromatin, such as heteropyknosis or C-bands (CRAIG and BICKMORE 1993; ENNIS 1974, 1976; VIRKKI 1983, 1984).
It is still largely unknown how the diphasic or “diphasoid” autosomal phenotype is expressed in female meiosis. It is commonly believed that the heterogametic sex, the male in this case, tends to a more restricted frequency and distribution of chias- mata (JONES 1987). However, the scanty informa- tion available for the Coleoptera does not seem to support this (SUOMALAINEN 1940, 1947; SMITH and VIRKKI 1978, p. 138). For structural reasons it seems feasible that diphasics must form rod biva- lents also in the female.
Phylogeny of Oedionychina
Fig. 13 represents a preliminary progress report of our views about the possible relationships among some genera in the Oedionychina based on cytoge- netic data. Although the tree is not fully resolved, we prefer to bring our model to scrutiny for it may have heuristic value. We are not aware of a phylo- genetic analysis for the genera in this putative tribe; nevertheless, the overall topology of the tree corresponds to our intuition of possible relation- ships between the genera. Of course, a more rigor-
ous examination of other genera and of other families of characters is needed to have a more compelling (i.e., better supported) model. If these are “natural” groups, then those other analyses would help to uniquely define the groups “Ornophoita and some Asphaera” as well as “most Alagoasa and most Walterianella”, both currently “defined” by 10 + X + y owing to our inability to find other defining cytological characters. Other problems with any tree are the difficulties in select- ing outgroups, the determination of homologies, and polarization of characters.
Aeknowledgmenrs. - The authors wish to express their gratitude to the late Dr Jan Bechyne and to Mrs Bohumila Springlova de Bechyne (Universidad Central de Venezuela, Maracay), to Dr Gerhard Scherer (Zoologische Staatssammlung, Miinchen, Ger- many) and to Dr David G. Furth (Harvard University, Cam- bridge, Mass., U S A . ) for their help in identifying some of the specimens. Author N.V. is grateful to all persons who helped him plan the practical phases of the collection, especially to Jan Bechyne and Felisberto Camargo (Rio de Janeiro), Ing. Fabian Portilla (Quito), Prof. Francisco Carrasco (Cuzco), and Ing. Luis Liceres Zarate (Tingo Maria, Peru). Dr. Amilton Ferreira (Cam- pus President, Universidade Estadual Paulista at Rio Claro, Brazil) offered laboratory facilities and additional help during a sabbatical of N.V., in 1980-81. Further thanks are due to the Brazilian mining company, “ICOMI”, who provided room, board, and transportation in Amapa during 1964. We are grateful for the kindness of Mss. Mdriko Yasuda and Rovene Prelock, who were responsible for most of the typing on this paper.

Hereditas 119 (1993) KARYOTYPE EVOLUTION IN NEOTROPICAL FLEA BEETLES 283
References BECHYNE, J. and SPRINGLOVA DE BECHYNB. 1966. Evidenz der
bisher bekannten Phenrica-Arten (Col. Phytophaga, Alti- cidae). - Entomol. Tidskr. 87: 142-170
CHARLESWORTH, B. 1985. Recombination, genome size and chro- mosome number. - In: The Evolution of Genome Size (ed K. CAVALIER-SMITH), Wiley & Sons, New York, p. 489-513
CRAIG, J. M. and BICKMORE, W. A. 1993. Chromosome bands - flavours to savour. - BioEssays 15 349-354
DOOLITTLE, W. F. 1985. The evolutionary significance of middle- repetitive DNAs. - In: The Evolution of Genome Size (ed K. CAVALIER-SMITH), Wiley & Sons, New York, p. 443-487
