in rabbits - ncbi
TRANSCRIPT

Journal of Physiology (1996), 495.1, pp. 159-168
Reversible inactivations of the cerebellum prevent theextinction of conditioned nictitating membrane responses
in rabbits
Narender Ramnani and Christopher H. Yeo
Neuroscience and Behaviour Group, Department of Anatomy and Developmental Biology,University College London, Gower Street, London WC1E 6BT, UK
1. Studies show that reversible inactivation of the anterior interpositus nucleus (AIP) of thecerebellum with muscimol (a GABAA agonist) prevents acquisition of the classicallyconditioned nictitating membrane response (NMR) in the rabbit. Here, we have usedreversible inactivations of the AIP with muscimol to investigate the role of the cerebellum inthe extinction of this response.
2. Experimental subjects were implanted with cannulae targeted to the AIP, through whichmuscimol could be infused via an injector cannula. This experiment was divided into threephases lasting 4 days, separated by 3 day intervals. Experimental and unoperated controlsubjects received acquisition training in phase 1; in phases 2 and 3 they received extinctiontraining.
3. Presentation of the conditioned stimulus (CS) alone in phase 2 produced normal extinction incontrol subjects. Muscimol inactivation of the AIP in experimental subjects during phase 2prevented extinction of conditioned responses (CRs), shown by initial high CR frequency inthe first post-drug session of phase 3, which then extinguished in a mannerindistinguishable from controls in phase 2.
4. Our findings support the suggestion that similar cerebellar circuitry is engaged in acquisitionand extinction ofNMR conditioning.
The neuronal architecture and connectivity of the cerebellarcortex have inspired several models that assign animportant motor learning function to the cerebellum (Marr,1969; Albus, 1971; Ito, 1972; Gilbert, 1974). These modelsare controversial, but recent evidence continues to indicatemotor learning functions in the cerebellum (see Thach,1996, for review).
The classically conditioned nictitating membrane response(NMR) of the rabbit has been used extensively to study theneural mechanisms of associative learning of a simple motorresponse (see Thompson & Krupa, 1994). A tactile orelectrical unconditional stimulus (US) applied to theperiorbital region always elicits an unconditioned reflex(UR) blink of the nictitating membrane. The NMR can beassociatively conditioned by presenting an auditoryconditioned stimulus (CS) that initially does not elicit aresponse, followed by the US. After repeated, pairedpresentations of these stimuli, the CS comes to evoke aclassically conditioned NMR.
Lesions of parts of the olivocerebellar system (includingcerebellar cortical lobule HVI) profoundly impair conditionedresponses (CRs) during acquisition training and retentiontesting (Yeo, Hardiman & Glickstein, 1985b; see Yeo, 1991;Thompson & Krupa 1994). Thus, the neural circuitry
essential for this simple form of associative learning includesthese parts of the cerebellum. Lesions of the cerebellum mayabolish CRs because a site of essential plasticity has beendestroyed. Alternatively, sites which are crucial for CRs andwhich are efferent from the cerebellum may requirecerebellar input to learn and/or express CRs (Welsh &Harvey, 1989). Studies employing permanent lesions cannotdifferentiate between the above possibilities and thereforehave not localized essential neural plasticity for NMRconditioning to the cerebellum.
Reversible lesions can overcome many of the drawbacksassociated with permanent lesions in studies of learning andmemory. If acquisition occurs while the cerebellum isinactivated, CRs will be evident when the blockade is lifted;CRs will not be evident if the cerebellum is essential foracquisition. Such a result does not prove that there isessential plasticity in the cerebellum but, assuming suchplasticity is not extensively distributed, further inactivationsin efferent and afferent pathways can be used to identify aset of candidate structures within which essential plasticitymust be present.
Reversible lesion studies have been used to test for acerebellar role in NMR conditioning. Some have found CRsafter training during cerebellar inactivation (Welsh & Harvey,
5364 159

160 N. Ramnani
1991), but others found no evidence for acquisition inpreviously naive subjects during cerebellar inactivations(Krupa, Thompson & Thompson, 1993; Nordholm,Thompson, Dersarkissian & Thompson, 1993); in addition,reversible inactivation of the brachium conjunctivum doesnot prevent NMR conditioning (Krupa & Thompson, 1995).These authors have concluded that the essential memorytrace for NMR conditioning is within the cerebellum (seeDiscussion).
One problem with these reversible lesion experiments is thatan essential contribution to learning of a particular brainregion is judged by the absence of learned responses afterthe inactivation. Other factors unrelated to learning mayproduce this result. If there are extended drug effects intothe post-inactivation phase and the target area is essentialfor response expression (as is the anterior interpositusnucleus, AIP), then initial failure to produce learnedresponses would relate to such extended drug effects andlearning may have occurred during the inactivation. Thisproblem may be overcome by analysing extinction learning.In NMR conditioning, extinction is observed as the gradualwaning of CRs with repeated presentation of the CS alonein the absence of the reinforcing US after previousacquisition. Several features of extinction learning, includingthe phenomenon of spontaneous recovery (see Mackintosh,1974) indicate that, like acquisition, extinction is an activelearning process. It is highly probable that the neuralcircuitry supporting extinction learning is closely related tocircuitry essential for acquisition learning.
In extinction learning, it is the absence of CRs whichdemonstrates learning, in contrast to acquisition, in whichthe presence of CRs indicates learning. If NMR extinctionlearning can be prevented using reversible inactivations inthe AIP then this will be revealed as the presence of CRs inthe post-inactivation phase. Such a result would rule outextended drug effects and demonstrate that the cerebellumis essential for extinction ofNMR conditioning.
The purpose of the present study is to examine the role ofthe cerebellum in extinction of NMR conditioning. Afteracquisition training, experimental subjects were given aperiod of extinction training during inactivation of the AIPwith muscimol, and then a period of extinction trainingwithout AIP inactivation. Control subjects received the samebehavioural training but without the muscimol blockade.Results are consistent with the suggestion that muscimolinactivation of the AIP prevents extinction of classicallyconditioned nictitating membrane (NM) responses.
METHODSSubjectsThe subjects were fifteen male Dutch belted rabbits weighing1 8-2-5 kg. They were housed individually, allowed food and waterad libitumn and maintained on a 12 h light-dark cycle.
ad C. H. Yeo J Phy8iol.495.1
SurgeryNine rabbits underwent surgery for the implantation of a guidecannula directed towards the right anterior interpositus nucleus(AIP). Each rabbit was anaesthetized using a fentanyl-fluanisonemixture (Hypnorm, Janssen, Oxford, UK; 0-1-5-0 mg kg-', I.M.)with a supplement of benzodiazepam (Valium, Roche, WelwynGarden City, UK; 0'5 mg kg-', I.v.) and was then intubated withan endotracheal tube and the head placed in a stereotaxicinstrument (Kopf, Tujunga, CA, USA), aligned with thestereotaxic co-ordinate system of Matricali (1961). Naloxonehydrochloride (Narcan, Du Pont; 5-7 mg kg-', i.v.) was used toantagonize the fentanyl and anaesthesia was maintainedthroughout the operation using halothane (1.5-2'5%) in a nitrousoxide-oxygen mixture (1: 3). The scalp was reflected and bone anddura removed to expose the right cerebellar cortex. Theapproximate position of the cerebellar nuclei was definedstereotaxically (A - 5 mm AP, A + 4 mm ML, A - 10 mm DV), andtheir exact position was then determined by electrophysiologicalrecording using tungsten microelectrodes. We looked for rapidcellular activity below the deepest cortical layer ventral to therostromedial part of the paramedian lobe. We then searched for aneyeblink-related area in the deep nuclei. Microelectrode tracks weremade until cellular activity that correlated with eyeblinks evokedby gentle tactile stimulation of the cornea periocular area wasrecorded. In order to evoke blinks whilst recording, we maintaineda level of anaesthesia that completely suppressed the toe-pinchresponse but which did not entirely suppress the corneal blinkreflex. A 24 gauge stainless-steel cannula guide was then implanted1 mm caudal and 1 mm dorsal to the position of the activated cells.The placement of the guide tip avoided damage to the connectionsof these cells and the orientation of the guide avoided damage toparts of the overlying cortex that are important for NMRconditioning (Yeo et al. 1985b). Exposed brain was covered withsterile absorbable gelatin foam and the cannula guide was fixed tothe skull with dental cement. The scalp was sutured around thecannula guide and the animal removed from the stereotaxicinstrument. Sodium chloride-glucose (0X18%-4%, 10-20 ml s.c.)was given to supplement fluid and electrolyte levels. Postoperativeanalgesia was provided by buprenorphine (Temgesic, Reckitt &Colman, Hull, UK; 50 ,ug kg-') each day for 3 days and antibioticcover was by chloramphenicol (chloromycetin succinate, FormitaliaCarlo Erba Ltd, Milton Keynes, UK; 7.5 mg kg-') twice per dayfor 3 days. All surgical subjects were maintained in a quiet recoveryroom separate from the main colony for these 3 days and theirbehaviour was closely monitored. All cannulated rabbits resumednormal feeding, drinking and grooming within 12 h and theyexhibited no obvious motor deficits. All subjects were allowed2 weeks for recovery.
Preparations for conditioningAll cannula-implanted and non-operated control rabbits were
prepared for measurement of nictitating membrane movements.Each subject was placed in a Perspex restraining stock and a rubberfoam-padded end-stop was positioned to hold each subject firmly.The ears were gently held between rubber foam pads to restrainhead movements. Two to three drops of ophthalmic anaesthetic(proxymetacaine hydrochloride; 0'5% w/v; Ophthaine, Squibb,Dublin, Ireland) were administered to the right eye so as to wet thesurface of the nictitating membrane and the sclera. After 5 min, a
sterile, fine monofilament suture (Ethicon, Prolene, 5/0) was passedthrough the superficial soft tissue on the lateral edge of the rightnictitating membrane and tied to form a very small loop (diameter,
a7i

Cerebellar inactivations prevent extinction
1P5 mm). This loop afforded a simple, atraumatic coupling for thenictitating membrane movement transducer (see below) and so it wasleft in place throughlout the experiment. Topical chloramphenicoleye ointment (1% w/w; Daniels Pharmaceuticals, Derby, UK) wasapplied to the right eye of each subject on each day throughout theexperiment and no subject developed infection or irritation of theeye or periocular areas. The subjects were then returned to theirhome cages where they resumed normal feeding and groomingbehaviours. We saw no evidence of discomfort caused by the sutureloop.
At the beginning of the adaptation session (see belo-w), twostandard 12 mm stainless-steel Michel clips served to deliverstimulation to the facial skin surrounding the eyelids. One wasplaced approximately 5 mm behind the lateral canthus of the eyeand the other was placed approximately 5 mm below the centre ofthe lower eyelid. The 12 mm clips were gently attached to thesuperficial skin layer using a standard applicator without the needto remove underlying fur. Again, when the subjects were returnedto their home cages at the end of the adaptation session, the rabbitsmade no attempts to remove the clips and we sawN no other evidenceof discomfort.
Conditioning apparatus and stimuliThe rabbits were trained using techniques similar to those firstdeveloped by Gormezano, Schneiderman, Deux & Fuentes (1962)and described in an earlier paper (Yeo & Hardiman, 1992). At thebeginning of each daily behavioural testing session, each subjectwas placed in the Perspex restraining stock (see above for details).Subjects adapted well to this restraint and remained passive duringthe behavioural testing sessions of up to 1 h duration. A lowt-torquepotentiometer mounted upon a simple alloy cradle was positionedabove the dorsal aspect of the head. Posteriorly, the cradle includeda smooth, curved alloy ring which passed around the base of theears and anteriorly, smooth alloy extensions stabilized the cradle oneach side of the muzzle. This arrangement allowed the body of thepotentiometer to be atraumatically fixed relative to the head. Theshaft of the potentiometer was then coupled to the nictitatingmembrane, via a lever and miniature universal joint, with a smallstainless-steel hook that w\as passed through the suture loop(Gormezano & Gibbs 1988). This coupling of the potentiometer andsuture allowed isotonic transduction of NM movement andeliminated the need for a restoring force on the transducer. Thisisotonic transduction is directly comparable with that used inrecent studies by Welsh & Harvey (1989, 1991) and with those fromour own laboratory (Gruart & Yeo, 1995).
