henrietteeae (melastomataceae): a new neotropical berry-fruited tribe
TRANSCRIPT

Systematic Botany (2010), 35(4): pp. 783–800© Copyright 2010 by the American Society of Plant TaxonomistsDOI 10.1600/036364410X539862
783
Miconieae DC. are the largest tribe of Melastomataceae Juss., with ca. 19–23 genera and 1,800 species ( Michelangeli et al. 2008 ). Miconieae are native to the Neotropics where they are frequently found in great abundance, from the low-lands to high elevation páramos. Miconia calvescens DC. and Clidemia hirta D. Don are notoriously invasive members of this tribe. Subsequent to their introductions, they have negatively impacted Pacific and Paleotropical ecosystems ( DeWalt and Hamrick 2004 ). This tribe of woody plants is generally char-acterized by 4- or 5-merous flowers, a prevalence of short to elongate and persistent external calyx teeth, usually glabrous petals with blunt to acute apices, stamens usually twice the number of petals with ovoid to elongated and usually at least slightly curved and more or less unappendaged anthers that open by apical pores or longitudinal slits, and baccate fruits with numerous minute seeds ( Judd 1989 ; Michelangeli et al. 2004 ). Among Neotropical melastomes, only the Blakeeae Benth. & Hook. f. are also berry-fruited, but this group is dis-tinguished from Miconieae by its prevailingly six-merous, axillary, and generally large flowers each subtended by two sets of decussate bracts ( Penneys et al. 2004a ; Penneys 2007 ; Morales-P. and Penneys 2010; Penneys and Judd in press ). In addition, a base chromosome number of x = 17 is characteris-tic of Miconieae ( Almeda 1997 ; Solt and Wurdack 1980 ).
Following recent revisions to the circumscription of the Miconieae, the tribe appears to be monophyletic ( Clausing and Renner 2001 ; Renner et al. 2001 ; Michelangeli et al. 2004 , 2008 ; Goldenberg et al. 2008 ). Recent analyses suggest that Eriocnema Naud. ( Fritsch et al. 2004 ; Michelangeli et al. 2004 ; Martin et al. 2008 ) and Physeterostemon R. Goldenb. & Amorim ( Amorim et al. 2009 ; Penneys unpubl. data), two Brazilian capsular-fruited genera, may be sister to the Miconieae, as traditionally circumscribed, and as such are intermediate between Miconieae and its sister tribe, Merianieae Triana ( Michelangeli et al. 2004 ). Chalybea Naud. and Huilaea Wurd. are now known to be members of the Blakeeae ( Penneys et al. 2004a ; Penneys 2007 ; Morales-P. and Penneys 2010 ),
rather than the Miconieae ( Triana 1871 ; Wurdack 1957 ; Judd 1989 ). The more or less unappendaged anthers and a chromo-some number of n = 17 have been suggested as putative syna-pomorphies for the Eriocnema + Miconieae s. s. clade ( Fritsch et al. 2004 ), suggesting that the berry fruits characteristic of the tribe are synapomorphic for the clade containing all gen-era of the Miconieae except Eriocnema and Physeterostemon .
The absence of convincingly delimited monophyletic groups within the Miconieae is well known to students of the family ( Cogniaux 1891 ; Macbride 1941 ; Wurdack 1972 ; Judd 1989 ; Judd and Skean 1991 ). Our view is that only a few of the currently recognized genera are likely to be monophyletic ( Goldenberg et al. 2008 ). Indeed, many are simply constructs of convenience or circumscriptions based on the examination of limited material (e.g. Cogniaux 1891 ). The circumscription of natural groups will be aided by ongoing phylogenetic anal-yses ( Michelangeli et al. 2004 , 2008 ; Penneys and Judd 2005 ; Judd 2007 ; Judd et al. 2008 ; Goldenberg et al. 2008 ; Bécquer-Granados et al. 2008 ; Martin et al. 2008 ) using morphological and molecular data.
Classifications of the Miconieae have included (among many others) these six genera, Bellucia Neck. ex Raf., LoreyaDC., Myriaspora DC., Henriettea DC., Henriettella Naud., and Llewellynia Pittier ( Naudin 1852 ; Bentham and Hooker 1867 ; Triana 1871 ; Baillon 1877 ; Cogniaux 1891 ; Krasser 1898 ), and their placement within this tribe has been accepted by later workers ( Gleason 1958 ; Wurdack 1980 ; Renner 1989 ; Almeda 2001 ). In an early cladistic analysis of the Miconieae, Judd (1989) found that these genera form a clade (including Kirkbridea Wurdack) sister to the rest of the Miconieae (or at least its axillary-flowered members).
In the course of our phylogenetic investigations of the Miconieae (e.g. Michelangeli et al. 2004 ; Penneys et al. 2004b ), it became apparent that Bellucia, Loreya, Henriettea , and Henriettella are not members of the Miconieae, nor are these genera (including, presumably, Kirkbridea, Llewelynia and Myriaspora ) sister to that tribe. Rather, the aforementioned
Henrietteeae (Melastomataceae): A New Neotropical Berry-Fruited Tribe
D. S. Penneys , 1 , 4 F. A. Michelangeli , 2 W. S. Judd , 3 and F. Almeda 1
1 Department of Botany, California Academy of Sciences, San Francisco, California 94118-4503, U. S. A.; e-mail: [email protected]; [email protected].
2 The New York Botanical Garden, 200th St. & Southern Blvd., Bronx, New York 10458, U. S. A.; e-mail: [email protected]. 3 Department of Biology, University of Florida, Gainesville, Florida 32611, U. S. A.; e-mail: [email protected].
4 Author for correspondence
Communicating Editor: Andrea Schwarzbach
Abstract— With nearly 2,000 species, all restricted to the Neotropics, Miconieae represent the largest radiation within Melastomataceae.Generic limits and relationships within the tribe have long been contested. Our molecular phylogenetic analyses of the tribe, based on sequences of nrITS and ndhF , resulted in consistent support for a clade comprising Bellucia, Loreya, Henriettea , and Henriettella, which is nei-ther nested within nor sister to the Miconieae. Furthermore, and based upon a series of presumably synapomorphic anatomical and mor-phological characters, e.g. megastyloid crystals, absence of both medullary and cortical vascular bundles, plinerved leaves, axillary or cauliflorous inflorescences, we consider three additional genera, Kirkbridea, Llewelynia , and Myriaspora , to belong to this clade, a newly recog-nized tribe, Henrietteeae . These seven genera had formerly been assigned to the Miconieae based on their having exappendiculate anthers (except Kirkbridea ) and berry fruits. Henriettella and Llewelynia are relegated to synonymy under Henriettea (25 new combinations are made under that genus: Henriettea boliviensis , H. bracteosa , H. caudata , H. duckeana , H. fissanthera , H. glabra , H. cogniauxiana , H. goudotiana , H. heteroneura , H. hondurensis , H. ininiensis , H. lawrancei , H. lundellii , H. maguirei , H. manarae , H. odorata , H. ovata , H. prancei , H. rimosa , H. steyermarkii , H. tachirensis , H. tobagensis , H. tovarensis , H. trachyphylla , H. williamsii ), and Loreya and Myriaspora are synonymized under Bellucia (with nine new combinations under that genus: Bellucia egensis , B. gracilis , B. klugii , B. nigricans , B. ovata , B. riparia , B. stri-gosa , B. subandina , B. wurdackiana ). A description of Henrietteeae along with a key to and brief descriptions of its constituent genera are pro-vided, in addition to necessary new combinations.
Keywords— homoplasy , Miconieae , ndhF , nrITS , phylogeny , taxonomy.

784 SYSTEMATIC BOTANY [Volume 35
cladistic analyses formed a distinct clade that diverged earlier in the evolutionary history of the family.
In this paper, we present phylogenetic analyses conducted in order to test the monophyly and phylogenetic position of the Henriettea clade. We sequenced the internal transcribed spacer (ITS) region of 18S – 26S nuclear ribosomal DNA as well as the plastid region, ndhF , then conducted both individ-ual and combined maximum parsimony (MP) and maximum likelihood (ML) analyses. The phylogeny recovered in the combined analysis was used as the basis for the recognition and description of the new tribe, Henrietteeae, and associated taxonomic changes. The taxonomic history of the constituent genera of the Henrietteeae, along with their putative pheno-typic synapomorphies, is also discussed.
Materials and Methods
Taxon Sampling— Flowering and vegetative material was collected in silica gel, and voucher specimens of all taxa utilized in the DNA analysis are deposited at A, ALBC, CAS, FLAS, INBIO, MO, NY, PMA, PORT, QCNE, VEN, US, and USZ (Appendix 1). Appendix 1 is a com-plete list of vouchers sequenced for this study; authorities given in the appendix are not repeated elsewhere in this paper. Leaf tissues in silica gel and DNA extractions were kept at -20°C for long-term storage. The molecular cladistic analysis included 14 OTU’s of the Henriettea alliance. Outgroup taxa included 32 species representing eight tribes ( Cogniaux 1891 ). Species of Pternandra Jack., putatively sister to the remaining Melastomatoideae ( Clausing and Renner 2001 ; Stone 2005 ), were used to root the cladograms.
Molecular Data Set— Each collection cited in Appendix 1 was sequenced for internal transcribed spacer regions of nuclear ribosomal DNA (nrITS) including portions of the 18S gene, the entire ITS 1, 5.8S cis-tron and ITS 2 regions, as well as the plastid ndhF .
Total genomic DNA was extracted either using Qiagen DNeasy kits (Qiagen, Inc., Santa Clarita, California) following the manufacturer’s instructions or from silica gel-dried leaves using a modified CTAB pro-cedure of Doyle and Doyle (1987) . The specimens were ground in mor-tars, or pulverized in a mixer mill, then incubated at 60°C for 30–120 minutes with 1.2 ml CTAB and either 8 μl of β-mercaptoethanol or 5 μlof Proteinase K, then transferred to labeled 1.5 ml tubes. DNA was pre-cipitated overnight with isopropanol at –20°C, centrifuged for 20 minutes at 13,000 rpm, washed twice with 70% ethanol, and dried. The DNA pel-let was then resuspended in 75 μl of 1 × TE (Tris-EDTA buffer, pH 8.5) and incubated at 60°C for approximately 30 minutes. DNA quality was checked by electrophoresis in a 1% agarose gel containing ethidium bro-mide in a Tris-Borate-EDTA Buffer (TBE). Preceding PCR amplification, total DNA samples were cleaned with Qiagen QIAquick PCR purification columns following the manufacturer’s instructions.
The internal transcribed spacer region of the nuclear ribosomal DNA (nrITS) was amplified using specific primers as detailed by Michelangeli et al. (2004) . PCR was performed in a 25 μl volume with the following reaction components: 0.8 μl template DNA (ca. 30 ng/μl), 2.0 μl 10 × Ex Taq buffer (Takara Bio Inc, Japan), 0.2 mM dNTP mixture, 6.5 mg BSA, 1 μmol of each primer, and 5% (v/v) of DMSO. The PCR parameters were 94°C for 5 min.; 35 cycles of 94°C for 10 sec, 50°C for 45 sec, 72°C for 50 sec, followed by 72°C for 10 min. Alternatively, some taxa were sequenced using the primers of Sun et al. (1994) and a touchdown thermal cycling program was used; the initial annealing temperature was 76°C, decreasing 1°C per cycle for 15 cycles, followed by 15 cycles at 61°C. Each step lasted for one minute.
The ndhF gene was amplified with primers developed by Olmstead and Sweere (1994) . Following Clausing and Renner’s study (2001) , we amplified the 3′ end of the gene between positions 972 (i.e. codon 305 of solanaceous sequences); and 1955, using forward primer ndhF-972F, reverse primer ndhF-1955R, and one or two pairs of internal primers (ndhF-1318F, ndhF-1318R, ndhF-1603F, and ndhF-1603R). Sequencing conditions and reagents were similar to those used for ITS, but the anneal-ing temperature was set to 48°C. Alternatively, a similar PCR program was run with 10 initial cycles with an annealing temperature of 45°C, fol-lowed by 25 cycles with an annealing temperature of 48°C. All PCR prod-ucts were purified using either MicroClean (Microzone Ltd., Haywoods Heath, U. K.), or QIAquick columns following the manufacturer’s instruc-tions and checked on an agarose gel for quality.
