review article - ncbi
TRANSCRIPT

THE EFFECTS OFBACTERIAL ENDOTOXINSON HOST MEDIATIONSYSTEMS
REVIEWARTICLE

The Effects of Bacterial Endotoxins on Host Mediation Systems
Htorical B od 527
Compostion of Bactei Endotons 529
Rela_onship dof Endoto2an Sbtctu to Biologic Act 532
Intracto of E oins With Host Mediation Systems 534
Humoral Mediaton Systems 535Serum Complement 535
Endotoxin-Complement Interactions In *iwvo 537Endotoxin-Complement Interactions In *Vitro 543
Coagulation Systems 550Intrinsic Coagulation Pathwav 552
Endotoxin Activation In *Vizo 552Endotoxin Activation In *itro 554
Extrinsic Coagulation Pathway 556Endotoxin Activation In Vivo 556Endotoxin Activation In Vitro 557
Cdul|ar Mediation System 560Platelets 560
Endotoxin-Platelet Interactions In *Vico 561Endotoxin-Platelet Interactions In *Vitro 568
Pol-morphonuclear Leukocy-tes (Neutrophils) 575Endotoxin-Neutrophil Interactions In *ivo 575Endotoxin-Neutrophil Interactions In *itro 580
\ acrophage M6\onocyte-Endotoxin Interactions 586Endothelial Cell-Endotoxin Interactions 594Mast Cells, Basophils 597
Mast Cell-Endotoxin Interactions In * iro 598Mast Cell-Endotoxin Interactions In V'itro 599
Sumnma ad Cond sn 601

The Effects of Bacterial Endotoxins on Host MediationSystems
A Review
D. C. Morrison, PhD, R. J. Ulevitch, PhD
Historical BackgroundIt has been recognized for almost 100 years that a number of bacterial
products have the capacity to exert profound effects on humans and otheranimals. One such class of bacterial products has been shown to normallyexist in close association with the bacterium and to be released only onbacterial Iysis. These toxic materials have been designated "endotoxins"to distinguish them from toxic substances synthesized and excreted by theintact bacterium, termed "exotoxins.''" Although endotoxins w%ere firstrecognized for their ability to induce fever (reviewed in Reference 2), theyhave since been shown to have a broad spectrum of biologic activities,manv of which will be reviewed here. Research by a number of investiga-tors has firmlv established that true endotoxins are derived only fromgram-negative bacteria and normally exist within the bacterium as in-tegral components of the bacterial cell wall.A schematic representation of the gram-negative bacterial cell wall is
shown in Text-figure 1. The bacterial surface basically consists of an innercytoplasmic membrane and a trilayer outer cell wall structure consistingof a mucopolvsaccharide-peptidoglycan layer, a phospholipid-proteinlayer, and the outermost lipopolysaccharide layer (LPS).3 Some strainsof gram-negative bacteria contain, in addition, a capsular polvsaccharidelaver. The LPS portion of the cell wall is chemically unique for each strainof bacteria and thus mav be used in the serologic classification of variousbacterial species (reviewed in Reference 4). On bacteriolysis, endotoxins,consisting of aggregates of lipopolysaccharides and protein (and, perhapsto a lesser extent, loosely bound lipids), are released from the bacteriuminto the surrounding medium.
From the Department of Immunopathology, Scripps Clinic and Research Foundation. La Jolla.California.
Supported in part by NIH Grants Al 13187, NHLBI 16411, and Al 07007. Dr. Morrison is therecipient of a USPHS Research Career Development Award IK04AI 00081.
This is publication No. 1513 from the Department of Immunopathology. Scripps Clinic andResearch Foundation, La Jolla, CA 92037.
Accepted for publication April 13, 1978.Address reprint requests to David C. Morrison, PhD. Department of Immunopathology. Scripps
Clinic and Research Foundation, 10666 Torrey Pines Road, La Jolla, CA 92037.0002-9440/78/01 19-0525/$01.00 527

528 MORRISON AND ULEVITCH American Journalof Pathology
TEXTr-F1GCRE 1-Structure ofthe bacterial cell membrane.
The selective extraction of these endotoxic materials from the bacte-rium established the basis for significant scientific advances in under-standing both the biology and the chemistry of these complex bacterialproducts. Two of the most popular of these extraction methods, which are
still extensively used for the preparation of endotoxins, are the proceduresdescribed by Boivin and Mesrobeanu 5 and Westphal, Liideritz, andBister.6 The former procedure (Boivin) involves extraction with ice-coldtrichloroacetic acid and results in endotoxic preparations which primarilyconsist of lipopolysaccharides but contain, in addition, protein and lipid.The latter procedure (Westphal) utilizes a hot aqueous phenol extractionof bacterial suspensions and yields a pure protein-free lipopolysaccharide,
which also manifests potent endotoxin activities. In addition to theseprocedures, other investigators have described a number of useful meth-ods to obtain endotoxic materials from gram-negative bacteria, includingthe diethylene glycol extraction procedure of Morgan,7 the aqueous etherextraction described by Ribi et al 8 the EDTA extraction procedure ofLeive et al,9 the method for selective extraction from rough mutantsdescribed by Galanos et al,10 and the recently described procedure ofMorrison and Leive " employing aqueous butanol. Each of these extrac-tion methods yields a biologically active material rich in lipo-polvsaccharide with, in many cases, variably lesser amounts of protein andlipid.The demonstration that virtually all of the biologic activities normally
attributable to bacterial endotoxins could also be elicited with isolatedchemically pure lipopolysaccharides (and, as will be indicated below, inmost instances, the lipid portion of the lipopolysaccharide molecule) hasprompted numerous investigators to utilize interchangeably the terms"'endotoxins" and "lipopolysaccharides" or "LPS.'" It is, however, ex-
tremely important to recognize that these two bacterial products may besignificantlyr different, both with respect to chemical composition and
Outer
luuer

Vol. 93, No. 2 BACTERIAL ENDOTOXINS 529November 1978
biologic activity. As defined by Braude in the ninth edition of the MerckIndex,'2 endotoxins are "lipopolysaccharide-protein complexes containedin cell walls of gram-negative bacteria including non-infectious gramnegatives," and thus the presence or absence of the protein componentserves to distinguish endotoxins from purified lipopolysaccharides.Equally as important as the chemical distinctions which serve to differen-tiate endotoxins and LPS are the potential differences in biologic activitiesof these bacterial products. Unfortunately, although originally recognizedby several early investigators as a major constituent of the "endotoxincomplex" isolated from gram-negative bacteria, the protein componenthas, in the past several decades, received considerably less attention thanthe lipopolysaccharide component. This in part reflects the significantadvances which have been made in defining the chemical structure of theLPS molecule and its relationship to endotoxic activity.
The Composition of Bactenal EndxinsAs indicated above, endotoxins isolated from gram-negative bacteria
primarily consist of lipopolysaccharides (LPS) with variable amounts ofprotein and lipid. The LPS molecule itself consists of a polysaccharideregion covalently bound to a lipid region, termed "lipid A" by Westphaland his associates, to distinguish it from noncovalent loosely bound lipidsassociated with the LPS molecule.s The polysaccharide portion of theLPS molecule is generally considered as consisting of two distinct regions:a "core" polysaccharide and an "O-antigen" polysaccharide. The corepolysaccharide region, which is often identical for large groups of bac-teria, contains a unique deoxysugar 2-keto 3-deoxy-octulosonate (KDO) aswell as heptose, phosphorylethanolamine, and several hexoses which to-gether define the core structure. The 0-antigen, in contrast, is chemicallyunique for each type of organism and LPS. The 0-antigens normally existas repeating oligosaccharide units containing three to four different hex-ose units each. The number of repeating oligosaccharide units can be asfew as two (semirough) or as many as 10 (smooth) even within the samebacterium.11"4l"o These basic LPS structural features are indicated dia-grammatically in Text-figure 2, and a more detailed chemical structure ofone such LPS molecule (from Escherichia coli 0111: B4) is shown in Text-figure 3.11.16 The LPS from other strains of bacteria would be structurallysimilar to that shown for E coli 0111:B4, although different hexoses wouldbe substituted in the core and 0-antigen region. Although shown asmonomeric structures, it should be noted that these molecules almostinvariably exist as high-molecular-weight aggregates (>106 daltons) andare often particulate.
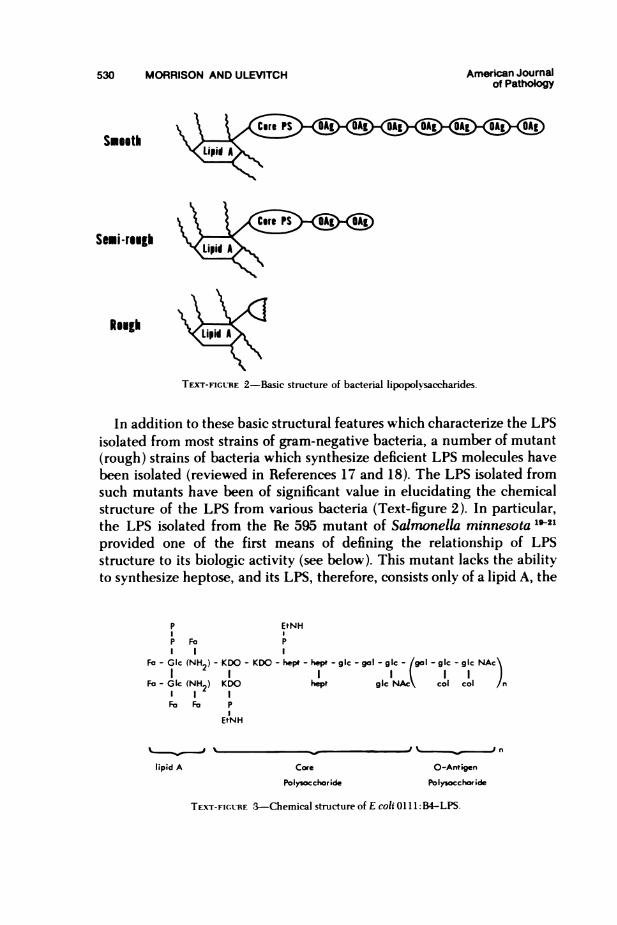
530 MORRISON AND ULEVITCH American Joumalof Pathology
Smooth
Semi-rough
Rough
TEXT-FIGURE 2-Basic structure of bacterial lipopolysaccharides.
In addition to these basic structural features which characterize the LPSisolated from most strains of gram-negative bacteria, a number of mutant(rough) strains of bacteria which synthesize deficient LPS molecules havebeen isolated (reviewed in References 17 and 18). The LPS isolated fromsuch mutants have been of significant value in elucidating the chemicalstructure of the LPS from various bacteria (Text-figure 2). In particular,the LPS isolated from the Re 595 mutant of Salmonella minnesota 19-21provided one of the first means of defining the relationship of LPSstructure to its biologic activity (see below). This mutant lacks the abilityto synthesize heptose, and its LPS, therefore, consists only of a lipid A, the
P EtNHI IP Fa PI I I
Fa - Glc (NH2) - KDO - KDO - hept - hept - gic - gal - gic - gal - gkc - gic NAc\I I I I IA I I J
Fa- Gk (NH2) KDO hept gicNAc col col nI I IFo Fo P
EtNH
lipid A Core O-Antigen
Polysocchoride Polysocckioide
TEXr-FIGc-RE 3-Chemical structure of E coli 011 1: B4-LPS.

Vol. 93, No. 2 BACTERIAL ENDOTOXINS 531November 1978
trisaccharide of KDO, and phosphorylethanolamine. More recently, heptose-deficient mutants have been characterized from several other strainsof bacteria.'=-'The lipid portion of the LPS molecule, ie, lipid A, is unique. As first
suggested by Burton and Carter and confirmed by a number of investi-gators, lipid A consists of a 1i-6 linked diglucosamine backbone con-taining both ester- and amide-linked long-chain fatty acids as well aspyrophosphate groups.' The basic structure of lipid A is indicated inText-figure 4. The ester-linked fatty acids appear to have no intrinsic spe-cificity, and both even- and odd-numbered long-chain hydrocarbons maybe present. The amide-linked fatty acids, in contrast, appear to be uni-formly ,B-OH substituted." The basic structure of the lipid A region ofLPS isolated from a broad spectrum of gram-negative bacteria is remark-ably similar.18''T' It might, therefore, not be surprizing to find that manyof the biologic activities of bacterial endotoxins are associated with thischemically unique amphipathic structure.
In contrast to the information available on the LPS portion of bacterialendotoxins, significantly less is known about the protein component (re-viewed in Reference 28). Although most of the earlier work attempted todefine the biologic properties of the protein component of endotoxins, anumber of investigators have recently examined the chemical nature ofthese proteins as well as their association with the LPS. Several recentreports '*-31 have suggested that protein may be covalently linked to theLPS molecule, perhaps through the lipid A region. It has recently beenshown ",32," that the protein in endotoxins prepared by, for example, theBoivin TCA I procedure or the aqueous butanol procedure 11 may be
i
I 04-
TEXr-FiGURE 4-Chemical structure of lipid A.I ",tAKDO]3
11P094-

532 MORRISON AND ULEVITCH American Joumalof Pathology
selectively extracted into the phenol phase on subsequent extraction withaqueous phenol." The endotoxin-associated proteins have recently beenshown to have a number of biologic activities and have been termed"' endotoxin protein" (EP)2 or "lipid A-associated protein" (LAP).33 Forthe purposes of this review, we will adopt the latter terminology.
Reatioship of Eoxin Sttue Bio c ActtThe experiments of Tal and Goebel 3 almost 30 years ago provided the
first information on the nature of the toxic component of bacterial endo-toxins. On the basis of limited acid and/or alkaline hydrolysis, theseinvestigators postulated a biologically active component (termed "com-ponent T") which endowed these preparations with toxicity. In the fol-lowing 2 decades, numerous studies attempted to correlate biologic activ-ity with the various physical and/or chemical properties of endotoxinpreparations with results implicating both the polysaccharide -" and thelipid " regions of the LPS component of bacterial endotoxins as havingthe predominant biologic activity. Other investigators " suggested thatneither fraction alone could elicit full biologic activity but rather that themacromolecular complex formed by the chemical bonds linking the lipidwith the polysaccharide was required for the manifestation of full biologicactivity. It was, therefore, not until the isolation and characterization of aheptose-deficient mutant strain of a gram-negative organism (the R595mutant of S minnesota) that the biologic activity of endotoxin prepara-tions was firmly established.l ll2" The demonstration by a number ofinvestigators that endotoxins (or LPS) prepared from these poly-saccharide-deficient (rough) mutants still manifest virtually all of theendotoxic properties of polysaccharide-containing endotoxins firmly es-tablished that lipid A plays a dominant role in endotoxicity.
In addition to the use of endotoxins from mutant strains of bacteria toestablish structure-function relationships, a number of physical-chemicalprocedures have been utilized to modify selective portions of the endo-toxin molecule. Significant advances in the procedures first used by Taland Goebels ' have yielded an abundance of information about thecomponents of the endotoxin complex required for biologic activity. Forexample, mild acid hydrolysis (usually in 1% glacial acetic acid at 100 Cfor 2 to 3 hours) cleaves the KDO bond linking the lipid A region of theLPS to the core polysaccharide.1 The lipid A, being hydrophobic, isinsoluble in aqueous solvents and thus may be easily separated from thepartially degraded, although antigenically intact, polysaccharide either bycentrifugation or extraction into organic solvents. Mild alkaline hydroly-sis 1° normally has employed conditions approximating 0.25N NaOH for
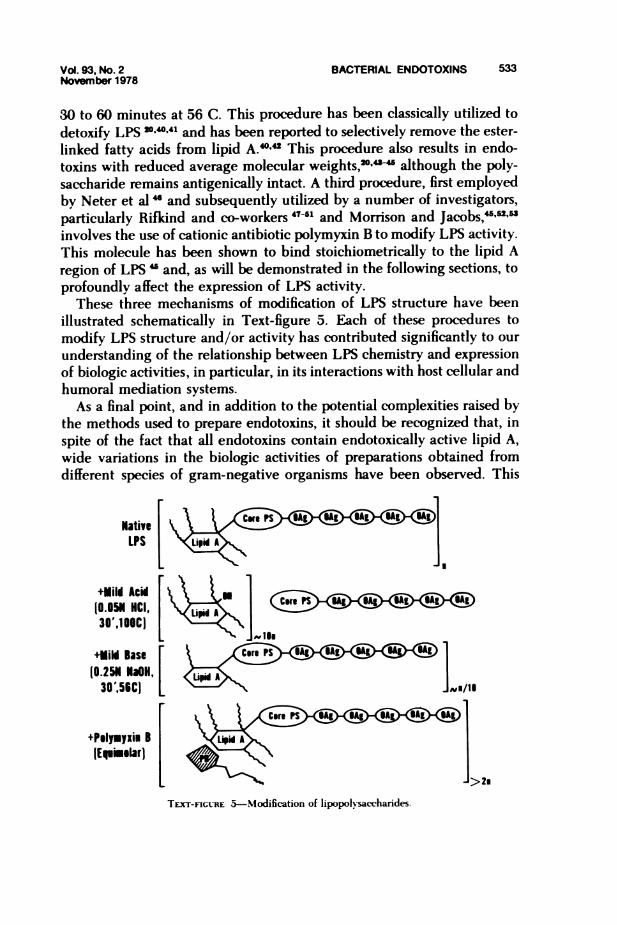
Vol. 93, No. 2 BACTERIAL ENDOTOXINS 533November 1978
30 to 60 minutes at 56 C. This procedure has been classically utilized todetoxify LPS 204041 and has been reported to selectively remove the ester-linked fatty acids from lipid A.4042 This procedure also results in endo-toxins with reduced average molecular weights,2'"- although the poly-saccharide remains antigenically intact. A third procedure, first employedby Neter et al " and subsequently utilized by a number of investigators,particularly Rifkind and co-workers 47-51 and Morrison and Jacobs,-"' "involves the use of cationic antibiotic polymyxin B to modify LPS activity.This molecule has been shown to bind stoichiometrically to the lipid Aregion of LPS " and, as will be demonstrated in the following sections, toprofoundly affect the expression of LPS activity.
These three mechanisms of modification of LPS structure have beenillustrated schematically in Text-figure 5. Each of these procedures tomodify LPS structure and/or activity has contributed significantly to ourunderstanding of the relationship between LPS chemistry and expressionof biologic activities, in particular, in its interactions with host cellular andhumoral mediation systems.As a final point, and in addition to the potential complexities raised by
the methods used to prepare endotoxins, it should be recognized that, inspite of the fact that all endotoxins contain endotoxically active lipid A,wide variations in the biologic activities of preparations obtained fromdifferent species of gram-negative organisms have been observed. This
NativeLPS
in
+Uili Aci10.051 HCI,30',10C)
+Uild Base(0.25N1 HON,
30'956C)
+PsIpyxi B [(Epimelarl
L >2m
NE/i
Avg/II
TEXr-FIGURE 5-Modification of lipopolvsaccharides.
- ----. - . -- .- .- IL, 0% 0%
0- - - --

534 MORRISON AND ULEVITCH American Journalof Pathology
would, in part, be reflective of a number of variables, including intrinsicactivity of LAP, differential modulation of lipid A activity by LAP and/orby polysaccharide, polysaccharide composition of the LPS, degree ofaggregation, association of metal ions, and, perhaps, the culture condi-tions for growth of the organisms. An investigator should therefore becautious in attributing a biologic activity to bacterial endotoxins on thebasis of an experiment performed with a single endotoxin preparationfrom one organism. This point would be especially true in the case ofbiologic activities ascribed to endotoxins versus those ascribed to purifiedlipopolysaccharides; several recent publications have underscored theprofound differences which can result when these two distinct prepara-tions from a single bacterial species are compared."'"
InteractI With Host i systemsA major function of the LPS on the bacterial cell surface is to serve as a
selective permeability barrier in controlling the transport of moleculesinto the cell." In addition, however, the location of the LPS on theoutermost surface of the bacterium allows it the maximal opportunity tointeract with the external environment. In many instances, this is theintestinal tract of most mammalian species, where gram-negative orga-nisms are normally a major constituent of the resident flora. To maintainthis symbiotic relationship it would thus appear essential for the host (andfor the bacterium) to utilize the full spectrum of its defense mechanisms toprotect against potential invasion of other tissues by the bacteria, whichwould lead to gram-negative sepsis and, ultimately, death.
It is in many cases, however, the host response to the gram-negativeorganism, rather than the organism itself, which poses the ultimate threatto host tissues. As pointed out by Thomas,57 "Our arsenals for fighting offbacteria are so powerful and involve so many different defense mecha-nisms, that we are in more danger from them than from the invaders." Hefurther observes that "these (endotoxin) macromolecules are read by ourtissues as the very worst of bad news. When we sense lipopolysaccharide,we are likely to turn on every defense at our disposal."
It is the purpose of this communication to review the studies whichhave contributed to our understanding of the mechanisms by which thesevarious host defense systems become turned on by endotoxins. Attentionhas focused on the molecular mechanisms of interaction of endotoxin withcellular and/or humoral mediation systems which lead to the generationof an immediate cell response. Although, for the most part, in vitroresponses will be considered separately from in vivo responses, the resultsobtained will, when relevant, be correlated. Because of the numerous

Vol. 93, No. 2 BACTERIAL ENDOTOXINS 535November 1978
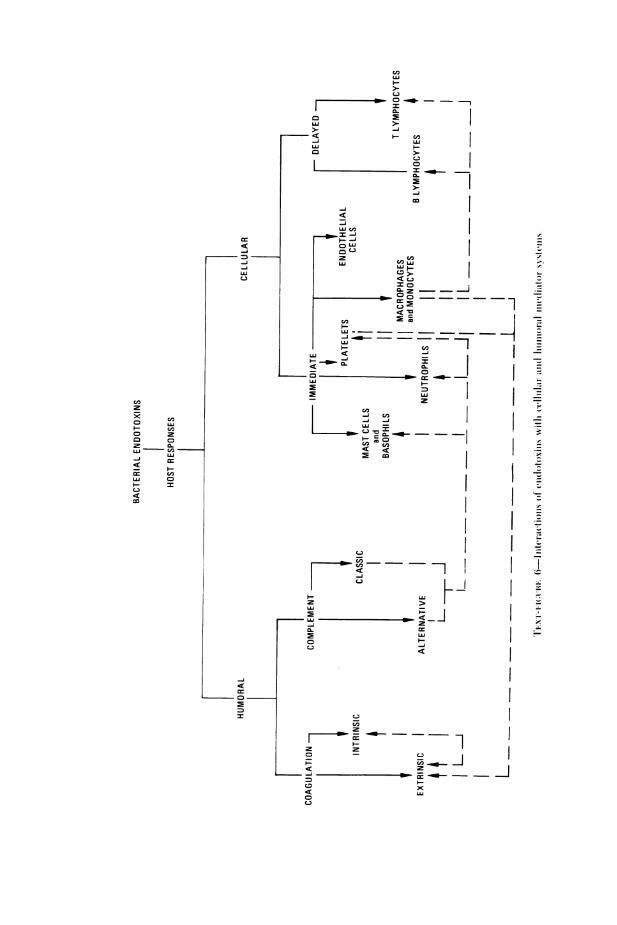
excellent review articles and symposiums which have been published onthe chemistry, biology, and immunology of lipopolysaccharides,4 16"the majority of the research covered in this review will deal with resultsobtained during the past 2 decades. Whereas much of the earlier datadefined in a phenomenologic sense the biologic activities of endotoxins,the past 20 years have witnessed a synthesis of the knowledge of thechemistry of LPS with its biologic activities and the beginning of theelucidation of the molecular mechanisms of endotoxin action.The interactions of endotoxins with cellular and humoral mediator
systems which will be covered in this review are outlined in Text-figure 6.The major humoral mediation systems, complement (both classic andalternative pathways) and coagulation (both extrinsic and intrinsic, in-cluding kinin-forming and fibrinolytic), will both be reviewed. In addi-tion, the effect of endotoxin-activated humoral components on cellularresponses will be explored. Studies of the direct effect of endotoxins oncells, including platelets, mast cells, basophils, neutrophils, monocytesand macrophages, and endothelial cells, will be discussed. Finally, theeffect of endotoxin-activated cell products on the humoral mediationsystems will be defined.One of the important areas of the interaction of endotoxins with cellular
mediator systems which will not be covered in this review is their ability toinitiate and to modulate lymphocyte responses, indicated in Text-figure 6as delayed cellular responses. The effect of endotoxins on lymphocytesand the immune response has been intensively studied during the past 2decades, resulting in a considerable body of knowledge concerning thesemultiple immunologic responses. A review of the pertinent literaturedealing with this subject will be published elsewhere."
As a final point, for the purpose of this review, we have adopted theunifying terminology of endotoxin to refer to studies performed both withendotoxins and with purified lipopolysaccharides. Wherever possible,however, we have indicated both the method of preparation of the endo-toxin and the organism from which it was derived. (In several instances,the biologic activity has been shown to depend critically on both thesource and method of extraction.) It is intended that this information mayallow the reader to account for potential variations in results obtained byindividual investigators.
Hunoral Mediatio SystemSW=u COMImpmt
The serum complement system of humans and other vertebrates repre-sents one of the major mediation systems in the circulating plasma.
cnLLJ
C-,l - o ~- - -~
0 2
-J LULJ
o~~~~~~~~c
LUJ L
LLJ F-
O U CD
O o
X~~~~
<-
ILCM
cnc sI
LL _° _
z- F-- _
I cn c<a- I_
LUI-
Z >L
-JCd
LLJ -4m IC2- _>
T-.j z
2 LUJ0I
,
0J - C- II
_ 2 _CC z I
C-
Cnzx n
0 LUF-cn
0 Z
0 0
z
LLI cn
LUJ 0_ F-
m
C-
~~02=C
-j

Vol. 93, No.2 BACTERIAL ENDOTOXINS 537November 1978
Initiation of the complement protein sequence may be accomplished byeither of two pathways: the classic pathway following initiation by antigenand IgG or IgM antibody and the altemative (properdin) pathway initi-ated by naturally occurring high-molecular-weight polysaccharides." Itmight not be surprising, therefore, to find that bacterial endotoxins wouldhave the capacitv to activate each of these complement pathways. Theresults of experiments performed during the past 2 decades have clearlydemonstrated this to be the case. The use of endotoxins was, in fact,instrumental in elucidating the mechanism of altemative complementpathway activation. Of significant interest, however, was the demonstra-tion that, in addition to altemative pathway activation by endotoxin,activation of the classic pathway by an antibody-independent mechanismcould also be accomplished. Considerable evidence has accumulated tosupport the concept that different portions of the endotoxin molecule areresponsible for these distinct anticomplementary activities, which aresummarized schematically in Text figure 7. The accumulated data demon-strate three distinct mechanisms for the activation of serum complement,onlv one of which requires antibodv to the endotoxin molecule. Theexperiments which led to these conclusions will be discussed in detailbelow. An excellent review on some of the earlier results to be discussedhas appeared previously.70
Endotoxin-Complement Interactions In Vivo
Perhaps the earliest studies detailing a relationship between the struc-ture of bacterial endotoxin and complement proteins in vivo were theexperiments of Landy and Pillemer.7' These authors tested 18 prepara-tions of endotoxin in mice and showed that the injection of endotoxin
Classic ComplementAlternative Complerent th^
PathAnti-Endotoxin
i Activtir Sura Antibodyiia+ > ENDOTOXINl r -
Fluid Phm C3b C1lI
K _ Antipn-AntibodyComplex
P+C3b+ B + D PBDC3b o C4, 2 C C4 + C2
C5-9
TE.XT-FIGtRE 7-Interaction of endotoxin mvith the complement system

538 MORRISON AND ULEVITCH American Joumalof Patholog
induced increased levels of circulating properdin (a major protein com-ponent of the alternative pathway) and that these levels correlated withincreased protection against infection. They noted that there was nocorrelation between the amounts of bound lipid or protein to the activitythey observed, and they thus concluded that it was the carbohydrateportion of their preparations which was responsible for the observedactivity. These authors further speculated that the spatial configuration ofthe sugar residues in the LPS portion of the endotoxin preparations wasalso of importance in determining the expression of activity. Hook et al 7
also noted alterations in mouse properdin levels following the administra-tion of S typhosa endotoxin, although these investigators found a muchmore rapid (2-hour) drop in properdin levels, which retumed to normalafter approximately 2 days.The classic experiments of Spink and Vick 74 provided evidence that
complement components may be involved in the lethal effects of endo-toxins in dogs. These authors were able to protect against the lethal effectsof endotoxins by transferring heated (complement-inactivated?) plasma todogs prior to administration of endotoxin. Additional studies by Spink etal 75 demonstrated rapid drops in complement titers (50 to 80%) in dogsgiven an LD,oo of endotoxin. These authors proposed an endotoxin-antibody anaphylactic mechanism to explain the initial rapid fall incomplement. These authors noted that, in dogs given an LD. dose ofendotoxin, the complement drops in the survivors were qualitatively sig-nificantlv less than those observed in the nonsurvivors.
Essentially the same results were obtained in rabbits, first by Gilbertand Braude 7" and subsequentlv by Pearlman et al.'" The results of theformer investigators demonstrated that administration of an LD,10 dose ofendotoxin (E coli, Proteus mirabilis, Boivin extract, 5.0 mg) caused rapid1- to 3-hour drops in levels of complement titers. However, administrationof endotoxin below the LDo0 level caused only sporadic alterations ofcomplement titers. Somewhat in contrast to these results (which may bereflective of the differences of intrinsic anticomplementary activity of theendotoxins used in these studies), Pearlman et al 77 showed that adjuvantamounts of endotoxins (S enteritidis, 5 Mg; S typhosa, 50 Ag) will reducerabbit complement levels as much as 33% over a 6-hour period. Theselatter authors argued that these effects could not be mediated by antibodybecause of the relatively slow drop in complement titers in comparisonwith antigen-antibody complexes.An important question with regard to mechanisms of endotoxin-initi-
ated complement activation was the potential role of natural antibody tothe endotoxin molecules. The presence of such antibody would allow the

