thalamic development induced by shh in the chick embryo
TRANSCRIPT

www.elsevier.com/locate/ydbio
Developmental Biology
Thalamic development induced by Shh in the chick embryo
Claudia Vieiraa, Ana-Lila Gardab, Kenji Shimamurac, Salvador Martineza,*
aNeuroscience Institute, Miguel Hernandez University N-332, Km 87, E-03550 Alicante, SpainbCentro IVI-Murcia, Navegante Macias 5, E-30007 Murcia Spain
cDivision of Morphogenesis, Department of Embryogenesis, Institute of Molecular Embryology and Genetics, Kumamoto University,
Honjo 2-2-1, Kumamoto 860-0811, Japan
Received for publication 18 February 2005, revised 23 May 2005, accepted 24 May 2005
Available online 18 July 2005
Abstract
Patterning of the early neural tube is achieved in part by the inductive signals, which arise from neuroepithelial signaling centers. The
zona limitans intrathalamica (ZLI) is a neuroepithelial domain in the alar plate of the diencephalon which separates the prethalamus from the
thalamus. The ZLI has recently been considered to be a possible secondary organizer, effecting its inductions via sonic hedgehog (Shh), a
signaling molecule which drives morphogenetic information for the thalamus. Using experimental embryological techniques involving the
generation of chimeric embryos, we show that the formation of the ZLI in the diencephalic alar plate is due to an interaction between the
prechordal and epichordal plate neuroepithelia. We also provide evidence that Shh expression in the ZLI underlies the morphogenetic activity
of this putative diencephalic organizer. Ectopic Shh led to the auto-induction of its own gene expression in host cells, as well as to the
expression of other genes involved in diencephalic regionalization and histogenesis. Analysis of long-term surviving embryos after Shh
ectopic expression demonstrated that Shh was able to induce thalamic structures and local overgrowth. Overall, these results indicate that Shh
expressed in the ZLI plays an important role in diencephalic growth and in the development of the thalamus.
D 2005 Elsevier Inc. All rights reserved.
Keywords: Chick; Neural tube; Diencephalon; Zona limitans intrathalamica; Sonic hedgehog; Thalamus; Secondary organizer
Introduction
The diencephalon is a cerebral region which develops
from the anterior part of the neural tube, the prosencepha-
lon. This area becomes subdivided into the telencephalon
and the hypothalamus at the anterior pole, and the
diencephalon at the posterior zone. The diencephalon is
limited by two boundaries, the most anterior one that lies
between the pedunculomammillary region and the eminentia
thalami and the posterior one that lies between the pretectum
and the mesencephalon (Martinez and Puelles, 2000; Puelles
and Rubenstein, 2003; Garcia-Lopez et al., 2004). The
diencephalon develops into two major neuroepithelial
domains known as the prethalamus and the thalamus
(Puelles and Rubenstein, 2003).
0012-1606/$ - see front matter D 2005 Elsevier Inc. All rights reserved.
doi:10.1016/j.ydbio.2005.05.031
* Corresponding author. Fax: +34 965 919 555.
E-mail address: [email protected] (S. Martinez).
The zona limitans intrathalamica (ZLI) is a central
boundary in the diencephalon which appears as a transverse
ventricular ridge between the prethalamus and the thalamus at
the neural tube stages (Figs. 1A–F; Martinez and Puelles,
2000; Echevarria et al., 2003; Puelles and Rubenstein, 2003;
Garcia-Lopez et al., 2004). The ZLI corresponds to an inter-
segmental boundary in the neuroepithelium, in which neuro-
epithelial cells show clonal restriction and a low proliferation
rate (Fig. 1F; Puelles et al., 1987; Figdor and Stern, 1993;
Martinez and Puelles, 2000; Martinez et al., 1992; Larsen et
al., 2001; Echevarria et al., 2003). The complex pattern of
regulatory molecules expressed by ZLI cells in the neural
tube during development is indicative of its importance as a
possible secondary morphogenetic organizer in diencephalic
histogenesis (Fig. 1G; Echevarria et al., 2003).
The dynamic expression of Shh in the ZLI (Figs. 1B, C)
may be a consequence of the interaction between the
prechordal and epichordal plate, which first specifies the
284 (2005) 351 – 363
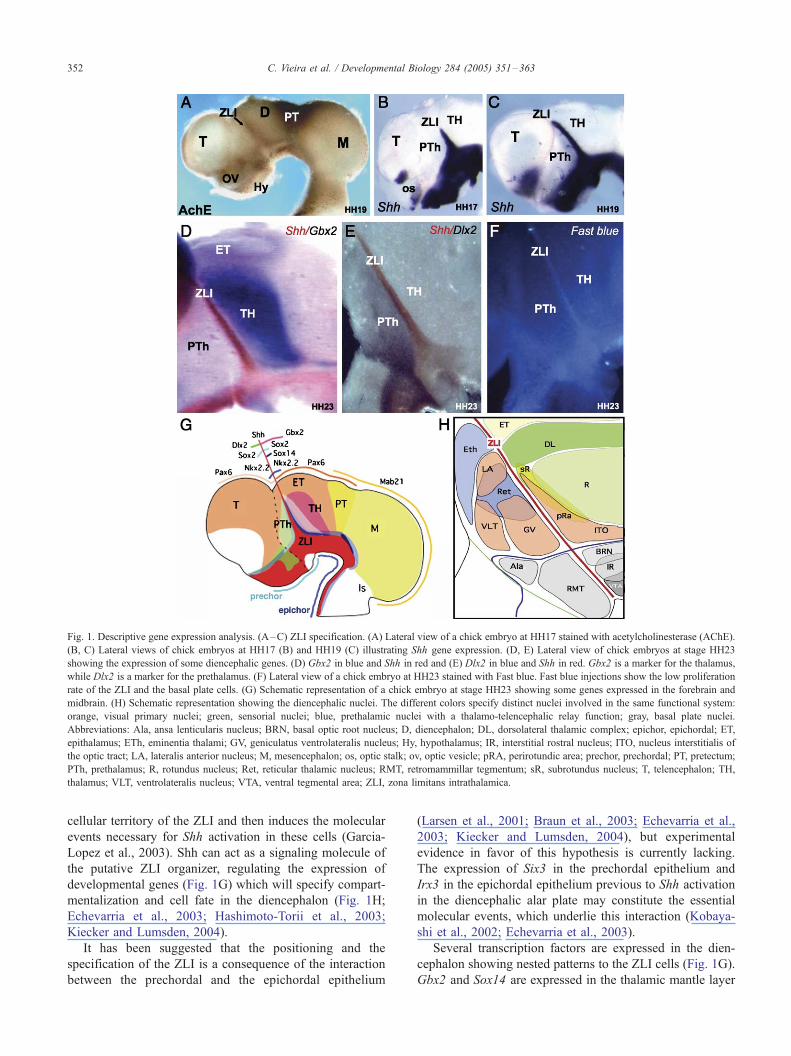
Fig. 1. Descriptive gene expression analysis. (A–C) ZLI specification. (A) Lateral view of a chick embryo at HH17 stained with acetylcholinesterase (AChE).
(B, C) Lateral views of chick embryos at HH17 (B) and HH19 (C) illustrating Shh gene expression. (D, E) Lateral view of chick embryos at stage HH23
showing the expression of some diencephalic genes. (D) Gbx2 in blue and Shh in red and (E) Dlx2 in blue and Shh in red. Gbx2 is a marker for the thalamus,
while Dlx2 is a marker for the prethalamus. (F) Lateral view of a chick embryo at HH23 stained with Fast blue. Fast blue injections show the low proliferation
rate of the ZLI and the basal plate cells. (G) Schematic representation of a chick embryo at stage HH23 showing some genes expressed in the forebrain and
midbrain. (H) Schematic representation showing the diencephalic nuclei. The different colors specify distinct nuclei involved in the same functional system:
orange, visual primary nuclei; green, sensorial nuclei; blue, prethalamic nuclei with a thalamo-telencephalic relay function; gray, basal plate nuclei.
Abbreviations: Ala, ansa lenticularis nucleus; BRN, basal optic root nucleus; D, diencephalon; DL, dorsolateral thalamic complex; epichor, epichordal; ET,
epithalamus; ETh, eminentia thalami; GV, geniculatus ventrolateralis nucleus; Hy, hypothalamus; IR, interstitial rostral nucleus; ITO, nucleus interstitialis of
the optic tract; LA, lateralis anterior nucleus; M, mesencephalon; os, optic stalk; ov, optic vesicle; pRA, perirotundic area; prechor, prechordal; PT, pretectum;
PTh, prethalamus; R, rotundus nucleus; Ret, reticular thalamic nucleus; RMT, retromammillar tegmentum; sR, subrotundus nucleus; T, telencephalon; TH,
thalamus; VLT, ventrolateralis nucleus; VTA, ventral tegmental area; ZLI, zona limitans intrathalamica.
C. Vieira et al. / Developmental Biology 284 (2005) 351–363352
cellular territory of the ZLI and then induces the molecular
events necessary for Shh activation in these cells (Garcia-
Lopez et al., 2003). Shh can act as a signaling molecule of
the putative ZLI organizer, regulating the expression of
developmental genes (Fig. 1G) which will specify compart-
mentalization and cell fate in the diencephalon (Fig. 1H;
Echevarria et al., 2003; Hashimoto-Torii et al., 2003;
Kiecker and Lumsden, 2004).
It has been suggested that the positioning and the
specification of the ZLI is a consequence of the interaction
between the prechordal and the epichordal epithelium
(Larsen et al., 2001; Braun et al., 2003; Echevarria et al.,
2003; Kiecker and Lumsden, 2004), but experimental
evidence in favor of this hypothesis is currently lacking.
The expression of Six3 in the prechordal epithelium and
Irx3 in the epichordal epithelium previous to Shh activation
in the diencephalic alar plate may constitute the essential
molecular events, which underlie this interaction (Kobaya-
shi et al., 2002; Echevarria et al., 2003).
Several transcription factors are expressed in the dien-
cephalon showing nested patterns to the ZLI cells (Fig. 1G).
Gbx2 and Sox14 are expressed in the thalamic mantle layer

C. Vieira et al. / Developmental Biology 284 (2005) 351–363 353
(Bulfone et al., 1993; Martinez-de-la-Torre et al., 2002;
Hashimoto-Torii et al., 2003), while Dlx2 is expressed in the
prethalamic mantle layer (Bulfone et al., 1993, Larsen et al.,
2001; Gonzalez et al., 2002). Nkx2.2 is expressed in the
neural epithelium flanking the expression of Shh in the ZLI
(Price et al., 1992, Shimamura et al., 1995).
Shh expression is a marker for the ZLI. Its expression
starts in the ventral edge of the ZLI, and then extends to all
the surface of the ZLI along the diencephalic alar plate
(Figs. 1B, C). In addition to the regionalization properties of
the ZLI, it has been postulated that Shh signaling plays a
role in neuroepithelial proliferation and thalamic growth; the
reduced size of the diencephalon in Shh mutants corrobo-
rates this hypothesis (Chiang et al., 1996). Thus, Shh
expression confers to the ZLI morphogenetic properties
which, as in the spinal cord, regulate the expression of
distinct sets of transcription factors necessary for the
sequential specification of neural progenitors in local
domains of the basal plate (Ericson et al., 1997; Briscoe et
al., 2000; for review see Ingham and McMahon, 2001;
Marti and Bovolenta, 2002). Evidence for the morphoge-
netic activity of Shh in the mesencephalic basal plate has
been reported by Agarwala’s group (Agarwala et al., 2001).
Indeed, Shh has been shown to act as a morphogen in the
spinal cord by inducing various genes in a concentration-
dependent manner (Roelink et al., 1995; Ericson et al.,
1997). Shh has also been proposed to act as a mitogen in the
expansion of cerebellar granule precursors (Dahmane and
Ruiz-i-Altaba, 1999; Wallace, 1999; Wechsler-Reya and
Scott, 1999; Oliver et al., 2005).
In the present paper, we demonstrate that the ZLI
develops in the neuroepithelium where the prechordal and
epichordal neural plates contact. We also show that Shh
expression is activated in ZLI cells and controls the
activation of several developmental genes, including itself,
involved in diencephalic regionalization and thalamic cell
fate. Ectopic expression of physiological levels of Shh
transforms mesencephalic and epithalamic regions into
organized thalamic structures. Finally, we provide evidence
that Shh signaling increases the growth of the thalamus.
Materials and methods
Microsurgery
Fertilized chick (Gallus gallus) and quail (Coturnix
coturnix japonica) eggs were incubated at 37-C in a forced
air incubator and staged according to Hamburger and
Hamilton (1951).
Heterotopic and isochronic grafts were performed using
quail or BrdU-labeled chick embryos as the source of the
prechordal neural epithelium to be implanted in the
epichordal epithelium of the host chick. Embryos were
operated at stage HH9 for chick and the equivalent stage for
quail embryos (7 somites). After microsurgery, the eggs
were sealed and kept in the incubator until HH23 (4 days of
incubation) or HH36 (10 days of incubation).
Cell aggregates of QT6-control quail fibroblasts and QT6
SHH-expressing cells (kindly provided by D. Duprez) were
implanted into the right side of the neural tube, after careful
opening of the neuroepithelium where cell aggregates were
inserted, at different positions in HH10–11 chicken
embryos. Cell aggregate preparation was performed in
accordance with Duprez et al. (Duprez et al., 1998).
Embryos were then left to develop until HH23 or HH36
and then fixed overnight in 4% paraformaldehyde in
phosphate-buffered saline solution (PBS 0.1 M, pH 7.4).
In situ hybridization
After fixation, embryos were rinsed in PBT (PBS with
0.1% Tween 20), dehydrated through an ascending meth-
anol series and stored in 100% methanol at �20-C before
being processed for in situ hybridization (ISH) as whole-
mount (HH23) or vibratome sections (HH36), as described
by Nieto et al. (Nieto et al., 1996). HH36 embryos were
embedded in 4% agarose in PBS and sectioned transversally
at 100 Am using a vibratome (Leica).
Digoxigenin and fluorescein-labeled RNA probes were
prepared from plasmids kindly provided by: J.L.R. Ruben-
stein (Dlx2, Nkx2.2, Shh, Pax6, Six3), A. Simeone (Gbx2),
G.G. Gonsalez (Mab21), G.R. Martin (Fgf8), A. McMahon
(Wnt8b), and M. Bronner-Fraser (Wnt3a). RNA-labeled
probes were detected by alkaline-phosphatase-coupled anti-
digoxigenin and anti-fluorescein antibodies (Roche Diag-
nostics, Mannheim, Germany) and NBT/BCIP was used as
a chromogenic substrate to detect the digoxigenin-labeled
probes (Boehringer, Mannheim, Germany), while INT/BCIP
was used for the detection of the fluorescein-labeled probes
(Roche Diagnostics, Mannheim, Germany). After ISH,
embryos were washed in PBT, photographed under a
dissecting microscope (Leica), and stored at 4-C in PBT
with 0.1% sodium azide. Vibratome sections were mounted
on glass slides, counterstained with neutral red and
dehydrated to absolute alcohol, cleared in xylene, and
mounted in Eukitt (O. Kindler GmbH and CO, Freiburg).
Histological analysis
Quail-chick chimeras were immunostained with a mono-
clonal anti-quail antibody (QCPN, Developmental Hybrid-
oma Bank, Iowa City, IA) like in Cobos et al. (2001).
Whole-mount embryos at HH19 were stained with
acetylcholinesterase (Martinez-de-la-Torre et al., 1990), and
embryos at HH23 were processed for Fast blue staining
dilution (Martinez et al., 1992).
BrdU staining was performed according to McConnell
and Kaznowski (1991).
Cryostat sections of short-term survival embryos with
SHH-expressing cells were counterstained with bisbenzi-
mide (Hoechst 33258, Sigma). Some chicken embryos at

C. Vieira et al. / Developmental Biology 284 (2005) 351–363354
HH36 were fixed overnight in Clarke’s fixative, dehydrated,
embedded in paraffin, and sectioned sagittally at 12 Am.
These sections were stained with cresyl violet. Subse-
quently, all sections were dehydrated to absolute alcohol,
cleared in xylene and mounted in Eukitt (O. Kindler GmbH
and CO, Freiburg), for further analysis and photography
under a dissecting microscope (Leica).
Results
Specification of the ZLI in the prosencephalic
neuroepithelium
The mechanisms which underlie the positioning of the
ZLI in the prosencephalic neuroepithelium are currently
unknown. It has been hypothesized that the ZLI develops in
the contacting zone between the prechordal and epichordal
neuroepithelium (Fig. 1; Larsen et al., 2001; Braun et al.,
2003; Echevarria et al., 2003; Hashimoto-Torii et al., 2003).
To explore this hypothesis, we grafted quail or BrdU-labeled
chick prechordal prosencephalic epithelium, exactly the
anlage of the dorsal pallium, into epichordal domains,
posterior diencephalon and mesencephalon of isochronic
chick embryos (Fig. 2). The grafts were excised with special
care with the help of our previously developed prosen-
cephalic fate maps, so as not to include epithelium which
expresses Fgf8 or Shh (Cobos et al., 2001; Garcia-Lopez et
al., 2004). As in our fate map experiments, we inserted a
grid with concentric circles into one ocular of the micro-
scope used to perform the grafts, so as to ensure the
exclusion of neuroepithelium expressing Fgf8 or Shh, as
well as to normalize the size and exact location of the graft
between different experiments. Of the 233 prechordal
transplants into the posterior diencephalon (n = 190) or
mesencephalon (n = 43), 152 cases were found to be
adequately integrated (120 in the diencephalon and 32 in the
mesencephalon). These cases were processed to study the
induction or repression of genes in the interface graft/host,
and in the grafted and host epithelial domains.
First, we analyzed the induction of Shh expression in the
contacting region 2 days after grafting. We observed that of
the 90 experimental cases, Shh expression was activated in
52 (Figs. 2A–K). Ectopically activated Shh was localized in
the graft/host interface, around the grafted epithelium,
contacting in most cases with the normal expression domain
of Shh in the basal plate (Figs. 2A–J). Anti-quail or anti-
BrdU immunostaining to detect donor cells showed that Shh
was induced in both donor and host interface cells (Fig. 2D).
The epichordal epithelium competent to express Shh was
mapped from the thalamus to the anterior rhombencephalic
region (isthmus-rhombomer1). On no occasion was Shh
expression induced in caudal graft/host interfaces localized
between r2 and r5 (Fig. 2K; n = 10). Some grafts of
epichordal into prechordal epithelium (n = 20) showed very
low integration and important structural anomalies, such as
dorsal midline opening, strong malformation, and reduction
of telencephalic derivatives. These cases were considered
inappropriate for analysis and interpretation.
It has been demonstrated that the expression of Fgf8 in
the isthmic organizer (IsO) is controlled by the interaction
of Otx2 and Gbx2 (Hidalgo-Sanchez et al., 1999; Joyner et
al., 2000; Garda et al., 2001; Martinez-Barbera et al.,
2001). Our next objective was to explore if the two kinds
of interactions, prechordal/epichordal and Otx2/Gbx2, can
act in parallel to regulate differential gene expression in
the contacting neuroepithelium. To this end, we grafted
prechordal-Otx2+ epithelium (Crossley et al., 2001) into
the posterior pole of the mesencephalic vesicle (epichor-
dal), including the isthmic constriction, a domain where
Gbx2 is strongly expressed (Garda et al., 2001). In seven
of ten cases, the graft perfectly integrated into the caudal
mesencephalon and the isthmic region. Neuroepithelial
expression of Shh and Fgf8 was detected by whole-mount
ISH in the chimeric neural tubes 48 h after grafting (Figs.
2I, J). We observed a clear induction of both genes in the
cells at the caudal graft/host interface, where the two
regulative effects have been established: prechordal–
epichordal interaction activated Shh in an anterior band
of cells and Otx2/Gbx2 interaction activated Fgf8 in a
posterior band of cells (Figs. 2I, J). When the interface
between donor and host epithelia developed as a normal or
ectopic choroidal structure, Shh induction in this interface
was not observed (Figs. 2C, G, and I).
Ectopic Shh induces molecular reorganization in the donor
and host epithelial grafts
Chimeric embryos presenting ectopic induction of Shh
were analyzed to explore additional modifications in the
molecular patterns of the grafted and host epithelium. In ‘‘ex
vivo’’ experiments and in electroporation assays, Shh has
been shown to modulate genetic expression at both sides of
the ZLI (Braun et al., 2003; Hashimoto-Torii et al., 2003;
Kiecker and Lumsden, 2004). However, the possible
inductive activity of Shh as a signaling molecule of the
ZLI has not yet been explored either at physiological levels
or in experiments in vivo. Thus, the induced expression of
Shh at the graft/host interface is a useful experimental model
to analyze effects of ectopic Shh in the diencephalic and
mesencephalic neuroepithelium.
In two cases, the graft included caudal prechordal
neuroepithelium, thus carrying some Fgf8, which caused
the growth of the grafted epithelium. These grafts developed
as a vesicle localized in the thalamus. This vesicle showed
an integration zone encircled by the expression of Shh. The
rostral component of this circle corresponded to the ZLI,
while ventrally the interface contacted with the basal plate
expression of Shh, and caudally a new Shh positive stripe
invaded the alar plate from the basal plate (Figs. 2F, G). The
normal antero-posterior pattern of expression of Pax6, i.e.,
positive in the thalamus and pretectum and negative in the
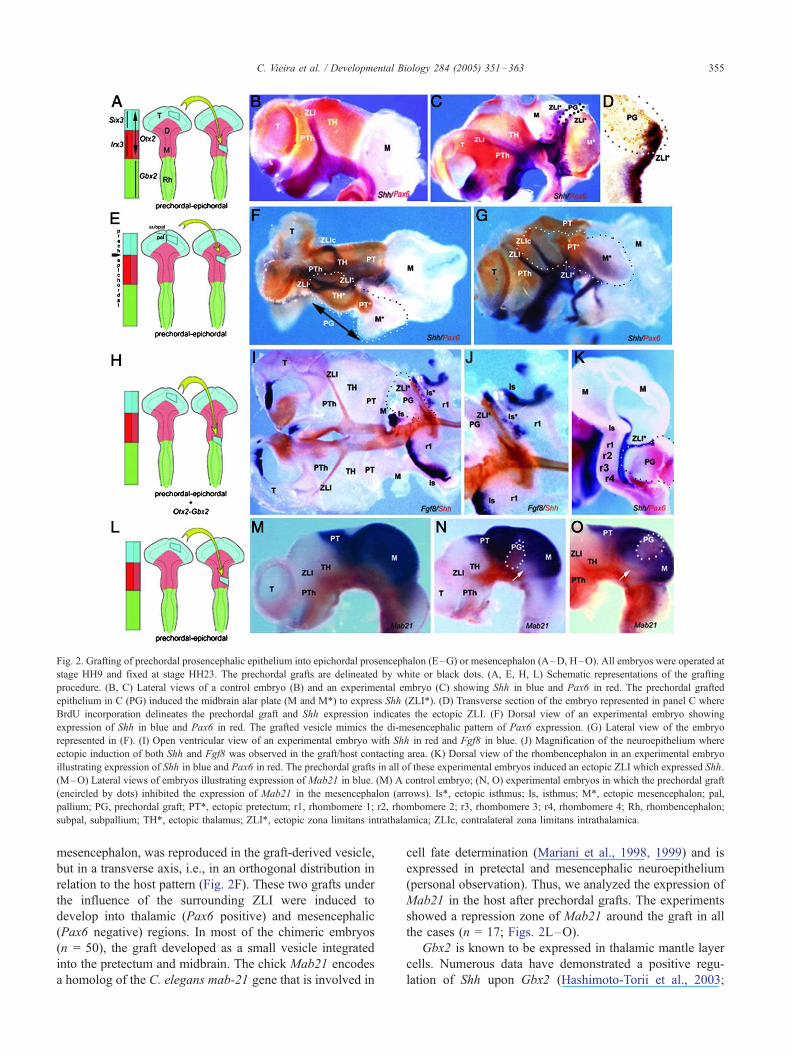
Fig. 2. Grafting of prechordal prosencephalic epithelium into epichordal prosencephalon (E–G) or mesencephalon (A–D, H–O). All embryos were operated at
stage HH9 and fixed at stage HH23. The prechordal grafts are delineated by white or black dots. (A, E, H, L) Schematic representations of the grafting
procedure. (B, C) Lateral views of a control embryo (B) and an experimental embryo (C) showing Shh in blue and Pax6 in red. The prechordal grafted
epithelium in C (PG) induced the midbrain alar plate (M and M*) to express Shh (ZLI*). (D) Transverse section of the embryo represented in panel C where
BrdU incorporation delineates the prechordal graft and Shh expression indicates the ectopic ZLI. (F) Dorsal view of an experimental embryo showing
expression of Shh in blue and Pax6 in red. The grafted vesicle mimics the di-mesencephalic pattern of Pax6 expression. (G) Lateral view of the embryo
represented in (F). (I) Open ventricular view of an experimental embryo with Shh in red and Fgf8 in blue. (J) Magnification of the neuroepithelium where
ectopic induction of both Shh and Fgf8 was observed in the graft/host contacting area. (K) Dorsal view of the rhombencephalon in an experimental embryo
illustrating expression of Shh in blue and Pax6 in red. The prechordal grafts in all of these experimental embryos induced an ectopic ZLI which expressed Shh.
(M–O) Lateral views of embryos illustrating expression of Mab21 in blue. (M) A control embryo; (N, O) experimental embryos in which the prechordal graft
(encircled by dots) inhibited the expression of Mab21 in the mesencephalon (arrows). Is*, ectopic isthmus; Is, isthmus; M*, ectopic mesencephalon; pal,
pallium; PG, prechordal graft; PT*, ectopic pretectum; r1, rhombomere 1; r2, rhombomere 2; r3, rhombomere 3; r4, rhombomere 4; Rh, rhombencephalon;
subpal, subpallium; TH*, ectopic thalamus; ZLI*, ectopic zona limitans intrathalamica; ZLIc, contralateral zona limitans intrathalamica.
C. Vieira et al. / Developmental Biology 284 (2005) 351–363 355
mesencephalon, was reproduced in the graft-derived vesicle,
but in a transverse axis, i.e., in an orthogonal distribution in
relation to the host pattern (Fig. 2F). These two grafts under
the influence of the surrounding ZLI were induced to
develop into thalamic (Pax6 positive) and mesencephalic
(Pax6 negative) regions. In most of the chimeric embryos
(n = 50), the graft developed as a small vesicle integrated
into the pretectum and midbrain. The chick Mab21 encodes
a homolog of the C. elegans mab-21 gene that is involved in
cell fate determination (Mariani et al., 1998, 1999) and is
expressed in pretectal and mesencephalic neuroepithelium
(personal observation). Thus, we analyzed the expression of
Mab21 in the host after prechordal grafts. The experiments
showed a repression zone of Mab21 around the graft in all
the cases (n = 17; Figs. 2L–O).
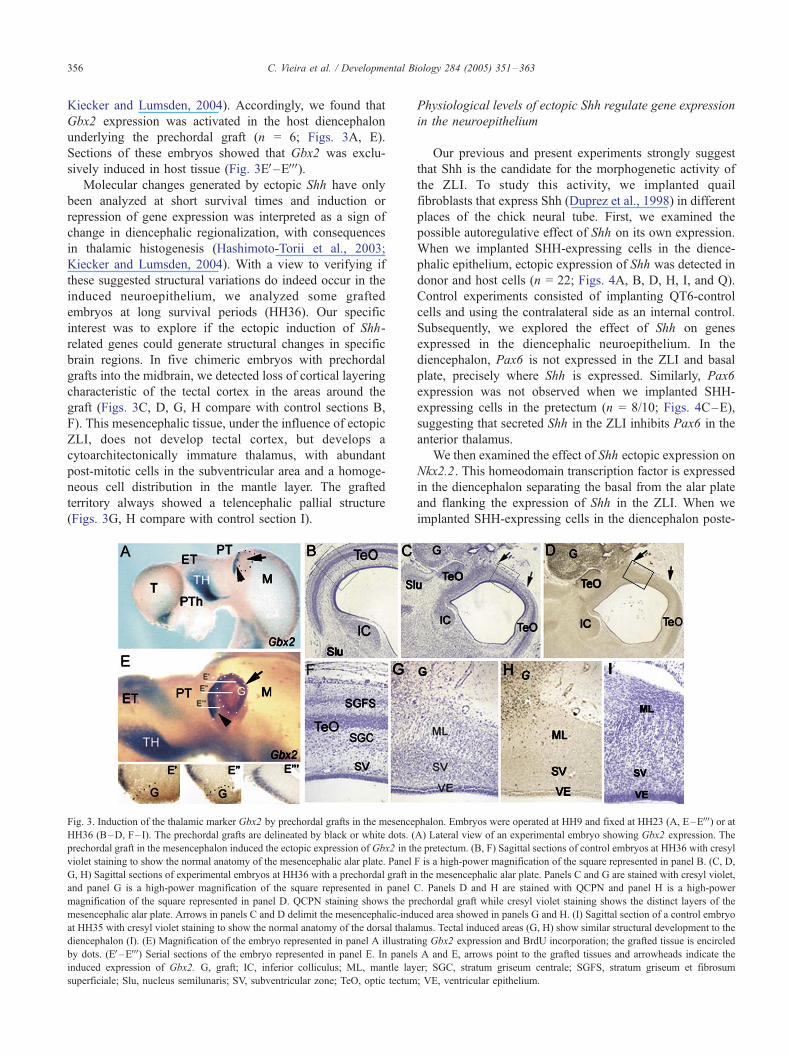
Gbx2 is known to be expressed in thalamic mantle layer
cells. Numerous data have demonstrated a positive regu-
lation of Shh upon Gbx2 (Hashimoto-Torii et al., 2003;
C. Vieira et al. / Developmental Biology 284 (2005) 351–363356
Kiecker and Lumsden, 2004). Accordingly, we found that
Gbx2 expression was activated in the host diencephalon
underlying the prechordal graft (n = 6; Figs. 3A, E).
Sections of these embryos showed that Gbx2 was exclu-
sively induced in host tissue (Fig. 3EV–EVVV).Molecular changes generated by ectopic Shh have only
been analyzed at short survival times and induction or
repression of gene expression was interpreted as a sign of
change in diencephalic regionalization, with consequences
in thalamic histogenesis (Hashimoto-Torii et al., 2003;
Kiecker and Lumsden, 2004). With a view to verifying if
these suggested structural variations do indeed occur in the
induced neuroepithelium, we analyzed some grafted
embryos at long survival periods (HH36). Our specific
interest was to explore if the ectopic induction of Shh-
related genes could generate structural changes in specific
brain regions. In five chimeric embryos with prechordal
grafts into the midbrain, we detected loss of cortical layering
characteristic of the tectal cortex in the areas around the
graft (Figs. 3C, D, G, H compare with control sections B,
F). This mesencephalic tissue, under the influence of ectopic
ZLI, does not develop tectal cortex, but develops a
cytoarchitectonically immature thalamus, with abundant
post-mitotic cells in the subventricular area and a homoge-
neous cell distribution in the mantle layer. The grafted
territory always showed a telencephalic pallial structure
(Figs. 3G, H compare with control section I).
Fig. 3. Induction of the thalamic marker Gbx2 by prechordal grafts in the mesence
HH36 (B–D, F– I). The prechordal grafts are delineated by black or white dots. (
prechordal graft in the mesencephalon induced the ectopic expression of Gbx2 in th
violet staining to show the normal anatomy of the mesencephalic alar plate. Panel
G, H) Sagittal sections of experimental embryos at HH36 with a prechordal graft in
and panel G is a high-power magnification of the square represented in panel
magnification of the square represented in panel D. QCPN staining shows the p
mesencephalic alar plate. Arrows in panels C and D delimit the mesencephalic-ind
at HH35 with cresyl violet staining to show the normal anatomy of the dorsal thala
diencephalon (I). (E) Magnification of the embryo represented in panel A illustrat
by dots. (EV–EVVV) Serial sections of the embryo represented in panel E. In panels
induced expression of Gbx2. G, graft; IC, inferior colliculus; ML, mantle lay
superficiale; Slu, nucleus semilunaris; SV, subventricular zone; TeO, optic tectum
Physiological levels of ectopic Shh regulate gene expression
in the neuroepithelium
Our previous and present experiments strongly suggest
that Shh is the candidate for the morphogenetic activity of
the ZLI. To study this activity, we implanted quail
fibroblasts that express Shh (Duprez et al., 1998) in different
places of the chick neural tube. First, we examined the
possible autoregulative effect of Shh on its own expression.
When we implanted SHH-expressing cells in the dience-
phalic epithelium, ectopic expression of Shh was detected in
donor and host cells (n = 22; Figs. 4A, B, D, H, I, and Q).
Control experiments consisted of implanting QT6-control
cells and using the contralateral side as an internal control.
Subsequently, we explored the effect of Shh on genes
expressed in the diencephalic neuroepithelium. In the
diencephalon, Pax6 is not expressed in the ZLI and basal
plate, precisely where Shh is expressed. Similarly, Pax6
expression was not observed when we implanted SHH-
expressing cells in the pretectum (n = 8/10; Figs. 4C–E),
suggesting that secreted Shh in the ZLI inhibits Pax6 in the
anterior thalamus.
We then examined the effect of Shh ectopic expression on
Nkx2.2. This homeodomain transcription factor is expressed
in the diencephalon separating the basal from the alar plate
and flanking the expression of Shh in the ZLI. When we
implanted SHH-expressing cells in the diencephalon poste-
phalon. Embryos were operated at HH9 and fixed at HH23 (A, E–EVVV) or atA) Lateral view of an experimental embryo showing Gbx2 expression. The
e pretectum. (B, F) Sagittal sections of control embryos at HH36 with cresyl
F is a high-power magnification of the square represented in panel B. (C, D,
the mesencephalic alar plate. Panels C and G are stained with cresyl violet,
C. Panels D and H are stained with QCPN and panel H is a high-power
rechordal graft while cresyl violet staining shows the distinct layers of the
uced area showed in panels G and H. (I) Sagittal section of a control embryo
mus. Tectal induced areas (G, H) show similar structural development to the
ing Gbx2 expression and BrdU incorporation; the grafted tissue is encircled
A and E, arrows point to the grafted tissues and arrowheads indicate the
er; SGC, stratum griseum centrale; SGFS, stratum griseum et fibrosum
; VE, ventricular epithelium.
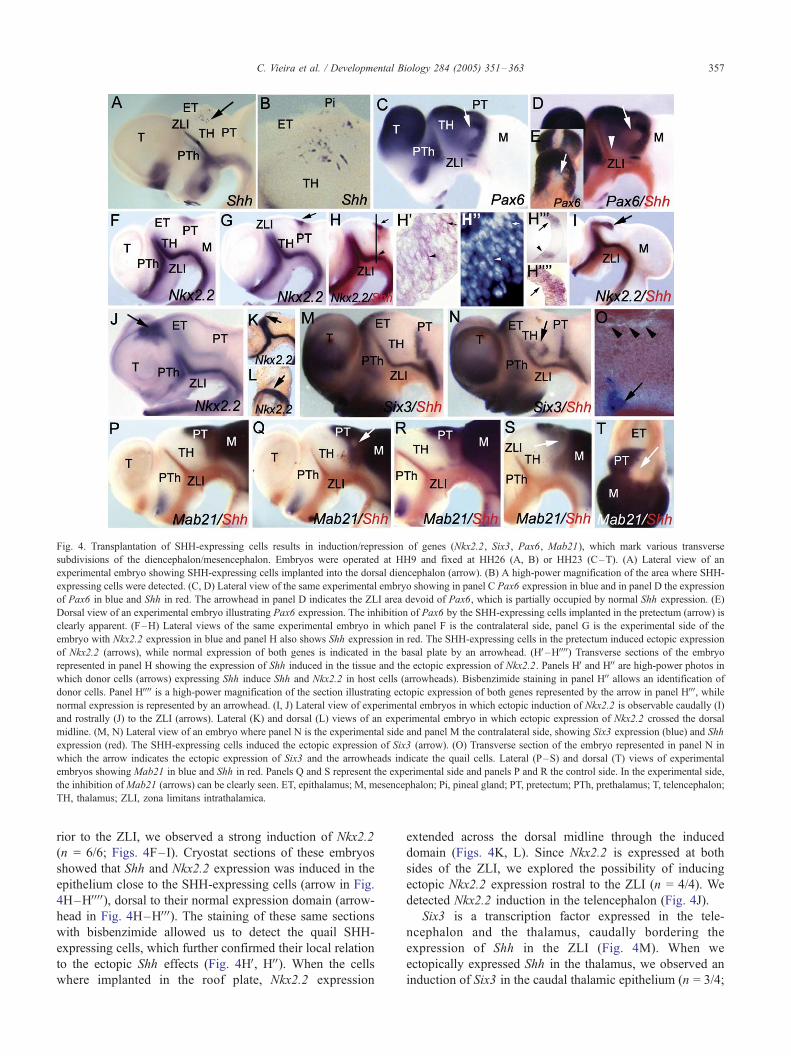
Fig. 4. Transplantation of SHH-expressing cells results in induction/repression of genes (Nkx2.2, Six3, Pax6, Mab21), which mark various transverse
subdivisions of the diencephalon/mesencephalon. Embryos were operated at HH9 and fixed at HH26 (A, B) or HH23 (C–T). (A) Lateral view of an
experimental embryo showing SHH-expressing cells implanted into the dorsal diencephalon (arrow). (B) A high-power magnification of the area where SHH-
expressing cells were detected. (C, D) Lateral view of the same experimental embryo showing in panel C Pax6 expression in blue and in panel D the expression
of Pax6 in blue and Shh in red. The arrowhead in panel D indicates the ZLI area devoid of Pax6, which is partially occupied by normal Shh expression. (E)
Dorsal view of an experimental embryo illustrating Pax6 expression. The inhibition of Pax6 by the SHH-expressing cells implanted in the pretectum (arrow) is
clearly apparent. (F–H) Lateral views of the same experimental embryo in which panel F is the contralateral side, panel G is the experimental side of the
embryo with Nkx2.2 expression in blue and panel H also shows Shh expression in red. The SHH-expressing cells in the pretectum induced ectopic expression
of Nkx2.2 (arrows), while normal expression of both genes is indicated in the basal plate by an arrowhead. (HV–HVVVV) Transverse sections of the embryo
represented in panel H showing the expression of Shh induced in the tissue and the ectopic expression of Nkx2.2. Panels HV and HVV are high-power photos inwhich donor cells (arrows) expressing Shh induce Shh and Nkx2.2 in host cells (arrowheads). Bisbenzimide staining in panel HVV allows an identification of
donor cells. Panel HVVVV is a high-power magnification of the section illustrating ectopic expression of both genes represented by the arrow in panel HVVV, whilenormal expression is represented by an arrowhead. (I, J) Lateral view of experimental embryos in which ectopic induction of Nkx2.2 is observable caudally (I)
and rostrally (J) to the ZLI (arrows). Lateral (K) and dorsal (L) views of an experimental embryo in which ectopic expression of Nkx2.2 crossed the dorsal
midline. (M, N) Lateral view of an embryo where panel N is the experimental side and panel M the contralateral side, showing Six3 expression (blue) and Shh
expression (red). The SHH-expressing cells induced the ectopic expression of Six3 (arrow). (O) Transverse section of the embryo represented in panel N in
which the arrow indicates the ectopic expression of Six3 and the arrowheads indicate the quail cells. Lateral (P–S) and dorsal (T) views of experimental
embryos showing Mab21 in blue and Shh in red. Panels Q and S represent the experimental side and panels P and R the control side. In the experimental side,
the inhibition ofMab21 (arrows) can be clearly seen. ET, epithalamus; M, mesencephalon; Pi, pineal gland; PT, pretectum; PTh, prethalamus; T, telencephalon;
TH, thalamus; ZLI, zona limitans intrathalamica.
C. Vieira et al. / Developmental Biology 284 (2005) 351–363 357
rior to the ZLI, we observed a strong induction of Nkx2.2
(n = 6/6; Figs. 4F–I). Cryostat sections of these embryos
showed that Shh and Nkx2.2 expression was induced in the
epithelium close to the SHH-expressing cells (arrow in Fig.
4H–HVVVV), dorsal to their normal expression domain (arrow-
head in Fig. 4H–HVVV). The staining of these same sections
with bisbenzimide allowed us to detect the quail SHH-
expressing cells, which further confirmed their local relation
to the ectopic Shh effects (Fig. 4HV, HVV). When the cells
where implanted in the roof plate, Nkx2.2 expression
extended across the dorsal midline through the induced
domain (Figs. 4K, L). Since Nkx2.2 is expressed at both
sides of the ZLI, we explored the possibility of inducing
ectopic Nkx2.2 expression rostral to the ZLI (n = 4/4). We
detected Nkx2.2 induction in the telencephalon (Fig. 4J).
Six3 is a transcription factor expressed in the tele-
ncephalon and the thalamus, caudally bordering the
expression of Shh in the ZLI (Fig. 4M). When we
ectopically expressed Shh in the thalamus, we observed an
induction of Six3 in the caudal thalamic epithelium (n = 3/4;
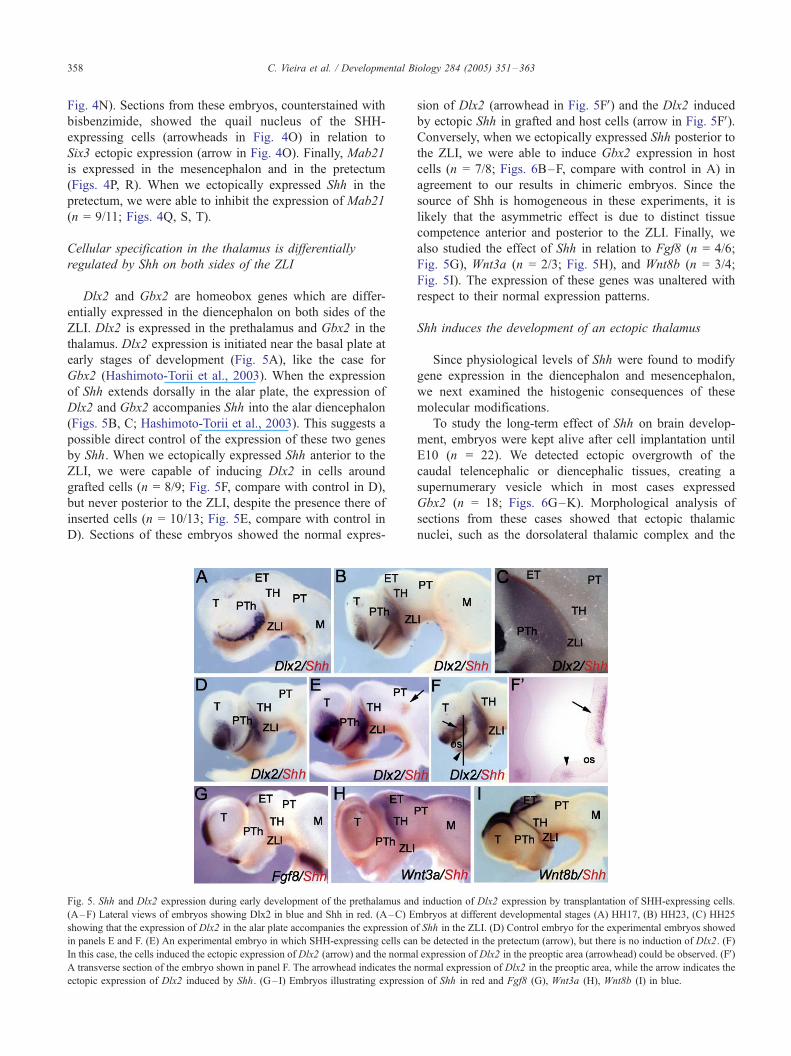
C. Vieira et al. / Developmental Biology 284 (2005) 351–363358
Fig. 4N). Sections from these embryos, counterstained with
bisbenzimide, showed the quail nucleus of the SHH-
expressing cells (arrowheads in Fig. 4O) in relation to
Six3 ectopic expression (arrow in Fig. 4O). Finally, Mab21
is expressed in the mesencephalon and in the pretectum
(Figs. 4P, R). When we ectopically expressed Shh in the
pretectum, we were able to inhibit the expression of Mab21
(n = 9/11; Figs. 4Q, S, T).
Cellular specification in the thalamus is differentially
regulated by Shh on both sides of the ZLI
Dlx2 and Gbx2 are homeobox genes which are differ-
entially expressed in the diencephalon on both sides of the
ZLI. Dlx2 is expressed in the prethalamus and Gbx2 in the
thalamus. Dlx2 expression is initiated near the basal plate at
early stages of development (Fig. 5A), like the case for
Gbx2 (Hashimoto-Torii et al., 2003). When the expression
of Shh extends dorsally in the alar plate, the expression of
Dlx2 and Gbx2 accompanies Shh into the alar diencephalon
(Figs. 5B, C; Hashimoto-Torii et al., 2003). This suggests a
possible direct control of the expression of these two genes
by Shh. When we ectopically expressed Shh anterior to the
ZLI, we were capable of inducing Dlx2 in cells around
grafted cells (n = 8/9; Fig. 5F, compare with control in D),
but never posterior to the ZLI, despite the presence there of
inserted cells (n = 10/13; Fig. 5E, compare with control in
D). Sections of these embryos showed the normal expres-
Fig. 5. Shh and Dlx2 expression during early development of the prethalamus an
(A–F) Lateral views of embryos showing Dlx2 in blue and Shh in red. (A–C) E
showing that the expression of Dlx2 in the alar plate accompanies the expression o
in panels E and F. (E) An experimental embryo in which SHH-expressing cells can
In this case, the cells induced the ectopic expression of Dlx2 (arrow) and the norma
A transverse section of the embryo shown in panel F. The arrowhead indicates the
ectopic expression of Dlx2 induced by Shh. (G–I) Embryos illustrating expressio
sion of Dlx2 (arrowhead in Fig. 5FV) and the Dlx2 induced
by ectopic Shh in grafted and host cells (arrow in Fig. 5FV).Conversely, when we ectopically expressed Shh posterior to
the ZLI, we were able to induce Gbx2 expression in host
cells (n = 7/8; Figs. 6B–F, compare with control in A) in
agreement to our results in chimeric embryos. Since the
source of Shh is homogeneous in these experiments, it is
likely that the asymmetric effect is due to distinct tissue
competence anterior and posterior to the ZLI. Finally, we
also studied the effect of Shh in relation to Fgf8 (n = 4/6;
Fig. 5G), Wnt3a (n = 2/3; Fig. 5H), and Wnt8b (n = 3/4;
Fig. 5I). The expression of these genes was unaltered with
respect to their normal expression patterns.
Shh induces the development of an ectopic thalamus
Since physiological levels of Shh were found to modify
gene expression in the diencephalon and mesencephalon,
we next examined the histogenic consequences of these
molecular modifications.
To study the long-term effect of Shh on brain develop-
ment, embryos were kept alive after cell implantation until
E10 (n = 22). We detected ectopic overgrowth of the
caudal telencephalic or diencephalic tissues, creating a
supernumerary vesicle which in most cases expressed
Gbx2 (n = 18; Figs. 6G–K). Morphological analysis of
sections from these cases showed that ectopic thalamic
nuclei, such as the dorsolateral thalamic complex and the
d induction of Dlx2 expression by transplantation of SHH-expressing cells.
mbryos at different developmental stages (A) HH17, (B) HH23, (C) HH25
f Shh in the ZLI. (D) Control embryo for the experimental embryos showed
be detected in the pretectum (arrow), but there is no induction of Dlx2. (F)
l expression of Dlx2 in the preoptic area (arrowhead) could be observed. (FV)normal expression of Dlx2 in the preoptic area, while the arrow indicates the
n of Shh in red and Fgf8 (G), Wnt3a (H), Wnt8b (I) in blue.
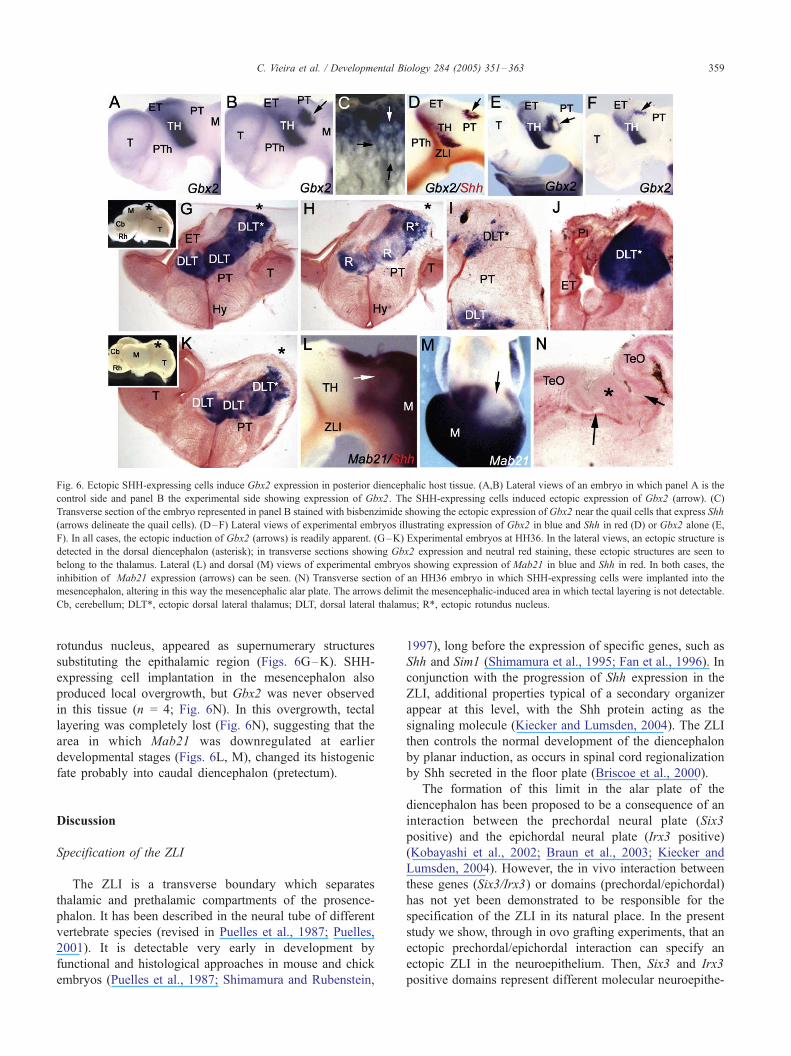
Fig. 6. Ectopic SHH-expressing cells induce Gbx2 expression in posterior diencephalic host tissue. (A,B) Lateral views of an embryo in which panel A is the
control side and panel B the experimental side showing expression of Gbx2. The SHH-expressing cells induced ectopic expression of Gbx2 (arrow). (C)
Transverse section of the embryo represented in panel B stained with bisbenzimide showing the ectopic expression of Gbx2 near the quail cells that express Shh
(arrows delineate the quail cells). (D–F) Lateral views of experimental embryos illustrating expression of Gbx2 in blue and Shh in red (D) or Gbx2 alone (E,
F). In all cases, the ectopic induction of Gbx2 (arrows) is readily apparent. (G–K) Experimental embryos at HH36. In the lateral views, an ectopic structure is
detected in the dorsal diencephalon (asterisk); in transverse sections showing Gbx2 expression and neutral red staining, these ectopic structures are seen to
belong to the thalamus. Lateral (L) and dorsal (M) views of experimental embryos showing expression of Mab21 in blue and Shh in red. In both cases, the
inhibition of Mab21 expression (arrows) can be seen. (N) Transverse section of an HH36 embryo in which SHH-expressing cells were implanted into the
mesencephalon, altering in this way the mesencephalic alar plate. The arrows delimit the mesencephalic-induced area in which tectal layering is not detectable.
Cb, cerebellum; DLT*, ectopic dorsal lateral thalamus; DLT, dorsal lateral thalamus; R*, ectopic rotundus nucleus.
C. Vieira et al. / Developmental Biology 284 (2005) 351–363 359
rotundus nucleus, appeared as supernumerary structures
substituting the epithalamic region (Figs. 6G–K). SHH-
expressing cell implantation in the mesencephalon also
produced local overgrowth, but Gbx2 was never observed
in this tissue (n = 4; Fig. 6N). In this overgrowth, tectal
layering was completely lost (Fig. 6N), suggesting that the
area in which Mab21 was downregulated at earlier
developmental stages (Figs. 6L, M), changed its histogenic
fate probably into caudal diencephalon (pretectum).
Discussion
Specification of the ZLI
The ZLI is a transverse boundary which separates
thalamic and prethalamic compartments of the prosence-
phalon. It has been described in the neural tube of different
vertebrate species (revised in Puelles et al., 1987; Puelles,
2001). It is detectable very early in development by
functional and histological approaches in mouse and chick
embryos (Puelles et al., 1987; Shimamura and Rubenstein,
1997), long before the expression of specific genes, such as
Shh and Sim1 (Shimamura et al., 1995; Fan et al., 1996). In
conjunction with the progression of Shh expression in the
ZLI, additional properties typical of a secondary organizer
appear at this level, with the Shh protein acting as the
signaling molecule (Kiecker and Lumsden, 2004). The ZLI
then controls the normal development of the diencephalon
by planar induction, as occurs in spinal cord regionalization
by Shh secreted in the floor plate (Briscoe et al., 2000).
The formation of this limit in the alar plate of the
diencephalon has been proposed to be a consequence of an
interaction between the prechordal neural plate (Six3
positive) and the epichordal neural plate (Irx3 positive)
(Kobayashi et al., 2002; Braun et al., 2003; Kiecker and
Lumsden, 2004). However, the in vivo interaction between
these genes (Six3/Irx3) or domains (prechordal/epichordal)
has not yet been demonstrated to be responsible for the
specification of the ZLI in its natural place. In the present
study we show, through in ovo grafting experiments, that an
ectopic prechordal/epichordal interaction can specify an
ectopic ZLI in the neuroepithelium. Then, Six3 and Irx3
positive domains represent different molecular neuroepithe-

C. Vieira et al. / Developmental Biology 284 (2005) 351–363360
lial areas that drive the necessary information to activate an
ectopic ZLI development. Moreover, we observed the
induction of Shh surrounding the graft and restricted to
the bordering graft cells. Although the graft was integrated
dorsally, far from the basal/alar plate limit, the expression of
Shh extended from the basal plate towards the area around
the graft. This demonstrates that the interaction between
these two domains induces a permissive property in the two
tissues to express Shh. Moreover, the expansion of the
expression domain of Shh to include the graft/host region
suggests that a possible repressive effect on Shh expression
in this domain had been reduced or blocked. Recently, it has
been proposed that the expression of Otx2 is fundamental to
restrict Shh expression to the basal plate of the midbrain
(Puelles et al., 2003). Therefore, we can speculate that, in
our experiments, Otx2 expression can be reduced between
the induced and the normal expression of Shh, suggesting
also a negative influence of Shh over Otx2 expression. The
induction of Shh in the alar plate, a primary Otx2 positive
epithelium, by ectopic SHH-expressing cells (Fig. 7A) and
the low expression of Otx2 in the Shh expressing domains
support this interpretation.
The ZLI as the thalamic organizer: molecular interactions
The dynamic expression of some genes which are
responsible for diencephalic regionalization and cell fate
could be dependent on Shh expression in the ZLI.
Shimamura’s group has demonstrated using ex vivo experi-
ments how Shh expression in the ZLI regulates Sox14 and
Gbx2 expression in the thalamic mantle layer (Hashimoto-
Torii et al., 2003). Recently, Kiecker and Lumsden (2004)
have partially reproduced these results in ovo, showing also
modifications of Pax6 expression by Shh ectopic over-
expression at both sides of the ZLI. However, it is difficult
to interpret their results since the corresponding control and
experimental embryos showed paradoxical gene expression
patterns in the basal plate that can be a source of artifactual
effects. Moreover, anatomical references were not evident in
the narrow fields shown in their figures. Our experimental
designs permit us to localize the area of ectopic Shh
expression, without affecting large neuroepithelial domains
and the basal plate. On the contrary, their experimental
approach, gene electroporation, usually generates gene over-
Fig. 7. Model of gene expression responses to signals from the zona
limitans intrathalamica (ZLI) and isthmic organizer (IsO). Single rectan-
gular boxes represent neuroepithelial cells and different colors label
heterogenic molecular domains in the neuroepithelium. (A) Schematic
interpretation of the observed effects generated by SHH-expressing cells.
(B) Schematic representation of the possible planar molecular interactions
at the level of neuroepithelial cells in the ZLI and IsO organizers.
Extracellular ‘‘diffusible’’ effects of signaling molecules (Shh and Fgf8) in
the experimental model (SHH-expressing cells) and secondary organizers
(ZLI and IsO) have been represented by interactive arrows (activation or
repression) in the lower part of each scheme. Intracellular effects have been
represented inside the schematized cells. Neuroepithelial position and
molecular background is represented by horizontal bars.

C. Vieira et al. / Developmental Biology 284 (2005) 351–363 361
expression in the electroporated epithelium that can also be
a source of additional effects. Our results are generated by
induction of Shh expression in both grafted cells and in the
host neuroepithelium, therefore not related to Shh over-
expression.
We set out to explore the effect of physiological levels of
Shh on the diencephalic alar epithelium. