the raf1 kinase associates with vimentin kinases and regulates the structure of vimentin filaments
TRANSCRIPT

The Raf-1 kinase associates with vimentin kinases andregulates the structure of vimentin filaments
PETRA JANOSCH,*,1 ARND KIESER,† MANFRED EULITZ,† JOSIP LOVRIC,† GUIDO SAUER,†
MANUELA REICHERT,‡ FOTINI GOUNARI,§ DIRK BUSCHER,¶ MANUELA BACCARINI,††
HARALD MISCHAK, ‡‡ AND WALTER KOLCH**The Beatson Institute for Cancer Research, Garscube Estate, Glasgow G61 1BD, U.K.; †GSF-Forschungszentrum fur Umwelt und Gesundheit, Institut fur Klinische Molekularbiologie undTumorgenetik, D-81377 Munchen, Germany; ‡Institute De Biochimie, Universite Lausanne, CH 1066Epalinges, Switzerland; §Dana-Farber Cancer Institute, Boston, Massachusetts 02115, USA; ¶The SalkInstitute, La Jolla, California 92037, USA; ††Institute of Microbiology and Genetics, Vienna Biocenter,A-1030 Vienna, Austria; and ‡‡Department of Nephrology, Medizinische Hochschule Hannover,30625 Hannover, Germany
ABSTRACT Using immobilized GST-Raf-1 as bait,we have isolated the intermediate filament proteinvimentin as a Raf-1-associated protein. Vimentincoimmunoprecipitated and colocalized with Raf-1 infibroblasts. Vimentin was not a Raf-1 substrate, butwas phosphorylated by Raf-1-associated vimentin ki-nases. We provide evidence for at least two Raf-1-associated vimentin kinases and identified one ascasein kinase 2. They are regulated by Raf-1, sincethe activation status of Raf-1 correlated with thephosphorylation of vimentin. Vimentin phosphoryla-tion by Raf-1 preparations interfered with its poly-merization in vitro. A subset of tryptic vimentinphosphopeptides induced by Raf-1 in vitro matchedthe vimentin phosphopeptides isolated from v-raf-transfected cells labeled with orthophosphoric acid,indicating that Raf-1 also induces vimentin phos-phorylation in intact cells. In NIH 3T3 fibroblasts,the selective activation of an estrogen-regulatedRaf-1 mutant induced a rearrangement and depoly-merization of the reticular vimentin scaffold similarto the changes elicited by serum treatment. Therearrangement of the vimentin network occurredindependently of the MEK/ERK pathway. Thesedata identify a new branch point in Raf-1 signaling,which links Raf-1 to changes in the cytoskeletalarchitecture.—Janosch, P., Kieser, A., Eulitz, M.,Lovric, J., Sauer, G., Reichert, M., Gounari, F.,Buscher, D., Baccarini, M., Mischak, H., Kolch, W.The Raf-1 kinase associates with vimentin kinasesand regulates the structure of vimentin filaments.FASEB J. 14, 2008–2021 (2000)
Key Words: Raf z phosphorylation z cytoskeleton z casein ki-nase 2
The physiological role of the Raf-1 kinase, thecellular homologue of the v-raf oncogene, has beena subject of intensive investigations in recent years. A
wide variety of growth factors and cytokines lead tothe activation of Raf-1 in many different cell types(reviewed in refs 1–3). The molecular mechanism ofRaf-1 activation is complex and still incompletelyunderstood, but in many situations involves the smallG-protein Ras. In its GTP-loaded state, Ras binds toRaf-1 with high affinity (reviewed in refs 4, 5) result-ing in the translocation of Raf-1 to the plasmamembrane (6, 7), where Raf-1 becomes activated bya still unknown mechanism, which involves phos-phorylation on tyrosine and/or serine residues (re-viewed in ref 8). Activation may be further enhancedby a lipid cofactor (9).
Activated Raf-1 phosphorylates and activates MEK(10–12), a dual specificity kinase that in turn phos-phorylates and activates MAPK/ERK, which propa-gates the signal to nuclear transcription factors, mostnotably the ternary complex factor, which is re-quired for the transcription of the c-fos gene (13,14). As constitutively activated MEK mutants cantransform NIH 3T3 cells and induce differentiationof PC12 cells, thereby mimicking the effects of v-raf,it has been suggested that activation of the MEK/ERK cascade is the main, if not sole, biologicalfunction of Raf-1 (15). Several lines of evidenceindicate, however, that Raf-1 may signal indepen-dently of the MEK/ERK pathway. First, v-raf cantransform Rat fibroblasts without activation of ERKs(16, 17). Second, Ras mutants have been isolatedthat still bind Raf-1 and consequently lead to ERKactivation, but fail to induce DNA synthesis or trans-formation (18, 19). Third, oncogenic Raf-1 caninduce the differentiation of 3T3L1 cells into adipo-cytes without activating ERKs (20, 21). Collectively,
1 Correspondence: The Beatson Institute for Cancer Re-search, Garscube Estate, Switchback Road, Bearsden, GlasgowG61 1BD, U.K. E-mail: [email protected]
2008 0892-6638/00/0014-2008/$02.25 © FASEB

these findings suggest the existence of other effec-tors and substrates of Raf-1.
Therefore, we set out to identify new Raf-1 signal-ing pathways and discovered that Raf-1 associateswith vimentin and vimentin kinases that regulate thearchitecture of vimentin filaments. Vimentin is themain constituent of intermediate filaments in mes-enchymal cells. Vimentin filaments are dynamicstructures that are constantly being rebuilt by anongoing exchange of vimentin protomers betweenthe soluble and polymeric phases (reviewed in refs22, 23). Although the exact function of the vimentinskeleton is still unknown, it is clear that its architec-ture is regulated in response to many extracellularstimuli and during the cell cycle. In part this regula-tion is exerted through phosphorylation, and vimen-tin has been described as a target for several kinases(reviewed in ref 24). Here we show that Raf-1 cansignal vimentin rearrangements via the regulation ofassociated vimentin kinases that are independent ofthe MEK/ERK pathway.
MATERIALS AND METHODS
Cell lines and reagents
HF-5, NIH 3T3, and COS-1 cells were maintained in Dulbec-co’s minimal essential medium (DMEM, Serva, Heidelberg,Germany) supplemented with glutamine and 10% fetal calfserum (FCS) from Seromed (Berlin, Germany) or Life Tech-nologies, Inc./BRL (Grand Island, N.Y.). The generation andmaintenance of BXB-ER cells were described previously (25).The MEK inhibitors PD098059 and U0126 were purchasedfrom New England Biolabs (Beverly, Mass.) and Promega(Madison, Wis.), respectively.
Isolation and identification of vimentin
3 3 109 logarithmically growing NIH 3T3 cells were lysed inTBS-1% Triton (TBST: 20 mM TrisHCl, pH 7.4, 150 mMNaCl, 2 mM EDTA, 1% Triton-X100, 1 mM PMSF, 10 mMleupeptin). The lysate was cleared by centrifugation at 21,000g for 10 min and incubated with glutathione Sepharose beads(Pharmacia, Piscataway, N.J.) for 1 h at 4°C. The beads wereloaded with ;5 mg of GST or GST-Raf-1 proteins that wereexpressed in the baculovirus/Sf-9 cell system and purified asdescribed (26). Beads were washed with TBST five timesbefore GST-Raf-1 and associated proteins were released withsodium dodecyl sulfate (SDS) gel sample buffer (3% SDS, 10mM DTT, 20 mM TrisHCl, pH 6.8), separated on a 10% SDSgel, and blotted onto nitrocellulose. The blot was stained withPonceau S. Proteins were cut out from the blot and digestedwith sequencing grade trypsin (Boehringer Mannheim,Mannheim, Germany) (27). Tryptic peptides were dissolvedin 60 ml of 0.1% trifluoroacetic acid and separated on anAquapore 300 Angstrom C8 column (1330 mm) using anApplied Biosystems Model 172 Microbore high-performanceliquid chromatography (HPLC). Peptide peaks were micro-sequenced in a Model 477A gas phase sequenator (AppliedBiosystems, Foster City, Calif.).
Vimentin phosphorylation and polymerization assays
Vimentin was expressed in Escherichia coli and purified asdescribed (28). Vimentin preparations in 8M urea weredialyzed against 10 mM TrisHCl, pH 7.4, 1 mM DTT, and 1mM EDTA. Insoluble material was removed by centrifugationat 21,000 g for 20 min. The supernatant containing solublevimentin protomers was incubated with purified GST-Raf-1proteins bound to glutathione Sepharose in a low-salt Raf-1kinase buffer (20 mM TrisHCl, pH 7.4, 20 mM NaCl, 10 mMMgCl2) supplemented with 2 mM ATP and 2.5 mCi of [32P]-g-ATP. In the course of experimentation, we discovered thatlow urea concentrations do not affect Raf-1 kinase activity andyielded identical results. Therefore, urea-containing vimentinpreparations were used in the kinase assays shown in Fig. 3.GST-Raf proteins were produced in Sf-9 cells, activated, andpurified as described (26). The protein kinase Ca (PKCa)-activated Raf preparations were devoid of detectable contam-ination by PKCa. Inclusion of a specific PKC inhibitor (1 mMGF203109X, Biomol, Plymouth Meeting, Pa.) did not alterRaf-1 mediated vimentin phosphorylation. Kinase reactionswere carried out as described previously (26). After thephosphorylation reaction, vimentin polymerization was in-duced by addition of NaCl to a final concentration of 150 mMand allowed to proceed for 30 min at room temperature.Vimentin polymers were pelleted by centrifugation at 21,000g for 20 min (28). The pellet was washed twice by vigorousresuspension in TBST before boiling in SDS gel samplebuffer. Samples were separated by SDS-gel electrophoresisand blotted onto nitrocellulose membranes. The blots wereautoradiographed and subsequently stained with vimentin orRaf antisera as indicated. Soluble activated Raf-1 was pro-duced as follows. GST-Raf-1 was activated by coexpression ofRas plus Lck in Sf-9 cells. After purification the Raf-1 portionwas released from the glutathione Sepharose beads by diges-tion with 0.5 mg thrombin. Thrombin was inactivated byaddition of 10 mg/ml leupeptin.
Phosphopeptide mapping
To assay vimentin phosphorylation in vivo, COS-1 cells weretransfected with a v-raf expression plasmid (29). For transfec-tion, 20–40% confluent COS-1 cells were incubated in 6-wellplates (Nunc, Roskilde, Denmark) containing 2 ml DMEMwith glutamine and 10% NU-serum (Collaborative Research,#55000) per well; 5 mg of 3611-v-raf plasmid DNA and 20 mlDC-mix (10.3 mg/ml chloroquin phosphate, Sigma, and 80mg/ml DEAE-dextrane, Sigma D-9885) were added to eachwell. After 4 h the medium was replaced by phosphate-buffered saline (PBS) containing 10% DMSO for 2 min. Thencells were fed with DMEM plus 10% FCS. Two days aftertransfection, cells were incubated in serum-free mediumovernight before harvest. Two days after transfection, COS-1cells were serum starved overnight, followed by additionalincubation in phosphate-free medium (Sigma, St. Louis, Mo.)for 3 h. Then cells were metabolically labeled by addition of0.5 mCi/ml [32P]-orthophosphoric acid for 3 h. Cells werelysed and vimentin was extracted as described (30). Inparallel, purified vimentin was phosphorylated by Raf-1 invitro. Samples were resolved by SDS-gel electrophoresis. Vi-mentin bands were cut out and digested with sequencinggrade trypsin (Promega) according to the instructions pro-vided by the manufacturer. Tryptic phosphopeptides wereprocessed for 2-dimensional peptide mapping as described(26) and spotted onto 20 3 20 cm phosphocellulose plates(Merck, Rahway, N.J.). In the first dimension, peptides wereresolved by electrophoresis in pH 8.9 buffer at 1250 volts for15 min; the second dimension was chromatography in n-butanol/pyridine/acetic acid/water (15/10/3/12) for 20 h.
2009RAF-1 REGULATES VIMENTIN KINASES

Immunoprecipitation and Western blotting
Approximately 1 3 107 growing NIH 3T3 cells were lysed inTBST buffer. To increase the yield of soluble vimentin, NaClwas omitted from the lysis buffer in some experiments.Lysates were cleared by centrifugation at 21,000 g for 10 minand immunoprecipitated with crafVI antiserum (1.5 ml per mlof lysate) or the corresponding preimmune serum. ThecrafVI serum was raised by immunizing rabbits with a peptidecorresponding to the unique carboxy-terminal 12 aminoacids of Raf-1 (26). Immunoprecipitates were washed fourtimes with TBST, then resolved by electrophoresis on 7.5%SDS gels and transferred to nitrocellulose membranes. Blotswere stained with anti-vimentin antibody (Vim3B4, Boehr-inger) using the enhanced chemiluminescence kit (Amer-sham, Arlington Heights, Ill.) as described previously (26).The antibodies were removed by incubating the membrane in3% SDS and 10 mM DTT for 15 min. Subsequently, Raf-1 wasdetected by staining with the PBB1 monoclonal Raf antibody(31). Soluble vimentin was precipitated with the V9 (DAKO,Carpinteria, Calif.) monoclonal antibody and Raf-1 was de-tected by staining with PBB1.
Cell fractionation
For the experiment shown in Fig. 8d, 1 3 106 COS-1 cells weretransfected with GST-BXB and MEK-DD (32) expressionplasmids using Superfect (Qiagen, Chatsworth, Calif.) accord-ing to the manufacturer’s instructions. Two days after trans-fection, the cells were treated with 50 mM U0126 (Promega)for 8 h and then lysed in 400 ml TBST. The lysates wereseparated into soluble and insoluble fractions by centrifuga-tion at 20,000 g for 30 min. The insoluble pellet was sonicatedand boiled in 400 ml SDS-gel sample buffer. Blots weresequentially stained with antibodies against vimentin (V9,DAKO), phospho-ERK (Sigma) and pan-ERK (Sigma). Forthe experiment shown in Fig. 8c, 1 3 107 BXB-ER cells werelabeled with 0.5 mCi/ml [32P]-orthophosphoric acid for 3 hbefore cells were treated with 1 mM estradiol (Sigma) asindicated. Lysates were prepared as above and vimentin wasimmunoprecipitated from the soluble fraction with the V9(DAKO) antibody.
In vitro vimentin binding experiments
For the experiments shown in Figs. 2d, e, ;1 3 108 logarith-mically growing NIH 3T3 cells were lysed in 12 ml of 20 mMTrisHCl, pH 7.5, 0.1 mM EDTA, 1% Triton X-100 plusphosphatase, and protease inhibitors. Lysates were cleared bycentrifugation at 20,000 g for 5 min and the supernatantswere incubated with the indicated GST-Raf proteins immobi-lized on glutathione agarose beads (Pharmacia) for 12 h at4°C. Beads were washed in lysis buffer four times, separatedon 12.5% SDS-gels, and sequentially immunoblotted withantibodies to vimentin (V9, DAKO; and goat anti-vimentinpolyclonal antibody, Sigma) and GST (Santa Cruz, SantaCruz, Calif.). GST, GRS, GNX, and GST-Raf proteins wereexpressed in Sf-9 insect cells and purified as described (26).For the experiments shown in Fig. 2e, the GST-tagged Raf-1kinase domain GNX was expressed in E. coli. The GNXmutants were generated by progressive deletions from thecarboxyl terminus and are described in (33). For the exper-iments shown in Fig. 2g, NIH cells were serum starvedovernight and stimulated with TPA (100 ng/ml) as indicated.Synthetic peptides were coupled to a chromatographic matrixusing the Ultralink kit (Pierce, Rockford, Ill.). The sequenceof the 259 peptide is QRQRSTS259TPNVHC, the sequence ofthe 621 peptide is KINRSAS621EPSLHRC. Corresponding
phosphopeptides were synthesized with phosphoserine atpositions 259 and 621, respectively. Incubation with celllysates was done as described above, but only for 2 h.Associated proteins were cut out from Coomassie-stained gels,digested with trypsin, and identified via mass determinationof tryptic peptides by mass spectrometry on a Bruker MALDI-TOF. MASCOT (www.matrixscience.com) was used for data-bases queries.
Immunofluorescence
For immunofluorescence experiments, human HF-5 ormouse NIH 3T3 fibroblasts were grown on chamber slides(Nunc), washed with PBS, and fixed in methanol for 10 minat 220°C. Samples were blocked with 10% serum in PBS for30 min and incubated with primary antibodies diluted in PBSplus 10% serum for 2 h at room temperature or for 16 h at4°C. FITC-coupled anti-mouse immunoglobulin G (IgG)(Jackson Laboratories, West Grove, Pa.) was used to detectanti-vimentin monoclonal antibodies and TRITC-coupledanti-rabbit IgG was used to visualize Raf-1. Between and afterincubations the slides were washed four times with PBS plus0.25% Triton X-100. The crafVI antiserum was used at a 1:100dilution or 1:1000 after affinity purification with the corre-sponding peptide, respectively. To ascertain antibody speci-ficity, the following control experiments were performed.When slides were incubated with mismatched combinationsof primary and secondary antibodies, i.e., crafVI (rabbit) andFITC-coupled anti-mouse IgG, or anti-vimentin (mouse) withTRITC-coupled anti-rabbit IgG, no staining was observed.Preincubation of the crafVI serum with the same volume ofthe peptide antigen (10 mg/ml) used for immunizationabolished the Raf stain. To assure the specificity of thevimentin stain, three different monoclonal vimentin antibod-ies were used including 7A3 (34), clone Vim3B4 (Boehr-inger) and clone 65E (Affinity, Neshanic Station, N.J.). Allantibodies used individually or as mixture gave indistinguish-able results. Photographs were taken at 10003 magnificationthrough a Zeiss Axiovert epifluorescence microscope. Toexclude ‘bleed through’ artifacts during photography, slidesstained with crafVI or vimentin antibodies alone were in-cluded as controls.
RESULTS
Isolation of vimentin as Raf-1-associated protein
Raf-1 associates with its substrate MEK quite stably(35, 36). Therefore, we tried to exploit this propertyto identify new Raf-1 substrates by isolating Raf-1-associated proteins. For this purpose, glutathione-S-transferase (GST) -tagged Raf-1 was expressed in thebaculovirus/Sf9 cell system and purified by adsorp-tion to glutathione-Sepharose beads. Subsequently,immobilized GST-Raf-1 and a control protein (GST)were incubated with NIH 3T3 lysates. After washeswith TBS-1% Triton, the GST-Raf-1 beads wereeluted with SDS, separated by SDS-polyacrylamidegel electrophoresis (PAGE), and blotted. Severalbands were found to specifically associate with Raf-1,but not with GST (Fig. 1a). The most abundant bandmigrated at 58 kDa. It was cut out from the blot,digested with trypsin, and the tryptic peptides were
2010 Vol. 14 October 2000 JANOSCH ET AL.The FASEB Journal

separated by reversed phase HPLC. Two of thecollected fractions that appeared to contain only asingle peptide were analyzed by microsequencing.The sequence of both tryptic peptides exhibited100% homology with mouse vimentin (Fig. 1b).