Vol. 93, No. 2 BACTERIAL ENDOTOXINS 539November 1978
formation of endotoxin-antibody complexes and thus provide a simplemechanism for the activation process. To address this problem, Miler etal 78 injected endotoxins (S paratyphi B) into precolostral newborn pigletswhose plasma did not contain detectable antibody. As controls, theseauthors injected saline or a high-titer rabbit antiendotoxin antibody priorto administration of endotoxin. The results of these experiments demon-strated that only those piglets which received antibody prior to endotoxinshowed drops in complement levels, suggesting a prominent role forantiendotoxin antibody in the anticomplementary activity of endotoxins.The results of similar in vitro experiments did not, however, support thesein vivo observations. (See Endotoxin-Complernent Interactions In Vitro.)The first unequivocal evidence for an in vivo antibody-independent
activation of serum complement by endotoxin was provided by Kane,May, and Frank.79 These authors injected endotoxin (E coli 0127:B8,Westphal) into normal guinea pigs and guinea pigs congenitally deficientin the fourth component of complement. These latter guinea pigs show anabsolute deficiency in the response of their serum complement to antigen-antibody complexes. When injected with endotoxin, however, significantdrops in the levels of C3-9 were observed in both groups of animals.These authors did, however, note several differences in the responses ofthese two groups of animals which were dependent on C4. (See Endo-toxin-Platelet Interaction In Vivo.) These studies thus provided unequiv-ocal evidence that bacterial endotoxins were capable of activating boththe classic and the alternative pathways of complement in guinea pigs,and these results have confirmed the earlier in vitro conclusions on alter-native pathway complement activation by endotoxin.Accompanying the increased recognition of the capacity of endotoxins
to interact with serum complement was the question of the potentialcontribution of such interactions to the pathophysiologic effects which arecharacteristic of endotoxin-mediated host responses. As discussed earlier,the experiments by Gilbert and Braude in rabbits 76 and Spink et al indogs 74 provided data which correlated significant drops in complementwith potential lethality in those animals given either an LD,o or an LDsoof endotoxin. More recent experiments, however, have established severalimportant model systems to assess more completelv the role of endotoxin-initiated complement activation on endotoxic activities in vivo. The firstof these has employed an anticomplementary protein isolated from thecobra venom of Naja naja.w This protein, termed "CoF" for cobra factorprotein, has been demonstrated to form a molecular complex with factor Bof the alternative pathway 81 and to efficiently consume CS and terminalcomponents both in vitro 8 and in vivo ° in a variety of species. When

540 MORRISON AND ULEVITCH American Journalof Pathdogy
appropriately administered to experimental animals, CoF will reducecirculating C3 levels to 5% of normal levels or less within 24 hours. Thisprocedure thus provided an experimental model to assess the effect of C3and terminal complement components in endotoxin-mediated patho-physiologic changes. The second model has employed the use of experi-mental animals genetically deficient in the synthesis of specific com-plement components, eg, mice deficient in C5, rabbits deficient in C6, or,as mentioned above, guinea pigs deficient in C4. Most studies havefocused on these latter two experimental animals.Of primary interest in these studies has been the assessment of the role
of C3 and terminal complement components in the pathophysiologicchanges induced following the administration of endotoxins. In this re-spect, the effect of complement on hemodynamic and hematologicchanges induced by endotoxin as well as on the local and generalizedShwartzman reaction and on lethality has been determined, often withless than uniform results. Particular emphasis has focused on changes inlevels of circulating formed elements such as platelets and white bloodcells and in plasma proteins such as the coagulation proteins. These willbe dealt with separately in the appropriate sections of this review.
Perhaps the first evidence to demonstrate directly a role for CS in thepathologic lesions induced by endotoxins was described by Polak andTurk.'" These authors used a number of different antiserums directedagainst various guinea pig components, including anti-T-cell, antilymphnode, antigranulocyte, anti-C3, anti-Y-globulin, and antiguinea pigserum, to deplete selectively guinea pigs. Significantly, only the anti-CSantibody suppressed the hemorrhagic lesions induced by bacterial endo-toxins.The experiments of From et al " provided support for a prominent role
of C3 and terminal components in the immediate hypotensive changesobserved following the administration of E coli endotoxin (LDSO, 750 ,g/kg) to dogs. Confirming earlier studies by several investigators, theseexperiments demonstrated moderate drops in complement titers afterinjection of endotoxin. Determinations of mean arterial blood pressures inthese animals showed a rapid drop in blood pressure following endotoxininjection, a partial recovery, and a second more gradual decrease. Ofsignificant interest, treatment with CoF abrogated the initial rapid drop inblood pressure but had no effect on the secondary blood pressure drop orthe ultimate endotoxin-induced mortality. Essentially identical results tothose obtained by From et al"u were also obtained in a feline model ofendotoxin shock described by Kitzmiller et al." Similar experiments byGarner, Chater, and Brown " in beagle dogs using E coli 026:B6 endo-

Vol. 93, No. 2 BACTERIAL ENDOTOXINS 541November 1978
toxin (4 mg/kg, LD1, within 6 hrs) yielded similar results. The animalsgiven endotoxin experienced a biphasic change in blood pressure whichwas abrogated if the animals were pretreated with CoF. In contrast to theearlier study, both the initial and the secondary changes in blood pressurewere altered by CoF pretreatment.The rabbit has also provided an interesting model for examination of
the relationship between complement- and endotoxin-induced patho-physiologic changes, and several investigators have examined variousaspects of the rabbit responses. Fong and GoodM* utilized CoF to depleterabbits of C3 and terminal complement components and then inducedlocalized and generalized Shwartzman reactions with two injections of Ecoli endotoxin (Boivin extract 100 to 270 Mg) 24 hours apart. (It may beimportant to note that the endotoxin preparation utilized in these experi-ments was found by these investigators to be highly anticomplementary,with as little as 1.4 gg of endotoxin reducing by 50% the hemolyticcomplement titer of a 1: 5 dilution of normal rabbit serum.) Of signifi-cance, only 1 of 15 of the complement-depleted rabbits developed eitherthe localized or the generalized Shwartzman reaction, in comparison tocontrol rabbits, 12 of 15 of which developed lesions. These data, therefore,suggest a role of C3 in the induction of the hemorrhagic necrosis charac-teristic of the Shwartzrnan reaction following the administration of endo-toxin and would be consistent with the earlier results obtained by Polakand Turk in the guinea pig.n The more recent experiments by Bergsteinand Michael,'" however, using the identical experimental system, yieldedcompletely opposite results. These investigators, using E coli 026: B6endotoxin (350 to 400 ,g Boivin), were completely unable to prevent thedevelopment of glomerular fibrin deposition or cortical necrosis in thegeneralized Shwartzman reaction by pretreatment with CoF to depleterabbits of CS and terminal components. The reasons for these disparateresults are unclear but may reflect differences in the types of endotoxinsused or the method by which the CoF was administered to the animals.The results of similar experiments by several other investigations, how-
ever, would also not be supportive of a major role for complement in thehemorrhagic lesions induced by endotoxins. Mulller-Berghaus and Loh-mann," examining the effect of infusion of S entettidis endotoxin (Boivinextract, 40 ug/kg/hr) into rabbits, were unable to detect any significantdifferences in either the hematologic or the histologic results obtainedwhen normal rabbits or rabbits congenitally deficient in C6 were com-pared. These authors concluded that C6 and terminal components werenot essential to the initiation of DIC by endotoxins. Similarly, Ulevitch etal * examined the effect of several different endotoxin preparations (E coli

542 MORRISON AND ULEVITCH American Journalof Pathology
0111: B4, Westphal, 50 Ag to 3 mg; S minnesota Re595, phenol-chloro-form-petroleum ether, 50 Ag to 3.3 mg; S marcescens, Westphal, 100 jgto 12.5 mg) on fibrin deposition in normal, C6-deficient, and CoF-treatedrabbits. Approximately 24 hours after a single injection of endotoxin,these authors observed that, although minor differences were observeddepending on the type of endotoxin administered, if anything, the ab-sence of C3 and terminal components enhanced the degree of fibrindeposition in comparison to the amount of deposition in control animals.The results of these studies did not demonstrate any differences in meanarterial blood pressure changes induced in the normal or complement-depleted rabbits after administration of endotoxin, independent of thetype of endotoxin employed.
However, in a more recent study by Ulevitch and Cochrane," using aBoivin extract of S marcescens (5.0 mg), it was shown that (in contrast tothe Westphal preparation used in the previous study) a biphasic change inmean arterial blood pressure could be elicited with this type of endotoxin,as was earlier demonstrated in dogs "' and cats." In addition, con-firming these earlier studies, the initial rapid change, but not the moregradual secondary change, could be abrogated by treatment of the rabbitswith CoF prior to administration of endotoxin. The authors further ob-served that up to 2.5 times as much of endotoxin from the identicalorganism prepared by the Westphal procedure did not induce the rapidhypotensive change but did produce identical secondary blood pressuredrops. These results underscore the significant variability in biologicactivity of endotoxins derived from a single organism by different extrac-tion procedures.
Ulevitch and Cochrane " also assessed in these experiments the effectof complement depletion on the lethal effects of the S marcescens Boivinendotoxin. They determined that intravenous injection of 5.0 mg ofendotoxin was lethal to 10 of 10 of the control rabbits and 10 of 10 of theCoF-treated rabbits. They thus concluded that C3 and terminal com-ponents did not play a significant role in the lethality in rabbits caused bybacterial endotoxins. These results contrast significantly with the earlierstudies of Brown and Lachmann." These authors, using apparently iden-tical preparations of endotoxin from the same source prepared in appar-ently the same manner and administered to the same species of rabbits,reported strikingly different results. Whereas 16 of 16 of the normalrabbits died following administration of endotoxin, it was found that 0 of12 of the CoF-treated rabbits died. These results suggest that C3 andterminal components play a critical role in manifesting the toxic proper-ties of bacterial endotoxins. The reasons for the different results obtained

Vol. 93, No.2 BACTERIAL ENDOTOXINS 543November 1978
in these two studies are unclear. Nevertheless, a third set of experimentalobservations published by Johnson and Ward 1 are relevant. These inves-tigators also examined the role of complement in endotoxin lethality inrabbits using both normal and C6-deficient animals. Following intra-venous injection of 100 gg of E coli 0111:B4 endotoxin (Boivin) intorabbits, 0 of 4 of the normal rabbits died (and only 1 of 8 of the C6heterozygotes) and 11 of 12 of the C6 homozygous deficient rabbits died.These results suggest a critical role for C6 (and perhaps terminal com-ponents) in the protection of the host against the lethal effects of bacterialendotoxins.
These completely contrasting results obtained by using dissimilar prep-arations in apparently similar experimental situations provide convincingevidence for the potentially broad spectrum of responses which may beattributable to endotoxins. They further make apparent the necessity todefine the endotoxin preparation used in a given experimental study andreiterate the potential fallacy of basing experimental conclusions on re-sults obtained with a single endotoxin preparation.
Although it seems very clear that the interaction of complement withbacterial endotoxins in vivo does, at least in part, contribute to the overallhost response to endotoxin (see also Platelets), the precise role of thisinteraction in the initiation of pathophysiologic changes leading to tissueinjury remains to be completely defined. Clearly, differences in endo-toxins from different species and from the same organism prepared bydifferent procedures, as well as inherent differences in these variousendotoxins to activate the various complement pathways, will contributeto their capacity to affect the complement system in vivo.
It is becoming increasingly clear that complement is consumed inpatients with shock due to gram-negative sepsis. Thus, McCabe I hasdemonstrated significantly decreased levels of CS in patients with shock(but not control patients or patients with bacteremia). More recent studiesby Fearon et al " and Robin et al " have confirmed these studies and, inaddition, provided evidence for alternative pathway activation leading toconsumption of complement components. These studies argue for a con-tinued effort to define the interaction of endotoxin and complement in thedevelopment of injury.
Endotoxin Complement Interactions In Vitro
Pillemer and his associates " demonstrated more than 20 years ago thatthe incubation of a number of naturally occurring microbial poly-saccharides, in particular a preparation of typhoid endotoxin, with normalhuman serum in vitro at concentrations of 1.5 to 3 mg/ml initiates the

544 MORRISON AND ULEVITCH American Journalof Pathology
inactivation of both properdin and complement component C3. Theseauthors showed further that the binding of endotoxin to properdin re-quired both complement components (presumably C3) and magnesium.Their data also suggested a prominent role for the polysaccharide portionof the endotoxin molecule in the interaction with properdin.
It had been suggested earlier,74 in part on the basis of the results of invivo experiments,78 that the anticomplementarv activity of endotoxins wasthe result of natural antiendotoxin antibodies in the serums of mostmammalian species. To probe this question, Kostka and Sterzl " exam-ined the capacitv of endotoxins to activate complement in serum fromprecolostral (IgG-free) piglets and adult pigs. Although both serums con-tained comparable levels of complement titers, they observed com-plement activation only in serums from adult pigs. The results of theirexperiments were supported by earlier in vivo experiments 78 and sug-gested a major role for antibody in the anticomplementary activity ofendotoxins.An important contribution to help resolve this question was provided by
the experiments of Muschel, Schmoker, and Webb."7 These authors dem-onstrated that the anticomplementary activity of endotoxin preparationswas unaffected in serums previously absorbed with either autologous orheterologous bacterial cells. In addition, they observed more activity inimmune serums, suggesting that the presence of antibody could contrib-ute to the anticomplementary activity of endotoxins. In contrast to theresults of Kostka and Sterzl," these authors demonstrated a significantlyenhanced sensitivity of piglet serum complement to endotoxin in com-parison with adult serums. The results of these experiments prompted theinvestigators to conclude that the anticomplementary activity of endo-toxin was not mediated by antigen-antibody complexes. Additional exper-iments demonstrated that, in terms of sensitivity to S typhosa endotoxin,human serum >>guinea pig serum >rabbit serum and that Boivin prepa-rations of endotoxin were considerably more anticomplementary thanWestphal preparations.A significant advance in the understanding of the biochemical mecha-
nism of interaction of bacterial endotoxins with serum complement camefrom a series of experiments by Gewurz, Mergenhagen, and their col-leagues." These investigators made the exciting observation that an endo-toxin preparation from Veillonella alcalescens, when incubated withguinea pig serum, promoted substantial consumption of CS without sig-nificant consumption of any of the classic pathway components Cl, 4, or2. Further studies demonstrated that large amounts of each of the sixterminal complement components C3, 5, 6, 7, 8, and 9 of guinea pig

Vol. 93, No.2 BACTERIAL ENDOTOXINS 545November 1978
serum could be consumed on interaction with endotoxic lipo-polysaccharide with virtually no consumption of Cl, 4, or 2.9 Additionalstudies by these investigators demonstrated that, unlike guinea pig serum,hamster serum did consume Cl, 4, and 2 after incubation with endo-toxin.100To address the question of the potential contribution of antiendotoxin
antibody in human serum, which might contribute to the anti-complementary activity, Gewurz et al 101 examined the effects of endo-toxins in a variety of immunoglobulin-deficient serums. These includedserums from representative subjects with developmental agammaglobuli-nemia (AGG), experimentally induced AGG, and clinical AGG. Theyconcluded from these experiments that endotoxins still possessed anti-complementary activity even in the presence of less than 2.5 X 10O mg/ml 7-globulin. However, in spite of these impressive observations, theseauthors were still reluctant to absolutely rule out the potential participa-tion of antibody.The observations demonstrating that endotoxins could consume CS and
terminal complement components without significant consumption of Cl,4, or 2 prompted several investigators to pursue this question in moredetail. Marcus et al 102 used a rabbit antiguinea pig C3 (which inhibitsEAC4 activity on C3) to demonstrate that inhibition of the CS convertaseformed by classic pathway components had no effect on the capacity ofValcalescens endotoxin-initiated C3 conversion in guinea pig serum.These authors concluded that the observed activity was ""due to anotherfactor which is presumably a component of the properdin system." Usinga different experimental approach, Frank et al,1" using guinea pig serum,confirmed the earlier observations of Gewurz, Mergenhagen, and col-leagues that endotoxins could consume CS and, later, components inguinea pig serum. Of considerable relevance, however, was the demon-stration that significant consumption of terminal components could bedemonstrated in guinea pig serum genetically deficient in C4 as well as innormal guinea pig serum. The authors concluded that these experimentsthus provided "unequivocal evidence of the function of an alternativecomplement pathway bypassing the early components." A third approachwas utilized by Gotze and Miuller-Eberhard 104 to examine this question.These authors demonstrated that a precursor protein in serum becomesenzymatically activated on incubation with naturally occurring plant orbacterial polysaccharides (or lipopolysaccharides). They further showedthat if serum was incubated with inulin (a yeast polysaccharide withanticomplementary properties similar to endotoxins) and the serum wasthen fractionated to purify this enzymatically activated protein, the iso-

546 MORRISON AND ULEVITCH American Journalof Pathology
lated protein had the capacity to directly cleave C3. The authors termedthis protein "CS proactivator" (Factor B) and suggested that activation ofthis second complement activation mechanism may be accomplished bynaturally occurring polysaccharides, including bacterial lipopoly-saccharides. These combined experiments thus provided convincing evi-dence that bacterial endotoxins did have the capacity to activate com-plement by an antibody-independent mechanism. They further allowed amechanistic explanation for the important earlier obserations of Gewurzand his collaborators,"'" demonstrating consumption of terminal com-ponents without concomitant early component consumption.The demonstration that bacterial endotoxins could activate an alterna-
tive pathway of complement prompted a number of investigators toexplore the biochemical basis for this anticomplementary activity. Theseexperiments may in part have been stimulated by the almost simultaneousdemonstration that preparations of endotoxin from rough Re strains ofbacteria, which contained almost no polysaccharide, were, nevertheless,fully capable of eliciting endotoxin responses. (See Relationship of Endo-toxin Structure to Biologic Activity.) It should be noted that the earlyexperiments of Pillemer and his associates 71,72 suggested that the poly-saccharide component of endotoxins was the dominant chemical moietyregulating anticomplementary activity. They further demonstrated thatboth magnesium and complement (CS?) were required for binding ofendotoxins to properdin. In view of these earlier studies, Mergenhagen etal 1" thus made the somewhat surprising observation that the endotoxinfrom a S minnesota (Re) mutant deficient in O-polysaccharide and hep-tose was equally as active as the endotoxin derived from the poly-saccharide-containing parent strain. These data thus suggested that largeamounts of polysaccharides were not an essential requirement for anti-complementary activity of bacterial endotoxins. However, since endo-toxins derived from mutant strains still contain small amounts of carbohy-drate, a polysaccharide requirement could not be completely excluded bythese experiments.An extensive series of experiments performed by Galanos and collabora-
tors examined the anticomplementary activity of a number of endo-toxin preparations isolated from both smooth (polysaccharide-containing)and rough (polysaccharide-deficient) strains of E coli and Salmonellaspecies. These authors observed that although all of the preparationstested had endotoxic activity, only a few had high anticomplementaryactivity. For example, the polysaccharide-containing endotoxins from Srmnnesota and S ruiA were highly anticomplementary, whereas thosefrom S milwaukee and S godesberg were virtually without activity. A

Vol. 93, No.2 BACTERIAL ENDOTOXINS 547Noveber 1978
similar spectrum of activities was obtained when a number of poly-saccharide-deficient endotoxins were examined. These experiments pro-vided additional support for the concept that the presence of 0-poly-saccharide or core polysaccharide was not an essential requirement foranticomplementary activity of endotoxins. The differences observed inactivity of the various strains were probably not the result of differentculture conditions or extraction methods since all bacterial cultures weretreated identically. However, some differences were observed with differ-ent batches of guinea pig serum, suggesting a possible role for antibody.A significant finding in the experiments of Galanos et al 106 was the
observation that preparations of isolated endotoxins, which themselveswere without detectable anticomplementary activity, nevertheless con-tained a lipid A portion, which after isolation from the native endotoxinby mild acid hydrolysis (see Relationship of Endotoxin Structure to Bio-logic Activity) was highly anticomplementary. These data suggest that thepresence of polysaccharide modulates the full expression of lipid A in itsanticomplementary activity. Although these experiments suggested thatanticomplementary activity of isolated lipid A required its being com-plexed to a soluble carrier such as bovine serum albumin, more recentexperiments by Morrison and Verroust 107 demonstrated that isolated lipidA alone under the appropriate conditions could be highly anti-complementary. In addition, both of these studies 106,107 demonstratedthat the anticomplementary activity of isolated lipid A was highly depen-dent on its degree of aggregation or solubility.The experiments of Mergenhagen et al 106 and Galanos et al,.16 demon-
strating the potent anticomplementary activity of lipid A and rough (Re)mutants, were confirmed and extended by Dierich et al."*6 These authorsshowed that lipopolysaccharides isolated from the smooth and Re mutantof S minnesota, as well as isolated lipid A, activated C3 in guinea pigserum. Endotoxin activation depended on Mg++ but not Ca++ and afactor which interacts with CoF (presumably Factor B of the alternativepathway). These data suggested that preparations of endotoxins from bothrough and smooth strains of bacteria could activate the alternative path-way independent of the presence or absence of polysaccharides in theendotoxin preparation. The available evidence therefore provided con-vincing evidence that many endotoxins had the capacity to activate thealtemative complement pathway.
However, the results of a number of experiments indicated that endo-toxins also interacted with classic pathway components. Muller-Eberhardet al 1'0 demonstrated by immunoprecipitation that several endotoxinpreparations could bind to purified human Clq. Similar results were

548 MORRISON AND ULEVITCH American Journalof Pathology
obtained by Loos et al 110 using guinea pig Clq and purified E coli 075endotoxin. These authors showed that many endotoxic preparations (par-ticularly lipid A and endotoxins from R mutants) could inhibit CT hemo-lytic function. The polysaccharide-containing endotoxins from E coli0111: B4 and S minnesota (smooth) were striking exceptions. The bindingof cr to sensitized erythrocytes prior to addition of endotoxin markedlyreduced the inhibition of hemolytic activity and yielded additional sup-port for a Clq binding site. These data thus provided evidence that, inaddition to alternative pathway activation, endotoxins may also activatethe classic pathway by an interaction with Cl. This concept receivedadditional support from the results published by Snyderman and Pike,l"'who measured the anticomplementary activity of Proteus vulgaris endo-toxin in the presence of chelators of Mg++, or Mg++ and Ca++. Theresults of these experiments suggested that maximal anticomplementarvactivity of endotoxins required both Mg++ and Ca++ and, by inference,an intact classic pathway.The first experiments to directly examine this question were performed
by Lachmann and Nicol.lU These authors compared the anti-complementary activity of isolated lipid A and endotoxin prepared from asmooth strain of E coli 026: B6. The results of these experiments sug-gested that lipid A could consume significant amounts of C1, 4, and 2 inhuman serum. In addition, lipid A could convert CS and factor B innormal but not in C(2-deficient serum. The polysaccharide-containing026: B6 endotoxin, in contrast, consumed minimal Cl, 4, and 2 and wasequally efficient at converting C3 and Factor B in both normal and C2-deficient serum. These data thus suggested that, in contrast to the resultsof Dierich et al,106 lipid A (and endotoxins from rough mutants) werealmost exclusively classic pathway activators and that alternative pathwayactivation was a property of polysaccharide-containing endotoxin prepara-tions.The experiments of Morrison and Kline 113 confirmed the observations
of Lachmann and Nicol 114 and, in addition, provided evidence to supportthe concept that altemative pathway activation does not require the activeparticipation of the lipid A region of the endotoxin molecule but rather isonly a function of the polysaccharide. In part, these experiments utilizedpolymyxin B which was found to completely abrogate lipid-A-mediatedclassic pathway activation 114 but had virtually no effect on poly-saccharide-mediated alternative pathway activation. The demonstrationthat lipid A and endotoxin from the Re 595 mutant of S minnesotarequired C2 to convert C3 and did not convert Factor B, however, wouldalso not support the concept of Dierich et al 106 that CS conversion by lipid

Vol. 93, No.2 BACTERIAL ENDOTOXINS 549November 1978
A was abrogated in Factor-B-depleted serum. Absorption experimentsusing whole bacteria as well as complement activation in agammaglobuli-nemic serum confirmed the earlier studies of Muschel et al 97 and Gewurzet al 101 and provided further evidence that natural anti-lipid-A antibodydid not play a prominent role in classic pathway activation by lipid A orendotoxins from rough mutants.
Recent experiments by Cooper and Morrison 115 have extended thestudies of Loos et al 110 to demonstrate directly that lipid A and roughendotoxins can bind to and initiate the activation of purified human Cl.In contrast to the earlier study, significant binding of endotoxin to pre-cursor Cl as well as CT was restricted to preparations of endotoxin fromrough strains and lipid A. The minimal binding of Cl to polysaccharide-containing endotoxin preparations was significantly inhibited by the pres-ence of LAP, suggesting that the binding was occurring through the lipidA region. As demonstrated earlier,', 1o2 binding was shown to occur viathe Clq portion of the Cl molecule. Of significance, attachment of thelipid A and rough endotoxins to precursor Cl (reconstituted from highlypurified Clq and proenzymes Cln and Cls) was demonstrated to be abiologically meaningful event because it led to the enzymatic conversionof proenzyme Cls and Cls. These studies thus demonstrated in a purifiedsystem that lipid A had the capacity to directly and efficiently activate theclassic pathway at the Cl step by an antibody-independent mechanism.The recent experiments by Galanos and Luderitz 116 have extended
their earlier examination of the physical and chemical state of lipo-polysaccharides required for complement activation. These studies em-ployed electrodialysis and subsequent conversion to uniform salt forms tocreate preparations of endotoxin with a broad spectrum of sedimentationcoefficients (and molecular weights). Polysaccharide-containing prepara-tions of endotoxin were found to lose anticomplementary activity if con-verted to the low-molecular-weight form with triethylamine. In contrast,endotoxins from Re mutants and lipid A were equally anticomplementarv,independent of salt form. Of interest, preparations of endotoxins fromintermediate rough strains of bacteria had low anticomplementary activ-ity under all conditions examined. Since, as was shown by Morrison andKline,11s the polysaccharide is responsible for altemative pathway activa-tion, these data might suggest that high-molecular-weight aggregates arerequired for efficient assembly of multiple altemative pathway conver-tases. Classic pathway activation by lipid A and Re mutants, in contrast,probably requires only binding of Cl.'11' The inability of endotoxinsfrom intermediate rough strains to activate complement would then beexplained by an insufficient amount of polvsaccharide to activate the

550 MORRISON AND ULEVITCH American Journalof Pathogy
alternative pathway. However, the presence of some polysaccharidemight be sufficient (as was earlier demonstrated for endotoxin fromsmooth strains 10S,113) to modulate the ability of lipid A to bind Cl andactivate the classic complement pathway.
In summary, the results of extensive experiments over the past 2 dec-ades have clearly established that the lipopolysaccharide portion of bacte-rial endotoxins plays a major role in the activation of serum complementby two antibody-independent mechanisms. The first involves the lipid Aregion of the LPS molecule which binds directly Cl, leading to anantibody-independent activation of the classic pathway. The presence ofpolysaccharide, which activates the alternative pathway by a lipid-A-independent antibody-independent mechanism, modulates the expressionof lipid A binding and activation of Cl. Endotoxins from semiroughstrains of bacteria have only low anticomplementary activity. Futureexperiments will be required to investigate the original postulate of Pille-mer 71-72 that "the spatial configuration of the sugar residues in the LPSmolecule is of importance in the expression of activity." The estab-lishment of such a postulate may clarify the observations of Galanos etal 1'* on the spectrum of anticomplementary activities of various endo-toxins prepared by identical procedures and provide a molecular basis foralternative pathway activation. The potential effect of lipid-A-associatedprotein on the expression of both alternative and classic pathway activa-tion should be explored. In this respect the observations of Muschel et al 7on the differential anticomplementary activity of endotoxins prepared byBoivin or Westphal extraction as well as the observation of Ulevitch andCochrane " on the similarly striking pronounced differences in com-plement-mediated responses in vivo are certainly worthy of further exper-imentation.
- -
The observations of Sanarelli 117 reported in 1924 and the similar reportof Shwartzman in 1928 11ll,19 of the pathologic effects of two properlytimed injections of culture filtrates from gram-negative organisms firstdemonstrated what we now know to be the extraordinary ability ofbacterial endotoxins to produce tissue injury through initiation of coagu-lative changes. These changes are generally referred to as disseminatedintravascular coagulation (DIC). Subsequent studies in the ensuing 5decades (reviewed in References 120 and 121) have extended these obser-vations and have resulted in two experimental models actively studiedtoday: the localized and the generalized Shwartzman phenomenon. Inboth of these experimental models, the formation of occlusive fibrin

Vol. 93, No. 2 BACTERIAL ENDOTOXINS 551November 1978
thrombi in the vessels of the affected tissues has been well documented. Itis now clear that even following a single intravenous injection of endo-toxin, coagulative changes are initiated, resulting in the deposition offibrin in a variety of tissues, including the lungs, liver, and spleen. It islikely that the underlying biochemical mechanisms of coagulativechanges following one or two properly spaced injections of endotoxin areidentical.One of the major unanswered questions regarding the mechanism of
endotoxin-induced coagulative changes involves a determination ofwvhether the activation of the prothrombin molecule occurs pre-dominantly by either the intrinsic or extrinsic coagulation pathway. Thus,investigations of the effect of endotoxin on Hageman factor activation andon the production of cell-derived procoagulant activities have been vigor-ously pursued. During the past several decades, numerous advances havebeen made in understanding the molecular biochemistry of blood coagu-lation, and, as a consequence, our knowledge of biochemical mechanismspotentially relevant to endotoxin-induced DIC has increased markedly.Based on our understanding of blood coagulation, the most likely siteswhere endotoxin might act to initiate coagulation are depicted in Text-figure 8.The molecular mechanism of activation of Hageman factor has recently
been investigated by Griffin and Cochrane '22 and new concepts con-cerning this process have been advanced. In particular, the critical role ofkallikrein and high-molecular-weight kininogen in Hageman factor acti-vation has been appreciated. Several reviews summarizing recent findings
Intrinsic Coauilation PathAy Extrirsic Coaguation Pathy
Hageman Factor (XII) Medator Cells (eg, monocyti)
-------END OTOXIN
Factor Xl Factor Vil
Act,vated Hpmnan _ *_ Release of Tisue Factor (TF)Factor Prottrombin
Factor Xla VI l-i- TF
Thrombin
Fibrinoq.. - w Fibrin
TEXT-FICGRE S-Interaction of endotoxin with the coagulation system.

552 MORRISON AND ULEVITCH American Journalof Pathology
concerning the chemistry and biology of the Hageman-factor-activatedpathways have appeared.mJl
In addition to the effects of Hageman factor on initiation of the intrinsiccoagulation system, several other important activities may be produced byactive Hageman factor. First, this enzyme can activate plasma pre-kallikrein to form kallikrein, a proteolytic enzyme capable of generatingbradykinin as well as activating plasminogen (reviewed in Reference 124).Second, it has recently been shown by Keisel et al 25 and Radcliffe et althat activated Hageman factor is capable of directly activating Factor VII.Although the physiologic significance of this is unknown, this newlyidentified activity of Hageman factor might provide a marked enhance-ment for endotoxin-induced coagulative changes. Investigation of thismechanism warrants study in in vivo models.The interaction of endotoxin with cells has also been suggested to result
in the elaboration of substances with procoagulant activity. In particular,platelets and blood leukocytes have been implicated in this event.The effects of endotoxin on platelets which might contribute to the
initiation of coagulative changes in vivo have been considered in thesection on endotoxin-complement interactions in vivo and will not bedescribed here. Recent studies of the mechanism of endotoxin-inducedDIC have focused on defining the mechanism(s) by which endotoxininjection results in activation of the prothrombin molecule. Specifically,the mechanism of activation of Hageman factor and the mechanism ofendotoxin action to initiate tissue factor production by blood leukocyteshas been studied in detail.
Intrinsic Coagulation Pathway-Hageman Factor Activation
Endotoxin Activation In Vivo. Evidence to support the proposal thatHageman factor is activated by endotoxin in vivo derives mainly fromindirect measurements believed to reflect Hageman factor activation.Since active Hageman factor is capable of activating prekallikrein, whichin turn can generate bradykinin from plasma kininogen, observations ofdecreased levels of plasma prekallikrein and kininogen or, alternatively,increased levels of bradykinin have been utilized to indicate Hagemanfactor activation. In this section, experimental observations derived fromstudies of patients with gram-negative bacillary infections will also becited as evidence to suggest that Hageman factor may be activated byendotoxin. Therefore, studies in humans 12 and in the subhuman pri-mate,"-' showing that blood bradykinin levels increase following theintravenous injection of endotoxin, have suggested the generation ofactivated Hageman factor by endotoxin. Kimball et al I2 showed that

Vol. 93, No.2 BACTERIAL ENDOTOXINS 553November 1978
injection of as little as 3 ng/kg of S abortus equi endotoxin into humansproduced a significant elevation in blood bradykinin levels within 30 to 60minutes after injection. These authors suggested that bradykinin mayserve as a trigger vasodilator in endotoxin shock. Other studies haveshown that in dogs 130 and in rabbits 131 total blood kininogen also de-creases following endotoxin injection.
Reichgott and his colleagues 132 examined the role of the lipid A regionof an E coli endotoxin (Westphal) on a variety of hemodynamic andhematologic changes produced in the rhesus monkey. To do this theytreated the endotoxin with 0.02N acetic acid at 100 C to prepare lipid A.The carbohydrate remaining in the supernatant was purified on charcoalto prepare two polysaccharide fractions, one containing 6.3% of total fattyacids (PS1) and one with less than 0.5% of the total fatty acids (PS2). It is ofinterest to note that the native endotoxin and PS1 both produced hypoten-sion and decreased peripheral resistance and, equally important, thatincreased levels of plasma bradykinin could be detected. When PS2 wasinjected, neither the hemodynamic changes characteristic of endotoxininjection nor increased bradykinin levels were observed. Thus, the poten-tial importance of the hpid A region of the endotoxin molecule in eventsleading to Hageman factor activation was suggested.
Experiments reported by Skjorten and Evensen 133 demonstrated that Smarcescens endotoxin did not induce DIC in Norwegian breed chickens,although infusions of tissue thromboplastin did result in accumulation offibrin in pulmonary vessels. Because it had been proposed that fowl aredeficient in Hageman factor,'" these results have also been cited asevidence for a critical role for Hageman factor in endotoxin-induced DIC.
Further support for the activation of Hageman factor derives fromclinical studies of patients with gram-negative bacillary infections. Masonand Colman 136 observed decreased levels of plasma Hageman factor andprekallikrein in patients with DIC in whom endotoxemia was thought tobe responsible for initiating the coagulative changes.
Robinson et al also provided evidence to suggest that Hageman factor isactivated by endotoxin released from bacteria.1" Patients undergoingtransurethral resection or cystoscopy were studied, and levels of bloodendotoxin were measured prior to and within 6 hours of these surgicalprocedures. In patients from whom positive endotoxin assays and evi-dence of gram-negative bacteremia were obtained, decreased levels ofplasma prekallikrein were observed. Decreased vascular resistance wasassociated with the appearance of the positive endotoxin test and thedecrease in plasma prekallikrein levels.
Thus, experimental evidence from studies of animal models as well as

554 MORRISON AND ULEVITCH American Journalof Pathology
clinical studies suggest that Hageman factor activation results from ex-posure of the host to endotoxin. However, direct quantitative measure-ments of Hageman factor activation either in human disease in whichendotoxin release from gram-negative bacteria is believed to initiate DICor in experimental models of endotoxin-induced coagulative changes havenot been reported. Direct biochemical measurements of Hageman factoractivation or activation of its substrates are essential to fully evaluate therole of these proteins in endotoxin-induced injury.
Evidence to suggest that initiation of coagulation through the intrinsiccoagulation system is not essential for endotoxin-induced DIC in a modelof the generalized Shwartzman reaction was reported by Shen et al.13" Forthis study, circulating coagulation Factor VIII (intrinsic pathway) inrabbits was depleted by prior injection of a cross-reactive human anti-serum against Factor VIII. Then a comparison of a variety of hematologicparameters was made between normal and Factor-VIII-depleted rabbitsinjected with a Boivin preparation of E coli 0111:B4 endotoxin. Thehematologic parameters studied included ""I-fibrinogen survival; fibrin-ogen levels; Factors VIII, VII, and V levels; white blood cells; platelets;and hematocrit. The kidneys were also examined histologically for thedeposition of fibrin. The results of these studies showed that Factor VIIIdepletion did not alter the endotoxin-induced consumption of fibrinogenand that the small decrease in Factors VII and V produced by endotoxinappeared to be the same in Factor-VIII-depleted and normal rabbits.These studies showed that fibrin deposition could be detected in thekidneys of Factor-VIII-depleted animals. Thus, from these results itwould appear that selective impairment of the intrinsic clotting system byFactor VIII depletion is not sufficient to prevent endotoxin-induced coag-ulative changes. However, since this study was performed using cortisone-treated rabbits to "prepare" for the generalized Shwartzman reaction, theprecise relationship of these conclusions to experiments employing eitherone or two properly timed injections of endotoxin remains to be estab-lished.
Endotoxin Activation In Vitro. Evidence to support a role for Hagemanfactor in endotoxin-induced coagulative changes has also derived from theresults of in vitro studies. Rodriguez-Erdmann 136 reported that the addi-tion of endotoxin markedly shortened the whole blood clotting time insiliconized glass as well as that of platelet-poor plasma. Direct activationof Hageman factor in plasma leading to bradykinin production was alsosuggested by the experiments of Nies and Melmon.' They reported thatthe addition of E coli 0127:B8 endotoxin to a final concentration of 200Ag/ml to either rhesus monkey or rabbit plasma reduced the plasma

Vol. 93, No.2 BACTERIAL ENDOTOXINS 555November 1978
kininogen level by at least 40% within 10 minutes. These authors sug-gested that kininogen was depleted through activation of Hageman fac-tor. However, because of the high affinity of kininogen for polyanionicsurfaces, the possibility that the kininogen was merely bound to theendotoxin and made inaccessible for the assay system cannot be excluded.In these studies, kininogen levels were determined by the addition oftrypsin and the total amount of bradykinin liberated.More recently, the studies of Morrison and Cochrane 1" have provided
insight into the mechanism of Hageman factor activation by endotoxins.These investigators demonstrated two important points using highly puri-fied preparations of human Hageman factor and endotoxin. First, activa-tion required a complex to be formed between the endotoxin and theHageman factor molecule. Second, the lipid A region of the endotoxinmolecule was required for activation and most likely represents the site ofbinding on the endotoxin for the Hageman factor. Several different exper-imental approaches were employed by Morrison and Cochrane 139 tosupport these conclusions. Evidence for the role of lipid A was obtained instudies which demonstrated that the activity of endotoxin preparationsfrom E coli 0111 :B4 (Westphal) was dependent on their content of lipid A.Further, evidence for the importance of lipid A was obtained from experi-ments using purified lipid A which showed that isolated lipid A didactivate Hageman factor directly. Additional experiments were performedwhich examined the effect of the removal of lipid A from the endotoxin onits subsequent ability to activate Hageman factor. In a kinetic experiment,a correlation between the presence of free KDO produced by mild acidhydrolysis (O.O1N H2SO, 100 C) of the endotoxin (see Relatiownship ofEndotoxin Structure to Biologic Activity) and the loss of the ability of thisendotoxin to activate Hageman factor was shown.
In terms of our current understanding of Hageman factor activation, itis likely that the endotoxin provides an appropriate negatively chargedsurface required to facilitate activation. Presumably, the negativelycharged phosphate residues of the endotoxin are important in providingthis charge density, and, thus, the polyanionic character of the endotoxinmolecule contributes significantly to this biologic effect. The importanceof the phosphate groups could be tested by examining the ability ofendotoxin from Chromobactenum violaceum to activate Hageman factorsince the phosphate groups of the endotoxins from this organism havebeen shown to be largely substituted with either 4-aminoarobinose orglucosamine. This endotoxin might also provide an alternative means ofevaluating the role of Hageman factor activation in the development ofendotoxin shock in the experimental animal models currently studied.

556 MORRISON AND ULEVITCH American Journalof Pathology
Extrinsic Coagulation Pathway-Tissue Factor Production
Endotoxin Activation In Vivo. A direct effect of endotoxin on theactivation of any of the known proteins of the extrinsic coagulationpathway has yet to be demonstrated. However, numerous observationshave shown that levels of coagulation Factor VII decrease following theinjection of endotoxin, indicating activation of the extrinsic coagulationpathwav (reviewed in References 120 and 121). The link between endo-toxin-induced coagulative changes and activation of the extrinsic coagu-lation system appears to be a cell-derived procoagulant; evidence tosupport the involvement of cells and the extrinsic coagulation pathwaywill be described below.
Approximately 30 years ago, Becker 140 demonstrated that the localizedShwartzman reaction could be suppressed by a number of agents that wenow know to produce marked leukopenia. It was, however, not until thestudies of Stetson and Good 141 and Thomas and Good 142 that the essentialrole of peripheral blood leukocytes in endotoxin-induced coagulativechanges was proposed. A comprehensive discussion of the role of thegranulocyte in the development of endotoxin-induced coagulativechanges has recently been presented by Horn.143
Compelling evidence for the essential role of the leukocyte in endo-toxin-induced coagulative changes in the generalized Shwartzman reac-tion in rabbits has derived from recent experiments of Bohn and Mflller-Berghaus.1" These investigators employed a Boivin preparation of Senteritidis endotoxin to examine the potential participation of plateletsand leukocytes in endotoxin-induced coagulative changes in the rabbit.For these experiments they administered busulfan to deplete rabbits ofperipheral blood platelets as well as leukocytes and demonstrated thatsuch treatment also inhibited the endotoxin-induced coagulative changes.Thev then examined the effect of transfusion of either platelets or perito-neal leukocvtes (see Platelets) on the development of endotoxin-inducedcoagulative changes in the busulfan-treated rabbits. These authorsshowed clearly that infusion of platelets did not render the busulfan-treated animals susceptible to development of microthrombi in thekidneys after endotoxin injection. In contrast, infusion of peritoneal leuko-cvtes did restore the susceptibility of the thrombocytopenic and gran-ulocytopenic rabbit to the coagulative changes induced by endotoxin. Ofinterest in these studies was the demonstration of the occurrence ofmicrothrombi in both lungs and spleen of busulfan-treated animals. Al-though not discussed by these investigators, these results suggest thateven in the absence of blood leukocvtes, endotoxin may trigger intra-vascular coagulation. Possible mechanisms might include endotoxin-in-

Vol. 93, No. 2 BACTERIAL ENDOTOXINS 557November 1978
duced Hageman factor activation or, alternatively, stimulation of tissue-fixed cells such as macrophages or other lymphoid cells by endotoxin toproduce a procoagulant activity. The identification of tissue-fixed cellswhich may participate in the initiation of endotoxin-induced coagulativechanges should prove to be an interesting area of research in the future.More direct evidence to suggest a role for the extrinsic coagulation
system in the development of endotoxin-induced DIC derives from exper-iments of Garner and Evensen.146 Using normal dogs and dogs with ahereditary deficiency of Factor VII, these investigators compared a varietvof hemodynamic, hematologic, and coagulative changes induced by aninfusion of 2.5 mg/kg of E coli 0111:B4 endotoxin. These investigatorsstated that histologic examination of tissues from endotoxin-treated nor-mal and Factor-VII-deficient dogs demonstrated significantly greaterdeposition of fibrin in tissues of normal dogs. All of the major organsshowed multiple thrombi and focal necrosis in the normal dogs; thesechanges were reported to be minimal or absent in the Factor-VII-deficient dogs. However, in a seemingly paradoxic observation, coagu-lation assays revealed few differences between normal and Factor-VII-deficient dogs when a variety of parameters were measured, including thekaolin-cephalin clotting time; fibrinogen levels; and Factor V, II, and Xlevels. Additionally, the Factor VII levels decreased markedlv in thenormal dogs given endotoxin. Thus, it would appear that the activation ofFactor VII following endotoxin injection plays a key role in the develop-ment of endotoxin-induced DIC, perhaps by accelerating the rate ofthrombin activation, although the data presented suggest that activationof the intrinsic coagulation system may also occur despite the absence ofFactor VII.Endotoxin Activation In Vitro. During the past 10 years, an understand-
ing of the molecular biochemistry of endotoxin-induced DIC via theblood leukocyte has emerged. Some of the experiments which have lead toour understanding of these events will be described.Niemetz and Fani 1" first demonstrated the marked ability of endotoxin
to enhance the normally minimal procoagulant activity of rabbit perito-neal leukocytes. Subsequently, Lerner et al 147 showed that human bloodleukocytes also possessed a minimal level of procoagulant activity, and theamount of procoagulant activity could be significantly enhanced by theaddition of endotoxin to the in vitro incubation mixtures. In these latterexperiments, a preparation of Serratia marcescens endotoxin was em-ployed. Garg and Niemetz I" subsequently showed that the rabbit perito-neal macrophage, elicited by prior injection of sterile starch, could bestimulated by endotoxin to produce tissue factor procoagulant activity

558 MORRISON AND ULEVITCH American Journalof Patogy
and, on a per cell basis, produced significantly more activity than rabbitperipheral blood leukocytes. These experiments were performed with anE coli 026:B6 endotoxin. From these and other studies 149 it was clear thatleukocytes could be stimulated by endotoxin to produce a procoagulantactivity which might be important in the pathogenesis of endotoxin-induced DIC.
Recent studies of the leukocyte procoagulant activity have focused ontwo problems: a) the identification of the cell responsible and b) a defini-tion of the endotoxin structural requirements for the stimulation of tissuefactor. In terms of the former, studies of purified populations of humanperipheral blood cells indicated that the monocyte was the principalsource of the endotoxin-induced procoagulant activity. Rivers et al 150fractionated human peripheral blood leukocytes by differential centrifu-gation techniques, finally separating lymphocytes and monocytes in 28%bovine serum albumin (diluted in RPMI medium) and centrifuged at844g for 30 minutes. The lymphocyte and monocyte cell fractions wereobserved to contain no polymorphonuclear leukocytes, and monocyteswere quantitated by neutral red uptake. Incubation of the purified cellsuspensions with 10 ug/ml of a Boivin preparation of E coli 0111:1B4endotoxin demonstrated that procoagulant activity was only generated bymonocyte pool. These authors concluded that previous studies of endo-toxin-induced tissue factor in lymphocyte suspensions and in polymorpho-nuclear cell suspensions could be explained by the presence of less than1% monocyte contamination.
Hiller et al 151 also demonstrated the critical role of the blood monocytein endotoxin-induced tissue factor generation. These authors employedFicoll/Urovision density centrifugation to separate peripheral blood lym-phocytes, monocytes, and granulocytes and examined the capacity of apreparation of S entenitidis endotoxin to induce tissue factor production inthese cell populations. These authors conclusively showed that the bloodmonocyte was the source of tissue factor when endotoxin was added to amixed leukocyte population. They demonstrated that the endotoxin-in-duced tissue factor activity derived from a cell density of 10 monocytes/,lexceeded that produced by a purified preparation of 10,000 granulocytes/Al. They further showed that within 3 hours after endotoxin addition,striking morphologic changes in monocytes occurred. These changes werecharacterized by the appearance of enlarged lysosomes and were consis-tent with previous suggestions that the tissue factor was derived from thecell lysosomes. Other studies by Rickles et al 152 have also suggested thatthe endotoxin-induced procoagulant activity of human blood leukocytesresides in the mononuclear cell fraction. However, in these studies no

Vol. 93, No. 2 BACTERIAL ENDOTOXINS 559November 1978
attempt was made to separate lymphocytes and monocytes. In a sub-sequent publication, Rickles et al 15 stated, without presenting data, thatthe human monocyte was the cell primarily responsible for endotoxin-induced tissue factor release but that human B lymphocytes were alsoactive in this regard. The human T lymphocyte appears not to be capableof producing tissue factor.'"
Studies by Rickles et al 15 and by Niemetz and Morrison 155 clearlydemonstrated that activation of tissue factor by endotoxin was associatedwith the lipid A region of the endotoxin molecule. Rickles et al 153 em-ployed endotoxin from wild type and both Re and Rc mutants of Styphimurium as well as isolated lipid A to show that lipid A was theessential structural feature of endotoxin for tissue factor activation. Theseauthors also provided evidence to support the essential role for lipid A inexperiments which examined preparations in which the lipid A was modi-fied. Either mild alkaline hydrolysis which removed acyl-linked fatty acidsor the addition of polymyxin B (see Relationship of Endotoxin Structure toBiologic Activity) both markedly reduced the ability of endotoxin or lipidA to activate mononuclear cells to produce tissue factor.-Niemetz and Morrison 1" utilized endotoxin isolated from the smooth
strains of E coli 0111:B4 or E coli 026:B6 from the rough strains E coli0111:B4J5 and S minnesota R595 and isolated lipid A to examine the roleof lipid A in the activation of tissue factor in human and rabbit peripheralblood leukocytes. A comparison of the different preparations indicatedthat free lipid A could activate blood leukocytes to produce tissue factorbut also showed that the presence of carbohydrate side chains couldmodulate the lipid-A-induced activation. For example, endotoxin from Ecoli 026:B6 was more active than isolated lipid A or endotoxin from E coli0111: B4, both of whic-h contain a greater percentage by weight of lipid A.The precise role of the carbohydrate in initiating tissue factor productionis unknown. Compelling evidence for the role of lipid A in the productionof tissue factor derived from two other experiments presented by Niemetzand Morrison. They showed that mild alkaline hydrolysis of endotoxinmarkedly reduced the ability of endotoxin to stimulate tissue factor pro-duction. Further evidence to support a critical role for lipid A was ob-tained in experiments in which polymyxin B was observed to markedlyinhibit the ability of lipid A to induce tissue factor production. Finally,experiments employing the native protoplasmic polysaccharide isolatedfrom E coli 0113,155 a polysaccharide with a carbohydrate compositionsimilar to LPS but devoid of any lipid A, showed that this material did notstimulate the production of tissue factor from rabbit peripheral bloodleukocytes.

560 MORRISON AND ULEVITCH American Journalof Pathology
Niemetz and Morrison also showed that alkali-treated endotoxin from Ecoli 0111: B4, when added simultaneously with or 1 hour after the additionof E coli 026:B6 endotoxin, would block the production of tissue factor.However, the addition of the alkali-treated endotoxin 3 hours after theaddition of 026:B6 endotoxin failed to block the induction of tissue factoractivity. These data suggested that events occurring during the first 2hours after addition of endotoxin commit the cells to production of tissuefactor. The ability of alkaline-treated endotoxin to block these eventsshould provide a useful tool to examine the interactions of endotoxins withcells, particularly at the cell membrane level, which lead to the initiationof tissue factor synthesis. The striking morphologic changes in humanmonocytes treated with endotoxin described by Hiller et al 151 at 3 hoursafter endotoxin stimulation also is good evidence for early changes in-duced by endotoxin prior to tissue factor production. Thus, future studiesemploying purified monocyte populations and endotoxin preparationswith different chemical characteristics should provide information aboutthe biochemical events occurring after the interaction of endotoxins withthe cell membrane which trigger the mononuclear cell to produce tissuefactor.
Ceular ation SystemsPlatelets
Recent experiments by a number of investigators have clearly estab-lished the profound influence which bacterial endotoxins can have on theplatelets of a number of species. These effects can be shown to occur bothin vivo and in vitro. The mechanisms of the platelet-endotoxin interac-tion, however, have been demonstrated to depend critically on the pres-ence or absence of immune adherence sites on the platelet membrane. Inthis respect, the platelets from rat, rabbit, dog, and guinea pig appear tobe much more responsive to endotoxins than those from primates sincethese latter cells lack immune adherence receptor sites. Nevertheless,responses of primate (and in particular human) platelets to endotoxinshave been reported by several investigators.
Platelet responses are usually characterized by aggregation, or clump-ing, and release of platelet constitutents, including ADP, platelet factor 3,and vasoactive amines, such as histamine and serotonin. The releaseresponse of platelets may be accomplished by either a lytic reaction inwhich both granule and cytoplasmic constituents are liberated or by asecretory response in which the integrity of the platelet membrane ismaintained and only granule constituents are released. In the lytic re-

Vol. 93, No. 2 BACTERIAL ENDOTOXINS 561November 1978
sponse of platelets to endotoxins, a prominent role of serum complementhas been suggested. In either case, however, the release of intracellularplatelet constituents initiated by endotoxins may contribute significantlyto the pathophysiologic effects of endotoxins, including hypotension re-sulting from increased vascular permeability and disseminated intra-vascular coagulation resulting from activation of the extrinsic clottingsequence. (See Coagulation Systems.) A critical evaluation of plateletfunction following the administration of endotoxins has, therefore, been aprimary objective of numerous investigators over the past 2 decades andwill be covered in this section.Endotoxin-Platelet Responses In Vivo
It has been recognized for some time that bacteria can initiate profoundalterations in the levels of circulating platelets. However, the extensiveexperiments of Stetson 157 first provided a firm basis for the concept of theparticipation of platelets in the pathophysiologic effects of endotoxins inexperimental animals. In these experiments, histologic examination oftissues documented the deposition of leukocyte-platelet thrombi withinsmall blood vessels following endotoxin administration to rabbits. Fromthese data it was postulated that this cellular accumulation might be thecause of hemorrhage and necrosis in the tissues. The results of studies byWeil and Spink 158 and McKay and Shapiro,'" demonstrating significantdrops in the levels of circulating platelets within 5 minutes to 1 hourfollowing the intravenous injection of endotoxin into dogs and rabbits,respectively, lent support for this concept. Subsequent experiments byCohen et al,160 using 51Cr-labeled platelets in dogs, demonstrated that theresponse to endotoxins could be biphasic. These authors observed a rapiddrop in circulating platelets, a partial return to the circulation, followedby a second drop in platelet levels. Experiments by Davis 161 and Spielvo-gel 162 provided electron micrographic evidence that the rapid plateletdrops in rabbits following administration of E coli endotoxin was accom-panied by large numbers of altered platelets. These platelet aggregateswere demonstrated to localize in pulmonary and hepatic capillaries, andextensive platelet destruction was evident. The latter study162 also showedthe particulate endotoxin in close association with the degranulated rabbitplatelets.
These suggestions of endotoxin-platelet interaction in vivo, based onthe experiments showing alterations in numbers of circulating plateletsand accumulation in tissues, were supported by other experiments demon-strating measurable free levels of platelet constituents. Thus, Shimamotoet al 16 and Davis et al,16 16 using dogs, demonstrated that simultaneouswith the rapid fall in platelets following injection of endotoxin was an

562 MORRISON AND ULEVITCH American Joumalof Pathology
equally rapid rise in the concentration of plasma serotonin levels, al-though the time course varied somewhat in these studies. In at least one ofthese studies,'" lethality was roughly proportional to platelet and sero-tonin changes; however, the authors cautioned that there was not neces-sarily a correlation between the two events and that other factors un-doubtedly also play a role in contributing to endotoxin lethality. Horowitzet al I" were able to show a transient increase in platelet factor 3 levels inrabbits given endotoxins, which correlated with the rapid disappearanceof platelets from the circulation. The authors' demonstration that theplatelet factor 3 so generated was associated with platelet-poor plasma ledthem to conclude that, on injection of endotoxin, platelet factor 3 wastransferred from the platelet to the plasma and was then either rapidlyinactivated or cleared from the circulation. The results of these experi-ments provide convincing evidence in support of the extensive in vivoeffects of bacterial endotoxins on circulating platelets.The relative contribution of these various platelet responses to the
injury produced by endotoxins has, however, been more difficult to accu-rately assess, and several approaches have been employed. In one ap-proach, several investigators have employed purified platelet factor 3 as asubstitute for the second injection of endotoxin in the induction of theShwartzman reaction. Rodriguiz-Erdmann 167 infused purified plateletfactor 3 into rabbits previously treated with Thorotrast (which has earlierbeen shown by Good and Thomas 166 to substitute for the first of twoendotoxin injections in the induction of the generalized Shwartzmanreaction) and demonstrated lesions similar to those normally obtainedwith the provocative endotoxin injection. In contrast to these results,Muiller-Berghaus et al,.i using a number of procedures to "prepare''rabbits or rats for the Shwartzman reaction, were uniformly unable toprovoke a response with a secondary injection of either purified bovineplatelet factor 3, rabbit brain tissue extract, or soya bean phospholipids.Differences between these two studies may in part be due to the sourceand amount of platelet factor 3 used in the experiments, although itshould be pointed out that in the latter study the amounts of plateletfactor 3 used in the experiments were three times the amount usuallyavailable in the normal rabbit. Mfiller-Berghaus et al concluded that, inaddition to platelet aggregation and release of platelet factor 3, otheractions of endotoxin were essential for the production of the generalizedShwartzman reaction.A somewhat different approach was taken by Evans et al 170 to assess the
role of platelets in lethal endotoxin shock in rabbits. These authors,utilizing sulfinpyrazone as an inhibitor of platelet aggregation and release

Vol. 93, No. 2 BACTERIAL ENDOTOXINS 563November 1978
of ADP, demonstrated a significant decrease both in the immediate hypo-tension and in the platelet disappearance compared with untreated rab-bits following endotoxin administration; the ultimate effect of the inhib-itor on endotoxin-induced lethality was, however, not indicated in thisstudy. These authors concluded from these studies that platelet aggrega-tion was of importance in the genesis of endotoxin shock and that endo-toxin-stimulated platelet responses involved the release of ADP.
As an alternative means of ascertaining the potential contribution ofplatelets to endotoxin-induced injury, a number of investigators havedepleted experimental animals of circulating platelets prior to administra-tion of endotoxins. Levin and Cluff 71 utilized guinea pig antirabbitplatelet antibody to deplete selectively rabbits of circulating platelets.The antiplatelet antiserum had no effect on circulating white blood cellsnor did unrelated antigen-antibody complexes affect their results. Theseauthors found that the induced thrombocytopenia did not inhibit thecutaneous hemorrhagic lesion of the localized Shwartzman reaction. Ad-ditional experiments by one of these investigators 172 using Thorotrast-treated rabbits demonstrated that, although anticoagulants were effectivein preventing the renal injury, depletion of platelets had no effect. Theseexperiments led the authors to conclude that platelets were not primarilyinvolved in the tissue damage induced by endotoxins.A similar experimental approach, but with some apparently significant
differences, was used by Margaretten and McKay 173 to examine the roleof the platelet in the endotoxin-induced generalized Shwartzman reactionin the rabbit. These authors used two intravenous injections of E coliendotoxin (200 ug) spaced 20 hours apart. Heparin was given 1 hour priorto antiplatelet antibody and 5 hours before the second injection of endo-toxin. (The anticoagulant was utilized to prevent the intravascular coagu-lation induced by the injection of platelet antiserum.) Under these cir-cumstances, none of the thrombocytopenic rabbits developed glomerularcapillary thrombi. In contrast, 6 of 10 of the control group of rabbits(given antiserum previously absorbed with rabbit platelets) developedlesions characteristic of the Shwartzman reaction. The authors concludedfrom these studies that platelets were essential for the development of thegeneralized Shwartzman reaction.A third experimental system also using antirabbit platelet antibody was
employed by Muiller-Berghaus and Kramer 174 to examine the role ofplatelets in the precipitation of soluble circulating fibrin by endotoxin. Forthese experiments, circulating platelets were depleted using goat anti-rabbit platelet antibody. Soluble fibrin was then generated by the intra-venous infusion of ancrod (which cleaves fibrinogen without activating

564 MORRISON AND ULEVITCH American JouMalof Pathology
the coagulation system). On administration of the second dose of endo-toxin to treated animals, no decrease (in fact, a slight enhancement) ofglomerular microclot formation was observed. The authors conclude thatplatelets do not contribute to the precipitation of soluble fibrin induced byendotoxin.A more recent approach to assess the role of platelets in tissue injury
induced by endotoxins has employed busulphan (methane sulfonic acid)to deplete rabbits differentially of platelets and granulocytes.1"'75 Thisexperimental approach has at least one advantage over the use of heterol-ogous antiplatelet antibody in that inadvertent activation of the com-plement and/or coagulation sequences in plasma following administra-tion of antibody (and generation of antigen-antibody complexes) isavoided. The subsequent injection of two sequential doses of endotoxinsinto such neutropenic and thrombocytopenic rabbits failed to inducedisseminated intravascular coagulation. Significantly, however, the re-placement of platelets by transfusion failed to restore the capacity ofrabbits to generate microthrombi. This was not the case, however, whengranulocytes were replaced. (See Polymorphonuclear Leukocytes [Neu-trophils4.) These results provide additional evidence that platelets are notessential participants in the activation of intravascular coagulation byendotoxin.Whereas the available evidence would not support a prominent role for
participation of the platelet in the in vivo intravascular coagulation initi-ated by endotoxins (in spite of the generation of platelet factor 3 activity),the available evidence would, in many instances, support a critical role forserum complement in the in vivo responses of platelets to endotoxins. Forexample, as first demonstrated by Fong and Good," only minor changes incirculating platelets were observed in rabbits depleted of C3 and terminalcomplement components (by pretreatment with CoF) compared withuntreated rabbits following the injection of endotoxin (E coli Boivin).Similar results were obtained by Brown and Lachmann 90,176 using rabbitseither genetically deficient in C6 or depleted of C3 and terminal com-ponents by CoF. These authors observed that normal rabbits injected witha lethal dose (5 mg) of S marcescens endotoxin (Boivin) manifest a rapiddrop in circulating platelets, the majority of which, the authors suggest,were acutely destroyed. These rapid platelet drops were virtually abro-gated in C3-9-depleted animals (CoF-treated). Of significant interest,administration of endotoxin to C6-deficient rabbits caused an acute se-questration of platelets which were, however, destroyed only to a variableextent. On return to the circulation, neither these platelets nor the plate-

Vol. 93, No. BACTERIAL ENDOTOXINS 565November 1978
lets in the C3-9-depleted animals showed the secondary, more gradualplatelet drops observed in normal animals. The results obtained by theseauthors confirmed the earlier results of Horowitz et al 16" on the genera-tion of platelet factor 3 activity in vivo and extended these observations todemonstrate the complement dependence of this response. Whereas goodplatelet factor 3 activity was observed in normal animals, virtually noactivity could be detected in either C6-deficient or C3-9-depleted ani-mals. These results indicate the requirement for C3 and terminal com-ponents in platelet factor 3 generation. Although no data were presented,these authors further reported the binding of 1'Cr-labeled endotoxin tothe platelets in uivo, results consistent with the morphologic observationsof Spielvogel.lU2
Miiller-Berghaus and Lohmann,66 however, reported that the responseof platelets to S enteritidis endotoxin in both normal and C6-deficientrabbits was virtually indistinguishable. However, because the earliesttime point examined by these authors was approximately 2 hours follow-ing endotoxin injection, they would have been unable to assess the veryearly thrombocytopenia observed by the earlier investigators.
Experiments in other species have also demonstrated complement-dependent platelet responses to endotoxins. The rapid drop in circulatingplatelets induced in dogs by endotoxin injection 166 was shown to bedependent on serum complement.' Similar experiments have also dem-onstrated a complement-dependent platelet response in cats." Of signifi-cance, experiments using C4-deficient guinea pigs have provided evi-dence that activation of the classic pathway of complement was requiredfor the initial rapid drop in guinea pig platelets following endotoxininjection.7' This result was somewhat surprising in view of the fact thatthe available in vitro data would support a prominent role for alternativepathway dependence of complement platelet interactions. (See Endo-toxin-Platelet Interactions In Vitro.) However, species differences, as wellas differences in sequestration vs lysis, may account for these differences.The accumulation of data demonstrating an immediate complement-
dependent drop in circulating platelets in dogs, rabbits, cats, and guineapigs, which was dependent on complement components at least throughC3,90,176 implicates this phenomenon as a characteristic in vivo response ofplatelets (containing immune adherence receptor sites) to endotoxin.Evidence suggesting that this was not always the case was first reportedby Levin and Beck,ln who observed only gradual drops in circulatingplatelets following injection of E coli 026:B6 endotoxin. Further informa-tion relating to this point was provided by the experiments of Ulevitch et

566 MORRISON AND ULEVITCH American Joumalof Pathoogy
al,"* using three different preparations of endotoxin: E coli 0111 : B4 (West-phal), S marcescens (Westphal), and S minnesota R595. Of importance,the platelet response in rabbits to these three preparations of endotoxinwas not identical. The endotoxin from E coli 0111:B4 did not induce arapid decrease in platelets; however, a gradual decrease which was unaf-fected by complement depletion was observed. In contrast, the latter twoendotoxins both induced rapid platelet drops as well as secondary plateletchanges which were similar to those observed with 0111: B4 endotoxin. Aswas true in the earlier studies, the early thrombocytopenia was abrogatedby complement depletion. However, the secondary platelet drops withthese endotoxin preparations were unaffected by complement depletion.These results contrast with the earlier studies by Brown and Lachmann "
but are consistent with the observations of Muiller-Berghaus and Loh-mann." The reasons for the differences in the results of this study andthose of Brown and Lachman 90 are unclear. These data, therefore, in-dicate that endotoxins can induce both complement-dependent and com-plement-independent responses in circulating rabbit platelets. The char-acteristics of the in vivo platelet response depend to a certain extent on thepreparation of endotoxin used.
These accumulated data therefore support the conclusion that the invivo responses of circulating platelets to endotoxins in animal specieswhose platelets are immune-adherence-receptor-positive are significant.In many cases the responses are dependent on complement. Although themajority of the available data suggest that platelets may not be essentialfor the initiation of many of the pathophysiologic changes induced byendotoxin, it would be difficult to conclude that their presence, coupledwith their demonstrated capacity to be affected by endotoxins, does notplay any role in the host response to endotoxin.The relationship of endotoxins and platelets in animal species whose
platelets lack immune adherence receptors, particularly humans and sub-human primates, is less defined. Although clearly of interest, the resultsobtained in the experiments summarized above are not particularly appli-cable to endotoxin-platelet interactions in humans. Unfortunately, but forobvious reasons, significantly less information is available on the in vivoeffect of endotoxins on primate platelets. Most experimental models haveused intravenous injections of relatively large (in comparison to, forexample, rabbits) amounts of endotoxin into monkeys, and the results ofmost studies have demonstrated only modest decreases in levels of circu-lating platelets. The results of one study in which S abortus equi endo-toxin (0.5 to 2 ng/kg) was injected intravenously into human volun-teers '7 did not demonstrate any consistent alterations in levels of

Vol. 93, No. 2 BACTERIAL ENDOTOXINS 567November 1978
circulating platelets. it should be emphasized, however, that the lack of ademonstration of alterations in levels of circulating platelet levels does notnecessarily reflect a lack of platelet-endotoxin interaction which mightresult in functional alterations in platelet activity. Unfortunately, suchstudies have not been performed in primates.The experiments of Hinshaw, Jordan, and Vick 178 first assessed the
effects of endotoxins on the level of circulating platelets of the primateCercocebus torquatus Atys (Sooty Mangabey). These authors demon-strated significant decreases (up to 70%) in platelets within 60 to 120minutes following the injection of 5.2 to 13 mg/kg of E coli endotoxin. (Inthis and similar studies, note the large amounts of endotoxins used.)Similar although less extensive decreases in circulating platelets werenoted bv Sheagren et al 179 in rhesus monkeys following the administra-tion of very high doses (1.25 to 12.5 mg/kg) of E coli or S typhosaendotoxins. In several animals, significant platelets drops were observedwithin 2 to 5 minutes following endotoxin injection. Very similar, rapidplatelet responses were observed in the macaque monkey 180 followinginjection of E coli endotoxin. Although these results are suggestive of theearly complement-dependent endotoxin-induced platelet changes in-duced in other species, there is insufficient evidence to correlate the tworesponses. More recent experiments by Ulevitch et al 181 have examinedthe effects of several endotoxins on circulating platelets in rhesus mon-keys. Although these authors did not observe an initial rapid drop inplatelets following the injection of 500 Ag/kg E coli 0111: B4 or S minne-sota R595 (rough) endotoxin, they did observe gradual platelet decreases,resulting in a reduction to approximately 50% during a 5-hour period. Itshould be noted that the concentration of endotoxin used in this study wassignificantly less than that employed in previous studies. Of considerableinterest in this study, the in- vivo platelet responses to endotoxin wereindependent of C3 and terminal complement components, a result similarto that reported earlier by several of these authors in studies performed inrabbits.
Morphologic evidence to support an in vivo platelet response to endo-toxin in primates has yielded less clear-cut evidence. Experiments byMcKay et al 182 suggested-that platelets may play a role in the formation ofmicrothrombi following injection of endotoxin (10 mg/kg) into rhesusmonkeys. These authors observed fibrin-platelet-leukocyte thrombi inhepatic sinusoids within 1 to 2 hours following endotoxin injection. Theyfurther reported that platelet aggregation in small vessels, particularlysplenic sinusoids, was another prominent anatomic change. These resultsprompted the authors to propose that such aggregation of platelets may

568 MORRISON AND ULEVITCH American Joumalof Pathology
account for the thrombocytopenia following administration of endotoxin.Contrasting results were published by Balis et al,7g who were unable todetect either platelet aggregates or any ultrastructural alterations in plate-lets in the lungs of rhesus monkeys receiving an acute lethal dose (5 to 10mg/kg) of E coli 026:B6 endotoxin. These authors concluded that plateletalterations leading to intravascular coagulation and release of vasoactiveamines did not play a significant initiating role in the development ofacute endotoxin-induced pulmonary changes (shock lung) in primates.
These combined studies suggest that bacterial endotoxins can, undercertain circumstances, affect the levels of circulating immune-adherence-receptor-negative platelets. However, the unusually high amount ofendotoxin uniformly described -as being required to induce platelet re-sponses in primates (5 to mg/kg) in relation to the amounts which aresufficient to induce fever in primates (5 to 10 ng/kg) provide support for aminimal role for primate platelets in response to bacterial endotoxins.
Endotoxin-Platelet Interactions In Vitro
The responses of platelets to bacterial endotoxins in vitro may also becatagorized into immune-adherence-receptor-positive platelet responsesand immune-adherence-receptor-negative platelet responses. Experi-ments utilizing the former system have focused almost entirely on therabbit platelet, whereas the latter system has described human platelet-endotoxin interactions almost exclusively. Since the platelet responses inthese two systems have been demonstrated to differ significantly in vitroas well as in vivo (see Endotoxin-Platelet Interactions In Vitro), they willbe considered separately in the following paragraphs.
In parallel with the early in vivo experiments on the effects of endo-toxins on circulating dog platelets,'" Davis, Meeker, and McQuarrie 185also examined the in vitro effects of endotoxins on platelets. These authorsdemonstrated the in vivo release of serotonin from anticoagulated rabbitblood following incubation with E coli endotoxins. Similarly, experimentsby Siqueira and Nelson 186 showed agglutination of guinea pig and rabbitplatelets using a soluble Boivin antigen of S typhosa in the presence ofantologous fresh serum or pooled complement. These authors suggestedthat agglutination of platelets was the result of the presence of naturalantibody in the serum capable of reacting with the S typhosa antigen.This interaction then yielded an immune complex which, in the presenceof complement, induced the platelet aggregation via the immune adher-ence sites.
Des Prez et al, 16187-191 in a comprehensive series of experiments, exten-sively characterized the in vitro response of rabbit platelets to endotoxins

Vol. 93, No. 2 BACTERIAL ENDOTOXINS 569November 1978
from E coli 0127:B8 (Boivin). (These authors noted that different lots ofendotoxin varied in their capacity to elicit platelet responses.) The resultsof these experiments clearly demonstrated that incubation of E coli endo-toxin (100 gg/ml) with platelet-rich rabbit plasma resulted in a time-dependent generation of platelet factor 3 activity,'"' release of sero-tonin,'" and platelet aggregation."8 Aggregation and release of serotoninwere shown to be inhibited by incubation at 4 C or by prior absorption ofthe plasma with heat-killed organisms. The release of serotonin variedlinearly as a function of time and was optimal (at the dose tested) after 3hours at 37 C. This observation, as well as the fact that thrombin but notendotoxin-induced responses were not inhibited by heparin, providedevidence that the mechanism of endotoxin-initiated platelet responses didnot involve the participation of thrombin.1' To exclude the potentialparticipation of natural antibody to the endotoxin (which would contrib-ute to immune-complex-mediated platelet damage) experiments wereperformed to examine the divalent cation requirements of both endo-toxin-and immune-complex-mediated platelet injury. Although both ofthese agents initiated release of serotonin in the presence of heparin,endotoxins but not immune complexes were capable of inducing release inplasma anticoagulated by either citrate (0.38%) or NaEDTA (0.1 %). Theobservation that neither agent was effective in 0.17% EDTA would,however, suggest an essential role for divalent cations in endotoxin-induced platelet damage."9 Additional experiments by these investigatorsdemonstrated that the endotoxin-initiated platelet injury was dependenton a heat-labile plasma factor.1"'m91 This factor was shown to be absorbedby zymosan under conditions which did not affect complement titers (16C), and the data suggested that the plasma factor affected in these twosystems was very similar. The serum hemolytic complement titers werenot reduced during the course of endotoxin-induced platelet injury. DesPrez suggested from these experiments the potential participation ofproperdin in modulating the plasma-dependent platelet response to endo-toxin.
Morphologic support for platelet-endotoxin interactions was providedby the elegant electron micrographs of Davis 161 and Spielvogel."6 UsingE coli 0127:B8 endotoxin at a concentration of 10 gg/ml in the presence ofheparinized plasma and rabbit platelets, Davis provided evidence forsignificant alterations in the limiting membranes of platelets after in-cubation at 37 C for as little as 90 seconds. After 30 minutes of incubation,significant numbers of large platelets aggregates were observed accom-panied by the appearance of a granular material in the medium. Similar tothe results obtained by Des Prez et al,8"7 no response was observed in

570 MORRISON AND ULEVITCH American Journalof Pathology
heparinized plasma at 4 C or, in addition, plasma anticoagulated withcitrate.
Spielvogel,'92 in a comprehensive study on the response of platelets froma number of species to endotoxin (E coli 0127:B8 Boivin), demonstratedsignificant differences in responses depending on the species of plateletsused. Rabbit, rat, dog, and guinea pig platelets (immune-adherence-positive) all aggregated in response to endotoxin in the presence ofplasma, although degranulation was limited to the first two platelet spe-cies. The electron micrographs provided evidence for adherence of theendotoxin particles to the platelet membrane, and, although in someinstances phagocytosis of the endotoxin particle by the platelet did occur,degranulation was not dependent on phagocytosis. In this respect, inhibi-tion of phagocytosis had no effect on endotoxin-induced degranulation.Spielvogel used a number of procedures, some of them similar to thosedescribed earlier by Des Prez et al,87 to demonstrate that aggregation anddegranulation were dependent on a functionally active complement sys-tem. The results of these experiments suggested that endotoxin-inducedplatelet damage was dependent on intimate contact between endotoxinsand platelets provided by immune adherence sites and required bothantibody and complement: these conclusions are similar to those sug-gested earlier by Siqueira and Nelson.1"
Experiments attempting to further define the mechanisms of endotoxin-initiated rabbit platelet damage were reported by Evans et al.70 Theseauthors used a number of platelet inhibitors of ADP release, includingacetylsalicylic acid, phenylbutazone, sulfinpyrazone, and sodium salicy-late, to inhibit endotoxin-induced rabbit platelet aggregation in hepari-nized plasma. These results suggested a direct effect on the release of ADPby endotoxin.Nagayama et al 192 examined a spectrum of bacterial endotoxins, includ-
ing E coli 026:B6, S typhosa 0901, S typhosa Ty58, S enteritidis, Smarcescens 08, and a detoxified preparation of S typhosa Ty58 (endo-toxoid), on a number of rabbit and human platelet functions, includingaggregation, release of serotonin, and generation of platelet factor 3activity in citrated platelet-rich plasma. The results of these experimentssuggested that all of the preparations tested, including the "detoxified"endotoxin, could, to a variable extent, affect each of the rabbit plateletfunctions tested. In one instance differing results were obtained usingendotoxins from the same organism but derived from different sources.However, since no indication was given as to how the endotoxins wereprepared, it is difficult to evaluate the significance of these differences.A significant contribution to the understanding of the mechanism of

Vol. 93, No. 2 BACTERIAL ENDOTOXINS 571November 1978
endotoxin-induced rabbit platelet injury was provided by experimentsreported by Siraganian.1I He demonstrated that a number of substances,including endotoxins, which normally induce plasma-dependent rabbitplatelet release reactions, were ineffective in plasma obtained from rabbitsgenetically deficient in C6. The critical dependence on C6 was establishedby the restoration of release responses by readdition of purified C6 to thedeficient plasma. Additional support for the participation of C3 andterminal complement components was provided by experiments demon-strating that plasma treated with the anticomplementary protein fromcobra venom (CoF) also did not support platelet-release reactions.The experiments of Brown and Lachmann " confirmed these studies,
demonstrating a requirement for terminal complement components inendotoxin-induced platelet release. These experiments also demonstratedthat generation of platelet factor 3 activity preceded the release of 51Cr (asan indicator of platelet release by complement-mediated lysis). Additionalexperiments by these authors 176 showed that, with low concentrations ofendotoxin (100 Ag/ml of S marcescens or E coli 0111 :B4), platelet factor 3activation required C3 and terminal components. However, at high con-centrations of endotoxin (1.0 mg/ml), although no activity was observedwith C3-9-depleted plasma, moderate activity could be generated in C6-deficient plasma. The specificity of the response was shown to reside in theplasma rather than in the platelet component of the system.
Using an endotoxin preparation from Acinetobacter 199A, Maclntyre etal 19 further explored the mechanism of endotoxin-initiated platelet re-sponses. These investigators provided evidence that the platelet aggrega-tion following endotoxin occurs in three distinct phases. They proposedthat the first phase involved binding of the endotoxin to the platelet whichoccurred in the absence of Ca++ or plasma proteins. An initial aggregationstep followed, which was not accompanied by secretion of platelet constit-uents but did require Ca++. A second phase of aggregation then ensued,which was accompanied by secretion of granule constituents and, to alesser extent, cytoplasmic leakage. They provided further evidence for atleast a partial secretory response by demonstrating that stimulation ofadenyl cyclase, inhibition of phosphodiesterase, or inhibition of prosta-glandin synthesis would inhibit the rabbit platelet responses. The authorssuggested, on the basis of plasma dilution and chelation studies, that theprimary phase required complement activation of C3 via the classicpathway. Additional experiments by these investigators 196 demonstratedalterations in platelet membrane protein components following additionof endotoxins.The experiments reported by Morrison et al,9m1"0 however, provided

572 MORRISON AND ULEVITCH American JouMalof Pathlogy
data which would support a prominent role for alternative, rather thanclassic, complement pathway activation leading to rabbit platelet lysis. Inaddition, in these experiments, the response was demonstrated to beentirely due to complement-mediated lysis, since identical release ofgranule localized serotonin and cytoplasmic lactic dehydrogenase wasobserved. These experiments employed a number of different endotoxinpreparations, as well as isolated lipid A, which have earlier been demon-strated to activate preferentially only one of the two complement path-ways. Significantly, only those preparations which had the capacity toactivate the alternative pathway were able to initiate rabbit platelet lysis.In this respect, isolated lipid A (which activates selectively the classiccomplement pathway by an interaction with Cl in t}ie absence of anti-body) (see Endotoxin-Complement Interactions In Vitro) did not initiaterabbit platelet lysis. Nevertheless, additional experiments by these investi-gators provided evidence that the lipid A portion of endotoxins does play arole in the platelet response, ie, by providing the means for attachment ofthe endotoxin molecule to the platelet, a phenomenon first observed bySpielvogel.1" Thus, agents such as polymyxin B, which bind to lipid A, ormild alkali, which hydrolyses selectively lipid-A-associated fatty acids (seeRelationship of Endotoxin Structure to Biologic Activity), abrogate thecapacity of endotoxin to initiate platelet lysis. The presence of lipid-A-associated protein (LAP, the protein component of bacterial endotoxins)enhanced by two orders of magnitude the capacity of endotoxins to causeplatelet lysis. Because the presence of the protein had no effect on altema-tive pathway activation by endotoxins (see Endotoxin-Complement Inter-actions In Vitro), this experiment suggested that LAP facilitated theattachment of the endotoxin to the platelet membrane. Further support-ive evidence was derived from experiments using endotoxins purified frommutant rough strains of bacteria. These endotoxins, which lack poly-saccharide but contain lipid A, did not initiate platelet lysis but wereeffective at blocking endotoxin responses to polysaccharide-containingendotoxins.
These accumulated data thus provide convincing evidence that im-mune-adherence-receptor-positive platelets can be profoundly altered invitro by bacterial endotoxins in the presence of complement. Plateletresponses are characterized by aggregation, generation of platelet factor 3activity, and release of intracellular constituents (by a complement-medi-ated platelet lysis and perhaps by noncytotoxic secretion). The plateletresponses are initiated by attachment of endotoxin to the platelet mem-brane involving a calcium- and plasma-protein-independent interactionvia the lipid A or lipid-A-associated protein region of the endotoxin

Vol. 93, No. 2 BACTERIAL ENDOTOXINS 573November 1978
molecule. This close association of the endotoxin with the platelet thenallows efficient assembly of alternative pathway complement componentson the polysaccharide portion of the endotoxin, which, in the micro-environment of the platelet membrane, generates terminal componentassembly and subsequent platelet lysis.
In addition to the complement-mediated platelet response to endo-toxin, a recent report by Morrison and Oades I" has demonstrated acomplement-independent secretory response of rabbit platelets to endo-toxins from rough polysaccharide-deficient strains of bacteria, eg, S min-nesota R595. Preincubation of washed rabbit platelets with such endo-toxins in Ca++-free buffer at 37 C followed by the addition of Ca++ (butnot Mg++ or Ba++) initiated an energy-dependent secretion of serotoninwithout concomitant release of cytoplasmic lactic dehydrogenase. A directcorrelation between amount of endotoxin added to platelets and sub-sequent association of radiolabeled calcium was noted, suggesting thatrough endotoxins may mediate calcium translocation in rabbit platelets.This response was not observed with polysaccharide-containing endo-toxins.The in vitro response of immune-adherence-receptor-negative platelets
(particularly human) to bacterial endotoxins is significntly less clearlydefined, and several conflicting reports have appeared in the literature.The experiments of Ream et al 200 first described in an in vitro system
the response of washed preparations of human platelets to a variety ofbacterial endotoxins. The authors observed that, in contrast to rabbitplatelets, in which the addition of endotoxin initiated massive aggregationin platelet-rich plasma, the response in human platelet-rich plasma was"'occasional and variable clumping." Nevertheless, these data providedmorphologic evidence for aggregation of human platelets in response to50 ug of E coli 0111:B4 endotoxin as well as endotoxins from a number ofother species. Evidence was presented which suggested that a heat-labileplasma factor (most probably Factor V) was the limiting factor in theaggregation response to endotoxins. Finally, these experiments demon-strated, with only one exception, a correlation between ability of endo-toxins to induce human platelet aggregation and their toxicity in the chickembryo lethality assay.
In contrast to these results, however, a number of investigators havebeen unable to initiate human platelet aggregation with a spectrum ofendotoxin preparations. 19,1 2N-2W Spielvogel '2 examined a Boivin ex-tract of E coli 0127:88 endotoxin and found that, although it inducedresponses in all immune-adherence-receptor-positive platelets, it had nodetectable effects on human or baboon platelets, either as assessed by

574 MORRISON AND ULEVITCH American Journalof Pathology
aggregation or degranulation. Similar results were obtained by Muller-Eckhardt and Lflscher,201 who reported no response of human platelets toeither E coli 055 or 0127 endotoxins (Boivin). These conclusions werederived from experiments utilizing both washed human platelets andhuman platelets suspended in pooled human plasma and were unalteredif the platelets were pretreated with a-chymotrypsin. These authors con-cluded that an immunologic injury of platelets by endotoxin comparablewith the effect of immune complexes or aggregated Y-globulin washighly improbable. More recent experiments by Pfueller and Liischerconfirmed these earlier studies."2 In these experiments, it was shown that500 ug/ml of endotoxin (E coli 026:B6 Boivin) had absolutely no detect-able effect on human platelet-rich plasma as assessed either by aggrega-tion or release of platelet constituents. Incubation of endotoxin withplasma did not inhibit the subsequent response of human platelet-richplasma to zymosan.
Experiments by Nagayama et al 192 examined the effects of severalendotoxin preparations on a number of human platelet responses, includ-ing aggregation, secretion of serotonin, and generation of platelet factor 3activity. These investigators found that none of the endotoxins testedinduced aggregation or platelet factor 3 activity. However, several of theendotoxins were able to initiate release of serotonin from human platelets,although whether this occurred by a lytic or secretory mechanism was notdetermined. The release of serotonin was shown by these authors not beinhibited by aspirin. In addition, in a recent study by Vermylen et al, "3eight endotoxins were tested in human platelet-rich plasma and in washedplatelet preparations. In no situation did the authors demonstrate signifi-cant aggregation or release of serotonin.
Several recent reports by Hawiger and his collaborators 2-"6 have,however, provided evidence for a secretory response of washed humanplatelets to a bacterial endotoxin preparation from S enteritidis (Boivin).The secretory response was demonstrated using this endotoxin at 200 ug/ml in plasma-free buffer. Secretion was characterized by the release ofdense granule localized serotonin and adenine nucleotides without releaseof the cytoplasmic enzymes lactic dehydrogenase or 0-glucuronidase.Significant generation of platelet factor 3 activity was also demonstratedand was shown to be associated with platelet membrane phospholipids.Neither hirudin nor heparin abolished the response, suggesting that theresponse was not dependent on thrombin. Of interest, and as was sug-gested by earlier studies,'" neither aspirin nor indomethicin blocked theresponse, suggesting that prostaglandins did not play a prominent role inthe response.
Additional experiments by these investigators demonstrated that the

Vol. 93, No. 2 BACTERIAL ENDOTOXINS 575Noveber 1978
platelet secretion in response to endotoxins could be blocked with aplatelet membrane fraction, suggesting a membrane site for the endo-toxin-platelet interaction. These results would be consistent with thoseobtained by Springer and Adye,"7 who also had demonstrated endotoxin-binding sites on the surface of human platelets. Additional studies bythese authors 20 have partially characterized the binding material andshown it to consist of selective phospholipids (primarily phosphatides).A report of a somewhat unusual effect of selected preparations of
endotoxins on human platelet secretory responses has recently been pub-lished by Ginsberg and Henson.210 These authors found that preparationsof endotoxins from polysaccharide-deficient (rough) strains of bacteria, aswell as isolated lipid A, significantly enhanced the secretory response ofwashed human platelets to aggregated immunoglobulins or to immunecomplexes. The endotoxin preparations by themselves were without de-tectable activity, as suggested by earlier investigations.10"a2012s0 Theenhancement was specific for aggregated immunoglobulins in that theendotoxins had no effect (and in fact were slightly inhibitory) on humanplatelet secretion induced by collagen, antiplatelet antibody, urate crys-tals, or thrombin. Although the mechanism of the observed enhancementwas not defined, the recently published evidence 211 demonstrating theformation of molecular complexes between such endotoxin preparationsand aggregated, but not monomeric, immunoglobulins suggests that theenhancement involves an interaction of the endotoxin with the aggregatedimmunoglobulins on the platelet surface.
These combined data support the concept that most endotoxins havelittle, if any, effect on human (or other immune-adherence-site-negative)platelets, either in platelet-rich plasma or in plasma-free buffer. Theseeffects include aggregation, release of intracellular constituents, and gen-eration of procoagulant activity. Such a generalization cannot be extendedto all endotoxins, however, since at least one preparation (S enteritidis,Boivin) has been clearly demonstrated to initiate a human platelet secre-tory response. The relevance of the human platelet response to thisendotoxin to more general mechanisms of human platelet-endotoxin in-teractions remains to be established. This would also apply to the capacityof selected endotoxin preparations to enhance secretory responses ofhuman platelets to aggregated immunoglobulins.
Endotoxin-Neutrophil Interactions In Vivo
Among the many physiologic manifestations produced by administra-tion of endotoxin, one of the most widely studied is the extreme leuko-

576 MORRISON AND ULEVITCH American Joumalof Patoogy
penia followed by leukocytosis. Because of the large number of descrip-tive experiments relating to the effects of endotoxins on polymorpho-nuclear leukocytes (neutrophils) in vivo, it is not possible to review thissubject exhaustively here. Rather, we will describe the pioneering experi-ments, emphasize studies related to mechanisms, and cite previousreviews for sources of information about various aspects of endotoxin/neutrophil interactions in vivo. An excellent historic summary of earlyobservations relating to the effects of bacterial products on circulatingblood leukocytes appeared in a review published in 1950 by Bennett andBeeson.2l2
Delaunay showed in 1943 that a glycolipid isolated from gram-negativebacteria produced a marked leukopenia when injected into guinea pigs.213He suggested that the leukopenia resulted from the trapping of these cellsin the capillaries of the lungs, liver, and spleen. In 1951, Stetson 157described the time course of the changes in levels of circulating neutro-phils following intravenous injection of a meningococcal endotoxin intorabbits. These changes were characterized by a rapid neutropenia occur-ring within minutes of injection and a subsequent period of 3 to 4 hoursduring which neutrophil levels remained at less than 10% of pre-endo-toxin levels. Recovery from this effect occurred, and by 6 hours afterinjection the blood leukocyte level was normal or elevated. This increasein circulating neutrophils was accounted for by a large number ofimmature cells. Stetson also confirmed previous suggestions that thedisappearance of the neutrophil was a result of sequestration of these cellsin capillaries, particularly in the lung. The marked effect of endotoxins oncirculating polymorphonuclear cells, resulting in the immediate neutro-penia followed by a granulocytosis, has been well documented in allspecies, including humans.214'215
Athens et a1214 employed kinetic techniques using 'H-diisopropylphos-phofluoridate-labeled neutrophils to show that the endotoxin-inducedleukopenia could be explained by a shift of these cells from the circulatingpool to the marginal granulocyte pool. These data are consistent with theearlier morphologic observations of neutrophil sequestration in capillarybeds. In the same study 214 the granulocytosis was shown to be secondaryto the release of granulocytes from the bone marrow reserves.
Recently it has been shown that products derived from complementactivation are capable of producing granulocytopenia as well as neutro-phil adherence in vitro.216 Whether a similar mechanism accounts forendotoxin-induced neutropenia and granulocyte adherence is unclear.Evidence to suggest that complement-independent mechanisms accountfor these changes derives from several investigations. Studies reported by

Vol. 93, No. 2 BACTERIAL ENDOTOXII;JS 577November 1978
Kane, May, and Frank 79 using C4-deficient guinea pigs showed that Ecoli 0127:B8 endotoxin (Westphal) produced a marked leukopenia despitethe absence of C4. They further showed that animals previously depletedof CS with CoF also demonstrated a rapid neutropenia, the kinetics andextent of which were indistinguishable from those observed in normo-complementemic or C4-deficient guinea pigs. Fong and Good " reportedthat the neutropenia in rabbits resulting from the injection of 270 Mg of aBoivin preparation of E coli endotoxin was unchanged by prior depletionof C3 with CoF. Studies by Miiller-Berghaus and Lohmann " demon-strated that the leukopenia resulting from infusion of 40 Mg/kg/hr of aBoivin preparation of S enteritidis endotoxin in normocomplementemicand C6-deficient rabbits was indistinguishable. Ulevitch et al8 alsoshowed that C3 depletion of rabbits failed to alter the kinetics or extent ofthe neutropenia produced by the injection of 100 Mg of E coli 0111:B4endotoxin (Westphal) or 50 Mg of S minnesota R595 endotoxin. Finally, ina recent report by Ulevitch et al 181 it was shown that CoF treatment ofrhesus monkeys did not alter the kinetics or extent of the neutropeniaproduced by the injection of 500 ug/kg of S minnesota R595 endotoxin.Therefore, it appears that in both nonprimate and primate species thepresence of an intact complement system is not essential for the endo-toxin-induced neutropenia.
However, in the report of Ulevitch et al 181 it was also shown thatfollowing the rapid injection of 5 mg/kg of S minnesota R595 endotoxininto rhesus monkeys, the rate of the neutropenia was significantly de-creased by prior depletion of C3. These results suggest that, under someconditions, complement-dependent reactions may contribute, in part, tothe neutropenia induced by endotoxin injection. This problem warrantsfurther study, and perhaps studies comparing endotoxins with differingcapacities for complement activation will clarify the role of complementin endotoxin-induced neutropenia.
MacGregor 217 investigated the mechanisms of the endotoxin-inducedneutropenia in rabbits injected with 0.1 Mg/kg of a Boivin preparation ofE coli 0127:88 endotoxin. He was able to show that, following endotoxininjection, the granulocytes demonstrated a marked enhancement of ad-herence when tested in an in vitro system. MacGregor showed that plasmaremoved 1 hour after endotoxin injection would significantly enhance theadhesiveness of PMN cells from a control animal. This factor was notpresent in serum and could be inhibited, in part, by heating the plasma at56 C for 30 minutes. Cells removed 1 hour after endotoxin injection, whenresuspended in normal plasma, demonstrated normal adherence. Theidentification and characterization of this substance might provide insight

578 MORRISON AND ULEVITCH American Joumalof Pathology
into the mechanism of endotoxin-induced granulocytopenia, and thiscertainly warrants further study.A role for lipid A in the induction of neutropenia following endotoxin
injection is suggested by the work of Corrigan and his collaborators. In1971, Corrigan and Bell 21 showed that pretreatment of a Boivin prepara-tion of E coli 0127:B8 endotoxin with polymyxin B (see Relationship ofEndotoxin Structure to Biologic Activity) significantly reduced the neutro-penia produced in rabbits injected with the endotoxin alone. More re-cently, Corrigan et al 29 examined the effect of polymyxin B on theneutropenia produced by intravenous injection into rabbits of typhoid-paratyphoid vaccine (a source of endotoxin). These experiments showedthat pretreatment of the vaccine with polymyxin B markedly inhibited theability of this endotoxin to produce the neutropenia. These data suggestthat the biochemical changes required for endotoxin-induced neutropeniaare triggered by a lipid A/mediation system interaction and imply that thecarbohydrate region of the endotoxin is not essential for this response.
Because of the marked effects of endotoxins on neutrophils in vivo,these cells became a focal point for studies designed to elucidate themechanisms of the various biologic and pathologic effects of endotoxin.For example, the neutrophil was regarded as the principal source ofpyrogen responsible for the endotoxin-induced febrile response. Wood, in1958, reviewed experimental-evidence supporting this concept.2t How-ever, subsequent studies suggested that alternative sources for endoge-nous pyrogen are present, and the blood monocyte and tissue macrophagehave been suggested to be major sources for this substance. Experimentalstudies of the pathogenesis of fever have been reviewed in 1973 byGriesman and Hornick 11 and in 1978 by Dinarello and Wolff.210The neutrophil was also suggested to participate in the pathogenesis of
endotoxin-induced coagulative changes (see Coagulation Systers andReference 143), particularly in the Shwartzman phenomenon. However,the blood monocyte now appears to be the principal source of tissuefactor, and the role of the neutrophil in the development of endotoxin-induced DIC is in question. Other studies performed by Miller, Reichgott,and Melmon suggest that endotoxin/neutrophil interactions may leadto hemodynamic changes principally through the generation of bradyki-nin.The demonstration of sequestration of blood neutrophils in pulmonary
capillaries following endotoxin injection led to suggestions that thesesequestered cells may contribute to the pulmonary injury accompanyingendotoxin shock (shock lung). Coalson et al 224 showed that as early as 15minutes following the injection into rhesus monkeys of 6 mg/kg of E coli

Vol. 93, No. 2 BACTERIAL ENDOTOXINS 579Novenber 1978
endotoxin, marked accumulation of neutrophils in pulmonary capillariesoccurred. The cells observed were often fragmented, and loss of thespecific granules was noted. Subsequently, at 1 hour after injection,marked degranulation of the cells was observed and the capillary endothe-lium appeared damaged at areas of leukocyte attachment. Four hoursafter injection, additional degranulation and fragmentation of neutrophilswere noted.To evaluate the role of the granulocytes in endotoxin-induced pul-
monary injury as well as hypotensive shock, Pingelton et al 2n studiedrhesus monkeys rendered leukopenic by total-body irradiation. Theseanimals had less than 3% of the normally observed white blood cell count.Animals were injected with 6 mg/kg of E coli endotoxin, and measure-ments of cardiac output, blood pressure, and blood bases were performed.A detailed examination of pulmonary tissue was also performed at thelight and electron microscopic levels. The physiologic changes induced byendotoxin were not significantly different in the leukopenic and normalmonkeys. The histologic changes induced by endotoxin in the pulmonarycapillaries were indistinguishable in the two groups of animals. Thus, incontrast to the results of Coalson et al,224 these studies suggest that thepathogenesis of endotoxin-induced changes in the rhesus monkey doesnot require the participation of blood leukocytes.
Finally, several studies have evaluated the extent to which endotoxinbinds to neutrophils following intravenous injection. Herring et al 226employed 51Cr-endotoxin prepared from E coli 026:B6 and examined thedistribution of the radiolabeled endotoxin among the formed elements ofblood at 1, 3, 5, and 8 minutes after injection of 3 to 4 gg/kg of 51Cr-endotoxin. Peripheral blood cells were separated into red blood cell,leukocyte, and platelet fractions. Analysis of these fractions for the con-tent of 51Cr-endotoxin demonstrated that the platelets contained at least97% of the total radioactivity found in the formed elements of blood.Thus, the granulocytes remaining in the circulation following endotoxininjection appeared not to bind significant amounts of endotoxin. Sub-sequently, Brunning et al 2" and Schrader et al 2" employed a preparationof 'H-endotoxin in both formed elements of blood as well as tissuesfollowing injection. In the former study,2' 1 mg of the 'H-endotoxin wasinjected and blood smears and tissues were examined by autoradiographyat different intervals after injection. In these studies it was reported thatuptake into the granulocyte population of blood cells could be detected.Neutrophils localized in the vasculature of the lung, spleen, liver, andkidney were also observed to have bound radiolabeled endodoxin.
In the latter study,20 the localization of 'H-endotoxin was studied when

580 MORRISON AND ULEVITCH American Journalof Patoogy
the radiolabeled endotoxin was injected either as a preparatory (250 Mg) orprovocative dose (400 Mg) for a localized Shwartzrnan reaction. When 3H-endotoxin was given intradermally as the preparatory dosage, it could bedemonstrated that infiltrating neutrophils rapidly took up the materialremaining at the injection site. By 16 hours after the neutrophils hadphagocytosed the 3H-endotoxin, the cells appeared to be degranulatedand began to lyse. In contrast, when the 3H-endotoxin was administeredas the provocative dosage, the localization of labeled neutrophils wasobserved to occur in pulmonary capillaries. These data support the pro-posal that neutrophils can, under the appropriate conditions, take upendotoxins. Because these studies were performed with a Boivin prepara-tion of E coli 0111:B4 endotoxin, the possibility that the presence of LAPenhanced the uptake by PMNs must be considered, especially in view ofthe marked effects of LAP on endotoxin binding to platelets reported byMorrison and Oades.196Although it is difficult to reconcile the findings of Brunning et al 27 and
Schrader et al 2" with those of Herring et al,2 these differences may beaccounted for by the dosage administered as well as the sensitivity of thedetection techniques. In any event, it seems clear that the interaction ofendotoxin with neutrophils in vivo and the effects of such interactions arenot completely understood and require further study.
Endotoxin-Neutrophil Interactions In Vito
In contrast to the in vivo effects of endotoxin on polymorphonuclearleukocytes (neutrophils), the in vitro responses of these cells to endotoxinsare significantly less well understood. Although it would appear on thebasis of a number of recent studies that the neutrophil is probably not thesource of procoagulant activity in preparations of endotoxin-stimulatedmixed leukocytes (see Coagulation Systems), a number of effects of endo-toxins on neutrophils have been noted. Unfortunately, relatively fewstudies have been devoted to an extensive investigation of these multiplepotential manifestations of endotoxins on neutrophils. The literature inthis area, as a consequence, suffers somewhat from a lack of the continuitywhich defines most of the other areas of endotoxin-mediator systeminteractions.The early experiments of Fritze and Wendt 29 examined the effects of
endotoxins on the capacity of human peripheral blood leukocytes tophagocytose bacteria. The results of these experiments demonstrated that,at concentrations of S abortus equi endotoxin from 10 to 1000 Mg/ml inanticoagulated peripheral blood, the phagocytic capacity of human leuko-cytes was significantly enhanced. However, the authors cautioned that the

Vol. 93, No. 2 BACTERIAL ENDOTOXINS 581November 1978
relatively high concentrations of endotoxins required to achieve theseeffects would make such a mechanism unlikely to have significant in vivorelevance.
Collins and Wood,mno who were engaged in an extensive series ofexperiments on the pathogenesis of fever, designed experiments to exam-ine whether stimulation of neutrophils by endotoxins could produce pyro-genic agents. These authors incubated rabbit peritoneal exudate neutro-phils in vitro with S flexneri endotoxin and demonstrated that, in thepresence of autologous serum, endotoxin induced selectively the release ofpyrogenic material. Of interest, these experiments did not detect anydifferences in endotoxin-induced pyrogen release from leukocytes ob-tained from tolerant rabbits compared with untreated control rabbits. Anadditional finding reported by these authors was that incubation of endo-toxins with neutrophils resulted in the inactivation of these molecules,presumably by the release of a factor(s) from the neutrophil.A comprehensive set of experiments performed by Cohn and Morse 231
examined the functional and metabolic alterations in rabbit peritonealexudate neutrophils following incubation with endotoxin (S abortus equi,Westphal). Pre-incubation of neutrophils with endotoxins for 20 minutessignificantly enhanced the ability of the treated neutrophils to kill S albusbacteria, although the magnitude of the effect was dependent on bothendotoxin and serum concentrations. Significant enhancement was ob-served at endotoxin concentrations as low as 0.1 ;g/ml. Additional datademonstrated that the enhanced killing activity was the result of en-hanced phagocytosis, a conclusion similar to that obtained by Fritze andWandt.29 Although no effects on 02 consumption were noted, these datadid provide evidence for significantly enhanced utilization of glucose byglycolysis and resultant production of lactic acid.
Similar effects of endotoxins on the basal metabolism of neutrophilswere noted by Hohnadel and Stjemholm 2n and Graham et al.n3 Theseinvestigators I demonstrated that S typhosa 0901 endotoxin did stimu-late the activity of the pentose cycle, both in resting neutrophils and inneutrophils undergoing phagocytosis. In contrast, no effect on the totalutilization of glucose was observed. Essentially equivalent results wereobtained in the latter study I using guinea pig peritoneal exudate neutro-phils stimulated by E coli endotoxin (50 gg/ml). Although a slight in-crease in respiration of neutrophils was observed following addition ofendotoxin, marked increases in glycolysis were observed. These effectswere inhibited by iodoacetamide and sodium fluoride but not by potas-sium cyanide. In addition to these metabolic changes, the authors alsoobserved increases in the uptake of extracellular fluids following endo-

582 MORRISON AND ULEVITCH American Journalof Pathology
toxin stimulation. There was, however, no definitive evidence by electronmicroscopy for increased pinocytosis or vesicularization induced in theneutrophils by endotoxins.An additional metabolic alteration induced in both rabbit and human
peripheral blood neutrophils, as well as rabbit peritoneal exudate neutro-phils, was described by Tenney and Rafter.28 Using a Boivin endotoxin at360 ,g/ml, these investigators showed a marked decrease in KCI-stimu-lated ATPase activity. This inhibition was observed in intact cells aswell as in cell-free preparations. Treatment of the endotoxin with hy-droxylamine (to remove lipid A-associated fatty acids) (see Relationship ofEndotoxin Structure to Biologic Activity) abolished the inhibitory activityof the endotoxin, suggesting a functional role for the lipid A portion of theendotoxin.
Further support for a major role for lipid A in the action of endotoxinson neutrophils has derived from the recent experiments of Bannatyne etal.235 These investigators demonstrated that the endotoxin (E coli0111:B4, 2 ug/ml)stimulated increase in hexose monophosphate shuntactivity of human neutrophils could be completely abrogated by poly-myxin B (which is known to bind to and block a number of lipid-A-mediated responses) (see Relationship of Endotoxin Structure to BiologicActivity). In addition, the release of several enzymes (ATPase, fl-gluc-uronidase, lysozyme) initiated by endotoxins was also reduced by treatmentwith polymyxin B. It would thus appear that the lipid A portion of theendotoxin preparation plays a prominent role in the interaction of endo-toxins with neutrophils.
Goodale et al 2" provided electron microscopic morphologic evidenceto support these studies on the effects of endotoxins on neutrophils.Following the incubation of whole human anticoagulated blood withendotoxin from S abortus equi, an electron-dense granular material wasdemonstrated to surround the leukocytes, particularly the neutrophils.This material appeared adjacent to the neutrophil cell membrane, and nodetectable damage to the cell was observed. These authors proposed thatthe dense material might be the endogenous pyrogen described earlier byCollins and Wood.tmA method for the indirect assessment of polymorphonuclear leukocyte
phagocytic capacity, employing the nitroblue tetrazolium (NBT) reduc-tion test, has also been utilized to assess neutrophil responses to endo-toxins. Following the addition of a phagocytic stimulus, or endotoxin, to asuspension of neutrophils, NBT is reduced to blue formazin by the neutro-phils. Formazin is a highly insoluble blue-black material which can beeither visualized microscopically or, following extraction, quantitated

Vol. 93, No. 2 BACTERIAL ENDOTOXINS 583November 1978
spectrophotometrically. The absolute number and proportion of "NBTpositive" neutrophils was presumably related to the phagocytic activity ofthese cells. As first demonstrated by Park and Good,"7 the in vitro simula-tion of normal human peripheral blood neutrophils in heparinized wholeblood for 5 to 10 minutes with 20 yg/ml of endotoxin caused approxi-mately 50% of the neutrophils to become NBT-positive. Additional exper-iments demonstrating significantly lower responses in whole blood anti-coagulated with EDTA suggested to these investigators the probableparticipation of complement in the response of endotoxins. The potentialdiagnostic value of this assay was further stressed by these authors.
These results were confirmed by the experiments of Matula and Pater-son,m2 who examined the NBT responses of peripheral blood neutrophilsof normal adults as well as patients with bacterial infection. Whereasneutrophils from the latter group responded spontaneously in the NBTreduction assay, this was not found in normal individuals. Neutrophilsfrom normal adults could, however, be stimulated either with gram-negative culture filtrates or with isolated endotoxin to reduce NBT,although the concentrations of endotoxins required (20 ug/ul) were some-what higher than reported in the previous study. Similar responses wereobtained by Goihman-Yahr et al,2" who examined neutrophil responses ofnormal individuals and of patients with leprosy to E coli 026:B6 (West-phal) endotoxins. These authors found that endotoxins stimulated thereduction of NBT by neutrophils from both groups of individuals. Incontrast to one of the earlier studies, these authors reported that thepresence of autologous serum inhibited the response to endotoxin, al-though the reasons for this were not clear.
In an attempt to elucidate the potential mechanism of the interaction ofendotoxins with neutrophils leading to NBT reduction, Corrigan et al 219examined the effect of the lipid A inhibitor, polymyxin B, on neutrophilresponses. These authors demonstrated that polymyxin B virtually elimi-nated the capacity of S parathyphi endotoxin to stimulate human periph-eral blood neutrophils to reduce NBT. These results would thus be consis-tent with the earlier observations of Tenney and Rafter 2" and Bannatyneet al,'m who examined the capacity of 3P2 and 4(C-labeled endotoxins tobind neutrophils. The results of these experiments demonstrated that thedegree of binding was proportional to the amount of endotoxin added,suggesting that there were multiple sites for endotoxin-neutrophil inter-actions. The binding interaction was found to be independent of temper-ature and not inhibited by iodoacetamide, N-ethylmalecimide, or sodiumcyanide. When considered with the results obtained by Graham et al, M
who demonstrated inhibition of endotoxin-stimulated glycolysis by iodo-

584 MORRISON AND ULEVITCH American Journalof Pathology
acetamide but not potassium cyanide, these results indicate that bindingof endotoxin to the neutrophil is a passive event and not itself a sufficientstimulus to initiate neutrophil responses. The binding was inhibited bynormal rabbit serum, sodium oxalate, and EDTA, results consistent withthe functional observations made by Park and Good 2" and Goihman-Yahr et al.2" It was further reported that the interaction of endotoxinswith neutrophils led to endotoxin detoxification, and a role of neutrophil-derived lipases was suggested. This conclusion is consistent with theearlier report of Collins and Wood 2m that neutrophils could inactivateendotoxins.
In experiments designed to probe the biochemical nature of the mate-rial in human neutrophils (and platelets and monocytes) responsible forthe binding of endotoxins, Springer and Adye 27extracted preparations ofhuman leukocytes with various solvents. These authors demonstrated thatthe majority of endotoxin-binding substances were primarily localized onthe surface membranes of the leukocytes and could be extracted inton-butanol, suggesting that neither proteins, carbohydrates, or nucleic acidswere involved in the binding. Further studies'20-2 have provided evi-dence for a major role of glycerophosphatides in the binding interaction.
In addition to these direct effects of endotoxins on neutrophils, theinteraction of endotoxins with serum can generate factors which influencethe response of neutrophils. In most instances these factors have beendemonstrated to be generated via endotoxin-initiated complement activa-tion. Thus, the experiments of Snyderman et al 241 provided evidence thatincubation of endotoxins from S marcescens (Boivin) or V alcalescens(Westphal) with a number of different mammalian serums generated alow-molecular-weight factor which was chemotactic for rabbit peritonealexudate neutrophils. Heat-stable chemotactic activity could be generatedwith concentrations of endotoxin in serum as low as 0.5 gg/ml, althoughthe endotoxin alone had no chemotactic activity. Incubation at 0 C or inEDTA or using serum preheated to 56 C for 30 minutes markedly reducedthe ability of endotoxin to generate chemotactic activity, and the extent ofactivity correlated well with loss of hemolytic activity, suggesting a rolefor complement components. Support for this conclusion was derivedfrom experiments using mouse serum deficient in C5, in which no chem-otactic activity was generated. No activity was generated on interaction ofdetoxified endotoxins with serum unless antibody specific to the endotoxinwas added. (Such detoxified preparations of endotoxin have no intrinsicanticomplementary activity. See Endotoxin-Complement Interactions InVitro). Further experiments by Snyderman et al "2 have provided evi-dence that the serum chemotactic factor generated by endotoxins is the

Vo. 93, No. 2 BACTERIAL ENDOTOXINS 585November 1978
anaphylatoxin polypeptide C5a derived from complement component C5.More recent experiments by Sveen,2S using endotoxins from Bacteriodesand Fusobacterium as well as Veillonella at concentrations as low as 0.01ug/ml, have confirmed these results.Although endotoxins have the capacity to induce neutrophil chemotaxis
by an interaction with serum complement, results obtained by severalinvestigators indicate that endotoxins can also interact directly with neu-trophils to inhibit chemotactic responses. As first demonstrated by Goetzland Austen,2" incubation of human neutrophils (or monocytes) with Ecoli 026:B6 (Boivin) endotoxin (0.2 ug/ml) for periods up to 1 hourinitiated the release of a mediator with the capacity to inhibit neutrophilchemotactic responses to a variety of diverse stimuli, including autologousserum, anaphylatoxin C5a, and kallikrein. This factor, termied "'NIF"'(neutrophil-immobilizing factor), was demonstrated to exist preformedwithin the cell, to have a molecular weight of approximately 5000, and tobe sensitive to proteolytic enzymes.More recently, Issekutz and Biggar 2" described the results of similar
experiments examining the effect of E coli 0111:B4 endotoxin on thechemotactic responses of human neutrophils. The results of these experi-ments suggested that exposure of human peripheral blood neutrophils toeven minute concentrations of endotoxin (as little as 0.1 ng/ml) for shortperiods desensitized them to a subsequent response to chemotactic factorsderived from incubation of aggregated human Y-globulin with serum.No effect of endotoxins was demonstrated on the chemotactic response ofneutrophils to bacterial chemotactic factors. In addition, these authorswere unable to detect any influence of endotoxin concentrations as high as0.2 jg/ml (which completely inhibited the chemotactic response to serumfactors) on hexose monophosphate shunt activity in either resting neutro-phils or neutrophils phagocytosing latex particles. The relationship of thisinhibitory activity induced by endotoxins to the neutrophil-immobilizingfactor described by Goetzl and Austen 2" remains, however, to be clearlyestablished.
It thus appears, on the basis of these in vitro studies, that endotoxinscan both elicit a variety of responses in neutrophils as well as alter normalneutrophil function. With regard to the former, endotoxins initiate neu-trophil metabolic activity and the expression and/or release of neutrophil-associated enzymes. Indirectly, via serum complement, endotoxins affectneutrophil mobility. Endotoxins alter the capacity of neutrophils tophagocytose and to respond to other chemotactic stimuli. Significantly,the amounts of endotoxin required to initiate neutrophil responses may beas low as those demonstrated to affect macrophages and monocytes. (See

586 MORRISON AND ULEVITCH American Joumalof Pathology
Macrophage/Monocyte-Endotoxin Interactions.) In addition, such con-centrations have been demonstrated in vivo to profoundly alter circulat-ing neutrophils. The exact nature of the relationship between these invitro neutrophil-endotoxin interactions to the in vivo host response tobacterial endotoxins remains yet to be fully clarified.
M_q~Mo diilwn
The mononuclear phagocyte, eg, the tissue macrophage or blood mono-cyte, appears to be crucial in "policing" the internal environment and,therefore, it is not surprising that bacterial endotoxins elicit markedresponses from these cells. Although the phagocytic properties of themononuclear cells have been recognized since Metchnikoffs time, it hasonly become apparent in recent years that the mononuclear phagocytecan be stimulated to release an impressive array of products which canmarkedly alter surrounding cells and tissues. Among the mediators re-leased from these cells in response to endotoxin challenge are pro-coagulant substances, collagenase, pyrogen(s), prostaglandins, colony-stimulating factor, and factors cytotoxic or cytolytic for tumor cells.Because of the extraordinary sensitivity of these cells to endotoxins, eg,many responses have been observed with the addition of submicrogramquantities of endotoxin in in vitro systems, these cells may play a crucialrole in vivo in the production of substances which mediate the diversebiologic effects of endotoxin. Thus, studies of the mechanism of endo-toxin/macrophage interactions are essential to the understanding of thehost response to endotoxin. A brief review of the effects of endotoxins onperitoneal cells (macrophages) was presented by Allison et al in 1973.2Recently an excellent review of studies of the biochemistry and physiologyof the secretory function of the mononuclear phagocyte has been pub-lished by Unanue.?47The first studies of mononuclear cell/endotoxin interaction focused on
the effect of endotoxin on morphologic and biochemical properties ofthese cells. In 1965, Cohn and Benson 2 demonstrated that, 4 days afterthe injection of 20 ug of S abortus equi into the peritoneal cavity of mice,the accumulation of a population of mononuclear phagocytes, largelymacrophages, was observed. The biochemical characterization of thesecells indicated that the content of at least three enzymes (acid phospha-tase, ,B-glucuronidase, and cathepsin) was increased markedly comparedwith controls, suggesting that the endotoxin had "activated" these cells.Whether these effects resulted from a direct action of endotoxin on themacrophage or were mediated through other endotoxin-induced changescould not be determined.

Vol. 93, No. 2 BACTERIAL ENDOTOXINS 587November 1978
However, in the same year, Bennett and Cohn,29 using cultured mono-cytes in vitro, showed that the addition of as little as 0.1 g/ml of Sabortus equi endotoxin to purified equine blood monocytes markedlyenhanced the ability of these cells to phagocytose bacteria and take upcolloidal gold. In addition, they reported that the endotoxin-treated cellsunderwent marked changes in both shape and cytoplasmic organization.Large numbers of dense granules were observed to form after the additionof endotoxin; these authors attributed this to increased pinocytoxic activ-ity. Endotoxin produced alterations in the cell membrane of the stimu-lated cell; these alterations were characterized by a marked ruffling of themembrane. These authors also suggested that endotoxin may stimulatethe formation of lysosomal hydrolases. Thus, these data showed that theeffects of endotoxin on the mononuclear phagocyte could be demon-strable in an in vitro system. However, in these studies, unheated 40%ofetal calf serum was included in the culture medium. Therefore, thepossibility that the endotoxin effect was not a direct effect on the cell butrather was induced by a product of an endotoxin/serum mediation systeminteraction, ie, complement, was not excluded.
In 1968, Wiener and Levanon 2 explored the effect of S abortus equiendotoxin on murine peritoneal monocytes in an in vitro system. Theseauthors employed as little as 5% or as much as 20% heat-activated (at 60 Cfor 20 minutes) fetal calf serum in the medium and showed that theaddition of submicrogram amounts of endotoxin markedly stimulated thesynthesis of acid phosphatase. Thus, these experiments provided betterevidence that the effect of endotoxin on the differentiation of the mono-cyte resulted from a direct action on the cells. In these studies the fetalcalf serum did not have a functionally active complement system. How-ever, these authors acknowledged the value of a serum-free in vitroculture system to study the effects of endotoxin on the mononuclearphagocyte.
Allison et al 2" further characterized the activation of murine peritonealexudate cells cultures in vitro using a Boivin preparation of E coli 055: B5endotoxin. These investigators showed that 72 hours after the addition of10 ,g/ml of endotoxin both the intracellular activity of certain enzymes aswell as the amount of enzyme released to the medium increased. Forexample, the total content of the lysosomal acid hydrolases acid napthyl-phosphatase and ,-N-acetylglucosaminidase was elevated 1.5- to 3-foldover control cell levels. In contrast, no changes in the content of two otherlysosomal enzymes, ,-glucuronidase and fl-galactosidase, was detected.Endotoxin treatment also led to a 2-fold stimulation in the levels of twocytoplasmic enzymes, lactic dehydrogenase and leucine naphthylamidase,

588 MORRISON AND ULEVITCH American Journalof Pathology
without concomitant loss to the medium. Thus, endotoxin appeared toselectively produce increased synthesis of certain enzymes and, in addi-tion, promoted the specific exocytosis of the lysosomal acid hydrolases.The release of these enzymes apparently was a function of viable cellssince no release of cytoplasmic enzymes, eg, lactic dehydrogenase, wasnoted.
Although the biologic significance of this mechanism was not clear, theauthors 2" suggested that this mechanism may be operative in diseases inwhich bacterial products play a major role.
In a similar series of experiments reported by Doe et al n' using murineperitoneal exudate cells, endotoxins from E coli 0111 : B4 or 0113 and fromS minnesota R595 were shown to be effective at concentrations from 0.2 to25 ug/ml in stimulating the release of lysosomal enzymes into the me-dium. They also reported that the intracellular lactic dehydrogenaseconcentration increased markedly and was not liberated into the mediumexcept at the highest endotoxin concentration. In contrast, lysozymesecretion was not altered by endotoxin; this finding is in keeping with theconcept that secretion of lysozyme is independent of the activity of thecell.20More recently, Bianco and Edelson 2'2 demonstrated an inability of S
abortus equi endotoxin to induce "activated" macrophages followingintraperitoneal injection of up to 300 Ag into the C3H/HeJ mouse. Incontrast, the intraperitoneal injection of Brewers thioglycollate medium,another agent commonly used to produce "activated" macrophages invivo, was effective in the C3H/HeJ mouse. The endotoxin employed heredid activate the resident mononuclear phagocytes of other strains, includ-ing the NCS, A/J, or C3H Feb/J mice, following intraperitoneal injection.The C3H/HeJ mouse has been shown to respond abnormally to manyendotoxin-induced defects. The abnormal response is due to a singlemutation 25 expressed as a cellular defect in response to endotoxin. Stud-ies of macrophages from the C3H/HeJ mouse may provide insight intothe nature of the interaction between the endotoxin and the macrophage,which leads to cell activation.The mechanism of endotoxin-induced macrophage activation remains
to be elucidated. Central to this question is the development of tech-niques to detect biochemical changes induced by endotoxin prior to thefinal expression of activation. However, despite this gap of knowledge,much is known about the types of mediators elaborated by the endotoxin-stimulated macrophage; some of these recent studies will be summarizedbelow. The capacity of endotoxin to induce tissue-factor production frommacrophages was discussed in the section Extrinsic Coagulation Pathway.

Vol. 93, No. 2 BACTERIAL ENDOTOXINS 589November 1978
Studies related to endotoxin-induced factors from macrophages whichmodulate the immune response will not be discussed here but will beconsidered in a forthcoming review."The macrophage lysosomal system has also been suggested as a partici-
pant in the detoxification of endotoxin. However, because this review isconfined to the production of mediators which may alter the cell andtissues in the vicinity of the macrophage or monocyte, endotoxin detoxifi-cation will not be discussed here.
Collagenase production and release by endotoxin-stimulated macro-phages has been reported in an elegant series of experiments performedby Wahl and his collaborators.2" They showed that guinea pig peritonealexudate cells, elicited by prior injection of Drakeol, could be stimulatedwith as little as 5 Ag/ml of the following endotoxin preparations: lipid Aproduced from acid hydrolysis of S typhimurium endotoxin, the poly-saccharide-deficient endotoxin from S minnesota R595, and the endotoxinisolated from the wild type S minnesota. The addition of a lipid-deficientpreparation of S typhimurium endotoxin prepared by alkaline hydrolysisaccording to the method of Tripodi and Nowotny did not stimulate theproduction of collagenase. These data further support the concept that aninteraction between the lipid region of the endotoxin and the macrophageis responsible for initiating collagenase production. The addition of 1 Ag/ml of cyclohexamide to the macrophages markedly suppressed the releaseof collagenase, clearly suggesting that protein synthesis was required forthe induction of collagenase.
These experiments were performed in serum-free medium. In addition,the authors stated that > 95% of the adherent cells employed in theseexperiments appeared to be macrophages, based on morphologic andfunctional criteria. Thus, the induction of collagenase in these cells mostlikely results from the direct interaction of endotoxin with the cell.More recently Wahl et al 2" demonstrated that prostaglandins may
regulate the endotoxin-induced synthesis of collagenase by peritonealmacrophages. The release of collagenase following the addition of endo-toxin to macrophage cultures was significantly inhibited by 10 AM indo-methacin. It was also shown that endotoxin (E coli 055: B5) stimulated therelease of PGE, by the macrophage, detectable as early as 4 hours afterendotoxin addition. These observations provide insight into the initialevents of endotoxin-induced macrophage activation. They further demon-strated the capacity of endotoxins to induce biologically important sub-stances, eg, prostaglandins, as an early event resulting from a mac-rophage/endotoxin interaction.Another macrophage enzyme induced by endotoxin is the plasminogen

590 MORRISON AND ULEVITCH American Journalof Patology
activator, a serine protease of molecular weight 48,000. The effect ofexposure to endotoxin on the enhancement of the production of plasmino-gen activator by macrophages has been described by Gordon et al.25These authors showed that intraperitoneal injection into female NCSRmice of endotoxin from S abortus equi, as well as isolated lipid A, couldprime peritoneal macrophages to subsequently release plasminogen acti-vator after challenge with latex beads. In addition, the endotoxin injectioncould significantly enhance the production and release of plasminogenactivator in the peritoneal mononuclear phagocytes, although the activityobtained was only 5 to 7% of that produced by the administration ofthioglycollate.
Alexander and Evans,.7 in a pioneering report, demonstrated thatperitoneal exudate cells from DBA/2 mice cultured in the presence of aslittle as 0.02 sg of endotoxin from Shigella or isolated lipid A preparedfrom wild type S minnesota were rendered cytotoxic for [51784 or SL2transplantable lymphoma cell lines. They reported that exposure of theadherent cells for as little as 30 minutes was sufficient to generate the fullycytotoxic macrophage. In these studies, a preparation of purified fungalvirus double-stranded RNA was reported to also render the macrophagescytotoxic. These authors showed that macrophages harvested from theperitoneal cavities of mice previously treated with BCG or thioglycollatewere "activated" by morphologic and biochemical criteria but did notbecome cytotoxic until exposure to endotoxin or double-stranded RNA.
Subsequently, Currie and Basham 2u showed that a population of ad-herent peritoneal exudate cells from Hooded and August rats could berendered cytolytic for rat sarcoma cells by prior exposure to endotoxin.For these experiments, 25 ug/ml of a Boivin preparation of S typhosaendotoxin was employed. These authors showed that the cytolytic activitywas only expressed against transformed cells and that a cytolytic factorcould be demonstrated in the culture supernatants from endotoxin-treatedcells.More recently, Doe and Henson 2' confirmed and extended the pre-
vious reports demonstrating that endotoxin will stimulate peritoneal ex-udate cells to become cytolytic for tumor cells. Doe and Henson employedendotoxin isolated from S minnesota R595 to show that prior exposure ofmurine peritoneal exudate cells to endotoxin stimulated these cells to killtumor cells in vitro. The lysis was nonspecific since both allogeneic andsyngeneic cells were susceptible. In this study it was established thatneither B or T lymphocytes were required to produce the endotoxinstimulation of the mononuclear phagocytes. These authors were unable todemonstrate the presence of a supernatant factor cytolytic for tumor cells.

Vol. 93, No. 2 BACTERIAL ENDOTOXINS 591November 1978
In an accompanying report, Doe et al "O investigated the structuralfeatures of the endotoxin required to generate a macrophage capable oflysing tumor cells. To do this, these investigators compared the capacity ofa variety of endotoxin preparations, as well as isolated lipid A preparedfrom either E coli 0111: B4 or S typhimurium R60 endotoxin, to stimulatemacrophages in vitro to kill tumor cells. These authors showed thatendotoxin preparations prepared by either phenol-water, butanol-water,phenol-chloroform-petroleum-ether or by Boivin extraction proceduresstimulated macrophages in vitro to kill tumor cells. Thus, this activityappeared to be a general property of all endotoxins and was not depen-dent on the method of endotoxin preparation. That a lipid A/macrophageinteraction was the stimulating signal to produce a macrophage withcytolytic activity was shown by three experimental means: First, lipid Aisolated from mild acid hydrolysis of endotoxin from S typhimurium R60as well as from E coli 0111: B4 stimulated the macrophages to becomecytolytic for tumor cells. Second, the stimulating effect of lipid A andendotoxins from S minnesota R595 and from E coli 0111 : B4 (Westphal)was abrogated by prior exposure of the endotoxin to polymyxin B. Finally,the authors showed that, although endotoxin from E coli 0113 was effec-tive at stimulating macrophages when added at concentrations as low as40 ng/ml, the lipid-free native protoplasmic polysaccharide from E coli0113 was not stimulatory. The native protoplasmic polysaccharide did notblock the stimulatory effect of the intact 0113 endotoxin. These combineddata provide evidence to support the concept that the lipid A/mac-rophage interaction is the determinant event. When partially purifiedpreparations of lipid A-associated protein (LAP) were examined for thecapacity to stimulate murine macrophages to become cytolytic for tumorcells, it was found that addition to 1 to 5 yg/ml of this material wasstimulatory. Thus, two components of the bacterial endotoxin complex,lipid-A-associated protein and lipid A, appear to stimulate murine macro-phages.Of great interest has been the recent report of Russell et al "I demon-
strating that marcophages obtained from BALBc/ANCr mice with pro-gressing or regressing Moloney sarcoma tumors were exquisitely sensitiveto the stimulatory effects of endotoxin added to these cells in vitro. In thecase of macrophages obtained from progressing tumors, as little as 1 ng/ml of the lipid-A-rich fraction of endotoxin prepared from E coli 0111: N4(Westphal) would markedly stimulate the cytolytic activity of these cells.Similar findings were obtained when macrophages explanted from re-gressing tumors were maintained in culture 24 hours prior to endotoxinaddition. Examination of freshly prepared tumor macrophages also dem-

592 MORRISON AND ULEVITCH American Journalof Pathology
onstrated that the basal level of cytolysis could be markedly enhanced byendotoxin addition. In contrast, thioglycollate elicited peritoneal exudatecells from normal mice or mice with 7- to 14-day progressing or regressingMoloney sarcomas were not stimulated by endotoxin until 100- to 1000-fold higher concentrations were employed. These experiments demon-strate the potent effect of endotoxin on an already primed macrophage, inthis case primed by undefined events occurring in the mileau of thetumor.
Hibbs et al 20 also have recently reported that macrophages from BCGor Toxoplasma gondii infected mice can be stimulated to kill tumor cellswhen exposed to 1 to 5 ng/ml of endotoxin from E coli 0128:B12. Ofinterest was the observation that the prior treatment of macrophages withtrypsin abolished the cytotoxic activity induced by endotoxin. Thus, atrypsin-sensitive cell component, possibly a membrane protein, may beessential to transmit the signal delivered to the macrophage by endotoxin.The exposure of murine blood monocytes, spleen cells, or peritoneal
exudate cells to endotoxin results in the secretion of a product, ie, colony-stimulating factor (CSF), necessary for the growth of colonies of gran-ulocytic and monocytic cells in vitro. In 1972, Hays et al 2" reported thatendotoxin injection into mice elicited the formation of CSF within min-utes after injection. They further reported that addition of endotoxin tothose fractions of blood leukocytes containing a monocyte populationresulted in CSF production.
Subsequently, two reports appeared in 1974 demonstrating that CSFcould be produced in vitro from a mononuclear phagocyte by exposure tosubmicrogram quantities of endotoxin. Eaves and Bruce 2" showed thatadherent peritoneal exudate cells from female A/KR mice would releaseCSF after exposure to as little as 0.01 Mg of endotoxin from a Boivinextract of S typhosa. The incubation of cells with endotoxin was per-formed in serum-free medium. They characterized the cell responsible asbelonging to a glass-adherent population of large mononuclear cellswhich were also capable of phagocytosing carbon particles. In addition,they showed that dissociated lung tissue was a rich source of cells whichcould be stimulated by endotoxin to produce CSF, leading to the specula-tion that alveolar macrophages might also produce CSF.
Ruscetti and Chervenick 2" showed that human blood monocytes couldbe stimulated in vitro by as little as 0.01 ;g of a Boivin preparation of Styphosa endotoxin. These experiments also provided evidence that neitherpolymorphonuclear leukocytes nor blood lymphocytes could be stimulatedby endotoxin in vitro to produce CSF.Apte et al investigated the cellular requirements for endotoxin-

Vol. 93, No. 2 BACTERIAL ENDOTOXINS 593November 1978
induced CSF secretion by murine spleen cells. Their results suggest thatCSF was generated by a spleen cell population characterized as a non-dividing, terminally differentiated cell that did not need active DNAsynthesis to produce CSF. They further demonstrated a requirement forboth active RNA and protein synthesis in this process. It is of interest tonote that a complex of endotoxin from E coli 055:B5 (Westphal) andpolymyxin B was effective at inducing CSF in in vitro cultures of murinespleen cells, although the mitogenic activity of the 055: B5 endotoxin wasabrogated by treatment with polymyxin B. These findings are of particularinterest but are difficult to reconcile with previous work by Apte, Galanos,and Pluznik 2a; they reported that the signal for CSF secretion resided inthe lipid A moiety of the endotoxin. However, as described below, a rolefor the carbohydrate portion of the endotoxin in CSF production has beenpreviously suggested.2" A final point made by Apte et al N6 suggested thatCSF production by spleen cells required cooperation by both spleniclymphocytes and macrophages, although no data were presented to sup-port this.A recent report by Nowotny et al 2w indicated that the carbohydrate
moiety of the LPS, in particular the core polysaccharide, was responsiblefor the induction of CSF following intraperitoneal injection of endotoxininto ICR mice. They prepared a lipid-A-free, polysaccharide-rich fractionfrom the supematant of an endotoxin preparation from S minnesota 1114treated with I.ON HCI at 100 C for 35 minutes. This material was fullyactive in the induction of CSF in vivo. These results demand that addi-tional experiments be performed to establish the chemical nature of thesignal delivered by endotoxin to initiate CSF production.
Several observations suggest that prostaglandins are released followingthe administration of endotoxin to experimental animals.2"'"' In a recentstudy, Fletcher et al V2 showed that concentrations of prostaglandin E andF were increased in arterial and mixed venous blood following the in-travenous injection of endotoxin in baboons. The cellular source(s) of thesesubstances has not been identified. However, recent studies of both mac-rophages and blood monocytes suggested that endotoxins can elicit thesynthesis of prostaglandins in these cell types.
Rosenstreich et al 273 showed that macrophages from C3H/HeN micesynthesized and released large amounts of both PGE2 and PGF2 28 hoursafter exposure to endotoxin prepared from E coli K235 endotoxin (West-phal). In contrast, macrophages from endotoxin nonresponder C3H/HeJmice did not produce either PGE, or PGF2, after the addition of K235endotoxin.
Kurland and Bockman 274 examined the ability of murine peritoneal

594 MORRISON AND ULEVITCH American Journalof Pahology
exudate cells and murine macrophage and nonmacrophage cell lines toproduce PGE1 and PGE2 after exposure to endotoxin. They demonstratedthat the adherent population of murine peritoneal exudate cells fromB6D2F1 mice, judged to be >99% macrophages by biochemical criteria,produced detectable PGE in response to as little as 0.1 sg of S typhosaendotoxin (Westphal). Maximal stimulation of 1 X 10' cells was obtainedwith 10 Ag of this endotoxin preparation. These authors further showedthat neoplastic mononuclear phagocyte cell lines could be stimulated toproduce PGE after exposure to 1 gg of S typhosa endotoxin. T- and B-celllymphoma cell lines were also examined and, in contrast, were not stimu-lated by endotoxin to produce PGE.
Kurland and Bockman "4 additionally showed that human blood mono-cytes are constitutive producers of PGE but that neither polymorphonu-clear leukocytes nor human blood lymphocytes performed this function invitro. Unfortunately, the effects of endotoxins on PGE synthesis by humanblood cells were not examined.
It is tempting to speculate that the macrophage or monocyte may play acentral role in the elaboration of mediators responsible for the manydiverse effects of endotoxins in vivo. Evidence for this derives from recentexperiments reported by Rosenstreich et al 'M and Glode et al,2'" in whichthey showed that the C3H/HeJ mouse could be rendered more sensitiveto the lethal effects of endotoxin by transfer of spleen cells from C3H/HeN mouse into a sublethally irradiated C3H/HeJ recipient. Rosen-streich et al 273 stated, without presenting the data, that the macrophagepopulation may be responsible for the enhanced lethality to endotoxins.These data should stimulate a great deal of experimentation designed toelucidate the role of the macrophage in the expression of the lethal effectsof endotoxin.Humes et al 276 showed that resident murine peritoneal macrophages
synthesize prostaglandins in response to inflammatory stimuli. Of particu-lar interest was the demonstration that the macrophage will convertarachidonic acid into PGI2 (prostacyclin); in fact, these authors suggestedthat this prostaglandin may be a major secretory product of these cells. Inview of the potent hemodynamic and hematologic effects of PGI2, anevaluation of the effect of endotoxin on PGI2 synthesis by macrophages iswarranted.
Eb-dof Ch4-ndotxi ~mita
In 1924, Sanarelli, describing the pathologic changes resulting from twoproperly timed injections of endotoxin, coined the term "'epithalaxie,'meaning breakdown of the endothelium, as a characteristic of the tissue

Vol. 93, No. 2 BACTERIAL ENDOTOXINS 595Novembe 1978
injury he observed. Whether this is a cause or a result of endotoxin-initiated changes is still unknown, although most workers would supportthe proposal that damage to endothelial cells could lead to the initiation ofa variety of pathologic and pathophysiologic changes. Evidence to suggesta role for the endothelial cell has derived from the results of two experi-mental approaches: a) the demonstration that endotoxins are localized inthe endothelial cells after intravenous injection and b) the demonstrationthat endothelial cells are damaged following the intravenous injection ofendotoxin. Studies which have implicated the endothelial cell in thepathogenesis of endotoxin-induced injury will be described below as willpossible mechanisms by which endotoxin/endothelial cell interaction maycontribute to injury.
Perhaps the first demonstration that endotoxins may interact directlywith endothelial cells was reported in 1957 by Tanaka et al,27 who showedby indirect immunofluorescence that a Boivin preparation of endotoxinfrom S entertidis could be detected in capillary endothelium of the liver,lungs, intestine, and, to a limited degree, kidney within 3 hours afterinjection into mice. These investigators also detected positive fluorescencein the endothelium of the major vessels of the liver. Five years later, in1962, Rubenstein et al,m employing a Boivin preparation of E coli0111:B4 endotoxin and indirect immunofluoresence techniques, examinedthe cellular localization of endotoxin following intravenous injection intodogs. These investigators described the localization of endotoxin in theendothelium of capillaries and of small and medium-sized veins of avariety of tissues, including liver, spleen, and jejunum as early as 10minutes after injection. It is of interest that no endotoxin was detected inthe major arteries and veins and that, although capillary and venuleendothelium was frequently involved, only rarely was endotoxin detectedin the arteriolar endothelium. Since both Tanaka et al 2 and Rubensteinand his co-workers 271 employed Boivin preparations, we cannot be certainif the immunofluorescent material localized in the endothelium representsLAP, LAP/endotoxin complexes, or endotoxin devoid of LAP. Studieswith protein-free LPS as well as isolated LAP might clarify this question.More recent work has focused on characterizing the extent and mecha-
nism of injury to the endothelium following endotoxin injection. McGrathand Stewart m showed that within 1 hour after the intracardiac injectionof 7.5 mg of endotoxin into rabbits, the endothelial cells of mesentericarteries were severely damaged. The damage was characterized by dis-torted nuclei, apparent nuclear vacuolization, and missing nuclei. Noattempt was made to detect the presence of endotoxin in the damagedcells; the large amount of endotoxin administered in these studies leaves

596 MORRISON AND ULEVITCH American Journalof Pathoogy
many questions about the pathogenic significance of the changes reportedhere.
However, Gaynor et al I reexamined this question using rabbits givena single intravenous injection of 40 to 120 ug/kg of E coli 0127:B8endotoxin. These investigators were able to demonstrate circulating endo-thelial cells in the blood within 5 minutes after endotoxin injection. Thisapparent stripping of endothelial cells from vessel walls could not beinhibited by concurrent heparin anticoagulation. Additional damage toendothelium was observed by histologic examination of segments of aorta;in 6 of 9 endotoxin-treated animals, morphologic abnormalities character-istic of endothelial damage were noted.
In contrast to these studies, McKay et al 182 were unable to detect anyevidence for endothelial cell damage produced by the injection of 10 mg/kg of endotoxin into rhesus monkeys. These authors examined tissuesobtained at 15 minutes and 1, 2, 3, and 4 hours after endotoxin and,although they demonstrated fibrin in a variety of tissues, electron micro-scopic examination failed to show endothelial cell damage. Whether theinability to detect damage to endothelial cells in the rhesus monkeyreflects species differences, differences between various preparations ofendotoxin employed, or other factors is not understood.
However, because of the relative lack of sensitivity in quantitatingendothelial injury solely by morphologic examination, alternative approaches have been developed. The use of 3H-thymidine and subsequentautoradiography to demonstrate endothelial cell turnover has been em-ployed by several investigators to assess endotoxin-induced injury,281'"and this technique has been shown to provide a more sensitive measure ofendothelial injury. Recently, Evensen et al 2" used this technique toexamine the role of the complement system in the endotoxin-inducedendothelial cell injury observed in the rat. As previously described, endo-toxin is a potent activator of the classic as well as the alternative pathwayof complement. Any one of the multiple biologic activities resulting fromcomplement activation could be responsible for the endotoxin-inducedendothelial cell injury. Evensen et al 2u compared the number of DNAsynthesizing cells in thoracic aorta strips in normocomplementemic andC3-depleted rats injected with either 50 or 500 ig of a Boivin preparationof E coli 0111:B4 endotoxin. They found that the number of endothelialcells demonstrating labeled nuclei were identical in the normocomple-mentemic and C3-depleted rats. Thus, neither direct effects of biologi-cally active products derived from complement activation or substancesreleased from other mediator cells in complement-dependent reactionsappear to be required for the endothelial injury. These findings are also

Vol. 93, No. 2 BACTERIAL ENDOTOXINS 597November 1978
consistent with the lack of effect of C3 depletion on a number of endo-toxin-induced pathophysiologic changes previously observed in the rab-bit, dog, and cat.More recently, the report of Raiji, Keane, and Michael 28demonstrated
that in situ perfusion of one kidney with endotoxin, followed by systemicadministration of a second dose of endotoxin in 24 hours, would elicit aunilateral Schwartzman reaction in the perfused kidney. These authorssuggested that the initial event in the development of renal corticalnecrosis might be a local effect of endotoxin in the vascular endotheliumof the kidney. This fascinating report should provide impetus to furtherstudies of the role of the endothelial cell in the development of endotoxin-induced injury.The biochemistry of the endothelial cell has been studied extensively
during the past several years, and it is apparent that this cell contains avariety of potentially injurious mediators. For example, Maclntyre et al 286have shown that the endothelial cell is capable of synthesizing pros-tacyclin (PGI2), a metabolite of prostaglandin endoperoxides, which is apotent inhibitor of platelet aggregation as well as a potent hypotensiveagent.2" The recent reports of Howes 2 and Fletcher et al 272 on theamelioratory effect of drugs which inhibit prostaglandin synthesis inexperimental models of endotoxin-induced shock and DIC suggest thatprostaglandins may be involved in the mediation of endotoxin-inducedinjury. The endothelial cell may play an important role in the elaborationof the prostaglandin mediators. Alternatively, damage to the endothelialcell could expose vascular basement membrane and provide a site atwhich Hageman factor could become activated. Should this occur, notonly would localized coagulation be initiated but also the elaboration ofpotent vasoactive substances such as bradykinin might result. Thus, theoriginal injury could be markedly amplified through a host of othermediators with injury to the endothelial cell playing a critical, primaryrole in these events.
M c _C. lD
The potential contribution of mast cells and basophils to the profoundvascular changes which occur during anaphylactic shock have been welldocumented.286 In spite of the very early suggestion of Sanarelli 117 thatendotoxin toxicity is not necessarily dependent on anaphylactic reactivity,it has been tempting to ascribe a contribution of mast cells and basophilsto the pathophysiologic alterations induced in vivo by endotoxins. Thishas, in part, resulted from the observations of many investigators 19
who have delineated a number of similarities between anaphylactic and

598 MORRISON AND ULEVITCH American Journalof Pathology
endotoxic shock. Nevertheless, in spite of a number of attempts to estab-lish such a contributory role of these cells in response to endotoxins invivo, the available evidence remains at best equivocable.
Mast Cell-Endotoxin Interactions In Vivo
Early experiments by Glick and Pothapragada 1 attempted to definealterations in metabolic processes in mast cells following intraperitonealinjection of endotoxin (E coli, 0.5 to 5 mg). Two hours after administra-tion of endotoxin, these authors observed significantly elevated levels ofsuccinic dehydrogenase in Ficoll-Hypaque purified mast cells comparedwith controls. It was, however, not ascertained whether these mast cellswere also degranulated, although it should be pointed out that similarincreases were observed in rats given compound 48/80, a mast cell de-granulating agent. Similarly, Antweiler and Hirsch 29 were unable todetect any direct disruptive effect on rat peritoneal mast cells followinginjection of 0.1 to 10 jig of S abortus equi or E coli 011l:B4 endotoxins.An extensive series of experiments by Gustafson and Cronberg 2" exam-
ined the effects of endotoxin (E coli 026:B6-Boivin) on mast cells andtheir relationship to the local Shwartzmnan reaction in the hamster. Theresults of these studies failed to demonstrate any effect of an intra-peritoneal injection of endotoxin on either peritoneal fluid or mesentericmast cells. Additionally, injection of endotoxin into hamster cheek pouchdid not alter either the histologic appearance or the number of residentmast cells. Additionally, injection of endotoxin into hamster cheek pouchdid not alter either the histologic appearance or the number of residentmast cells. In the local Shwartzman reaction induced in hamster cheekpouch, these authors did observe mast cell disruption between 5 and 6experiments were performed to induce the local Shwartzman in the cheekpouch previously depleted of mast cells by treatment with distilled water.No differences were observed between mast-cell-depleted and controlreactions. The authors concluded from these studies that mast cell altera-tions did not seem to be an essential or a primary effect in endotoxinreactions. Somewhat contrasting results were obtained by Hook et al,2'4who observed a decline in both mast cell count and available histaminecontent of hamster peritoneal cells 4 to 14 days following injection ofV alcalescens (Westphal) endotoxin.
Several more investigators, in addition, have detected alterations inmast cells following administration of endotoxins. Urbaschek,m9 for ex-ample, noted degranulation of 'mesenteric mast cells following intra-venous injection of endotoxins. Experiments by Jokay and Karczag.96suggested that, following intraperitoneal injection of endotoxin into mice,

Vol. 93. No. 2 BACTERIAL ENDOTOXINS 599November 1978
the number of mast cells recovered from the peritoneal cavity decreasesapproximately 70% over an 18-hour period. Further studies by theseauthors "7 indicated that over a 5-hour period the number of "non-intact"mast cells in the peritoneal cavity increased significantly, These eventswere accompanied by an increase in permeability of the peritoneal cavity.Because of the significantly delayed response observed in comparison toother mast cell stimuli, eg, antigen-IgE antibody, the authors concludedthat the endotoxin-mediated effects on the peritoneal mast cells were mostlikely not direct.
Mast Cell-Endotoxin Interactions In Vitro
There is virtually no evidence to indicate that bacterial endotoxins haveany direct effect on mast cells or basophils in vitro. Unfortunately, how-ever, in vitro systems to study mast cell and basophil responses (degranu-lation and release of vasoactive amines) have generally focused on rat ormouse peritoneal cells and peripheral blood leukocytes, respectively.Since these particular cells from various species may respond differently toendotoxins, the results of the limited studies reviewed here should not beconsidered as definitive negative evidence for mast cell/basophil re-sponses to endotoxins.The experiments of Asboe-Hansen and Glick "' first examined the in
vitro response of rat peritoneal mast cell suspensions to an E coli (Boivin)endotoxin. These authors were unable to detect any influence of endo-toxin on mast cells. Similar results were obtained by Sandusky et al '9using either an E coli 0127:B8 (Boivin) or (Westphal) prepared endotoxin,even at concentrations as high as 1.0 mg/ml. These conclusions werereached whether the investigators used rat peritoneal cells or chopped ratlung fragments as a source of mast cells. Further support for theseconclusions was provided by the experiments of Morrision and Betz,',who similarly showed that concentrations of E coli 0111:B4 endotoxin(Westphal or Butanol) even as high as 1.0 mg/ml were ineffective atinitiating secretion from rat peritoneal mast cells.
Additional experiments by these latter investigators,' however, dem-onstrated that, although endotoxins themselves were unable to stimulaterat peritoneal mast cells, the isolated protein component (LAP-lipid-A-associated protein) of the endotoxin was, at the low concentration (1 to 10;g/ml), able to initiate the noncytotoxic secretion of vasoactive amines.The secretion required calcium and cellular energy and was inhibited at47 C, demonstrating a true secretory response. The mast cell response wasshown to be independent of lymphocytes or macrophages in peritonealfluids, suggesting a direct effect on the mast cells. At high concentrations

600 MORRISON AND ULEVITCH American Journalof Pathology
(>100 gg/ml), LAP was shown to be cytotoxic to the mast cells (as well asthe other cells in the peritoneal fluids). These authors postulated that theappropriate in vivo dissociation of endotoxins into the constituent proteinand LPS fractions might provide a potent mast cell degranulating agent.
In addition to direct effects of endotoxins on mast cells, indirect effectsmodulated by serum proteins, in particular the anaphylatoxin poly-peptides C3a and C5a, generated during the activation of serum com-plement would also be expected to contribute to mast cell and basophilresponses. These polypeptides are known to initiate secretion from mastcells and basophils,Im and the ability of endotoxins to activate com-plement has been well characterized. (See Serum Complement). The firstdemonstration of anaphylatoxin production by the interaction of endo-toxin with rat plasma was reported by Greisman,30 who showed thegeneration of a factor in serum which could contract guinea pig ileum.Studies by Lichtenstein et al,302 using V alcalescens endotoxin in guineapig serum further characterized the anaphylatoxin production. Theseauthors showed that no activity was produced in heated serum, in serumincubated at 0 C, or in the presence of EDTA. Additional studies charac-terized the guinea pig ileum response as tachyphylactic and inhibitable byantihistamines. These studies were later confirmed by Dierich et al,1°" whoshowed that anaphylatoxin production was independent of the presenceof polysaccharide in the endotoxin preparation.The relationship of anaphylatoxin production by endotoxins to activa-
tion of mast cells or basophils has not been pursued in great detail. Theexperiments of Antweiler and Hirsch,292 using concentrations of endotoxin(S abortus equi, E. coli 0111:B4 0.1 to 10 Mg!5 ml), which they estimatedto be in excess of those which might reasonably be found in serum, wereunable to detect any release of histamine from rabbit anticoagulatedwhole blood. They concluded that histamine release from formed ele-ments was probably not a significant event in endotoxin interactions withthe host. Sandusky et al,2"9 who demonstrated no direct effect of endo-toxins on rat peritoneal mast cells or chopped lung fragments, were able toinduce histamine release in the latter but not the former system usingserum which had previously been incubated at 37 C with endotoxins for60 minutes.
Another approach to this problem was examined by Hook et al,29 whodemonstrated that the incubation of V alcalescens endotoxin with hamsterserum resulted in the generation of a factor which initiated histaminerelease from hamster peritoneal mast cells following a second incubation.The fact that generation of the factor in serum was inhibited in thepresence of EDTA or in serum heated to 56 C for 30 minutes suggested a

Vol. 93, No. 2 BACTERIAL ENDOTOXINS 601November 1978
relationship to serum complement activation by endotoxin. This con-clusion was supported by studies using detoxified preparations of endo-toxins. These investigators further correlated the generation of histamine-releasing activity with loss of hemolytic complement activity. Additionalstudies by these investigators 3 have shown the active material genera-tion in serum to be either an a2- or pI-globulin with an approximatemolecular weight of 60,000. These characteristics would clearly differenti-ate this material from the classic anaphylatoxins C3a and C5a normallygenerated in serum which, as indicated earlier, are potent mast cellstimuli. The precise nature of the active material and its relationship tocomplement components, particularly the anaphylatoxins, is unclear.More recent experiments by Glauser et al 3 have confirmed these
earlier observations. These authors have, however, provided evidence fora role of cAMP in the histamine release response of the mast cells. In thisrespect, it was shown that sodium dichromoglyeate, isoproterenol, andanimophylline, antagonists of cAMP, significantly inhibited the hamstermast cell response to the endotoxin-generated serum factor.The majority of the available evidence, therefore, would not support a
primary role for mast cell/basophil responses to endotoxins by a directendotoxin cell interaction. The potential participation of these cells inendotoxin-host mediation system interactions via indirect mechanisms, inparticular, the complement system, would be suggested on the basis ofseveral experiments. The net contribution of such interactions to bepathophysiologic changes brought about by endotoxins remains to becritically evaluated.
SUmary and Concusions
The accumulated data which we have endeavored to summarize in thisreview delineate the profound multiple biologic effects which bacterialendotoxins can exert on host mediation systems and reaffirm the observa-tion of Thomas57 that endotoxins will "turn on every defense at ourdisposal." Significant effects of endotoxins on coagulation and comple-ment, on platelets, polymorphonuclear leukocytes and endothelial cells,and especially on macrophages/monocytes and perhaps to a lesser extentmast cells have been confirmed both in vitro and in vivo. Responses havebeen demonstrated to be the result of both direct endotoxin/mediatorsystem interactions as well as indirect effects. Significant variations havebeen demonstrated in the biologic responses to the endotoxins preparedfrom different gram-negative organisms as well as to endotoxins preparedfrom the same organism by different extraction methods. These observa-

602 MORRISON AND ULEVITCH American-Journalof Pathology
tions emphasize the inherent differences in various endotoxins and under-score the need to define endotoxin preparations used in different experi-mental situations.
Virtually all of the mediation systems examined appear to require thelipid A region of the lipopolysaccharide portion of the endotoxin complex.This would include activation of the classic pathway of complement by aninteraction with Cl, activation of the intrinsic coagulation pathway by aninteraction with Factor XII, binding to platelets, and stimulation ofneutrophils and macrophages/monocytes. Notable exceptions to this arealternative complement pathway activation and generation of colony-stimulating factor, in whichla-prominent role for polysaccharide has beensuggested. The current understanding of the multiple biologic activities ofthe lipid A portion of bacterial endotoxins has benefitted tremendouslyfrom the comprehensive chemical studies on lipid A performed by West-phal, Luderitz, and their co-workers. These investigators have defined thechemical structure of the lipid A molecule. Without this information,studies to define the molecular mechanisms of endotoxin action would bevirtually impossible.
Nevertheless, in spite of the impressive accomplishments which havedefined and characterized the biologic responses of host-mediation sys-tems to endotoxins, much remains to be done. In this respect, two majorareas of research are essential: The first involves a more precise delinea-tion of the potential contribution of the interaction of the various hostmediation systems to the in vivo pathophysiologic effects of endotoxins. Inview of the multiple biologic effects of endotoxins on these systems, thiswill not be an easy task. However, significant advances have been made,and firmly based concepts on the potential role of at least one suchmediation system, ie, serum complement, have been reasonably wellestablished. The second area involves a critical examination of the molec-ular parameters which regulate the interactions between endotoxins andplasma proteins (such as Factor XII and Cl) or mediator cell membranes(eg, platelets, macrophages). Since a prominent role for lipid A has beendemonstrated, the recent advances in the knowledge of lipid A structuremake such research goals highly promising. In addition, the recent re-recognition of the potent biologic activities of the lipid-A-associatedprotein fraction of bacterial endotoxins makes further research on thiscomponent imperative, both with regard to its intrinsic activity as well asits capacity to modulate the expression of lipid-A-initiated endotoxinresponses.

Vol. 93, No. 2 BACTERIAL ENDOTOXINS 603November 1978
Rgf s1. Pfeiffer R: Untersuchungen uber das Cholera gift. Zeitschrift Hyg Infektionskran
11:393-412, 18922. Westphal 0, Westphal U, Sommer T: History of pyrogen research. Microbiology
1977. Edited by D Schiessinger. Washington, D.C., The American Society ofMicrobiology, 1977, pp 221-238
3. Braun V: Molecular organization of the rigid layer and the cell wall of Escherichiacoli. Bacterial iUpopolysaccharides. Edited by EH Kass, SM Wolff. Chicago, TheUniversity of Chicago Press, 1973, pp 1-8
4. Lfuderitz 0, Jann K, Wheat R: Somatic and capsular antigens of gram-negativebacteria. Comprehensive Biochemistry, Section 5: Chemical Biology. Extracellularand Supporting Structures, Vol 26, Part A. New York, Elsevier Publishing Co., 1968,pp 105-228
5. Boivin A, Mesrobeanu L: Recherches sur les antigenes somatiques et sur lesendotoxines des bacteries. I. Considerations generales et expose des techniquesutilisees. Rev Immunol (Paris) 1:553-569, 1935
6. Westphal 0, Lilderitz 0, Bister F: Uber die Extraktion von Bakterien mit Phenol-Wasser. Z Naturforsch 7B:148-155, 1952
7. Morgan WTJ: The isolation and properties of a specific antigenic substance fromB. dysenteriae (Shiga). Biochem J 31:2003-2021, 1937
8. Ribi E, Milner KC, Perrine TD: Endotoxic and antigenic fractions from the cellwall of Salmonella enteitidis: Methods for separation and some biologic activities. jImmunol 82:75-84, 1959
9. Leive L, Shovlin VK, Mergenhagen SE: Physical, chemical, and immunologicalproperties of lipopolysaccharide released from Escherichia coli by ethyl-enediaminetetraacetate. J Biol Chem 243:6384-691, 1968
10. Galanos C, Ltlderitz 0, Westphal 0: A new method for the extraction of Rlipopolysaccharides. Eur J Biochem 9:945-249, 1969
11. Morrison DC, Leive L: Fractions of lipopolysaccharide from Escherichia coli0111:B4 prepared by two extraction procedures. J Biol Chem 250:2911-2919, 1975
12. Windholz M (editor): The Merck Index, Ninth edition. Rahway, N.J., Merck &Co., Inc., 1976, p 469
13. Westphal 0, Lflderitz 0: Chemische Erforschung von Lipopolysacchariden gram-negativer Bakterien. Agnew Chem [Engl] 66:407-417, 1954
14. Jann B, Reske K, Jann K: Heterogeneity of lipopolysaccharides. Analysis of poly-saccharide chain lengths by sodium dodecylsulfate-polyacrylamide gel elec-trophoresis. Eur J Biochem 60:239-246, 1975
15. Gmeiner J: The isolation of two different lipopolysaccharide fractions from variousProteus rmrabilis strains. Eur J Biochem 58:621-626, 1975
16. Edstrom RD, Heath EC: The biosynthesis of cell-wall lipopolysaccharide in Es-cherichia coli. VII. Studies on the structure of the 0-antigenic polysaccharide. JBiol Chem 242:4125-4133, 1967
17. Lfderitz 0, Westphal 0, Staub AM, Nikaido H: Isolation and chemical andimmunological characterization of bacterial lipopolysaccharides. Microbial Toxins:A Comprehensive Treatise. Bacterial Endotoxins, Vol 4. Edited by SJ Ajl, GWeinbaum, S Kadis. New York, Academic Press, Inc., 1971, pp 145-234
18. Galanos C. Lflderitz 0, Rietschel E, Westphal 0: Newer aspects of the chemistrvand biology of bacterial lipopolysaccharides, with special reference to their lipid Acomponent. Biochemistry of Lipids II, Vol 14. (International Review of Biochemis-try.) Edited by TW Goodwin. Baltimore, University Park Press, 1977, pp 239-335
19. Luderitz 0, Westphal 0: The significance of enterobacterial mutants for the

604 MORRISON AND ULEVITCH American Journalof Pathology
chemical investigation of their cell-wall polvsaccharides. Angeu Chemie (InternatiEdit) 5:198-210, 1966
20. Tripodi D, Nowotny A: Relation of structure to function in bacterial 0-antigens.V. Nature of active sites in endotoxic lipopolvsaccharides of Serratia marcescens.Ann NY Acad Sci 133:604-621, 1966
21. Kim YB, Watson DW: Biologically active endotoxins from Salmonella mutantsdeficient in 0- and R-polysaccharides and heptose. J Bacteriol 94:1320-1326, 1967
22. Irvin RT, Chatterjee AK, Sanderson KE, Costerton JW: Comparison of the cellenvelope structure of a lipopolysaccharide-defective (heptose-deficient) strain and asmooth strain of Salmonella typhimunum. J Bacteriol 124:930-941, 1975
23. Koplow J, Goldfine H: Alterations in the outer membrane of the cell envelope ofheptose-deficient mutants of Escherichia coli. J Bacteriol 17:527-543, 1974
24. Burton AJ, Carter HE: Purification and characterization of the lipid A componentof the lipopolvsaccharides from Eschenchia coli. Biochemistry 3:411418, 1964
25. Gmenier J, Luderitz 0, Westphal 0: Biochemical studies on lipopolysaccharidesof Salmonella R mutants. VI. Investigations on the structure of the lipid A com-ponent. Eur J Biochem 7:370-379, 1969
26. Rietschel ET, Hase S, King MT, Redmond J. Lehmann V: Chemical structure oflipid A.2 pp 262-268
27. Hase S, Rietschel ET: Isolation and analysis of the lipid A backbone: Lipid Astructure of lipopolvsaccharides from various bacterial groups. Eur J Biochem63:101-107, 1976
28. Lulderitz 0, Galanos C, Lehmann V, Nurminen NI, Rietschel ET, Rosenfelder GCSimon NI, Westphal 0: Lipid A: Chemical structure and biological activity.' pp9-21
29. Wober N%V, Alaupovi6 P: Studies on the protein moiety of endotoxin from gram-negative bacteria. Characterization of the protein moiety isolated by phenol treat-ment of endotoxin from Serratia marcescens 08 and Escherichia coli 0 141: K85(B).Eur J Biochem 19:340-356, 1971
30. Wober W, Alaupovic P: Studies on the protein moiety of endotoxin from gram-negative bacteria: Characterization of the protein moiety isolated bv acetic acidhydrolysis of endotoxin from Serratia marcescens 08. Eur J Biochem 19:357-367,1971
31. Wu M-C, Heath EC: Isolation and characterization of lipopolvsaccharide proteinfrom Escherichia coli. Proc Natl Acad Sci USA 70:2572-2576, 1973
32. Sultzer BNI, Goodman GW: Endotoxin protein: A B-cell mitogen and polvclonalactivator of C3H!/HeJ lymphoc`tes. J Exp Med 144:821-827, 1976
33. Nlorrison DC, Betz SJ, Jacobs DNI: Isolation of a lipid A-bound polypeptideresponsible for "LPS-initiated" mitogenesis of C3H/HeJ spleen cells. J Exp NIed144:840-846, 1976
34. Tal C, Goebel, WF: On the nature of the toxic component of the somatic antigenof Shigella paradysenteriae type Z (Flexner). J Exp NIed 92:25-34, 1950
35. Ribi E, Anacker RL, Fukushi K, Haskins WT, Landy M, Nlilner KC: Relationshipof chemical composition to biological activity. Bacterial Endotoxins. Edited by NILandy, W Braun. Rahwav, N.J., Quinn & Boden Co., 1964, pp 16-28
36. Westphal 0, Nowotny A, Luderitz 0, Hurni H, Eichenbergen E, Schonholzer G:Die Bedeuting der Lipoid Komponente (Lipoid A) fur die biologischen Wirkun-gen bakterieller Endotoxine (Lipopolysaccharide). Pharm Acta Helv ,33:401-411,1958
37. Kasai N: Chemical studies on the lipid component of endotoxin, with specialemphasis on its relationship to biological activities. Ann NY Acad Sci 133:486-507.1966
38. Westphal 0, Gmeiner J, Luderitz 0, Tanaka A, Eichenberger E: Chemistry and

Vol. 93, No. 2 BACTERIAL ENDOTOXINS 605November 1978
biology of the lipid-A component of enterobacterial lipopolvsaccharides. Colloq IntCNRS Paris 174:69-78, 1969
39. Kasai N, Nowotnv A: (1967) Endotoxic glycolipid from a heptoseless mutant ofSalmonella minnesota. J Bacteriol 94:1824-1836, 1967
40. Neter E, Westphal 0, Liuderitz 0, Gorzynski EA. Eichenberger E: Studies ofenterobacterial lipopolysaccharides. Effects of heat and chemicals on ervthroc%-te-modifying, antigenic, toxic and pvTogenic properties. J Immunol 76:377-7385, 1956
41. Now-otnv A: Endotoxoid preparations. Nature 197:721-722, 196342. Rietschel ET. Gottert H, Liuderitz 0, Westphal 0: Nature and linkages of the
fatty acids present in the lipid-A component of Salnonella lipopolvsaccharides. EurJ Biochem 28:166-173, 1972
4,3. Niwa NM, Milner KC, Ribi E, Rudbach JA: Alteration of physical, chemical, andbiological properties of endotoxin by treatment with mild alkali. J Bacteriol97:1069-1077, 1969
44. Goodman GWN, Sultzer BM: Mild alkaline hydrolvsis of lipopolysaccharide endo-toxin enhances its mitogenicity for murine B cells. Infect Immun 17:205-214. 1977
45. Morrison DC, Jacobs DN: Binding of polyrmnxin B to the lipid A portion of bacteriallipopolysaccharides. Immunochemistry 13:813-818, 1976
46. Neter E, Gorzmnski EA, Westphal 0, Luideritz 0: The effects of antibiotics onenterobacterial lipopolysaccharides (endotoxins), hemagglutination and hemolysis.J Immunol 80:66-72, 1958
47. Rafkind D: Prevention by polymrxin B of endotoxin lethality in mice. J Bacteriol93:1463-1464, 19671
48. Rifkind D: Studies on the interaction between endotoxin and polymyxin B. JInfect Dis 117:433-438, 1967
49. Rifkind D, Hill RB Jr: Neuralization of the Shwartzman reactions by polyrmvxin B.J Immunol 99:564-569, 1967
50. Rifkind D, Palmer JD: Neutralization of endotoxin toxicity in chick embryos byantibiotics. J Bacteriol 92:813-819, 1966
51. Palmer JD, Rifkind D: Neutralization of the hemodynamic effects of endotoxin bvpolymvxin B. Surg Gvnecol Obstet 138:755-759. 1974
52. Jacobs DN. Morrison DC: Dissociation between mitogenicitx and immuno-genicity of TNP-lipopolvsaccharide, a T-independent antigen. J Exp Med141:1453-1458, 1975
53. Jacobs DNM, Mlorrison DC: Inhibition of the mitogenic response to lipo-polvsaccharide (LPS) in mouse spleen cells by polymrnxin B. J Immunol 118:21-27.1977
54. Skidmore BJ, Morrison DC, Chiller JM. Weigle WO: Immunologic properties ofbacterial lipopolvsaccharide (LPS). II. The unresponsiveness of C3H/HeJ mousespleen cells to LPS-induced mitogenesis is dependent on the method used to extractLPS. J Exp Med 142:1488-1508, 1975
55. Ulevitch RJ, Cochrane CG: Role of complement in lethal bacterial lipo-polvsaccharide-induced h-potensive and coagulative changes. Infect Immun19:204-211, 1978
56. Leive L: Release of lipopolvsaccharide by EDTA treatment of E. coli. BiochemBiophys Res Commun 21:290-296, 1965
57. Thomas L: The Lives of a Cell: Notes of a Biology W'atcher. New York, The'Viking Press, 1974, p 78
58. Landy NI (Convener): Symposium on Bacterial Endotoxins. Bacteriol Rev 25:427-458. 1961
59. Landv NI, Braun VV (editors): Bacterial Endotoxins. Rahwav, N.J.. Quinn & BodenCo.. Inc., 1964

606 MORRISON AND ULEVITCH American Journalof Pathology
60. Nowotny A (consulting editor): Molecular biology of gram-negative bacterial lipo-polvsaccharides. Ann NY Acad Sci 133:277-786, 1964
61. Luiaeritz 0, Staub AM, Westphal 0: Immunochemistrv of 0 and R antigens ofSalmonella and related enterobactenaceae. Bacterial Rev 30:192-255, 1966
62. Nowotny A: Molecular aspects of endotoxic reactions. Bacterial Rev 33:72-98,1969
63. Weinbaum G, Kadis S, Ajl SJ (editors): Microbial Toxins: A ComprehensiveTreatise. Bacterial Endotoxins, Vol 4. New York, Academic Press, Inc., 1971
64. Kadis S, Weinbaum G, Ajl SJ (editors): Microbial Toxins: A ComprehensiveTreatise. Bacterial Endotoxins, Vol 5. New York, Academic Press, Inc., 1971
65. Kass EH, Wolff SM (editors): Bacterial Upopolvsacchrides. Chicago, Universitvof Chicago Press, 1973
66. Westphal 0: Bacterial endotoxins. Int Arch Allerg Appl Immunol 49:1-43, 197567. Schlessinger D (editor): Microbiology 1977. Washington, D.C., The American
Society of Microbiology, 1977, pp 217-43068. Morrison DC, Ryan JF: Modulation of host defense mediator systems by bacterial
endotoxins. Adv Immunol (In preparation)69. Muller-Eberhard JH: The serum complement system. Textbook of Immuno-
pathology, Second edition, Vol 1. Edited by PA Miescher, HJ Muller-Eberhard.New York, Grune & Stratton, 1976, pp 45-73
70. Mergenhagen SE, Snydermnan R, Gewurz H, Shin HS: Significance of com-plement to the mechanism of action of endotoxin. Curr Top Microbiol Immunol50:37-77, 1969
71. Landy M, Pillemer L: Increased resistance to infection and accompanying altera-tion in properdin levels following administration of bacterial lipopolysaccharides. JExp Med 104:33409, 1956
72. Landy M, Pillemer L: Elevation of properdin levels in mice following administra-tion of bacterial lipopolysaccharides. J Exp Med 103:823-833, 1956
73. Hook WA, Carey WF, Muschel LH: Alterations in serum Ivsozyme and properdintiters of mice following x-irradiation or treatment with zymosan or endotoxin. JImmunol 84:569-575, 1960
74. Spink WW, Vick J: A labile serum factor in experimental endotoxin shock: Cross-transfusion studies in dogs. J Exp Med 114:501-508, 1961
75. Spink WW, Davis RB, Potter RK Chartrand S: The initial stage of canine endo-toxin shock as an expression of anaphylactic shock: Studies on complement titersand plasma histamine concentrations. J Clin Invest 43:696-704, 1964
76. Gilbert VE, Braude AI: Reduction of serum complement in rabbits after injectionof endotoxin. J Exp Med 116:477-490, 1962
77. Pearlman DS, Sauers JB, Talmage DW: The effect of adjuvant amounts of endo-toxins on the serum hemolytic complement activity in rabbits. J Immunol 91:748-756, 1963
78. Miler J, Tlaskalova H, Kostka J, Jilek M: Effect of endotoxin on the complementlevel in sera of precolostral newborn pigs. Folia Microbiol (Praha) 11:475-478, 1966
79. Kane MA, Mav JE, Frank MM: Interactions of the classical and alternate com-plement pathway with endotoxin lipopolysaccharide: Effect on platelets and bloodcoagulation. J Clin Invest 52:370-376, 1973
80. Cochrane CG, Mflller-Eberhard HJ, Aiken BS: Depletion of plasma complementin vio by a protein of cobra venom: Its effect on various immunologic reactions.J Immunol 105:55-69, 1970
81. Cooper NR: Formation and function of a complex of the C3 proactivator with aprotein from cobra venom. J Exp Med 137:451-460, 1973
82. Polak L, Turk JL: Suppression of the haemorrhagic component of the Shwartz-man reaction by anti-omplement serum. Nature 223:738-739, 1969

Vol. 93, No. 2 BACTERIAL ENDOTOXINS 607November 1978
83. From AHL, Gewurz H, Gruninger RP, Pickering RJ, Spink WW: Complement inendotoxin shock: Effect of complement depletion on the early hypotensive phase.Infect Immunol 2:38-41, 1970
84. Kitzmiller JL, Lucas WE, Yelenosky PF: The role of complement in feline endo-toxin shock. Am J Obstet Gvnecol 112:414-421, 1972
85. Garner R, Chater BV, Brown DL: The role of complement in endotoxin shock anddisseminated intravascular coagulation: Experimental observations in the dog. Br JHaematol 28:393-401, 1974
86. Fong JSC, Good RA: Prevention of the localized and generalized Shwartzmanreactions by an anticomplementary agent, cobra venom factor. J Exp NMed 134:642-655, 1971
87. Bergstein JM, Mlichael AF Jr: Failure of cobra venom factor to prevent thegeneralized Shwartzrnan reaction and loss of renal cortical fibrinolytic activity. Am JPathol 74:19-30, 1974
88. Muller-Berghaus G, Lohmann E: The role of complement in endotoxin-induceddisseminated intravascular coagulation: Studies in congenitally Cs-deficient rabbits.Br J Haematol 28:403-418, 1974
89. Ulevitch RJ, Cochrane CG, Henson PM, Morrison DC, Doe WF: Mediationsvstems in bacterial lipopolvsaccharide-induced hypotension and disseminated in-travascular coagulation. I. The role of complement. J Exp NMed 142:1570-1590,1975
90. Brown DL, Lachmann PJ: The behavior of complement and platelets in lethalendotoxin shock in rabbits. Int Arch Allergy Appl Immunol 45:193-205, 1973
91. Johnson KJ, Ward PA: Protective function of C6 in rabbits treated with bacterialendotoxin. J Immunol 106:1125-1127, 1971
92. NMcCabe WR: Serum complement levels in bacteremia due to gram negativeorganisms. N Engl J Med 288:21-23, 1973
93. Fearon DT, Ruddy S, Schur PH, NMcCabe WR: Activation of the properdin path-way of complement in patients with gram-negative bacteremia. N Engl J Med292:937-940, 1975
94. Robin MI, Intrator L, Andre C. Lagrue G, Rapin MI: Complement abnormalitiesand infection-related shock. Nouv Presse Med 3:883-884. 1974
95. Pillemer L, Schoenberg MD, Blum L, Wurz L: Properdin system and immunity.II. Interaction of the properdin system with polysaccharides. Science 122:545-549.1955
96. Kostka J, Sterzl J: The action of endotoxin on complement. Folia Microbiol(Praha) 7:191-192, 1962
97. Muschel LH, Schmoker K, Wtebb PM: Anticomplementary action of endotoxin.Proc Soc Exp Biol Med 117:639-643, 1964
98. Gewurz H, Mergenhagen SE, Nowotny- A, Phillips EJK: Interactions of the com-plement system with native and chemically modified endotoxins. J Bacteriol95:397-405, 1968
99. Gewurz H, Shin HS, Mergenhagen SE: Interactions of the complement systemwith endotoxic lipopolvsaccharide: Consumption of each of the six terminal com-plement components. J Exp Med 128:1049-1057, 1968
100. Hook WA, Snvderman R, NMergenhagen SE: Consumption of hamster com-plement by bacterial endotoxin. J Immunol 105:268-270, 1970
101. Ge\urz H, Pickering RJ, Snvderman R, Lichtenstein LM. Good RA, NlengenhagenSE: Interactions of the complement system with endotoxic lipopol saccharides inimmunoglobulin-deficient sera. J Exp Med 131:817-831, 1970
102. NMarcus RL, Shin HS, NMaver M\M: An alternative complement pathway: C3cleaving activity not due to C4,2a, on endotoxic lipopolysaccharide after treatment

608 MORRISON AND ULEVITCH American Journalof Pathology
with guinea pig serum: Relation to properdin. Proc Natl Acad Sci USA 68:1351-1354. 1971
103. Frank MM, May J, Gaither T. Ellman L: In vitro studies of complement functionin sera of C4-deficient guinea pigs. J Exp Med 134:176-187, 1971
104. Gotze 0, MillIer-Eberhard HJ: The C3-activator system: An alternative path-way of complement activation. J Exp Med 134(Suppl):90s-108s, 1971
105. NMergenhagen SE, Gewurz H, Bladen HA, Nowotny A, Kasai N. Lulderitz 0:Interactions of the complement system with endotoxins from a Salmonella min-nesota mutant deficient in 0-polysaccharide and heptose. J Immunol 100:227-229, 1968
106. Galanos C, Rietschel ET. Lulderitz 0. Westphal 0: Interaction of lipo-polvsaccharides and lipid A with complement. Eur J Biochem 19:143-152. 1971
107. NMorrison DC, V'erroust P: Anticomplementarv activity of lipid A isolated fromlipopolysaccharides. Proc Soc Exp Biol Med 143:1025-1030, 197-3
108. Dierich NIP, Bitter-Suermann D, Konig W, Hadding U, Galanos C, RietschelET: Analysis of bypass activation of C3 by endotoxic LPS and loss of this potency.Immunology 24:721-733, 1973
109. NIiller-Eberhard HJ, Bokisch VA, Budzko DB: Studies of human anaphylatoxinsand of their physiological control mechanism. Sixth International Symposium onImmunopathology: Immunopathology: Antigen Cell Interaction and the Control ofAntibody Synthesis. Edited by PA NMiescher. Basel, Schwabe & Co.. 1971, pp 191-200
110. Loos NI, Bitter-Suermann D, Dierich N: Interaction of the first (CI), the second(C2) and the fourth (C4) component of complement with different preparations ofbacterial lipopolvsaccharides and with lipid A. J Immunol 112:935-940, 1974
111. Snvderman R, Pike MC: Interaction of complex polysaccharides with the com-plement system: Effect of calcium depletion on terminal component consumption.Infect Immun 11:273-279, 1975
112. Lachmann PJ, Nicol PAE: Studies on the C3b feedback cycle. Advances in theBiosciences, Vol 12. Edited by G Raspe, S Bernhard. Pergammon Press, Vieweg.Braunschweig, Germany, 1974, pp 262-269
113. Nlorrison DC, Kline LF: Activation of the classical and properdin pathways ofcomplement by bacterial lipopol%saccharides (LPS). J Immunol 118:362-368, 197
114. Nlorrison DC, Jacobs DM: Inhibition of lipopolysaccharide-initiated activation ofserum complement by polvmyxin B. Infect Immun 13:298-01, 1976
115. Cooper NR, Nlorrison DC: Binding and activation of the first component ofhuman complement by the lipid A region of lipopolysaccharide. J Immunol 120:1862-1868, 1978
116. Galanos C, Lulderitz 0: The role of the physical state of lipopolysaccharides in theinteraction with complement. High molecular weight as prerequisite for the ex-pression of anticomplementarv activity. Eur J Biochem 65:403-408. 1976
117. Sanarelli G: De la pathogenie du chol&ra (neuvieme memoire). Le choleraexperimental. Ann Inst Pasteur Lille 38:11-72, 1924
118. Shwartzman G: Studies on Bacillus typhosus toxic substances. I. Phenomenon oflocal skin reactivity to B. typhosus culture filtrate. J Exp NMed 48:247-268, 1928
119. Shwartzman G: A new phenomenon of local skin reactivity to B. typhosus culturefiltrate. Proc Soc Exp Biol NMed 25:560-561, 1928
120. Beller FK: The role of endotoxin in disseminated intravascular coagulation.Thromb Diath Haemorrh 36:(Suppl):125-149, 1969
121. NMcKav DG: Vessel wall and thrombogenesis-endotoxin. Thromb Diath Hae-morrh 29:11-26. 1973
122. Griffin JH, Cochrane CG: NMechanisms for the involvement of high molecularweight kininogen in surface-dependent reactions of Hageman factor. Proc NatlAcad Sci USA 73:2554-2558, 1976

Vol. 93, No. 2 BACTERIAL ENDOTOXINS 609November 1978
123. Colman RW, Wong PY: Participation of Hageman factor dependent pathways inhuman disease states. Thromb Haemostas 38:751-775, 1977
124. Revak SD, Cochrane CG, Griffin JH: Initiation of coagulation by contact activa-tion of Hageman factor. Kidney Disease: Hematologic and Vascular Problems.Edited bv RM McIntosh, SJ Guggenheim, RW Schrier. New York, John Wiley &Sons, Inc., 1977, pp 29-40
125. Kisiel W, Fujikawa K, Davie EW: Activation of bovine factor VII (proconvertin).by factor XIIa (activated Hageman factor). Biochemistry 16:4189-4194, 1977
126. Radcliffe R, Bagdasarian A, Colman R, Nemerson Y: Activation of bovine factorVII by Hageman factor fragments. Blood 50:611-617, 1977
127. Kimball HR, Melmon KL, Wolff SM: Endotoxin-induced kinin production inman. Proc Soc Exp Biol Med 139:1078-1082, 1972
128. Nies AS, Forsyth RP, Williams HE, Melmon KL: Contribution of kinins toendotoxin shock in unanesthetized rhesus monkevs. Circ Res 22:155-164. 1968
129. Nies AS, Melmon KL: Variation in endotoxin-induced kinin production and effectbetween the rabbit and the rhesus monkey. Am J Phvsiol 225:230-233, 1973
130. Diniz CR, Reis ML, Conrado AP: Bradvkinin and related kinins. Edited byNI Rocha, E Silva, HA Rothschild. New York, Plenum Publishing Corp., 1967, p 15
131. Erdos EG, Miwa I: Effect of endotoxin shock on the plasma kallikrein-kininsystem of the rabbit. Fed Proc 27:92-95, 1968
132. Reichgott MJ, Melmon KL, Forsvth RP, Greineder D: Cardiovascular and meta-bolic effects of whole or fractionated gram-negative bacterial endotoxin in theunanesthetized rheusus monkey. Circ Res 33:346--352, 1973
133. Skjlrten F, Evensen SA: Induction of disseminated intravascular coagulation inthe factor XII-deficient fowl: Morphological effects of liquoid, bacterial endotoxinand tissue thromboplastin in the normal anticoagulated fowl. Thromb DiathHaemorrh 30:25-35, 1973
134. Soulier J-P, Wartelle 0, Menache D: Hageman trait and PTA deficiency: The roleof contact of blood with glass. Br J Haematol 5:121-138, 1959
1.35. Mason JW, Colman RW: The role of Hageman factor in disseminated intra-vascular coagulation induced by septicemia, neoplasia or liver disease. ThrombDiath Haemorrh 26:325-331, 1971
136. Robinson JA, Klodnycky ML, Loeb HS, Racic MR, Gunnar RNM: Endotoxin.prekallikrein, complement and systemic vascular resistance: Sequential measure-ments in man. Am J Med 59:61-67, 1975
137. Shen SM-C, Rapaport SI, Feinstein DI: Intravascular clotting after endotoxin inrabbits with impaired intrinsic clotting produced by a factor VIII antibody. Blood42:523-534, 1973
1.38. Rodriguez-Erdmann F: Studies on the pathogenesis of the generalized Shwartz-man reaction. III. Trigger mechanism for the activation of the prothrombin mole-cule. Thromb Diath Haemorrh 12:471-483, 1964
139. Morrison DC, Cochrane CG: Direct evidence for Hageman factor (factor XII)activation by bacterial lipopolvsaccharides (endotoxins). J Exp Med 140:797-811,1974
140. Becker RNI: Suppression of local tissue reactivitv (Shwartzrnan Phenomenon) bynitrogen mustard, benzol, and x-ray irradiation. Proc Soc Exp Biol Med 69:247-250.1948
141. Stetson CA, Good RA: Studies on the mechanism of the Shwartzman phenome-non. Evidence for the participation of polymorphonuclear leucocytes in the phe-nomenon. J Exp Med 93:49-64, 1951
142. Thomas L, Good RA: Studies on the generalized Shwartzman reaction. I. Generalobservations concerning the phenomenon. J Exp Med 96:605-624, 1952
143. Horn RG: Evidence for participation of granulocytes in the pathogenesis or thegeneralized Shwartzman reaction: A review.' pp 125-135

610 MORRISON AND ULEVITCH Amrican Jouralof Pahogy
144. Bohn E, Mulller-Berghaus G: The effect of leucocyte and platelet transfusion onthe activation of intravascular coagulation by endotoxin in granulocytopenic andthrombocytopenic rabbits. Am J Pathol 84:239-258, 1976
145. Gamer R, Evensen SA: Endotoxin-induced intravascular coagulation and shock indogs: The role of factor VII. Br J Haematol 27:655-668, 1974
146. Niemetz J, Fani K: Role of leucocytes in blood coagulation and the generalizedShwartzrnan reaction. Nature [New Biol] 232:247-248, 1971
147. Lemer RG, Goldstein R, Cummings G: Stimulation of human leucocyte thrombo-plastic activity by endotoxin. Proc Soc Exp Biol Med 138:145-148, 1971
148. Garg SK, Niemetz J: Tissue factor activity of normal and leukemic cells. Blood42:729-735, 1973
149. Niemetz J, Fani K: Thrombogenic activity of leucocytes. Blood 42:47-59, 1973150. Rivers RPA, Hathaway WE, Weston WL: The endotoxin-induced coagulant ac-
tivity of human monocytes. Br J Haematol 30:311-316, 1975151. Hiller E, Saal JG, Ostendorf P, Griffiths GW: The procoagulant activity of human
granulocytes, lymphocytes and monocytes stimulated by endotoxin. Klin Woch-enschr 55:751-757, 1977
152. Rickles FR, Levin J, Hardin JA, Barr CF, Conrad ME Jr: Tissue factor generationby human mononuclear cells: Effects of endotoxin and dissociation of tissue factorgeneration from mitogeric response. J Lab Clin Med 89:792-803, 1977
153. Rickles FR, Rick PD: Structural features of Salmonella typhimuium lipo-polysaccharide required for activation of tissue factor in human mononuclear cells.J Clin Invest 59:1188-1195, 1977
154. Rickles FR, Bobrove AM: Tissue factor in mononuclear cell cultures. Blood46:1046, 1975 (Abstr)
155. Niemetz J, Morrison DC: Lipid A as the biologically active moiety in bacterialendotoxin (LPS)initiated generation of procoagulant activity by peripheral bloodleucocytes. Blood 49:947-956, 1977
156. Rudback JA, Anacker RL, Haskins WT, Milner KC, Ribi E: Physical structure ofnative protoplasmic polysaccharide from Escherichia coli. J Immunol 98:1-7, 1967
157. Stetson CA Jr: Studies on the mechanism of the Shwartzman phenomenon. Cer-tain factors involved in the production of the local hemorrhagic necrosis. J Exp Med93:489-504, 1951
158. Weil MH, Spink WW: A comparison of shock due to endotoxin with anaphylacticshock. J Lab Clin Med 50:501-515, 1957
159. McKay DG, Shapiro SS: Alterations in the blood coagulation system induced bybacterial endotoxin. I. In vivo (generalized Shwartman reaction). J Exp Med107:353-367, 1958
160. Cohen P, Braunwald J, Gardner FH: Destruction of canine and rabbit plateletsfollowing intravenous administration of carbon particles or endotoxin. J Lab ClinMed 66:263-271, 1965
161. Davis RB: Electron microscopic changes in blood platelets induced by bacteriallipopolysaccharide. Exp Mol Pathol 5:559-574, 1966
162. Spielvogel AR: An ultrastructural study of the mechanisms of platelet-endotoxininteraction. J Exp Med 126:235-250, 1967
163. Shimamoto T, Yamazaki H, Sagawa N, Iwahara S, Konishi T, Maezawa H: Effectof bacterial endotoxins on platelets and release of serotonin (5-hydroxy tryptamine).Proc Jpn Acad 34:450-455, 1958
164. Davis RB, Meeker WR Jr, Bailey WL: Serotonin release by bacterial endotoxin.Proc Soc Exp Biol Med 108:774-776, 1961
165. Davis RB, Meeker WR McQuarrie DG: Immediate effects of intravenous endo-toxin on serotonin concentrations and blood platelets. Circ Res 8:234-239, 1960
166. Horowitz HI, Des Prez RM, Hook EW: Effects of bacterial endotoxin on rabbit

Vol. 93, No. 2 BACTERIAL ENDOTOXINS 611November 1978
platelets. II. Enhancement of platelet factor 3 activity in vivo and in vitro. J ExpMed 116:619-633, 1962
167. Rodriguez-Erdmann F: Intravascular activation of the clotting system with phos-pholipids: Production of the generalized Shwartzman reaction with platelet factor3. Blood 26:541-553, 1965
168. Good RA, Thomas L: Studies on the generalized Shwartzman reaction. II. Theproduction of bilateral cortical necrosis of the kidneys by a single injection ofbacterial toxin in rabbits previously treated with thorotrast or trypan blue. J ExpMed 96:625-641, 1952
169. Muller-Berghaus G, Goldfinger D, Margaretten W, McKay DG: Platelet factor 3and the generalized Shwartzman reaction. Thromb Diath Haemorrh 18:726-735,1967
170. Evans G, Lewis AF, Mustard JF: The role of platelet aggregation in the develop-ment of endotoxin shock. Br J Surg 56:624, 1969 (Abstr)
171. Levin J, Cluff LE: Platelets and the Shwartzman phenomenon. J Exp NMed121:235-246, 1965
172. Levin J, Beck E: A re-appraisal of the role of blood coagulation and platelets in thegeneralized Shwartzxnan phenomenon. Thomb Diath Haemorrh 15:519-38, 1966
173. Margaretten W, McKay DG: The role of the platelet in the generalized Shwartz-man reaction. J Exp Med 129:585-590, 1969
174. Muller-Berghaus G, Kramer W: Effect of platelet antiserum on the precipitationof soluble fibrin by endotoxin. Thromb Haemostas 35:237-248, 1976
175. Muller-Berghaus G, Bohn E, Hobel W: Activation of intravascular coagulation byendotoxin: The significance of granulocytes and platelets. Br J Haematol 33:213-220, 1976
176. Brown DL, Lachmann PJ: The relationship of complement-mediated plateletdamage to intravascular coagulation: Comparison between endotoxin and insulin inthe rabbit.lu pp
177. Wolff SM, Rubenstein M, Mulholland JH, Alling DW: Comparison of hemato-logic and febrile response to endotoxin in man. Blood 26:190-201, 1965
178. Hinshaw LB, Jordan MM, Vick JA: Histamine release and endotoxin shock in theprimate. J Clin Invest 40:1631-1637, 1961
179. Sheagren JN, Wolff SM, Shulman NR: Febrile and hematologic responses ofrhesus monkeys to bacterial endotoxin. Am J Physiol 212:884-890, 1967
180. Dinbar A, Rapaport SI, Patch MJ, Grant W, Fonkalsrud EW: Hematologic effectsof endotoxin on the macaque monkey. Surgery 70:596-63, 1967
181. Ulevitch RJ, Cochrane CG, Bangs K, Herman CM, Fletcher JR, Rice CL: Theeffect of complement depletion on bacterial lipopolysaccharide (LPS-induced he-modynamic and hematologic changes in the rhesus monkey. Am J Pathol 92:227-240, 1978
182. McKay DG, Margaretten W, Csavossy I: An electron microscope study of endo-toxin shock in rhesus monkeys. Surg Gvnecol Obstet 125:825-832, 1967
183. Balis JU, Gerber LI, Rappaport ES, Neville WE: Mechanisms of blood-vascularreactions of the primate lung to acute endotoxemia. Exp Mol Pathol 21:123-137,1974
184. Davis RB, McQuarrie DG, Meeker WR: Immediate effects of Escherichia coliendotoxin administration on platelets and serotonin concentration. Fed Proc18:211, 1959
185. Davis RB, Meeker WR Jr, McQuarrie DG: Early changes in intravascular pres-sures, platelets, and serotonin after E. coli endotoxin administration. Surg Forum10:401-405, 1959
186. Siqueira M, Nelson RA Jr: Platelet agglutination by immune complexes and itspossible role in hypersensitivity. J Immunol 86:516-525, 1961

612 MORRISON AND ULEVITCH Arnerican Joumalof Pathoogy
187. Des Prez RM, Horowitz HI, Hook EW: Effects of bacterial endotoxin on rabbitplatelets. I. Platelet aggregation and release of platelet factors in vitro. J Exp Med114:857-874, 1961
188. Horowitz HI, Des Prez RM, Hook EW: Activation of platelet factor 3 by bacterialendotoxin and by immune reactions. Blood 20:760, 1962 (Abstr)
189. Des Prez RM: Effects of bacterial endotoxin on rabbit platelets. III. Comparisonof platelet injury induced by thrombin and by endotoxin. J Exp Med 120:305-313,1964
190. Des Prez RM, Bryant RE: Effects of bacterial endotoxin on rabbit platelets. IV.The divalent ion requirements of endotoxin-induced and immunologically inducedplatelet injury. J Exp Med 124:971-982, 1966
191. Des Prez RM: The effects of bacterial endotoxin on rabbit platelets. V. Heat labileplasma factor requirements of endotoxin-induced platelet injury. J Immunol99:966-973, 1967
192. Nagayama M, Zucker MB, Beller FK: Effects of a variety of endotoxins on humanand rabbit platelet function. Thromb Diath Haemorrh 26:467-473, 1971
193. Siraganian RP: Platelet requirement in the interaction of the complement andclotting systems. Nature [New Biol] 239:208-210, 1972
194. Maclntyre DE, Allen AP, Thorne KJI, Glauert AM, Gordon JL: Endotoxin-induced platelet aggregation and secretion. I. Morphological changes and pharma-cological effects. J Cell Sci 28:211-223, 1977
195. Thorne KJI, Oliver RC, Maclntyre DE, Gordon JL: Endotoxin-induced plateletaggregation and secretion. II. Changes in plasma membrane proteins. J Cell Sci28:225-236, 1977
196. Morrison DC, Henson PM, Kline LF: Lipopolysaccharide (LPS) initiate lipid-Aindependent (alternative) and lipid A-dependent (classical) activation of com-plement in plasma mediated platelet lysis. Fed Proc 35:655, 1976 (Abstr)
197. Morrison DC, Kline LF, Oades ZG, Henson PM: Mechanisms of lipo-polysaccharide-initiated rabbit platelet responses. I. Altemative complement path-way dependence of the lytic response. Infect Immun 20:744-751, 1978
198. Morrison DC, Oades ZG: Mechanisms of lipopolysaccharide-initiated rabbitplatelet responses. II. Evidence that lipid A is responsible for binding of lipo-polysaccharide to the platelet. J Immunol (In press)
199. Oades ZG, Morrison DC: Lipopolysaccharide-induced calcium transport initiatesselective release of serotonin from rabbit platelets. Am Soc Microbiol (Abstr) 1978
200. Ream VJ, Deykin D, Gurewich V, Wessler S: The aggregation of human plateletsby bacterial endotoxin. J Lab Clin Med 66:245-252, 1965
201. M&ller-Eckhardt CH, Luscher EF: Immune reactions on human blood platelets.IV. Investigations of the problem of an immunologically induced effect of endo-toxin on human platelets. Thromb Diath Haemorrh 20:336-344, 1968
202. Pfueller SL, Lfscher EF: Studies of the mechanisms of the human platelet releasereaction induced by immunologic stimuli. II. The effects of zymosan. J Immunol112:1211-1218, 1974
203. Vermylen J, Fumarola D, Semeraro N: The effect of endotoxins on generation ofthrombin in the platelet atmosphere. Thromb Diath Haemorrh 34:315, 1975 (Abstr)
204. Hawiger J, Hawiger A, Ttmmons S: Endotoxin-sensitive membrane component ofhuman platelets. Nature 256:125-127, 1975
205. Hawiger A, Hawiger J, Timmons S, Steckley S, Cheng C: The role of membranereceptors in pathways of human platelet activation by endotoxin and other micro-bial products. Thromb Diath Haemorrh 34:594, 1975 (Abstr)
206. Hawiger J, Hawiger A, Steckley S, Timmons S, Cheng C: Membrane changes inhuman platelets induced by lipopolysaccharide endotoxin. Br J Haematas 35:285-299, 1977

Vol. 93, No. 2 BACTERIAL ENDOTOXINS 613November 1978
207. Springer GF, Adve JC: Endotoxin-binding substances from human leukocytes andplatelets. Infect Immun 12:978-986, 1975
208. Springer GF, Adye JC: Endotoxin interaction with phosphatides from humanleukocytes and platelets and with lvmphoid cells of susceptible and resistant mice.2pp 326-329
209. Adye JC, Springer GF: Binding of endotoxin (LPS) by glycerophosphatides fromhuman platelets and leukocytes. Naturw-issenschaften 64:130-156, 1977
210. Ginsberg MH, Henson PM: Enhancement of platelet response to immune com-plexes and IgG aggregates by lipid A-rich bacterial lipopolvsaccharides. J Exp Med147:207-218, 1978
211. Ginsberg MH, M1orrison DC: The selective binding of aggregated IgG to lipid A-rich bacterial lipopolysaccharides. J Immunol 120:317-319, 1978
212. Bennett IL Jr, Beeson PB: The properties and biologic effects of bacterial pyro-gens. Medicine 29:363-400, 1950
213. Delaunav A: Mise en evidence d'une nouvelle propriete des antigenes glucidolipi-diques: Leur pouvoir leucopenisant. C R Soc Biol [Paris] 137:389-590. 194:3
214. Athens JW, Haab OP, Raab SO, Mauer AM, Ashenbrucker H, Cartwright GE.Wintrobe NMM: Leukokinetic studies. I'. The total blood, circulating and mar-ginal granulocvte pools and the granulocyte turnover rate in normal subjects. J ClinInvest 40:989-995, 1961
213. Mechanic RC, Frei E III, Landy MI, Smith WWN%: Quantative studies of humanleucocytic and febrile response to single and repeated doses of purified bacterialendotoxin. J Clin Invest 41:162-172, 1961
216. Fehr J, Jacob HS: In vitro granulocyte adherence and in vivo margination: Tw-oassociated complement-dependent functions. J Exp NMed 146:641-652. 1977
217. MacGregor RR: Granulocyte adherence changes induced by hemodialysis. endo-toxin, epinephrine, and glucocorticoids. Ann Intern Med 86:35-39, 19.7
218. Corrigan JJ Jr, Bell BM: Comparison between the polvmvxins and gentamicin inpreventing endotoxin-induced intravascular coagulation and leucopenia. InfectImmun 4:563-566, 1971
219. Corrigan JJ Jr, Sieber OF Jr, Ratajczak H, Bennett BB: Modification of humanneutrophil response to endotoxin with polymvxin B sulfate. J Infect Dis 130:384-387, 1974
220. Wood WBJr: Studies on the cause of fever. N Engl J Med 258:1023-1031 1938221. Greisman SE, Hornick RB: Nlechanisms of endotoxin tolerance with special refer-
ence to man.' pp 25 -268222. Dinarello CA, Wolff SM: Pathogenesis of fever in man. N Engl J Med 298:607-
612, 1978223. Nliller RL, Reichgott NMJ, Mlelmon KL: Biochemical mechanisms of generation of
bradvkinin by endotoxin.s pp 136-148224. Coalson JJ, Hinshaw LB, Guenter CA: The pulmonary untrastructure in septic
shock. Exp 0ol Pathol 12:84-103, 1970225. Pingelton WW, Coalson JJ, Guenter CA: Significance of leucocvtes in endotoxic
shock. Exp Biol Pathol 22:183-195, 1975226. Herring WB, Herion JC, Walker RI, Palmer JG: Distribution and clearance of
circulating endotoxin. J Clin Invest 42:79-87, 1963227. Brunning RD, Woolfrev BF, Schrader WNH: Studies with tritiated endotoxin. I1.
Endotoxin localization in the formed elements of the blood. Am J Pathol 44:401-409, 1964
228. Schrader WH, Woolfrey BF, Brunning RD: Studies with tritiated endotoxin. III.The local Schwartzman reaction. Am J Pathol 44:597-611, 1964
229. Fritze E, Wendt F: Zum Wirkungsmechanismus bakterieller Lipopolysaccharide.

614 MORRISON AND ULEVITCH American Journalof Patoogy
II. Die Phagocytoseaktivitat menschlicher Blutzgranulocyten unter den EinflussbakterieUer Lipopolysaccharide in vitro. Klin Wochenschr 33:575-576, 1955
230. Collins RD, Wood WB Jr: Studies on the pathogenesis of fever. VI. The inter-action of leukocytes and endotoxin in vitro. J Exp Med 110:1005-1016, 1959
231. Cohn ZA, Morse SI: Functional and metabolic properties of polymorphonuclearleukocytes. II. The influence of a lipopolysaccharide endotoxin. J Exp Med111:689-704, 1960
232. Hohnadel J, Stjernholm R: The influence of endotoxin on the glucose metabolismof normal human polymorphonuclear leukocytes. Fed Proc 25:760, 1966 (Abstr)
233. Graham RC Jr, Karnovsky MJ, Shafer AW, Glass EA, Karnovsky ML: Metabolicand morphological observations on the effect of surface-active agents on leukocvtes.J Cell Biol 32:629-647, 1967
234. Tenney SR, Rafter GW: Leukocyte adenosine triphosphatases and the effect ofendotoxin on their activity. Arch Biochem Biophys 126:53-8, 1968
235. Bannatyne RM, Harnett NM, Lee KY, Biggar WD: Inhibition of the biologiceffects of endotoxin on neutrophils by polymyxin B sulfate. J Infect Dis 136:469-474, 1977
236. Goodale F, Fillmore RK Hiulman E: Electron microscopic observations of humanleukocytes. I. Response in vitro to bacterial endotoxin. Exp Mol Pathol 1:229-250,1962
237. Park BH, Good RA: NBT test stimulated. Lancet 2:616, 1970238. Matula G, Paterson PY: Spontaneous in vitro reduction of nitroblue tetrazolium
by neutrophils of adult patients with bacterial infection. N Engl J Med 285:311-317, 1971
239. Goihman-Yahr M, Rodriguez-Ochoa G, Aranzazu N, Convit J: Polvmorpho-nuclear activation in leprosy. I. Spontaneous and endotoxin-stimulated reductionof nitroblue tetrazolium: Effects of serum and plasma on endotoxin-inducedactivation. Clin Exp Immunol 20:257-264, 1975
240. Gimber PE, Rafter GW: The interaction of Escherichia coli endotoxin with leuko-cytes. Arch Biochem Biophys 135:14-20 1969
241. Snyderman RK Gewurz H, Mergenhagen SE: Interactions of the complementsystem with endotoxic lipopolysaccharide. Generation of a factor chemotactic forpolymorphonuclear leukocytes. J Exp Med 128:259-275, 1968
242. Synderman R, Shin HS, Phillips JK, Gewurz H, Mergenhagen SE: A neutrophilchemotactic factor derived from C'5 upon interaction of guinea pig serum withendotoxin. J Immunol 103:413-422, 1969
243. Sveen K: Rabbit polymorphonuclear leukocyte migration in vitro in response tolipopolysaccharides from bacteroides, fusobacterium and veillonella. Acta PatholMicrobiol Scand [B] 85:374-380, 1977
244. Geotzl EJ, Austen KF: A neutrophil-immobiizing factor derived from humanleukocytes. I. Generation and partial characterization. J Exp Med 136:1564-1580,1972
245. Issekutz AC, Biggar WD: Influence of serum-derived chemotactic factors andbacterial products on human neutrophil chemotaxis. Infect Immun 15:212-220,1977
246. Allison AC, Davies P, Page RC: Effects of endotoxin on macrophages and otherlymphoreticular cells. J Infect Dis 128:S212-S219, 1973
247. Unanue ER: Secretory function of mononuclear phagocytes: A review. Am JPathol 83:396-417, 1976
248. Cohn ZA, Benson B: The differentiation of mononuclear phagocytes: Morphol-ogy, cytochemistry, and biochemistry. J Exp Med 121:153-170, 1965
249. Bennett WE, Cohn ZA: The isolation and selected properties of blood monocytes.J Exp Med 123:145-160, 1966

Vol. 93, No. 2 BACTERIAL ENDOTOXINS 615November 1978
250. Wieher E, Levanon D: The in vitro interaction between bacterial lipo-polysaccharide and differentiating monocytes. Lab Invest 19:584-590, 1968
251. Doe W, Yang D, Henson P: Bacterial lipopolysaccharide (LPS-induced mac-rophage (MO) stimulation: Selective lysosomal enzyme release. Fed Proc 36:1263,1977 (Abstr)
252. Bianco C, Edelson PJ: Macrophage activation in C3H/HeJ mice. Fed Proc36:1263, 1977 (Abstr)
253. Watson J, Riblet R: Genetic control of responses to bacterial lipopolysaccharidesin mice. I. Evidence for a single gene that influences mitogenic and immunogenicresponses to lipopolysaccharides. J Exp Med 140:1147-1161, 1974
254. Wahl LM, Wahl SM, Mergenhagen SE, Martin GR: Collagenase production byendotoxin-activated macrophages. Proc Natl Acad Sci USA 71:3598-3601, 1974
255. Wahl LM, Olsen CE, Sandberg AL, Mergenhagen SE: Prostaglandin regulationof macrophage collagenase production. Proc Natl Acad Sci USA 74:4955-4958,1977
256. Gordon S, Unkeless JC, Cohn ZA: Induction of macrophage plasminogen activa-tor by endotoxin stimulation and phagocvtosis: Evidence for a two-stage process. JExp Med 140:995-1010, 1974
257. Alexander P, Evans R: Endotoxin and double stranded RNA render macrophagescvtotoxic. Nature [New Biol] 232:76-78, 1971
258. Currie GA, Basham C: Activated macrophages release a factor which lyses malig-nant cells but not normal cells. J Exp Med 142:1600-1605, 1975
259. Doe WF, Henson PM: Macrophage stimulation by bacterial lipopolysaccharides.I. Cvtolvtic effect on tumor target cells. J Exp Med 148:544-556, 1978
260. Doe WF, Yang ST, Morrison DC, Betz SJ, Henson PMI: Macrophage stimulationby bacterial lipopolysaccharides. II. Evidence for independent differentiation sig-nals delivered by lipid A and a protein rich fraction of LPS. J Exp Med 148:557-568, 1978
261. Russell SW, Doe WF, Nlcintosh AT: Functional characterization of a stable,noncytolytic stage of macrophage activation in tumors. J Exp Med 146:1511-1520,1977
262. Hibbs JB, Jr, Taintor RR, Chapman HA Jr, Weinberg JB: Macrophage tumorkilling: Influence of the local environment. Science 197:279-282, 1977
263. Hays EG, Forsen NR, Rodensky D, Craddock CG: The effect of endotoxin oncolony stimulating factor (CSF) release from granulocvtes and monocytes in vitro.Blood 40:949, 1972 (Abstr)
264. Eaves AC, Bruce WR: In vitro production of colony-stimulating activity. I. Ex-posure of mouse peritoneal cells to endotoxin. Cell Tissue Kinet 7:19-0, 1974
265. Ruscetti FW, Chervenick PA: Release of colony-stimulating factor from mono-cytes by endotoxin and polyinosinic-polycytidylic acid. J Lab Clin Med 83:64-72,1974
266. Apte RN, Hertogs CF, Pluznik DH: Regulation of lipopolysaccharide-inducedgranulopoiesis and macrophage formation b) spleen cells. I. Relationship betweencolony-stimulating factor release and lvmphocyte activation in vitro. J Immunol118:1435-1440, 1977
267. Apte RN, Galanos C, Pluznik DH: Lipid A, the active part of bacterial endotoxinsin inducing serum colony stimulating activity and proliferation of splenic gran-ulocyte/macrophage progenitor cells. J Cell Physiol 87:71-117, 1976
268. Nowotnv A, Behling UH, Change HL: Relation of structure to fuinction in hacterialendotoxins. VIII. Biological activities in a polysaccharide-rich fraction. J Ininitinol115:199-203, 1975
269. Anderson FL, Tsagaris TJ, Jubiz W, Kuida H: Prostaglandin F aind E levels

616 MORRISON AND ULEVITCH American Journalof Pathology
during endotoxin-induced pulmonary hypertension in calves. Am J Physiol228:147-9-1482, 1973
270. Anderson FL, Jubiz WV, Tsagaris TJ, Kuida H: Endotoxin-induced prostaglandinE and F release in dogs. Am J Physiol 228:410-414, 1975
271. Herman AG, V-ane JR: Release of renal prostaglandins during endotoxin-inducedhypotension. Eur J Pharmacol 39:79-90, 1976
272. Fletcher JR, Ramwell PWT, Herman CM: Prostaglandins and the hemodynamiccourse of endotoxin shock. J Surg Res 20:389-594, 1976
273. Rosenstreich DL, Glode LM, WVahl LM, Sandberg AL, MergenhagenSE: Analysis of the cellular defects of endotoxin unresponsive C3H/ HeJ mice.2pp314-320
274. Kurland JL, Bockrnan R: Prostaglandin E production by human blood monocytesand mouse peritoneal macrophages. J Exp Med 147:952-957, 1978
275. Glode LM, Mergenhagen SE, Rosentreich DL: Significant contribution of spleencells in mediating the lethal effects of endotoxin in civo. Infect Immun 14:626-630,1976
276. Humes JL, Bonney RJ, Pelus L, Dahlgren ME, Sadowski SJ, Kuehl FA Jr, DaviesP: Macrophages synthesize and release prostaglandins in response to inflamma-tory stimuli. Nature 269:149-131, 1977
277. Tanaka N, Nishimura T, Yoshiyuki T: Histochemical studies on the cellulardistribution of endotoxin of Salmonella enteritidis in mouse tissues. Jpn J Mlicrobiol3:191-199, 1959
278. Rubenstein HS, Fine J, Coons AH: Localization of endotoxin in the walls of theperipheral vascular system during lethal endotoxemia. Proc Soc Exp Biol Med111:458-467, 1962
279. McGrath JM, Stewart GJ: The effects of endotoxin on vascular endothelium. JExp Med 129:833-840, 1969
280. Gay-nor E, Bouvier C, Spaet TH: Vascular lesions: Possible pathogenetic basis ofthe generalized Shwartzman reaction. Science 170:986-988, 1970
281. Gaynor E: Increased mitotic activity in rabbit endothelium after endotoxin: Anautoradiographic study. Lab Invest 24:318-320, 197-1
282. Gerrity RG, Caplan BA, Richardson NI, Cade JF, Hirsh J, SchwartzCJ: Endotoxin-induced endothelial injury and repair. I. Endothelial cell turnoverin the aorta of the rabbit. Exp Mol Pathol 23:379-85, 1975
2&3. Evensen SA, Pickering RJ, Batbouta J, Shepro D: Endothelial injury induced bybacterial endotoxin: Effect of complement depletion. Eur J Clin Invest 3:463-469,1975
284. Raij L, Keane WNF, Michael AF: Unilateral Shwartzman reaction: Cortical necro-sis in one kidney following in vivo perfusion with endotoxin. Kidney Int 12:91-95,1977
285. Naclntvre DE, Pearson JD, Gordon JL: Localization and stimulation of pros-tacyclin production in vascular cells. Nature 271:549-551, 1978
286. Fitzpatrick TNM, Alter I, Corey EJ, Ramwell PWN, Rose JC, Kot PA: Cardiovascularresponses to PGI2 (prostacyclin) in the dog. Circ Res 42:192-194, 1978
287. Howes EL Jr, Kwok N\IT, NIcKay DG: The effects of indomethacin on the general-ized Shwartzman reaction. Am J Pathol 90:7-22, 1978
288. Bach NIK (editor): Immediate Hvpersensitiv-itv-N\lodern Concepts and Develop-ments. New York, NMarcel Dekkar, Inc., 1978
289. Nlaclean LD, XVeil NH: Hvpotension (shock) in dogs produced by Escherichiacoli endotoxin. Circ Res 4:546-536, 1956
290. Hinshaw LB: The relase of vasoactive agents by endotoxin. pp 118-125291. Glick D, Pothaprogada S: Succinic dehydrogenase system in isolated mast cells
from the rat. Proc Soc Biol MIed 106:359-362, 1961

Vol. 93, No. 2 BACTERIAL ENDOTOXINS 617November 1978
292. Antweiler H, Hirsch E: Zur Frage einer akuten Mastzellschadigung und Histaminfreisetzung durch Endotoxine. Naunyn Schmiedebergs Arch Pharmacol 241:213-214, 1961
293. Gustafson GT, Cronberg S: Effect of endotoxin on mast cells and the extension ofthe local Shwartzman reaction to the hamster. Acta Pathol Microbiol Scand 59:21-31, 1963
294. Hook WA, Snyderman R, Mergenhagen SE: Histamine-releasing factor gener-ated by the interaction of endotoxin with hamster serum. Infect Immun 2:462-467,1970
295. Urbaschek B: Zur Pathophysiologie der initialen Wirkung bakterieller Endo-toxine. Zentralbl Bakteriol [Orig A] 212:537-544, 1969
296. J6kay I, Karczag E: Cellular and permeability changes in the peritoneal cavity ofmice after injection of endotoxin. Z Immunitaetsforsch 148:414-424, 1975
297. Jokay I, Karczag F: Effect of endotoxin on the peritoneal mast cells in mice. ActaBiol Acad Sci Hung 27:119-124, 1976
298. Asboe-Hansen G, Glick D: Influence of various agents on mast cells isolated fromrat peritoneal fluid. Proc Soc Exp Biol Med 98:458-461, 1958
299. Sandusky CB, Johnson AR, Moran NC: Effect of endotoxin and anaphylatoxin onrat mast cells and chopped rat lung. Proc Soc Exp Biol Med 143:764-768, 1973
300. Morrison DC, Betz SJ: Chemical and biological properties of a protein-rich frac-tion of bacterial lipopolysaccharides. II. The in vitro rat peritoneal mast cellresponse. J Immunol 119:1790-1795, 1977
301. Greisman SE: Activation of histamine-releasing factor in normal rat plasma by E.coli endotoxin. Proc Soc Exp Biol Med 103:628-632, 1960
302. Lichtenstein LM, Gewurz H, Adkinson NF Jr, Shin HS, MergenhagenSE: Interactions of the complement system with endotoxic lipopolysaccharide:The generation of an anaphylatoxin. Immunology 16:327-336, 1969
303. Hook WA, Snyderman R, Mergenhagen S: Further characterization of a factorfrom endotoxin-treated serum which releases histamine and heparin from mastcells. Infect Immun 5:909-914, 1972
304. Glauser FL, Palmer J, Cecconi S, Schoolcraft W, Wells I, Novey H, Egan P,Smeltzer D: The effect of endotoxin on the mast cell AMP system. Ann Allergy38:104-107, 1977
Adm m .ntThe authors express their deep appreciation to Ms. Monica Bartlett, Betty Goddard, and Margaret
Stone for their patience and assistance during the preparation of this review.

618 MORRISON AND ULEVITCH Amerkcan Joumalof Patology
[End ofArticle]