Each subject Nwas then placed in a ventilated, sound-attenuatingchamber facing a loudspeaker mounted centrally. The conditionedstimulus (CS) wNas a 1 kHz sine wave tone of 310 ms duration andan intensity of 63 dB (A scale) (reference 20 ,uN m-2). Backgroundnoise produced by-ventilation fans was 54 dB (reference 20,uN m-2).The unconditioned stimulus (US) was periorbital electricalstimulation. Each US was a 60 ms train of three biphasic pulses ofintensity 2-5 mA applied to the periorbital region of the facethrough the stainless-steel clips described above. On paired trialsthe interstimulus interval between the CS and US onset wAas250 ms. The intertrial interval was randomly selected between 25and 35 s. This US level was sufficient to evoke a clear blink of theexternal eyelids and a closure of the nictitating membrane that didnot habituate with repeated presentations. On initial stimulation,small movements of the neck and limb muscles could sometimes beseen but these habituated rapidly over a fewr trials.
Behavioural trainingSession types. There were three different types of sessions.
(i) Adaptation. Subjects were placed in the restraining stock, fittedw-ith the nictitating membrane transducer cradle and the Michelclips (see above) and placed in the conditioning chamber for 1 h -equivalent to the duration of one conditioning session. There wTereno presentations of either the CS or the US during this period.
(ii) Acquisition training. Subjects were placed in the restrainingstock, fitted with the nictitating membrane transducer cradle andplaced in the conditioning chamber. Each session consisted of 100trials. In ninety trials the CS and US w,ere paired and in ten trialsthe CS was presented alone. A CS alone was presented in everytenth trial.
(iii) Extinction training. Subjects were placed in the restrainingstock, fitted with the nictitating membrane transducer cradle andplaced in the conditioning chamber. Each session consisted of 100trials. The CS was presented alone in every trial.
Experimental design. Two weeks after surgery, the monofilamentloop was sutured in the right nictitating membrane and 3 days laterall subjects underwent a session of adaptation. On the follo-wingday, they entered the experiment in phase 1. There were threephases, and 3 days between each phase.
(i) Phase 1. All subjects received four daily sessions of acquisition.
(ii) Phase 2. All subjects received four daily sessions of extinctiontraining. Experimental subjects received muscimol infusion prior toeach session; 1-54 nmol muscimol in 1 jul solution (muscimolhydrobromide; RBI; in 50 mm phosphate-buffered saline, pH 7 4)was injected over 1 min into the right cerebellum of eachcannulated subject. The injection -was via a 36 gauge cannulainserted through the 24 gauge implanted guide cannula andprotruding 05 mm below the tip of the guide. The injection was,therefore, 05 mm above cells with activity related to eyeblinks.The cannulated subjects were then left for 1 h in their home cagebefore training.
(iii) Phase 3. All subjects received four daily sessions of extinctiontraining. If experimental subjects had extinguished in phase 2,they would show the same frequency of CRs as control subjects inphase 3. However, if muscimol infusions prevented extinction, thenthe subjects would extinguish at the same rate as controlsextinguishing for the first time (control subjects in phase 2).
Subject groups. Nine subjects underwent surgery in which a guidecannula was implanted above the right cerebellar nuclei. Fivesubjects in the cannulated group had very low frequencies of CRsduring the first extinction session with muscimol blockade in phase2 (see Results) and so were assigned to the 'effective' group. Theremaining four animals had much higher CR frequencies duringthis session (see Results) and were assigned to the 'ineffective'group (in = 4). The control group consisted of six subjects.
HistologyAt the end of the experiment rabbits -with implanted cannulae wvereinjected with heparin sodium (500 units kg-', i.v.) and an overdoseof pentobarbitone sodium (90 mg kg-', i.v.). Each rabbit wN-asperfused through the aorta with 09% saline followed by 4%formaldehyde and the brain was removed from the skull. The brainwas then embedded in 10% gelatin and placed in a solution of 20%sucrose formalin for 3 days. Frozen sections, 60 /tm thick, were cutin the transverse plane. Alternate sections were mounted ontogelatinized slides and stained for Nissl substance with Cresyl Violet.
J Phy8iol.495.1 161

N Ramnani and C. H. Yeo
Data analysisConditioned responses. A conditioned response (CR) was definedas a nictitating membrane response with amplitude greater than orequal to 0 5 mm and with onset latency greater than 35 ms fromCS onset (Yeo, Hardiman & Glickstein, 1985a). All responsesoccurring from CS onset to 1000 ms after CS onset were recorded.Responses on paired and unpaired trials occurring between 35 and250 ms from CS onset were defined as CRs. The frequency ofresponses whose latency to onset occurred between 250 and1000 ms in the extinction phases was calculated for each group, inorder to determine whether response onset latency changed duringextinction training. These were expressed as a function of thenumber of trials presented. In order to show within-session effects,data were plotted graphically in ten trial blocks (9 paired trials + 1CS-alone trial for phase 1, and 10 unpaired trials for phases 2 and3). Where necessary, within-session frequency graphs weresuperimposed on block frequency graphs. Data from the paired andunpaired trials were treated similarly, i.e. occurrence of CRs couldonly be determined within the CS-US interval. We applied thesame analysis to extinction learning trials which, althoughconsisting entirely of CS-alone presentations, were analysed for CRfrequency during the same time window. Statistical analysis wasperformed on session and block CR frequency.