Interaction between Raf-1 and vimentin
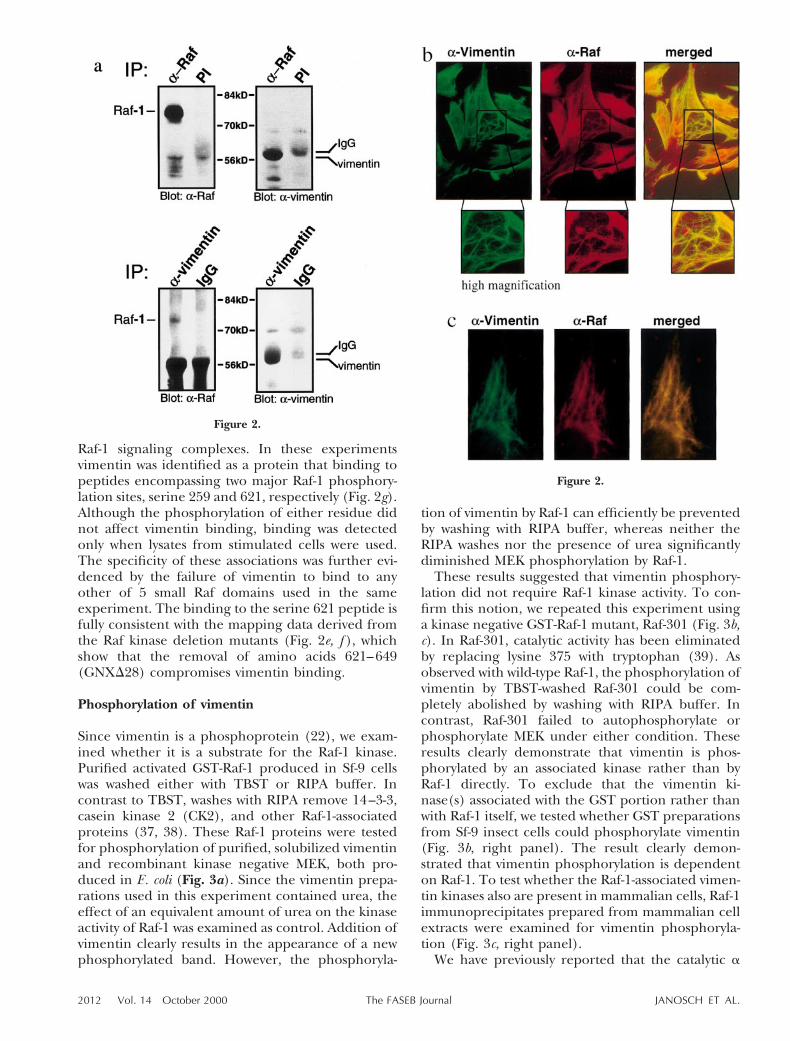
To test whether Raf-1 and vimentin are associated incells, NIH 3T3 lysates were immunoprecipitated withanti-Raf-1 (crafVI) or the corresponding preimmuneserum. The Immunoprecipitates were resolved by
SDS-PAGE, blotted, and stained with a monoclonalanti-vimentin antibody. Subsequently, the blot wasstripped and stained with Raf-1 specific antiserum(Fig. 2a). As evident from Fig. 2a, vimentin wasfound in the Raf-1 immunoprecipitate. Due to itssimilar size, vimentin comigrated with the immuno-globulin heavy chains, which somewhat distorted thevimentin band. Therefore, the specificity of thevimentin stain was confirmed by using several differ-ent anti-vimentin antibodies (data not shown). Inthe reciprocal experiment, Raf-1 was coimmunopre-cipitated by anti-vimentin antibodies but not controlimmunoglobulins.
The Raf-1 association with vimentin in intact cellswas further corroborated by immunofluorescenceexperiments in human and mouse fibroblasts, wherecells were simultaneously stained with anti-raf-1 andanti-vimentin antibodies (Fig. 2b, c). The specificityof the antibodies was assured as described in Mate-rials and Methods. Vimentin forms a filamentousreticular network throughout the cytoplasm. Raf-1was diffusely distributed in the cytosol, but alsodetected in fibrillar structures that showed extensiveoverlap with vimentin filaments. The spread-outmorphology and larger size of HF-5 facilitated visu-alization of the colocalization (Fig. 2b), but consis-tent findings were obtained in NIH 3T3 cells (Fig.2c) and after nocodazole treatment of NIH 3T3 andHF-5. Nocodazole disrupts the ordered architectureof intermediate filaments, resulting in a cotton ball-like appearance of vimentin filaments, which alsocolocalized with Raf-1 (data not shown). These dataindicate that Raf-1 can bind to vimentin in intactcells and colocalizes with the vimentin scaffold.
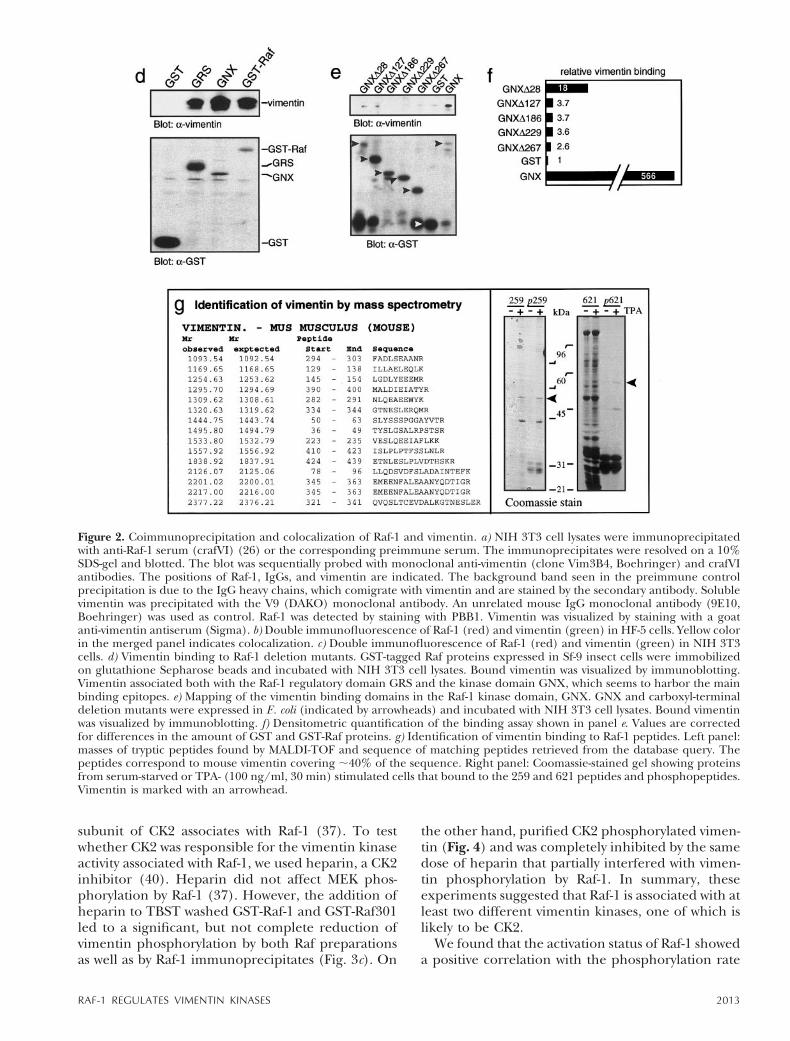
However, in vitro experiments using highly puri-fied vimentin and Raf-1 proteins produced in E. colior insect cells failed to reveal interactions (data notshown), suggesting that the interaction betweenRaf-1 and vimentin was indirect and mediated by ayet unknown component. Since vimentin binding toRaf-1 was readily observed in crude cell lysates, weused NIH 3T3 cell lysates to map the vimentinbinding domains in Raf-1. Although vimentin boundto both the regulatory and catalytic portions of Raf-1,the main binding domain appeared to be located inthe kinase domain (Fig. 2d). To refine the bindingsite in the Raf kinase domain, we used deletionmutants (Fig. 2e, f ). Vimentin binding was severelyreduced by the deletion of 28 carboxyl-terminalRaf-1 amino acids and virtually eliminated by furtherdeletion, indicating that the predominant vimentinbinding domain resides in the extreme carboxylterminus of Raf-1. The existence of at least twovimentin binding sites in Raf-1 was serendipitouslyand independently confirmed in the course of ex-periments where we used small peptide domains asaffinity baits to isolate the protein components of
Figure 1. Isolation of vimentin as Raf-1-associated protein. a)GST-tagged Raf-1 was immobilized on glutathione Sepharosebeads and incubated with NIH 3T3 cell lysates. Proteins wereeluted with SDS-gel sample buffer, separated on a 10% SDSgel, transferred to nitrocellulose, and stained with Ponceau S.Lane 1, protein marker; lane 2, proteins eluted from theGST-Raf-1 beads; lane 3, proteins eluted from the GSTcontrol beads. The positions of GST-Raf-1 and vimentin areindicated. b) The Raf-1-associated 58 kDa band was cut outfrom the blot and digested with trypsin. Tryptic peptides wereseparated by reversed phase HPLC and individual peaks weremicrosequenced. Complete sequence data could be obtainedfrom two tryptic peptides that showed 100% homology tomurine vimentin. They appear in boldface and underlined inthe mouse vimentin sequence.
2011RAF-1 REGULATES VIMENTIN KINASES
Raf-1 signaling complexes. In these experimentsvimentin was identified as a protein that binding topeptides encompassing two major Raf-1 phosphory-lation sites, serine 259 and 621, respectively (Fig. 2g).Although the phosphorylation of either residue didnot affect vimentin binding, binding was detectedonly when lysates from stimulated cells were used.The specificity of these associations was further evi-denced by the failure of vimentin to bind to anyother of 5 small Raf domains used in the sameexperiment. The binding to the serine 621 peptide isfully consistent with the mapping data derived fromthe Raf kinase deletion mutants (Fig. 2e, f ), whichshow that the removal of amino acids 621–649(GNXD28) compromises vimentin binding.
Phosphorylation of vimentin
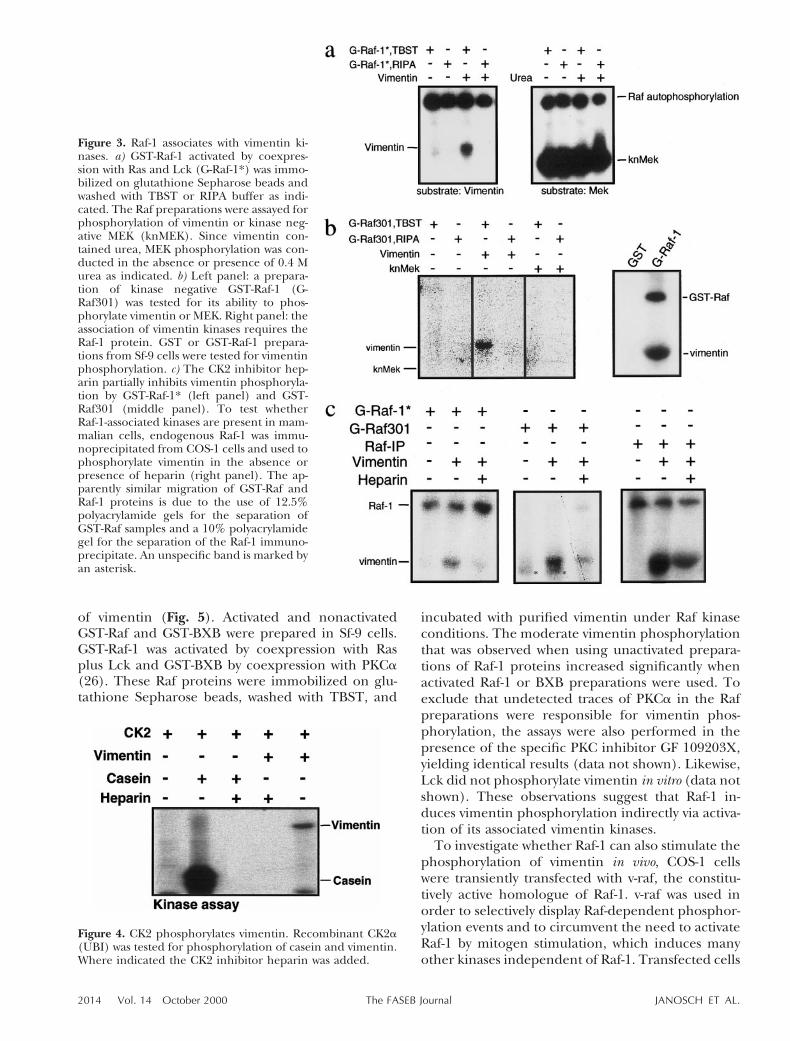
Since vimentin is a phosphoprotein (22), we exam-ined whether it is a substrate for the Raf-1 kinase.Purified activated GST-Raf-1 produced in Sf-9 cellswas washed either with TBST or RIPA buffer. Incontrast to TBST, washes with RIPA remove 14–3-3,casein kinase 2 (CK2), and other Raf-1-associatedproteins (37, 38). These Raf-1 proteins were testedfor phosphorylation of purified, solubilized vimentinand recombinant kinase negative MEK, both pro-duced in E. coli (Fig. 3a). Since the vimentin prepa-rations used in this experiment contained urea, theeffect of an equivalent amount of urea on the kinaseactivity of Raf-1 was examined as control. Addition ofvimentin clearly results in the appearance of a newphosphorylated band. However, the phosphoryla-
tion of vimentin by Raf-1 can efficiently be preventedby washing with RIPA buffer, whereas neither theRIPA washes nor the presence of urea significantlydiminished MEK phosphorylation by Raf-1.
These results suggested that vimentin phosphory-lation did not require Raf-1 kinase activity. To con-firm this notion, we repeated this experiment usinga kinase negative GST-Raf-1 mutant, Raf-301 (Fig. 3b,c). In Raf-301, catalytic activity has been eliminatedby replacing lysine 375 with tryptophan (39). Asobserved with wild-type Raf-1, the phosphorylation ofvimentin by TBST-washed Raf-301 could be com-pletely abolished by washing with RIPA buffer. Incontrast, Raf-301 failed to autophosphorylate orphosphorylate MEK under either condition. Theseresults clearly demonstrate that vimentin is phos-phorylated by an associated kinase rather than byRaf-1 directly. To exclude that the vimentin ki-nase(s) associated with the GST portion rather thanwith Raf-1 itself, we tested whether GST preparationsfrom Sf-9 insect cells could phosphorylate vimentin(Fig. 3b, right panel). The result clearly demon-strated that vimentin phosphorylation is dependenton Raf-1. To test whether the Raf-1-associated vimen-tin kinases also are present in mammalian cells, Raf-1immunoprecipitates prepared from mammalian cellextracts were examined for vimentin phosphoryla-tion (Fig. 3c, right panel).
We have previously reported that the catalytic a
Figure 2.
Figure 2.
2012 Vol. 14 October 2000 JANOSCH ET AL.The FASEB Journal
subunit of CK2 associates with Raf-1 (37). To testwhether CK2 was responsible for the vimentin kinaseactivity associated with Raf-1, we used heparin, a CK2inhibitor (40). Heparin did not affect MEK phos-phorylation by Raf-1 (37). However, the addition ofheparin to TBST washed GST-Raf-1 and GST-Raf301led to a significant, but not complete reduction ofvimentin phosphorylation by both Raf preparationsas well as by Raf-1 immunoprecipitates (Fig. 3c). On
the other hand, purified CK2 phosphorylated vimen-tin (Fig. 4) and was completely inhibited by the samedose of heparin that partially interfered with vimen-tin phosphorylation by Raf-1. In summary, theseexperiments suggested that Raf-1 is associated with atleast two different vimentin kinases, one of which islikely to be CK2.
We found that the activation status of Raf-1 showeda positive correlation with the phosphorylation rate
Figure 2. Coimmunoprecipitation and colocalization of Raf-1 and vimentin. a) NIH 3T3 cell lysates were immunoprecipitatedwith anti-Raf-1 serum (crafVI) (26) or the corresponding preimmune serum. The immunoprecipitates were resolved on a 10%SDS-gel and blotted. The blot was sequentially probed with monoclonal anti-vimentin (clone Vim3B4, Boehringer) and crafVIantibodies. The positions of Raf-1, IgGs, and vimentin are indicated. The background band seen in the preimmune controlprecipitation is due to the IgG heavy chains, which comigrate with vimentin and are stained by the secondary antibody. Solublevimentin was precipitated with the V9 (DAKO) monoclonal antibody. An unrelated mouse IgG monoclonal antibody (9E10,Boehringer) was used as control. Raf-1 was detected by staining with PBB1. Vimentin was visualized by staining with a goatanti-vimentin antiserum (Sigma). b) Double immunofluorescence of Raf-1 (red) and vimentin (green) in HF-5 cells. Yellow colorin the merged panel indicates colocalization. c) Double immunofluorescence of Raf-1 (red) and vimentin (green) in NIH 3T3cells. d) Vimentin binding to Raf-1 deletion mutants. GST-tagged Raf proteins expressed in Sf-9 insect cells were immobilizedon glutathione Sepharose beads and incubated with NIH 3T3 cell lysates. Bound vimentin was visualized by immunoblotting.Vimentin associated both with the Raf-1 regulatory domain GRS and the kinase domain GNX, which seems to harbor the mainbinding epitopes. e) Mapping of the vimentin binding domains in the Raf-1 kinase domain, GNX. GNX and carboxyl-terminaldeletion mutants were expressed in E. coli (indicated by arrowheads) and incubated with NIH 3T3 cell lysates. Bound vimentinwas visualized by immunoblotting. f) Densitometric quantification of the binding assay shown in panel e. Values are correctedfor differences in the amount of GST and GST-Raf proteins. g) Identification of vimentin binding to Raf-1 peptides. Left panel:masses of tryptic peptides found by MALDI-TOF and sequence of matching peptides retrieved from the database query. Thepeptides correspond to mouse vimentin covering ;40% of the sequence. Right panel: Coomassie-stained gel showing proteinsfrom serum-starved or TPA- (100 ng/ml, 30 min) stimulated cells that bound to the 259 and 621 peptides and phosphopeptides.Vimentin is marked with an arrowhead.
2013RAF-1 REGULATES VIMENTIN KINASES
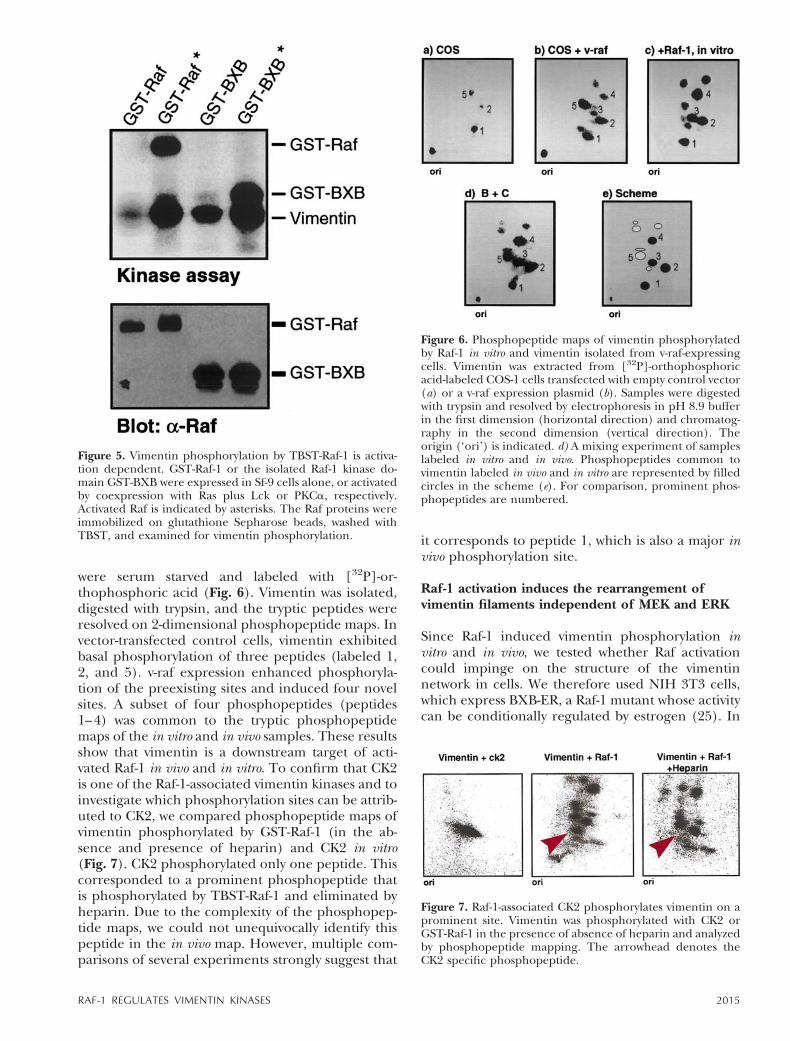
of vimentin (Fig. 5). Activated and nonactivatedGST-Raf and GST-BXB were prepared in Sf-9 cells.GST-Raf-1 was activated by coexpression with Rasplus Lck and GST-BXB by coexpression with PKCa(26). These Raf proteins were immobilized on glu-tathione Sepharose beads, washed with TBST, and
incubated with purified vimentin under Raf kinaseconditions. The moderate vimentin phosphorylationthat was observed when using unactivated prepara-tions of Raf-1 proteins increased significantly whenactivated Raf-1 or BXB preparations were used. Toexclude that undetected traces of PKCa in the Rafpreparations were responsible for vimentin phos-phorylation, the assays were also performed in thepresence of the specific PKC inhibitor GF 109203X,yielding identical results (data not shown). Likewise,Lck did not phosphorylate vimentin in vitro (data notshown). These observations suggest that Raf-1 in-duces vimentin phosphorylation indirectly via activa-tion of its associated vimentin kinases.
To investigate whether Raf-1 can also stimulate thephosphorylation of vimentin in vivo, COS-1 cellswere transiently transfected with v-raf, the constitu-tively active homologue of Raf-1. v-raf was used inorder to selectively display Raf-dependent phosphor-ylation events and to circumvent the need to activateRaf-1 by mitogen stimulation, which induces manyother kinases independent of Raf-1. Transfected cells
Figure 3. Raf-1 associates with vimentin ki-nases. a) GST-Raf-1 activated by coexpres-sion with Ras and Lck (G-Raf-1*) was immo-bilized on glutathione Sepharose beads andwashed with TBST or RIPA buffer as indi-cated. The Raf preparations were assayed forphosphorylation of vimentin or kinase neg-ative MEK (knMEK). Since vimentin con-tained urea, MEK phosphorylation was con-ducted in the absence or presence of 0.4 Murea as indicated. b) Left panel: a prepara-tion of kinase negative GST-Raf-1 (G-Raf301) was tested for its ability to phos-phorylate vimentin or MEK. Right panel: theassociation of vimentin kinases requires theRaf-1 protein. GST or GST-Raf-1 prepara-tions from Sf-9 cells were tested for vimentinphosphorylation. c) The CK2 inhibitor hep-arin partially inhibits vimentin phosphoryla-tion by GST-Raf-1* (left panel) and GST-Raf301 (middle panel). To test whetherRaf-1-associated kinases are present in mam-malian cells, endogenous Raf-1 was immu-noprecipitated from COS-1 cells and used tophosphorylate vimentin in the absence orpresence of heparin (right panel). The ap-parently similar migration of GST-Raf andRaf-1 proteins is due to the use of 12.5%polyacrylamide gels for the separation ofGST-Raf samples and a 10% polyacrylamidegel for the separation of the Raf-1 immuno-precipitate. An unspecific band is marked byan asterisk.
Figure 4. CK2 phosphorylates vimentin. Recombinant CK2a(UBI) was tested for phosphorylation of casein and vimentin.Where indicated the CK2 inhibitor heparin was added.
2014 Vol. 14 October 2000 JANOSCH ET AL.The FASEB Journal
were serum starved and labeled with [32P]-or-thophosphoric acid (Fig. 6). Vimentin was isolated,digested with trypsin, and the tryptic peptides wereresolved on 2-dimensional phosphopeptide maps. Invector-transfected control cells, vimentin exhibitedbasal phosphorylation of three peptides (labeled 1,2, and 5). v-raf expression enhanced phosphoryla-tion of the preexisting sites and induced four novelsites. A subset of four phosphopeptides (peptides1–4) was common to the tryptic phosphopeptidemaps of the in vitro and in vivo samples. These resultsshow that vimentin is a downstream target of acti-vated Raf-1 in vivo and in vitro. To confirm that CK2is one of the Raf-1-associated vimentin kinases and toinvestigate which phosphorylation sites can be attrib-uted to CK2, we compared phosphopeptide maps ofvimentin phosphorylated by GST-Raf-1 (in the ab-sence and presence of heparin) and CK2 in vitro(Fig. 7). CK2 phosphorylated only one peptide. Thiscorresponded to a prominent phosphopeptide thatis phosphorylated by TBST-Raf-1 and eliminated byheparin. Due to the complexity of the phosphopep-tide maps, we could not unequivocally identify thispeptide in the in vivo map. However, multiple com-parisons of several experiments strongly suggest that
it corresponds to peptide 1, which is also a major invivo phosphorylation site.
Raf-1 activation induces the rearrangement ofvimentin filaments independent of MEK and ERK
Since Raf-1 induced vimentin phosphorylation invitro and in vivo, we tested whether Raf activationcould impinge on the structure of the vimentinnetwork in cells. We therefore used NIH 3T3 cells,which express BXB-ER, a Raf-1 mutant whose activitycan be conditionally regulated by estrogen (25). In
Figure 5. Vimentin phosphorylation by TBST-Raf-1 is activa-tion dependent. GST-Raf-1 or the isolated Raf-1 kinase do-main GST-BXB were expressed in Sf-9 cells alone, or activatedby coexpression with Ras plus Lck or PKCa, respectively.Activated Raf is indicated by asterisks. The Raf proteins wereimmobilized on glutathione Sepharose beads, washed withTBST, and examined for vimentin phosphorylation.
Figure 6. Phosphopeptide maps of vimentin phosphorylatedby Raf-1 in vitro and vimentin isolated from v-raf-expressingcells. Vimentin was extracted from [32P]-orthophosphoricacid-labeled COS-1 cells transfected with empty control vector(a) or a v-raf expression plasmid (b). Samples were digestedwith trypsin and resolved by electrophoresis in pH 8.9 bufferin the first dimension (horizontal direction) and chromatog-raphy in the second dimension (vertical direction). Theorigin (‘ori’) is indicated. d) A mixing experiment of sampleslabeled in vitro and in vivo. Phosphopeptides common tovimentin labeled in vivo and in vitro are represented by filledcircles in the scheme (e). For comparison, prominent phos-phopeptides are numbered.
Figure 7. Raf-1-associated CK2 phosphorylates vimentin on aprominent site. Vimentin was phosphorylated with CK2 orGST-Raf-1 in the presence of absence of heparin and analyzedby phosphopeptide mapping. The arrowhead denotes theCK2 specific phosphopeptide.
2015RAF-1 REGULATES VIMENTIN KINASES
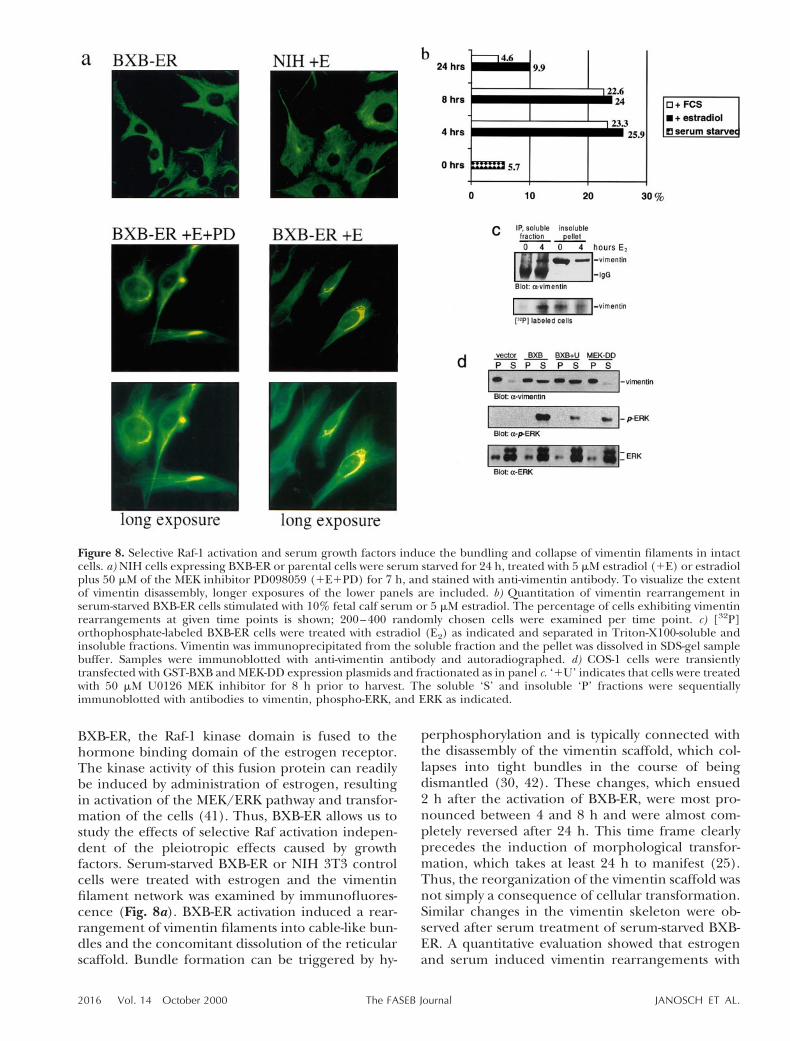
BXB-ER, the Raf-1 kinase domain is fused to thehormone binding domain of the estrogen receptor.The kinase activity of this fusion protein can readilybe induced by administration of estrogen, resultingin activation of the MEK/ERK pathway and transfor-mation of the cells (41). Thus, BXB-ER allows us tostudy the effects of selective Raf activation indepen-dent of the pleiotropic effects caused by growthfactors. Serum-starved BXB-ER or NIH 3T3 controlcells were treated with estrogen and the vimentinfilament network was examined by immunofluores-cence (Fig. 8a). BXB-ER activation induced a rear-rangement of vimentin filaments into cable-like bun-dles and the concomitant dissolution of the reticularscaffold. Bundle formation can be triggered by hy-
perphosphorylation and is typically connected withthe disassembly of the vimentin scaffold, which col-lapses into tight bundles in the course of beingdismantled (30, 42). These changes, which ensued2 h after the activation of BXB-ER, were most pro-nounced between 4 and 8 h and were almost com-pletely reversed after 24 h. This time frame clearlyprecedes the induction of morphological transfor-mation, which takes at least 24 h to manifest (25).Thus, the reorganization of the vimentin scaffold wasnot simply a consequence of cellular transformation.Similar changes in the vimentin skeleton were ob-served after serum treatment of serum-starved BXB-ER. A quantitative evaluation showed that estrogenand serum induced vimentin rearrangements with
Figure 8. Selective Raf-1 activation and serum growth factors induce the bundling and collapse of vimentin filaments in intactcells. a) NIH cells expressing BXB-ER or parental cells were serum starved for 24 h, treated with 5 mM estradiol (1E) or estradiolplus 50 mM of the MEK inhibitor PD098059 (1E1PD) for 7 h, and stained with anti-vimentin antibody. To visualize the extentof vimentin disassembly, longer exposures of the lower panels are included. b) Quantitation of vimentin rearrangement inserum-starved BXB-ER cells stimulated with 10% fetal calf serum or 5 mM estradiol. The percentage of cells exhibiting vimentinrearrangements at given time points is shown; 200–400 randomly chosen cells were examined per time point. c) [32P]orthophosphate-labeled BXB-ER cells were treated with estradiol (E2) as indicated and separated in Triton-X100-soluble andinsoluble fractions. Vimentin was immunoprecipitated from the soluble fraction and the pellet was dissolved in SDS-gel samplebuffer. Samples were immunoblotted with anti-vimentin antibody and autoradiographed. d) COS-1 cells were transientlytransfected with GST-BXB and MEK-DD expression plasmids and fractionated as in panel c. ‘1U’ indicates that cells were treatedwith 50 mM U0126 MEK inhibitor for 8 h prior to harvest. The soluble ‘S’ and insoluble ‘P’ fractions were sequentiallyimmunoblotted with antibodies to vimentin, phospho-ERK, and ERK as indicated.
2016 Vol. 14 October 2000 JANOSCH ET AL.The FASEB Journal

similar efficiencies and kinetics (Fig. 8b). The MEKinhibitor PD98059 did not interfere with BXB-ER-induced vimentin rearrangement (Fig. 8a), indicat-ing that it was not due to the MEK/ERK pathway butoccurred independently. In addition, neither MEKnor MAPK could phosphorylate vimentin in vitro,further confirming that the rearrangement of vimen-tin in the cell was not mediated by MEK or ERK (datanot shown).
To verify that the collapse of vimentin filaments inresponse to activation of BXB-ER was accompanied byphosphorylation and solubilization, we labeledBXB-ER cells with [32P]-orthophosphoric acid andseparated the lysates into detergent-soluble and insol-uble fractions (Fig. 8c). The amount of vimentin thatcould be immunoprecipitated from the soluble frac-tion increased dramatically upon BXB-ER activation,whereas insoluble vimentin filaments decreased. Solu-bilization was connected with phosphorylation as indi-cated by the preferential incorporation of 32P labelinto soluble vimentin. These results suggest thatBXB-ER causes vimentin rearrangements by inductionof vimentin phosphorylation, which leads to the solu-bilization of filaments. To further corroborate thishypothesis we transiently transfected COS-1 cells withBXB or an activated MEK-1 mutant, MEK-DD (32), andfractionated the cells as above (Fig. 8d). BXB led to apronounced redistribution of vimentin from the insol-uble pellet to the soluble phase, whereas MEK-DD wasineffective. The BXB-mediated vimentin solubilizationwas completely resistant to the MEK inhibitor U0126,although this inhibitor markedly suppressed ERK acti-vation by BXB. In summary, these experimentsdemonstrate a strong correlation in vivo between Rafactivity, vimentin phosphorylation, and vimentin depo-lymerization, which, however, is independent of MEKand ERK.
The above experiments pointed to a new effectorthat links Raf-1 to vimentin, but of course could notreveal how many steps downstream of Raf-1 thiseffector was. The simplest and immediately testablepossibility was that the effector(s) was one or more ofthe Raf-1-associated vimentin kinases. Therefore, weassayed whether phosphorylation by TBST-washedRaf-1 preparations could affect vimentin filaments invitro. Polymerized vimentin filaments were incubatedwith activated GST-Raf-1 or purified CK2 under Rafkinase conditions (Fig. 9a). At different time points,the samples were separated into soluble and insolu-ble fractions by centrifugation and the distributionof vimentin was examined by Western blotting.Quantitative evaluation of the Western blots by laserscanning densitometry showed that incubation withactivated Raf-1 resulted in the progressive redistribu-tion of vimentin into the soluble phase. In contrast,CK2 caused no significant redistribution of vimentin,
and thus is unlikely to mediate the effects of Raf-1 tothe vimentin scaffold.
Figure 9. Raf-1 induces vimentin phosphorylation and inhib-its polymerization. a) Polymerized vimentin filaments wereincubated with soluble activated Raf-1 (Raf-1*) or recombi-nant CK2a in Raf kinase buffer supplemented with 100 mMATP. In the ‘w/o’ lane kinases were omitted. At the timesindicated, aliquots were withdrawn and separated into solubleand insoluble fractions by centrifugation. The amount ofvimentin present in these fractions was detected by Westernblotting and quantitated by laser densitometry using a Cy-bertech Laserscan. Results are shown as the ratio betweensoluble and insoluble vimentin. b) Soluble vimentin wasphosphorylated with GST-Raf-1 (‘Raf’)or GST-Raf-1 that hadbeen activated by coexpression of PKCa in Sf-9 cells (Raf*).After the kinase reaction, the GST-Raf-1 beads were removedby centrifugation. The supernatant was further incubated 30min at room temperature in the presence of 150 mM NaCl toinduce vimentin polymerization. Vimentin polymers werepelleted by centrifugation (28). The supernatant and pelletfractions were boiled in SDS-sample buffer, separated bySDS-PAGE, and blotted. The blot was autoradiographed andthe distribution of vimentin was visualized by immunostainingwith anti-vimentin antibody.
2017RAF-1 REGULATES VIMENTIN KINASES

Vimentin filaments are dynamic structures that areconstantly rebuilt due to the exchange of solubletetramers with polymerized filaments (22, 24). Theslow kinetics of the reaction further suggested thatthe Raf-1 induced phosphorylation of vimentin didnot disrupt vimentin polymers directly, but ratherprohibited the reintegration of phosphorylated sol-uble vimentin into the filaments. To test this hypoth-esis, soluble vimentin oligomers were phosphory-lated with Raf-1 and activated Raf-1 (Fig. 9b).Immediately after the kinase reaction, vimentin po-lymerization was induced by raising the NaCl con-centration from 20 to 150 mM. Polymerized vimen-tin was separated from soluble vimentin bycentrifugation. The major fraction of phosphory-lated vimentin failed to polymerize and was recov-ered in the soluble fraction. In contrast, unactivatedRaf-1 induced little vimentin phosphorylation anddid not appreciably inhibit polymerization. These invitro results are consistent with the in vivo effects ofRaf on vimentin filaments, since increasing the rateof disassembly causes the loss of the reticular struc-ture of the vimentin network and its collapse intotight bundles.
Furthermore, these data suggested that the effectsof Raf-1 on vimentin are mediated by a Raf-1-acti-vated and -associated vimentin kinase other thanCK2. Several kinases have been described that regu-late vimentin by direct phosphorylation. These in-clude PKCa (43), cdc2 (44), and PKA (30, 43, 45).To examine whether any of these could be theassociated vimentin kinases of Raf-1, the pattern ofphosphorylation sites in vimentin induced by thesekinases were compared with the pattern induced byRaf-1 (data not shown). The phosphopeptide mapsshowed clearly that the phosphorylation sites invimentin induced by Raf-1 do not match with thephosphorylation sites in vimentin induced by PKCa,PKA, and cdc2. Thus, these kinases are unlikelycandidates for the Raf-1-associated vimentin ki-nase(s).
DISCUSSION
In this study we present evidence that the Raf-1kinase is physically linked to the vimentin scaffoldand can alter its structure by phosphorylation. In abiochemical screen for Raf-1-associated proteins, weisolated vimentin as a Raf-1-associated protein fromNIH 3T3 cell lysates. Immunofluorescence experi-ments revealed that a fraction of Raf-1 colocalizedwith vimentin filaments. This is consistent with theemerging hypothesis that Raf-1 participates in differ-ent signaling complexes with distinct subcellulardistribution. For instance, Raf-1 targeted to theplasma membrane efficiently activated the MEK/
ERK pathway, resulting in proliferation and sensiti-zation to myc-induced apoptosis (46), whereas Raf-1targeted to the mitochondrial membrane failed tostimulate the MEK/ERK cascade but prevented apo-ptosis by inducing the phosphorylation and inactiva-tion of BAD, a proapoptotic molecule (47). Thus, itappears that Raf-1 is distributed into different poolswithin the cell and that the coupling of Raf-1 tosubstrates may be regulated by subcellular colocal-ization.
We found that Raf-1 can induce vimentin phos-phorylation in vitro and in vivo. This activity is due toat least two different Raf-1-associated kinases ratherthan to Raf-1 catalytic activity. We could tentativelyidentify one of these kinases as CK2. The removal ofCK2 activity from Raf-1 kinase preparations by RIPAbuffer (37) or inhibition with heparin resulted in asignificant reduction in the phosphorylation of vi-mentin. As shown by phosphopeptide mapping, CK2phosphorylated vimentin only on one peptide invitro, which is also phosphorylated in vivo. Thus, wehave identified a novel vimentin kinase that on thebasis of its biochemical behavior most likely corre-sponds to CK2. Inhibition of CK2 diminished, but itdid not abolish vimentin phosphorylation by TBST-Raf-1 in vitro. As revealed by phosphopeptide map-ping, TBST-Raf-1 induced vimentin phosphorylationon multiple sites, some of which were also induced inv-raf-transfected cells. Therefore, we conclude thatRaf-1 associates with at least one other vimentinkinase besides CK2. We tried to identify the otherkinase(s) by comparing the phosphorylation sitepattern induced by TBST-Raf-1 with the sites phos-phorylated by known vimentin kinases. Several ki-nases have been reported to phosphorylate vimentinin vitro including cdc2 (44), mos (48), PKC (43),cAMP dependent kinase, PKA (30, 43, 45), cGMPdependent kinase (49), and calmodulin-regulatedkinase II (50). Since the roles of PKA, PKC, and cdc2in vimentin phosphorylation are much better char-acterized than those of other kinases, we focused onthese three. However, a comparison of the phos-phopeptide maps suggests that we can excludePKCa, PKA, and cdc2 as Raf-1-associated vimentinkinases, especially since we also were unable todetect coprecipitation or in vitro association (datanot shown).
Vimentin phosphorylation by TBST-Raf-1 was de-pendent on its activation, suggesting that Raf-1 canactivate one or more of its associated vimentinkinases. Indeed, we observed that Raf-1 was able toenhance the activity of recombinant CK2a in vitro.This activation increased with the amount of Raf-1added, but was independent of phosphorylation byRaf-1 (data not shown). An activation of CK2 by Raf-1is also suggested by the observation that v-Raf expres-sion enhanced the phosphorylation of vimentin on a
2018 Vol. 14 October 2000 JANOSCH ET AL.The FASEB Journal

peptide that is phosphorylated by CK2 in vitro. Sincev-raf expression also enhanced vimentin phosphory-lation on other sites, we believe that Raf-1 can alsoactivate vimentin kinases in cells.
These kinases seem to regulate the architecture ofvimentin filaments. The activation of a conditionalRaf-1 kinase, BXB-ER, in fibroblasts induces therearrangement of the vimentin scaffold. This was nota consequence of morphological transformation,which ensues hours later and is not caused bychanges in the actin cytoskeleton to which vimentinis linked, because BXB-ER does not induce actinrearrangement (25). In this study, a bundling ofmicrotubules was observed in response to BXB-ERactivation that slightly resembles the bundling ofvimentin filaments seen here. The effect on themicrotubules, however, was mediated via the MEK/ERK pathway and could be blocked by MEK inhibi-tion (25). In contrast, vimentin rearrangement inresponse to BXB-ER activation is not affected byMEK inhibition and therefore occurs independentlyof microtubule rearrangement. Our in vitro experi-ments indicate that this effect is due to Raf-1-associ-ated kinases that phosphorylate vimentin andthereby interfere with polymerization. Due to thecomplex phosphorylation pattern, we have not beenable to determine which phosphorylation sites areresponsible. CK2 phosphorylation does not seem toplay a role, since it fails to affect vimentin filamentsin vitro.
