differentiated evolutionary pathways in haemulidae (perciformes): karyotype stasis versus...
TRANSCRIPT

RESEARCH PAPER
Differentiated evolutionary pathways in Haemulidae(Perciformes): karyotype stasis versus morphologicaldifferentiation
C. C. Motta Neto • P. A. Lima-Filho •
W. C. Araujo • L. A. C. Bertollo • W. F. Molina
Received: 8 July 2011 / Accepted: 20 September 2011
� Springer Science+Business Media B.V. 2011
Abstract Extensive phenotypic diversity (size, col-
ors and shapes) among species of Haemulidae is
practically dissociated from the conservative cytoge-
netic pattern observed in this family. Detailed analyses
indicate that karyotypic stasis is maintained even under
the scrutiny of different chromosome investigation
methods. Chromosomal banding patterns (endophe-
notype) of five Atlantic species are presented here:
Conodon nobilis, Pomadasys corvinaeformis, Haem-
ulon aurolineatum, H. plumierii and H. steindachneri,
obtained by incorporating the base analog 5-BrdU,
C-banding and staining with base-specific fluoro-
chromes. Despite a few chromosomal specificities,
relative karyotypic conservation was confirmed, cor-
roborating earlier studies on this family. On the other
hand, phenotypic patterns (exophenotype), identified
by geometric morphometrics contrast visibly with the
chromosomal conservation of this group. As such, the
evolutionary rates of chromosomes and body mor-
phology demonstrate clear asynchrony. Possible
causes of karyotypic stasis in Haemulidae are
discussed as well as the sharing of this condition with
other Perciformes.
Keywords Cytogenetics � Geometric
morphometrics � Evolutionary divergence �Chromosome stasis
Introduction
In many groups of marine fish, the speciation process
is not followed by significant karyotype differentia-
tion. This is observed in several Perciformes families
(Molina et al. 2002; Molina 2007), characterized by a
common, extensive and phyletically dispersed karyo-
type composed of 2n = 48 chromosomes predomi-
nantly acrocentric (Klinkhardt et al. 1995; Brum and
Galetti 1997). This karyotypic pattern, considered
basal for this fish order (Galetti et al. 2000), is also
shared by the family Haemulidae (Accioly and Molina
2007; Nirchio et al. 2007). Large panmictic popula-
tions, which would predispose to the maintenance of
intragroup genetic cohesion, or even intrinsic chro-
mosomal characteristics, could be related to karyo-
typic stasis seen in several fish groups (Molina 2007;
Araujo et al. 2010). In many cases, such as in
Haemulidae, this karyotypic stability contrasts with
conspicuous morphological patterns, especially in the
adult phases of development.
A large portion of fish species exhibits rela-
tively small chromosomes, poor in longitudinal bands
C. C. Motta Neto � P. A. Lima-Filho �W. C. Araujo � W. F. Molina (&)
Departamento de Biologia Celular e Genetica, Centro de
Biociencias, Universidade Federal do Rio Grande do
Norte, Natal, RN, Brazil
e-mail: [email protected]
L. A. C. Bertollo
Departamento de Genetica e Evolucao, Universidade
Federal de Sao Carlos, Sao Carlos, SP, Brazil
123
Rev Fish Biol Fisheries
DOI 10.1007/s11160-011-9236-4

and heterochromatic segments. Replication bands,
obtained by incorporating of the base analog
5-Bromo-20-deoxyuridine (5-BrdU) during DNA rep-
lication, may provide resolutive chromosomal char-
acters for identifying cryptic rearrangements, often not
identified by other methodologies (Kasahara 2009).
Indeed, this type of banding has been informative in
the karotypic analysis of a number of groups, such as
Cyprinidae (Zhang and Wu 1985; Hellmer et al.
1991); Salmonidae (Delany and Bloom 1984; Pendas
et al. 1993); Muraenidae (Salvadori et al. 2003),
Scorpaenidae (Giles et al. 1988) and several Charac-
iformes species (Bertollo et al. 1997; Maistro et al.
1999; Daniel-Silva and Almeida-Toledo 2005; Molina
and Galetti 2007), also contributing to phylogenetic
and evolutionary studies (Boron 2003; Sumner 2003).