Cycle sequencing was performed using the primers cited above in 20 μl reactions including 3 μl of PCR product template and 17 μl of master mix. The thermocycler program used for cycle sequencing began at 96°C for two minutes, followed by 25 cycles of 10 sec at 96°C, 5 sec at 50°C, and 4 minutes at 60°C. Upon completion of this program, the product was held at 4°C. Cycle sequencing products were cleaned with either Exosap (Amersham Biosciences, Piscataway, New Jersey), or a 70% ethanol pre-cipitation procedure.
The cycle sequencing product was analyzed on Applied Biosystems ABI 377 or 3100 sequencers at the DNA Sequencing Core Lab at the University of Florida or the New York Botanical Garden using the man-ufacturer’s protocols for the dye-terminator reaction. Sequencher 4.7 (GeneCode Corp., Ann Arbor, Michigan) was used to make appropriate judgments by visually inspecting every base position before assembling the electropherograms. The ends of matrices were trimmed to exclude sequencing artifacts. Edited sequences were manually aligned in Se-Al ( Rambaut 1996 ). Molecular matrices (ITS, ndhF , and ITS + ndhF ) are avail-able on TreeBASE (study number S10711).
Phylogenetic Search Strategies— The program PAUP* 4.0b10 ( Swofford 2002 ) was used to reconstruct phylogenetic relationships among all taxa in the ITS, ndhF , and combined MP analyses. In all cases, heuristic analyses of equally weighted characters were conducted with 10,000 random-addi-tion replicates with tree-bisection-reconnection (TBR) branch swapping, character states unordered, gaps treated as missing data, MaxTrees setting = 100, and MulTrees on. The strength of support for the cladistic relationships found in the initial analyses was assessed using bootstrap support (BS; Felsenstein 1985 ). For each analysis, a BS analysis was per-formed with 10,000 resampling replicates, a heuristic search, TBR, with MulTrees in effect.
For ML analyses, the data set was first examined using Modeltest 3.7 ( Posada and Crandall 1998 ), which, based upon the Akaike Information Criterion (AIC), identified as most appropriate models of evolution the general time reversible (GTR; 4-nucleotide frequency state parameters; 6 substitution rate parameters), plus proportion of invariable sites (I), plus a gamma distribution (Γ), for both the ITS and combined ITS + ndhFmatrices, and the transversion model (TVM), plus I, plus Γ for the ndhFmatrix. ML trees were generated using GARLi ver. 0.951 ( Zwickl 2006 ) with branch support calculated from 500 bootstrap replicates. Missing data in the aligned matrices totaled 13.74%, 4.37%, and 9.17% in the ITS, ndhF , and combined analyses, respectively.
Results
ITS— The range of lengths of all ITS 1, ITS 2 spacers and the 5.8S cistron is similar to that found in other angiosperms (Baldwin 1992 ). The 5.8S cistron is 163 bp in length for almost all taxa. For the entire ITS region, 982 nucleotide sites ( Table 1 ) were unambiguously aligned and included in the analysis. Of these sites, 546 were constant, 436 were variable, and 329 were phylogenetically informative in the parsimony analysis.
The MP analysis of ITS resulted in 43 most parsimonious trees of 1,241 steps, CI = 0.55, and RI = 0.74. The tree ( Fig. 1 ) was rooted with Pternandra spp. The Tibouchineae + Microlicieae clade receives BS 82, and comprises ( Desmoscelis
Table 1. Summary statistics for phylogenetic analyses of molecular data sets for the Henrietteeae with additional outgroups.
ITS ndhF ITS + ndhF
Alignment length (bp) 982 934 1,964No. of constant characters 546 662 1,092No. of variable characters 436 272 872No. of informative characters 329 177 658No. of MP trees 43 2,233 41Tree length 1,241 474 2,482CI 0.55 0.73 0.55RI 0.74 0.86 0.74ML model GTR+ I+Γ TVM+ I+Γ GTR+I+ ΓLn −7,572.9644 −4,098.3530 −15,145.9287Fig. MP/ML 1/2 3/4 5/6
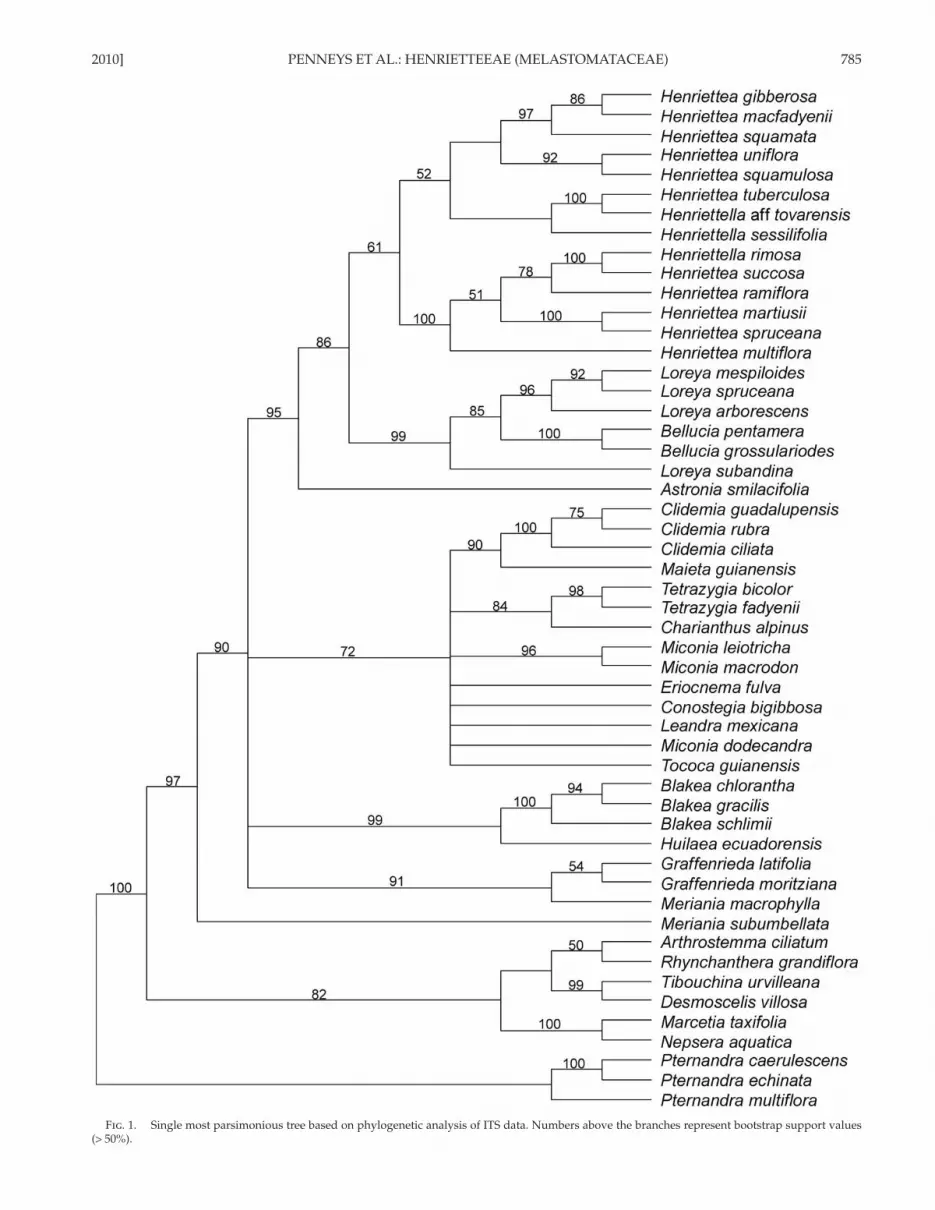
2010] PENNEYS ET AL.: HENRIETTEEAE (MELASTOMATACEAE) 785
Fig . 1. Single most parsimonious tree based on phylogenetic analysis of ITS data. Numbers above the branches represent bootstrap support values (> 50%).

786 SYSTEMATIC BOTANY [Volume 35
villosa + Tibouchina urvilleana , BS 99) + ( Arthrostemma cilia-tum + Rhynchanthera grandiflora , BS 50) + ( Nepsera aquatica + Marcetia taxifolia , BS 100). Meriania subumbellata is sister (BS 97) to a clade containing all remaining taxa, which form a polytomy (BS 90) comprising Blakeeae (BS 99), Miconieae (BS 72), and Merianieae (BS 91). Astronia smilacifolia (Astronieae) is sister (BS 95) to the members of the Henriettea alliance (BS 86). Within the latter clade, Bellucia spp. + Loreya spp. receives strong support (BS 99), while Henriettea spp. + Henriettellaspp. is only weakly supported (BS 61).
In the ML ITS analysis, the GTR + I + Γ model was statisti-cally the most appropriate DNA substitution model ( Table 1 ). The ML tree is shown as Fig. 2 . Pternandra spp. were used to root the tree and the remaining taxa form a dichotomy com-posed of Astronia smilacifolia sister (BS 51) to the Henrietteaalliance (BS 53), and a clade (BS 57) comprising the Blakeeae (BS 85), Merianieae (BS 52), Tibouchineae + Microlicieae clade (BS 100), and Miconieae (BS 76). Within the Henriettea alli-ance, Bellucia spp. + Loreya spp. receives strong support (BS 89), while Henriettea spp. + Henriettella spp. form a weakly supported clade (BS 56).
ndhF— For the ndhF marker, 934 nucleotide sites were unambiguously aligned and included in the analysis. Of these sites, 662 were constant, 272 were variable, and 177 were phy-logenetically informative in the parsimony analysis.
The MP analysis of the aligned ndhF matrix resulted in 2,233 most parsimonious trees, CI = 0.73, and RI = 0.86 ( Table 1 ). The strict consensus tree ( Fig. 3 ) illustrates a polytomy com-posed of Astronia smilacifolia , the Henriettea alliance (BS 96), and the remaining taxa (BS 59). There is little resolution within the Henriettea alliance, though the Bellucia spp. + Loreyaspp. clade is weakly supported (BS 63). With BS 97, Blakeeae (BS 99) is strongly supported as sister to the Tibouchineae + Microlicieae clade (BS 91), and Miconieae (BS 96) is supported as sister (BS 78) to the Merianieae (BS 79).
In the ML analysis of ndhF , the TVM + I + Γ model was statistically the most appropriate DNA substitution model ( Table 1 ). The ML tree (-lnL = -4,098.3530; Fig. 4 ) shows the Pternandra clade as sister (BS 100) to the clade composed of the Henriettea alliance (BS 90), plus the remaining taxa (BS 52). In the latter grouping, Astronia smilacifolia is sister to the clade (BS 66) composed of the Blakeeae (BS 100) + Tibouchineae + Microlicieae (BS 92) which together receive strong statistical support (BS 98), plus the clade (BS 80) Miconieae (BS 95) + Merianieae (BS 84). Within the Henriettea alliance, Belluciaspp. + Loreya spp. form a weakly supported clade (BS 57).
ITS + ndhF— For the combined ITS + ndhF analysis, 1,964 nucleotide sites were included. Of these sites, 1,092 were con-stant, 872 were variable, and 658 were phylogenetically infor-mative in the parsimony analysis ( Table 1 ).
The MP analysis of the ITS + ndhF matrix resulted ( Fig. 5 ) in 41 most parsimonious trees of 2,482 steps, CI = 0.55, and RI = 0.74. With minor exceptions, the combined ML analy-sis (using the GTR + I + Γ model) resulted in a tree ( Fig. 6 ) with nearly the same topology as MP, thus relationships and bootstrap statistics are discussed here jointly. These analy-ses show Pternandra spp. as sister (MPBS 100/MLBS 100) to the remaining taxa, and Astronia smilacifolia sister (MPBS 100/MLBS 54) to a clade composed of the Henriettea alli-ance (MPBS 96/MLBS 63). Meriania subumbellata is sister (MPBS 100) to a grouping of all other taxa (MPBS 99/MLBS 60). Within the Henriettea alliance, Bellucia spp. + Loreyaspp. is well supported (MPBS 100/MLBS 91) and resolved,
while the Henriettea spp. + Henriettella spp. clade is moder-ately supported (MPBS 73/MLBS 69) and well resolved. The Tibouchineae + Microlicieae clade (MPBS 92/MLBS 100) appears between Pternandra spp. and the clade composed of the remaining taxa in the parsimony analysis, whereas in the likelihood analysis, it is one element of a polytomy containing Blakeeae (MPBS 100/MLBS 87), Merianieae (MPBS 96/MLBS 63), and Miconieae (MPBS 86/MLBS 98).
