the chondrocyte cytoskeleton in mature articular cartilage: structure and distribution of actin,...
TRANSCRIPT

© The Histochemical Society, Inc.
0022-1554/00/$3.30
1307
ARTICLE
Volume 48(10): 1307–1320, 2000The Journal of Histochemistry & Cytochemistry
http://www.jhc.org
The Chondrocyte Cytoskeleton in Mature Articular Cartilage: Structure and Distribution of Actin, Tubulin, and Vimentin Filaments
Eve Langelier, Rosmarie Suetterlin, Caroline D. Hoemann, Ueli Aebi, and Michael D. Buschmann
Biomedical and Chemical Engineering, Ecole Polytechnique, Montreal, Quebec, Canada (EL,MDB); M.E. Müller Institutefor Structural Biology, Biozentrum, University of Basel, Basel, Switzerland (RS,UA); and BioSynTech, Ltd., Scientific and Hi-Technology Park, Laval, Quebec, Canada (CDH)
SUMMARY
We investigated the structure of the chondrocyte cytoskeleton in intact tissuesections of mature bovine articular cartilage using confocal fluorescence microscopy com-plemented by protein extraction and immunoblotting analysis. Actin microfilaments werepresent inside the cell membrane as a predominantly cortical structure. Vimentin and tubu-lin spanned the cytoplasm from cell to nuclear membrane, the vimentin network appearingfiner compared to tubulin. These cytoskeletal structures were present in chondrocytes fromall depth zones of the articular cartilage. However, staining intensity varied from zone tozone, usually showing more intense staining for the filament systems at the articular sur-face compared to the deeper zones. These results obtained on fluorescently labeled sec-tions were also corroborated by protein contents extracted and observed by immunoblot-ting. The observed cytoskeletal structures are compatible with some of the proposedcellular functions of these systems and support possible microenvironmental regulation ofthe cytoskeleton, including that due to physical forces from load-bearing, which are knownto vary through the depth layers of articular cartilage.
(J Histochem Cytochem 48:1307–1320, 2000)
T
he chondrocyte cytoskeleton
is a three-dimen-sional (3D) network composed of three types of pro-tein networks: actin microfilaments, microtubules,and intermediate filaments. Studies of the major com-ponents of the cytoskeleton in other cell types haveshown that actin microfilaments play roles in a tre-mendous number of cellular events, including migra-tion (Heath and Holifield 1991), adhesion (Turnerand Burridge 1991), shape alteration (Sims et al. 1992),cell signaling (Juliano and Haskill 1993), organellemovements (Simon and Pon 1996), endocytosis (Riez-man et al. 1996), secretion (Sontag et al. 1988; Koukou-ritaki et al. 1996), contractile ring formation (Otto andSchroeder 1990), myocyte contraction (Cooke 1986), ex-
tracellular matrix assembly (Hayes et al. 1999), andthe regulation of chondrocyte hypertrophy (Hirsch et al.1996,1997). Other studies have shown that constantlyrenewing microtubules contribute to intracytoplasmictransport (Vale 1987), mitotic spindle formation(Mitchison et al. 1986), and ciliary movement (Gib-bons 1981), and can be upregulated in chondrocytehypertrophy (Farquharson et al. 1999). Finally, inter-mediate filaments, in their polymeric coiled-coil config-uration, have been predominantly implicated in the me-chanical integration of cellular space, and in the spatialcoordination of mechanical events (Lazarides 1980) aswell as signal transduction (Traub 1995). Furthermore,all three cytoskeletal systems are believed to participatein intracellular mRNA transport and targeting (Glotzerand Ephrussi 1996).
Articular chondrocytes do not divide or migrate inhealthy adult cartilage, even in response to growth fac-tor stimulation. Therefore, the likely roles for micro-filaments in chondrocytes are in cell–matrix interactions,
Correspondence to: M. D. Buschmann, Biomedical Engineer-ing Inst., Ecole Polytechnique of Montreal, PO Box 6079, StationCentre-Ville, Montreal, Quebec, Canada H3C 3A7. E-mail: [email protected]
Received for publication December 6, 1999; accepted April 7,2000 (9A5156).
KEY WORDS
actin
tubulin
vimentin
cytoskeleton
confocal microscopy
chondrocyte
cartilage
by guest on July 3, 2015jhc.sagepub.comDownloaded from

1308
Langelier, Suetterlin, Hoemann, Aebi, Buschmann
cell signaling, differentiation, intracellular transport,control of secretion/endocytosis and in resiliency ofcell shape. Microfilaments (MFs) have been clearlyimplicated in control of chondrocyte phenotype(Brown and Benya 1988; Mallein–Gerin et al. 1991;Martin et al. 1999). In chondrocytes, a most likelyfunction for microtubules (MTs) is intracytoplasmictransport, whereas intermediate filaments (IFs) of chon-drocytes could be involved in mechanical as well astransport and signaling functions. Mechanical func-tions of IFs in chondrocytes are supported by previousobservations of increased IFs in weight-bearing re-gions of articular cartilage (Eggli et al. 1988) and incompression-bearing regions in fibrocartilage (Ralphset al. 1991,1992). A potential role of IFs in osteoar-thritis was also suggested by the loss of IF expressionin a mouse model of osteoarthritis before any other de-tectable extracellular changes (Benjamin et al. 1995).
Given the clear significance of cytoskeletal architec-ture in the physiology of chondrocytes and cartilage, itis important to develop techniques to assess the pres-ence and organization of cytoskeletal components inchondrocytes in situ. Fluorescence confocal micros-copy has been used after cryofixation to obtain confo-cal images of microfilaments and intermediate fila-ments, but not microtubules, in intact rat articularcartilage (Durrant et al. 1999). There have been noprevious studies reporting the maintenance and obser-vation of all three cytoskeletal systems in chondro-cytes in intact articular cartilage (for review see Ben-jamin et al. 1994). Therefore the purpose of this studywas to describe the distribution and physiological 3-Dorganization of chondrocyte cytoskeletal networks(MFs, MTs, and vimentin IFs) in situ, which is essentialfor a better understanding of the roles of the cytoskele-ton in cartilage physiology, mechanotransduction, anddisease, such as osteoarthritis. Distinct cell and extra-cellular matrix (ECM) differences are known to existamong different depth zones of articular cartilage(Muir et al. 1970; Speer and Dahners 1979; Bayliss etal. 1983; Schenk et al. 1986; Eggli et al. 1988; Wonget al. 1996), as are depth-dependent patterns of themechanical parameters of stress and strain (Kempsonet al. 1973; Woo et al. 1976; Roth and Mow 1980;Guilak 1995; Schinagl et al. 1996; Jurvelin et al. 1997;Kolmonen et al. 1997). Given these known cell, ECM,and mechanical zonal dependencies, we hypothesizedthat cytoskeletal structure and composition may alsodisplay zonal dependencies. We therefore requiredthat the cytoskeleton be assessed in each of the classi-cally defined noncalcified zones: superficial (or tan-gential), transitional, and radial (just before the calci-fied cartilage zone). Confocal immunofluorescenceand epifluorescence techniques were used to charac-terize the morphology and zonal dependence of thechondrocyte cytoskeletal components actin, tubulin,
and vimentin. The observed zonal distributions andmorphology of the three cytoskeletal filament systemslend support to some of their potential functionalroles in chondrocytes and provide a reference pointwith which alterations can be compared.