Morphometric analyses, which enables identifica-
tion of the simultaneous variation of traits related to a
complex body structure (Rohlf and Marcus 1993;
Monteiro and Reis 1999), has contributed to evolu-
tionary biology (Blackith and Reyment 1971), popu-
lation and phylogenetic studies, sex differentiation as
well as intra and interspecific variation (Strauss and
Fuiman 1985; Ehlinger 1991; Vidales et al. 1997;
Fairbairn 1997; Baras 1999; Fulford and Rutherford
2000; Aguirre and Shervette 2005; Rapp Py-Daniel
and Cox Fernandes 2005). The association between
genetic and morphological data has been increasingly
used to understand the processes involved in phylo-
genetic diversification of different groups of organ-
isms (Cheverud 1988; Doebley and Stec 1993; Larson
1998).
The present study investigates chromosomal char-
acters in species of three genera of Haemulidae,
obtained by replicating bands, associated to body
patterns revealed by geometric morphometrics.
Materials and methods
Cytogenetic analyses were carried out with Conodon
nobilis (n = 10, 6 females, 3 males and 1 immature),
Pomadasys corvinaeformis (n = 12, 3 females, 7
females and 2 immatures) and three species of
Haemulon: H. aurolineatum (n = 9, 3 females and 6
males), H. plumierii (n = 8, two females, three males
and three immatures) and H. steindachneri (n = 8,
three females, three males and two immatures),
collected on the coast of Rio Grande do Norte state
(northeastern of Brazil). Samples of C. nobilis,
P. corvinaeformis, H. aurolineatum and H. plumierii
were obtained from two different geographic
areas (5�1301.7300S, 35�9057.8500W and 5�44050.5000S,
35�12010.5200W), while samples of H. steindachneri
were collected in an area further to the south
(5�13011.9700S, 35�2503.8200W).
Specimens were submitted to mitotic stimulation in
vivo with combined antigens (Molina 2001; Molina
et al. 2010), anesthetized with clove oil (Eugenol) and
sacrificed. Mitotic metaphases were obtained from the
anterior portion of the kidney, following the in vivo
preparation method described by Gold et al. (1990).
Heterochromatic chromosomal regions were identi-
fied by C-banding (Sumner 1972) and GC- or AT-rich
segments by CMA3/DAPI fluorochrome staining
(Barros-e-Silva and Guerra 2009). Briefly, slides aged
for 3 days were stained with CMA3 (0.1 mg/ml) for
60 min, restained with DAPI (1 lg/ml) for 30 min,
mounted in glycerol:McIlvaine buffer pH 7.0 (1:1) and
analyzed under epifluorescence microscope equipped
with the appropriate filter set.
Replication bands using the thymine analogue,
5-BrdU were obtained using methodology developed
by Giles et al. (1988). Specimens were submitted to
intraperitoneal inoculation of 5-BrdU (5 mg/ml in
0.9% NaCl solution) at a ratio of 1 ml/100 g of
body weight, 6 h before being sacrificed to obtain
mitotic chromosomes. FPG (Fluorochrome Photolysis
Giemsa) staining was used to reveal RBG (Replication
Bands by Bromodeoxyuridine using Giemsa) bands.
Chromosomal preparations, with base analogue incor-
poration, were stained with Hoescht 33258 solution
(Sigma; 1 mg of Hoescht in 1 ml of methanol and
100 ml of 0.5XSSC) for 40 min in a dark chamber,
washed in distilled water, recovered with a 2XSSC
film and irradiated with ultraviolet light (254 gm) at a
distance of 10 cm, for 1 h, then stained with a 5%
Giemsa solution diluted in phosphate buffer, pH 6.8.
Approximately thirty metaphases were analyzed for
specimen. The best metaphases were photographed
under an Olympus BX50 epifluorescence microscope
equipped with an Olympus DP70 digital image capture
system. The chromosomes acrocentrics were arranged
in the karyotype in descending order of size.
Morphometric analyses were conducted in the fol-
lowing adult specimen samples: C. nobilis (n = 30),
P. corvinaeformis (n = 24), H. aurolineatum (n = 50),
H. plumierii (n = 30) and H. steindachneri (n = 39).
Rev Fish Biol Fisheries
123
The left side of the body was photographed with a Sony
H10 digital camera (8.1 megapixels) mounted on a
tripod at standardized distance and position, eliminating
possible distortions in shape and body size. With respect
to body patterns, eleven common landmarks among
the species were defined. These were located on the
initial (1) and terminal (11) portion of the upper maxilla;
at the anterior (2) and posterior (3) portion at the base of
hard dorsal fin rays; final insertion at the base of the soft
dorsal fin; (4); posterior (5) and anterior (6) base of
the anal fin; insertion of ventral (7) and pectoral fins (8)
and posterior (9) and anterior (10) end of the ocular
cavity.
