the etiology of paraneoplastic autoimmunity
TRANSCRIPT

The Etiology of Paraneoplastic Autoimmunity
Emanual Maverakis & Heidi Goodarzi & Lisa N. Wehrli &Yoko Ono & Miki Shirakawa Garcia
Published online: 19 January 2011# Springer Science+Business Media, LLC (outside the USA) 2011
Abstract Although they may sometimes appear similar,paraneoplastic autoimmunity has a unique pathogenesis,different from the classical autoimmune diseases notassociated with cancer. When distinguished clinically,paraneoplastic autoimmunity is more severe and oftenpresents with a broader range of clinical signs andsymptoms. Management of these patients is difficult andis usually centered in part on treatment of the underlyingmalignancy. Self-antigens recognized in the setting ofparaneoplastic autoimmunity can be diverse, and thenumber of determinants recognized within a single antigencan be numerous. This review uses prototypic examples ofparaneoplastic immune-mediated diseases and their associatedmalignancies to describe the mechanisms by which immunedysregulation can occur in the setting of cancer. Specificdiseases covered include paraneoplastic pemphigus, Sweet’ssyndrome, pyoderma gangrenosum, thymoma-associatedmultiorgan autoimmunity, myasthenia gravis, autoimmunehemolytic anemia, immune thrombocytopenia, and theparaneoplastic neurological syndromes. The malignanciesdiscussed include thymoma, non-Hodgkin’s lymphoma,and chronic lymphocytic leukemia, among others. Themechanisms by which cancers induce autoimmunity are
broken down into the following categories: disruption ofcentral tolerance, peripheral immune dysregulation, andalteration of self-antigens. For each category, examplesof paraneoplastic autoimmune diseases and their associ-ated malignancies are discussed. Finally, mechanisms bywhich cancer treatment can lead to autoimmunity andexamples of polymorphisms that are linked to bothcancer and autoimmunity are discussed.
Keywords Paraneoplastic pemphigus .Myasthenia gravis .
Autoimmunity . Self . Non-Hodgkin’s lymphoma . CLL .
Pyoderma gangrenosum . Sweet’s syndrome . Thymoma .
Pemphigus vulgaris
Similar in Appearance But Distinct MolecularMechanisms
It is sometimes clinically and histologically challenging todistinguish a paraneoplastic autoimmune phenomenon fromrun of the mill autoimmune disease. Using paraneoplasticpemphigus as a prototypic example, patients may presentwith oral erosions and flaccid blisters that are indistin-guishable from pemphigus vulgaris, another severe auto-immune blistering disease. However, concluding that theunderlying neoplasm simply induced the development ofpemphigus vulgaris would be incorrect. At an elementarylevel, the pathophysiological mechanisms of the diseasesmight seem very similar; both paraneoplastic pemphigusand pemphigus vulgaris can have autoreactive antibodiesdirected against cell surface adhesion molecules of thecadherin family, desmoglein 1 (160 kDa) and desmoglein 3(130 kDa) [1]. In each case, binding of autoreactiveantibodies to the keratinocyte desmogleins leads to epider-mal separation (acantholysis) and blistering, but the events
E. Maverakis (*) :H. Goodarzi : L. N. Wehrli :Y. Ono :M. S. GarciaDepartment of Dermatology, University of California Davis,School of Medicine,3301 “C” Street, Suite 1400,Sacramento, CA 95816, USAe-mail: [email protected]
E. MaverakisDepartment of Veteran Affairs Northern CaliforniaHealth Care System,Sacramento, CA 95655, USA
Clinic Rev Allerg Immunol (2012) 42:135–144DOI 10.1007/s12016-010-8248-5

that result in the formation of these antibodies are likelyvery distinct.
Evidence in support of this theory comes from thedetailed characterization of the autoreactive antibodies. Inpemphigus vulgaris, autoreactive antibodies are mainly ofthe IgG4 subclass and are directed against the N-terminalextracellular domain of desmoglein 3. In contrast, theautoreactive antibodies in paraneoplastic pemphigus arepredominantly of the IgG1 subclass with some being IgG2,and the epitopes recognized by these are distributedthroughout the extracellular domain of desmoglein 3, ratherthan being concentrated just at its N terminus [2]. There isalso a greater diversity of antigens recognized by theautoreactive immune response in patients with paraneo-plastic pemphigus. Other common autoantigens includedesmoplakin I (250 kDa), desmoplakin II (210 kDa),bullous pemphigoid antigen (230 kDa), envoplakin(210 kDa), periplakin (190 kDa), and plectin (500 kDa),among others. Thus, a greater degree of “determinantspreading” occurs in the setting of this malignancy-induced autoreactive immune response [3, 4]. There arenot only more autoantigens being recognized, but there arealso a greater number of epitopes recognized within theindividual autoantigens (Fig. 1).
Paraneoplastic Autoimmune Diseases May Havea Unique and Severe Presentation
The management of paraneoplastic autoimmunity is oftencentered on treatment of the underlying malignancy, withclinical improvement usually noted after successful chemo-therapy or surgical excision of the tumor [5]. In the absenceof these options, aggressive immunosuppression is fre-quently required to control the aberrant immune response.In addition to being more refractory to therapy, the clinicalpresentations of paraneoplastic immune-mediated diseasesare often more severe than their non-malignancy-inducedcounterparts. For example, patients with paraneoplasticpemphigus will often develop a severe stomatitis that isrefractory to aggressive therapy [6–8]. These patients mayalso develop bronchiolitis obliterans and die of respiratoryfailure [9, 10]. Given the severe and diverse presentations,it has been suggested that the paraneoplastic diseases ofepithelial adhesion be known as paraneoplastic autoimmunemultiorgan syndrome [11].
In addition to autoreactive immune responses, there are avariety of other immune-mediated diseases that haveincreased severity when associated with a malignancy.Sweet’s syndrome (i.e., acute febrile neutrophilic dermato-sis) is characterized by fever, neutrophilia, and edematouscutaneous lesions. On histological examination, theselesions are filled with neutrophils and karyorrhexis in the
absence of vasculitis. Sweet’s syndrome can be associatedwith inflammatory bowel disease, infections, pregnancy, ormedications. It is also associated with hematologic malignan-cies, usually acute myelogenous leukemia or myelodysplasticsyndrome [12]. When associated with malignancy, Sweet’ssyndrome can have a more severe presentation with bullaeand ulcerations of the skin [13, 14]. Oddly enough, theseneutrophilic lesions can occur in leukemic patients who areseverely neutropenic. The paraneoplastic form of Sweet’ssyndrome is also more likely to have extracutaneousmanifestations including musculoskeletal symptoms, conjunc-tivitis, episcleritis, glomerulonephritis, pulmonary infiltrates,and mucous membrane ulcers.
Pyoderma gangrenosum (PG) is another neutrophilicdisease that often has a unique presentation in the setting ofmalignancy. PG is an ulcerative skin disease that is mostfrequently associated with inflammatory bowel disease orrheumatoid arthritis [15–19]. When associated with myelo-dysplastic syndrome, myeloma, paraproteinemia, or leuke-mia, PG lesions may have an atypical vesiculobullouspresentation and the distribution of the ulcers can includeatypical sites such as the hands [8, 20, 21]. The immuno-logic mechanism that results in PG is not well understood,but gross elevations of numerous cytokines and proteinshave been reported, including: IL-1β, IL-6, IL-8, IFN-γ, G-CSF, TNF-α, MMP-9, MMP-10, and Elafin [22–27].Moreover, patients with PG respond well to TNF-αblockade with adalimumab and infliximab [28]. Althoughthe skin lesions of PG are dominated by neutrophils, thesepatients are known to have T cell clonal expansions andlymphocytes are dominant in very early PG lesions [29–31].
“Central” Immune Dysregulation in the Settingof Malignancy
Educating the immune system to distinguish self from non-self is a highly sophisticated process that involves bothcentral and peripheral safeguards. A malignancy can disruptthis process at several points, the most basic of which isthymic education. The thymus is comprised of a centralmedulla, peripheral cortex, and an outer capsule. Thestromal cells of the thymus include thymic corticalepithelial cells, thymic medullary cells, and dendritic cells.The cortex is comprised of lymphoid cells and epithelialreticular cells, which form an adventitia to the bloodvessels. It is within the cortex that T cell receptor (TCR)gene rearrangements take place and where the T cellsbecome positively selected. In positive selection, T cellsbearing TCRs with affinity for peptide-bound majorhistocompatibilty complex (MHC) class I or II moleculesare selected for further development. T cells with non-functional TCR gene rearrangements, or those that cannot
136 Clinic Rev Allerg Immunol (2012) 42:135–144

Fig. 1 The pathogenesis of paraneoplastic autoimmunity is unique. aYoung boy with paraneoplastic pemphigus presented with severeintractable stomatitis and flaccid blisters on his trunk and extremities.b General histology of the biopsy specimen revealed a vesiculobullousdermatitis with neutrophils and eosinophils. Direct immunofluores-cence (not shown here) exhibited variable linear and granulardeposition of IgG and C3 along the dermal epidermal junction. c CTscan revealed an abdominal mass (arrow), which on biopsy wasconsistent with Castleman’s disease. d Normal keratinocytes areconnected by desmosomes (i.e., macula adherens). These structures
are specialized for cell-to-cell adhesion. Desmogleins and desmocol-lins, all part of the cadherin family, are transmembrane proteins thatbridge the extracellular space between keratinocytes. These proteinsare then linked via plakoglobin, desmoplakin, and plakophilin tointracellular keratin intermediate filaments. e In pemphigus vulgarisautoreactive antibodies (IgG4) bind to the N-terminal extracellulardomain of desmoglein proteins, causing acantholysis of the keratino-cytes (arrow in b). f In paraneoplastic pemphigus, autoreactiveantibodies (IgG1 and IgG2) bind to multiple desmoglein determinantsas well as several other desmosome and hemidesmosome proteins
Clinic Rev Allerg Immunol (2012) 42:135–144 137

interact with a peptide-bound MHC complex, are deleted.Even a single peptide-bound MHC complex is capable ofselecting for a reasonably diverse T cell repertoire [32].
The medulla is the predominant site for T cell negativeselection, which removes autoreactive T cells from themature repertoire. T cells that interact too strongly withpeptide-bound MHC complexes are deleted. In order forthis process to effectively purge autoreactive T cells fromthe mature T cell repertoire, a diverse set of organ-specificself-antigens must be expressed in the thymus, a processcontrolled by the autoimmune regulator (AIRE). When self-antigens are expressed in the thymus, they are processedand presented to maturing thymocytes. Each determinant isprocessed differently, resulting in a hierarchy of determi-nant display. Thymic T cells expressing TCRs with highaffinity for well-processed self-determinants undergo neg-ative selection. The ability of a self-determinant to inducenegative selection is directly related to its expression levelin the thymus. T cells expressing high affinity TCRsspecific to poorly presented determinants can avoidnegative selection and escape to the periphery [33–36].The importance of thymic education is highlighted byautoimmune polyendocrinopathy-candidiasis-ectodermaldystrophy, a severe autoimmune syndrome that occurswhen there is a genetic defect in AIRE. It is thereforelogical that any malignancy-induced change in eitherpositive selection, negative selection, antigen expression,or antigen processing and presentation would have pro-found effects on the circulating mature T cell repertoire.
The most obvious way a malignancy can affect thymiceducation is through presentation in the thymus itself.Thymoma is the most common neoplasm arising from thethymus; other tumors include: lymphomas, germ celltumors, thymic carcinoma, carcinoid tumors, and othermalignancies that can metastasize to the site. Thymomashave an incidence of 0.15 cases per 100,000 and occur inpatients between 40 and 60 years old [37]. Histologically,thymomas are composed of thymic epithelial cells and non-neoplastic lymphocytes. Therefore, the non-neoplasticlymphocytes are maturing in an abnormal environment[38], incapable of appropriately orchestrating central toler-ance. Many of the T cells that are normally deleted in thethymus interact too strongly with self-MHC (HLA-A, -B,and -C and HLA-DQ, -DR, and -DP). In thymoma-associated multiorgan autoimmunity (TAMA), these strong-ly interacting T cells fail to be deleted and, once they escapeto the periphery, they act similarly to grafted allogeniclymphocytes following an unmatched bone marrow trans-plant. The resulting disease is similar, clinically andhistologically, to graft-versus-host disease. Patients withTAMA present with a morbiliform skin eruption, chronicdiarrhea, and abnormal liver enzymes (Fig. 2). The diseaseis often fatal [39].
A more common autoimmune disease associated withthymoma is myasthenia gravis (MG). This disease resultsfrom autoantibodies directed against the acetylcholinereceptors (AChR), located in the postsynaptic villi ofvoluntary muscles. In some patients with MG, the autor-eactive B cells reside directly within the thymoma. Amalignant thymus might be ideal for development of anautoreactive antibody response as both AChR-specific Tcells and AChR protein have been shown to reside there[40]. This is also evident by the fact that removal of thethymoma usually results in improvement of thymoma-associated MG. Other examples of thymoma-associatedautoimmune diseases include pure red blood cell aplasia,hypogammaglobulinemia, and paraneoplastic pemphigus[41–46]. The finding of antibody producing B cells residingin the tumor itself is not limited to malignancies involving thethymus. In paraneoplastic pemphigus, autoantibodies havebeen elicited in both the serum of patients [47] andfrom cultured tumor cells [48, 49]. Specifically, antibodiesto desmoplakins I and II, periplakin, plectin, envoplakin,170-kDa antigen, and desmogleins 1 and 3 have beendetected [10, 47–49]. Although the mechanism of paraneo-plastic pemphigus is not clear, it has been hypothesized thatthe cells in the tumors can produce the autoantibodies [10,50]. Likewise, defects in T cell development are not limitedto malignancies involving the thymus. There are likely manytypes of tumors that can also support the development ofimmature (CD3, TdT, CD4/CD8 double positive) T cells,setting the stage for autoimmunity in these patients. At leastone such case has been reported in the literature [51], but thislikely presents a common mechanism for paraneoplasticautoimmunity. Immature T cells migrating to a tumor willnot undergo appropriate negative selection.
“Peripheral” Immune Dysregulation in the Settingof Malignancy
Although an essential mechanism, central tolerance alone isinsufficient to prevent the induction of autoimmunity. Addi-tional peripheral safe guards are therefore required to limit theaberrant activation of T cells, the most important of which arethe co-stimulatory requirements for T cell activation [52]. TCRspecificity for a peptide-bound MHC complex alone isinsufficient to induce optimal T cell activation; naïve T cellsrequire a co-stimulatory signal in addition to TCR engage-ment. In the absence of this co-stimulation, T cells becomeunresponsive when they encounter APCs bearing cognateMHC-bound peptide antigens. The classic co-stimulatorysignal is provided by CD28, which is constitutively expressedby T cells [52]. The ligands for CD28 are B7-1 (CD80) andB7-2 (CD86). With the exception of dendritic cells, B7expression by immature APCs is low. However, when anAPC
138 Clinic Rev Allerg Immunol (2012) 42:135–144

Fig. 2 Autoimmunity as a result of central immune dysregulation.Left panel depicts a patient with TAMA, which is similar, clinicallyand histologically, to graft-versus-host disease. Patients present with amorbiliform skin eruption, chronic diarrhea, and abnormal liverenzymes. Diffuse erythematous plaques with scale are distributedover the patient’s a back and b chest. c Palms with diffuse erythemaand some focal areas with scale. d Normal thymocyte development.Common lymphoid progenitor cells migrate to the thymus as early Tcell lineage progenitors (ETP) cells. Once in the thymus, ETP cells(CD3-CD4-CD8-CD25-CD44hiKIT+) progress through differentstages of double negative cells DN2 (CD4-CD8-CD25+CD44+),DN3 (CD4-CD8-CD25+CD44-), and then finally DN4 cells (CD4-CD8-CD25-CD44-), which express a pre-TCR. They then becomeCD4+CD8+ double positive cells and somatic DNA rearrangementgenerates a functional T cell receptor (TCR), allowing the cell to gothrough positive selection. Selected CD4+ or CD8+ single positivecells enter the medulla where they will stay for prolonged periods oftime before being released into the periphery. Importantly, in thethymus expression of organ-specific antigens is controlled by theautoimmune regulator (AIRE), a gene essential for normal thymiceducation. Multiple antigens expressed in the medulla are depicted as
the multicolor epithelial cells in the background of the panel.Autoreactive T cells (shown as red-fragmented cells in this figure)undergo apoptosis and thereby are purged from the T cell repertoirebefore entering the peripheral circulation. e In the setting of thymoma,AIRE continues to allow expression of organ-specific self-antigens butthe thymic epithelial cells are now transformed and the carefullyorchestrated thymocyte developmental pathway is altered. Non-malignant ETP cells continue to enter the thymus but theirdevelopment is affected by the malignancy. Any change in antigenprocessing, organ-specific protein expression, or co-stimulatorysignals could allow the non-malignant thymocytes to escape negativeselection. Autoreactive T cells (shown as red cells in this figure) canbe either double positive or single positive. When they enter theperipheral circulation, these autoreactive T cells act in a similarfashion as allogenic T cells after a bone marrow transplantation.Sometimes, the autoreactive T cells remain in the thymus, where theyactivate autoreactive B cells in the presence of self antigen, whichmight be provided by AIRE. In such cases, removal of the thymomacan lead to improvement of the patient’s malignancy-associatedautoimmunity
Clinic Rev Allerg Immunol (2012) 42:135–144 139
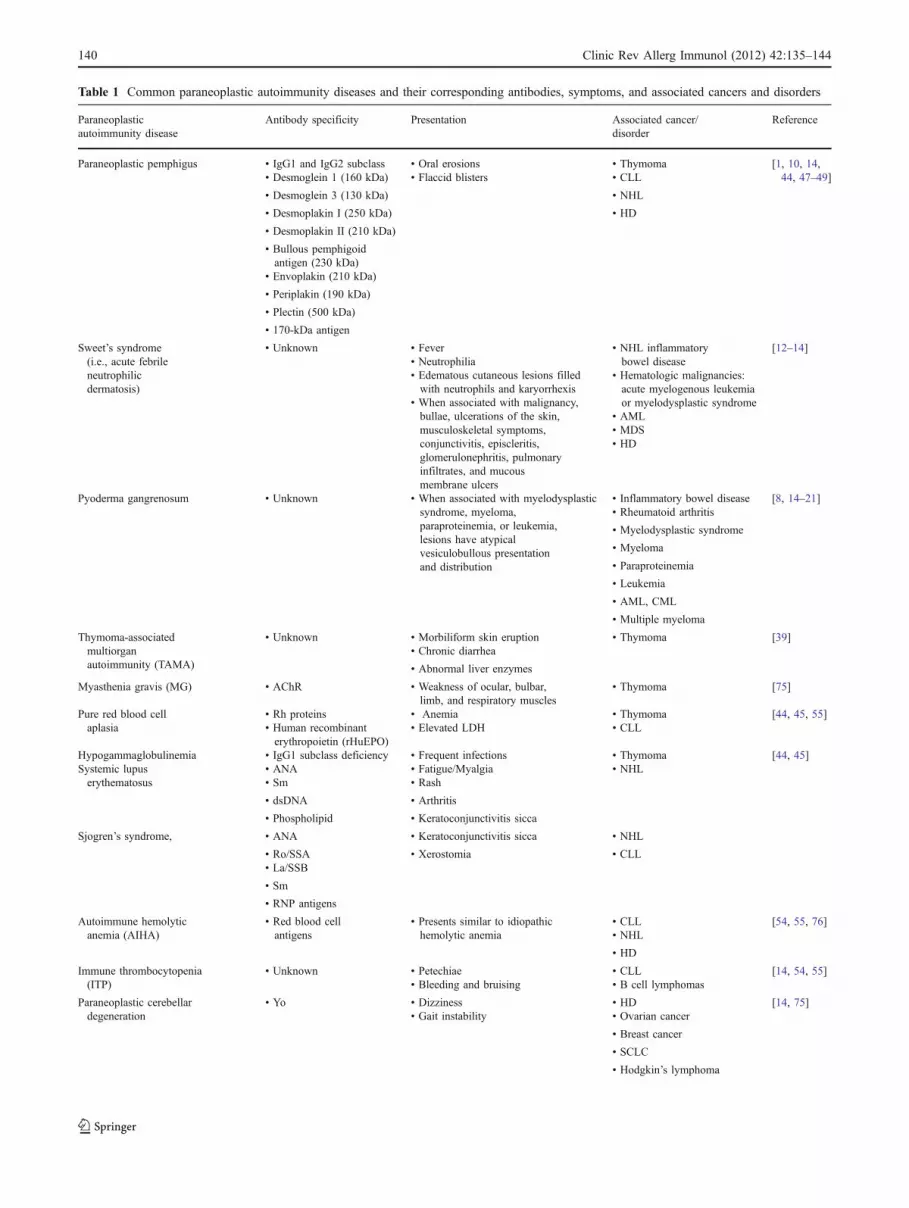
Table 1 Common paraneoplastic autoimmunity diseases and their corresponding antibodies, symptoms, and associated cancers and disorders
Paraneoplasticautoimmunity disease
Antibody specificity Presentation Associated cancer/disorder
Reference
Paraneoplastic pemphigus • IgG1 and IgG2 subclass • Oral erosions • Thymoma [1, 10, 14,44, 47–49]• Desmoglein 1 (160 kDa) • Flaccid blisters • CLL
• Desmoglein 3 (130 kDa) • NHL
• Desmoplakin I (250 kDa) • HD
• Desmoplakin II (210 kDa)
• Bullous pemphigoidantigen (230 kDa)
• Envoplakin (210 kDa)
• Periplakin (190 kDa)
• Plectin (500 kDa)
• 170-kDa antigen
Sweet’s syndrome(i.e., acute febrileneutrophilicdermatosis)
• Unknown • Fever • NHL inflammatorybowel disease
[12–14]• Neutrophilia
• Hematologic malignancies:acute myelogenous leukemiaor myelodysplastic syndrome
• Edematous cutaneous lesions filledwith neutrophils and karyorrhexis
• AML• When associated with malignancy,bullae, ulcerations of the skin,musculoskeletal symptoms,conjunctivitis, episcleritis,glomerulonephritis, pulmonaryinfiltrates, and mucousmembrane ulcers
• MDS• HD
Pyoderma gangrenosum • Unknown • When associated with myelodysplasticsyndrome, myeloma,paraproteinemia, or leukemia,lesions have atypicalvesiculobullous presentationand distribution
• Inflammatory bowel disease [8, 14–21]• Rheumatoid arthritis
• Myelodysplastic syndrome
• Myeloma
• Paraproteinemia
• Leukemia
• AML, CML
• Multiple myeloma
Thymoma-associatedmultiorganautoimmunity (TAMA)
• Unknown • Morbiliform skin eruption • Thymoma [39]• Chronic diarrhea
• Abnormal liver enzymes
Myasthenia gravis (MG) • AChR • Weakness of ocular, bulbar,limb, and respiratory muscles
• Thymoma [75]
Pure red blood cellaplasia
• Rh proteins • Anemia • Thymoma [44, 45, 55]• Human recombinanterythropoietin (rHuEPO)
• Elevated LDH • CLL
Hypogammaglobulinemia • IgG1 subclass deficiency • Frequent infections • Thymoma [44, 45]Systemic lupuserythematosus
• ANA • Fatigue/Myalgia • NHL• Sm • Rash
• dsDNA • Arthritis
• Phospholipid • Keratoconjunctivitis sicca
Sjogren’s syndrome, • ANA • Keratoconjunctivitis sicca • NHL
• Ro/SSA • Xerostomia • CLL• La/SSB
• Sm
• RNP antigens
Autoimmune hemolyticanemia (AIHA)
• Red blood cellantigens
• Presents similar to idiopathichemolytic anemia
• CLL [54, 55, 76]• NHL
• HD
Immune thrombocytopenia(ITP)
• Unknown • Petechiae • CLL [14, 54, 55]• Bleeding and bruising • B cell lymphomas
Paraneoplastic cerebellardegeneration
• Yo • Dizziness • HD [14, 75]• Gait instability • Ovarian cancer
• Breast cancer
• SCLC
• Hodgkin’s lymphoma
140 Clinic Rev Allerg Immunol (2012) 42:135–144

is activated, B7 becomes upregulated, providing the appropriatesecondary signals for T cell stimulation. Once activated, CD4+T cells will express CD40L, an important molecule for T celleffector function. CD40L in turn binds to CD40 on the surfaceof APCs including dendritic cells, macrophages, and B cells.Ligation of CD40 on the surface of APCs greatly increases theAPCs’ antigen presentation and co-stimulatory capacity.