Statistical analysis. A priori comparisons of CR frequencies ofgroups across blocks and sessions was achieved using three-wayanalysis of variance (ANOVA). In addition, a two-way ANOVAand the Student-Newman-Keuls test were applied for post hocanalysis where required.
RESULTSThe principal findings of this study were that muscimolinfusions into the anterior interpositus nucleus (AIP)completely prevented the extinction of conditioned responses.
Histological verification and group assignmentThere were nine cannulated subjects. Five cannulatedsubjects (El-E5) were included in the effective group. Theyshowed very few CRs during the first extinction session ofphase 2 (range, 0-2-5 %). Cannula placements of thesesubjects were located between approximately 0 5 and1-5 mm from anterior parts of the interpositus nucleus (seeFigs 1 and 2). In four of these subjects (E1-E4), the guidecannulae traversed white matter, slightly dorsal to or justbelow the surface of posterior parts of the interpositusnucleus. We have previously shown that damage in this areadoes not impair conditioned responses (Yeo et al. 1985a). Inone other case (E5), the cannula tip was located in thecaudal aspect of the interpositus nucleus.
The remaining four subjects showed high CR frequencies insession 1 of phase 2 (range, 42-100%). Cannula placementsof these subjects were located between 2 and 5 mm from theAIP. Placements in two of these subjects were located closeto the surface of the cerebellum, in the white matterbetween vermis and paramedian lobe, dorsal to the AIP (I6and 17). In another subject, a cannula tip was located in thecerebellar cortex of the paramedian lobe (18), and another atthe base of lobule VI of the cerebellar cortex (I9). Thesesubjects constituted the ineffective group.
Behavioural results. All probability values result fromanalysis of variance tests unless stated otherwise.
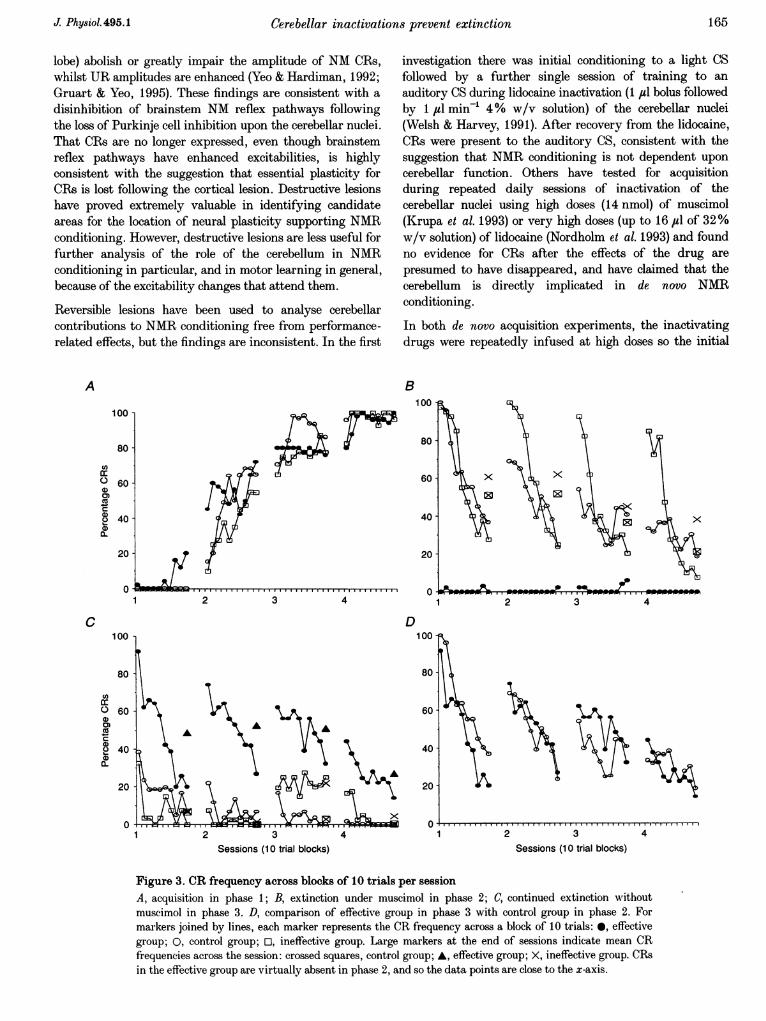
Phase 1. There were significant effects of session (F333 =40 5; P< 0 05) and block (F, 18= 6-15; P< 0 05), consistentwith increases in CR frequency during learning. Groups(Fig. 3A; F21 = 026, P < 0-05) and all interactions werenot significant factors, indicating that the three groupslearned at similar rates.
Phase 2. Before each session of phase 2 all cannulatedsubjects received injections of muscimol. Figure 3B showsthat subjects in the effective group produced very few CRsduring this stage (052 %).
Subjects in the control group showed a pattern of extinctiontypical for this type of conditioning. There was nosignificant group-by-session interaction between the CRfrequencies of the ineffective group and the control group(F324 = 022; P> 0 05), but there was a significant group-by-block interaction (F3,72= 7 10; P<005). Therefore,subjects in the ineffective group extinguished normally oversessions (see session CR frequencies in Fig. 3B), but haddifferent patterns of CR frequencies within the sessions.Inspection of Fig. 3B reveals that the ineffective group hadelevated levels of spontaneous recovery at the beginning ofeach session (see block CR frequencies in Fig. 3B).
Phase 3. All subjects were given continued extinctiontraining for four more sessions (Fig. 3C). The cannulatedsubjects did not receive muscimol in this phase.
At the beginning of session 1, the ineffective group andcontrol group showed low CR frequencies, which extinguishedto baseline levels by session 4. However, subjects in theeffective group showed very high CR frequencies, whichthen extinguished with continued training (see comparisonwith control group, below). There were significant maineffects for groups (F2 11 = 8 14; P < 0 05), confirming thatCR frequencies were different between groups.
Consistent with the very low session CR frequencies in boththe ineffective and control groups in this phase, the maineffect for sessions was not significant (F3 33 = 2-58;P> 0 05), and there was no significant interaction betweengroups and sessions (F6,33 = 1-44; P> 0'05). The significantmain effect for blocks (F9 99 = 1055; P< 0 05) andsignificant interaction between groups and blocks(F18,99= 322; P < 0'05) are clearly due to large within-session differences across groups - the cannulated groupextinguished from high CR frequency to a low CR frequency,in contrast to the CR frequencies of the ineffective andcontrol groups, which showed low levels of extinction withineach session.
By inspection of Fig. 3C, it is clear that the main effect ofgroups was due to greater CR frequencies produced by theeffective group. Over sessions, a Student-Newman-Keulstest revealed that there was no significant difference betweenthe ineffective and the control group (two-way ANOVA,F258 = 0-025, P< 0 05; Student-Newman-Keuls, p = 2,
J. Physiol. 495.1

Cerebellar inactivations prevent extinction
25 mm
3-0
3.5
Figure 1. Reconstructions of cannulated subjectsTransverse sections through the cerebellum andbrainstem at 0 5 mm intervals. Section identifier isdistance from A. Each symbol represents the site ofinjection in a cannulated subject. 0, effectiveplacements; A, ineffective placements. DPFL, dorsalparaflocculus; crll, crus II; HVI, lobule HVI; ND,dentate nucleus; NF, fastigial nucleus; NI, interpositusnucleus; PM, paramedian lobe; VPFL, ventralparaflocculus.
PM~
DPL(
4*0
4.5
5-0
(PFL IX
163J. Physiol. 495.1
VI
... .--' C.-I-7----"..

N Ramnani and C. H. Yeo
Table 1. Frequency of responses occurring between 250 and 1000 ms as percentages + S.D.of all responses
Control group Ineffective group Effective group
Phase 2 1-04 + 2'11 0 95 +1P65 241 + 3O08Phase3 0O4+0-64 1-27 + 1'25 0O5+1'52
q = 004, P> 005), but there was a significant differencebetween the effective and control groups (Student-Newman-Keuls, p = 3, q = 4 07, P< 0 05), and betweenthe effective and ineffective groups (Student-Newman-Keuls, p= 2, q= 360, P< 005).
Comparison of effective group (phase 3) and controlgroup (phase 2). A comparison between the effective groupin phase 3 and the control group in phase 2 (Fig. 3D)revealed no significant main group effect (F1 9 = 0f2,P> 005). There were also no group-by-session(F327 = 1f29, P> 0 05) or group-by-block interactions(Fg,81 = 07, P> 0 05). There were significant block effects(F981= 16 7; P<0 05) and session effects (F327= 4-76;P< 005). Therefore, subjects in the effective groupextinguished at a rate very similar to that of control groupsubjects extinguishing for the first time.
Analysis of responses occurring between 250 and1000 ms. The number of responses occurring after stimulusoffset was very low (Table 1). The tendency for CR onsetlatency to change in the extinction phases was not significant.
In summary, these results show that the muscimol blockadeof the cerebellum appears to prevent the extinction ofconditioned NMRs.
DISCUSSIONIt is now well established that destructive lesions of thecerebellum and its associated circuitry can abolish or impairconditioned reflex responses of the nictitating membrane(for reviews, see Thompson, 1986 and Yeo, 1991). Whethercerebellar lesions selectively impair learning or whether theyproduce more general deficits in the performance ofconditioned responses is controversial. It is clear thatdestructive lesions of the cerebellar nuclei can lead tochanges in the reflex NM response to periocular stimulation.These changes include an extension of the response rise timeand decreased response frequencies at low stimulusintensities close to threshold (Welsh & Harvey, 1989). Thepresence of these deficits may relate to depressedexcitabilities of the brainstem reflex pathways for motorexpression of both conditioned and unconditioned NMresponses. So the interpretation that permanent lesions ofthe cerebellar nuclei produce only learning deficits isconfounded.
Studies using cerebellar cortical lesions have producedevidence that supports the cerebellar learning hypothesismore clearly because the lesion effects upon CRs and URsare strongly dissociated. Lesions of lobules HVI, and partsof HVII and HVIII (medial Crus I and rostral paramedian
Figure 2. Photomicrograph of transverse, Nissl-stained section through the cerebellum andbrainstem of subject El (approximately 3-5 mmcaudal to A)The cannula track penetrated through the paramedianlobe, terminating just above lateral parts of posteriorinterpositus nucleus. Scale bar, 2 mm.
J. Physiol. 495.1164
Cerebellar inactivations prevent extinction
lobe) abolish or greatly impair the amplitude of NM CRs,whilst UR amplitudes are enhanced (Yeo & Hardiman, 1992;Gruart & Yeo, 1995). These findings are consistent with adisinhibition of brainstem NM reflex pathways followingthe loss of Purkinje cell inhibition upon the cerebellar nuclei.That CRs are no longer expressed, even though brainstemreflex pathways have enhanced excitabilities, is highlyconsistent with the suggestion that essential plasticity forCRs is lost following the cortical lesion. Destructive lesionshave proved extremely valuable in identifying candidateareas for the location of neural plasticity supporting NMRconditioning. However, destructive lesions are less useful forfurther analysis of the role of the cerebellum in NMRconditioning in particular, and in motor learning in general,because of the excitability changes that attend them.
Reversible lesions have been used to analyse cerebellarcontributions to NMR conditioning free from performance-related effects, but the findings are inconsistent. In the first
A
100 -
80
0
0
a-
20
*72
3..
3 4
investigation there was initial conditioning to a light CSfollowed by a further single session of training to anauditory CS during lidocaine inactivation (1 ul bolus followedby 1 ,td min-' 4% w/v solution) of the cerebellar nuclei(Welsh & Harvey, 1991). After recovery from the lidocaine,CRs were present to the auditory CS, consistent with thesuggestion that NMR conditioning is not dependent uponcerebellar function. Others have tested for acquisitionduring repeated daily sessions of inactivation of thecerebellar nuclei using high doses (14 nmol) of muscimol(Krupa et al. 1993) or very high doses (up to 16 jul of 32%w/v solution) of lidocaine (Nordholm et al. 1993) and foundno evidence for CRs after the effects of the drug arepresumed to have disappeared, and have claimed that thecerebellum is directly implicated in de novo NMRconditioning.
In both de novo acquisition experiments, the inactivatingdrugs were repeatedly infused at high doses so the initial
B100
80
60 X
40
20
3 4
D100
80
60
3Sessions (10 trial blocks)
1 2 3Sessions (10 trial blocks)
4
Figure 3. CR frequency across blocks of 10 trials per sessionA, acquisition in phase 1; B, extinction under muscimol in phase 2; C, continued extinction withoutmuscimol in phase 3. D, comparison of effective group in phase 3 with control group in phase 2. Formarkers joined by lines, each marker represents the CR frequency across a block of 10 trials: 0, effectivegroup; 0, control group; U, ineffective group. Large markers at the end of sessions indicate mean CRfrequencies across the session: crossed squares, control group; A, effective group; X, ineffective group. CRsin the effective group are virtually absent in phase 2, and so the data points are close to the x-axis.
C100
80
cciro 6000
0)2- 4000L
20
0
m I-% I0 I -T-r-7-1
165J Physiol. 495.1
1

1N Ramnani and C. H. Yeo
absence of CRs after training during cerebellar inactivationsmight be due to a carry-over of residual drug effects into thefollowing acquisition sessions, rather than an absence oflearning under the blockade. In the present experiment we
have tested this possibility by examining extinctionlearning. In extinction it is the absence of CRs whichdemonstrates learning, in contrast to acquisition, in whichthe presence of CRs represents learning. We found thatsubjects given four sessions of extinction training undermuscimol blockade of the cerebellum displayed highfrequencies of NM CRs after the effects of the drug haddissipated and that four more sessions of continued extinctiontraining subsequent to the blockade resulted in behaviourwhich was similar to that of controls, extinguishing ovrerfour sessions for the first time. The presence of CRs afterextensive extinction training indicates firstly, that followingrepeated infusions at low doses, there are no extendedresidual effects of muscimol on the performance of CRs andsecondly, that no learning takes place during the blockade.
The most direct interpretation of our findings is thatextinction learning of the classically conditioned NMR iscritically dependent upon the cerebellum and so is preventedby cerebellar inactivation. But there are other possibilities.Drugs may have three kinds of effect on classicalconditioning. Firstly, they may impair motor expression ofCRs through effects on motor systems. Secondly, they mayalter the sensory properties of the CS and US, which wouldin turn affect learning and performance of CRs. Thirdly,they may impair learning-related plasticity (Schindler &Harvey, 1990). We consider our results in the context ofthese possibilities.
Does muscimol infusion in the cerebellar nuclei affectthe motor expression of CRs?All subjects in the effective group, with cannula locationsclose to the AIP, were trained to asymptotic levels of CRfrequency in phase 1 before administration of the muscimol.In all these cases, inuscimol infusions in phase 2 completelyabolished performance of CRs. It is most likely that duringphase 2, motor expression of CRs was prevented. However,the high frequency of CRs present immediately upon
testing in phase 3, after the effects of muscimol haddissipated, indicates that there were no deficits of motorexpression extending into phase 3 and validates thereversible lesion technique as free from performance deficitsduring the post-inactivation phase. It is not possible toextend this particular validation to an earlier study ofacquisition using higher doses of muscimol (Krupa et al.1993) but a recent study from our laboratory shows theproduction of new CRs is prevented with intracerebellarinfusions of muscimol at the same low dose as used here(Hardiman, Ramnani & Yeo, 1995).
The ineffective group extinguished at the same rate as thecontrol group, but showed enhanced spontaneous recovery atthe beginning of each session n phase 2, though not in phase
3. The cannula placements in this group were not as close to
the AIP as those in the effective group and were mainlydorsal to it. Muscimol infusions would have invaded corticalareas dorsal to the AIP more quickly in the ineffective groupand agonized GABAA-mediated transmission by corticalinterneurons. We suggest that muscimol spread was notoptimally localized to block cortical function in all NMresponse-related areas but was sufficient to produce somedisinhibition of the NM-related areas in the cerebellar nuclei,resulting in small enhancements of reflex expression andthus the enhanced spontaneous recovery. The increasedspontaneous recovery is apparent and similar to'performance' changes that have been noted in other studies.
Does muscimol infusion in the cerebellar nuclei affectsensory properties of the CS or US?We have considered and rejected the possibility of residualmuscimol effects upon motor expression in the critical post-inactivation sessions. The remaining possibility, other thanthat the drug impaired learning-related plasticity, is that thedrug affected the sensory properties of the CS and US. In thepresent extinction learning study, there is no presentationof the US during the muscimol phase, so any possible effectsthat muscimol may have upon the sensory properties of theUS would not be relevant to extinction learning.
If muscimol affected the sensory properties of the CS, thenstate-dependent learning (SDL) may have occurred. Here,subjects learn in the drug condition but cannot retrieve thisinformation unless they are returned to the drug condition.If a drug alters the properties of the CS, subjects will firstassociate the CS and US. But then, after the drug effectshave dissipated, the CS will be perceived as being novel,and subjects will need to re-learn the association in thedrug-free state. Two independent sets of associations maytherefore be formed - one in the drug condition, and theother in the no-drug condition.