Discussion
Henrietteeae and Its Two Subclades— Our phylogenetic investigations of the Miconieae have illuminated errors in previous classifications made as a result of misleading and homoplasious patterns of morphological variation. In light of the results presented here ( Figs. 1 – 6 ), Bellucia, Loreya, Henriettea,and Henriettella, and also their putative relatives, Kirkbridea,Llewelynia, and Myriaspora, are removed from the Miconieae and placed in a newly recognized tribe, Henrietteeae, which is formally described below. This alliance of ca. 100 species is found to be monophyletic in phylogenetic analyses of ITS, ndhF , and the combined molecular analyses. The new tribe is consistently well-supported as a clade distinct from (and not sister to) the Miconieae. Retaining these taxa within the Miconieae would render that tribe biphyletic. Although our sampling of putatively early divergent Asian melastome out-group genera such as Pternandra , Astronia Blume, AstrocalyxMerr., Astronidium A. Gray, and Beccarianthus Cogn., remains sparse, the Henrietteeae appear to represent (see Figs. 1 – 6 ) an early-divergent lineage within the family.
The circumscription of the Miconieae has recently under-gone another contraction, namely, the removal of Huilaea and Chalybea to the Blakeeae ( Penneys et al. 2004a ; Penneys 2007 ; Morales-P. and Penneys 2010; Penneys and Judd in press ), as well as a minor expansion as a result of the inclusion of the two species of Eriocnema ( Fritsch et al. 2004 ; Michelangeli et al. 2004 ; Andrade et al. 2007a ; Martin et al. 2008 ; Goldenberg et al. 2008 ) and three species of Physeterostemon ( Amorim et al. 2009 ). The former change reduces somewhat the variability of anatomical and floral characters (see Morales-P. and Penneys 2010 ). Eriocnema adds the perennial herbaceous habit and a capsular fruit type ( Andrade et al. 2007b ) to an otherwise woody, berry-fruited tribe. The species of Physeterostemonare also capsular, but these plants are shrubs to subshrubs. Likewise, removing the Henriettea alliance causes the remain-ing taxa to share a reduced complement of anatomical and reproductive characters, for example, taxa with megasty-loid crystals and lacking medullary bundles are excluded (see below). These modifications to the circumscription of the Miconieae have been discussed previously ( Michelangeli et al. 2004 , 2008 ; Goldenberg et al. 2008 ) and will be described by our group in greater detail in forthcoming publications along with further insights into internal tribal relationships, generic circumscriptions, and character evolution.
The Henrietteeae are clearly monophyletic as demon-strated in the individual ITS, ndhF , and combined molecular analyses ( Figs. 1 – 6 ). In a morphology-based cladistic analy-sis, Judd (1989) identified a clade composed of the seven genera here considered as the tribe Henrietteeae, and is thus supportive of our molecular results. Several morphological and anatomical characters consistently separate this group from the Miconieae. Whether these characters are synapo-morphic for the group, or plesiomorphic in relation to the
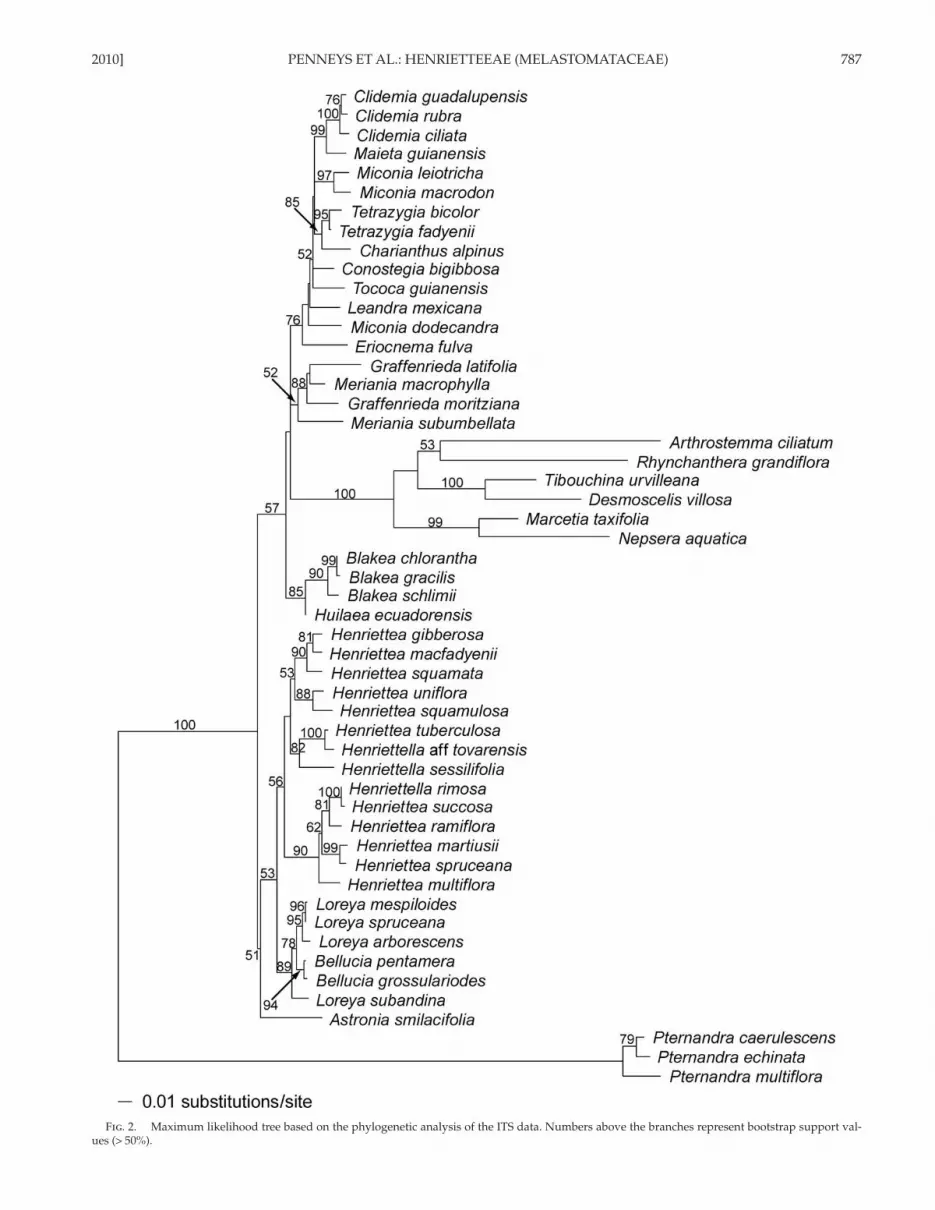
2010] PENNEYS ET AL.: HENRIETTEEAE (MELASTOMATACEAE) 787
Fig . 2. Maximum likelihood tree based on the phylogenetic analysis of the ITS data. Numbers above the branches represent bootstrap support val-ues (> 50%).
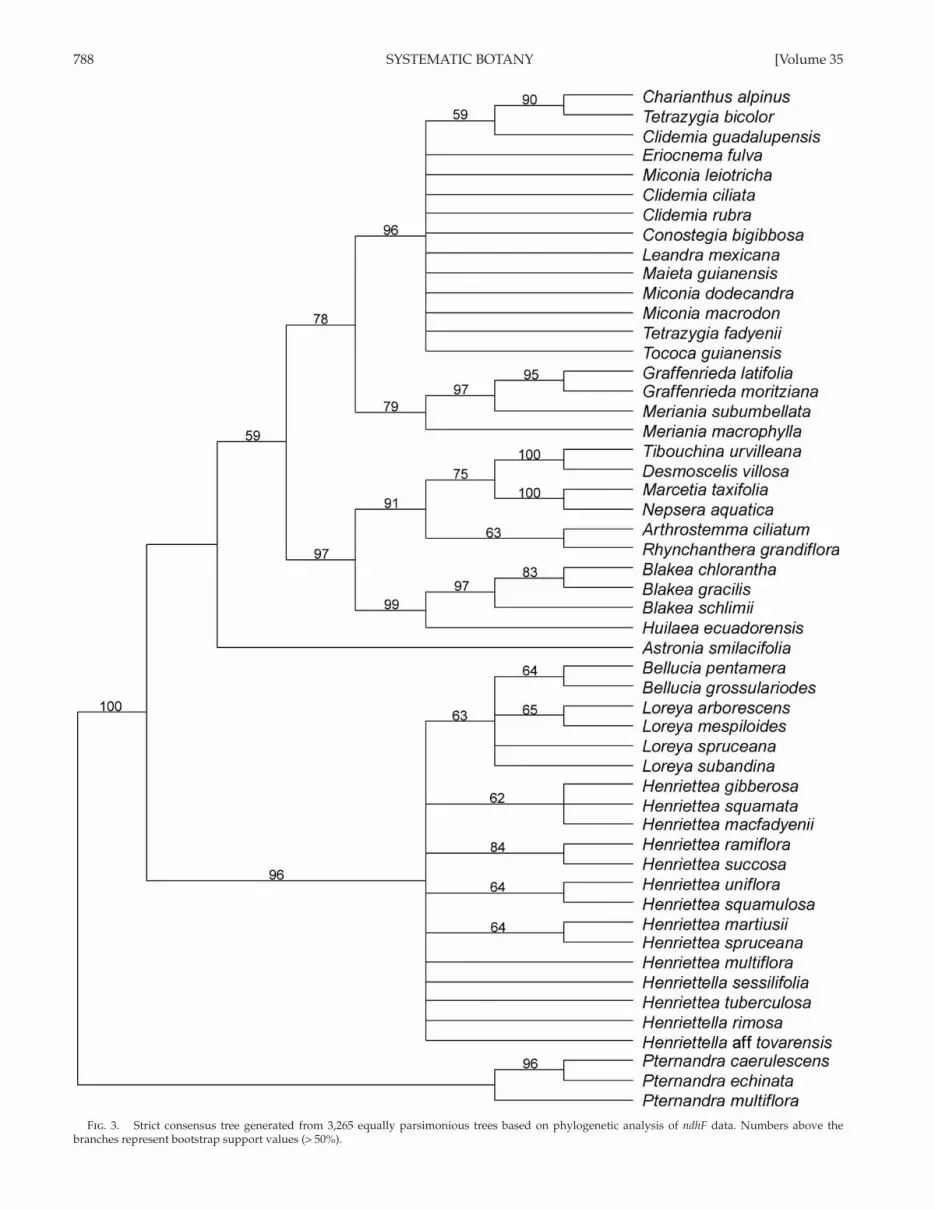
788 SYSTEMATIC BOTANY [Volume 35
Fig . 3. Strict consensus tree generated from 3,265 equally parsimonious trees based on phylogenetic analysis of ndhF data. Numbers above the branches represent bootstrap support values (> 50%).

2010] PENNEYS ET AL.: HENRIETTEEAE (MELASTOMATACEAE) 789
Fig . 4. Maximum likelihood tree based on phylogenetic analysis of ndhF data. Numbers above the branches represent bootstrap support values (> 50%).