In the cell, vimentin filaments are not static struc-tures but are continuously rebuilt due to the incor-poration of soluble vimentin tetramers into thepolymers. This exchange seems to be important formaintaining the structure of the vimentin cytoskele-ton. The assembly and higher order arrangementare altered in response to cell cycle or differentiationspecific cues (22). These processes are regulated atleast partly by vimentin phosphorylation (22, 51, 52).Only the role of the cell cycle-regulated cdc-2 kinasein the disassembly of vimentin filaments duringmitosis is well established (44, 53–55). Mitosis isaccompanied by an increase in vimentin phosphor-ylation. Depending on the cell type, the vimentinscaffold undergoes a major rearrangement or com-plete disintegration during mitosis (44). These dif-ferences may be due to variations in the stoichiom-etry of vimentin phosphorylation by cdc-2 or couldreflect the cell type-specific participation of othervimentin kinases (44). Consistent with the latterhypothesis, PKC (56) and v-mos (57) have beenreported as mitotic vimentin kinases.
The physiological meaning of vimentin phosphor-ylation during interphase is less clear and may alsobe cell type dependent (22, 58). This uncertaintystems primarily from the incomplete knowledgeabout the precise functions of intermediate fila-
ments. Evidence suggests that intermediate filamentsdo not just serve “as mechanical integrators of cellu-lar space” (59), but may also be involved in regula-tory processes such as transformation (60), differen-tiation (61–64), cellular senescence (65), secretion(66), and even control of gene expression (67).Given the diverse functions of intermediate fila-ments, however, it appears plausible that their struc-tural organization is adapted to specific cellularrequirements by regulatory mechanisms, which con-ceivably could involve different kinases. Under invitro conditions, vimentin filaments can be disruptedby PKC or cAMP-dependent kinase (PKA) -mediatedphosphorylation (68, 69) of vimentin on sites otherthan cdc-2 (44). Microinjection of the catalytic sub-unit of PKA into rat embryo fibroblasts caused thedisassembly and collapse of vimentin filaments intotight bundles (30), and stimulation of Swiss 3T3 cellswith cAMP agonists resulted in a similar redistribu-tion of vimentin filaments (70). Unlike many othercell lines, including NIH 3T3, cAMP acts as a growth-promoting factor in Swiss 3T3 cells, suggesting thatvimentin rearrangement may be part of the mito-genic response. This idea is supported by our obser-vation that in NIH 3T3 cells stimulation with serumgrowth factors as well as the selective activation ofBXB-ER by estrogen both trigger vimentin bundlingwith similar kinetics.
Taken together, our data demonstrate that activa-tion of the Raf-1 kinase induces extensive vimentinreorganization similar to that seen during growthfactor stimulation. Whether the changes of the vi-mentin scaffold are required for the execution of themitogenic response has to await future studies. Theimportant finding of our study, however, is theidentification of a novel branch point in Raf-1 sig-naling that links Raf-1 with the cytoskeleton viaassociated vimentin kinases.
We thank Drs. P. Chambon for the estrogen receptorcDNA, G. Johnson for bacteria expressing MEK and ERK, M.Weber for the MEK-DD expression plasmid, and G. Multhofffor supplying the human fibroblast cells. We thank V. O’Brienfor critical reading of the manuscript and members of thelaboratory for helpful discussions. This work was supported bythe Cancer Research Campaign and DFG grant Ko-1492 toW.K.
REFERENCES
1. Heidecker, G., Kolch, W., Morrison, D. K., and Rapp, U. R.(1992) The role of Raf-1 phosphorylation in signal transduction.Adv. Cancer Res. 58, 53–73
2. Williams, N. G., and Roberts, T. M. (1994) Signal transductionpathways involving the Raf proto-oncogene. Cancer MetastasisRev. 13, 105–116
3. Daum, G., Eisenmann-Tappe, I., Fries, H.-W., Troppmair, J., andRapp, U. R. (1994) The ins and outs of Raf kinases. TrendsBiochem. Sci. 19, 474–480
4. Moodie, S. A., and Wolfman, A. (1994) The 3Rs of life: Ras. Rafand growth regulation. Trends. Genet. 10, 44–48
2019RAF-1 REGULATES VIMENTIN KINASES

5. Avruch, J., Zhang, X. F., and Kyriakis, J. M. (1994) Raf meetsRas: completing the framework of a signal transduction path-way. Trends. Biochem. Sci. 19, 279–283
6. Stokoe, D., Macdonald, S. G., Cadwallader, K., Symons, M., andHancock, J. F. (1994) Activation of Raf as a result of recruitmentto the plasma membrane. Science 264, 1463–1467
7. Leevers, S. J., Paterson, H. F., and Marshall, C. J. (1994)Requirement for Ras in Raf activation is overcome by targetingRaf to the plasma membrane. Nature (London) 369, 411–414
8. Morrison, D. K., and Cutler, R. E. (1997) The complexity ofRaf-1 regulation. Curr. Opin. Cell Biol. 9, 174–179
9. Dent, P., Reardon, D. B., Morrison, D. K., and Sturgill, T. W.(1995) Regulation of Raf-1 and Raf-1 mutants by Ras-dependentand Ras-independent mechanisms in vitro. Mol. Cell. Biol. 15,4125–4135
10. Dent, P., Haser, W., Haystead, T. A., Vincent, L. A., Roberts,T. M., and Sturgill, T. W. (1992) Activation of mitogen-activatedprotein kinase kinase by v-Raf in NIH 3T3 cells and in vitro [seecomments]. Science 257, 1404–1407
11. Kyriakis, J. M., App, H., Zhang, X. F., Banerjee, P., Brautigan,D. L., Rapp, U. R., and Avruch, J. (1992) Raf-1 activates MAPkinase-kinase. Nature (London) 358, 417–421
12. Howe, L. R., Leevers, S. J., Gomez, N., Nakielny, S., Cohen, P.,and Marshall, C. J. (1992) Activation of the MAP kinase pathwayby the protein kinase raf. Cell 71, 335–342
13. Kortenjann, M., Thomae, O., and Shaw, P. E. (1994) Inhibitionof v-raf-dependent c-fos expression and transformation by akinase-defective mutant of the mitogen-activated protein kinaseErk2. Mol. Cell. Biol. 14, 4815–4824
14. Gille, H. G., Sharrocks, A. D., and Shaw, P. E. (1992) Phosphor-ylation of transcription factor p62TCF by MAP kinase stimulatesternary complex formation at c-fos promoter. Nature (London)358, 414–417
15. Cowley, S., Paterson, H., Kemp, P., and Marshall, C. J. (1994)Activation of MAP kinase kinase is necessary and sufficient forPC12 differentiation and for transformation of NIH 3T3 cells.Cell 77, 841–852
16. Kizaka Kondoh, S., and Okayama, H. (1993) Raf-1 is not a majorupstream regulator of MAP kinases in rat fibroblasts. FEBS Lett.336, 255–258
17. Gallego, C., Gupta, S. K., Heasley, L. E., Qian, N. X., andJohnson, G. L. (1992) Mitogen-activated protein kinase activa-tion resulting from selective oncogene expression in NIH 3T3and rat 1a cells. Proc. Natl. Acad. Sci. USA 89, 7355–7359
18. White, M. A., Nicolette, C., Minden, A., Polverino, A., Van Aelst,L., Karin, M., and Wigler, M. H. (1995) Multiple Ras functionscan contribute to mammalian cell transformation. Cell 80,533–541
19. Joneson, T., White, M. A., Wigler, M. H., and Bar-Sagi, D. (1996)Stimulation of membrane ruffling and MAP kinase activation bydistinct effectors of RAS. Science 271, 810–812
20. Porras, A., Muszynski, K., Rapp, U. R., and Santos, E. (1994)Dissociation between activation of Raf-1 kinase and the 42-kDamitogen-activated protein kinase/90-kDa S6 kinase (MAPK/RSK) cascade in the insulin/Ras pathway of adipocytic differen-tiation of 3T3 L1 cells. J. Biol. Chem. 269, 12741–12748
21. Porras, A., and Santos, E. (1996) The insulin Ras pathway ofadipocytic differentiation of 3T3 L1 cells: dissociation betweenRaf-1 kinase and the MAPK/RSK cascade. Int. J. Obes. 20 (Suppl.3), S43–S51
22. Fuchs, E., and Weber, K. (1994) Intermediate filaments: struc-ture, dynamics, function, and disease. Annu. Rev. Biochem. 63,345–382
23. Evans, R. M. (1998) Vimentin: the conundrum of the interme-diate filament gene family. Bioessays 20, 79–86
24. Inagaki, M., Matsuoka, Y., Tsujimura, K., Ando, S., Tokui, T.,Takahashi, T., and Inagaki, N. (1996) Dynamic properties ofintermediate filaments: regulation by phosphorylation. Bioessays18, 481–487
25. Lovric, J., Dammeier, S., Kieser, A., Mischak, H., and Kolch, W.(1998) Activated raf induces the hyperphosphorylation of stath-min and the reorganization of the microtubule network. J. Biol.Chem. 273, 22848–22855
26. Hafner, S., Adler, H. S., Mischak, H., Janosch, P., Heidecker, G.,Wolfman, A., Pippig, S., Lohse, M., Ueffing, M., and Kolch, W.(1994) Mechanism of inhibition of Raf-1 by protein kinase A.Mol. Cell. Biol. 14, 6696–6703
27. Luo, K., Hurley, T. R., and Sefton, B. M. (1991) Cyanogenbromide cleavage and proteolytic peptide mapping of proteinsimmobilized to membranes. Methods Enzymol. 201, 149–152
28. Kouklis, P. D., Hatzfeld, M., Brunkener, M., Weber, K., andGeorgatos, S. D. (1993) In vitro assembly properties of vimentinmutagenized at the beta-site tail motif. J. Cell Sci. 106, 919–928