DOVER, G. A. 1980. Ignorant DNA?- Nature 2 8 5 619-620 ENNIS, T. J. 1974. Chromosome structure in Chilocorus (Cole-
optera: Coccinellidae). I . Fluorescent and Giemsa banding patterns. - Can. J . Genet. Cytol. 16: 651-661
ENNIS, T. J. 1976. Chromosome structure in Chilocorus (Cole- optera: Coccinellidae). 11. The asynchronous replication of constitutive heterochromatin. - Can. J . Genet. Cytol. I8 85-91
EUTENEUER, U. and MCINTOSH, J. R. 1981. Structural polarity of the kinetochore microtubules in PtK, cells. - J . Cell Biol. 89 338-345
GASSNER, G. and KLEMETSON, D. J. 1974. A transmission electron microscopic examination of hemipteran and lepidopteran gonial centromeres. - Can. J . Genet. Cytol. 16: 457-464
HOLM, P. B. and RASMUSSEN, S. W. 1980. Chromosome pairing, recombination nodules and chiasma formation in diploid Bom- byx males. - Carlsberg Res. Commun. 45: 483-548
JONES, R. N. 1987. Chiasmata. - In: Meiosis (ed P. B. MOENS), Acad. Press, New York, p. 213-244
JONES, R. N. and REES, H. 1982. B Chromosomes. - Acad. Press, New York
MANNA, G. K. and SMITH, S. G. 1959. Chromosomal polymor- phism and interrelationships among bark weevils of the genus Pissodes Gemdr. - Nucleus 2 179-208
MCINTOSH, J. R. and MCDONALD, K. L. 1989. The mitotic spindle. - Sci. Am. 261: 48-56
MESA, A. and FONTANETTI, C. S. 1983. Mecanismo sexual complexo, polimorfismo autosomal e numero alto de cromosso- mos S en Enchroma gigantea (L. 1758) (Coleoptera, Bupresti- dae). - Abstr. 35. Reun. Anual Soc. Brasil. Progr. Cienc.
NOKKALA, S. 1985. Restriction of kinetic activity of holokinetic chromosomes in meiotic cells and its structural basis. - Hered- itas 102: 85-88
NOKKALA, S. and NOKKALA, C. 1985. Mitotic and meiotic behavionr of axial core structure of holokinetic chromosomes. - Hereditas 103: 107-110
NOKKALA, C. and NOKKALA, S. 1989. Characteristics of B-chro- mosomes in Golerucella nymphaeae complex (Coleoptera, Chrysomelidae). - Hereditas I I I : 179- 189
PETITPIERRE, E., SEGARRA, C., YADAV, J. S. and VIRKKI, N. 1988. Chromosome numbers and meioformulae of Chrysomelidae. - In: Biology qf the Chrysomelidae (eds P. JOLIVET, E. PETIT- PIERRE, and T. H. HSIAO), Kluwer Academic Publishers, Dordrecht, The Netherlands, p. 161 ~ 1x6
SCHUBERT, I., SCHRIEVER-SCHWEMMER, G., WERNER, T. and ADLER, I.-D. 1992. Telomeric signals in Robertsonian fusion and fission chromosomes: implications for the origin of pseudo- aneuploidy. - Cytogenet. Cell Genet. 59: 6-9
SEENO, T. N. and WILCOX, J . A. 1982. Leafbeetle genera (Coleoptera: Chrysomelidae). - Entomography I : 1 - 121
SMITH, S. G. 1965. Heterochromatin, colchicine, and karyotype. - Chromosoma 16: 162-165
SMITH, S. G. 1966. Natural hybridization in the coccinellid genus Chiiocorus. - Chromosoma 18: 380-406
SMITH, S. G. 1969. Nucleolar organisation and chromocentre formation in Chilocorus. - In: Chromosomes Today / I (eds C. D. DARLINGTON and K. R. LEWIS), Oliver &Boyd Ltd., Edinburgh, p. 41-43
SMITH, S. G. 1970. Chromosomal polymorphism in North Amer- ican Pissodes weevils: structural isomerism. - Can. J . Genet. Cytol. 12: 506-540
SMITH, S . G . 1973. Chromosomal polymorphism and interrelation- ships in Pissodes weevils: Additional cytogenetical evidence on synonymy. - Can. J . Genet. Cytol. 15 83-100
SMITH, S. G. and VIRKKI, N. 1978. Animal Cytogenetics: Coleoptera (ed B. JOHN). - Borntraeger, Stuttgart-Berlin