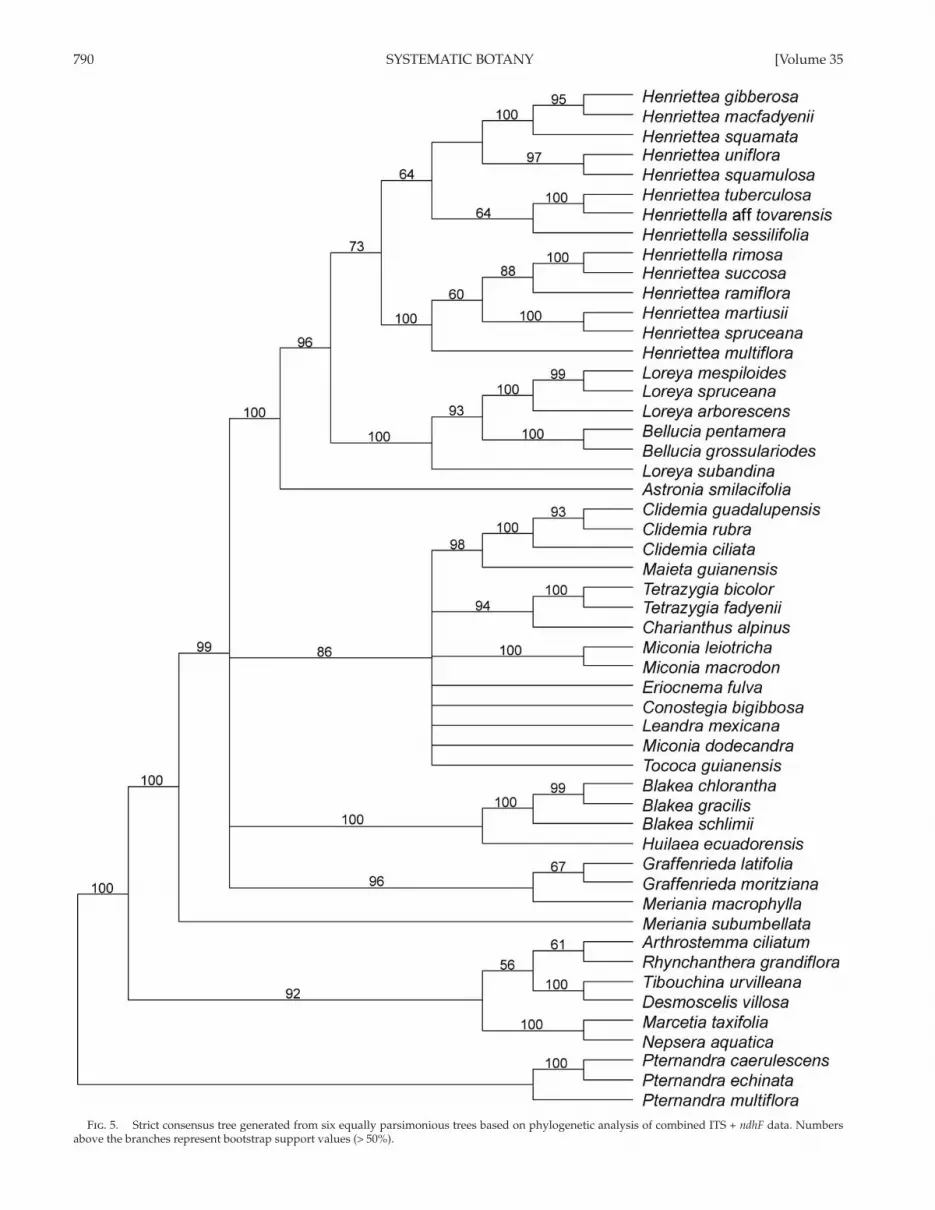
790 SYSTEMATIC BOTANY [Volume 35
Fig . 5. Strict consensus tree generated from six equally parsimonious trees based on phylogenetic analysis of combined ITS + ndhF data. Numbers above the branches represent bootstrap support values (> 50%).

2010] PENNEYS ET AL.: HENRIETTEEAE (MELASTOMATACEAE) 791
Fig . 6. Maximum likelihood tree based on phylogenetic analysis of combined ITS + ndhF data. Numbers above the branches represent bootstrap support values (> 50%).

792 SYSTEMATIC BOTANY [Volume 35
various potential outgroups, such as Pternandra and, more importantly, the Astronieae, however, needs to be more rig-orously tested. Megastyloids are large, elongated crystals of calcium oxalate; they are located in the secondary xylem ( Solereder 1908 ; ter Welle and Mennega 1977 ) but also occur in other tissues; they are especially conspicuous in the leaves ( Judd et al. 2008 ). They are present in nearly all species of the Henriettea alliance and are particularly conspicuous in Henriettea . They are lacking in the Miconieae, the members of which instead typically produce druse crystals, but megasty-loids also occur in some Astronieae ( Baas 1981 ). Van Tieghem (1891a , 1891b ) found that Loreya, Myriaspora , Henriettea , and Henriettella all lack both medullary and cortical vascular bun-dles, and they were therefore removed from the Miconieae in his anatomy-based classification. Cogniaux (1891) sig-naled in the final two paragraphs of his monograph that he had recently received van Tieghem’s (1891a) publication and accepted the findings. In Loreya, Myriaspora , Henriettea , and Henriettella , van Tieghem (1892) found that superficial cork-formation occurs as the outermost cell-layer of the primary cortex becomes the phellogen. Ter Welle and Koek-Noorman (1981) found exclusively septate fibers in Bellucia , Loreya and Henriettella. In other Neotropical genera they sampled, both septate and nonseptate fibers were found. Anomocytic sto-mata also characterize the Henrietteeae plus Astronieae ( Baas 1981 ) in contrast to the polocytic or diacytic stomata typical of the Miconieae.
Additional potential synapomorphies for the Henrietteaalliance include plinervy ( Penneys et al. 2004b ), axillary or cauliflorous inflorescences ( Judd 1989 ), and pubescent petals ( Judd 1989 ). Elongated hairs with roughened, often dendritic bases are synapomorphic for Henriettea , though not found in all species ( Wurdack 1986 ; Judd 1989 ; Penneys et al. 2004b ; Judd et al. 2008 ).
Henrietteeae have berry fruits derived from inferior ova-ries, a condition that may have evolved independently in the Miconieae, Dissochaeteae Triana, and Blakeeae, though it has been noted ( Clausing et al. 2000 ) that the fruit of Pternandrais a fleshy capsule/dry berry. Placentation is parietal in Pternandra , basal to basal-axile in Astronieae ( Clausing et al. 2000 ; Clausing and Renner 2001 ), and axile to intruded axile in Henrietteeae (and most Miconieae). The unappendaged (except Kirkbridea ), porose anthers ( Fig. 7 ) of the Henrietteeae are generally similar to the anthers of Miconieae, though the anthers of some species of Bellucia closely resemble those of Blakea . Our ongoing investigations into family-level phy-logenetics should clarify the level of universality of these characters.
The relatively few available chromosome counts also dis-tinguish the Henrietteeae from the Miconieae. The base num-ber in Miconieae is x = 17 ( Almeda 1997 ; Almeda and Chuang 1992 ). Henriettella fascicularis and Henriettella ramiflora (Sw.) Naudin both have n = 28, in Myriaspora egensis Mart. ex DC. n = 15, and in Bellucia pentamera n = 20 (21) ( Solt and Wurdack 1980 ). Judd (1989) noted that Loreya, Bellucia, and Myriasporaare united by the putative synapomorphies of a chromo-some number of n = 15 or 20, and Henriettea, Henriettella, and Llewelynia form a clade based upon a chromosome number of n = 28, though these observations are based on a paucity of chromosome counts for these genera.
The MP and ML analyses of ITS sequences as well as both analyses of ITS + ndhF sequence data ( Figs. 1 , 2 , 5 , 6 ) support two major subclades within the Henrietteeae, i.e. a Bellucia
+ Loreya clade and a Henriettea + Henriettella clade, although the monophyly of the latter clade does not receive strong sta-tistical support. The morphology-based cladistic analysis of Judd (1989) also supported these clades, with the Bellucia + Loreya clade (which also included Myriaspora ) united by the synapomorphies of flowers lacking external calyx teeth, pos-sessing robust fleshy anthers, and an expanded and radiately ridged stigma, and the Henriettea + Henriettella clade (includ-ing Llewelynia and with Kirkbridea sister to these three gen-era) supported by the presence of multicellular strigose hairs with a stellate, radiate, irregularly branched, or suprabasally expanded base (or hair types clearly modified from these con-ditions), petals with an apical apiculum on the abaxial sur-face, and fasciculate inflorescences. The taxonomic history of, and phylogenetic patterns within, each of these two clades are discussed below.
Bellucia, Loreya, and Myriaspora— The intergeneric rela-tionships within the Henrietteeae are obscured due to non-discrete generic boundaries. Cogniaux (1891) , referring to the Miconieae, in which these genera were then placed, stated that, “The delimitation of genera is arbitrary enough; one could easily distinguish more of them, at the same time one could justify the reunion of many of those which are gener-ally admitted today”. The close relationship between Belluciaand Loreya has long been remarked upon by students of the Melastomataceae. In the most recent monograph of these plants, Renner (1989) maintained the two genera, but explic-itly accepted the resultant paraphyly of Loreya . Gleason and Smith (1933) noted that the genera were weakly separated, and also that the petals of Loreya in bud are imbricated in such a way as to suggest the related genus, Henriettella . Wurdack (1965 , 1973 , 1980 ) maintained Loreya and Bellucia and consid-ered the closest generic affinities of Loreya to be with Henrietteaand Henriettella (1965), although we note that he may have been impressed by symplesiomorphic similarities.
The relationship between Bellucia and Loreya is so intimate that some workers ( Baillon 1877 ; Judd 1989 ) have considered them to be congeneric. In the treatment of the Melastomataceae for the Flora of Peru, Macbride (1941) , synonymized Loreyaunder Bellucia rather than “straining at a gnat” to recognize both. Renner (1989) found that the only consistent and dis-crete character used to distinguish Bellucia from Loreya is the presence of a glaucous, papillose epidermis on the abaxial leaf surface in the former (clearly synapomorphic, when com-pared with outgroups) vs. the smooth, nonglaucous abaxial leaf surfaces of the latter (retained plesiomorphies).
Flower merosity, the number of stigma lobes, and ovary cells have been emphasized in earlier classifications (e.g. Renner 1989 ) though these characters exhibit considerable overlap between the putative genera, e.g. Loreya subrotundifolia(Wurdack) S. Renner and L. riparia S. Renner link with Belluciaon the basis of higher merosity. In most species of Bellucia,the 6–8-merous flowers have calyx lobes that cover the devel-oping petals until just prior to anthesis, and in some species, e.g. B. acutata Pilger, B. beckii Renner, B. dichotoma Cogn., and B. grossularioides , the calyx is calyptrate ( Judd 1989 ; Renner 1989 ). In Loreya , the 5-(6–7)-merous flowers have calyx lobes that are vestigial and the developing petals quickly protrude beyond them, a condition also found in B. huberi (Wurdack) Renner, a species originally described in Loreya ( Renner 1989 ). Renner (1989) noted that the seed coat of Bellucia is irregularly sculptured with bulging periclinal cell walls, while in Loreyait is regular-tubercular.

2010] PENNEYS ET AL.: HENRIETTEEAE (MELASTOMATACEAE) 793
Fig . 7. Stamens of representative Henrietteeae species in dorsal, side, and ventral views. A: Henriettea odorata . B: H. fissanthera . C: H. tuberculosa . D: H. lundellii . E: Llewelynia williamsii (left, dorsal view; right, side view). F: H. seemannii . G: H. ramiflora . H: H. strigosa . I: H. succosa . J: Myriaspora egensis . K: Loreya spruceana . L: Bellucia pentamera .

794 SYSTEMATIC BOTANY [Volume 35
The number of anther pores ( Fig. 7 ) has also been used to separate Bellucia and Loreya , with two pores in Bellucia, and one or two pores in Loreya ( Cogniaux 1891 ; Gleason and Smith 1933 ; Renner 1989 ). However, Penneys (2007) found this character to be remarkably labile in the Blakeeae with the uniporate condition arising multiple times within that tribe, and there is little to suggest that it could be of greater predic-tive value in related tribes.
The above described mosaic pattern of variation in mor-phological characters fits well with our molecular phyloge-nies ( Figs. 1 – 6 ; and available morphology-based analyses see Renner 1989 ; Judd 1989 ), all of which suggest that Bellucia is nonmonophyletic if Loreya is segregated.
The systematic affinities of Myriaspora also need to be clari-fied, and it is unfortunate that we were not able to include this taxon in our analyses. However, the recognition of Myriaspora,a monotypic, Amazonian genus of shrubs and trees, also likely renders Loreya (as traditionally circumscribed) para-phyletic. Renner (1989) , on the basis of a morphological anal-ysis, explicitly noted that the maintenance of Myriaspora and Bellucia as separate genera causes Loreya to be paraphyletic, but considered such an action to be justifiable on the basis of convenience and tradition. Based on his analyses, Judd (1989) considered these three groups to be congeneric, as he consid-ered the recognition of a nonmonophyletic Loreya to be mis-leading and inappropriate following a phylogenetic approach. The characters, i.e. shrubby habit and flowers borne in the axils of extant leaves rather than on old wood, purportedly
distinguishing Myriaspora from Bellucia and Loreya can all be found in the latter genera. In such characters as the smooth, abaxial foliar epidermis, 5-merous flowers, truncate calyx, abaxially strigulose petals, 5-lobed stigmas, 5-celled ovaries, uniporate anthers, and highly regular tuberculate seed coats, Myriaspora closely resembles Loreya ( Judd 1989 ; Renner 1989 ). The calyx lobes of Myriaspora are fused into an unusual calyp-triform, irregularly dehiscent cap that may not be homolo-gous with the cap of some species of Bellucia ( Judd 1989 ).