Materials and Methods
Tissue Explant Isolation and Culture
A recently described cartilage–bone tissue explant system(Dumont et al. 1999) was used in these studies. The uniquefeatures of this disk explant system compared to most previ-ously used cartilage explant systems (Grodzinsky 1990) arethe presence of an intact articular surface and a thin layer ofsubchondral bone, thus minimizing tissue cutting and swell-ing in culture. The lack of swelling allows easier mainte-nance of homeostasis by limiting diffusional loss of matrixmolecules. We showed stable cell viability, collagen and pro-teoglycan content, synthesis, and loss during 3-week cultureusing defined serum-free medium. We used this cultured ex-plant system in the current study to allow future studies in-volving application of mechanical loads in vitro and to pro-vide a postmortem stabilization period of several days inculture. Briefly, cartilage–bone cores were isolated from theload-bearing areas of the humeral heads of 1–2-year-oldsteers using an orthopedic coring bit (050.720; StraumannCanada, Toronto, ONT, Canada) under constant cooling ir-rigation with HBSS (14060; Gibco BRL, Rockville, MD).The location of isolated explants within the load-bearingcontact area was noted. In a grid of
z
30 disks taken fromone load-bearing region of the humeral head, in contactwith the scapular articular cartilage, there were 18 disks cir-cumscribing the periphery and 12 disks clustered more cen-trally within the load-bearing region. The explants were cutjust beneath the bone–cartilage interface with a specializeddevice to retain a thin layer of subchondral bone, thus pre-serving the natural cartilage–bone interface. The resultingdisks (4-mm diameter, 1.5-mm thick) contained the entirethickness of articular cartilage (
z
1.2-mm thick) attached toa layer of subchondral bone (
z
0.3-mm thick). Disk diame-ter was reduced to 3 mm using a biopsy punch (Miltex 15-33-33; Apocom, Montreal, PQ, Canada) cutting from thecartilage surface through the bone. After isolation, the diskswere washed five times with HBSS containing antibiotics,transferred to 96-well round-bottom microplates (5850-96;Corning Costar, Cambridge, MA), and cultured in 250
m
l/well of DME/F12 (pH 7.4) (D8900) supplemented with 50
m
g/ml gentamycin (G1272), 0.01% BSA (A8412), and 20
m
g/ml ascorbate (A4034; all from Sigma, St Louis, MO) at37C in 95% air, 5% CO
2
. Medium was changed every 24 hr,with ascorbate prepared fresh immediately before mediumchange.
Cell Viability
Cell viability was determined as described previously (Du-mont et al. 1999). Cartilage sections (75-
m
m thick) obtainedwith a Tissue-Chopper (McTwain; Brinkmann, Westbury,NY) were submitted to an assay solution consisting of PBS
by guest on July 3, 2015jhc.sagepub.comDownloaded from

Chondrocyte Cytoskeleton in Articular Cartilage
1309
with 1
m
M calcein AM, and 1.2
m
M ethidium homodimer-1(L-3224 LIVE/DEAD viability/Cytotoxicity Kit; MolecularProbes, Eugene, OR) to detect green fluorescing live and redfluorescing dead cells. Samples were incubated for 30 min orlonger at 37C, after which green (live) or red (dead) cellswere visualized with a Zeiss inverted fluorescence micro-scope and documented using a digital camera and NorthernEclipse software from Empix Imaging (Mississauga, ONT,Canada). Image manipulation and 3D reconstruction wereachieved using Scion Image from Scion (Frederick, MD).
Tissue Preparation for Fluorescence Microscopy
The cytoskeletal stabilization medium during processing was amodified Hank’s balanced salt solution (mHBSS): 136.9 mMNaCl, 5.36 mM KCl, 2 mM MgCl
2
, 0.336 mM Na
2
HPO
4
,0.44 mM KH
2
PO
4
, 4 mM NaHCO
3
, 2 mM EGTA, 5.55 mM
d
-glucose, 0.11% w/v 2-morphilino ethanesulfonic acid(MES), pH 6.5 [prepared from HBSS 10X (14185; GibcoBRL) by adding 2 mM MgCl
2
(M2393)
1
4 mM NaHCO
3
(S7277)
1
2 mM EGTA (E4378)
1
0.11 % w/v 2-morphil-ino ethanesulfonic acid (MES) (M5287); all from Sigma](Baschong et al. 1997). The low pH, chelation of calcium,and presence of magnesium has been found to aid in thepreservation of cytoskeletal structures during processing forseveral cell types (Bacallao et al. 1995). Samples were re-moved after 1–3 days of culture and placed on filter paper(bone layer down) on the stage of a Tissue Chopper with4% low melting temperature agarose (Seaplaque agarose;FMC, Rockland, ME) solubilized in mHBSS. (At contact,the agarose temperature is between 34 and 38C.) Thickslices (100
m
m) were taken, slicing from the articular surfacethrough the thin bone layer. The resulting slices were a rect-angular cross-section containing the entire articular cartilagelayer, the subtending calcified layer, and bone. Slices werethen permeabilized immediately after slicing in mHBSS con-taining 2% Octyl Poe (P-1140; Bachem Bioscience, King ofPrussia, PA) (Garavito and Rosenbusch 1986) and 0.125%gluteraldehyde (1152; Mecalab, Montreal, PQ, Canada) for20 min, washed in mHBSS for 5 min, fixed in 1% gluteral-dehyde in mHBSS for 30 min, all at 37C, and finally washedthree times for 5 min in mHBSS and stored in mHBSS at 4C(Baschong et al. 1997). An autofluorescence block (Weber etal. 1978) is performed on the same day or on the day afterthe fixation using 5 mg/ml NaBH
4
(S9125; Sigma) in mH-BSS on ice twice for 30 min, followed by washing four timesfor 5 min in mHBSS, and the slices are stored in mHBSS at4C. To allow antibody penetration, slices were digested for17 hr at 37C with 200 mU/ml chondroitinase ABC (100332,Seikagaku; PDI BioScience, Aurora, ONT, Canada) and 400mU/ml keratanase (100810; Seikagaku) in Tris-buffered sa-line (TBS; 100 mM NaCl, 100 mM Tris-HCl (7732; Mallin-ckrodt, Paris, KY), pH 7.4) with 0.01% BSA, followed bywashing in mHBSS. Actin MFs were directly labeled withTRITC–phalloidin (P1951, Sigma; diluted 1:1000 in mHBSS)in sealed containers for 4 hr. Vimentin IFs were also directlylabeled with a monoclonal antibody (MAb) anti-vim-Cy3(C9080, Sigma; diluted 1:700) in sealed containers for 4 hr.For
b
-tubulin, slices were incubated with a primary MAb (1111 876; Boehringer Mannheim, Laval, PQ, Canada; di-luted 1:50) for 4 hr, followed by secondary donkey anti-
mouse coupled to Cy2 (PA42003, Amersham, Poole, UK; di-luted 1:300) or secondary goat anti-mouse coupled to Alexa488 (A-11001, Molecular Probes; diluted 1:300). Slices werewashed four times over 5 min in mHBSS with agitation andwere immersed for 5 min in an anti-fading reagent, 0.18 mg/ml catalase (106 810; Boehringer Mannheim)
1
0.5 mg/mlglucose oxidase (646 431; Boehringer Mannheim)
1
0.1 mg/ml glucose (G7520; Sigma). Individual slices were mountedwith Mowiol (20 g Mowiol (475904; Calbiochem, San Di-ego, CA) and 0.3 g
N
-propyl-gallat (P3130; Sigma) in 40 mlglycerin (G6279; Sigma)) on slides with coverslips.
Confocal scanning microscopy was performed using argonlaser excitation on a Leica TCS 4-D CLSM (Leica; Heidel-berg, Germany) equipped with PL Apo 100
3
and Apochro-mat 63
3
1.4 NA oil immersion objectives. Optical sectionsat 0.5
m
m increments were obtained throughout the 100-
m
msection or were obtained for regions limited to particularcells and cell groups. Epifluorescence images were acquiredusing an inverted florescence Zeiss microscope equipped witha digital camera and image acquisition software (see above).To reduce background for epifluorescence, thinner 50-
m
msections from the tissue chopper were used. Cartilage–bonecore samples (before cutting off of the subchondral bone)were also sectioned using a vibratome (Vibratome 1000; Mi-cle Lab Engineering, Gomshall, Surrey, UK) because this sec-tioning procedure was seen to be gentler and to reduce celldeath due to sectioning. In that case, the long core samplewas held in place by the bone core with a custom mini-viseinstalled on the stage of the vibratome. Slices 25 and 50
m
mthick were taken, slicing from the articular surface through athin layer of bone. A razor blade was then used to detach theslices from the remaining bone core. During slicing with thevibratome, the sample was bathed in mHBSS at room tem-perature (RT).
Protein Extraction and Western Blotting Analysis
Western blotting analysis on individual disks was performedto complement observed depth-dependent fluorescence la-beling. Cultured disks were glued bone surface down withcyanoacrylate on the stage of the vibratome and bathed inmHBSS, pH 6.5, at RT. The disks were cut transversely into100-
m
m circular serial sections, the first section representingthe articular surface and the last the deep radial zone. Sec-tions were placed in mHBSS, pH 6.5, in individual wells of a96-well plate at RT. Each disk yielded 8–12 tangential sec-tions, which were subsequently divided into three equal pools(top
ù
tangential, middle
ù
transitional, bottom
ù
radial)representing the three zones of articular cartilage. Pooled sec-tions were weighed and immediately flash-frozen with liquidnitrogen. Each pool of sections was rendered into powder bypulverizing (BioPulverizer; Biospec Products, Rijswik, TheNetherlands) over liquid nitrogen, and the powder extractedwith 100
m
l RIPA buffer [0.1% SDS (4095-02; JT Baker,Phillipsburg, NJ), 1.0% desoxycholate (AC2849; Anachemia,Montreal, PQ, Canada), 1% Triton X-100 (161-0407; Bio-Rad, Hercules, CA), 10 mM Tris (161-0716; Bio-Rad), pH7.4, 150 mM NaCl] with protease inhibitors aprotinin(Sigma; A4529), PMSF (Sigma; P7626), iodoacetamide (Sigma;I6125), pepstatin A (Sigma; P5318), TPCK (Sigma; T4376), atconcentrations recommended by the manufacturer. Extracts
by guest on July 3, 2015jhc.sagepub.comDownloaded from

1310
Langelier, Suetterlin, Hoemann, Aebi, Buschmann
were cleared at 15,800
3
g (13 krpm) at 4C for 10 min. Ap-proximately equal volumes of RIPA-extracted supernatant(85
6
3
m
l) were obtained from each group of layers of alldisks analyzed. Proportionally equal volumes of supernatantwere loaded on three 10% SDS-PAGE gels and transferredto Immobilon-P membrane (IPVH00010; Millipore, Bed-ford, MA). To generate protein extracts of RIPA-insolubleproteins, the pellets from cleared RIPA extracts were ex-tracted in 100
m
l 4 M guanidine-HCl (4078-01; JT Baker)with protease inhibitors for 60 hr at 4C, followed by Centri-con concentration with RIPA as the exchange buffer, andvolumes were made to equal 100
m
l with RIPA before sub-jecting equal proportions of each sample to SDS-PAGE andimmunoblotting transfer. In parallel, a smaller volume ofeach extract was subjected to SDS-PAGE followed by Coo-massie Blue staining to control for total protein loaded fromeach extract. The resulting membranes were immunoblottedwith one of three antisera, including anti-vimentin mouseMAb V9 (V6630, Sigma; 1:2500), anti-
b
-tubulin MAb (1111 876, Boehringer; 1:350), or anti-actin MAb (A4700,Sigma; 1:500), followed by anti-mouse HRP-conjugated sec-ondary MAb (PI-2000; Vector, Burlingame, CA, 1:2500)and ECL chemiluminescence (RPN 2106; Amersham).
Time Lapse Imaging
A custom-made microscopy chamber was used to observechondrocyte morphology in permeabilization and fixationsolutions to ensure a minimum of structural alterations dur-ing these procedures. The microscopy chamber consisted ofa stainless steel plate with the dimensions of a slide. Theplate was pierced in the middle and two coverslips were usedas transparent bottom and top for the chamber that could beused on an inverted microscope. Thick slices (100
m
m) weretaken from fresh and cultured (up to 3 days) disks using atissue chopper as described above and were placed inmHBSS in the microscopy chamber. Differential interfer-ence contrast (DIC) images were taken of the fresh and cul-tured sections using an inverted fluorescence Zeiss micro-scope equipped with a
3
100 oil immersion objective. DICtime-lapse images of cultured sections were taken duringthe 20-min permeabilization period (2% Octyl Poe and0.125% glutaraldehyde in mHBSS) and the 30-min fixationperiod (1% gluteraldehyde in mHBSS) to observe potentialalterations in chondrocyte morphology due to those proce-dures.
Results
Reported results for cell viability, low-magnificationepifluorescence microscopy, confocal microscopy, andtime-lapse imaging represent common characteristicsseen in groups containing four to eight samples.
Cell Viability
Chondrocytes were viable in all zones at the time oftissue isolation and throughout the culture period of1–3 days, with the exception of a thin layer of cells at
the articular surface and the cut radial edge of thedisks (as seen previously in Figure 3 of Dumont et al.1999). The occasional dead cells in zones other thanthe most superficial layer were found to be those cutby the tissue chopper during sectioning.
Labeling Intensity and Western Blotting Analysis Showed a Heterogeneous Content of Cytoskeletal Components in the Different Depth Zonesof Articular Cartilage
Low-magnification epifluorescence images of the im-munostained cartilage sections displayed distinctivezonal dependencies of labeling intensity of the compo-nents of the chondrocyte cytoskeleton (Figure 1). Vi-mentin IF and MT signal intensity was higher in thesuperficial zones compared to deeper zones, whereassignal intensity for actin MFs was more uniform.Sharp vimentin and tubulin filament network gradientswere observed in those disks situated near the periph-ery of the load-bearing region, whereas more centrallylocated disks showed only slight depth-dependent ex-pression gradients. These zonal characteristics wereobserved both in sections taken from disks using thetissue chopper and in those prepared by the more gen-tle cutting of the vibratome on cartilage–bone corescontaining long bone stems.
Immunoblotting analysis showed that, in general,RIPA-solubilized cytoskeletal subunit levels were ele-vated in the most superficial layer relative to the mid-dle and deep zones (Figure 2). No major differenceswere observed between central and peripheral disksfor the RIPA extracts. However, GuCl extracts fromRIPA-insoluble pellets contained amounts of vimentinthat appeared to match the histochemical vimentingradients observed in adjacent disks. In the GuCl ex-tracts of centrally located disks, vimentin levels werehigher in the tangential and radial layers compared tothe transitional region, similar to the immunohis-tochemical signals (Figure 1). For peripherally locateddisks, there was a monotonic gradient in vimentin dis-tribution which was also similar to the immunohis-tochemical signals (Figure 1), with increasing levels ofvimentin content from the radial to the tangentialzone. Neither actin nor tubulin was detected in theGuCl extracts. The SDS-PAGE stained with CoomassieBlue showed protein loading to be globally equal forextracts from different layers (data not shown), whereseveral bands appeared uniformly present in differentdepths, whereas others manifested gradients of eitherdecreasing (as for cytoskeletal proteins) or increasingintensity with depth. An example of the latter was ob-served with blotting with an antibody specific for theC-propeptide of collagen Type II (data not shown),confirming the efficiency of protein extraction and re-covery from different layers and the presence of physi-
by guest on July 3, 2015jhc.sagepub.comDownloaded from

Chondrocyte Cytoskeleton in Articular Cartilage
1311
ological gradients in protein concentrations vs depthfrom the articular surface.
Specific Structural Morphologies for Actin, Tubulin, and Vimentin Filament Systems in ChondrocytesIn Situ Were Observed Using Confocal Microscopy
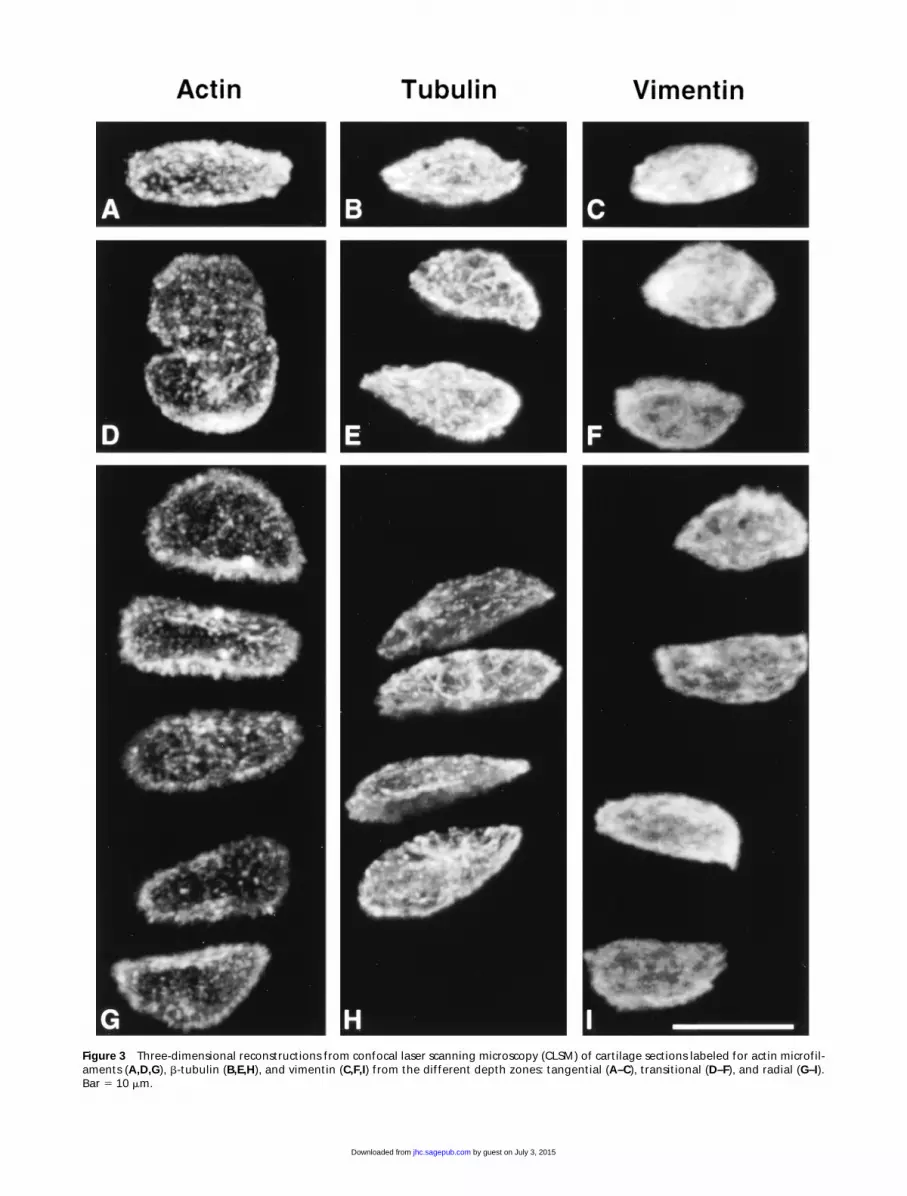
Cytoskeletal morphology within chondrocytes indi-cated specific structural characteristics for the three
filament systems in all depth zones (Figure 3). Actinlabeling was dense, punctate, and cortical, predomi-nantly located just inside the cell membrane (Figure4). MTs formed a loose basket-like mesh uniformlydistributed in the cytoplasm (Figure 5). Vimentin IFsdisplayed a tighter mesh that also traversed the cyto-plasm from the plasma to the nuclear membrane (Fig-ure 6). Regions of cells that were devoid of any label(including double-labeled specimens) were often rec-
Figure 1 Low-magnification epifluorescence images of cartilage sections labeled for actin microfilaments (A,D), b-tubulin (B,E), and vimen-tin (C,F). Sections were obtained from disks situated in the load-bearing contact region, some with more central (A–C) and some with moreperipheral (D–F) locations. The division of the cartilage into three zones, tangential at the articular surface followed by the transitional andfinally the deep radial zone, is shown by the dotted lines. Bar 5 250 mm.
by guest on July 3, 2015jhc.sagepub.comDownloaded from

1312
Langelier, Suetterlin, Hoemann, Aebi, Buschmann
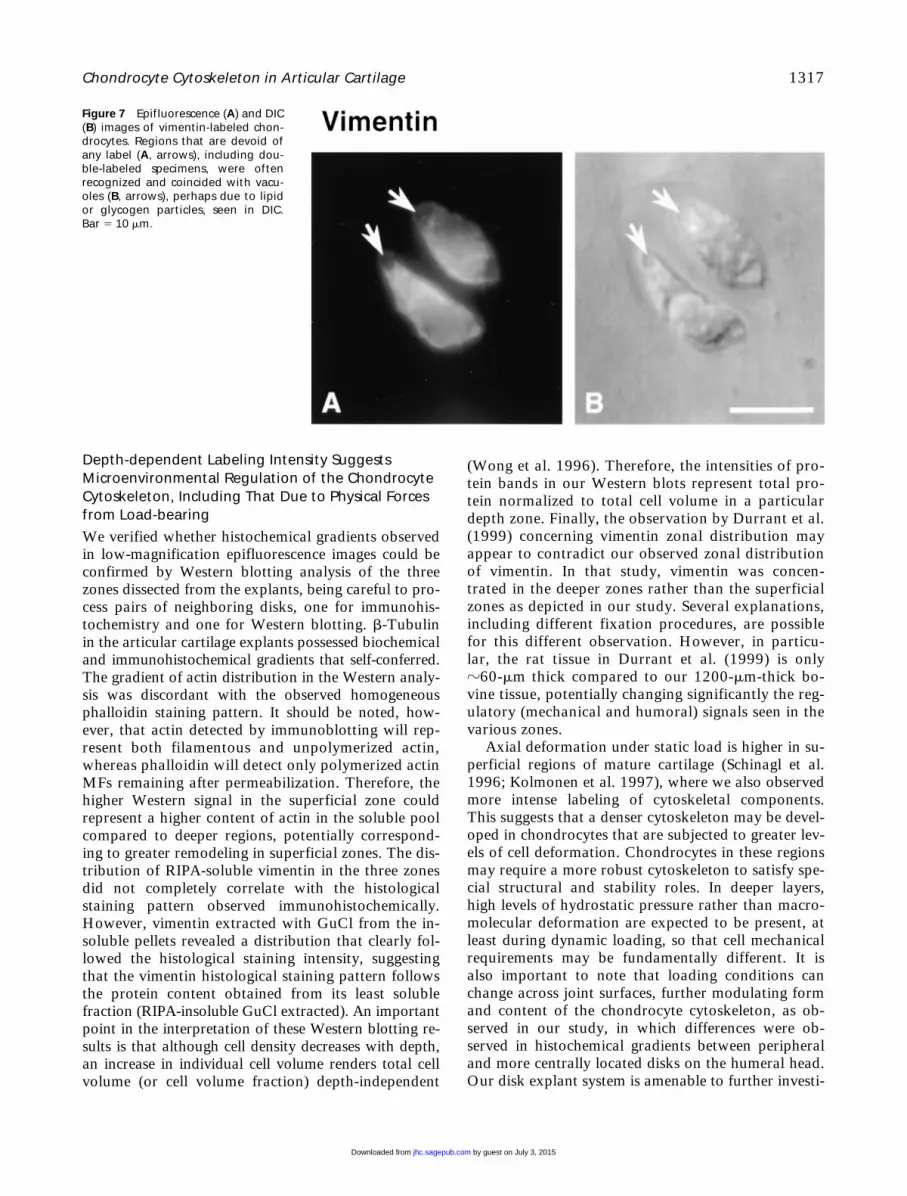
ognized (Figure 7). This absence of staining for cyto-skeletal components co-localized with vacuolar struc-tures seen using differential interference contrast (DIC)optics.