Procrustes analyses (Rohlf and Slice 1990) were
developed using TPSDig2 software (Rohlf 2006) to
locate landmarks (11 points). Coordinates represent-
ing the spatial position of each landmark were
overlapped using CoordGen software. Analyses of
canonical variables (CV) and MANOVA allowed
morphometric comparisons of the species. Allocation
and cluster tests, using the Jackknife method and
CVAGen6 software, were used to correctly classify
specimens into their respective species. A comparative
deformation matrix of morphological differences
among species was obtained from the mean distribu-
tion of canonic variables through the MorphoJ 1.02b
software (Klingenberg 2011).
Results
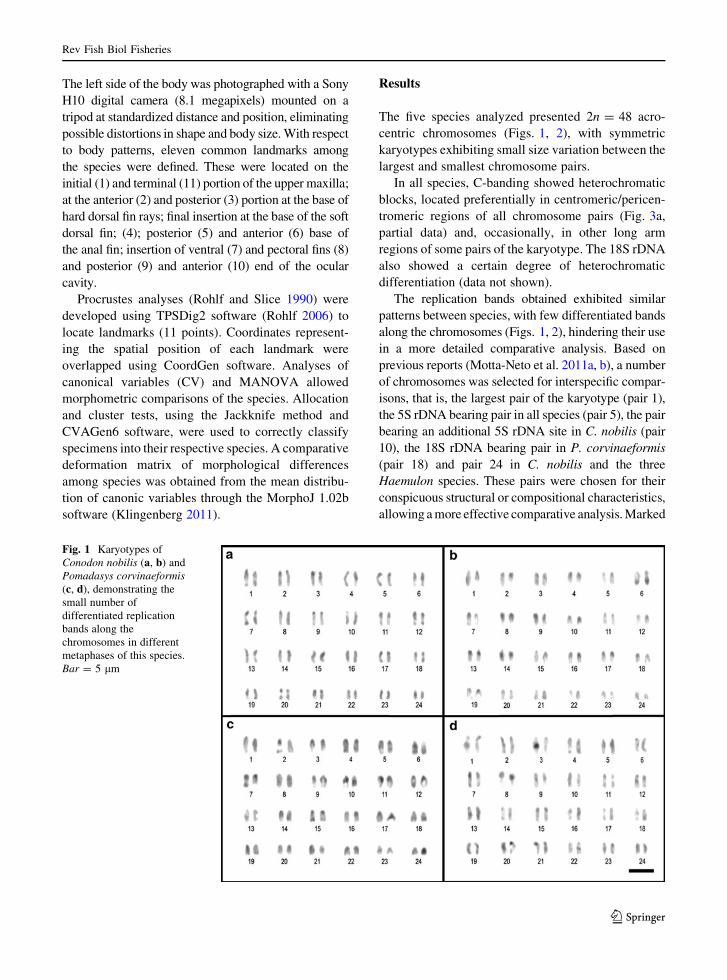
The five species analyzed presented 2n = 48 acro-
centric chromosomes (Figs. 1, 2), with symmetric
karyotypes exhibiting small size variation between the
largest and smallest chromosome pairs.
In all species, C-banding showed heterochromatic
blocks, located preferentially in centromeric/pericen-
tromeric regions of all chromosome pairs (Fig. 3a,
partial data) and, occasionally, in other long arm
regions of some pairs of the karyotype. The 18S rDNA
also showed a certain degree of heterochromatic
differentiation (data not shown).
The replication bands obtained exhibited similar
patterns between species, with few differentiated bands
along the chromosomes (Figs. 1, 2), hindering their use
in a more detailed comparative analysis. Based on
previous reports (Motta-Neto et al. 2011a, b), a number
of chromosomes was selected for interspecific compar-
isons, that is, the largest pair of the karyotype (pair 1),
the 5S rDNA bearing pair in all species (pair 5), the pair
bearing an additional 5S rDNA site in C. nobilis (pair
10), the 18S rDNA bearing pair in P. corvinaeformis
(pair 18) and pair 24 in C. nobilis and the three
Haemulon species. These pairs were chosen for their
conspicuous structural or compositional characteristics,
allowing a more effective comparative analysis. Marked
Fig. 1 Karyotypes of
Conodon nobilis (a, b) and
Pomadasys corvinaeformis(c, d), demonstrating the
small number of
differentiated replication
bands along the
chromosomes in different
metaphases of this species.
