the journal of physiology, vol. 243, no. 1 frontispiece - ncbi
TRANSCRIPT

The Journal of Physiology, Vol. 243, No. 1 Frontispiece
eI
r
sT
'A
./or
.s> jr j __
A. F. HUXLEY

J. Physiol. (1974), 243, pp. 1-43 1With 1 plate and 12 text-figure8Printed in Great Britain
REVIEW LECTURE
MUSCULAR CONTRACTION
GIVEN AT THE MEETING OF THE PHYSIOLOGICAL SOCIETY AT
LEEDS UNIVERSITY ON 14--15 DECEMBER 1973
BY A. F. HUXLEYFrom the Department of Physiology,
University College London, Gower Street, London WC1E 6BT
INTRODUCTION
It is a particular pleasure to me that this Review Lecture is being heldin the university where the late Professor W. T. Astbury worked. He wasamong the first to investigate muscles by means of X-ray diffraction, andhis successes and his enthusiasm prepared the way for this technique tobecome the most powerful method for investigating the submicroscopicstructure of living muscle and its changes during physiological activity.
In this lecture, I shall mainly be going over the development, during thelast 20 years, of ideas about the origin of the force produced by striatedmuscle. In the same period there have also been great advances in otheraspects of muscle physiology, and by way of introduction I will first runover the steps that are now believed to lead up to the contractile processitself. Throughout, I shall confine myself to the twitch fibres of striatedmuscle of amphibians and mammals, and I shall concentrate on experi-ments with living muscle, especially those on isolated fibres. Structuraland biochemical investigations have been at least equally productive,but I am not able to do justice to these aspects, and in any case, thephysiological side provides more than enough material for a single lecture.
Activation of the contractile materialExcitation. An action potential in an individual muscle fibre is set up at
any point where the internal potential is raised by some 40 mV above itsresting level (about -90 mV relative to the surrounding fluid). This mayoccur either at the end-plate (when stimulation is through the motor nervefibre) or opposite the cathode if direct stimulation is employed. The actionpotential propagates in both directions to the ends of the fibre, by amechanism which differs only in details from that which operates in nervefibres (Hodgkin, 1951).
I -2

It is the change of membrane potential itself (not, for example, anyassociated longitudinal current in the interior of the fibre) that is effectivein causing the fibre contents to contract (Kuffler, 1946). The relationbetween membrane potential change and mechanical response is graded(apart from the phenomena mentioned in the next section), and the all-or-none character of the twitch is a consequence of the all-or-nonecharacter of the electrical change at the surface membrane (E. D. Adrian,1922; Gelfan, 1931, 1933).Inward spread. As well as propagating along the surface membrane,
however, this potential change also spreads inwards along the mem-branes of the tubules, invaginated from the surface membrane, whichconstitute the 'T' (transverse) system. These tubules run in the spacesbetween the myofibrils, forming transverse networks across the fibre atdefinite positions in the striation pattern, and they are open to theextracellular space (Endo, 1964, 1966; H. E. Huxley, 1964a; Page, 1964;D. K. Hill, 1964; Franzini-Armstrong & Porter, 1964), although theiropenings have seldom been clearly seen in electron micrographs ofmammal-ian or adult amphibian muscle. In amphibians there is one transversenetwork per sarcomere, at the level of the Z line, while in mammals thereare two per sarcomere, one near each boundary between A and I bands;in a bird, both of these patterns have been seen in different muscles (Page,1969).
If the lumen of one of these networks is made electrically negative byapplying an electrical pulse to a micropipette in contact with the surfaceof the fibre over one of the openings, a highly localized contraction occurs,spreading inwards for a substantial distance but confined to a single Iband in muscles whose transverse tubules are at the level of the Z line(A. F. Huxley & R. E. Taylor, 1955, 1958), or to a single half-sarcomereif there are two sets of tubules per sarcomere (A. F. Huxley & R. W.Straub, 1958). The electrical conduction in the transverse tubules hasrecently been shown to be a regenerative process (Costantin, 1970;Costantin & S. R. Taylor, 1971), similar to the longitudinal propagationalong the surface membrane but with much lower safety factor (R. H.Adrian & Peachey, 1973), and this accounts for the remarkable speed withwhich the onset of shortening spreads inwards within each fibre (Gonzalez-Serratos, 1971).In the local-stimulation experiments with a small-diameter pipette,
one of these two-dimensional networks is being stimulated through asingle tubule connecting to the outside of the fibre; this situation is un-favourable for propagation and, together with the low safety-factor forpropagation when the network is simultaneously stimulated at all of itsconnexions to the outside, probably accounts for the fact that the
2 A. F. HUXLEY

MUSCULAR CONTRACTION
response did not spread inwards beyond 10 Iam or so. With larger pipettes,Strickholm (1962, 1966) did sometimes obtain responses in which severalsarcomeres contracted over the whole cross-section of an isolated fibre,and the graded local contractions obtained by Gelfan (1933) in responseto a long-lasting positive potential applied through a pipette of 15 /tmdiameter contained an initial twitch-like component. There is no explana-tion yet for the remarkable responses recorded by Sugi & Ochi (1967),in which a ring-shaped contraction encircling the fibre took place when arather strong negative pulse was applied to a pipette in contact with thefibre surface.
Intracellular calcium release. For much of their length, these transversetubules are flanked on both sides by vesicles belonging to the sarcoplasmicreticulum, known as the lateral cisternae. The membrane potential changein the tubules of the T system somehow causes calcium ions to be liber-ated from the lateral cisternae; the mechanism is not known but smallmembrane currents probably associated with this 'gating' process haverecently been described (Schneider & Chandler, 1973). The release ofcalcium ions was first inferred from indirect evidence, but has now beendemonstrated by two elegant direct methods. Jdbsis & O'Connor (1966)succeeded in getting the calcium-sensitive dye, murexide, to enter thecells of toad muscle through their surface membrane, and recorded changesof light absorption that indicated a transient rise in intracellular calciumconcentration when the muscle was stimulated. The second method makesuse of aequorin, a protein isolated from a jellyfish, Aequorea (Shimomura,Johnson & Saiga, 1962), which luminesces when exposed to calcium ionseven at concentrations of the order of 10-6 M. A solution of aequorin isinjected into a muscle fibre, and when the fibre is stimulated an emissionof light occurs. This experiment was first performed by Ridgway & Ashley(1967; Ashley & Ridgway, 1970) in the very large muscle fibres of abarnacle, and the remarkable feat of doing the same thing in an isolatedfrog muscle fibre was reported to the Society earlier this year (Rudel &S. R. Taylor, 1973). Both methods show that after a single stimulus thefree calcium concentration reaches its maximum early in the rising phaseof tension, and has dropped practically back to its resting level by thetime the tension reaches its peak.These calcium ions diffuse into the interior of the myofibrils, where they
bring the contractile proteins into the active state by combining with theregulatory protein 'troponin' (Ebashi, 1963; Ebashi & Kodama, 1965).This protein, itself consisting of three separable components, is a consti-tuent of the thin filaments, and appears to exert its control on the inter-actions of myosin and actin through yet another component of thethin filaments, the native form of the protein tropomyosin (Bailey, 1948).
3

Relaxation is brought about by active transport of calcium into longi-tudinal elements of the sarcoplasmic reticulum, lowering the free calciumconcentration so that calcium dissociates from the troponin, and the restingstate of the contractile material is restored.The role of calcium in the regulation of contraction was very well
reviewed a few years ago by Ebashi and his colleagues (Ebashi & Endo,1968; Ebashi, Endo & Ohtsuki, 1969).
THE MECHANISM OF CONTRACTION
The position in 1945-1953The accepted background. By the end of the second world war, a large
body of knowledge of the biochemical and physiological aspects of muscu-lar contraction had been built up. Dr Dorothy Needham's 'MachinaCarnis' (1971) provides a most valuable account of this, as well as of morerecent work, while the numerous contributions of A. V. Hill and his schoolare beautifully reviewed in his 'Trails and Trials in Physiology' (1965).Some of the most important points are summarized in the following
paragraphs; subsequent work has confirmed all of them, with quantitativemodifications in some respects. They still provide a background to alltheorizing about the processes which actually give rise to tension andshortening.A ATP is the link between metabolism and contraction in the sensethat (a) this substance is synthesized from ADP and inorganic phosphateas a result of glycolytic and oxidative metabolism within each musclecell and (b) shortening of the muscle is coupled to the reconversion ofthis substance to ADP+Pi. Phosphorylcreatine (PCr) acts as a buffer,enabling relatively large quantities of ATP to be made available on ashort time scale by the Lohmann reaction:
ADP + PCr = ATP + creatine.
B 'Myosin' (Ktihne, 1864) is itself an ATPase (Engelhardt & Lyubi-mova, 1939).C The physical properties of myosin threads (Engelhardt, Lyubimova &Meitina, 1941) and of myosin in solution (Needham, Shen, Needham &Lawrence, 1941; Dainty et al. 1944) are drastically altered when ATP ispresent.D KUhne's 'myosin' can be separated into two proteins, 'actin', andwhat is now known as myosin (F. B. Straub, 1943). In this separation,the proteins are dissociated from a complex, 'actomyosin', by the actionof ATP (A. Szent-Gybrgyi, 1943).E When a muscle is fully activated by tetanic stimulation, it will
4 A. F. HUXLEY
MUSCULAR CONTRACTION
1 *0'
08
0
C-
06
04
02
1-4
1-2
02 04 06 08 1-0V/Vmax
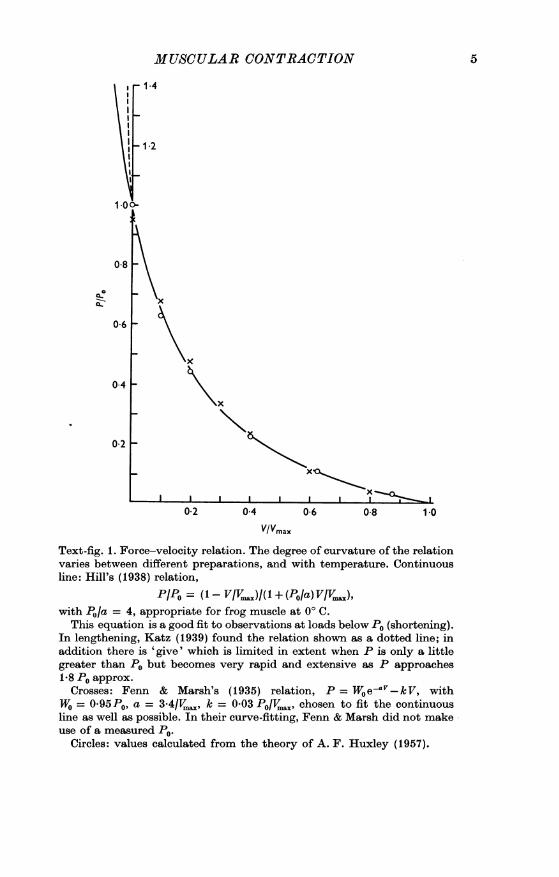
Text-fig. 1. Force-velocity relation. The degree of curvature of the relationvaries between different preparations, and with temperature. Continuousline: Hill's (1938) relation,
P/PO = (1-VVIV.)I(l + (PO/a) VIV..),with P./a = 4, appropriate for frog muscle at 0° C.
This equation is a good fit to observations at loads below P0 (shortening).In lengthening, Katz (1939) found the relation shown as a dotted line; inaddition there is 'give' which is limited in extent when P is only a littlegreater than P0 but becomes very rapid and extensive as P approaches1-8 P0 approx.
Crosses: Fenn & Marsh's (1935) relation, P = Woe aV-kV, withWO = 0*95P0, a = 3.4/VI,, k = 0-03 P0/V., chosen to fit the continuousline as well as possible. In their curve-fitting, Fenn & Marsh did not makeuse of a measured P0.
Circles: values calculated from the theory of A. F. Huxley (1957).
5
shorten at a speed which is a definite function of the load applied toit (Fenn & Marsh, 1935). A. V. Hill (1938) showed that to a sufficientdegree of approximation, this function is part of a rectangular hyperbola(Text-fig. 1).F When a load greater than the isometric tetanus tension (PO) isapplied to a stimulated muscle, the steady speed of lengthening is muchsmaller than would be expected from an extrapolation from speeds ofshortening at loads below P0 (Hill, 1938; Katz, 1939; Text-fig. 1). Inaddition, the muscle 'gives' when the load is first applied; the extent ofthis 'give' is small when the load is only a little greater than P0 but be-comes very large, simulating complete relaxation, as the load approachesa value of approximately 1-8 P0 (Katz, 1939).
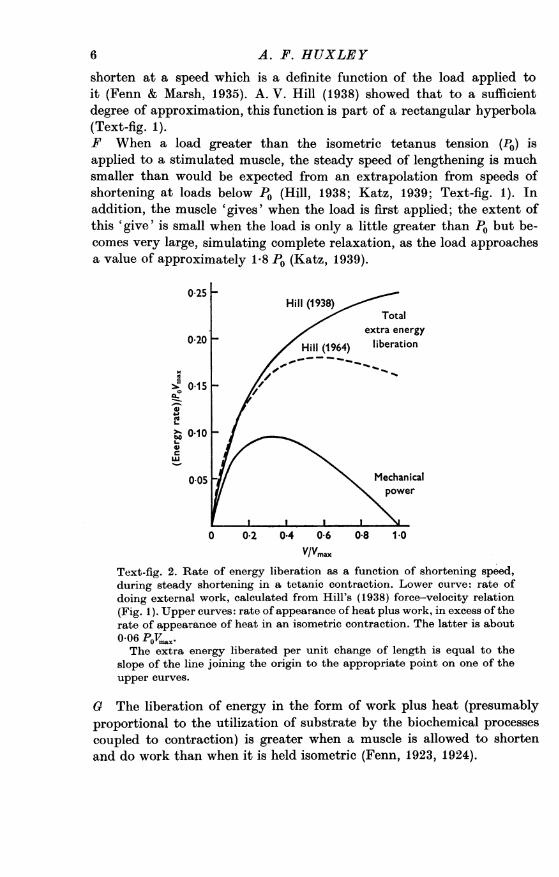
025Hill (1938)
Total
0.20extra energy
Hill (1964) liberation
> 0110C
0 05 M mechanicalg S~~~~~~~ower
0 0.2 0.4 0.6 0-8 1.0VIVmax,
Text-fig. 2. Rate of energy liberation as a function of shortening speed,during steady shortening in a tetanic contraction. Lower curve: rate ofdoing external work, calculated from Hill's (1938) force-velocity relation(Fig. 1). Upper curves: rate of appearance of heat plus work, in excess of therate of appearance of heat in an isometric contraction. The latter is about0-06 PoV,,,X.The extra energy liberated per unit change of length is equal to the
slope of the line joining the origin to the appropriate point on one of theupper curves.
G The liberation of energy in the form of work plus heat (presumablyproportional to the utilization of substrate by the biochemical processescoupled to contraction) is greater when a muscle is allowed to shortenand do work than when it is held isometric (Fenn, 1923, 1924).
A. F. HUXLEY6
MUSCULAR CONTRACTIONH This increase in the rate of energy liberation flattens off at higherspeeds of shortening (A. V. Hill, 1938; Text-fig. 2), i.e. the energy liberatedper unit change of length becomes less as the speed of shortening isincreased.I During forced lengthening, the rate of energy liberation (heat producedminus work done on the muscle) is much less than the rate during iso-metric contraction (Fenn, 1924; Hill, 1938; Aubert, 1944a, b, 1948).J The wide-angle X-ray diffraction picture is not very different in activemuscle from what it is in relaxed muscle (Astbury, 1947).
100
SPC.0
0 50
a ~~~~~~~~A4
0 Length (%/) 50 100 150 200
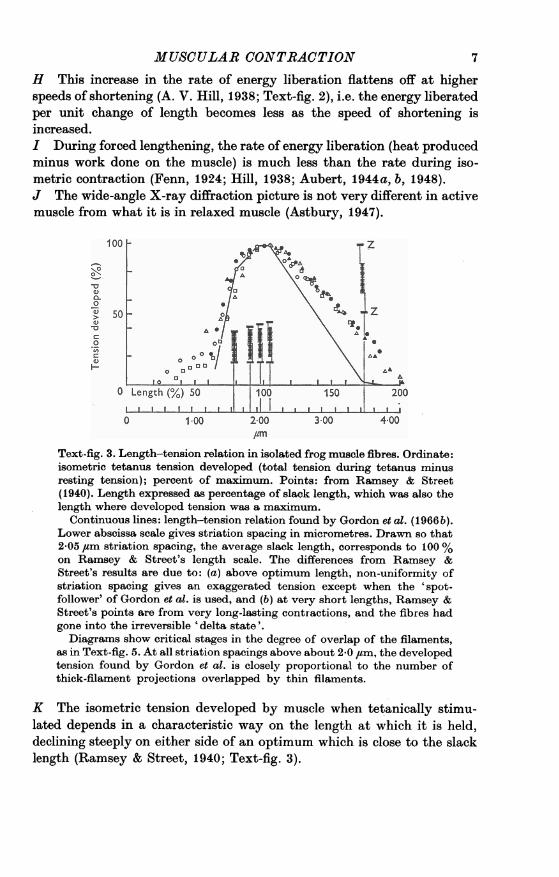
01.00 2-00 3-00 4-0Text-fig. 3. Length-tension relation in isolated frog muscle fibres. Ordinate:isometric tetanus tension developed (total tension during tetanus minusresting tension); percent of maximum. Points: from Ramsey & Street(1940). Length expressed as percentage of slack length, which was also thelength where developed tension was a maximum.
Continuous lines: length-tension relation found by Gordon et al. (1966 b).Lower abscissa scale gives striation spacing in micrometres. Drawn so that2-05 ,um striation spacing, the average slack length, corresponds to 100 %on Ramsey & Street's length scale. The differences from Ramsey &Street's results are due to: (a) above optimum length, non-uniformity ofstriation spacing gives an exaggerated tension except when the 'spot-follower' of Gordon et al. is used, and (b) at very short lengths, Ramsey &Street's points are from very long-lasting contractions, and the fibres hadgone into the irreversible 'delta state'.Diagrams show critical stages in the degree of overlap of the filaments,
as in Text-fig. 5. At all striation spacings above about 2-0 /tm, the developedtension found by Gordon et al. is closely proportional to the number ofthick-filament projections overlapped by thin filaments.
K The isometric tension developed by muscle when tetanically stimu-lated depends in a characteristic way on the length at which it is held,declining steeply on either side of an optimum which is close to the slacklength (Ramsey & Street, 1940; Text-fig. 3).
7

Confirmation of these points. The evidence quoted in the last section alldates from before 1945. Among more recent papers which confirm andextend the conclusions are the following:A The role of ATP was originally postulated on very indirect evidence.Conversion of ATP to ADP during contraction could not be demonstratedexperimentally (except in extreme fatigue), but this was attributed torapid re-synthesis of the ATP by transphosphorylation from phosphoryl-creatine. Since the war, utilization of ATP has been repeatedly demon-strated in preparations where the contractile material is directly exposedto an artificial medium (glycerinated fibres; fragmented myofibrils; thestripped- or skinned-fibre preparation of Natori; see books by Weber(1958) and Arronet (1973)). Direct evidence ofATP utilization in excitable,intact muscle was at last obtained by Cain, Infante & Davies (1962), whoused fluorodinitrobenzene as an inhibitor of creatine phosphokinase toprevent the rephosphorylation of the ADP that was formed.B There is now an enormous literature on the ATPase activity of myosin,of proteolytic fragments of myosin, and of actomyosin. A recent review isthat of E. W. Taylor (1972).C and D The kinetics of the dissociation of actomyosin through inter-action with ATP have been investigated with rapid-reaction techniquesby several groups (Tonomura & Watanabe, 1952; Barany, Edman &Palis, 1952; Tonomura, 1972; Mommaerts, 1956; Lymn & Taylor, 1971).G and H Energy liberation as a function of shortening speed has beenfurther investigated by Hill (1949a, b, 1964), using measurements of therate of production of heat and of doing work. There has been controversywhether 'shortening heat' appears as a term in the total energy liberatedin a cycle of contraction and relaxation, or whether it might be a reflexionof some process that is reversed during relaxation. It is now seems clearthat much, but not all, of 'shortening heat' does appear as a contributionto the total energy liberated (Aubert & Lebacq, 1971; Dickinson &Woledge, 1973).
Hill (1964) showed that the rate of energy liberation falls off moresharply with increase of shortening speed than indicated in his 1938 paper;indeed, the rate passes through a maximum when the speed of shorteningis about 0-6 Vmax (Text-fig. 2).The energy which appears as heat is far from being accounted for by
the measured changes in ATP and PCr concentrations in the muscleduring the contraction (Kushmerick, Larson & Davies, 1969; Gilbert,Kretzschmar, Wilkie & Woledge, 1971).I The drastic fall in energy liberation when a muscle is stretched duringcontraction has been repeatedly confirmed both by myothermal methods(Abbott, Aubert & Hill, 1951; Abbott & Aubert, 1951; Hill & Howarth,
A. F. HUXLEY8

MUSCULAR CONTRACTION1959; Wilkie, 1968) and by measurement of phosphorylcreatine utilization(Wilkie, 1968) and appearance of inorganic phosphate (Curtin & Davies,1973). It is not yet clear whether the net energy liberation in a completetwitch can drop to zero, as suggested by the results of Hill & Howarth(1959) or whether there is always a residual energy liberation at leastequal to the 'activation heat'. The former would require that the energy-yielding reactions associated with the activation process can actually bereversed by stretch during the contraction; the latter would only requirethat stretching suppresses the ATP splitting by the contractile proteins.J Interest in X-ray diffraction studies of muscle has switched from thewide-angle pattern (related to spacings of less than 1 nm) to the low-anglepattern (related to spacings of 5-50 nm). H. E. Huxley has been thechief pioneer in this work (see, for example, H. E. Huxley (1953a) andH. E. Huxley & Brown (1967)). An excellent review was published in 1968by Professor Jean Hanson, whose death this summer was a severe andunexpected blow to her many friends, and also to the progress of researchon muscle. Much of the more recent X-ray work is reviewed in the articlesin the section entitled 'Muscle Structure' in the volume produced fromthe 1972 Cold Spring Harbor Symposium (vol. 37, 1973). Many significantchanges in the diffraction pattern have been found, but the absence ofany suggestion of major change in the backbone structure of the filamentshas been confirmed throughout.K The length-tension curve was re-investigated by Gordon, A. F. Huxley& Julian (1966b), who confirmed the main features reported by Ramsey &Street (1940) but eliminated an error due to the non-uniformity of musclefibres when they are greatly stretched (see p. 17).
Continuous-filament theories. For many years, it had been generallysupposed that the contractile material was in the form of continuousfilaments that shortened by some kind of folding or coiling. Points A-Gin the preceding paragraphs were readily assimilated into this frameworkof ideas, and early electron microscope investigations of muscle structurewere claimed as confirming the existence of continuous filaments (Hall,Jakus & Schmitt, 1946; Rozsa, Szent-Gy6rgyi & Wyckoff, 1950). Duringthe period 1945-1953 it was almost universally supposed that the sequenceof events in a contraction was as follows.
1. Actin and myosin exist as separate proteins in the resting musclefibre (cf. point D).
2. When the fibre is stimulated, the actin and myosin interact to formcontinuous filaments of 'actomyosin'.
3. This actomyosin acts as an ATPase (point B), the action of the enzymeincluding a step which causes or allows the length of the filament to bereduced (point C).
9

This theory left open many questions about the intimate nature of theshortening process. Does the hydrolysis of ATP occur during the con-traction itself, or is contraction a spontaneous process, the actin andmyosin being dissociated by ATP during relaxation? Does the tendencyto shorten arise from the formation of new bonds in the shortened state('internal energy' theories: Meyer, 1929; Buchthal & Kaiser, 1951;Polissar, 1952), or is it attributable to an increased freedom of the fila-ments to coil up under the influence of thermal agitation ('entropytheories': Wohlisch, 1940; Pryor, 1950; Morales & Botts, 1952)?
In so far as ATP splitting is coupled to shortening, continuous-filamenttheories automatically account for point G. However, they provide nobasis for point H, i.e. chemical change per unit change of length is lessthe higher the speed of shortening (Needham (1950) pointed out that thisfeature might be explained by cyclic action of the contractile units withina single contraction (see p. 20), but this idea is difficult to incorporate intoany continuous-filament theory). The very specific theory worked out byPolissar (1952) accounted adequately for the force-velocity relation (pointE); he claimed to explain the rate of energy liberation (point H) but seemsto have mistaken A. V. Hill's 'shortening heat' for the total energyliberated in the shortening (the work done is an additional element ofliberated energy). The absence of change in the X-ray diffraction pattern(point I) could not be easily reconciled with the idea of folding of thefilaments.
Sliding filamentsThe continuous-filament view of muscle was upset by three discoveries
made just 20 years ago.1. The myosin is confined to the A bands, i.e. the bands with the higher
refractive index (Hasselbach, 1953; Hanson & H. E. Huxley, 1953).2. The filament array seen with the electron microscope within each
myofibril consists of two interdigitating sets of filaments (H. E. Huxley,1953b).
3. The length of the A bands does not alter when the muscle isstretched or shortened (Harman, 1954; H. E. Huxley & Hanson, 1954;A. F. Huxley & Niedergerke, 1954).
I shall not spend time either in discussing these experiments, as theyare well known and have been reviewed many times (e.g. Hanson &H. E. Huxley, 1955; A. F. Huxley, 1957; H. E. Huxley, 1960), or inreviewing the extent to which their results had been foreshadowedin forgotten work by nineteenth century microscopists and chemists(see A. F. Huxley, 1957). They led two of the groups of workers concernedto the conclusion that length changes in striated muscle take place
A. F. HUXLEY10

MUSCULAR CONTRACTION
by a relative sliding motion between two sets of filaments, each ofwhich retains a substantially unaltered length (H. E. Huxley, 1953b;H. E. Huxley & Hanson, 1954; A. F. Huxley & Niedergerke, 1954). This isrepresented in the familiar diagrams of Text-figs. 4 and 5. On the whole,
E
o
UI IL W 1 b, 2 05 ,um
,lllllludii11 1 1111,,,,,, ,,,r I"1 ,.l,,, 1,1.11.11,
...,,......... ,H ,,,4 B........1.|||1|||l111111111111Id lii .lll§11|1Ill'Il~lW§|ll
I*- a, 1 60 Aim 01005 jmPM
1*- A band lIband -01 H zone -,
| ~ ~ ~~~II I I
Text-fig. 4. Relation of striation pattern to the double system of inter-digitating filaments found by H. E. Huxley (1953b). The high refractiveindex and the birefringence of the A bands are due to the thick filaments,composed principally of myosin. The 'heavy meromyosin' part of eachmyosin molecule projects, but aregion at the middle ofeach thick filament isfree from these projections.The thin filaments are principally composed of actin but contain also
the tropomyosin and the troponin. The H zone is recognized in freshmaterial as a region in the middle of the A band with lower refractive indexthan the outer parts. The diagram also shows the definitions of the dimen-sions that are used in constructing Text-fig. 5.
Additional filaments, which are seen when the myosin is dissolved awayor when the muscle is stretched so far that there is a gap between thethick and thin filaments (p. 14), are not shown because it is not yet knownwhere they attach to the other structures in the filament array. (FromGordon et at. 1966b.)
this proposal was quickly accepted, and some observations which at firstseemed not to fit in with it have been satisfactorily explained. It is clearlyright to adopt it as a first approximation to what occurs when a musclechanges its length; the following sections will discuss how far it can beregarded as a complete account.How nearly constant are the filament lengths? Although both light and
electron microscopy showed that the filament lengths stay approximatelyconstant, neither method would be capable of detecting changes of say afew per cent if they were to occur either during passive length changesor when a muscle is activated. The resolving power of the light micro-scope is not sufficient, and electron microscope preparations are subject
11

to uncertain amounts of shrinkage. The best available measurements arethose made by low-angle X-ray diffraction: H. E. Huxley & W. Brown(1967) found no detectable change (i.e. less than a few tenths of one percent) in the thin-filament spacings, but an increase of about 1 % in thethick-filament spacings, when the pattern from a muscle in isometriccontraction is compared with one from resting muscle. The filamentsmust, of course, be extensible in some degree, and even an extensionof as little as 01% as a direct consequence of the isometric tension
3:65 ,um (a+b)
-. 2-20-2 25 /4m (b+c)-*-
2-... ... ... ...
-. 2 05 4um (b) -i .......................
3
4 1 85-1-900m (b -c)-
1.65 uim (a+z)
105ma R
i (b+z)6 - M2
Text-fig. 5. Critical stages in the increase of filament overlap as a fibreshortens. The symbols for the dimensions are defined in Text-fig. 4. Thestriation spacings for each stage of shortening are calculated from the fol-lowing values of the dimensions, all for frog muscle: Thick filamentlength, a: 1-60 ,um (Page & H. E. Huxley, 1963). Length of thin filamentsand Z line, b: 2-05 ,um (Page & H. E. Huxley, 1963). Thickness of Z line,z: 0 05 /tm. Region of thick filament free from projections, c: 0-15-0-20 #rm(H. E. Huxley, 1963).More recent work suggests that the thin filaments are somewhat shorter,
probably about 1-95 #rm (Page, 1968). (From Gordon et al. 1966 b.)
carried by the filaments would be appreciable when one is trying tointerpret the mechanical properties of active muscle (see Text-fig. 8).
There is no clear evidence about the significance of the increase in thick-filament spacings. Possible interpretations are:
(1) stretch caused by the isometric tension;(2) elongation related to the activation process, perhaps causing the
latency relaxation, and(3) a change of position of the myosin heads (generally assumed to be
12 A. F. HUXLEY

MUSCULAR CONTRACTIONthe actual scatterers on the thick filament) relative to the backbone of thefilament.
Events in extreme shortening. Frog striated muscle shortens readilyto a striation spacing of about 1X3 ,tm (Goydon, A. F. Huxley & Julian,1966b), and in long-lasting contractions it will shorten to about 1 jtm(Ramsey & Street, 1940), though in this case the shortening is not fullyreversible ('delta state'). The approximate filament lengths shown onText-fig. 5 make it clear that long before the spacing of 1-3 /tm is reachedsomething beyond the most straightforward sliding must occur: the thinfilaments will have met at the centre of the sarcomere, and the ends ofthe thick filaments will have come into contact with the Z lines.As regards the thin filaments, it was shown by electron microscopy
(H. E. Huxley, 1964b) that their ends slide past each other, giving aregion of 'double overlap' at the centre of the sarcomere (Text-fig. 5),which accounts for the CM band which appears in the light microscopewhen vertebrate striated muscle shortens actively (P1. 1). This band, aswell as the better known Cz contraction band, has been seen in fixed andstained preparations (Jordan, 1934), in living isolated fibres (A. F. Huxley& Niedergerke, 1954, 1958; A. F. Huxley & Gordon, 1962; Plate 1) and inseparated myofibrils (H. E. Huxley & Hanson, 1954).On further shortening, after the Z lines reach the ends of the thick
filaments, the other contraction band (Cz) appears, as a region of highprotein concentration or strong staining centred on the position of the Zline. As regards vertebrate muscle, no one has yet deciphered the tangledmass of material that is seen at this position under the electron microscope.
In frog muscle, at any rate, contraction bands are formed only duringactive shortening. Fibrils that are passively shortened below their slacklength, either by active shortening of another part of the cross-sectionof the same fibre (A. F. Huxley & Gordon, 1962; P1. 1) or by longitudinalcompression of an isolated fibre set in gelatin (Gonzalez-Serratos, 1971),are thrown into folds, and the sliding motion of filaments does not gobeyond (and may not even reach) the point where the thin filaments meetat the centre of the sarcomere (L. M. Brown, Gonzalez-Serratos & A. F.Huxley, 1970). On relaxation after active shortening below this length,the fibrils re-extend themselves, either throwing the whole fibre into curves(Ramsey & Street, 1940) or, if this is prevented by surrounding materials,making the fibrils go wavy within the straight cylindrical outline of thewhole fibre (Gonzalez-Serratos, 1971). A whole muscle can remain short-ened as regards its overall dimensions after relaxation, but a subsequentcontraction has a long latent period corresponding to the straighteningout of the fibrils before they can begin to exert tension (Gonzalez-Serratos,1971).
13

Other filamentous constituents. H. E. Huxley & Hanson (1954, 1957)gave clear evidence that after the thick filaments had been dissolvedaway, some continuous structure remained in the myofibril; filamentousmaterial has been seen with the electron microscope in this situation(Walcott & Ridgway, 1967). When a muscle is so far stretched that thereis a gap between the ends of the thick and thin filaments, some filamentousmaterial is seen crossing the gap (Carlsen, Knappeis & Buchthal, 1961;A. F. Huxley & Peachey, 1961; Sjbstrand, 1962; McNeill & Hoyle, 1967;Hoyle, 1968). It is not yet clear what these additional filaments arecomposed of, or where they attach to the main system of thick and thinfilaments. It is natural to suppose that they carry much or all of thatpart of the resting tension which is attributable to the fibre contents;this is substantial although the sarcolemma makes a large contributionat high degrees of extension (Casella, 1950; Podolsky, 1964; Fields &Faber, 1970; Rapoport, 1972).
In the fibrillar muscles of insects there are well defined connexions fromthe ends of the thick filaments to the Z lines (Auber & Couteaux, 1963)but there is no reason to think that vertebrate muscle contains similarstructures.
Cause of the sliding movementAlternative possibilities. As I mentioned earlier, it was soon accepted
that the shortening of vertebrate striated muscle took place by a relativesliding motion of two sets of interdigitating filaments. This was an essen-tial step that had to be taken before a realistic theory of contractioncould be developed, but in itself the 'sliding-filament theory' is not atheory of contraction. The question to which we want to find the answeris, what makes the filaments slide? The position is analogous to thatwhich existed in 1950, when the question was, what causes the filaments(assumed continuous) to fold?There are many kinds of interaction between two sorts of filaments
that might lead to movement which increases their overlap. The followingare some of those which have actually been proposed.
(1) On activation. the end of the thin filament becomes attached to anadjacent thick filament, and the part of the thin filament which is in theoverlap zone shortens (Podolsky, 1959).
(2) Some kind of 'vernier' action takes place between sites on the thickand thin filaments whose spacings are slightly different (Spencer &Worthington, 1960; H. E. Huxley, 1964b).
(3) One of the types of filament undergoes small cyclic changes oflength, the other type of filament being temporarily attached to it duringthe shortening phase of each cycle (Hanson & H. E. Huxley, 1955,
14 A. F. HUXLEY

MUSCULAR CONTRACTION
pp. 254-5; H. E. Huxley, 1960, p. 475; Asakura, Taniguchi & Oosawa,1963).
(4) Superthin (T) filaments, claimed to run continuously from Z to Zbetween the well known actin and myosin filaments, shorten under theinfluence of the ATPase activity of myosin and actin (McNeill & Hoyle,1967; Hoyle, 1968).
(5) A lateral repulsive force is generated between the filaments (Elliott,Rome & Spencer, 1970), or the Z disks expand laterally (Ullrick, 1967),and the consequent expansion of the lattice is coupled to shortening in theway that is actually observed (by low-angle X-ray diffraction) when intactmuscle changes its length.
(6) The filaments move so as to increase the number of sites at whichsome chemical interaction between actin and myosin can exist (H. E.Huxley & Hanson, 1954).
(7) Electric charges of opposite sign, distributed along the two typesof filament, generate a net longitudinal force tending to increase theamount of overlap (Yu, Dowben & Kornacker, 1970).
(8) Sites of interaction between the two types of filament, each genera-ting a force in the shortening direction, are distributed within each overlapzone (A. F. Huxley & Niedergerke, 1954). Examples of widely differenttheories within this general category are those of H. H. Weber (1956, 1958),A. F. Huxley (1957), Tonomura et al. (1961), Davies (1963), H. E. Huxley(1969), A. F. Huxley & Simmons (1971b) and McClare (1972).The first of these classes of theories (folding of thin filaments in overlap
zone) has been excluded because electron microscopy shows that the endsof the thin filaments move inwards, and may overlap each other at thecentre of the A band, during active contraction (H. E. Huxley, 1964b).The theories under headings (2), (3) and (4) have not been developed
to the point of making quantitative predictions which can be testedagainst measurements of tension, heat production etc., and in most of thecases there is no direct evidence for the events proposed as the origin of theforce. For these reasons they are not generally in favour, although theycannot be rigorously excluded.
Theories (5) which depend on lateral expansion as the primary eventare made unlikely by a recent observation by Matsubara & Elliott(1972). They measured the lateral spacing between filaments in fibresfrom which the sarcolemma had been removed, and found that the depend-ence on sarcomere length that had long been known in intact muscle didnot occur. Since these skinned fibres are perfectly capable of contraction,this makes it very improbable that lateral expansion is a causal link in thechain of events between chemical reaction and shortening.Maximization of number of interaction sites (6). The basis of this theory
15

is that the potential energy of the system is decreased by a definite amount(say /E) for each actin-myosin interaction that is formed. The number ofinteraction sites per unit cross-sectional area in one sarcomere is propor-tional to the length x of each overlap zone. Now x =a+m - s2, wherea = actin filament length (from Z line to its end), m = half myosin fila-ment length, and s = sarcomere length. Hence the potential energy of thesystem can be written as
E = EO-kxlAE= EO-kLXE(a+ m-s2).
The tension (per unit cross-section) generated by the tendency to moveso as to reduce the potential energy of the system is dE/ds, i.e. kAE/2.This is independent of the amount of overlap, in contradiction to thedirect proportionality suggested by the length-tension curve of Ramsey& Street (1940) (confirmed later by Gordon, A. F. Huxley & Julian,1966b; Text-fig. 3).An alternative way of reaching this result is as follows. Forces at inter-
action sites within the overlap zone will be as much in the lengthening asin the shortening direction and will thus cancel out; it is only at the endsthat there is a net attractive force which acts in the shortening direction.The situation at the ends is, to a first approximation, the same whetherthere is much or little overlap, so the force would not be expected to dependgreatly on overlap.A further disadvantage of this type of theory is that it does not account
for shortening beyond the stage where the whole of each thin filamentoverlaps with thick filament. This occurs at a sarcomere length of 1-6 or1-7 Mum, but, as mentioned on p. 13, an isolated frog muscle fibre shortensactually down to a sarcomere length of 1-3 or even 1 0 /hm.Yet another difficulty is that this type of theory leads to the expectation
that the net chemical change associated with shortening, and hence thetotal energy released as heat and work, will simply be proportional to theincrease in number of interaction sites and hence to the amount of shorten-ing. This is contrary to the observations of A. V. Hill (1938), mentionedas point H, p. 7. The disagreement is even more striking if the comparisonis made with Hill's more recent measurements (Hill, 1964; Fig. 2).
Electrostatic theories (7). These may be regarded as a sub-class of thetype of theory (6) discussed in the previous section, and they suffer fromall three of the difficulties pointed out there - failure to explain (a) theproportionality between tension and overlap, (b) the ability to shortenbeyond the length where overlap is complete, and (c) the decrease of energyliberation per unit change of length, as the speed of shortening is increased.Electrostatic theories, however, meet an additional difficulty in that
A. F. HUXLEY16

MUSCULAR CONTRACTIONthere is a high concentration of potassium ions within the muscle fibre,which would be expected to act as gegenions, and thus to screen any fixednegative charges on one of the filaments.As regards the dependence of tension on sarcomere length, Yu et al.
(1970) did introduce an auxiliary assumption into their electrostatictheory to make tension decline in proportion to overlap. I do not feelsatisfied by this way of dealing with the difficulty: the decrease of tensionwith increasing stretch is a feature which gives rise to several unpleasanttypes of instability, and one would not expect it unless it was a directconsequence of some inescapable aspect of the contraction mechanism.
Independent force-generators. Theories that come under the heading(8) have the immediate attraction that they lead to a direct proportion-ality between generated tension and overlap. This was suggested byRamsey & Street's (1940) length-tension curve, as was pointed out byA. F. Huxley & Niedergerke (1954). Ramsey & Street's curve, however,does not fall to zero until the degree of stretch is substantially greaterthan ought to be needed to bring overlap to zero (compare Text-figs. 3and 5). This point was investigated by Peachey and myself (A. F. Huxley& Peachey, 1961). We found a simple explanation for the discrepancy:when these isolated fibres from frog muscle are stretched, the ends do notelongate as much as the middle, so that a situation is reached where thestriation spacing is perhaps 3-8 Mum, with no overlap, for the main part ofthe fibre length, but drops to perhaps 3 0 Mm or less at the ends, so thatthere is appreciable overlap for the last few tenths of a millimetre of fibreat each end. When the fibre is stimulated with the tendons held, theseend regions shorten, stretching the middle part and causing the tensionto rise. In isotonic contractions, however, we found that no shorteningoccurred in the parts of the fibre that were sufficiently elongated toeliminate overlap. Podolsky (1964) also found that the stripped-fibrepreparation of Natori does not shorten in response to applied calcium ifit is stretched so far that no overlap is to be expected.
This non-uniformity introduces serious difficulties into determinationsof the true isometric length-tension curve. We overcame these by usinga photo-electric servo device to keep constant the distance between twomarkers, stuck on to an isolated fibre, which defined a part of the fibrelength within which the striation spacing was nearly uniform (Gordon,A. F. Huxley & Julian, 1966a, b). With this apparatus we obtained iso-metric tetanus tensions (Text-fig. 3) that were closely proportional to-overlap calculated from (a) the filament lengths obtained by Page &H. E. Huxley (1963) and (b) the distribution of projections on the thickfilaments (H. E. Huxley, 1957, 1963). The 'overlap' to which tension wasproportional was therefore the overlap of thin filaments with that part of
17

the thick filaments which carries projections, confirming not only the ideathat the interaction sites act as independent force generators, but also thesuggestion of H. E. Huxley (1957) that the projections from the thickfilaments, or 'cross-bridges', actually are the interaction sites.
It now appears (Page, 1968) that the thin-filament length of 2-05 gmgiven by Page & Huxley (1963) was too great, and that the true value(for twitch fibres of frog muscle) is probably -about 1-95 ,tm. This meansthat the fit between our tension measurements and the overlap calculatedfrom our striation spacings is not as good as it seemed at the time. Thepoint deserves re-investigation, although the discrepancy is not beyond therange of possible errors due to non-uniformity etc., combined withuncertainty in the thick-filament length.Another feature of contraction due to independent force-generators
in the overlap zone is that the speed of shortening under zero load oughtto be independent of the amount of overlap: the speed of sliding thatreduces the net power output from any cross-bridge (and therefore itscontribution to tension) to zero will be the same however many othercross-bridges are active in the same overlap zone. One aspect of thiswould show up in comparisons between muscle types with differentsarcomere (and filament) lengths: a muscle with shorter sarcomeres has alarger number of overlap zones per millimetre of fibre length, and shouldtherefore shorten more rapidly than a muscle with longer sarcomeres. Arelation of this kind has long been known (Jasper & Pezard, 1934; Atwood,Hoyle & Smyth, 1965) in arthropods, where sarcomere lengths vary greatlyfrom one muscle to another, and Niedergerke and I (A. F. Huxley &Niedergerke, 1954) pointed out that this observation was consistent withthe idea of independent force generators in each overlap zone. Amongvertebrates there is almost no variation of sarcomere length, so a compari-son of this kind cannot be made, but we found (A. F. Huxley & Julian,1964; Gordon et al. 1966b) that reducing the overlap by increasing theinitial length of an isolated frog muscle fibre had practically no effect onthe speed of active shortening under very light load.The idea of independent force generators distributed over each overlap
zone therefore provides straightforward explanations for two strikingfeatures of contraction: isometric tetanus tension proportional to overlap,and speed of unloaded shortening independent of overlap. In my view,these points are sufficient grounds for provisionally assuming that themechanism of force generation is of this kind.
It should be said that these simple effects of changing the amount ofoverlap apply only in the range of lengths above the slack length of thefibre. Below the slack length, both isometric tension and speed of un-loaded shortening fall off steeply, and although some of the features of this
18 A. F. HUXLEY