To verify that vacuolar structures were not artifac-tual, either induced by culture or by permeabilizationand fixation procedures, DIC images from sections offresh and cultured disks, as well as time-lapse imagingof sections undergoing permeabilization and fixation,were obtained. Vacuolar structures were visible onfreshly isolated and cultured disks (Figures 8A and 8B).Time-lapse imaging under DIC also revealed that cellmorphology, including vacuolar inclusion size andshape, was maintained throughout the permeabilizationand fixation procedures (Figures 8C–8F). These resultsconfirm the absence of marked morphological changescaused by culture, permeabilization, or fixation, sug-gesting that the observed cytoskeletal structures wererepresentative of in situ structures. The observed vacu-olar structures could be remnants of lipid droplets (Col-lins et al. 1965; Stockwell 1967; Eggli et al. 1988) orglycogen particles (Stockwell 1967; Eggli et al. 1988).
Discussion
Structural studies of the cytoskeleton of chondrocytesin intact tissue sections of mature articular cartilagewere undertaken to elucidate the in situ morphologyof these systems in the different depth zones of articu-lar cartilage. The dependence of morphology and la-beling intensity on the different zones, and the poten-tial roles and regulation of these filament systems inchondrocytes within mature articular cartilage, were
investigated. A uniform tissue preparation method wasfound to preserve in situ structures of all three fila-ment systems, based on a non-ionic detergent–glutaral-dehyde preparation of thick tissue sections, followedby autofluorescence blocking and glycosaminoglycandigestion to allow for antibody penetration. We ob-served that each filament system possessed a distinctstructure and that the intensity of labeling dependedon the depth zone of the labeled chondrocyte. Somefunctional roles and cytoskeletal regulatory factors arecompatible with these observations.
The Specific Structures Pertaining to Actin, Tubulin, and Vimentin Filament Systems Highlight Certain Functional Roles of These Systems in Chondrocytes of Articular Cartilage
The three cytoskeletal filament networks, MFs, MTs,and vimentin IFs, exhibited specific structural charac-teristics that can be related to their distinctive func-tions. Actin MF labeling was dense and punctate atthe peripheral cortex, similar to that reported by Dur-rant et al. (1999). The localization of actin just insidethe cell membrane has been observed in other celltypes and is consistent with roles of secretion and en-docytosis. The MF network could further offer struc-tural protection against shear stress at the cell cortexbecause, at low strains, MF networks offer greater re-sistance to shear stress compared to IF and MT net-works (Janmey et al. 1991). In articular cartilage, fluidflow occurs during dynamic compression, and thisflow could impose shear stress on the chondrocytecortex via the cell membrane (Kim et al. 1995). Thesefocal points of actin at cell membranes of chondro-cytes in situ were recently seen to co-localize with vincu-lin (Durrant et al. 1999) and could therefore representsites of adhesion to the ECM. This cortical distributionof actin and apparent lack of a direct link between theMFs and the nucleus argues against direct transmissionof mechanical deformation to the nucleus via microfila-ments (Guilak 1995). [It is also worth noting that cy-tochalasin D does not specifically alter cytoskeletal ac-tin independently of nuclear actin (Karim et al. 1992)or other cytoskeletal components.] Instead, the link-age between actin MFs and the nucleus could be indi-rect, involving other cytoskeletal proteins such as IFs.Finally, the importance of microfilament architecturein the control of chondrocyte phenotype cannot be un-derestimated (Brown and Benya 1988; Mallein–Gerinet al. 1991). Unlike chondrocytes grown in a mono-layer, chondrocytes in situ contained no stress fibers,further emphasizing the importance of developingthese in situ observation techniques.
MTs formed a loose, basket-like mesh spanningthroughout the cytoplasm. This structure is in agreementwith the intracytoplasmic transport functions associ-
Figure 2 Western blotting analysis of the three different zones(tangential, transitional, and radial) performed on disks situated inthe load-bearing contact region, with more central (left) or moreperipheral locations (right). Disks were chosen adjacent to the disksused for low-magnification epifluorescence images (Figure 1).Equal proportions of RIPA extracts and GuCl extracts of RIPA-insol-uble proteins were subjected to SDS-PAGE and immunoblottingtransfer. Membranes were immunoblotted for actin, b-tubulin, andvimentin.
by guest on July 3, 2015jhc.sagepub.comDownloaded from
Chondrocyte Cytoskeleton in Articular Cartilage
1313
Figure 3
Three-dimensional reconstructions from confocal laser scanning microscopy (CLSM) of cartilage sections labeled for actin microfil-aments (
A,D,G
),
b
-tubulin (
B,E,H
), and vimentin (
C,F,I
) from the different depth zones: tangential (
A–C
), transitional (
D–F
), and radial (
G–I
).Bar
5
10
m
m.
by guest on July 3, 2015jhc.sagepub.comDownloaded from

1314
Langelier, Suetterlin, Hoemann, Aebi, Buschmann
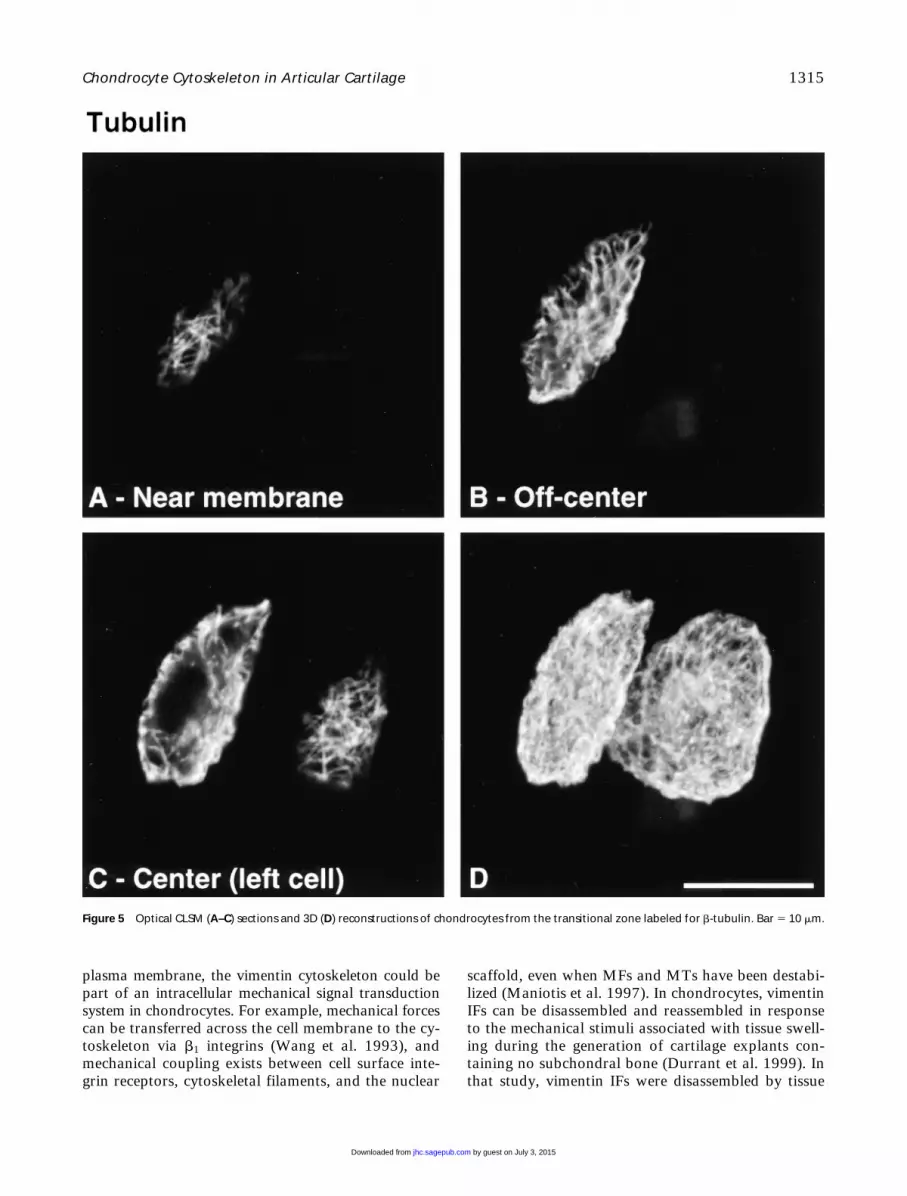
ated with MTs. In addition, the suggestion that MTscould serve as stabilizing elements for other cytoskele-tal systems (Janmey et al. 1991; Maniotis et al. 1997)is compatible with the MT organization observed inour study.
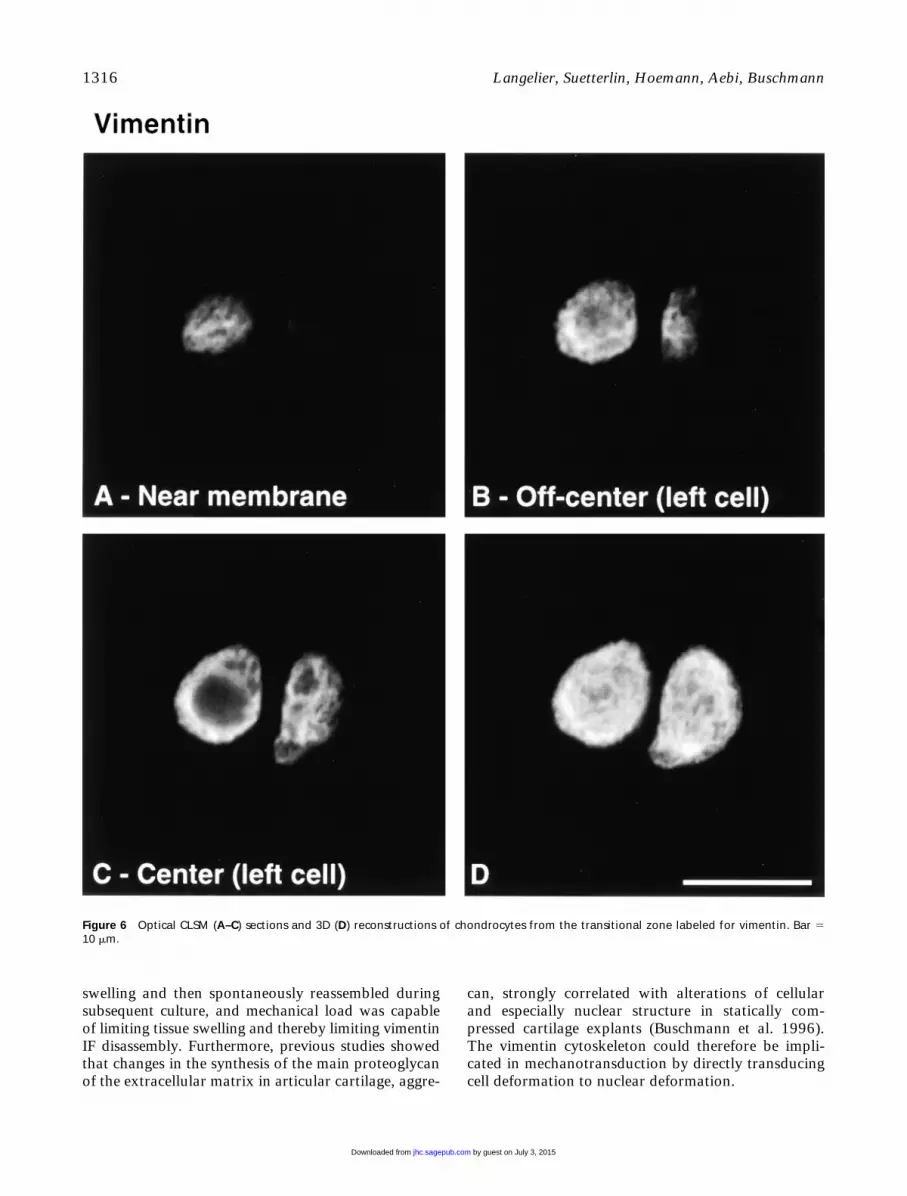
Vimentin IFs displayed a tighter, yet finer, meshthan MTs that also traversed the cytoplasm from the
plasma membrane to the nuclear membrane. Intracel-lular mRNA transport and targeting functions of theIF cytoskeleton are well served by this spanning orga-nization of vimentin, which is also consistent with thenotion that IFs may serve as mechanical integrators ofcellular space (Lazarides 1980). In agreement with itsnetwork organization spanning from the nuclear to the
Figure 4 Optical CLSM (A–C) sections and 3D (D) reconstructions of chondrocytes from the transitional zone labeled for actin microfila-ments. Bar 5 10 mm.
by guest on July 3, 2015jhc.sagepub.comDownloaded from
Chondrocyte Cytoskeleton in Articular Cartilage
1315
plasma membrane, the vimentin cytoskeleton could bepart of an intracellular mechanical signal transductionsystem in chondrocytes. For example, mechanical forcescan be transferred across the cell membrane to the cy-
toskeleton via
b
1
integrins (Wang et al. 1993), andmechanical coupling exists between cell surface inte-grin receptors, cytoskeletal filaments, and the nuclear
scaffold, even when MFs and MTs have been destabi-lized (Maniotis et al. 1997). In chondrocytes, vimentinIFs can be disassembled and reassembled in responseto the mechanical stimuli associated with tissue swell-ing during the generation of cartilage explants con-taining no subchondral bone (Durrant et al. 1999). Inthat study, vimentin IFs were disassembled by tissue
Figure 5 Optical CLSM (A–C) sections and 3D (D) reconstructions of chondrocytes from the transitional zone labeled for b-tubulin. Bar 5 10 mm.
by guest on July 3, 2015jhc.sagepub.comDownloaded from
1316
Langelier, Suetterlin, Hoemann, Aebi, Buschmann
swelling and then spontaneously reassembled duringsubsequent culture, and mechanical load was capableof limiting tissue swelling and thereby limiting vimentinIF disassembly. Furthermore, previous studies showedthat changes in the synthesis of the main proteoglycanof the extracellular matrix in articular cartilage, aggre-
can, strongly correlated with alterations of cellularand especially nuclear structure in statically com-pressed cartilage explants (Buschmann et al. 1996).The vimentin cytoskeleton could therefore be impli-cated in mechanotransduction by directly transducingcell deformation to nuclear deformation.