Bar = 5 lm
Rev Fish Biol Fisheries
123

interspecific conservation was generally observed in
the replication bands on these marker chromosomes,
corroborated by the similar pattern in their C-positive
bands and the co-location of GC-rich regions with 18S
rDNA sites (Fig. 3). The only GC-rich chromosomal
regions (CMA3?/DAPI-) corresponded to 18S rDNA
sites (NORs) present on pair 18 of P. corvinaeformis and
pair 24 of the remaining species.
Cluster analyses of canonical variables indicate that
the variance is distributed mainly by axis 1 (61.51%), 2
(21.39%), 3 (11.41%) and 4 (5.68%). Species belong-
ing to the genus Haemulon were perfectly discrimi-
nated amongst themselves, by both canonical axis 1
and 2 which, taken together, account for 82.90%
of total variance. The greatest morphometric proximity
was sequentially identified between H. plumierii
and both H. steindachneri and H. aurolineatum
(Fig. 4a). Body morphometrics showed signifi-
cant differences (MANOVA axis 1—Lambda =
0.0001, X2 = 1,585.43, df = 72, P \ 0.0001; Axis
2—Lambda = 0.0020, X2 = 998.93, df = 51,
P \ 0.0001), allowing correct discrimination of all
individuals and their respective species, applying the
Jackknife test. Main inter-species differences were
primarily related to head and body height measures.
Thin-plate spline deformation grids for the mean
measures of each species were examined to anatom-
ically describe inter-species shape variations (Fig. 4c).
Fig. 2 Karyotypes of
Haemulon aurolineatum(a, b), H. plumierii (c, d) and
H. steindachneri (e, f),showing the small number
of differentiated replication
bands along the
chromosomes in different
metaphases of these species.
Bar = 5 lm
Rev Fish Biol Fisheries
123

Discussion
The family Haemulidae is characterized by a symmet-
rical and conservative karyotypic pattern (Accioly and
Molina 2007; Nirchio et al. 2007), with little inter-
species chromosome variability. This conservation is
also found in different markers previously analyzed in
this group. In effect, all Haemulidae species analyzed
belonging to the genera Conodon, Pomadasys and
Haemulon exhibit reduced heterochromatic blocks,
located mainly in centromeric/pericentromeric regions
(Nirchio et al. 2007; Motta-Neto et al. 2011a, b),
following the common pattern detected for numerous
Perciformes species (Molina 2007). The same pattern
is repeated in the species investigated here, indicating
their limited applicability in karyotypic differentiation
analysis of Haemulidae. This contrasts with other fish
groups where C-band distribution in chromosomes
may represent of effective population markers (e.g.
Souza et al. 1996). Indeed, this homogeneous pattern in
heterochromatin differs substantially from other fish
groups, where this genome component had an impor-
tant role in karyotypic evolution, as occurs in different
Characiformes species (e.g. Vicari et al. 2008; Cioffi
et al. 2009), revealing polymorphisms and a high
degree of heterochromatin heterogeneity. This evi-
dence corroborates the idea that heterochromatin is not
characterized merely by its degree of condensation, but
also by its organization, molecular composition, asso-
ciated proteins and function (Sumner 2003; Horn and
Peterson 2006).
No base-specific richness was detected in the
studied species, except for nucleolus organizer regions
whose pattern is GC?. In lower invertebrates, ribo-
somal sites, particularly those related to NORs, exhibit
a positive response to GC-specific fluorochromes
(Schmid and Guttenbach 1988), often associated to
C-positive band segments in fish, as occurs in Haemu-
lidae. This occurrence, along with similar chromo-
somal bands obtained by digestion with a number of
restriction endonucleases (Motta-Neto et al. 2011a, b),
suggests marked heterochromatic homogeneity in
these species. A particularly interesting question that
remains unanswered is whether this uniform pattern,
related to repetitive DNA regions, would be associated
to simultaneous chromosomal rearrangements, as
proposed for Pomacentridae species (Molina and
Galetti 2002, 2004), as well as to karyotypic orthose-
lection processes of Perciformes (Molina 2007).