29. Rapp, U. R., Goldsborough, M. D., Mark, G. E., Bonner, T. I.,Groffen, J., Reynolds, F. H., Jr., and Stephenson, J. R. (1983)Structure and biological activity of v-raf, a unique oncogenetransduced by a retrovirus. Proc. Natl. Acad. Sci. USA 80, 4218–4222
30. Lamb, N. J., Fernandez, A., Feramisco, J. R., and Welch, W. J.(1989) Modulation of vimentin containing intermediate fila-ment distribution and phosphorylation in living fibroblasts bythe cAMP-dependent protein kinase. J. Cell Biol. 108, 2409–2422
31. Mischak, H., Kolch, W., Hofer, F., Weissinger, E., Gessl, A.,Davidson, W. F., Aiello, F. B., Blaas, D., and Rapp, U. R. (1990)A raf/myc virus immortalized macrophage cell line whichsupports the growth of B-cell and B-cell hybridomas. Oncogene 5,1377–1382
32. Catling, A. D., Schaeffer, H. J., Reuter, C. W., Reddy, G. R., andWeber, M. J. (1995) A proline-rich sequence unique to MEK1and MEK2 is required for raf binding and regulates MEKfunction. Mol. Cell. Biol. 15, 5214–5225
33. Yeung, K., Janosch, P., McFerran, B., Rose, D. W., Mischak, H.,Sedivy, J. M., and Kolch, W.(2000). The mechanism of suppres-sion of the Raf/MEK/ERK pathway by the RKIP inhibitorprotein. Mol. Cell. Biol. In press
34. Papamarcaki, T., Kouklis, P. D., Kreis, T. E., and Georgatos,S. D. (1991) The ‘lamin B-fold’. Anti-idiotypic antibodies reveala structural complementarity between nuclear lamin B andcytoplasmic intermediate filament epitopes. J. Biol. Chem. 266,21247–21251
35. Kolch, W., Philipp, A., Mischak, H., Dutil, E. M., Mullen, T. M.,Feramisco, J. R., Meinkoth, J. L., and Rose, D. W. (1996)Inhibition of Raf-1 signaling by a monoclonal antibody, whichinterferes with Raf-1 activation and with MEK substrate binding.Oncogene 13, 1305–1314
36. Huang, W., Alessandrini, A., Crews, C. M., and Erikson, R. L.(1993) Raf-1 forms a stable complex with Mek1 and activatesMek1 by serine phosphorylation. Proc. Natl. Acad. Sci. USA 90,10947–10951
37. Janosch, P., Schellerer, M., Seitz, T., Reim, P., Eulitz, M.,Brielmeier, M., Kolch, W., Sedivy, J. M., and Mischak, H. (1996)Characterization of IkappaB kinases—IkappaB-a is not phos-phorylated by Raf-1 or protein kinase C isozymes, but is a caseinkinase II substrate. J. Biol. Chem. 271, 13868–13874
38. Michaud, N. R., Fabian, J. R., Mathes, K. D., and Morrison, D. K.(1995) 14–3-3 is not essential for Raf-1 function: identificationof Raf-1 proteins that are biologically activated in a 14–3-3- andRas-independent manner. Mol. Cell. Biol. 15, 3390–3397
39. Kolch, W., Heidecker, G., Lloyd, P., and Rapp, U. R. (1991)Raf-1 protein kinase is required for growth of induced NIH/3T3cells. Nature (London) 349, 426–428
40. Graves, P. R., Haas, D. W., Hagedorn, C. H., DePaoli-Roach,A. A., and Roach, P. J. (1993) Molecular cloning, expression,and characterization of a 49-kilodalton casein kinase I isoformfrom rat testis. J. Biol. Chem. 268, 6394–6401
41. Samuels, M. L., Weber, M. J., Bishop, J. M., and McMahon, M.(1993) Conditional transformation of cells and rapid activationof the mitogen-activated protein kinase cascade by an estradiol-dependent human Raf-1 protein kinase. Mol. Cell. Biol. 13,6241–6252
42. Turowski, P., Myles, T., Hemmings, B. A., Fernandez, A., andLamb, N. J. (1999) Vimentin dephosphorylation by proteinphosphatase 2A is modulated by the targeting subunit B55. Mol.Biol. Cell 10, 1997–2015
43. Inagaki, M., Nishi, Y., Nishizawa, K., Matsuyama, M., and Sato, C.(1987) Site-specific phosphorylation induces disassembly ofvimentin filaments in vitro. Nature (London) 328, 649–652
44. Chou, Y. H., Bischoff, J. R., Beach, D., and Goldman, R. D.(1990) Intermediate filament reorganization during mitosis ismediated by p34cdc2 phosphorylation of vimentin. Cell 62,1063–1071
45. O’Connor, C. M., Gard, D. L., and Lazarides, E. (1981) Phos-phorylation of intermediate filament proteins by cAMP-depen-dent protein kinases. Cell 23, 135–143
2020 Vol. 14 October 2000 JANOSCH ET AL.The FASEB Journal

46. Kauffmann Zeh, A., Rodriguez Viciana, P., Ulrich, E., Gilbert,C., Coffer, P., Downward, J., and Evan, G. (1997) Suppression ofc-Myc-induced apoptosis by Ras signalling through PI(3)K andPKB. Nature (London) 385, 544–548
47. Wang, H. G., Rapp, U. R., and Reed, J. C. (1996) Bcl-2 targetsthe protein kinase Raf-1 to mitochondria. Cell 87, 629–638
48. Singh, B., and Arlinghaus, R. B. (1989) Vimentin phosphoryla-tion by p37mos protein kinase in vitro and generation of a50-kDa cleavage product in v-mos-transformed cells. Virology173, 144–156
49. Wyatt, T. A., Lincoln, T. M., and Pryzwansky, K. B. (1991)Vimentin is transiently co-localized with and phosphorylated bycyclic GMP-dependent protein kinase in formyl-peptide-stimu-lated neutrophils. J. Biol. Chem. 266, 21274–21280
50. Ando, S., Tokui, T., Yamauchi, T., Sugiura, H., Tanabe, K., andInagaki, M. (1991) Evidence that Ser-82 is a unique phosphor-ylation site on vimentin for Ca21-calmodulin-dependent pro-tein kinase II. Biochem. Biophys. Res. Commun. 175, 955–962
51. Eriksson, J. E., Opal, P., and Goldman, R. D. (1992) Intermedi-ate filament dynamics. Curr. Opin. Cell Biol. 4, 99–104
52. Heins, S., and Aebi, U. (1994) Making heads and tails ofintermediate filament assembly, dynamics and networks. Curr.Opin. Cell Biol. 6, 25–33
53. Chou, Y. H., Ngai, K. L., and Goldman, R. (1991) The regula-tion of intermediate filament reorganization in mitosis. p34cdc2phosphorylates vimentin at a unique N-terminal site. J. Biol.Chem. 266, 7325–7328
54. Chou, Y. H., Rosevear, E., and Goldman, R. D. (1989) Phos-phorylation and disassembly of intermediate filaments in mi-totic cells [published erratum appears in Proc. Natl. Acad. Sci.USA, 1989, vol. 86, p. 3214]. Proc. Natl. Acad. Sci. USA 86,1885–1889
55. Ando, S., Tsujimura, K., Matsuoka, Y., Tokui, T., Hisanaga, S.,Okumura, E., Uchiyama, M., Kishimoto, T., Yasuda, H., andInagaki, M. (1993) Phosphorylation of synthetic vimentin peptidesby cdc2 kinase. Biochem. Biophys. Res. Commun. 195, 837–843
56. Takai, Y., Ogawara, M., Tomono, Y., Moritoh, C., Imajoh-Ohmi,S., Tsutsumi, O., Taketani, Y., and Inagaki, M. (1996) Mitosis-specific phosphorylation of vimentin by protein kinase C cou-pled with reorganization of intracellular membranes. J. Cell Biol.133, 141–149
57. Bai, W., Arlinghaus, R. B., and Singh, B. (1993) Association ofv-Mos with soluble vimentin in vitro and in transformed cells.Oncogene 8, 2207–2212
58. Klymkowsky, M. W. (1995) Intermediate filaments: new proteins,some answers, more questions. Curr. Opin. Cell Biol. 7, 46–54
59. Lazarides, E. (1980) Intermediate filaments as mechanicalintegrators of cellular space. Nature (London) 283, 249–256
60. Eiden, M. V., MacArthur, L., and Okayama, H. (1991) Suppres-sion of the chemically transformed phenotype of BHK cells by ahuman cDNA. Mol. Cell. Biol. 11, 5321–5329
61. Ngai, J., Capetanaki, Y. G., and Lazarides, E. (1984) Differenti-ation of murine erythroleukemia cells results in the rapidrepression of vimentin gene expression. J. Cell Biol. 99, 306–314
62. Gard, D. L., and Lazarides, E. (1982) Cyclic AMP-modulatedphosphorylation of intermediate filament proteins in culturedavian myogenic cells. Mol. Cell. Biol. 2, 1104–1114
63. Shea, T. B., Beermann, M. L., and Fischer, I. (1993) Transientrequirement for vimentin in neuritogenesis: intracellular deliv-ery of anti-vimentin antibodies and antisense oligonucleotidesinhibit neurite initiation but not elongation of existing neuritesin neuroblastoma. J. Neurosci. Res. 36, 66–76
64. Capetanaki, Y., Smith, S., and Heath, J. P. (1989) Overexpres-sion of the vimentin gene in transgenic mice inhibits normallens cell differentiation. J. Cell Biol. 109, 1653–1664
65. Satoh, Y., Kashimura, M., Kaneko, S., Karasaki, Y., Higashi, K.,and Gotoh, S. (1994) Cloning of cDNAs with possible associa-tion with senescence and immortalization of human cells.Mutat. Res. 316, 25–36
66. Bertrand, F., Veissiere, D., Hermelin, B., Paul, A., Capeau, J.,Picard, J., and Cherqui, G. (1994) Phosphorylation of vimentinis an intermediate step in protein kinase C-mediated glycocon-jugate secretion. Am. J. Physiol. 266, C611–C621
67. Traub, P., and Shoeman, R. L. (1994) Intermediate filamentproteins: cytoskeletal elements with gene-regulatory function?Int. Rev. Cytol. 154, 1–103
68. Geisler, N., Hatzfeld, M., and Weber, K. (1989) Phosphorylationin vitro of vimentin by protein kinases A and C is restricted tothe head domain. Identification of the phosphoserine sites andtheir influence on filament formation. Eur. J. Biochem. 183,441–447
69. Ando, S., Tanabe, K., Gonda, Y., Sato, C., and Inagaki, M. (1989)Domain- and sequence-specific phosphorylation of vimentininduces disassembly of the filament structure. Biochemistry 28,2974–2979
70. Escribano, J., and Rozengurt, E. (1988) Cyclic AMP increasingagents rapidly stimulate vimentin phosphorylation in quiescentcultures of Swiss 3T3 cells. J. Cell. Physiol. 137, 223–234
Received for publication October 25, 1999.Revised for publication March 23, 2000.
2021RAF-1 REGULATES VIMENTIN KINASES