Unlike immature APCs, a variety of lymphomas,including non-Hodgkin’s lymphoma (NHL), can expressB7 and thereby act as excellent APCs, effectively bypassingan important autoimmunity safeguard. In addition, it hasbeen shown that tumors can directly support naïve T cellinfiltration, activation, and differentiation into effector cells[53]. Thus, naïve T cells can bypass the lymph nodes alltogether and migrate straight into the tumor. Autoimmunediseases associated with NHL include paraneoplasticpemphigus, Sweet’s syndrome, systemic lupus erythemato-sus, and Sjogren’s syndrome, among others.
Autoimmune complications also occur in 10% to 25% ofpatients with chronic lymphocytic leukemia (CLL). Auto-immune hemolytic anemia (AIHA) and immune thrombo-cytopenia are the most common types [54, 55]. Moreover,non-hematological autoimmunity occurs in 1% to 16% ofpatients with CLL. Sjogren’s syndrome is the mostcommon in these cases, but other forms of autoimmunitysuch as paraneoplastic pemphigus have also been reported.As was the case in NHL, it is likely that CLL serves insome capacity as an APC, directly activating autoreactiveimmune responses. In favor of this mechanism, CLL canexpress MHC, B7-1, B7-2, CD83, and CD54 [56, 57], andhigher β2 microglobulin expression correlates with anincreased incidence of autoimmunity. Furthermore, CD5+
B-CLL cells have been shown to process and presentaberrant red blood cell epitopes and induce AIHA [58].
In addition to activating immune responses, CLL cansuppress some immune functions, a potential mechanism toprevent anti-leukemic immune responses. Specifically, non-malignant CD4+ and CD8+ T cells co-cultured with CLL havedecreased cytoskeleton remodeling and decreased vesicletrafficking, resulting in impaired immune synapse formation[59, 60]. Theoretically, such pathways could lead to impairmentof regulatory T cell function and induction of autoimmunity.
Altered Self and Cancer
The immune system is a final safeguard to the developmentof cancer, as illustrated by the increased numbers ofaggressive skin cancers seen in immunosuppressed indi-viduals. Thus, the ability of the immune system torecognize a cancer as something “foreign” is paramount tosurvival of the individual. In addition to NK cells, theadaptive immune response also plays a role in cancersurveillance. As part of a healthy cell’s transformation intoa cancer cell, multiple genetic mutations occur, some ofwhich result in amino acid alterations, thereby creatingnovel antigens. Malignancies can also result in aberrantupregulation of fetal proteins or overexpression of proteinsusually sequestered in immunoprivileged sites, such as thecentral nervous system. Finally, tumor necrosis can result ininflammation and release of self-antigens. The end result istumor reactive antibodies which can recognize a variety ofantigens including: oncoproteins P185, 1-myc, and c-myb; thetumor suppressor gene P53; proliferation-associated antigens
Table 1 (continued)
Paraneoplasticautoimmunity disease
Antibody specificity Presentation Associated cancer/disorder
Reference
Paraneoplastic encephalomyelitis • Hu (ANNA-1) • Subacute sensory neuronopathy • SCLC [75]
Paraneoplastic anti-NMDARencephalitis
• NMDAR • Confusion • Teratoma, frequentlyovarian
[77]• Restlessness
• Agitation
• Frequent paranoid ordelusional thoughts
• Seizures
Stiff person syndrome • Gephyrin • Rigidity of musculature • Breast cancer [14, 78]• Amphiphysin • Spasms • HD
• SCLC
Cancer-associated retinopathy (CAR) • Recoverin (antigen) • Vision loss due to retinal degeneration • SCLC [75, 79]• Breast cancer
Paraneoplastic opsoclonus–myoclonus ataxia (POMA)
• NOVA-1 • Oscillopsia • Breast cancer [80]• Myoclonus
Lambert–Eaton syndrome • Anti-N-type calciumchannel
• Muscle fatigue • SCLC [75, 81]• Depressed tendon reflexes
CLL chronic lymphocytic leukemia, NHL non-Hodgkin’s lymphoma, HD Hodgkin’s lymphoma, SCLC single cell lung carcinoma
Clinic Rev Allerg Immunol (2012) 42:135–144 141

cyclin A, B1, D1, E CENP-F, CDK, and U3-RNP; onconeuralantigens Hu, Yo, Ri, and Tr; cancer/testis antigens MAGE,GAGE, BAGE, SSX, ESO, SCP, and CT7; and rheumatologicantigens RNP and Sm [61]. Table 1 lists several paraneoplasticautoimmune diseases and their associated autoantibodies.
Paraneoplastic neurological syndromes are a result ofimmune-mediated damage to neural tissue. Examples ofthis include paraneoplastic cerebellar degeneration, limbicencephalitis, stiff person syndrome, sensorimotor peripheralneuropathy, and certain inflammatory myopathies [14]. Inthese diseases, tumors express antigens that are normallyrestricted to neurons. Antibodies produced in response tothese onconeural antigens then cause pathology at distantsites in the central or peripheral nervous system. Impor-tantly, the autoreactive immune response suppresses theunderlying tumor growth and patients with malignancy-associated paraneoplastic neurological syndromes often dobetter than patients without these syndromes [14].
Treatment-Induced Autoimmunity
Cytotoxic agents that are used to treat cancer can causewidespread tissue damage, resulting in the release of autoan-tigens. In general, these regimens are globally immunosuppres-sive, but not all immune cells are affected equally. It has beensuggested that Tregs are relatively more susceptible to somechemotherapy regimens. Fludarabine and chlorambucil arethought to be especially active against Tregs [62–64]. Interest-ingly, peripheral blood mononuclear cells isolated from CLLpatients express CD154 when cultured in vitro with purineanalogues [65] and CD154 expression has been associated invivo with development of autoimmunity and resistance tofludarabine [65]. The fact that there have been several cases ofparaneoplastic pemphigus following fludarabine therapy isevidence in favor of this hypothesis [66]. As immunotherapyfor cancer becomes more popular, cases of treatment-inducedautoimmunity will become more common. For instance,ipilimumab blocks CTLA4, which allows for unopposed Tcell activation. Patients with metastatic melanoma haveexperienced improved survival with ipilimumab; however, itis known to cause autoreactive immune responses [67].
Polymorphisms Linked to Cancer Can Also Be Linkedto Autoimmunity
Patients with rheumatoid arthritis and other autoimmunediseases have an excess risk of cancer including lympho-mas, leukemia, and myelomas. Many factors may contrib-ute to this risk including therapeutic medications andchronic immune activation. However, many polymorphismsthat predispose patients to autoimmunity also predispose
them to the development of cancer. For example, SNPanalysis showed that interferon regulatory factor 4(IRF4) polymorphisms are linked to an excess risk ofCLL [68, 69]. IRF4 is also important in B cell activation,IgM secretion, thymocyte differentiation, and macrophagepolarization. It has been shown that phosphorylation ofIRF4 by ROCK2 regulates IL-17 and IL-21 productionand the development of autoimmunity in mice [70]. Otherexamples include elevation of B cell-activating factor andpolymorphisms in tumor necrosis factor (BAFF;TNFSF13B), both of which are associated with lymphomaand autoimmunity [71, 72]. With respect to BAFF, CD19+cells from SLE patients have it aberrantly upregulation[73], and in CLL cells, BAFF can enhance expression ofc-MYC via activation the canonical IkappaB kinase/NF-kappaB pathway, which is important for leukemia-cellsurvival [74].
Acknowledgments EM holds career awards from the BurroughsWellcome Fund and the Howard Hughes Medical Institute.
References
1. Amagai M, Nishikawa T, Nousari HC, Anhalt GJ, Hashimoto T(1998) Antibodies against desmoglein 3 (pemphigus vulgarisantigen) are present in sera from patients with paraneoplasticpemphigus and cause acantholysis in vivo in neonatal mice. J ClinInvest 102:775–782
2. Futei Y, Amagai M, Hashimoto T, Nishikawa T (2003) Confor-mational epitope mapping and IgG subclass distribution ofdesmoglein 3 in paraneoplastic pemphigus. J Am Acad Dermatol49:1023–1028
3. Lehmann PV, Forsthuber T, Miller A, Sercarz EE (1992)Spreading of T-cell autoimmunity to cryptic determinants of anautoantigen. Nature 358:155–157
4. Bowen GM et al (2000) Lichenoid dermatitis in paraneoplasticpemphigus: a pathogenic trigger of epitope spreading? ArchDermatol 136:652–656
5. Wang L, Bu D, Yang Y, Chen X, Zhu X (2004) Castleman’stumours and production of autoantibody in paraneoplasticpemphigus. Lancet 363:525–531
6. Anhalt GJ et al (1990) Paraneoplastic pemphigus. An autoimmunemucocutaneous disease associated with neoplasia. N Engl J Med323:1729–1735
7. Sehgal VN, Srivastava G (2009) Paraneoplastic pemphigus/para-neoplastic autoimmune multiorgan syndrome. Int J Dermatol48:162–169
8. Ehst BD, Minzer-Conzetti K, Swerdlin A, Devere TS (2010)Cutaneous manifestations of internal malignancy. Curr Probl Surg47:384–445
9. Martinez De Pablo MI et al (2005) Paraneoplastic pemphigusassociated with non-Hodgkin B-cell lymphoma and good responseto prednisone. Acta Derm Venereol 85:233–235
10. Mimouni D et al (2002) Paraneoplastic pemphigus in children andadolescents. Br J Dermatol 147:725–732
11. Billet SE, Grando SA, Pittelkow MR (2006) Paraneoplasticautoimmune multiorgan syndrome: review of the literature andsupport for a cytotoxic role in pathogenesis. Autoimmunity39:617–630
142 Clinic Rev Allerg Immunol (2012) 42:135–144

12. Buck T, Gonzalez LM, Lambert WC, Schwartz RA (2008)Sweet’s syndrome with hematologic disorders: a review andreappraisal. Int J Dermatol 47:775–782
13. Zappasodi P, Forno C, Corso A, Lazzarino M (2006) Mucocuta-neous paraneoplastic syndromes in hematologic malignancies. IntJ Dermatol 45:14–22
14. Sands J, Tuscano JM (2010) Geoepidemiology and autoimmunemanifestations of lymphoproliferative disorders. Autoimmun Rev9:A335–A341
15. Perry HO, Brunsting LA (1957) Pyoderma gangrenosum; aclinical study of nineteen cases. AMA Arch Derm 75:380–386
16. Bernstein CN, Blanchard JF, Rawsthorne P, Yu N (2001) Theprevalence of extraintestinal diseases in inflammatory bowel disease:a population-based study. Am J Gastroenterol 96:1116–1122
17. Tan MH, Gordon M, Lebwohl O, George J, Lebwohl MG (2001)Improvement of Pyoderma gangrenosum and psoriasis associatedwith Crohn disease with anti-tumor necrosis factor alphamonoclonal antibody. Arch Dermatol 137:930–933
18. Veloso FT, Carvalho J, Magro F (1996) Immune-related systemicmanifestations of inflammatory bowel disease. A prospectivestudy of 792 patients. J Clin Gastroenterol 23:29–34
19. Brunsting LA, Goeckerman WH, O’Leary PA (1930) Pyoderma(echthyma) gangrenosum—clinical and experimental observationsin five cases occurring in adults. Arch Dermatol Syph 22:655–680
20. Bennett ML et al (2000) Pyoderma gangrenosum. A comparisonof typical and atypical forms with an emphasis on time toremission. Case review of 86 patients from 2 institutions.Medicine (Baltimore) 79:37–46
21. Duguid CM, O’Loughlin S, Otridge B, Powell FC (1993) Paraneo-plastic pyoderma gangrenosum. Australas J Dermatol 34:17–22
22. Oka M et al (2000) Interleukin-8 overexpression is present inpyoderma gangrenosum ulcers and leads to ulcer formation inhuman skin xenografts. Lab Invest 80:595–604
23. Oka M (2007) Pyoderma gangrenosum and interleukin 8. Br JDermatol 157:1279–1281
24. TanakaN, FujiokaA, Tajima S, Ishibashi A, Hirose S (2000) Elafin isinduced in epidermis in skin disorders with dermal neutrophilicinfiltration: interleukin-1 beta and tumour necrosis factor-alphastimulate its secretion in vitro. Br J Dermatol 143:728–732
25. Saito S et al (2006) CD30+ anaplastic large cell lymphomacomplicated by pyoderma gangrenosum with increased levels ofserum cytokines. Eur J Haematol 77:251–254
26. Kawakami T, Yamazaki M, Soma Y (2009) Reduction ofinterleukin-6, interleukin-8, and anti-phosphatidylserine-prothrombin complex antibody by granulocyte and monocyteadsorption apheresis in a patient with pyoderma gangrenosum andulcerative colitis. Am J Gastroenterol 104:2363–2364
27. Bister V, Makitalo L, Jeskanen L, Saarialho-Kere U (2007)Expression of MMP-9, MMP-10 and TNF-alpha and lack ofepithelial MMP-1 and MMP-26 characterize pyoderma gangreno-sum. J Cutan Pathol 34:889–898
28. Brooklyn TN et al (2006) Infliximab for the treatment ofpyoderma gangrenosum: a randomised, double blind, placebocontrolled trial. Gut 55:505–509
29. Mittal S, Milner BJ, Vickers MA (2005) Pyoderma gangrenosumas a cause of splenomegaly and association with a T-cell clone.Clin Lab Haematol 27:402–404
30. Brooklyn TN, Williams AM, Dunnill MG, Probert CS (2007) T-cell receptor repertoire in pyoderma gangrenosum: evidence forclonal expansions and trafficking. Br J Dermatol 157:960–966
31. Su WP, Schroeter AL, Perry HO, Powell FC (1986) Histopatho-logic and immunopathologic study of pyoderma gangrenosum. JCutan Pathol 13:323–330
32. Gapin L et al (1998) Quantitative analysis of the T cell repertoireselected by a single peptide-major histocompatibility complex. JExp Med 187:1871–1883
33. Maverakis E et al (2003) Autoreactive T cells can be protectedfrom tolerance induction through competition by flanking determi-nants for access to class IIMHC. ProcNatl Acad Sci USA 100:5342–5347
34. Sercarz EE, Maverakis E (2003) Mhc-guided processing: bindingof large antigen fragments. Nat Rev Immunol 3:621–629
35. Maverakis E, Beech JT, Schneider S, Sercarz EE (2008)Presentation of a determinant by MHC class II can be preventedthrough competitive capture by a flanking determinant on amultideterminant peptide. J Autoimmun 31:59–65
36. Gammon G, Sercarz E (1989) How some T cells escape toleranceinduction. Nature 342:183–185
37. Engels EA, Pfeiffer RM (2003) Malignant thymoma in the UnitedStates: demographic patterns in incidence and associations withsubsequent malignancies. Int J Cancer 105:546–551
38. Melms A et al (2006) Thymus and myasthenia gravis: antigenprocessing in the human thymus and the consequences for thegeneration of autoreactive T cells. Acta Neurol Scand Suppl183:12–13
39. Wadhera A et al (2007) Thymoma-associated multiorgan autoim-munity: A graft-versus-host-like disease. J Am Acad Dermatol 57(4):683–689
40. Nagvekar N et al (1998) A pathogenetic role for the thymoma inmyasthenia gravis. Autosensitization of IL-4- producing T cellclones recognizing extracellular acetylcholine receptor epitopespresented by minority class II isotypes. J Clin Invest 101:2268–2277
41. Moschella SL (1967) Pyoderma gangrenosum. A patient success-fully treated with intralesional injections of steroid. ArchDermatol 95:121–123
42. Jennings JL (1983) Pyoderma gangrenosum: successful treatmentwith intralesional steroids. J Am Acad Dermatol 9:575–580
43. Mrowietz U, Christophers E (1991) Clearing of pyodermagangrenosum by intralesional cyclosporin A. Br J Dermatol125:499
44. Rosenow EC 3rd, Hurley BT (1984) Disorders of the thymus. Areview. Arch Intern Med 144:763–770
45. Lara PN Jr (2000) Malignant thymoma: current status and futuredirections. Cancer Treat Rev 26:127–131
46. Good RA (1954) Agammaglobulinaemia: a provocative experi-ment of nature. Bull Univ Minn Hosp 26:1–19
47. Fujimoto W et al (2002) Paraneoplastic pemphigus associatedwith Castleman’s disease and asymptomatic bronchiolitis obliter-ans. Eur J Dermatol 12:355–359
48. Wang J et al (2005) Autoantibody production from a thymomaand a follicular dendritic cell sarcoma associated with paraneo-plastic pemphigus. Br J Dermatol 153:558–564
49. Wang J et al (2005) Paraneoplastic pemphigus associated withCastleman tumor: a commonly reported subtype of paraneoplasticpemphigus in China. Arch Dermatol 141:1285–1293
50. Nikolskaia OV, Nousari CH, Anhalt GJ (2003) Paraneoplasticpemphigus in association with Castleman’s disease. Br J Dermatol149:1143–1151
51. KimWY, Kim H, Jeon YK, Kim CW (2010) Follicular dendritic cellsarcoma with immature T-cell proliferation. Hum Pathol 41:129–133
52. Harding FA, McArthur JG, Gross JA, Raulet DH, Allison JP (1992)CD28-mediated signalling co-stimulates murine T cells and preventsinduction of anergy in T-cell clones. Nature 356:607–609
53. Thompson ED, Enriquez HL, Fu YX, Engelhard VH (2010)Tumor masses support naive T cell infiltration, activation, anddifferentiation into effectors. J Exp Med 207:1791–1804
54. Hamblin TJ (2006) Autoimmune complications of chroniclymphocytic leukemia. Semin Oncol 33:230–239
55. Dearden C (2008) Disease-specific complications of chroniclymphocytic leukemia. Hematology Am Soc Hematol EducProgram, pp 450–456
Clinic Rev Allerg Immunol (2012) 42:135–144 143

56. Tomic J et al (2006) Sensitization of IL-2 signaling through TLR-7 enhances B lymphoma cell immunogenicity. J Immunol176:3830–3839
57. Jablonska E et al (2005) TNF family molecules in the serum ofpatients with B-cell chronic lymphocytic leukemia (B-CLL). LeukLymphoma 46:1307–1312
58. Hall AM, Vickers MA, McLeod E, Barker RN (2005) Rhautoantigen presentation to helper T cells in chronic lymphocyticleukemia by malignant B cells. Blood 105:2007–2015
59. Gorgun G, Holderried TA, Zahrieh D, Neuberg D, Gribben JG(2005) Chronic lymphocytic leukemia cells induce changes ingene expression of CD4 and CD8 T cells. J Clin Invest 115:1797–1805
60. Ramsay AG et al (2008) Chronic lymphocytic leukemia Tcells showimpaired immunological synapse formation that can be reversed withan immunomodulating drug. J Clin Invest 118:2427–2437
61. Abu-Shakra M, Buskila D, Ehrenfeld M, Conrad K, Shoenfeld Y(2001) Cancer and autoimmunity: autoimmune and rheumaticfeatures in patients with malignancies. Ann Rheum Dis 60:433–441
62. Cines DB, Bussel JB, Liebman HA, Luning Prak ET (2009) TheITP syndrome: pathogenic and clinical diversity. Blood 113:6511–6521
63. Beyer M et al (2005) Reduced frequencies and suppressivefunction of CD4+CD25hi regulatory T cells in patients withchronic lymphocytic leukemia after therapy with fludarabine.Blood 106:2018–2025
64. Dearden C et al (2008) The prognostic significance of a positivedirect antiglobulin test in chronic lymphocytic leukemia: a beneficialeffect of the combination of fludarabine and cyclophosphamide onthe incidence of hemolytic anemia. Blood 111:1820–1826
65. Citores MJ et al (2010) CD154 expression triggered by purineanalogues in vitro: correlation with treatment response andautoimmune events in chronic lymphocytic leukemia. ExpHematol 38:165–173
66. Gooptu C et al (2001) Paraneoplastic pemphigus: an associationwith fludarabine? Br J Dermatol 144:1255–1261
67. Hodi FS et al (2010) Improved survival with ipilimumab inpatients with metastatic melanoma. N Engl J Med 363:711–723
68. Di Bernardo MC et al (2008) A genome-wide association studyidentifies six susceptibility loci for chronic lymphocytic leukemia.Nat Genet 40:1204–1210
69. Shaffer AL et al (2008) IRF4 addiction in multiple myeloma.Nature 454:226–231
70. Biswas PS et al (2010) Phosphorylation of IRF4 by ROCK2regulates IL-17 and IL-21 production and the development ofautoimmunity in mice. J Clin Invest 120:3280–3295
71. Novak AJ et al (2009) Genetic variation in B-cell-activating factoris associated with an increased risk of developing B-cell non-Hodgkin lymphoma. Cancer Res 69:4217–4224
72. Juszczynski P et al (2002) Human leukocyte antigens class II andtumor necrosis factor genetic polymorphisms are independentpredictors of non-Hodgkin lymphoma outcome. Blood 100:3037–3040
73. Chu VT et al (2009) Systemic activation of the immune systeminduces aberrant BAFF and APRIL expression in B cells inpatients with systemic lupus erythematosus. Arthritis Rheum60:2083–2093
74. Zhang W et al (2010) B-cell activating factor and v-Mycmyelocytomatosis viral oncogene homolog (c-Myc) influenceprogression of chronic lymphocytic leukemia. Proc Natl AcadSci USA 107:18956–18960
75. Lang B, Vincent A (1996) Autoimmunity to ion-channels andother proteins in paraneoplastic disorders. Curr Opin Immunol8:865–871
76. Ehrenfeld M, Abu-Shakra M, Buskila D, Shoenfeld Y (2001) Thedual association between lymphoma and autoimmunity. BloodCell Mol Diseases 27:750–756
77. Dalmau J et al (2007) Paraneoplastic anti-N-methyl-D-aspartatereceptor encephalitis associated with ovarian teratoma. AnnNeurol 61:25–36
78. Butler MH et al (2000) Autoimmunity to gephyrin in Stiff–Mansyndrome. Neuron 26:307–312
79. Polans AS et al (1995) Recoverin, a photoreceptor-specificcalcium-binding protein, is expressed by the tumor of a patientwith cancer-associated retinopathy. Proc Natl Acad Sci USA92:9176–9180
80. Buckanovich RJ, Yang YYL, Darnell RB (1996) The onconeuralantigen Nova-1 is a neuron-specific RNA-Binding protein, theactivity of which is inhibited by paraneoplastic antibodies. JNeurosci 16:1114–1122
81. Lennon VA et al (1995) Calcium-channel antibodies in theLambert–Eaton syndrome and other paraneoplastic syndromes.N Engl J Med 332:1467–1474
82. Yu Z et al (2001) CRMP-5 neuronal autoantibody: marker of lungcancer and thymoma-related autoimmunity. Ann Neurol 49:146–154
144 Clinic Rev Allerg Immunol (2012) 42:135–144