MUSCULAR CONTRACTION
decline seemed to be related to the length of the filaments (Gordon etal. 1966b), it is now clear that a failure of inward spread of activation isat least part of the explanation (S. R. Taylor & RUdel, 1970; RUdel &S. R. Taylor, 1971).
Nature of the force generatorsIf on this basis we accept the idea that the relative force between thick
and thin filaments is made up of contributions from independent forcegenerators distributed within each overlap zone, and that these forcegenerators are to be identified with the projections seen by H. E. Huxley(1957, 1963) on the thick filaments, our next question is: how does each oneof them generate its force? Most of the ideas now current assume that thecross-bridges operate by some cycle of straightforward chemical inter-actions - formation and breakage of bonds of various kinds - betweenmyosin, actin and ATP. McClare (1971), however, claims that all suchtheories contain hidden assumptions and are contrary to the second lawof thermodynamics. He therefore asserts (1972) that it is necessary tosuppose that force is generated by some direct quantum-mechanicalinteraction between excited states in the parts of the filaments that areundergoing relative motion. I am not impressed by his arguments againstorthodox chemical theories: for example, he claimed that a theory ofmine (A. F. Huxley, 1957) would lead to a very low overall efficiency buthis argument depends on postulating an unreasonably large effect of ATPin changing the equilibrium between the attached and detached conditionsof a cross-bridge. There is at present no observation that suggests posi-tively that muscle is operated by a 'molecular energy mechanism' suchas McClare proposes, and until there is some such evidence, it remainsprofitable to follow up the possibilities of contraction mechanisms thatdepend on chemical processes that are better understood.The following are some points that are suggested by evidence that
already existed when sliding filaments were proposed.(a) Cyclic action. Each cross-bridge is presumably part of a myosin
molecule, and the total length of a myosin molecule is around 0-2 ,tm(Portzehl, 1950; Mommaerts, 1951; since confirmed by electron micro-scopy: Rice, 1961; Zobel & Carlson, 1963; H. E. Huxley, 1963; Rowe,1964). The extent of sliding that may occur in a single contraction may,however, be as great as 1 4am. It is therefore unlikely that any one cross-bridge can remain attached, and exerting force, throughout such a contrac-tion, and the natural conclusion is that, even within a single contraction,individual cross-bridges undergo cycles in which they attach, generate force,and detach.
(b) Detachment is presumably brought about by interaction with ATP.
19

A. F. HUXLEY
Szent-Gy6rgyi (1943) and Straub (1943) had shown that myosin and actinare dissociated from each other on addition of ATP (point D, p. 4),and this action has recently been shown to be very rapid (Lymn &E. W. Taylor, 1971). It is an open question whether detachment is theonly change brought about by ATP, or whether it also re-sets somecondition in the myosin molecule which enables it to generate force ina subsequent cycle.
(c) To explain point G (p. 6; increased energy release when shorteningis permitted), it is necessary to suppose that detachment (and thereforeATP utilization) is slow unless shortening occurs and the sliding move-ment allows the cross-bridge to complete the working part of its stroke.Theories which do not incorporate a feature of this kind (e.g. the theory ofOplatka, 1972) lead to a rate of energy liberation which is greatest in theisometric state, i.e. the reverse of the 'Fenn effect'.
(d) The rate constant for attachment of a free cross-bridge must beassumed to be moderate (in relation to speed of shortening) in order toexplain point H (p. 7), i.e. as speed of shortening increases, the energyliberated per unit change of length becomes smaller. With this assumption,the general explanation offered by Needham (1950) will operate: at lowspeeds of shortening, each cross-bridge will have time to attach to eachsite on the thin filament that comes within range as the filaments slidepast each other, but at higher speeds of sliding the rate constant forattachment becomes rate-limiting, and the cross-bridge will miss some ofits opportunities for undergoing a cycle of attachment to a thin-filamentsite.
(e) To explain point I (reduced energy liberation during stretch), theattachment process must be assumed reversible, without splitting of ATP,if the cross-bridge is prevented from going through its working stroke.At several points, these arguments depend on the assumption that the
measured rate of liberation of heat plus work at each instant is an indica-tion of the rate of consumption of ATP and PCr. This has not been con-firmed by direct measurements of the change in concentration of thesesubstances (Kushmerick et al. 1969; Gilbert et al. 1971), and the discrep-ancy between thermal and chemical measurements has not yet beenresolved. Caution is therefore necessary in accepting theoretical schemes(such as my own: A. F. Huxley, 1957) that are based on this assumption.
A. F. Huxley's 1957 theoryIn 1954-55 I worked out a theory of contraction (A. F. Huxley, 1957;
Text-fig. 6) which incorporated the features listed in the last section.Within this framework the structural and chemical assumptions were the
20

MUSCULAR CONTRACTION
simplest I could think of that would cause contraction to occur. Theassumptions about rate constants for attachment and detachment ofcross-bridges (both dependent on the position of the cross-bridge on thethick filament relative to the thin-filament site) were also of an elementarykind. With appropriate values for these rate constants and for the stiffnessand range of action of the cross-bridges, this theory gave an adequatefit both to the force-velocity curve and to the relations between load andrate of energy liberation found by A. V. Hill (1938). (Deshcherevskii(1968) has indeed shown that a closely related, though less complete,
Myosinfilament
ActinAfilament
Equilibrium position laxlyofM site - I I
Text-fig. 6. Mechanism of contraction proposed by A. F. Huxley (1957).The 'side-piece' M, elastically connected to the thick filament, is assumedcapable of binding to sites A (only one shown) on the thin filament.Attachment occurs spontaneously but may be reversible; detachment occursprincipally by a process which involves the hydrolysis of an ATP molecule.Attachment is accelerated by an enzyme attached to the thick filament,placed to the right of 0 and therefore effective only at moments when Mis displaced in this direction by Brownian motion. As soon as an attach-ment is made, the elastic connexion exerts a force in the direction tendingto shorten the muscle. The detachment process (utilizing ATP) is acceleratedby another enzyme attached to the thick filament, placed to the left of 0.Detachment therefore occurs rapidly after the sliding movement has broughtthe AM complex to positions where the elastic connexion exerts a forcewhich resists shortening.
kinetic theory leads exactly to the algebraic expressions fitted by Hill(1938) to his results.) I regard this as good grounds for thinking that thegeneral outline of the theory is correct - cross-bridges operating in acycle consisting of attachment with moderate rate constant, exertionof force, and detachment with high rate constant after sliding motion ofthe filaments sufficient to bring the force near to zero. The agreement withexperimental results would remain if these kinetic features were retainedbut the structural and chemical assumptions were changed; therefore,the agreement is evidence in favour of the kinetic features but not of thestructural and chemical ones.The theory leads naturally to an explanation of the discontinuity
21

between shortening and lengthening in the force-velocity curve (point F,p. 6; Text-fig. 1).
Since this theory was published, two kinds of experimental results havebeen obtained which are not explained by it. One of these was A. V. Hill'sfinding (1964) that 'shortening heat' was not simply proportional to theamount of shortening, as had appeared from his results published in 1938,but was relatively smaller when the speed of shortening was high (Text-fig. 2). The new results implied (Woledge, 1968) that, as speed of shorteningis raised, the total rate of energy liberation actually passes through amaximum, and falls at the highest shortening speeds. Chaplain & From-melt (1971) pointed out, correctly, that this result could not be accom-modated in the kinetic scheme of my 1957 theory. There are several waysin which this result might be explained; one which involves only a smallchange to the 1957 theory is to assume that attachment is not a single-stage but a two-stage process (A. F. Huxley, 1973), the cross-bridge beingeasily detached, without net chemical change, until the second step hasoccurred.The other group of experimental findings that are not accounted for
by the 1957 theory are the 'transient' responses that can be recordedwhen either the load on a muscle fibre, or its length, is suddenly changed.These are the subject of the following section.
Mechanical transient responses and their interpretationThe idea of investigating the contraction process by imposing a sudden
change of mechanical conditions during a contraction is not new. Experi-ments of this kind were done by Gasser & Hill (1924) and by Levin &Wyman (1927), and led to the representation of active muscle as twocomponents in series, a 'contractile component' and a 'series elasticcomponent'. Of these, the former was defined as having a definite force-velocity curve, i.e. at any instant its speed of shortening was determinedentirely by the load on the muscle at that instant. Similarly, the 'serieselastic component' was defined as having a length that was entirelydetermined by the load at that instant. The behaviour of a system consist-ing of these two elements in series is indeed a first approximation to thebehaviour of active muscle, but recent experiments with the much im-proved time resolution that has become available have shown that theresponses of muscle to sudden changes of load or length are in fact muchmore complicated (and correspondingly more interesting) than those ofthe two-component model.
' Velocity transients' and 'tension transients'. The first experiments toshow this more complicated behaviour were those of Podolsky (1960;Civan & Podolsky, 1966). A bundle of fibres was stimulated and held, at
A. F. HUXLEY22
MUSCULAR CONTRACTION 23
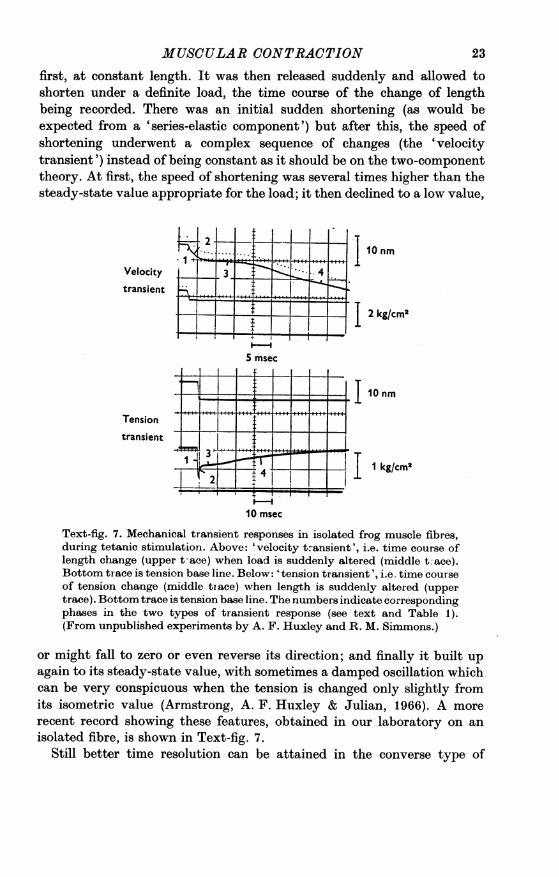
first, at constant length. It was then released suddenly and allowed toshorten under a definite load, the time course of the change of lengthbeing recorded. There was an initial sudden shortening (as would beexpected from a 'series-elastic component') but after this, the speed ofshortening underwent a complex sequence of changes (the 'velocitytransient') instead of being constant as it should be on the two-componenttheory. At first, the speed of shortening was several times higher than thesteady-state value appropriate for the load; it then declined to a low value,
Velocity 3X 4 0ntransient
-
2
2kg/cm2
5 msec.i
II -.. I II- qw
i ! i t....l....i....Tension
I110nr
transient ~ 3
X{l kg/cm2
10 msec
Text-fig. 7. Mechanical transient responses in isolated frog muscle fibres,during tetanic stimulation. Above: 'velocity transient', i.e. time course oflength change (upper t-ace) when load is suddenly altered (middle t ace).Bottom trace is tension base line. Below: 'tension transient', i.e. time courseof tension change (middle trace) when length is suddenly altered (uppertrace). Bottom trace is tension base line. The numbers indicate correspondingphases in the two types of transient response (see text and Table 1).(From unpublished experiments by A. F. Huxley and R. M. Simmons.)
or might fall to zero or even reverse its direction; and finally it built upagain to its steady-state value, with sometimes a damped oscillation whichcan be very conspicuous when the tension is changed only slightly fromits isometric value (Armstrong, A. F. Huxley & Julian, 1966). A morerecent record showing these features, obtained in our laboratory on anisolated fibre, is shown in Text-fig. 7.
Still better time resolution can be attained in the converse type of
A. F. HUXLEY
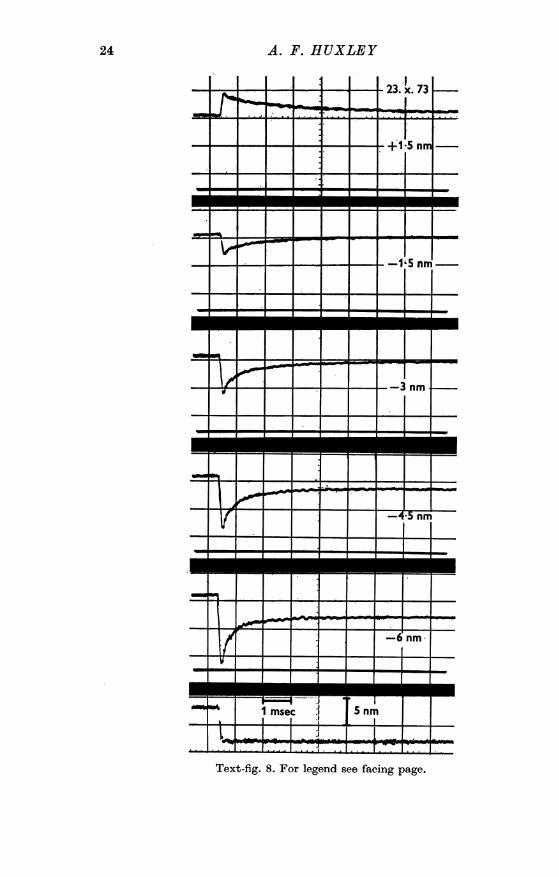
Text-fig. 8. For legend see facing page.
24

MUSCULAR CONTRACTION 25
experiment, in which a sudden small change of length is imposed during anisometric contraction, and the ensuing tension changes are recorded(Text-figs. 7 and 8). Again there is a tension change simultaneous with thelength change, as expected from a series-elastic component, and thesubsequent tension recovery - the 'tension transient' - is much morecomplex than the nearly exponential time course expected on the two-component theory. The phases of the recovery are set out in Table 1,which also shows the way in which they correspond to the phases of thevelocity transient in the other type of experiment.
TABLE 1. Phases of the transient response to sudden reduction of length('tension transient') or of load '(velocity transient')
Time of Events in Events inPhase occurrence 'tension transient' 'velocity transient'
1 During applied step Simultaneous drop of Simultaneous shorten-change tension ing
2 Next 1-2 ms Rapid early tension Rapid early shorteningrecovery
3 Next 5-20 ms Extreme reduction or Extreme reduction oreven reversal of rate even reversal ofof tension recovery shortening speed
4 Remainder of the Gradual recovery of Shortening at steadyresponse tension, with asymp- speed sometimes with
totic approach to superposed dampedisometric tension oscillation
The stated times are appropriate for frog muscle at 50 C approx.
Phase 1 can be attributed to a more or less instantaneous elasticity inthe fibre, though the evidence presented so far does not show where itresides or whether it is truly 'in series' with a separate contractilecomponent.
Phases 3 and 4 can be accounted for by detachment and re-attachmentof cross-bridges with kinetics not unlike those postulated in the 1957theory. Phase 3 would be a period in which both processes are actively at
Text-fig. 8. Family of transients. The records are obtained in the same wayas the bottom two traces in Text-fig. 7, but the time base is 10 times fasterand the step change of length (bottom panel) is complete in 0-2 ms whilethe step in Text-fig. 7 took approx. 1 ins.The distance specified in each of the upper panels is the amount of the
imposed length change in each half-sa-comere. (This Figure, and Text-figs. 9 and 10, are from recent unpublished experiments by L. E. Ford,A. F. Huxley and R. M. Simmons.)

work. Detachment comes to an end first, since it has the higher rate con-stant (points (c) and (d), p. 20), leaving attachment as the predominantprocess in Phase 4. In order for detachment to cause a tendency for tensionto fall (Phase 3), it is necessary to suppose that accelerated detachmentbegins when the cross-bridge is still in a position where it exerts positivetension (A. F. Huxley & Simmons, 1973, pp. 673-4); this is a point ofdifference from the 1957 theory. Julian, Sollins & Sollins (1973) haveworked out the time course of tension in Phase 3 and Phase 4 on a theoryof this kind, and obtained curves which resemble those given by realmuscle fibres very closely. I think that this kind of explanation of Phase 3may well prove correct, though it is not excluded that there may be amore direct 'de-activation by release' such as is widely postulated toexplain the oscillatory behaviour of the asynchronous flight muscles ofcertain insects (Pringle. 1949, 1967).
Phase 2, the early tension recovery in the tension transient, is the aspectof these phenomena to which our laboratory has so far been paying mostattention. It will be discussed in the next section.
The early tension recovery. Two main types of explanation for this phasehave been put forward. Podolsky and his colleagues (Podolsky, Nolan &Zaveler, 1969; Podolsky & Nolan, 1973) propose that during an isometriccontraction, many cross-bridges are not attached because thin-filamentsites are not available at the required positions. The sliding movement ofthe filaments during the shortening step brings some of these cross-bridgeswithin range of thin-filament sites; these cross-bridges attach rapidly andexert tension as soon as they are attached. There is evidence (Huxley &Simmons, 1971a; Text-fig. 10) that much or all of the instantaneouscompliance (phase 1 of the transients) resides in the cross-bridges, so thatthe increase in number of attached cross-bridges postulated by Podolskyshould be accompanied by a decrease in this compliance. Tests made byapplying a second step when the early recovery phase is nearly completehave shown that the stiffness is no greater than in the isometric condition(Ford, A. F. Huxley & Simmons, 1974), so we are inclined to think thatlittle, if any, of the early tension recovery can be attributed to attachmentof cross-bridges that were free up to the moment of the original step changeof length.The instantaneous tension drop followed by the early tension recovery
is roughly similar to the response of a 'Voigt element', i.e. a spring inseries with a parallel combination of spring and dashpot. If both of thesprings obey Hooke's law, straight lines will be obtained when T1 (theextreme tension reached simultaneously with the length step) and T2(the tension approached during the early recovery) are plotted against y,the amount of the length change. Text-fig. 9 shows that the T1 curve
26 A. F. HUXLEY
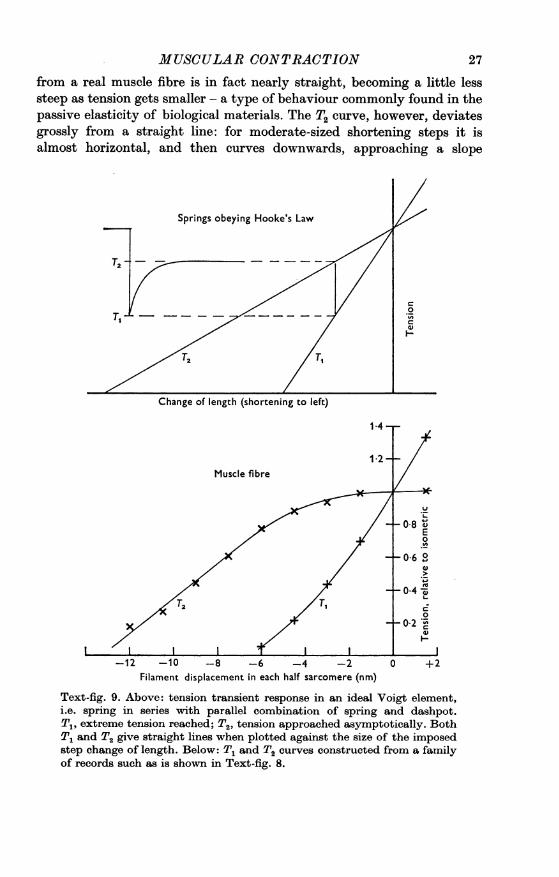
MUSCULAR CONTRACTIONfrom a real muscle fibre is in fact nearly straight, becoming a little lesssteep as tension gets smaller - a type of behaviour commonly found in thepassive elasticity of biological materials. The T2 curve, however, deviatesgrossly from a straight line: for moderate-sized shortening steps it isalmost horizontal, and then curves downwards, approaching a slope
Springs obeying Hooke's Law
1 4
1 2Muscle fibre
-10 -8 -6 -4 -2Filament displacement in each half sarcomere (nm)
U
08 EE0
06 °
0 4 0
+2
Text-fig. 9. Above: tension transient response in an ideal Voigt element,i.e. spring in series with parallel combination of spring and dashpot.T., extreme tension reached; T2, tension approached asymptotically. BothT1 and T2 give straight lines when plotted against the size of the imposedstep change of length. Below: T1 and T2 curves constructed from a familyof records such as is shown in Text-fig. 8.
27
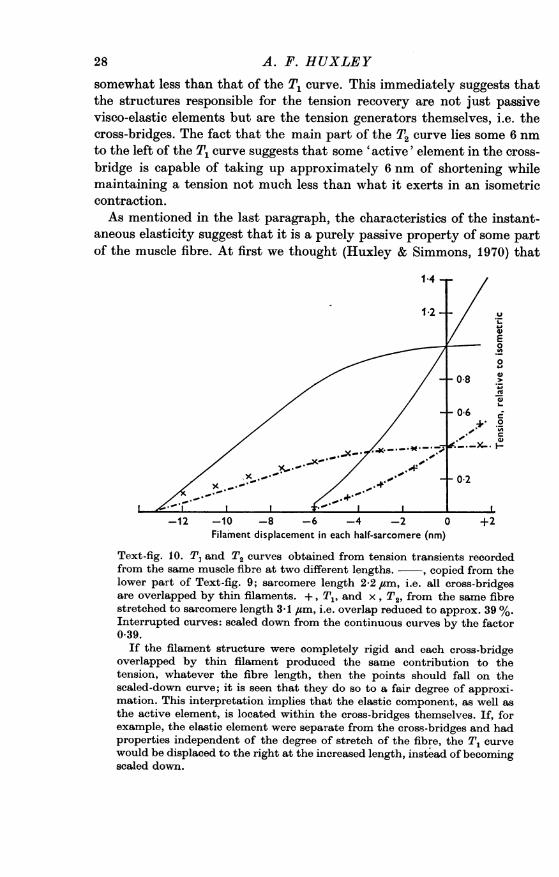
somewhat less than that of the T1 curve. This immediately suggests thatthe structures responsible for the tension recovery are not just passivevisco-elastic elements but are the tension generators themselves, i.e. thecross-bridges. The fact that the main part of the T2 curve lies some 6 nmto the left of the T1 curve suggests that some 'active' element in the cross-bridge is capable of taking up approximately 6 nm of shortening whilemaintaining a tension not much less than what it exerts in an isometriccontraction.As mentioned in the last paragraph, the characteristics of the instant-
aneous elasticity suggest that it is a purely passive property of some partof the muscle fibre. At first we thought (Huxley & Simmons, 1970) that
1-4
E0
0
-12 -10 -8 -6 -4 -2 0 +2Filament displacement in each half-sarcomere (nm)
Text-fig. 10. T, and T2 curves obtained from tension transients recordedfrom the same muscle fibre at two different lengths. - , copied from thelower part of Text-fig. 9; sarcomere length 2-2 /tm, i.e. all cross-bridgesare overlapped by thin filaments. +, T1, and x, T2, from the same fibrestretched to sarcomere length 3-1 ,tm, i.e. overlap reduced to approx. 39 %.Interrupted curves: scaled down from the continuous curves by the factor0 39.
If the filament structure were completely rigid and each cross-bridgeoverlapped by thin filament produced the same contribution to thetension, whatever the fibre length, then the points should fall on thescaled-down curve; it is seen that they do so to a fair degree of approxi-mation. This interpretation implies that the elastic component, as well asthe active element, is located within the cross-bridges themselves. If, forexample, the elastic element were separate from the cross-bridges and hadproperties independent of the degree of stretch of the fibre, the T, curvewould be displaced to the right at the increased length, instead of becomingscaled down.
28 A. P. HUXLEY

MUSCULAR CONTRACTIONit resided in the filaments and perhaps the Z line, constituting a true 'serieselastic component' whose properties were independent of the state of theforce generators themselves. There was, however, nothing to exclude thepossibility that it was due to some part of the cross-bridges. In order todecide between these possibilities, we recorded tension transients duringtetani with the fibre stretched to different lengths, i.e. with reducedamounts of overlap of thick and thin filaments. The result was simple(Huxley & Simmons, 1971 a; Text-fig. 10): both the T1 and T2 curvesscaled down in direct proportion to the amount of overlap. The straight-forward interpretation is that the instantaneous elasticity, as well as theearly recovery, is attributable to the cross-bridges, the filament arraysbeing almost completely rigid. The filaments must of course possess somecompliance, and it is difficult to put a limit to its possible amount, but wethink it unlikely to be large enough to lead to serious error if, as a firstapproximation, it is disregarded. We therefore assume, provisionally,that the whole of the instantaneous compliance resides in some componentof the cross-bridges.Two components within each cross-bridge. Thus, each cross-bridge seems
to contain (1) an instantaneous elastic element, and (2), in series with it,an element which can maintain tension while taking up limited but sub-stantial amounts of length change. The total range over which element(2) actively generates movement is probably 10-12 nm; it is likely to begreater than the 6 nm mentioned above because, after attaching, itpresumably has to generate a movement of the order of 5 nm so as tostretch the elastic element, even in an isometric contraction. This estimateof the range of action agrees as well as can be expected with the valuescalculated by H. E. Huxley (1960, p. 452) and by Davies (1963) from esti-mates of the tension per cross-bridge and the work that can be done permole of ATP utilized. They assume (as do most authors) that one moleculeof ATP is hydrolysed per cross-bridge cycle. Tonomura (1972, p. 390),however, proposes a cycle that utilizes two molecules of ATP, which wouldrequire the range of action to be twice as great.Can a cross-bridge exert negative tension? Another interesting feature of
the T1 curve is that it approaches the base line at a definite angle and nottangentially or asymptotically. The measured T. values are all higherthan they would be if a truly instantaneous length step were applied, sincethe early recovery begins before the step is complete, and this effect isrelatively greater in the large steps because the early recovery is morerapid (see below). The true T1 curve is therefore straighter (and steeper)than what we have been able to record, and it would approach theabscissa in an even more angular manner than is seen in Text-fig. 9.Blange, Karemaker & Kramer (1972) claimed that the curve approached
29

30 A. F. HUXLEYthe base line tangentially, and interpreted this by supposing that cross-bridges could not support negative tension and that more and more ofthem became slack as the size of shortening step was increased, reducingthe stiffness of the fibre and therefore the slope of the T1 curve. Ourresults do not confirm Blange et al. (1972), and the fact that we find asharp approach to the base line is evidence that the cross-bridges are ableto exert negative tension.
Evidence for stepwise action. When the time course of the early tensionrecovery is examined (Text-fig. 8), another interesting non-linear feature
a
I
Thick filamentLMM S
s-I1 2
Thin filament-.-
I
11Thick filament -
LMM I- 2
s-I 2
Thin filament-
11
3
Text-fig. 11. a, diagram of active cross-bridges adapted from H. E. Huxley(1969). He proposed that the force originates in a tendency for the myosinhead (S.1) to rotate relative to the thin filament, and is transmitted to thethick filament by the S-2 portion of the myosin molecule acting as aninextensible link. Flexible points at each end of S-2 permit S-1 to rotate,and allow for variations in the separation between filaments.
b, development from a proposed by A. F. Huxley & Simmons (1971 b)to incorporate the elastic and stepwise-shortening elements for which theygave evidence derived from tension transients. The strength ofbinding oftheattached sites are higher in position 2 than position 1, and in position 3than position 2. During isometric contraction the myosin head oscillatesrapidly between its three stable positions. The myosin head can be detachedfrom position 3 with the utilization of a molecule of ATP; this is thepredominant process during shortening. During stretch, the myosin headcan dissociate from position 1 without utilization of ATP.

MUSCULAR CONTRACTIONmakes itself evident. The half-time of the early tension recovery is shortestin a large release, becoming progressively longer as the size of the release isdecreased, and longer still in stretches, the variation being continuous overthe whole range that can be investigated. The following explanation wasproposed by A. F. Huxley & Simmons (1971 b). Shortening of element (2)of the cross-bridge takes place by rapid switching from one to the nextof a series of perhaps 2, 3 or 4 states, which have progressively lowerpotential energy. These switching movements are reversible, and in theisometric state an equilibrium is set up, each cross-bridge dividing itstime between the various states under the influence of (a) the potentialenergy differences, tending to cause forward movement and (b) the tensionin the elastic element (1) tending to cause backward movement. To make aforward switching movement, the cross-bridge requires activation energynot only to escape from the state it is already in, but also to do the workof stretching the elastic element far enough for it to get into the new state.The work term therefore appears as a component of the activation energyfor a forward movement; the higher the tension, the greater the activationenergy and therefore the slower the rate of transfer to the next position.The greater a shortening step, the less the tension in each elastic element,and therefore the less the activation energy for the forward steppingmovement of element (2). This motion therefore occurs more rapidly,and shows itself as an early recovery phase with shorter half-time.
Relation to structural and chemical information. The most widely acceptedview of the structure of a cross-bridge is that proposed by H. E. Huxley(1969, 1971) on the basis of observations (largely his own) by electronmicroscopy and low-angle X-ray diffraction. Text-fig. 11 a is an adaptationof his diagram. An important and suggestive feature of his proposal isthat the force originates in a tendency for the myosin head to rotate aboutits point of attachment to the thin filament, and is transmitted to the thickfilament by 'sub-fragment 2' of the myosin molecule which acts as alink. In H. E. Huxley's account, this link is assumed inextensible,and the connexions at its ends to the other parts of the myosin moleculeare assumed freely flexible.Can the instantaneous elastic element (1) and the stepwise shortening
element (2), whose existence is suggested by the tension transients, beidentified with particular parts of this structure? Simmons and I (Huxley& Simmons, 1971 b) tentatively proposed the arrangement shown inText-fig. 11 b, i.e. the elastic element is in S-2 and the stepping move-ment occurs by rotation of the whole myosin head relative to the thinfilament. I do not know of any evidence against this suggestion, and Istill use it in my own thinking about the contractile mechanism. But it isimportant to remember that there is no specific evidence in favour of it,
31
2 P H Y 243
32 A. F. HUXLEY
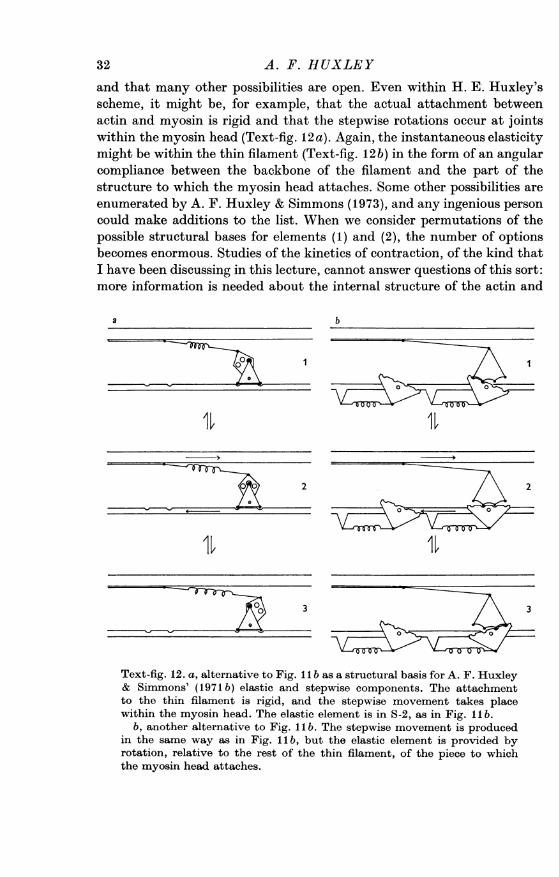
and that many other possibilities are open. Even within H. E. Huxley'sscheme, it might be, for example, that the actual attachment betweenactin and myosin is rigid and that the stepwise rotations occur at jointswithin the myosin head (Text-fig. 12 a). Again, the instantaneous elasticitymight be within the thin filament (Text-fig. 12b) in the form of an angularcompliance between the backbone of the filament and the part of thestructure to which the myosin head attaches. Some other possibilities areenumerated by A. F. Huxley & Simmons (1973), and any ingenious personcould make additions to the list. When we consider permutations of thepossible structural bases for elements (1) and (2), the number of optionsbecomes enormous. Studies of the kinetics of contraction, of the kind thatI have been discussing in this lecture, cannot answer questions of this sort:more information is needed about the internal structure of the actin and
a b
f0011 1 11
totetin fiamn irgd, an th,,pie oeettae lc
wihn h yoi_ ha.Th_ latcelmn 0a
Texant-fg.12ra alternative to Fig. 1 1 b asha strucurabsismoveen is produxled
in the same way as in Fig. II b, but the elastic element is provided byrotation, relative to the rest of the thin filament, of the piece to whichthe myosin head attaches.

MUSCULAR CONTRACTION
myosin molecules, and about the positions of the parts of the myosinmolecule during contraction. The latter may come from X-ray diffractionor from electron microscopy, or from optical or spin-resonance measure-ments of the orientation of components of the myosin molecule or of smallmolecules attached to it (Seidel & Gergely, 1973; Botts et al. 1973; Steigeret al. 1973; Carlson et at. 1973).
Relation to chemical events. If the idea of a stepwise movement of (orwithin) the myosin molecule is correct, it becomes necessary to supposethat the stepping is somehow coupled to the hydrolysis of ATP. Much isnow known about intermediate stages in the enzymic hydrolysis of ATPby the actomyosin ATPase, both from steady-state studies (e.g. Moos,1973) and from kinetic experiments (reviewed by Tonomura (1972),E. W. Taylor (1972) and Bagshaw et al. (1973)). As has already beenmentioned, one of the conspicuous actions of ATP is to cause actomyosinto dissociate; on a scheme such as the one shown in Text-fig. 11 b this actionwould be sufficient to drive the system, if it is assumed that this disso-ciation can only be brought about when the myosin head has moved toits final position (most clockwise position in Text-fig. 1 b). If, however,the stepping movement involves some internal re-arrangement within themyosin molecule (as in Text-fig. 12 a), it becomes necessary to postulateeither that this rearrangement is coupled to some stage of the hydrolysisof the ATP molecule, or else that the rearrangement occurs spontaneouslyand is reversed before the next cycle by coupling to ATP hydrolysis. Theproposal of Tonomura et al. (1961) requires the hydrolysis of two ATPmolecules per cycle, one to drive the conformational change and the otherto detach the myosin head from the thin filament.
There is at present no evidence to show which (if any) of these ways inwhich ATP hydrolysis might be coupled to movement is correct. I thinkthat most biochemists at the present day would favour the idea that some' conformational change' occurs in the myosin molecule soon after it bindsan ATP molecule, bringing it into a state which can do mechanical workwhen it becomes attached to a thin filament. The meaning of the word'conformational ', however, has become so broad that this statement wouldbe true whatever the nature of the effect on a myosin molecule of bindingan ATP - whether it was a gross shape change whose reversal could domechanical work, or a shift of electrons that leads to a change of affinityfor binding sites on the thin filament. I am reminded of the remark attribu-ted to Dean Inge (Mencken, 1936), that the word 'bloody' had become'simply a sort of notice that a noun may be expected to follow': 'con-formational' has become simply a notice that the word 'change' may beexpected to follow.
In this sense, the theory that the key event is a 'conformnational change'2-2
33

induced in the myosin molecule by some stage of the process of ATPhydrolysis, will almost inevitably be confirmed. Meanwhile, it is an un-helpful hypothesis because it does not specify what sort of change is tobe looked for.
CONCLUSION
In the summer of 1972, a highly successful symposium on 'The Mech-anism of Muscle Contraction' was held at Cold Spring Harbor. Everyonewho was there felt at the end that he knew a lot more about musclecontraction than he had known before the meeting, and the book producedfrom the contributions is a most valuable record of the present positionin most aspects of the field. But I was horrified to hear several of theparticipants saying that 'the problem of muscle contraction is solved inprinciple'. Even if the points set out in the Summary (para. 4) all turnout to be true, they only take one perhaps halfway to a real understandingofhow muscle contracts: for example, we do not know any ofthe following.
1. Which structure is the elastic element in the cross-bridge.2. What is the nature of this elasticity.3. What structure undergoes the stepwise change.4. Whether the attachment of the myosin head is to a single actin
monomer, or to two (or more) monomers within the thin filament.5. What kind of bonds hold the myosin head to the thin filament.6. How binding of ATP causes myosin to dissociate from actin.7. What is the significance of the fact that each myosin molecule has
two heads.
And, quite apart from the fact that all these things are unknown, thewhole history of theories of muscular contraction during the last halfcentury shows that even when a set of ideas seems to be well established,there is a large chance that it will be overthrown by some unexpecteddiscovery.
SUMMARY
1. A resume of the events in excitation and in excitation-contractioncoupling is given.
2. The theory that length changes in striated muscle take place pre-dominantly by relative sliding movements of two sets of submicroscopicfilaments was proposed in 1953-54 and was soon firmly established, dis-placing the idea of continuous filaments of actomyosin that shortened byfolding.
3. Many proposals have been made for the mechanism by which thesliding movement is brought about during contraction.
34 A. F. HUXLEY

MUSCULAR CONTRACTION4. A coherent, though still incomplete, theory is presented. Evidence
(in many cases dating from before 1945) is given for each of the followingfeatures:
a, shortening and the development of tension are produced by inde-pendent force-generators distributed in each zone of overlap,
b, these force-generators are identified with the cross-bridges seen inthe electron microscope,
c, within a single contraction, each force-generator may act repeatedlyin cycles of attachment, pulling, and detachment,
d, detachment is brought about during a contraction by interactionwith ATP,
e, detachment by ATP is slow until shortening has allowed the cross-bridge to perform the working part of its stroke,
f, the rate constant for attachment is moderate,g, attachment is reversible, without utilization of ATP, if the cross-
bridge is prevented from performing its working stroke,h, each cross-bridge contains a non-linear element which takes up some
6 nm of shortening in I.1Ims,i, within each cross-bridge there is an elastic element in series with this
non-linear element,j, the filaments themselves are so stiff that their compliance is near to
being negligible, andk, the non-linear element in each cross-bridge makes its movement in a
small number of steps.5. None of these points should be regarded at the present day as being
established beyond doubt, and many important matters are quite un-known, e.g. the structural identification of the elements postulated frommechanical observations, and the nature of the bonds between actin andmyosin.
Of the observations and ideas discussed in this review, those which originatedin my laboratory owe much to a long series of collaborators, whose names appearin the References.
I also wish to thank Dr A. M. Gordon for letting me reproduce the photo-graphs which appear as PI. 1, and my present co-workers Dr R. M. Simmons andDr L. E. Ford for letting me include some unpublished records from our latest seriesof experiments.
I gratefully acknowledge permission from the Central Office of Information,London. to reproduce the frontispiece.
35

A. F. HUXLEY
REFERENCES
ABBOTT, B. C. & AuIBERT, X. M. (1951). Changes of energy in a muscle during veryslow stretches. Proc. R. Soc. B 139, 104-117.
ABBOTT, B. C., AUBERT, X. M. & HILL, A. V. (1951). The absorption of work by amuscle stretched during a single twitch or a short tetanus. Proc. R. Soc. B 139,86-104.
ADRIAN, E. D. (1922). The relation between the stimulus and the electric responsein a single muscle fibre. Archs ngerl. Physiol. 7, 330-332.
ADRIAN, R. H. & PEACHEY, L. D. (1973). Reconstruction of the action potential offrog sartorius muscle. J. Physiol. 235, 103-131.
ARMSTRONG, C. M., HUXLEY, A. F. & JULIAN, F. J. (1966). Oscillatory responses infrog skeletal muscle fibres. J. Physiol. 186, 26-27P.
ARRONET, N. I. (1973). Motile Muscle and Cell Models (translated from Russian byB. Haigh). New York and London: Consultants Bureau.
ASAKURA, S., TANIGUCHI, M. & OOSAWA, F. (1963). Mechano-chemical behaviourof F-actin. J. molec. Biol. 7, 55-69.
ASHLEY, C. C. & RIDOWAY, E. B. (1970). On the relationships between membranepotential, calcium transient and tension in single barnacle muscle fibres. J. P. hysiol.209, 105-130.
ASTBURY, W. T. (1947). On the structure of biological fibres and the problem ofmuscle. Proc. R. Soc. B 134, 303-328.
ATWOOD, H. L., HOYLE, G. & SMYTH, T. Jr. (1965). Mechanical and electricalresponses of single innervated crab-muscle fibres. J. Physiol. 180, 449-482.
AUBER, J. & COUTEAUX, R. (1963). Ultrastructure de la strie Z dans des musclesde dipteres. J. Microscopie 2, 309-324.
AUBERT, X. (1944a). La chaleur degagee par un muscle soumis A un travail en cycle.C. r. Sganc. Soc. Biol. 138, 1011-1013.
AUBERT, X. (1944b). L'influence du delai de relachement sur la chaleur d6gag6epar un muscle soumis a un travail en cycle. C. r. Sganc. Soc. Biol. 138, 1048-1050.
AUBERT, X. (1948). Reversibilite partielle de la contraction musculaire au cours del'absorption du travail en cycle. Archs int. Phy8iol. 55, 348-361.
AUBERT, X. & LEBACQ, J. (1971). The heat of shortening during the plateau oftetanic contraction and at the end of relaxation. J. Physiol. 216, 181-200.
BAGSHAW, C. R., ECCLESTON, J. F., TRENTHAM, D. R., YATES, D. W. & GOODY, R. S.(1973). Transient kinetic studies of the Mg++-dependent ATPase of myosin andits proteolytic subfragments. Cold Spring Harb. Symp. quant. Biol. 37, 127-135.
BAILEY, K. (1948). Tropomyosin: a new asymmetric protein component of themuscle fibril. Biochem. J. 43, 271-279.
BARANY, E. H., EDMAN, K. A. P. & PALIS, A. (1952). The influence of electrolyteson the rate of viscosity drop in ATP-actomyosin mixtures. Acta physiol. scand.24, 361-367.
BLANCo, T., KAREMAKER, J. M. & KRAMER, A. E. J. L. (1972). Elasticity as anexpression of cross-bridge activity in rat muscle. Pflugers Arch. 336, 277-288.
BOTTS, J., COOKE, R., DOS REMEDIOS, C., DUKE, J., MENDELSON, R., MORALES,M. F., TOKIWA, T. & VINIEGRA, G. (1973). Does a myosin cross-bridge progressa-m-over-arm on the actin filament? Cold Spring Harb. Symp. quant. Biol. 37,195-200.
BROWN, L. M., GONZALEZ-SERRATOS, H. & HUXLEY, A. F. (1970). Electron micro-scopy of frog muscle fibres in extreme passive shortening. J. Physiol. 208, 86-88P.
BUCHTHAL, F. & KAISER, E. (1951). The rheology of the cross striated muscle fibre(pp. 233-291). Biol. meadr 21, no. 7.
36

MUSCULAR CONTRACTIONCAIN, D. F., INFANTE, A. A. & DAVIES, R. E. (1962). Chemistry of muscle contrac-
tion. Adenosine triphosphate and phosphorylcreatine as energy supplies for singlecontractions of working muscle. Nature, Lond. 196, 214-217.
CARLSEN, F., KNAPPEIS, G. G. & BUCHTHAL, F. (1961). Ultrastructure of the restingand contracted striated muscle fiber at different degrees of stretch. J. biophy8.biochem. Cytol. 11, 95-117.
CARLSON, F. D., BONNER, R. & FRASER, A. (1973). Intensity fluctuation auto.correlation studies of resting and contracting frog sartorius muscle. Cold SpringHarb. Symp. quant. Biol. 37, 389-396.
CASELLA, C. (1950). Tensile force in total striated muscle, isolated fibre and sarco-lemma. Acta physiol. 8cand. 21, 380-401.
CHAPLAIN, R. A. & FROMMELT, B. (1971). A mechanochemical model for muscularcontraction. I. The rate of energy liberation at steady state velocities of shorteningand lengthening. J. Mechanochem. Cell Motility 1, 41-56.
CIVAN, M. M. & PODOLSKY, R. J. (1966). Contraction kinetics of striated musclefibres following quick changes in load. J. Physiol. 184, 511-534.
COSTANTIN, L. L. (1970). The role of sodium current in the radial spread of con-traction in frog muscle fibers. J. gen. Physiol. 55, 703-715.
COSTANTIN, L. L. & TAYLOR, S. R. (1971). Active and passive shortening in voltage-clamped frog muscle fibres. J. Physiol. 218, 13-15P.
CURTIN, N. A. & DAVIES, R. E. (1973). Chemical and mechanical changes duringstretching of activated frog skeletal muscle. Cold Spring Harb. Symp. quant. Biol.37, 619-626.
DAINTY, M., KLEINZELLER, A., LAWRENCE, A. S. C., MIALL, M., NEEDHAM, J.,NEEDHAM, D. M. & SHEN, S.-C. (1944). Studies on the anomalous viscosity andflow-birefringence of protein solutions. III. Changes in these properties of myosinsolutions in relation to adenosinetriphosphate and muscular contraction. J. gen.Physiol. 27, 355-399.
DAVIES, R. E. (1963). A molecular theory of muscle contraction: calcium-dependentcontractions with hydrogen bond formation plus ATP-dependent extensions ofpart of the myosin-actin cross-bridges. Nature, Lond. 199, 1068-1074.
DESHCHEREVSKII, V. J. (1968). Two models of muscular contraction. Biophysics 13(1969), 1093-1101.
DICKINSON, V. A. & WOLEDGE, R. C. (1973). The thermal effects of shortening intetanic contractions of frog muscle. J. Physiol. 233, 659-671.
EBASHI, S. (1963). Third component participating in the superprecipitation of'natural actomyosin'. Nature, Lond. 200, 1010.
EBASHI, S. & ENDO, M. (1968). Calcium ion and muscle contraction. Prog. Biophys.molec. Biol. 18, 123-183.
EBASHI, S., ENDO, M. & OHTSUKI, I. (1969). Control of muscle contraction. Q. Rev.Biophys. 2, 351-384.
EBASHI, S. & KODAMA, A. (1965). A new protein factor promoting aggregation oftropomyosin. J. Biochem. 58, 107-108.
ELLIOTT, G. F., RoME, E. M. & SPENCER, M. (1970). A type of contraction hypothesisapplicable to all muscles. Nature, Lond. 226, 417-420.
ENDO, M. (1964). Entry of a dye into the sarcotubular system of muscle. Nat'gre,Lond. 202, 1115-1116.
ENDO, M. (1966). Entry of fluorescent dyes into the sarcotubular system of the frogmuscle. J. Physiol. 185, 224-238.
ENGELHARDT, V. A. & LyuBIMOVA, M. N. (1939). Myosine and adenosinetriphos-phatase. Nature, Lond. 144, 668-669.
37

ENGELHARDT, V. A., LYUBIMOVA, M. N. & MEITINA, R. A. (1941). Chemistry andmechanics of the muscle studied on myosin threads. Dokl. Akad. Nauk SSSR 30,644.
FENN, W. 0. (1923). A quantitative comparison between the energy liberated andthe work performed by the isolated sartorius muscle of the frog. J. Phy8iol. 58,175-203.
FENN, W. 0. (1924). The relation between the work performed and the energyliberated in muscular contraction. J. Physiol. 58, 373-395.
FENN, W. 0. & MARSH, B. S. (1935). Muscular force at different speeds of shortening.J. Phyaiol. 85, 277-297.
FIELDS, R. W. & FABER, J. J. (1970). Biophysical analysis of the mechanicalproperties of the sarcolemma. Can. J. Phy8iol. Pharmacol. 48, 394-404.
FORD, L. E., HUXLEY, A. F. & SIMMONS, R. M. (1974). Mechanism of early tensionrecovery after a quick release in tetanized muscle fibres. J. Phy8iol. 240, 42-43P.
FRANZINI-ARMSTRONG, C. & PORTER, K. R. (1964). Sarcolemmal invaginationsconstituting the T system in fish muscle fibers. J. Cell Biol. 22, 675-696.
GASsER, H. S. & HILL, A. V. (1924). The dynamics of muscular contraction. Proc.R. Soc. B 96, 398-437.
GELFAN, S. (1931). Studies of single muscle fibres. III. Further evidence of gradedresponses in single fibres. Am. J. Phy8iol. 96, 16-20.
GELFAN, S. (1933). The submaximal responses of the single muscle fibre. J. Phy8iol.80, 285-295.
GILBERT, C., KRETZSCHMAR, K. M., WILKIE, D. R. & WOLEDGE, R. C. (1971).Chemical change and energy output during muscular contraction. J. Physiol.218, 163-193.
GONZkLEZ-SERRATOS, H. (1971). Inward spread of activation in vertebrate musclefibres. J. Physiol. 212, 777-799.
GORDON, A. M., HUXLEY, A. F. & JULIAN, F. J. (1966a). Tension development inhighly stretched vertebrate muscle fibres. J. Physiol. 184, 143-169.
GORDON, A. M., HUXivY, A. F. & JULIAN, F. J. (1966b). The variation in isometrictension with sarcomere length in vertebrate muscle fibres. J. Phy8iol. 184, 170-192.
HALL, C. E., JAKus, M. A. & SCHMITT, F. 0. (1946). An investigation of crossstriations and myosin filaments in muscle. Biol. Bull. mar. biol. Lab. Wood8 Hole90, 32-50.
HANSON, J. (1968). Recent X-ray diffraction studies of muscle. Q. Rev. Biophy&. 1,177-216.
HANSON, J. & HUXLEY, H. E. (1953). Structural basis of the cross-striations inmuscle. Nature, Lond. 172, 530-532.
HANSON, J. & HUXLEY, H. E. (1955). The structural basis of contraction in striatedmuscle. Symp. Soc. exp. Biol. 9, 228--264.
HARMAN, J. W. (1954). Contractions of skeletal muscle myofibrils by phase micro-scopy. Fedn Proc. 13, 430.
HASSELBACH, W. (1953). Elektronenmikroskopische Untersuchungen an Muskel-fibrillen bei totaler und partieller Extraktion des L-Myosins. Z. Naturf. 8b,449-454.
HILL, A. V. (193S). The heat of shortening and the dynamic constants of muscle.Proc. R. Soc. B 126, 136-195.
HILL, A. V. (1949 a). The heat of activation and the heat of shortening in a muscle.twitch. Proc. R. Soc. B 136, 195-211.
HILL, A. V. (1949 b). Work and heat in a muscle twitch. Proc. R. Soc. B 136, 220-228.HILL, A. V. (1964). The effect of load on the heat of shortening of muscle. Proc. R.
Soc. B 159, 297-318.
A. F. HUXLEY38

MUSCULAR CONTRACTION 39HILL, A. V. (1965). Trails and Trials in Physiology. London: Arnold.HruL, A. V. & HOWARTH, J. V. (1959). The reversal of chemical reactions in con-
tracting muscle during an applied stretch. Proc. R. Soc. B 151, 169-193.HILL, D. K. (1964). The space accessible to albumin within the striated muscle
fibre of the toad. J. Physiol. 175, 275-294.HODGKIN, A. L. (1951). The ionic basis of electrical activity in nerve and muscle.
Biol. Rev. 26, 339-409.HOYLE, G. (1968). In Symposium on Mwucle, ed. ERNST, E. & STRAUB, F. B.,
pp. 34-48. Budapest: Akad6mia-i Kiad6.HUXLEY, A. F. (1952). Applications of an interference microscope. J. Physiol. 117,
52-53P.HUXLEY, A. F. (1954). A high-power interference microscope. J. Physiol. 125,
11-13P.HUXLEY, A. F. (1957). Muscle structure and theories of contraction. Prog. Biophys.
biophys. Chem. 7, 255-318.HUXLEY, A. F. (1973). A note suggesting that the cross-bridge attachment during
muscle contraction may take place in two stages. Proc. R. Soc. B 183, 83-86.HUXLEY, A. F. & GORDON, A. M. (1962). Striation patterns in active and passive
shortening of muscle. Nature, Lond. 193, 280-281.HUXLEY, A. F. & JuLIAN, F. J. (1964). Speed of unloaded shortening in frog striated-
muscle fibres. J. Physiol. 177, 60-61 P.HUXLEY, A. F. & NIEDERGERKE, R. (1954). Interference microscopy of living
muscle fibres. Nature, Lond. 173, 971-973.HUXLEY, A. F. & NIEDERGERKE, R. (1958). Measurement of the striations of isolated
muscle fibres with the interference microscope. J. Physiol. 144, 403-425.HUXLEY, A. F. & PEACHEY, L. D. (1961). The maximum length for contraction in
vertebrate striated muscle. J. Physiol. 156, 150-165.HUXLEY, A. F. & SImMONs, R. M. (1970). A quick phase in the series-elastic com-
ponent of striated muscle, demonstrated in isolated fibres from the frog. J. Physiol.208, 52-53P.
HUXLEY, A. F. & SIMMONs, R. M. (1971a). Mechanical properties ofthe cross-bridgesof frog striated muscle. J. Physiol. 218, 59-60P.
HUXLEY, A. F. & SIMMONs, R. M. (1971 b). Proposed mechanism of force generationin striated muscle. Nature, Lond. 233, 533-538.
HUXLEY, A. F. & SIMMONs, R. M. (1973). Mechanical transients and the origin ofmuscular force. Cold Spring Harb. Symp. quant. Biol. 37, 669-680.
HUXLEY, A. F. & STRAUB, R. W. (1958). Local activation and interfibrillar structuresin striated muscle. J. Physiol. 143, 40-41P.
HUXLEY, A. F. & TAYLOR, R. E. (1955). Function of Krause's membrane. Nature,Lond. 176, 1068.
HUXLEY, A. F. & TAYLOR, Et. E. (1958). Local activation of striated muscle fibres.J. Physiol. 144, 426-441.
HUXLEY, H. E. (1953a). X-ray analysis and the problem of muscle. Proc. R. Soc.B 141, 59-62.
HUXLEY, H. E. (1953b). Electron microscope studies of the organisation of thefilaments in striated muscle. Biochim. biophys. Acta 12, 387-394.
HUXLEY, H. E. (1957). The double array of filaments in cross-striated muscle.J. biophys. biochem. Cytol. 3, 631-648.
HUXLEY, H. E. (1960). Muscle Cells. In The Cell, vol. iv, ed. BRACHET, J. & MIRSKY,A. E., pp. 365-481. New York: Academic Press.
HUXLEY, H. E. (1963). Electron microscope studies on the structure of naturaland synthetic protein filaments from striated muscle. J. molec. Biol. 7, 281-308.