To observe SDL, behaviour in the drug condition is oftencompared with behaviour afterwards in the no-drugcondition. However in our study, CR performance wasabolished by the drug in the effective group so overtbehaviour during muscimol inactivation could not beobserved. If SDL had taken place, then subjects in theeffective group may have extinguished covertly undermuscimol in a state-dependent manner in phase 2, and maynot have shown retention for the extinction of CRs becausethere was no muscimol in phase 3. Thus, CRs may havebeen unextinguished at the beginning of phase 3.Alternatively, the subjects may not have extinguished inphase 2 because muscimol inactivation of the cerebellumimpaired learning-related plasticity. In choosing betweenthese options, we consider the following points.
SDL could not have occurred due to muscimol effects inareas surrounding the target area or through the circulationinto more widespread brain areas because the extinctionbehaviour of the ineffective group during and aftermuscimol infusion was similar to that of the control group(see above). In phase 2, CRs of the ineffective group were
J Physiol.495.1166

Cerebellar inactivations prevent extinction1
not abolished by muscimol and extinguished like those ofcontrol subjects in plhases 2 and 3. If muscimol had producedSDL effects in the ineffective group, then subjects wouldhave extinguished in phase 2, and would showN no signs ofhaving extinguislhed in phase 3 (i.e. high CR frequencieswhich would have declined with further training). However,there was a continual decline in CR frequency in phases 2and 3, so extinction in phase 2 was continuous with, and notdissociated from, extinction in phase 3.
A final alternative possibility is that SDL effects may! bepurely local and restricted to the target area of the infusion.In the present experiment, extinction learning may haveoccurred during muscimol inactivation of the AIP withinthe olivocorticonuclear compartment disrupted by druginfusion but in a state-dependent manner. Thus, withinactivation studies, we cannot distinguish between thepossibility that there is SDL within cerebellar circuitry orthe possibility that leaining within cerebellar circuitry isdisrupted. But for the purposes of identifying the locationsof plasticity essential for NMIR conditioning, the distinctionbetween these possibilities is of little importance since bothindicate plasticity within the cerebellum.
Anatomical and temporal specificity of inactivationSince providing anatomical evidence for the convergence ofNAIR conditioning-related information in lobule HVI of thecerebellar cortex (Yeo et al. 1985 b) Mwe have tested forcerebellar cortical mechanisms for NAIR conditioning(Hardiman & Yeo, 1992; Yeo & Hardiman, 1992; Gruart &Yeo, 1995). Our findings are consistent with other models ofmotor learning in the cerebellar cortex (Marr, 1969; Albus,1971; Ito, 1972; Gilbert, 1974) and with recentphysiological studies of cerebellar cortex during eyeblinkconditioning in decerebrate cats (Hesslow, 1994a, b). Our useof muscimol inactivations of the cerebellar nuclei to testcerebellar involvement in the NMR extinction learning may,therefore, appear surprising. Howrever, we suggest thatinactivation of the cerebellar nuclei provides an effectivedisruption of all rielated cerebellar cortical and olivaryregions through losses of nucleo-olivary inhibition anddisruption of nucleocortical inputs.
WTe have assessed the approximate spread of muscimol andalso the time taken for behavioural effects to disappear. W;Vehave considered eviclence from our onwn laboratory and othersources. Mluscimol doses varied, but in all cases, theinjection volumes were 1 jl. AIink & Thach (1991) injected8 8 nmol of muscimiol into the dentate nucleus of monkeys(approximately 6 times the dose used in our study).Behavioural effects began about 3 min after the injectionand disappeared between 7 and 12 h later. Robinson, Straube& Fuchs (1993) also injected 8-8 nmol muscimol into thecaudal fastigial nucleus of monkeys and found behaviouralchanges starting 10 min after injection and lasting for2-3 h, wsith no residual effects 24 h later. Their estimatedradius of spreal was up to 25 mm. In their analysis ofNAIR conditioning in rabbits, Krupa et al. (1993) injectecl
tritiated muscimol into the AIP and autoradiographyrevealed that their dose of 14 nmol did not spread outsidethe cerebellum within 2 h.
Since the dose used in our study (IP54 nmol) is only 11 % ofthe dose used by Krupa et al. (1993), our dose too wouldnot have spread outside the cerebellum in significantconcentrations during a conditioning session. Wheninuscimol was injected up to 2 mm from the AIP, CRs wereabolished consistent with permanent lesion studies (Yeo etal. 1985a). However, if muscimol was injected further awayfromn the AIP, CRs were not abolished. Assuming thatmuscimol abolishes CRs by acting at the AIP, it is highlyunlikely that the injection bolus spread beyond 2 mm or insignificant concentrations beyond the cerebellum. Ourpreliminary data indicate that an injection of 1P54 nmoltakes effect within 5 min. Behavioural effects (depressed CRfrequency or absent CRs) persist for approximately 7 h inovertrained animals, after which CR,s recover completely,and there are no effects on CRs after 24 h (N. Ramnani,unpublished observation).
Therefore, the behavioural time course of muscimol in ourstudy was considerably less than 24 h, and the approximatespread of muscimol was approximately 2 nun. The effects ofmuscimol in one session of training could not have intrudedinto the next.
Muscimol infusion in the cerebellar nuclei preventsplasticity for extinction learning of the classicallyconditioned NMRKrupa et al. (1993) prevented the development of CRs withdoses of muscimol infusions in the AIP much higher thanthose used in the present experiment. If such doses do notproduce SDL or significant post-inactivation performancedeficits, then the cerebellum is clearly implicated in theacquisition of NMR conditioning. Here we havedemonstrated that, at low doses, muscimol inactivations ofthe AIP do not produce general state-dependent learningeffects or significant post-inactivation performance deficitsand so we conclude that our inactivations have preventedthe plasticity associated with the extinction of NMRconditioning. In a related acquisition study (Hardiman,Ramnani & Yeo, 1995) we have also shown that the same lowdose of muscimol prevents the development of CRs in naiveanimals.