It has been suggested that the systematic occurrence
of certain rearrangements along the chromosomal
evolution by orthoselective processes could lead to
fixation of symmetrical karyotypes, exhibiting simi-
lar-sized chromosomes with the same morphology
Fig. 3 Chromosome pairs 1, 5, 10, 18 and 24 of Conodonnobilis (Cn), Pomadasys corvinaeformis (Pc), Haemulonaurolineatum (Ha), H. plumierii (Hp) and H. steindachneri(Hs), used for inter-species comparative marker analysis.
a C-banding showing the preferential location of C-positive
heterochromatin in the centromeric/pericentromeric region of
the chromosomes; b CMA3 fluorochrome staining depicting
GC-positive regions in pair 18 of P. corvinaeformis and pair 24
of the remaining species; c replication bands showing a similar
pattern among species. Bar = 5 lm
Rev Fish Biol Fisheries
123

(White 1973; King 1981). By contrast, similar kary-
otypes, extensively distributed among species of a
given group, could also indicate the relatively recent
divergence of this group, with insufficient time to
fixate particular chromosomal rearrangements (Sola
et al. 1981). Although this does not explain karyotypic
stasis of several marine families of Perciformes that
emerged in the Tertiary period, this situation cannot be
ruled out for species of Haemulon, whose diversifica-
tion is more recent (Cequea and Perez 1971).
It is well known that several chromosomal banding
techniques are less resolutive in lower than in higher
vertebrates. Thus, replication bands have been used as
a substitute for conventional banding G, Q or R
procedures, which are generally not informative in
fish. In several cases the patterns generated contribute
not only to karyotypic pairing and definition, but also
to sex chromosome analysis, detection of interspecific
homeologies, cytotaxonomy and phylogenetic inves-
tigations (Bertollo et al. 1997; Maistro et al. 1999;
Boron 2003; Daniel-Silva and Almeida-Toledo 2001,
2005; Molina and Galetti 2007; Araujo et al. 2010).
In the Haemulidae analyzed here, where karyotypes
are highly symmetrical with small chromosomes,
replication bands generated a common pattern, reflect-
ing only a slight variation in chromosomal framework
without conspicuous cytotaxonomic markers. How-
ever, in spite of this limitation, data suggest greater
replication band similarity among species of Haemu-
lon, as phylogenetically expected, evidenced by the
relatively higher number of bands.
Nevertheless, individual analysis of some poten-
tially equivalent chromosomes among species, such as
pairs 1, 5, 10, 18 and 24, allowed more effective
interspecific comparison, indicating the maintenance
of large homeologous blocks between them. Despite
the presence of similar structural and replication bands
among species, pair 1 exhibited variable size, around
2 lm in P. corvinaeformis and H. plumierii and 3 lm
in C. nobilis, H. aurolineatum and H. steindachneri,
two or three times larger than the smallest pairs,
respectively. Thus, despite greater karyotypic similar-
ities, some internal reorganization has also taken place
in the chromosomes during the evolutionary process.
Taken together, data obtained by different methodol-
ogies, such as replication banding, base-specific
fluorochrome staining, C-banding and restriction
endonuclease digestion patterns (Nirchio et al. 2007;
Accioly and Molina 2007; Motta-Neto et al. 2011a, b),
suggest that broad structural segments are shared by
Haemulidae genomes. This condition, which has been
indicated for this family, could correspond to a
karyotypic model also valid for other Perciformes
groups. Although information on the maintenance of
phylogenetically syntenic segments shared by Neo-
tropical species is still limited, available data for some
genera (Maistro et al. 1999; Souza et al. 1996, 2001;
Daniel-Silva and Almeida-Toledo 2001, 2005; Vicari
et al. 2008) indicate this may occur.
In contrast to cytogenetic patterns, the morpholog-
ical patterns of Haemulidae species, measured by
geometric morphometrics, demonstrated perfect inter-
species discrimination. The first two canonical vari-
ables explained 82.9% of total variance. Species of
Haemulon were discriminated in relation to both CV
Fig. 4 Geometric morphometrics of analyzed Haemulidae
species. a Species ordering on the first two canonical axes;
b landmarks used in morphometric analyses of the species
(bar = 2 cm); c deformation grid indicating shape variation
between consensus configurations for the species. Vectors show
the deformation direction at each landmark
Rev Fish Biol Fisheries
123

axes. However, it is suggested that the species
analyzed be separated into two groups, one composed
of congeneric species of Haemulon (H. aurolineatum,
H. plumierii and H. steindachneri), and the other, less
discriminated by canonical axis 2, consisting of
representatives of the remaining genera (C. nobilis
and P. corvinaeformis). Morphological data obtained
corroborate the general patterns described for these
species, in which Haemulon is characterized as having
long oblong bodies, while C. nobilis and P. corvinae-
formis have a robust to moderately robust elongated
body (Lindeman and Toxey 2002). Clusters formed
also agree with molecular analyses performed on the
genus Haemulon (Rocha et al. 2008). Graphic repre-
sentation of the data on CV 2 axis, distinguishes
between species with similar feeding habits. Thus, the
group composed of C. nobilis, P. corvinaeformis,
H. plumierii and H. steindachneri, with an imbricated
pattern in relation to CV 2 axis, encompasses species
that feed strictly on macroinvertebrates. The other
more discriminated group consists only of H. auro-
lineatum, which displays more generalist feeding
habits. These are predominantly planktivore, but also
with a macroinvertebrate diet (Lindeman and Toxey
2002). In this case, in addition to reflecting phyloge-
netic patterns, the deformation grid also seems to
indicate functional ecomorphological adaptations,
related to variations primarily in the anterior region
of the body. Indeed, phylogenetic analyses based on
nuclear and mitochondrial genes, associated to the
ecological characteristics of Haemulon species, cor-
roborate historical changes, irrespective of feeding
habits, even among closely related species (Rocha
et al. 2008).
Differentiated evolutionary rates may be attributed
to cytogenetic (endophenotype) and morphological
(exophenotype) divergence in the Haemulidae family,
revealing techytelic evolution (fast-evolving) in terms
of ecomorphological traits. This condition had previ-
ously been determined for Serranidae species (Molina
et al. 2002), whose marked morphological differenti-
ations are present in cytogenetically conservative
species. These data are strongly compatible with the
occurence of ecological speciation in some Haemuli-
dae species (Rocha et al. 2008). In addition to
population isolation, due to strict biogeographic
barriers, evolution in this family may have occurred,
in part, through rapid changes in ecological adapta-
tion after niche fractioning, resulting in particular
morphoecological modifications, without significant
alterations in the karyotypic macrostructure of species.
The karyotypic patterns of Haemulidae presented
here demonstrate extensive chromosomal conserva-
tion, even after different cytogenetic investigation
methodologies, suggesting diversification of shapes
without concomitant fixation of conspicuous chromo-
somal rearrangements. This may also be extensive for
other families of Perciformes, which have also
revealed similar chromosomal patterns to those of
the family Haemulidae.
Acknowledgments The authors thank the Federal University
of Rio Grande do Norte, CNPq (Process No. 556793/2009-9),
and IBAMA (Process No. 19135/1) for supporting this study,
Jose Garcia for taxonomic identification of the specimens and
Pablo Martinez for assistance with geometric morphometrics
analysis.
References
Accioly IV, Molina WF (2007) Contribuicao a citogenetica dos
generos Pomadasys e Anisostremus (Haemulidae, Perci-
formes). Publica III:36–44
Aguirre WE, Shervette VR (2005) Morphological diversity of
the Cynoscion group (Perciformes: Sciaenidae) in the Gulf
of Guayaquil, Ecuador (Perciformes: Sciaenidae), a com-
parative approach. Environ Biol Fish 73:403–413
Araujo WC, Martinez PA, Molina WF (2010) Mapping of
ribosomal DNA by FISH, EcoRI digestion and replication
bands in the cardinalfish Apogon americanus (Percifor-
mes). Cytologia 75(1):109–117
Baras E (1999) Functional implications of early sexual growth
dimorphism in vundu. J Fish Biol 54:119–124
Barros e Silva E, Guerra M (2009) The meaning of DAPI bands
observed after C-banding and FISH procedures. Biotech
Histochem 4:1–11
Bertollo LAC, Fontes MS, Fenocchio AS, Cano J (1997) The
X1X2Y sex chromosome system in the fish Hoplias mala-baricusi. I. G-, C- and chromosome replication banding.
Chromosome Res 5:493–499
Blackith RE, Reyment RA (1971) Multivariate morphometrics.