SUOMALAINEN, E. 1940. Beitrage zur Zytologie der partheno- genetischen Insekten. I . Coleoptera. - Ann. Acad. Sci. Fenn. A I V : 1-143
SUOMALAINEN, E. 1947. Parthenogenese und Polyploidie bei Riisselkafern (Curculionidae). - Hereditas 33: 425-456
VIDAL, 0. R. 1984. Chromosome numbers of Coleoptera from Argentina. - Genetica 6 5 235-239
VIRKKI, N. 1963. High chromosome number and giant postreduc- tional sex chromosomes in the beetle Walterianella uenusta Scbaufuss (Chrysomelidae, Alticinae). - J . Agric. Uniu. Puerto Rico 47: 151-163
VIRKKI, N. 1964. On the cytology of some neotropical chrysomelids (Coleoptera). -Ann. Acad. Sci. Fenn. A I V : 1-24
VIRKKI, N. 1967. Orientation and segregation of asynaptic multiple sex chromosomes in the male of Omophoita clerica Erichson (Coleoptera: Alticidae). - Heredifas 5 7 276-288
VIRKKI, N. 1968. Regular segregation of seven asynaptic sex chromosomes in the male of Asphaera daniela Bechyne (Coleoptera, Alticidae). - Caryologia 21: 47-51
VIRKKI, N. 1970. Sex chromosomes and karyotypes of the Alticidae (Coleoptera). - Hereditas 64 267-282
VIRKKI, N. 1972. Contraction stage and formation of the distance sex bivalent in Oedionychina (Coleoptera, Alticidae). - Hered- itas 71: 259-288
VIRKKI, N. 1980. Fleabeetles, especially Oedionychina, of a Puerto Rican marshland in 1969-72. - J . Agric. Uniu. Puerto Rico 64: 63-92
VIRKKI, N. 1983. Banding of Oedionychina (Coleoptera) chromo- somes. - J . Agric. Univ. Puerto Rico 6 7 221-255
VIRKKI, N. 1984. Chromosomes in evolution of Coleoptera. - In: Chromosomes in Evolution of Eukaryotic Groups, Vol. II (ed.7 A. K. SHARMA and A. SHARMA), CRC Press, p. 41-76
VIRKKI, N. 1988a. Cytotaxonomy of Alticinae. - In: Biology of Chrysomelidae (eds P. JOLIVET, E. PETITPIERRE, and T. H. HSIAO), Kluwer Acad. Publishers, p. 187-203
VIRKKI, N. 1988b. The sex chromosomes of Disonychina (Coleoptera, Alticinae): Xy + nX systems. - Cytobios 53: 43- 55
VIRKKI, N. 1989. Proximal vs. distal collochores in Coleopteran chromosomes. - Hereditas 1 1 0 101-107
VIRKKI, N. 1990. What happens in the clump stage of spermato- genesis? - Nucleus 33: 41 -43
VIRKKI, N. and KIMURA, M. 1978. Distribution of nuclear pores and perinuclear dense substances in spermatocytes of some Oedionychina fleabeetles. - Biosystems 1 0 213-225
VIRKKI, N. and REYES-CASTILLO, P. 1972. Cytotaxonomy of Passalidae (Coleoptera). - An. Esc. Nac. Cienc. Bid . Mixico 1 9 49-83
VIRKKI, N., SANTIAGO-BLAY, J. A. and CLARK, S. M. 1991. Chromosomes of some Puerto Rican Disonychina and Oe- dionychina (Coleoptera: Chrysomelidae: Alticinae: Oediony- chini): Evolutionary implications. - Psyche 98 373-395
VIRKKI, N., SANTIAGO-BLAY, J. A. and RILEY, E. G. 1992. Chromosomes of Puerto Rican Hispinae and Cassidinae (Coleoptera: Chrysomelidae). - Coleops Bull. 4 6 29-42
VIRKKI, N. and ZAMBRANA, I. 1980. Demes of a Puerto Rican fleabeetle, Alagoasa bicoior (L.) differing mainly in mean body size and foodplant association. - J . Agric. Univ. Puerto Rico 64: 264-274
WAGENAAR, E. 1969. End-to-end chromosome attachments in mitotic interphase and their possible significance to meiotic chromosome pairing. - Chromosoma 26 410-426
WHITE, M. J. D. 1973. Animal Cytology and Evolution. Third Edition. - Cambridge Uniu. Press, Cambridge, Great Britain
WOLF, K. W., NOVAK, K., and MAREC, F. 1992. Chromosome structure in spermatogenesis of Anabolia furcata (Trichop- tera). -Genome 35 46-52
WU, T.-P. 1992. B chromosomes in Sorghum stipoideum. - Hered- ity 68: 457-463