On this evidence alone, we cannot conclude that essentialplasticities for the acquisition and extinction of NMRconditioning are confined to the cerebellum. The cerebellummay supply inputs essential for acquisition and extinction toefferent targets but recent work has shown that thispossibility is unlikely. Reversible inactivation withtetrodotoxin of the brachium conjunctivum, the majorefferent pathwvay from the cerebellum, does not- preventacquisition (Krupa & Thompson, 1995). These findings areconsistent with the suggestion that acquisition learning, andnow extinction learning too, are critically dependent uponcerebellar mechanisms.
J Phlysiol. 495.1 167

168 N Ramnani and C. H. Yeo J Physiol.495.1
ALBUS, J. S. (1971). A theory of cerebellar function. MlathematicalBioscien ces 10, 25-6 1.
GILBERT, P. F. (1974). A tlheory that explains the function andstructure of the cerebellum. Brain Research 70, 1-18.
GORMEZANO, I. & GIBBS, C. M. (1988). Transduction of the rabbit'snictitating membrane response. Behaviour Research MIethods,Instruments and Apparatus 20, 18-21.
GORMEZANO, I., SCHNEIDERMAN, N., DEUX, E. & FUENTES, I. (1962).Nictitating memblrane: Classical conditioning and extinction in thealbino rabbit. Science 138, 33-34.
GRUART, A. & YEO, C. H. (1995). Cerebellar cortex and eyeblinkconditioning: bilateral regulation of conditioned responses.Experimental Brain Research 104, 431-448.
HARDIMAN, M. J., RAMNANI, N. & YEO, C. H. (1995). Reversibleinactivation of the cerebellum prevents the acquisition andextinction of conditioned nictitating membrane response. BrainResearch Associationi A bstracts 12, 111.
HARDIMAN, M. J. & YEo, C. H. (1992). The effect of kainic acid lesionsof the cerebellar cortex on the conditioned nictitating membraneresponse in the rabbit. European Journal of N"euroscience 4,966-980.
HESSLOW, G. (1994a). Inhibition of classically conditioned eyeblinkresponses by stimulation of the ceiebellar cortex in the decerebratecat. Journal of Physiology 476, 245-256.
HESSLOW, G. (1994b). Correspondence between climbing fibre inputand motor output in eveblink-related areas in cat cerebellar cortex.Journal of Physiology 476, 229-244.
ITO, Al. (1972). Neural design of the cerebellar motor control system.Brain Research 40, 81-84.
KRUPA, D. J. & THOMPSON, R. F. (1995). Inactivation of the superiorcerebellar peduncle blocks expression but not acquisition of therabbit's classically conditioned eye-blink response. Proceedings ofthe National Academy of Sciences of the USA 92, 5097-5101.
KRUPA, D. J., THOMPSON, J. K. & THOMPSON, R. F. (1993).Localization of a memory trace in the mammalian brain. Scienice260, 989-991.
MACKINTOSH, N. J. (1974). The Psychology of Animal Learnting.Academic Press, London.
MIARR, D. (1969). A theory of cerebellar cortex. Journal of Physiology202, 437-470.
MATRICALI, B. (1961). A new stereotaxic co-ordinate system for therabbit's brain stem. The medial lemniscus in the rabbit: its coursethirough the brain stem and the origin, fibre content andtermination of its compionents. PhD Thesis, University of Leiden.
AIINK, J. W & THACH, T. WV. (1991). Basal ganglia motor control. III.Pallidal ablation: normiial reaction time, muscle cocontraction, andslow movement. Journal of Neurophysiology 62, 330-351.
NORDHOLM, A. F., THOMPSON, J. K., DERSARKISSIAN, C. & THOMPSON,R. F. (1993). Lidocaine infusion in a critical region of the cerebellumcompletely prevents learning of the conditioned eyeblink response.Behavioural Neuroscietnce 107, 882-886.
ROBINSON, F. R., STRAUBE, A. & FUCHS, A. F. (1993). Role of thecaudal fastigial nucleus in saccade generation. II. Effects ofmuscimol inactivation. Journal of Nleurophysiology 70, 1741-1758.
SCHINDLER, C. NV. & HARVEY, J. A. (1990). Use of classicalconditioning proceduures in behavioural pharmacology. DrugDevelopmnent Research 20, 169-187.
THACH, WV. T. (1996). On the specific role of the cerebellum in motorlearning and cognition: clues fiom pet activation and lesion studiesin man. Behavioural andl Brain1 Sciences (in the Press).
THOMPSON, R. F. (1986). The neurobiology of learning and memory.AScience 223, 941-947.
THOMPSON, R. F. & KRUPA, D. J. (1994). Organization of memorytraces in the mammalian brain. Anlnual Review of Neuroscience 17,519-549.
WELSH, J. P. & HARVEY, J. A. (1989). Cerebellar lesions and thenictitating membrane reflex: performance deficits of the conditionedand unconditioned response. Journal of Neuroscience 9, 299-311.
WELSH, J. P. & HARVEY, J. A. (1991). Pavlovian conditioning in therabbit during inactivation of the interpositus nucleus. Journal ofPhysiology 444, 459-480.
YEO, C. H. (1991). Cerebellum and classical conditioning of motorresponses. Proceedings of the New York Academy of Sciences 627,292-304.
YEO, C. H. & HARDIMAN, Al. J. (1992). Cerebellar cortex and eyelidconditioning - A reexamination. Experimental Brain Research 88,623-638.
YEO, C. H., HARDIMAN, Ml. J. & GLICKSTEIN. Ml. (1985a). Classicalconditioning of the nictitating membrane response of the rabbit: Ilesions of the cerebellar nuclei. Experimental Brain Research 60,87-98.
YEO, C. H., HARDIMAN, AM. J. & GLICKSTEIN, M. (1985b). Classicalconditioning of the nictitating membrane response of the rabbit: IIIConnections of cerebellar lobule HVI. Experimental Brain Research60,114-126.
AcknowledgementsThis research is supported by the Medical Research Council and bya graduate studentship from the Department of Anatomy andDevelopmental Biology, University College London, UK.
Author's email addressN. Ramnani: [email protected]
Received 9 January 1996; accepted 30 April 1996.