40 A. F. HUXLEYHUXLEY, H. E. (1964a). Evidence for continuity between the central elements of
the triads and extracellular space in frog sartorius muscle. Nature, Lond. 202,1067-1071.
HUXLEY, H. E. (1964b). Structural arrangements and the contraction mechanism instriated muscle. Proc. R. Soc. B 160, 442-448.
HUXLEY, H. E. (1969). The mechanism of muscular contraction. Science, N.Y. 164,1356-1366.
HUXLEY, H. E. (1971). The structural basis of muscular contraction. Proc. R. Soc.B 178, 131-149.
HUXLEY, H. E. & BROWN, W. (1967). The low-angle X-ray diagram of vertebratestriated muscle and its behaviour during contraction and rigor. J. molec. Biol. 30,383-434.
HUXLEY, H. E. & HANSON, J. (1954). Changes in the cross-striations of muscleduring contraction and stretch and their structural interpretation. Nature, Lond.173, 973-976.
HUXLEY, H. E. & HANSON, J. (1957). Quantitative studies on the structure ofcross-striated myofibrils. I. Investigations by interference microscopy. Biochim.biophy8. Acta 23, 229-249.
JASPER, H. H. & PEZARD, A. (1934). Relation entre la rapidity d'un musclestrike et sa structure histologique. C. r. hebd. SNanc. Acad. Sci., Paria 198, 499-501.
JOBSIS, F. F. & O'CONNOR, M. J. (1966). Calcium release and reabsorption in thesartorius muscle of the toad. Biochem. biophys. Re8. Commun. 25, 246-252.
JORDAN, H. E. (1934). Structural changes during contraction in the striped muscleof the frog. Am. J. Anat. 55, 117-133.
JULIAN, F. J., SOLLINS, K. R. & SOLLINS, M. R. (1973). A model for musclecontraction in which cross-bridge attachment and force generation are distinct.Cold Spring Harb. Symp. quant. Biol. 37, 685-688.
KATZ, B. (1939). The relation between force and speed in muscular contraction.J. Physiol. 96, 45-64.
KUFFLER, S. W. (1946). The relation of electric potential changes to contracture inskeletal muscle. J. Neurophysiol. 9, 367-377.
KtHNE, W. (1864). Untersuchungen uiber das Protoplama und die Contractilitdt.Leipzig: Engelmann.
KUSHMERICK, M. J., LARSON, R. E. & DAVIES, R. E. (1969). The chemical energeticof muscle contraction. I. Activation heat, heat of shortening and ATP utilizationfor activation-relaxation processes. Proc. R. Soc. B 174, 293-313.
LEVIN, A. & WYMAN, J. (1927). The viscous elastic properties of muscle. Proc. R.Soc. B 101, 218-243.
LYMN, R. W. & TAYLOR, E. W. (1971). Mechanism of adenosine triphosphatehydrolysis by actomyosin. Biochemistry 10, 4617-4624.
MCCLARE, C. W. F. (1971). Chemical machines, Maxwell's demon and livingorganisms. J. theor. Biol. 30, 1-34.
MCCLARE, C. W. F. (1972). A 'molecular energy' muscle model. J. theor. Biol. 35,569-595.
MONEILL, P. A. & HOYLE, G. (1967). Evidence for superthin filaments. Am. Zool. 7,483-498.
MATSUBARA, I. & ELLIOTT, G. F. (1972). X-ray diffraction studies on skinned singlefibres of frog skeletal muscle. J. molec. Biol. 72, 657-669.
MENCKEN, H. L. (1936). The American Language, 4th edn. p. 312. New York:A. A. Knopf.
MEYER, K. H. (1929). 'Ober Feinbau, Festigkeit und Kontraktilitat tierischerGewebe. Biochem. Z. 214, 253-281.

MUSCULAR CONTRACTION 41
MOMMAERTS, W. F. H. M. (1951). The scattering of light in myosin solutions.I. The angular dissymmetry and the molecular length. J. biol. Chem. 188,553-557.
MOMMAERTS, W. F. H. M. (1956). The effect of adenosinetriphosphate upon acto-myosin solutions, studied with a recording dual beam light-scattering photometer.J. gen. Physiol. 39, 821-830.
Moos, C. (1973). Actin activation ofheavy meromyosin and subfragment-1 ATPases;steady state kinetics studies. Cold Spring Harb. Symp. quant. Biol. 37, 137-143.
MORALES, M. & BOTTS, J. (1952). A model for the elementary process in muscleaction. Archs Biochem. Biophys. 37, 283-300.
NEEDHAM, D. M. (1950). Myosin and adenosinetriphosphate in relation to musclecontraction. Biochim. biophys. Acta 4, 42-49.
NEEDHAM, D. M. (1971). Machina Carnis. The Biochemistry of Muscular Contractionin its Historical Development. Cambridge: University Press.
NEEDHAM, J., SHEN, S.-C., NEEDHAM, D. M. & LAWRENCE, A. S. C. (1941). Myosinbirefringence and adenylpyrophosphate. Nature, Lond. 147, 766-768.
OPLATKA, A. (1972). On the mechanochemistry of muscular contraction. J. theor.Biol. 34, 379-403.
PAGE, S. (1964). The organization of the sarcoplasmic reticulum in frog muscle.J. Physiol. 175, 10-1IP.
PAGE, S. G. (1968). Fine structure of tortoise skeletal muscle. J. Physiol. 197,709-715.
PAGE, S. G. (1969). Structure and some contractile properties of fast and slowmuscles of the chicken. J. Physiol. 205, 131-145.
PAGE, S. G. & HUXLEY, H. E. (1963). Filament lengths in striated muscle. J. CellBiol. 19, 369-390.
PODOLSiY, R. J. (1959). The chemical thermodynamics and molecular mechanismof muscular contraction. Ann. N.Y. Acad. Sci. 72 (12), 522-537.
PODOLSxY, R. J. (1960). Kinetics of muscular contraction: the approach to thesteady state. Nature, Lond. 188, 666-668.
PODOLSKY, R. J. (1964). The maximum sarcomere length for contraction of isolatedmyofibrils. J. Physiol. 170, 110-123.
PODOLSKY, R. J. & NOLAN, A. C. (1973). Muscle contraction transients, cross-bridgekinetics, and the Fenn effect. Cold Spring Harb. Symp. quant. Biol. 37, 661-668.
PODOLSKY, R. J., NOLAN, A. C. & ZAVELER, S. A. (1969). Cross-bridge propertiesderived from muscle isotonic velocity transients. Proc. natn. Acad. Sci. U.S.A.64, 504-511.
PoLissAR, M. J. (1952). Physical chemistry of contractile process in muscle. I-IV.Am. J. Physiol. 168, 766-811.
PORTZEHL, H. (1950). Masse und Masze des L-Myosins. Z. Naturf. 5b, 75-78.PRINGLE, J. W. S. (1949). The excitation and contraction of the flight muscles of
insects. J. Physiol. 108, 226-232.PRiNGLE, J. W. S. (1967). The contractile mechanism of insect fibrillar muscle.
Prog. Biophys. molec. Biol. 17, 1-60.PRYOR, M. G. M. (1950). Mechanical properties of fibres and muscles. Prog. Biophys.
biophys. Chem. 1, 216-268.RAMSEY, R. W. & STREET, S. F. (1940). The isometric length-tension diagram of
isolated skeletal muscle fibers of the frog. J. cell. comp. Physiol. 15, 11-34.RAPOPORT, S. I. (1972). Mechanical properties of the sarcolemma and myoplasm in
frog muscle as a function of sarcomere length. J. gen. Physiol. 59, 559-585.RIcE, R. V. (1961). Conformation of individual macromolecular particles frommyosin solutions. Biochim. biophys. Acta 52, 602-604.

42 A. F. HUXLEYRIDGWAY, E. B. & ASHLEY, C. C. (1967). Calcium transients in single muscle fibers.
Biochem. biophys. Re8. Commun. 29, 229-234.ROWE, A. J. (1964). The contractile proteins of skeletal muscle. Proc. R. Soc. B 160,
437-441.RozSA, G., SZENT-GYORGYI, A. & WYCKOFF, R. W. G. (1950). The fine structure of
myofibrils. Expi Cell Re8. 1, 194-205.RUDEL, R. & TAYLOR, S. R. (1971). Striated muscle fibers: facilitation of contraction
at short lengths by caffeine. Science, N.Y. 172, 387-388.RUDEL, R. & TAYLOR, S. R. (1973). Aequorin luminescence during contraction of
amphibian skeletal muscle. J. Phy8iol. 233, 5-6 P.SCHNEIDER, M. F. & CHANDLER, W. K. (1973). Voltage dependent charge movement
in skeletal muscle: a possible step in excitation-contraction coupling. Nature,Lond. 242, 244-246.
SEIDEL, J. C. & GERGELY, J. (1973). Investigation of conformational changes inspin-labeled myosin: implications for the molecular mechanism of musclecontraction. Cold Spring Harb. Symp. quant. Biol. 37, 187-193.
SHIMOMURA, O., JOHNSON, F. H. & SAIGA, Y. (1962). Extraction, purification andproperties of aequorin, a bioluminescent protein from the luminous hydromedusan,Aequorea. J. cell. comp. Phyajol. 59, 223-239.
SJOSTRAND, F. S. (1962). The connections between A- and I-band filaments instriated frog muscle. J. Ultrastruc. Res. 7, 225-246.
SPENCER, M. & WORTHINGTON, C. R. (1960). A hypothesis of contraction in striatedmuscle. Nature, Lond. 187, 388-391.
STEIGER, G. J., RUEGG, J. C., BOLDT, K. M., LUBBERS, D. W. & BREULL, W. (1973).Changes in the polarization of tryptophan fluorescence in the actomyosinsystem of working muscle fibers. Cold Spring Harb. Symp. quant. Biol. 37,377-378.
STRAUB, F. B. (1943). Actin. Stud. Inst. med. Chem. Univ. Szeged (1942) 2, 3-15.STRICKHOLM, A. (1962). Excitation currents and impedence of a small electrically
isolated area of the muscle cell surface. J. cell. comp. Physiol. 60, 149-167.STRICKHOLM, A. (1966). Local sarcomere contraction in fast muscle fibres. Nature,
Lond. 212, 835-836.SUGI, H. & OCHI, R. (1967). The mode of transverse spread of contraction initiatedby local activation in single frog muscle fibers. J. gen. Physiol. 50, 2167-2176.
SZENT-GYORGYI, A. (1943). Discussion. Stud. Int. med. Chem. Univ. Szeged (1941-2)1, 67-71.
TAYLOR, E. W. (1972). Chemistry of muscle contraction. A. Rev. Biochem. 41,577-616.
TAYLOR, S. R. & RUDEL, R. (1970). Striated muscle fibers: inactivation ofcontractioninduced by shortening. Science, N.Y. 167, 882-884.
TONOMURA, Y. (1972). Muscle Proteins, Muscle Contraction and Cation Transport.Tokyo: University of Tokyo Press.
TONOMURA, Y. & WATANABE, S. (1952). Effect of adenosine triphosphate on thelight-scattering of actomyosin solution. Nature, Lond. 169, 112-113.
TONOMURA, Y., YAGI, K., Ku-Bo, S. & KITAGAWA, S. (1961). A molecular mechanismof muscle contraction. J. Res. Inst. Catalysis, Hokkaido Univ. 9, 256-286.
ULLRICK, W. C. (1967). A theory of contraction for striated muscle. J. theor. Biol.15, 53-69.
WALCOTT, B. & RIDGWAY, E. B. (1967). The ultrastructure of myosin-extractedstriated muscle fibers. Am. Zool. 7, 499-504.
WEBER, H. H. (1956). In Verhandlungen der Deutachen orthopadiachen Gesellachaft,44th Congress, pp. 13-27. Stuttgart: Enke.
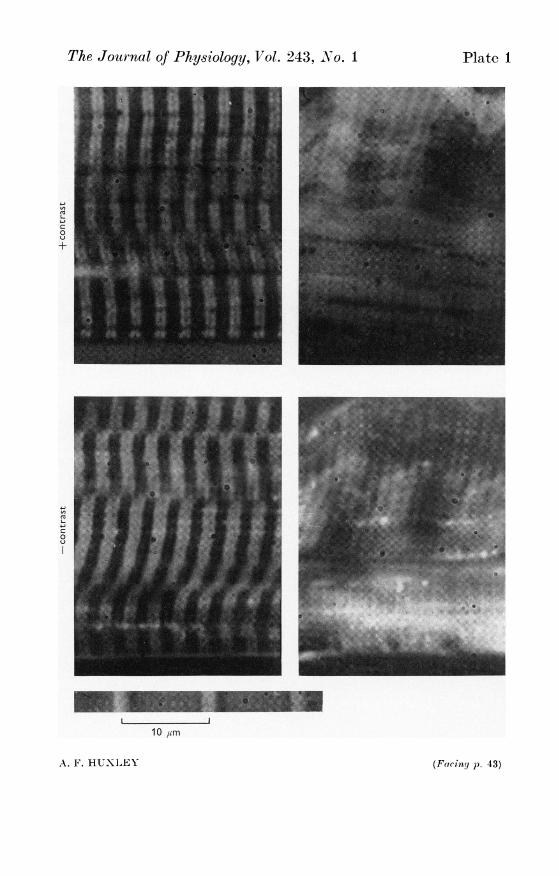
The Journal of Physiology, Vol. 243, NS o. 1 Plate 1
+
0
u
IA
_,-_i.
--Snat110 Jim
A. F. HUXLEY (Facing p. 43)

MUSCULAR CONTRACTIONWEBER, H. H. (1958). The Motility of Mu8cle and CelI, pp. 33-36. Cambridge,
Mass.: Harvard University Press.WILKIE, D. R. (1968). Heat work and phosphorylcreatine break-down in muscle.
J. Physiol. 195, 157-183.WOHLISCH, E. (1940). Muskelphysiologie vom Standpunkt der kinetischen Theorie
der Hochelastizitat und der Entspannungshypothese des Kontr ktionsmechanis-mus. Naturwuien9chaften 28, 305-312 and 326-335.
WOLEDGE, R. C. (1968). The energetics of tortoise muscle. J. Phy8iol. 197, 685-707.Yu, L. C., DOWBEN, R. M. & KORNACKER, K. (1970). The molecular mechanism
of force generation in striated muscle. Proc. natn. Acad. Sci. U.S.A. 66, 1199-1205.ZOBEL, C. R. & CARLSON, F. D. (1963). An electron microscopic investigation ofmyosin and some of its aggregates. J. molec. Biol. 7, 78-89.
EXPLANATION OF PLATE
Active and passive shortening within a living isolated frog muscle fibre The fibreis stimulated by a slowly increasing current; the cathode is near the edge of thefibre which is seen near the bottom of each micrograph. The fibre does not giveaction potentials in these responses.
The micrographs are single frames from a cine film taken through an interferencemicroscope (A. F. Huxley, 1952, 1954). The upper pair of pictures are taken withthe microscope adjusted so as to give positive contrast (dark indicates high refractiveindex, i.e. A bands, Z lines and contraction bands are dark). For the contractionshown in the lower pair of pictures, the microscope has been readjusted to givenegative. contrast (A bands, Z lines and contraction bands light).
In each pair of pictures, the left-hand one is before, and the right-hand one during,the stimulus. The right-hand pictures show that during the shortening the myofibrilsnear the depolarized surface of the fibre remain straight, and must therefore becontracting actively, while the myofibrils nearer the centre of the fibre (upper partof picture) are thrown into waves and must therefore be shortening passively. Inthese passively shortened fibrils, the resting pattern of dark and light bands persists(the A bands appear somewhat narrowed, but this is due to the obliquity of thefibrils). In the actively contracting fibrils near the edge, however, the pattern haschanged to one with narrow dense lines. These are the C. contraction bands, placedat the centre of the original A bands; on further shortening the Cz contractionbands, at the position of the Z lines, appear as a second set of thin dense linesmidway between the successive C. bands.From a film taken by A. F. Huxley and A. M. Gordon, 3 November 1961; cf.
Huxley & Gordon (1962).
43