Currently available morphology-based phylogenetic analy-ses ( Judd 1989 ; Renner 1989 ) indicate that no apomorphies unite the species of Loreya when certain species are segregated as either Bellucia or Myriaspora . The paraphyly ( Figs. 1 – 6 ) of Loreya is also evident in our DNA-based phylogenetic analyses, despite the modest taxon sampling. When the vari-able morphological characters historically employed in the diagnosis of Bellucia, Loreya , and Myriaspora , are considered, our data dictate the recognition of a broadly circumscribed Bellucia , rather than maintaining two (or possibly even more) genera of doubtful monophyly.
Henriettea, Henriettella, Kirkbridea, and Llewelynia–—Henriettea and Henriettella have been variously combined and (weakly) separated since shortly after Naudin (1852) pro-posed the latter genus. This group of shrubs and small trees commonly have plinerved leaves, planar to bullate adaxial leaf surfaces, stellate-based setae ( Fig. 8 ), and clustered or fas-ciculate inflorescences emanating from leaf axils or old wood (sometimes cauliflorous to ground level). Their flowers are
Fig . 8. Hairs of representative Henrietteeae species. A: Henriettea ramiflora . B: H. lundellii . C: H. seemannii . D: H. strigosa . E: H. tuberculosa .

2010] PENNEYS ET AL.: HENRIETTEEAE (MELASTOMATACEAE) 795
4–5-merous, with uniporose or biporose anthers ( Fig. 7 ), and truncate stigmas. The fruits are many-seeded berries derived from inferior ovaries.
De Candolle (1828) erected Henriettea to accommodate H. succosa (first described as Melastoma succosa Aubl.), a spe-cies that he distinguished from other melastome genera in his Prodromus on the basis of the plinerved leaves, axillary inflo-rescences, velvety petals, basally bifid, uniporose, and apically rostrate anthers ( Fig. 7 ), a pubescent style, capitate stigma, and juicy berry. Henriettea currently comprises ca. 15 species of shrubs and small trees that mostly have plinerved leaves with bulla-based hairs on the adaxial surface. The flowers are mostly 2–3 cm in diameter, with apically obtuse and basally clawed petals, and the anthers are often purple and apically attenuate-rostrate or oblong-attenuate.
Henriettella was proposed by Naudin (1852) , and in that publication, the characters used to justify the recognition of this new genus were restricted to its smaller flowers and non-rostrate anthers. Naudin (1852) listed three species, Henriettellagoudotiana Naud., H. patrisiana (DC.) Naud., and H. seeman-nii Naud., and later Britton and Wilson (1925) selected the latter species as the lectotype. The ca. 40 species of shrubs and small trees comprising Henriettella have flowers mostly 0.5–1 cm in diameter, with apically acute petals, and anthers that are often yellow and oblong to obovate.
Those students of the Melastomataceae who have main-tained the separation between Henriettea and Henriettellainclude Bentham and Hooker (1867) , Triana (1866 , 1871 ), Cogniaux (1891) , Krasser (1898) , Gleason (1932 , 1958 ), Wurdack (1973 , 1980 ), Wurdack et al. (1993) , Renner (1989) , Almeda (2001 , 2009 ), and Mendoza and Ramirez (2006) . Although Wurdack separated the two genera in his family treatments for the floras of Venezuela (1973), Ecuador (1980), and French Guiana ( Wurdack et al. 1993 ), he noted elsewhere ( Wurdack 1972 ) that Henriettella ovata Cogn. and Henrietteapatrisiana form a generic transition zone. Almeda (2009) also admitted that in his treatment the genera were separated not out of firm taxonomic conviction, but rather to avoid intro-ducing nomenclatural changes in advance of a complete revi-sion. Other systematists, including Grisebach (1864) , Baillon (1877) , Macbride (1941) , Howard (1989) , Williams (1963) , Liogier (1995) , Proctor (1972) , Standley and Williams (1963) , Judd (1986 , 1989 ), and Judd et al. (2008) , have considered Henriettella to be synonymous with Henriettea , and some have made the appropriate combinations to reflect that view. Macbride (1941 : 500), in fact, stated that “there seems to be no significant concomitance of the characters of petals, anthers or pubescence that have been used to maintain two similar but distinct genera”. The evidence for combining the two genera has been discussed in detail by Judd (1986 , 1989 ) who found that a narrowly defined Henriettea could not be diagnosed by any autapomorphies and that its species intergrade with Henriettella , while Henriettea s. l. (including Henriettella ) is sup-ported by several distinctive morphological synapomorphies (see above). Judd (1989) also enumerated a variety of charac-ters and species that bridge supposed generic boundaries.
The monotypic Llewelynia Pittier, which unfortunately was not included in our analyses, is characterized by a calyptrate calyx and pleiostemony (with 35–50 stamens per flower). However, in all other aspects, it closely conforms with other species of the Henriettea alliance. Judd (1989) noted that the calyces of H. spruceana , H. succosa , H. stellaris Berg ex Triana, and H. ramiflora (Sw.) DC. also form caps, and, although
dehiscence in the latter species is longitudinal rather than circumscissile, they are likely homologous. Furthermore, calyptrate calyces have evolved independently numerous times within other genera of Melastomataceae, e.g. Blakea,Clidemia, Huilaea, Mecranium J. D. Hooker, and Miconia Ruiz & Pav. Presumably basal genera of melastomes, e.g. Pternandra,Astronia, and Astronidium likewise frequently have calyptrae ( Maxwell and Veldkamp 1990a , 1990b ). Pleiostemony is spo-radic among melastome tribes, e.g. Astronieae ( Astrocalyx ), Dissochaeteae ( Plethiandra Hook. f.), and Miconieae ( ClidemiaD. Don, Conostegia D. Don, Miconia ) ( Kadereit 2006 ). Finally, Llewelynia shares more anatomical features with Henrietteathan Miconieae ( ter Welle and Koek-Noorman 1981 ). Given the labile and homoplasious nature of these character states, and the placement of Llewelynia williamsii Pittier in a clade with Henriettea and Henriettella in the cladistic analyses of Judd (1989) , we see no justification for maintaining this spe-cies as distinct from Henriettea .
In his description of the new genus Kirkbridea , Wurdack (1976) noted that it differs from the closely related gen-era, Loreya, Henriettea , and Myriaspora , in having prominent external calyx teeth, dimorphic stamens with bilobed ven-tral appendages, and ovaries that are 1/2–2/3 inferior and crowned with strigose pubescence. Judd (1989) found that Kirkbridea is closely related to Henriettea s. l., being linked to that clade by the presence of megastyloids, multicellular stri-gose hairs with an expanded or stellate base, 4- or 5-merous flowers, and petals with an apical apiculum on the abaxial surface. Molecular evidence confirming this placement is greatly desired, but on the strength of the aforementioned characters, we feel justified in including Kirkbridea in the Henrietteeae, though its exact placement within the tribe remains uncertain.
Taxonomic Treatment
Henrietteeae Penneys, Michelangeli, Judd, and Almeda, tri-bus novum. Type genus: Henriettea DC., Prod. 3. 178. 1828.
Tribus novum, Astronieae affine, a quo antheris porosis et fructibus baccatis differt, et Miconieae affine, a quo crystallis megastyliformibus (vs. crystallis solum globosis) et fasciculis vascularibus medullosis et corticeis absentibus (vs. praesenti-bus) differt. Type species: Henriettea succosa (Aubl.) DC.
Evergreen, terrestrial shrubs, treelets, or trees; various tis-sues with megastyloid crystals, druses sometimes also pres-ent; stems lacking both medullary and cortical vascular bundles. Indumentum variable, the hairs sparsely to densely distributed, uniseriate to multiseriate, elongate, eglandular or gland-tipped, often with roughened or dendritic-stellate basal region, strigulose, granulose, furfuraceous, or glabres-cent. Twigs compressed, quadrate, to terete in cross-section. Stipules absent. Petioles terete to adaxially canaliculate. Leaves opposite, decussate, equal to slightly anisophyllous, blade chartaceous to coriaceous, planar, smooth to minutely papillose on abaxial epidermis, the apex acute to broadly rounded, often abruptly short to long acuminate, the base acute to obtuse or rounded, margin plane to revolute, entire to variously toothed; venation acrodromous, mostly plinerved, some basally-nerved, with prominent midvein (primary vein) and 2–7 pairs of secondary veins (including a pair of weak, submarginal veins), tertiary veins numerous, ± perpendicular

796 SYSTEMATIC BOTANY [Volume 35
to midvein, adaxial surface glabrescent, or with persistent hairs (as above), abaxial surface essentially glabrous to densely pubescent with various hair types (as above). Inflorescences fasciculate, short cymose, or flowers solitary at each node, pedunculate or ± sessile, axillary in distal nodes, on old wood below the leaves, or cauliflorous, bracts and bracteoles cadu-cous (rarely persistent). Flowers perfect, mostly 4–8-mer-ous, pedicellate, actinomorphic, with androecium and/or style commonly declinate. Hypanthium narrowly to broadly globose, cylindrical to conical, terete, the outer surface gla-brous or pubescent (as above), the inner surface glabrous, obscurely to prominently ridged. External calyx lobes sub-apical, conic, short-subulate to elongate or obsolete, inter-nal calyx lobes calyptrate or valvate, rupturing regularly or irregularly at anthesis, the lobes shallowly ovate to triangu-lar or depressed semicircular, the margin entire. Petals imbri-cate in bud, symmetrical or oblique, reflexed, or rotate, often fleshy, oblong, ovate, elliptic, lanceolate, obovate, to rhombic, often clawed basally, the apex acute, obtuse, rounded, trun-cate, to emarginate; often with adaxial callose ridges, the mar-gins entire or with one to several marginal claws on at least one side, often cucullate distally with an external subapical tooth, often puberulent, white, cream, pale yellow, pink, or lavender-tinged. Stamens mostly twice as many as the petals, rarely pleiostemonous, incurved in bud, isomorphic (dimor-phic in Kirkbridea ), glabrous, filaments terete, stout to slender; anthers white, cream, yellow, lavender, sometimes changing
color postpollination, free, anther sacs rounded to deeply cleft ventrally, laterally compressed or rounded, linear to obtuse, apically rounded to rostrate, opening by one or two apical pores, or more or less rimose, connective thickened dorsally, not or scarcely prolonged below the anther sacs or shortly bilobed at the base (prominently so in Kirkbridea ). Ovary (3-)4–8(-15)-loculate, partially to completely inferior, apically glabrous or rarely glandular- or strigose-pubescent, smooth to ridged; placentation axile to intruded axile, the ovules numerous, style elongate, terete, cylindrical, glabrous or glandular-pubescent, white, cream, pink, or lavender; stigma clavate, truncate, capitellate, or capitate, often lobed or fluted. Berries ± globose to hemispheric, greenish when immature, becoming yellowish-green, red, or deep purple at maturity, glabrous to pubescent (as above), the exocarp thin to leathery. Seeds obovate-angulate, obovate-rounded, or pyriform, with irregular shallow grooves, tuberculate, vaguely foveolate, or muriculate.
Karyology—Henriettella fascicularis (Sw.) Triana and Henriettella ramiflora (Sw.) Naudin, n = 28; Myriaspora egen-sis 2 n = 30; Bellucia axinanthera Triana n = 20 (21) ( Solt and Wurdack 1980 ).
Distribution— Southern Mexico through Central America, Bolivia, Brazil, northern South America, the Greater and Lesser Antilles. Especially diverse in the Amazon Basin and Greater Antilles. Occurring from sea level to ca. 3,000 m elevation.
Genera—Bellucia Raf., Henriettea DC., Kirkbridea Wurd.