Figure 6 Optical CLSM (A–C) sections and 3D (D) reconstructions of chondrocytes from the transitional zone labeled for vimentin. Bar 510 mm.
by guest on July 3, 2015jhc.sagepub.comDownloaded from
Chondrocyte Cytoskeleton in Articular Cartilage
1317
Depth-dependent Labeling Intensity Suggests Microenvironmental Regulation of the Chondrocyte Cytoskeleton, Including That Due to Physical Forces from Load-bearing
We verified whether histochemical gradients observedin low-magnification epifluorescence images could beconfirmed by Western blotting analysis of the threezones dissected from the explants, being careful to pro-cess pairs of neighboring disks, one for immunohis-tochemistry and one for Western blotting.
b
-Tubulinin the articular cartilage explants possessed biochemicaland immunohistochemical gradients that self-conferred.The gradient of actin distribution in the Western analy-sis was discordant with the observed homogeneousphalloidin staining pattern. It should be noted, how-ever, that actin detected by immunoblotting will rep-resent both filamentous and unpolymerized actin,whereas phalloidin will detect only polymerized actinMFs remaining after permeabilization. Therefore, thehigher Western signal in the superficial zone couldrepresent a higher content of actin in the soluble poolcompared to deeper regions, potentially correspond-ing to greater remodeling in superficial zones. The dis-tribution of RIPA-soluble vimentin in the three zonesdid not completely correlate with the histologicalstaining pattern observed immunohistochemically.However, vimentin extracted with GuCl from the in-soluble pellets revealed a distribution that clearly fol-lowed the histological staining intensity, suggestingthat the vimentin histological staining pattern followsthe protein content obtained from its least solublefraction (RIPA-insoluble GuCl extracted). An importantpoint in the interpretation of these Western blotting re-sults is that although cell density decreases with depth,an increase in individual cell volume renders total cellvolume (or cell volume fraction) depth-independent
(Wong et al. 1996). Therefore, the intensities of pro-tein bands in our Western blots represent total pro-tein normalized to total cell volume in a particulardepth zone. Finally, the observation by Durrant et al.(1999) concerning vimentin zonal distribution mayappear to contradict our observed zonal distributionof vimentin. In that study, vimentin was concen-trated in the deeper zones rather than the superficialzones as depicted in our study. Several explanations,including different fixation procedures, are possiblefor this different observation. However, in particu-lar, the rat tissue in Durrant et al. (1999) is onlyz60-mm thick compared to our 1200-mm-thick bo-vine tissue, potentially changing significantly the reg-ulatory (mechanical and humoral) signals seen in thevarious zones.
Axial deformation under static load is higher in su-perficial regions of mature cartilage (Schinagl et al.1996; Kolmonen et al. 1997), where we also observedmore intense labeling of cytoskeletal components.This suggests that a denser cytoskeleton may be devel-oped in chondrocytes that are subjected to greater lev-els of cell deformation. Chondrocytes in these regionsmay require a more robust cytoskeleton to satisfy spe-cial structural and stability roles. In deeper layers,high levels of hydrostatic pressure rather than macro-molecular deformation are expected to be present, atleast during dynamic loading, so that cell mechanicalrequirements may be fundamentally different. It isalso important to note that loading conditions canchange across joint surfaces, further modulating formand content of the chondrocyte cytoskeleton, as ob-served in our study, in which differences were ob-served in histochemical gradients between peripheraland more centrally located disks on the humeral head.Our disk explant system is amenable to further investi-
Figure 7 Epifluorescence (A) and DIC(B) images of vimentin-labeled chon-drocytes. Regions that are devoid ofany label (A, arrows), including dou-ble-labeled specimens, were oftenrecognized and coincided with vacu-oles (B, arrows), perhaps due to lipidor glycogen particles, seen in DIC.Bar 5 10 mm.
by guest on July 3, 2015jhc.sagepub.comDownloaded from

1318 Langelier, Suetterlin, Hoemann, Aebi, Buschmann
gation of the roles and dynamics of the chondrocytecytoskeleton and, in particular, its alteration due toload. Such studies may have implications for the abil-ity of chondrocytes to structurally withstand certainlevels of mechanical load and subsequently to prop-erly maintain a functional tissue or lead to a cellular
malfunction and net degeneration of articular carti-lage. Furthermore, the techniques developed in ourstudy could be applied to observe the chondrocyte cy-toskeleton in cartilage tissue from animal models ofdisease or from humans, to aid understanding of carti-lage pathology.
Figure 8 DIC images of cartilage sec-tions. Vacuolar structures (arrows) arevisible on freshly isolated disks (A)and on cultured disks (B). In addition,as observed by time-lapse imaging,cell morphology, including vacuoleand nuclear size and shape, wasmaintained throughout the perme-abilization (C,D) and fixation (E,F)processing procedures. Bar 5 10 mm.
by guest on July 3, 2015jhc.sagepub.comDownloaded from

Chondrocyte Cytoskeleton in Articular Cartilage 1319
AcknowledgmentsSupported by the Arthritis Society (Canada), by Fonds
pour la Formation de Chercheurs et l’Aide à la Recherche(scholarship to EL), by the Swiss National Foundation forScientific Research, by the M.E. Müller Foundation of Swit-zerland, and by the Canton Basel Stadt.
Literature CitedBacallao R, Kiai K, Jesaitis L (1995) Guiding principles of speci-
men preservation for confocal fluorescence microscopy. InPawley JB, ed. Handbook of Biological Confocal Microscopy.New York, Plenum Press, 311–325
Baschong W, Sutterlin R, Aebi U (1997) Punch-wounded, fibro-blast populated collagen matrices—a novel approach forstudying cytoskeletal changes in three dimensions by confocallaser scanning microscopy. Eur J Cell Biol 72:189–201
Bayliss MT, Venn M, Maroudas A, Ali SY (1983) Structure of pro-teoglycans from different layers of human articular cartilage.Biochem J 209:387–400
Benjamin M, Archer CW, Ralphs JR (1994) Cytoskeleton of car-tilage cells. Microsc Res Tech 28:372–377
Benjamin M, Ralphs JR, Archer CW, Mason RM, Chambers M(1995) Cytoskeletal changes in articular fibrocartilage are anearly indicator of osteoarthritis in STR/ORT mice. Trans Or-thop Res Soc 41:246
Brown PD, Benya PD (1988) Alterations in chondrocyte cyto-skeletal architecture during phenotypic modulation by retinoicacid and dihydrocytochalasin B-induced reexpression. J CellBiol 106:171–179
Buschmann MD, Hunziker EB, Kim YJ, Grodzinsky AJ (1996) Al-tered aggrecan synthesis correlates with cell and nucleus structurein statically compressed cartilage. J Cell Sci 109(pt 2)499–508
Collins DH, Ghadially FN, Meachim G (1965) Intra-cellular lip-ids of cartilage. Ann Rheum Dis 24:123–135
Cooke R (1986) The mechanism of muscle contraction. CRCCrit Rev Biochem 21:53–118
Dumont J, Ionescu M, Reiner A, Poole AR, Tran–Khanh N,Hoemann CD, McKee M, Buschmann MD (1999) Maturefull-thickness articular cartilage explants attached to bone arephysiologically stable over long-term culture in serum-freemedia. Connect Tissue Res 40:259–272
Durrant LA, Archer CW, Benjamin M, Ralphs JR (1999) Organ-isation of the chondrocyte cytoskeleton and its response tochanging mechanical conditions in organ culture. J Anat 194(pt 3):343–353
Eggli PS, Hunziker EB, Schenk RK (1988) Quantitation of struc-tural features characterizing weight- and less-weight-bearing re-gions in articular cartilage: a stereological analysis of medial femo-ral condyles in young adult rabbits. Anat Rec 222:217–227
Farquharson C, Lester D, Seawright E, Jefferies D, Houston B(1999) Microtubules are potential regulators of growth-platechondrocyte differentiation and hypertrophy. Bone 25:405–412
Garavito RM, Rosenbusch JP (1986) Isolation and crystallizationof bacterial porin. Methods Enzymol 125:309–328
Gibbons IR (1981) Cilia and flagella of eukaryotes. J Cell Biol91(3 pt 2):107s–124s
Glotzer JB, Ephrussi A (1996) mRNA localization and the cy-toskeleton. Semin Cell Dev Biol 7:357–365
Grodzinsky AJ (1990) Mechanical and electrical properties andtheir relevance to physiological processes. In Maroudas A, Kuett-ner ICE, eds. Methods in Cartilage Research. New York, Aca-demic Press, 275–281
Guilak F (1995) Compression-induced changes in the shape andvolume of the chondrocyte nucleus. J Biomech 28:1529–1541
Hayes AJ, Benjamin M, Ralphs JR (1999) Role of actin stress fi-bres in the development of the intervertebral disc: cytoskele-tal control of extracellular matrix assembly. Dev Dyn 215:179–189.