Academic Press, London
Boron A (2003) Replication banding patterns in the spined
loach, Cobitis taenia L. (Pisces, Cobitidae). Genetica
119:51–55
Brum MJI, Galetti PM Jr (1997) Teleostei plan ground karyo-
type. J Comput Biol 2:91–102
Cequea H, Perez JE (1971) Variacion intra e interespecıfica de
hemoglobina y proteınas del plasma en algunas especies
del genero Haemulon. Bol Inst Oceanograf Univ Oriente
10(2):79–85
Cheverud JM (1988) A comparison of genetic and phenotypic
correlations. Evolution 42(5):958–968
Cioffi MB, Martins C, Centofante L, Jacobina U, Bertollo
LAC (2009) Chromosomal variability among allopatric
Rev Fish Biol Fisheries
123

populations of Erythrinidae fish Hoplias malabaricus:
mapping of three classes of repetitive DNAs. Cytogenet
Genome Res 125:132–141
Daniel-Silva MFZ, Almeida-Toledo LF (2001) Chromosome
R-banding pattern and conservation of a marker chromo-
some in four species, genus Astyanax (Characidae, Te-
tragonopterinae). Caryologia 54:209–215
Daniel-Silva MFZ, Almeida-Toledo LF (2005) Chromosome
evolution in fish: BrdU replication patterns demonstrate
chromosome homeologies in two species of the genus
Astyanax. Cytogenet Genome Res 109:497–501
Delany ME, Bloom SE (1984) Replication banding patterns in
the chromosomes of the rainbow trout. J Hered 75:431–434
Doebley J, Stec A (1993) Inheritance of morphological differ-
ences between maize and teosinte: comparison of results
for two F2 populations. Genetics 134:559–570
Ehlinger TJ (1991) Allometry and analysis of morphometric
variation in the bluegill, Lepomis macrochirus. Copeia
2:347–357
Fairbairn DJ (1997) Allometry for sexual size dimorphism:
pattern and process in the coevolution of body size in males
and females. Annu Rev Ecol Syst 28:659–687
Fulford RS, Rutherford DA (2000) Discrimination of larval
Morone geometric shape differences with landmark mor-
phometrics. Copeia 4:965–972
Galetti PM Jr, Aguilar CT, Molina WF (2000) An overview on
marine fish cytogenetics. Hydrobiologia 420:55–62
Giles V, Thode G, Alvarez MC (1988) Early replication bands in
two scorpion fishes, Scorpaena porcus and S. notata (Order
Scorpaeniformes). Cytogenet Cell Genet 47:80–83
Gold JR, Li C, Shipley NS, Powers PK (1990) Improved methods
for working with fish chromosomes with a review of
metaphase chromosome banding. J Fish Biol 37:563–575
Hellmer A, Voiculescu I, Schempp W (1991) Replication
banding studies in two cyprinid fishes. Chromosoma
100:524–553
Horn PJ, Peterson CL (2006) Heterochromatin assembly: a new
twist on an old model. Chromosome Res 14:83–94
Kasahara S (2009) Introducao a pesquisa em citogenetica de
vertebrados. Sociedade Brasileira de Genetica, Ribeirao
Preto
King M (1981) Chromosome change and speciation in lizards.
In: Atchley WR, Woodruff D (eds) Evolution and specia-
tion. Essays in honor of M. J. D. White. Cambridge Uni-
versity Press, London, pp 262–285
Klingenberg CP (2011) MORPHOJ: an integrated software
package for geometric morphometrics. Mol Ecol Res
11:353–357
Klinkhardt M, Tesche M, Greven H (1995) Database of
fish chromosomes. Westarp Wissenschaften (1st edn).
Magdeburg
Larson A (1998) The comparison of morphological and
molecular data in phylogenetic systematics. In: DeSalle R,
Schierwater B (eds) Molecular approaches to ecology and
evolution. Birkhauser Verlag, Basel, pp 275–296
Lindeman KC, Toxey C (2002) Haemulidae. In: Carpenter KE
(ed) FAO Species identification guide for fishery purposes.
The living marine resources of the western central Atlantic.
Food and Agriculture Organization of the United Nations,
Rome, pp 1522–1550
Maistro EL, Foresti F, Oliveira C (1999) R- and G-band patterns
in Astyanax scabripinnis paranae (Pisces, Characiformes,
Characidae). Genet Mol Biol 22:201–204
Molina WF (2001) An alternative method for mitotic stimula-
tion in fish cytogenetics. Chromosome Sci 5:149–152
Molina WF (2007) Chromosomal changes and stasis in marine
fish groups. In: Pisano E, Ozouf-Costaz C, Foresti F, Ka-
poor BG (eds) Fish cytogenetics. Science Publishers,
Enfield, pp 69–110
Molina WF, Galetti PM Jr (2002) Robertsonian rearrangements
in the reef fish Chromis (Perciformes, Pomacentridae)
involving chromosomes bearing 5 s rRNA genes. Genet
Mol Biol 25(4):373–377
Molina WF, Galetti PM Jr (2004) Karyotypic changes associ-
ated to the dispersive potential on Pomacentridae (Pisces,
Perciformes). J Exp Mar Biol Ecol 309:109–119
Molina WF, Galetti PM Jr (2007) Early replication banding in
Leporinus species (Osteichthyes, Characiformes) bearing
differentiated sex chromosomes (ZW). Genetica 130:
153–160
Molina WF, Maia-Lima FA, Affonso PRAM (2002) Divergence
between karyotypical pattern and speciation events in
Serranidae fish (Perciformes). Caryologia 55:299–305
Molina WF, Alves DEO, Araujo WC, Martinez PA, Silva MFM,
Costa GWWF (2010) Performance of human immuno-
stimulating agents in the improvement of fish cytogenetic
preparations. Genet Mol Res 9:1807–1814
Monteiro LR, Reis SF (1999) Princıpios de morfometria ge-
ometrica. Ed. Holos, Ribeirao Preto
Motta-Neto CC, Cioffi MB, Bertollo LAC, Molina WF (2011a)
Extensive chromosomal homologies and evidence of
karyotypic stasis in Atlantic grunts of the genus Haemulon(Perciformes). J Exp Mar Biol Ecol 401:75–79
Motta-Neto CC, Cioffi MB, Bertollo LAC, Molina WF (2011b)
Molecular cytogenetic analysis of Haemulidae fish (Perc-
iformes): evidence of evolutionary conservation. J Exp
Mar Biol Ecol 407:97–100
Nirchio M, Gaviria JI, Oliveira C, Ferreira IA, Martins C (2007)
Cytogenetic analysis of three species of the genus Haem-ulon (Teleostei: Haemulinae) from Margarita Island.