Key to the Genera of Henrietteeae
1. Inflorescence clearly pedunculate; stamens dimorphic; anthers with bilobed, ventral appendages; ovaries 1/2–2/3 inferior, crowned with strigose pubescence . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Kirkbridea
1. Plants without the above combination of characters . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2 2. Indument of slender to stout, elongate, multicellular hairs with radiate to stellate-globular basal portion, or derived types,
e.g. stellate hairs, peltate scales, elongate hairs with minute projections near base, or elongate hairs with a suprabasal expanded region and narrowed basal attachment; flowers with external calyx teeth (although sometimes inconspicuous); anthers nonfleshy; ovary (3-)4–6 loculate; stigma truncate or only slightly capitate-expanded, nonridged . . . . . . . . . . . . . . . . . . . . . . Henriettea
2. Indument of slender to stout, elongate, unbranched, multicellular hairs; flowers lacking external calyx teeth; anthers robust, fleshy; ovary 5–15 loculate; stigma expanded and clearly radiately ridged . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Bellucia
B ellucia Rafinesque, Sylva tellur. 92. 1838, nom. cons.—TYPE: Bellucia grossularioides (L.) Triana.
Webera J. F. Gmelin, Syst. Nat. 2 (1): 776, 820. Sep-Nov 1791, non Schreber, May 1791, nec Hedwig, 1801.—TYPE: Webera quinquenervia (Aublet) J. F. Gmelin Basionym: Blakea quinquenervia Aubl.
Apatitia Desvaux in Hamilton, Prodr. Pl. Ind. Occid. 42. 1825.—TYPE: Apatitia blakeoides Desvaux.
Myriaspora DC. Prodr. 3: 165. 1828.—TYPE: Myriaspora egen-sis DC.
Loreya De Candolle, Prodr. 3: 178. 1828.—TYPE: Loreya arbore-scens (Aublet) DC.
Oxisma Raf., Sylva Tellur. 94. 1838.—TYPE: Oxisma arborescens(Aublet) Raf.
Axinanthera Karsten, Linnaea 30: 157. Post Apr 1859.—TYPE: Axinanthera macrophylla Karsten.
Heteroneuron Hooker f. in Bentham & Hooker, Gen. pl. 1(3): 768. 1867.—TYPE: Heteroneuron nigricans Hooker f.
Shrubs to trees; indumentum of slender to stout, elongate, unbranched, multicellular hairs, but sometimes nearly gla-
brous with age. Leaves mostly plinerved, but some basally-nerved, with abaxial epidermis smooth to minutely papillose. Inflorescences of solitary flowers, small cymes, or fascicles, in leaf axils or at nodes of older branches. Flowers 5–8-merous; hypanthium ± cylindrical, to hemispheric, campanulate, or conical; external calyx lobes absent; internal calyx lobes thick to thin, calyptrate (and rupturing circumscissily or irregu-larly in bud) or valvate, or the calyx lobes ± free in bud, the lobes well-developed and triangular to shallowly ovate or depressed semicircular, or nearly obsolete (i.e. not distinguish-able from the calyx tube); petals oblong, ovate to obovate, to rhombic, often clawed basally, usually ± pubescent, lacking an apical apiculum on the abaxial surface; anthers robust and fleshy, oblong to ovate-oblong, laterally compressed, with 1 or 2 minute pores, the thecae white, cream or yellow; ovary 5–15-loculate, the placentation axile to intruded axile, and placenta sometimes highly divided; style terete, glabrous; stigma expanded-capitate or peltate, clearly radiately ridged. Berries subglobose to hemispheric.
Species Requiring New Combinations—
Bellucia egensis (DC.) Penneys, Michelangeli, Judd, and Almeda, comb. nov. Myriaspora egensis DC., Prod. 3. 165. 1828.
Myriaspora decipiens Naud. Ann. Sci. Nat., Bot. sér: 16. 1851.

2010] PENNEYS ET AL.: HENRIETTEEAE (MELASTOMATACEAE) 797
Bellucia gracilis (Renner) Penneys, Michelangeli, Judd, and Almeda, comb. nov. Loreya gracilis Renner, Mem. New York Bot. Gard. 50: 45. 1989.
Bellucia klugii (Renner) Penneys, Michelangeli, Judd, and Almeda, comb. nov. Loreya klugii Renner, Mem. New York Bot. Gard. 50: 52. 1989.
Bellucia nigricans (Hooker f.) Penneys, Michelangeli, Judd, and Almeda, comb. nov. Loreya nigricans (Hooker f.) Triana, Trans. Linn. Soc. 28. 142. 1871. Heteroneuron nigri-cans Hooker f., Benth. and Hook. f. Gen. 1. 768. 1867.
Bellucia ovata (O. Berg ex Triana) Penneys, Michelangeli, Judd, and Almeda, comb. nov. Loreya ovata O. Berg ex Triana, Trans. Linn. Soc. 28. 142. 1871.
Loreya minor Cogniaux in Martius, Mart. Fl. Bras. 14. 4. 522. 1888.
Bellucia riparia (Renner) Penneys, Michelangeli, Judd, and Almeda, comb. nov. Loreya riparia Renner, Mem. New York Bot. Gard. 50: 46. 1989.
Bellucia strigosa (Gleason) Penneys, Michelangeli, Judd, and Almeda, comb. nov. Loreya strigosa Gleason, Bull. Torr. Bot. Cl. 60: 385. 1933.
Loreya quadrifolia Gleason, Bull. Torr. Bot. Cl: 60. 386. 1933.
Bellucia subandina (Wurdack) Penneys, Michelangeli, Judd, and Almeda, comb. nov. Loreya subandina Wurdack, Phytologia, 18: 161.1969.
Bellucia wurdackiana (Renner) Penneys, Michelangeli, Judd, and Almeda, comb. nov. Loreya wurdackiana Renner, Mem. New York Bot. Gard. 50: 42. 1989.
HENRIETTEA DC., Prod. 3. 178. 1828.—TYPE: Henriettea succosa(Aubl.) DC. Melastoma succosa Aubl.
Henriettella Naudin, Ann. Sci. Nat., Bot. sér. 3, 18: 107. 1852.—TYPE: Henriettella seemanii Naudin.
Llewelynia Pittier, Bol. Soc. Venez. Cienc. Nat. 5. 307. 1939.—TYPE: Llewelynia williamsii Pittier.
Shrubs to trees; indument of slender to stout, elongate, multicellular hairs with radiate to stellate-globular basal por-tion, or derived types, e.g. stellate hairs, peltate scales, elon-gate hairs with minute projections near base, or elongate hairs with a suprabasal expanded region and narrowed basal attachment. Leaves usually plinerved, with abaxial epidermis smooth. Inflorescences usually fascicles, occasionally 3-flow-ered cymes, or reduced to a solitary flower, usually at nodes of older branches, but occasionally in leaf axils. Flowers 4–6-merous; hypanthium ± cylindrical, to hemispheric, or campanulate; external calyx lobes present, conspicuous to inconspicuous; internal calyx lobes ± thin, calyptrate and opening circumscissilly or longitudinally and irregularly, or the lobes distinct, elongate-triangular to triangular or only shallowly so, to nearly obsolete and the calyx ± “open” in bud; petals oblong, ovate to obovate, to rhombic, often nar-rowed basally, usually ± pubescent, usually with an apical apiculum on the abaxial surface; anthers nonfleshy, attenuate-rostrate to elongate-ovate, or shorter and oblong or obovate, laterally compressed to nearly terete, with 1 or 2 pores, these minute, to greatly expanded and then ± rimose, the thecae cream, yellow, or purple; ovary (3-) 4–6-loculate, the placen-
tation axile to intruded axile, and placenta not divided; style terete, glabrous or pubescent; stigma ± truncate, nonridged or only very obscurely so. Berries globose to hemispheric.
Species Requiring New Combinations—
Henriettea boliviensis (Cogn.) Penneys, Michelangeli, Judd, and Almeda, comb. nov. Henriettella boliviensis Cogn. in DC., Monogr. Phan. 7: 1045. 1891.
Henriettea bracteosa (Wurdack) Penneys, Michelangeli, Judd, and Almeda, comb. nov. Henriettella bracteosa Wurdack, Phytologia 22: 417. 1972.
Henriettea caudata (Gleason) Penneys, Michelangeli, Judd, and Almeda, comb. nov. Henriettella caudata Gleason, Recueil Trav. Bot. Neerl. 32: 210. 1935.
Henriettea duckeana (Hoehne) Penneys, Michelangeli, Judd, and Almeda, comb. nov. Henriettella duckeana Hoehne, Anexos Mem. Inst. Butantan, Sec. Bot. 1, fasc. 5: 170, pl. 20. 1922.
Henriettea fissanthera (Gleason) Penneys, Michelangeli, Judd, and Almeda, comb. nov. Henriettella fissantheraGleason, Phytologia 1: 35. 1933.
Henriettea glabra (Vell.) Penneys, Michelangeli, Judd, and Almeda, comb. nov. Melastoma glabra Vell., Fl. Flum. 4: 171. Table 121. 1829. Henriettella glabra (Vell.) Cogn. in Mart., Fl. Bras. (Martius) 14, pt. 4: 538. 1888.
Henriettea cogniauxiana Penneys, Michelangeli, Judd, and Almeda, comb. et nom. nov., for Henriettella glaziovianaCogn. in Mart., Fl. Bras. (Martius) 14, pt. 4: 539, pl. 114. 1888. A new name for this species is needed because the epithet is occupied by Henriettea glazioviana Cogn. in Mart., Fl. Bras. (Martius) 14, pt. 4: 532. 1888.
Henriettea goudotiana (Naudin) Penneys, Michelangeli, Judd, and Almeda, comb. nov. Henriettella goudotianaNaudin, Ann. Sci. Nat., Bot. sér. 3, 18: 108. 1852.
Henriettea heteroneura (Gleason) Penneys, Michelangeli, Judd, and Almeda, comb. nov. Henriettella heteroneuraGleason, Mem. New York Bot. Gard. 8: 143. 1953.
Henriettea hondurensis (Wurdack) Penneys, Michelangeli, Judd, and Almeda, comb. nov. Henriettella hondurensisWurdack, Wrightia 6: 22. 1978.
Henriettea ininiensis (Wurdack) Penneys, Michelangeli, Judd, and Almeda, comb. nov. Henriettella ininiensisWurdack, Phytologia 64: 301. 1988.
Henriettea lawrancei (Gleason) Penneys, Michelangeli, Judd, and Almeda, comb. nov. Henriettella lawrancei Gleason, Phytologia 1: 31. 1933.
Henriettea lundellii (Wurdack) Penneys, Michelangeli, Judd, and Almeda, comb. nov. Henriettella lundellii Wurdack, Wrightia 6: 21. 1978.
Henriettea maguirei (Wurdack) Penneys, Michelangeli, Judd, and Almeda comb nov. Henriettella maguirei Wurdack, Phytologia 13: 80. 1966.
Henriettea manarae (Wurdack) Penneys, Michelangeli, Judd, and Almeda, comb. nov. Henriettella manarae Wurdack, Phytologia 24: 293. 1972.

798 SYSTEMATIC BOTANY [Volume 35
Henriettea odorata (Markgr.) Penneys, Michelangeli, Judd, & Almeda comb. nov. Henriettella odorata Markgr., Notizbl. Bot. Gart. Berlin-Dahlem 15: 380. 1941.
Henriettea ovata (Cogn.) Penneys, Michelangeli, Judd, and Almeda, comb. nov. Henriettella ovata Cogn. in Mart., Fl. Bras. (Martius) 14, pt. 4: 540, pl. 115. 1888.
Henriettea prancei (Wurdack) Penneys, Michelangeli, Judd, and Almeda, comb. nov. Henriettella prancei Wurdack, Phytologia 24: 206. 1972.
Henriettea rimosa (Wurdack) Penneys, Michelangeli, Judd, and Almeda, comb. nov. Henriettella rimosa Wurdack, Acta Bot. Venez. 4(1–4): 65. 1970.
Henriettea steyermarkii (Wurdack) Penneys, Michelangeli, Judd, and Almeda, comb. nov. Henriettella steyermarkiiWurdack, Bol. Soc. Venez. Ci. Nat. 25: 54. 1963.
Henriettea tachirensis (Wurdack) Penneys, Michelangeli, Judd, and Almeda, comb. nov. Henriettella tachirensisWurdack, Phytologia 55: 143. 1984.
Henriettea tobagensis (Wurdack) Penneys, Michelangeli, Judd, and Almeda, comb. nov. Henriettella tobagensisWurdack, Phytologia 7: 243. 1960.
Henriettea tovarensis (Cogn.) Penneys, Michelangeli, Judd, and Almeda, comb. nov. Henriettella tovarensis Cogn. in DC., Monogr. Phan. 7: 1044. 1891.
Henriettea trachyphylla (Triana) Penneys, Michelangeli, Judd, and Almeda, comb. nov. Henriettella trachyphyllaTriana, Trans. Linn. Soc. London 28: 144. 1871 [1872].
Henriettea williamsii (Pittier) Penneys, Michelangeli, Judd, and Almeda, comb. nov. Llewelynia williamsii Pittier, Bol. Soc. Venez. Ci. Nat. 5: 308. 1939.
KIRKBRIDEA Wurdack, Brittonia 28: 141. 1976.—TYPE: Kirkbridea tetramera Wurdack.
Shrubs to small trees; indument of appressed to incurved, slender, smooth to setulose hairs 1–4 mm, inconspicuously caducous-glandular. Leaves 5–7-plinerved or somewhat pseudoplinerved, membranaceous, ovate to oblong elliptic, apex acuminate to mucronate, base acute to obtuse, margins entire to distinctly ciliate-serrate. Inflorescences pedunculate, (1–)3(–5)–flowered, paired in leaf axils. Flowers pedicellate, 4–5-merous; bracteoles subpersistent; hypanthium terete; external calyx lobes present, conspicuous; internal calyx lobes apiculate-prolonged to acuminate; petals oblong-ovate, api-culate, unguiculate, abaxial surface with a line of strigose hairs; stamens dimorphic, thecae subulate, uniporose, pore ventrally inclined; ventral anther connective appendages paired, elongate, apically emarginate (larger stamens), or blunt (smaller stamens); ovary 4–5-loculate, 1/2–2/3 inferior, densely crowned with sericeous-strigose hairs; style glabrous; stigma subcapitate. Fruit a berry.
No new combinations are required under this genus.
Acknowledgments . We thank the following individuals for provid-ing plant material: Mac Alford, Chris Baraloto, Eldis Bécquer-Granados, Mario Blanco, J. Dan Skean, Jr., and Karen Redden. We also would like to thank two anonymous reviewers for their helpful comments and insights on this manuscript. We acknowledge the following institutions: CAS, FLAS, JBSD, NY, S, and US for their loans and processing of her-barium material consulted in connection with this research. Funding for
this research was provided by the National Science Foundation (grants DEB-0508582 to DSP, DEB-0515665 to FAM, and DEB-0515636 to WSJ). The Lewis B. and Dorothy Cullman program for Molecular Systematics Studies at The New York Botanical Garden supported sequencing efforts, and the Center for Comparative Genomics at the California Academy of Sciences provided computer support.
Literature Cited
Almeda, F. 1997 . Chromosome numbers and their evolutionary sig-nificance in some neotropical and paleotropical Melastomataceae . BioLlania. Edición Especial No. 6: 167 – 190 .
Almeda, F. 2001 . Melastomataceae . Pp. 1339 – 1419 in Flora de Nicaragua vol. 2 , eds. W. D. Stevens , C. Ulloa Ulloa , A. Pool , and O. M. Montiel . Monographs in Systematic Botany vol. 85. St. Louis : Missouri Botanical Garden .
Almeda, F. 2009 . Melastomataceae . Pp. 164 – 338 in Flora Mesoamericana vol. 4 , eds. G. Davidse , M. Sousa-Sanchez , S. Knapp , and F. Chiang . Mexico City : Universidad Nacional Autónoma de México .
Almeda, F. and T. I. Chuang . 1992 . Chromosome numbers and their sys-tematic significance in some Mexican Melastomataceae . SystematicBotany 17: 583 – 593 .
Amorim, A. A., R. Goldenberg , and F. A. Michelangeli . 2009 . A new species of Physeterostemon (Melastomataceae) from Bahia, Brazil, with notes on the phylogeny of the genus . Systematic Botany 34: 324 – 329 .
Andrade, P. M., E. R. Forni-Martins , and F. R. Martins . 2007a . Size structure and fertility in an Eriocnema fulva Naudin (Melastomataceae) popula-tion in Southeastern Brazil . Brazilian Journal of Biology 67: 685 – 693 .
Andrade, P. M., E. R. Forni-Martins , and F. R. Martins . 2007b . Reproductive system of Eriocnema fulva Naudin (Melastomataceae), an endemic species of Minas Gerais state, SE Brazil . Brazilian Journal of Biology 67: 313 – 319 .
Baas, P. 1981 . A note on stomatal types and crystals in the leaves of Melastomataceae . Blumea 27: 475 – 479 .
Baillon, H. E. 1877 . Monographie des Mélastomacées . Pp. 1 – 65 in Histoire des plantes vol. 7 . Paris : Hachette .
Baldwin, B. G. 1992 . Phylogenetic utility of the internal transcribed spacers of nuclear ribosomal DNA in plants: an example from the Compositae . Molecular Phylogenetics and Evolution 1: 3 – 16 .
Bécquer-Granados, E. R., K. M. Neubig , W. S. Judd , F. A. Michelangeli , J. R. Abbott , and D. S. Penneys . 2008 . Preliminary molecular phyloge-netic studies in Pachyanthus (Miconieae, Melastomataceae) . BotanicalReview 74: 37 – 52 .
Bentham, G. and J. D. Hooker . 1867 . Melastomaceae . Pp. 725 – 773 in GeneraPlantarum vol. 1 . London : Reeve and Co .
Britton, N. L. and P. Wilson . 1925 . Melastomataceae . Pp. 1 – 18 in Scientificsurvey of Porto Rico and the Virgin Islands , vol. 2 . New York : New York Academy of Sciences .
Clausing, G. and S. S. Renner . 2001 . Molecular phylogenetics of Melastomataceae and Memecylaceae: implications for character evo-lution . American Journal of Botany 88: 486 – 498 .
Clausing, G., K. Meyer , and S. S. Renner . 2000 . Correlations among fruit traits and evolution of different fruits within Melastomataceae . Botanical Journal of the Linnean Society 133: 303 – 326 .
Cogniaux, A. 1891 . Mélastomacées . Pp. 1 – 1256 in Monographiae Phaner-ogamarum vol. 7 . eds. A. P. de Candolle , and C. de Candolle . Paris : G. Masson .
de Candolle, A. P. 1828 . Melastomaceae . Pp. 99 – 202 in Prodromus systematis naturalis regni vegetabilis vol 3 . ed. A. P. de Candolle . Paris : Treuttel & Würtz .
DeWalt, S. J. and J. L. Hamrick . 2004 . Genetic variation of introduced Hawaiian and native Costa Rican populations of an invasive tropical shrub, Clidemia hirta (Melastomataceae) . American Journal of Botany 91: 1155 – 1162 .
Doyle, J. J. and J. L. Doyle . 1987 . A rapid DNA isolation procedure for small quantities of fresh leaf tissue . Phytochemical Bulletin 19: 11 – 15 .
Felsenstein, J. 1985 . Confidence limits on phylogenies: an approach using the bootstrap . Evolution 39: 783 – 791 .
Fritsch, P. W., F. Almeda , S. S. Renner , A. B. Martins , and B. C. Cruz . 2004 . Phylogeny and circumscription of the near-endemic Brazilian tribe Microlicieae (Melastomataceae) . American Journal of Botany 91: 1105 – 1114 .
Gleason, H. A. 1932 . A synopsis of the Melastomataceae of British Guiana . Brittonia 1: 127 – 184 .
Gleason, H. A. 1958 . Flora of Panama: Melastomataceae . Annals of the Missouri Botanical Garden 45: 203 – 304 .

2010] PENNEYS ET AL.: HENRIETTEEAE (MELASTOMATACEAE) 799
Gleason, H. A. and A. C. Smith . 1933 . Plantae Krukovianae (Concluded) . Bulletin of the Torrey Botanical Club 60: 379 – 396 .
Goldenberg, R., D. S. Penneys , F. Almeda , W. S. Judd , and F. A. Michelangeli . 2008 . Phylogeny of Miconia (Melastomataceae): pat-terns of stamen diversification in a megadiverse neotropical genus . International Journal of Plant Sciences 169: 963 – 979 .
Grisebach, A. H. R. 1864 . Melastomaceae . Pp. 243 – 269 in Flora of the British West Indian Islands . London : L. Reeve .
Howard, R. A. 1989 . Melastomataceae . Pp. 532 – 579 in Flora of the Lesser Antilles vol. 5 . Jamaica Plain, Massachusetts : Arnold Arboretum, Harvard University .
Judd, W. S. 1986 . Taxonomic placement of Calycogonium squamulosum(Melastomataceae: Miconieae) . Brittonia 38: 238 – 242 .
Judd, W. S. 1989 . Taxonomic studies in the Miconieae (Melastomataceae). III. Cladistic analysis of axillary-flowered taxa . Annals of the Missouri Botanical Garden 76: 476 – 495 .
Judd, W. S. 2007 . Revision of Miconia sect. Chaenopleura (Miconieae, Melastomataceae) in the Greater Antilles . Systematic Botany Monographs 81: 1 – 235 .
Judd, W. S. and J. D. Skean Jr . 1991 . Taxonomic studies in the Miconieae (Melastomataceae). IV. Generic realignments among terminal-flowered taxa . Bulletin of the Florida Museum of Natural History 36: 25 – 84 .
Judd, W. S., J. D. Skean Jr. , D. S. Penneys , and F. A. Michelangeli . 2008 . A new species of Henriettea (Melastomataceae) from the Sierra de Baoruco, the Domincan Republic . Brittonia 60: 217 – 227 .
Kadereit, G. 2006 . Revision of Plethiandra Hook. f.: a polystaminate, East Asian genus of Melastomataceae . Edinburgh Journal of Botany 62: 127 – 144 .
Krasser, F. 1898 . Melastomataceae . Pp. 130 – 199 in Die Natürlichen Pflanzenfamilien vol. 3 , eds. A. Engler and K. Prantl . Leipzig : Verlag .
Liogier, H. A. 1995 . Melastomataceae . Pp. 17 – 67 in Descriptive flora of Puerto Rico and adjacent islands vol 4 , ed. H. A. Liogier . Editorial de La Universidad de Puerto Rico. Rio Piedras : Universidad de Puerto Rico .
Macbride, J. F. 1941 . Flora of Peru (Melastomataceae) . Fieldiana. Botany 13: 249 – 521 .
Martin, C. V., D. Little , R. Goldenberg , and F. A. Michelangeli . 2008 . A preliminary phylogenetic analysis of the polyphyletic genus Leandra (Miconieae, Melastomataceae) . Cladistics 24: 315 – 327 .
Maxwell, J. F. and J. F. Veldkamp . 1990a . Notes on the Astronieae (Melastomataceae) – I. Astrocalyx, Astronia . Blumea 35: 71 – 114 .
Maxwell, J. F. and J. F. Veldkamp . 1990b . Notes on the Astronieae (Melastomataceae) – II. Astronidium, Beccarianthus . Blumea 35: 115 – 164 .
Mendoza, H. and B. Ramirez . 2006 . Guía ilustrada de géneros de Melastomataceae y Memecylaceae de Colombia . Bogotá : Instituto de Investigación de Recursos Biológicos Alexander von Humboldt; Univerisad del Cauca .
Michelangeli, F. A., D. S. Penneys , J. Giza , D. Soltis , M. H. Hils , and J. D. Skean Jr . 2004 . A preliminary phylogeny of the tribe Miconieae (Melastomataceae) based on nrITS sequence data and its implications on inflorescence position . Taxon 53: 279 – 290 .
Michelangeli, F. A., W. S. Judd , D. S. Penneys , J. D. Skean Jr. , E. R. Bécquer-Granados , R. Goldenberg , and C. V. Martin . 2008 . Multiple events of dispersal and radiation of the tribe Miconieae (Melastomataceae) in the Caribbean . Botanical Review 74: 53 – 77 .
Morales-P., M. E. and D. S. Penneys . 2010 . New species of Chalybea Naud. and Huilaea Wurd. (Melastomataceae) . Brittonia 62: 26 – 34 .
Naudin, C. V. 1849–1853 . Melastomacearum monographicae descriptio-nis . Annales des Sciences Naturelles, Botanique, Series III , tom. xii–xviii, consolidated reprint.
Olmstead, R. G. and A. Sweere . 1994 . Combining data in phylogenetic sys-tematics: an empirical approach using three molecular data sets in the Solanaceae . Systematic Biology 43: 467 – 481 .
Penneys, D. S. 2007 . Phylogeny and character evolution in the Blakeeae (Melastomataceae): Neotropical hemiepiphytes with mite and ant domatia . Ph. D. dissertation. Gainesville, Florida : University of Florida .
Penneys, D. S. and W. S. Judd . 2005 . A systematic revision and cladistic analysis of Charianthus (Melastomataceae) using morphological and molecular characters . Systematic Botany 30: 559 – 584 .
Penneys, D. S. and W. S. Judd . In press. Phylogenetics and morphology in the Blakeeae (Melastomataceae) . International Journal of Plant Sciences .
Penneys, D. S., M. W. Whitten , N. H. Williams , and W. S. Judd . 2004a . Huilaeaand the Blakeeae (Melastomataceae): phylogenetic relationships recon-sidered . Abstract ID no. 136. Botany 2004. Alpine diversity: adapted to the peaks. July 31- August 5. Salt Lake City, Utah. Available at:
http://2004.botanyconference.org/engine/search/index.php?func=detail&aid=136.
Penneys, D. S., F. A. Michelangeli , W. S. Judd , and J. D. Skean Jr . 2004b . Henrietteeae, a new tribe of Neotropical Melastomataceae . Abstract ID no. 226. Botany 2004. Alpine diversity: adapted to the peaks. July 31- August 5. Salt Lake City, Utah. Available at: http://2004.botanyconference.org/engine/search /index.php?func=detail&aid=226 .
Posada, D. and K. A. Crandall . 1998 . MODELTEST: Testing the model of DNA substitution . Bioinformatics 14: 817 – 818 .
Proctor, G. R. 1972 . Melastomataceae . Pp. 529 – 549 in Flowering plants of Jamaica , ed. C. D. Adams . Mona, Jamaica : University of the West Indies .
Rambaut, A. 1996 . Se-Al: Sequence Alignment Editor . Available at http://evolve.zoo.ox.ac.uk/ .
Renner, S. S. 1989 . Systematic studies in the Melastomataceae: Bellucia,Loreya , and Macairea . Memoirs of the New York Botanical Garden 50: 1 – 112 .
Renner, S. S., G. Clausing , and K. Meyer . 2001 . Historical biogeography of Melastomataceae: the roles of Tertiary migration and long-distance dispersal . American Journal of Botany 88: 1290 – 1300 .
Solereder, H. 1908 . Systematic anatomy of the dicotyledons . Oxford : Clarendon Press .
Solt, M. L. and J. J. Wurdack . 1980 . Chromosome numbers in the Melastomataceae . Phytologia 47: 199 – 230 .
Standley, P. C. and L. O. Williams . 1963 . Flora of Guatemala (Melastomaceae) . Fieldiana. Botany 24: 407 – 525 .
Stone, R. D. 2005 . Phylogeny of major lineages in Melastomataceae, subfamily Olisbeoideae: utility of nuclear glyceraldehyde 3-phos-phate dehydrogenase ( GapC ) gene sequences . Systematic Botany 31: 107 – 121 .
Sun, Y., D. A. Skinner , G. H. Liang , and S. H. Hulbert . 1994 . Phylogenetic analysis of Sorghum and related taxa using internal transcribed spac-ers of nuclear ribosomal DNA . Theoretical and Applied Genetics 89: 26 – 32 .
Swofford, D. L. 2002 . PAUP* 4.0b10 Phylogenetic Analysis Using Parsimony (*and other methods) . Sunderland : Sinauer Associates, Inc .
Triana, J. 1866 . Dispositio Melastomacearum . Bulletin du Congrès International de Botanique d’Horticulture réuni à Amsterdam 457 – 461 .
Triana, J. 1871 . Les Mélastomacées . Transactions of the Linnaean Society of London. Botany 28: 1 – 188 .
van Tieghem, M. Ph . 1891a . Classification anatomique des Mélastomacées . Bulletin de la Société Botanique de France 38: 114 – 124 .
van Tieghem, M. Ph . 1891b . Sur la structure et les affinities des Mémécylées . Annales des Sciences Naturelles 13: 23 – 92 .
van Tieghem, M. Ph . 1892 . Deuxième addition aux recherches sur la struc-ture et les affinities des Mélastomacées . Annales des Sciences Naturelles 7: 369 – 380 .
ter Welle, B. J. H. and A. M. W. Mennega . 1977 . On the presence of large styloids in the secondary xylem of the genus Henriettea(Melastomataceae) . IAWA Bulletin 2: 31 – 35 .
ter Welle, B. J. H. and J. Koek-Noorman . 1981 . Wood anatomy of the Neotropical Melastomataceae . Blumea 27: 335 – 394 .
Williams, L. O. 1963 . Tropical American plants V . Fieldiana. Botany 29: 545 – 586 .
Wurdack, J. J. 1957 . Certamen Melastomataceis IV . Brittonia 9: 101 – 109 . Wurdack, J. J. 1965 . Certamen Melastomataceis IX . Phytologia 11: 377 – 400 . Wurdack, J. J. 1972 . Certamen Melastomataceis XX . Phytologia 24:
195 – 208 . Wurdack, J. J. 1973 . Melastomataceae . Pp. 1 – 819 in Flora de Venezuela vol 8 ,
ed. T Lasser . Caracas : Instituto Botánico . Wurdack, J. J. 1976 . Endemic Melastomataceae of the Sierra Nevada de
Santa Marta, Colombia . Brittonia 28: 138 – 143 . Wurdack, J. J. 1980 . Melastomataceae . Pp. 1 – 406 in Flora of Ecuador vol 13 ,
eds. G. Harling and B. Sparre . Göteborg : University of Göteborg . Wurdack, J. J. 1986 . Atlas of hairs for Neotropical Melastomataceae .
Smithsonian Contributions to Botany 63: 1 – 80 . Wurdack, J. J., T. Morley , and S. S. Renner . 1993 . Melastomataceae . Pp. 1 –
427 in Flora of the Guianas vol 99 , ed. A. R. A. G. van Rijn . Koenigstein : Koeltz .
Zwickl, D. J. 2006 . Genetic algorithm approaches for the phylogenetic analysis of large biological sequence datasets under the maximum likelihood criterion . Ph.D. dissertation. Austin : The University of Texas at Austin .
Appendix 1 . Taxon sampling, collector name and number, and GenBank accession information for specimens used in this study. All vouchers are deposited at A, ALBC, CAS, FLAS, INBIO, MJG, MO, NY, PMA, PORT, QCNE, VEN, US, and/or USZ. The information is listed

800 SYSTEMATIC BOTANY [Volume 35
as: taxon, collector, collector number, GenBank accession numbers (ITS, ndhF ). Newly generated sequences begin with GU968 .
Arthrostemma ciliatum Pav. ex D. Don — Skean 3806 (FLAS); AY460429, GU968804. Astronia smilacifolia Triana ex C. B. Clarke — Clausing 189 (MJG); EF683153, AF215549. Bellucia grossulariodes (L.) Triana —Redden 1111 (US); GU968787, EU711372. Bellucia pentamera Naud. — Michelangeli 690 (NY); GU968788, GU968805. Blakea chlorantha Almeda — Penneys 1512 (FLAS); GU968789, GU968806. Blakea gracilis Hemsl. — Boyle 6631 (INBIO); AY460445, GU968807. Blakea schlimii Triana —Michelangeli 726 (NY); AY460441, EU711373. Charianthus alpinus (Sw.) R.A. Howard — Penneys 1301 (FLAS); AY460463, EU055902. Clidemia ciliata D. Don — Michelangeli 700 (NY); AY460472, EU055907. Clidemia guadalupensis Griseb. — Penneys 1313 (FLAS); EF418802, EU055917. Clidemia rubra (Aubl.) Mart. — Michelangeli 384 (VEN); AY460481, AF215579. Conostegia bigibbosa Cogn. — Michelangeli 841 (INBIO); AY460485, EU055932. Desmoscelis villosa (Aubl.) Naud. — Michelangeli 827 (NY); AY460430, EU711374. Eriocnema fulva Naud. — Almeda 8416(CAS); EF418811, AY553781. Graffenrieda latifolia (Naud.) Triana —Penneys 1303 (FLAS); EF683143, EU055943. Graffenrieda moritzianaTriana — Michelangeli 832 (NY); AY460451, EU055944. Henriettea gib-berosa (Urb.) Alain — Bécquer-G. 81149 (FLAS); GU968790, GU968808. Henriettea macfadyenii (Triana) Alain — Skean 3795 (FLAS); GU968791, GU968809. Henriettea martiusii (DC.) Naud. — Michelangeli 412 (VEN); AY460564, EU711375. Henriettea multiflora Naud. — Redden 2129 ; GU968792, GU968810. Henriettea ramiflora (Sw.) DC. — Bécquer-G. 82451 (FLAS); GU968793, GU968811. Henriettea spruceana Cogn. —Michelangeli 390 (NY); AY460565, GU968812. Henriettea squamata (Alain) Alain — Bécquer-G. 82268 (FLAS); GU968794, GU968813. Henriettea squamulosa (Cogn.) Judd — Skean 3807 (FLAS); EF683151, GU968814. Henriettea succosa (Aubl.) DC. — Michelangeli 685 (NY); AY460568, GU968815. Henriettea tuberculosa (Donn.Sm.) L. O. Williams — (ITS)
Almeda 7663 (CAS); AY460566, ( ndhF ) Blanco 2274 (FLAS); GU968816. Henriettea uniflora Judd, Skean, Penneys, & Michelangeli — Skean 4298 (FLAS); GU968795, GU968817. Henriettella rimosa Wurdack —Michelangeli 684 (NY); AY460567, GU968818. Henriettella sessilifolia(L.) Alain — Michelangeli 798 (VEN); GU968796, GU968819. Henriettellaaff. tovarensis Cogn. — Michelangeli 741 (VEN); GU968797, GU968820. Huilaea ecuadorensis Wurdack — Penneys 1589 (FLAS); GU968798, GU968821. Leandra mexicana (Naud.) Cogn. — Skean 4005 (FLAS); GU968799, AF215580. Loreya arborescens DC. — (ITS) Baraloto 2823(FLAS); EF683147, ( ndhF ) Renner 2152B (MO); EU711377. Loreya mespi-loides Miq. — Michelangeli 606 (NY); GU968800, GU968822. Loreya spruceana Triana — (ITS) Penneys 1854 (FLAS); EF683149, ( ndhF ) Renner 237 (MO); GU968823. Loreya subandina Wurdack — Alford 3107 (BH); AY460563, GU968824. Maieta guianensis Aubl. — Michelangeli 499 (NY); AY460498, AF215581. Marcetia taxifolia (A. St.-Hil.) DC. — Michelangeli 680 (NY); AY460431, GU968825. Meriania macrophylla (Benth.) Triana — Michelangeli 829 (NY); AY460455, AM235414. Meriania subumbellataCogn. — Michelangeli 819 (NY); AY460457, EU055972. Miconia dode-candra Cogn. — Michelangeli 758 (NY); EU055769, EU056026. Miconia leiotricha Wurdack — Michelangeli 775 (VEN); AY460461, EU056049. Miconia macrodon (Naud.) Wurdack — Michelangeli 797 (NY); AY460516, EU056058. Nepsera aquatica (Aubl.) Naud. — Struwe 1158(NY); AY460433, AF215569. Pternandra caerulescens Jack. — (ITS) Chen 543 (FLAS); EF683154, ( ndhF ) Clausing s. n. (MJG); AF215558. Pternandra echinata Wall. — (ITS) Chen 542 (FLAS); EF683155, ( ndhF ) Clausing s.n. (MJG); AF215559. Pternandra multiflora Cogn. — Clausing 142(MJG); GU968801, AF215560. Rhynchanthera grandiflora DC. — (ITS) Michelangeli 541 (NY); AY460436, ( ndhF ) Prévost 3281 (NY); AF215584. Tetrazygia bicolor Cogn. — Zomlefer 739 (FLAS); AY460543, EU056130. Tetrazygia fadyenii Hook. — Williams & Whitten 99106 (FLAS); AY460545, EU056133. Tibouchina urvilleana (DC.) Cogn. — (ITS) Hils 1382 (FLAS); AY460439, ( ndhF ) Renner s.n. (MJG); AF272820. Tococa guianensis Aubl. — Michelangeli 703 (NY); AY460554, EU056136.