Heath JP, Holifield BF (1991) Cell locomotion: new research testsold ideas on membrane and cytoskeletal flow. Cell Motil Cy-toskel 18:245–257
Hirsch MS, Cook SC, Killiany R, Svoboda KKH (1996) In-creased cell diameter precedes chondrocyte terminal differen-tiation, whereas cell-matrix attachment complex proteins ap-pear constant. Anat Rec 244:284–296
Hirsch MS, Lunsford LE, Trinkaus–Randall V, Svoboda KKH(1997) Chondrocyte survival and differentiation in situ are in-tegrin mediated. Dev Dyn 210:249–263
Janmey PA, Euteneuer U, Traub P, Schliwa M (1991) Viscoelas-tic properties of vimentin compared with other filamentousbiopolymer networks. J Cell Biol 113:155–160
Juliano RL, Haskill, S (1993) Signal transduction from the extra-cellular matrix. J Cell Biol 120:577–585
Jurvelin JS, Buschmann MD, Hunziker EB (1997) Optical andmechanical determination of Poisson’s ratio of adult bovinehumeral articular cartilage. J Biomech 30:235–241
Karim OM, Seki N, Pienta KJ, Mostwin JL (1992) The effect ofage on the response of the detrusor to intracellular mechani-cal stimulus: DNA replication and the cell actin matrix. J CellBiochem 48:373–384
Kempson GE, Muir H, Pollard C, Tuke M (1973) The tensileproperties of the cartilage of human femoral condyles relatedto the content of collagen and glycosaminoglycans. BiochimBiophys Acta 297:456–472
Kim YJ, Bonassar LJ, Grodzinsky AJ (1995) The role of cartilagestreaming potential, fluid flow and pressure in the stimulationof chondrocyte biosynthesis during dynamic compression. JBiomech 28:1055–1066
Kolmonen P, Hunziker EB, Buschmann MD, Jurvelin JS (1997) De-termination of the interstitial deformation of articular cartilage inunconfined compression. Trans Orthop Res Soc 22:2279
Koukouritaki SB, Theodoropoulos PA, Margioris AN, GravanisA, Stournaras C (1996) Dexamethasone alters rapidly actinpolymerization dynamics in human endometrial cells—evi-dence for nongenomic actions involving cAMP turnover. JCell Biochem 62:251–261
Lazarides E (1980) Intermediate filaments as mechanical integra-tors of cellular space. Nature 283:249–256
Mallein–Gerin F, Garrone R, van der Rest M (1991) Proteoglycanand collagen synthesis are correlated with actin organization indedifferentiating chondrocytes. Eur J Cell Biol 56:364–373
Maniotis AJ, Chen CS, Ingber DE (1997) Demonstration of me-chanical connections between integrins, cytoskeletal filaments,and nucleoplasm that stabilize nuclear structure. Proc Natl AcadSci USA 94:849–854
Martin I, Vunjak–Novakovic G, Yang J, Langer R, Freed LE(1999) Mammalian chondrocytes expanded in the presenceof fibroblast growth factor 2 maintain the ability to differentiateand regenerate three-dimensional cartilaginous tissue. Exp Cell Res253:681–688
Mitchison T, Evans L, Schulze E, Kirschner M (1986) Sites ofmicrotubule assembly and disassembly in the mitotic spindle.Cell 45:515–527
Muir H, Bullough P, Maroudas A (1970) The distribution ofcollagen in human articular cartilage with some of its physio-logical implications. J Bone Joint Surg [B] 52:554–563
Otto JJ, Schroeder TE (1990) Association of actin and myosin inthe contractile ring. Ann NY Acad Sci 582:179–184
Ralphs JR, Benjamin M, Thornett A (1991) Cell and matrix biol-ogy of the suprapatella in the rat: a structural and immunocy-tochemical study of fibrocartilage in a tendon subject to com-pression. Anat Rec 231:167–177
Ralphs JR, Tyers RN, Benjamin M (1992) Development of func-tionally distinct fibrocartilages at two sites in the quadricepstendon of the rat: the suprapatella and the attachment to thepatella. Anat Embryol (Berl) 185:181–187
Riezman H, Munn A, Geli MI, Hicke L (1996) Actin–, myosin– andubiquitin–dependent endocytosis. Experientia 52:1033–1041
Roth V, Mow VC (1980) The intrinsic tensile behavior of the ma-
by guest on July 3, 2015jhc.sagepub.comDownloaded from

1320 Langelier, Suetterlin, Hoemann, Aebi, Buschmann
trix of bovine articular cartilage and its variation with age. JBone Joint Surg [Am] 62:1102–1117
Schenk RK, Eggli PS, Hunziker EB (1986) Articular cartilage mor-phology. In Kuettner KE, Schleyerbach R, Hascall VC, eds. Articu-lar Cartilage Biochemistry. New York, Raven Press, 3–22
Schinagl RM, Ting MK, Price JH, Sah RL (1996) Video microscopyto quantitate the inhomogeneous equilibrium strain within artic-ular cartilage during confined compression. Ann Biomed Eng24:500–512
Simon VR, Pon LA (1996) Actin–based organelle movement. Expe-rientia 52:1117–1124
Sims JR, Karp S, Ingber DE (1992) Altering the cellular mechanicalforce balance results in integrated changes in cell, cytoskeletaland nuclear shape. J Cell Sci 103:1215–1222
Sontag JM, Aunis D, Bader MF (1988) Peripheral actin filamentscontrol calcium-mediated catecholamine release from streptolysin-O-permeabilized chromaffin cells. Eur J Cell Biol 46:316–326
Speer DP, Dahners L (1979) The collagenous architecture of articu-lar cartilage. Correlation of scanning electron microscopy andpolarized light microscopy observations. Clin Orthop 139:267–275
Stockwell RA (1967) The lipid and glycogen content of rabbit ar-
ticular hyaline and non-articular hyaline cartilage. J Anat 102:87–94
Traub P (1995) Intermediate filaments and gene regulation. PhysiolChem Phys Med NMR 27:377–400
Turner CE, Burridge K (1991) Transmembrane molecular as-semblies in cell-extracellular matrix interactions. Curr OpinCell Biol 3:849–853
Vale RD (1987) Intracellular transport using microtubule-basedmotors. Annu Rev Cell Biol 3:347–378
Wang N, Butler JP, Ingber DE (1993) Mechanotransduction across thecell surface and through the cytoskeleton. Science 260:1124–1127
Weber K, Rathke PC, Osborn M (1978) Cytoplasmic microtubularimages in glutaraldehyde-fixed tissue culture cells by electron mi-croscopy and by immunofluorescence microscopy. Proc NatlAcad Sci USA 75:1820–1824
Wong M, Wuethrich P, Eggli P, Hunziker E (1996) Zone-spe-cific cell biosynthetic activity in mature bovine articular carti-lage: a new method using confocal microscopic stereologyand quantitative autoradiography. J Orthop Res 14:424–432
Woo SL, Akeson WH, Jemmott GF (1976) Measurements ofnonhomogeneous, directional mechanical properties of artic-ular cartilage in tension. J Biomech 9:785–791
by guest on July 3, 2015jhc.sagepub.comDownloaded from