Venezuela. Genetica 131:135–140
Pendas AM, Moran P, Garcia-Vazquez E (1993) Replication
banding patterns in Atlantic salmon (Salmo salar). Gen-
ome 36:440–444
Rapp Py-Daniel LH, Cox Fernandes C (2005) Dimorfismo
sexual em Siluriformes e Gymnotiformes (Ostariophysi) da
Amazonia. Acta Amaz 35(1):97–110
Rocha LA, Lindeman KC, Rocha CR, Lessios HA (2008) His-
torical biogeography and speciation in the reef fish genus
Haemulon (Teleostei: Haemulidae). Mol Phylogenet Evol
48:918–928
Rohlf FJ (2006) TpsDig, version 2.10. Department of Ecology
and Evolution, State University of New York, Stony Brook
Rohlf FJ, Marcus LF (1993) A revolution in morphometrics.
Trends Ecol Evol 8(4):129–132
Rohlf FJ, Slice DE (1990) Extensions of the procrustes method
for the optimal superimposition of landmarks. Syst Zool
39:40–59
Salvadori S, Coluccia E, Cannas R, Cau A, Deiana AM (2003)
Replication banding in two Mediterranean moray eels:
Rev Fish Biol Fisheries
123

chromosomal characterization and comparison. Genetica
119:253–258
Schmid M, Guttenbach M (1988) Evolutionary diversity of
reverse (R) fluorescent chromosome bands in vertebrates.
Chromosoma 97:101–114
Sola L, Cataudella S, Capanna E (1981) New developments in
vertebrate cytotaxonomy III. Karyology of bony fishes: a
review. Genetica 54:285–328
Souza IL, Moreira-Filho O, Galetti PM Jr (1996) Heterochro-
matin differentiation in the characid fish Astyanax scabri-pinnis. Braz J Genet 19:405–410
Souza IL, Galian J, De La Rua P, Bertollo LAC, Moreira-Filho
O (2001) Non-random distribution of the GC-rich hetero-
chromatin and nucleolar rDNA sites on Astyanax scabri-pinnis chromosomes. Cytologia 66:637–655
Strauss RE, Fuiman LA (1985) Quantitative comparisons of
body form and allometry in larval and adult Pacific sculpins
(Teleostei: Cottidae). Can J Zool 63:1582–1589
Sumner AT (1972) A simple technique for demonstrating cen-
tromeric heterochromatin. Exp Cell Res 75:304
Sumner AT (2003) Chromosomes: organization and function.
Blackwell, North Berwick
Vicari MR, Artoni RF, Moreira-Filho O, Bertollo LAC (2008)
Colocalization of repetitive DNAs and silencing of major
rRNA genes. A case report of the fish Astyanax janeiro-
ensis. Cytogenet Genome Res 122:67–72
Vidales K, Markakis G, Tsimenides N (1997) Discrimination
between populations of picare Spicara smaris Linnaeus,
1758 in the Aegean Sea, using multivariate analysis of
phenetic characters. Fish Res 30:191–197
White MJD (1973) Animal cytology and evolution, 3rd edn.
Cambridge University Press, Cambridge
Zhang R, Wu H (1985) A study of sex chromosome in Carassiusauratus by BrdU Hoechst 33258 Giemsa techniques. Acta
Genet Sin 12:373–378
Rev Fish Biol Fisheries
123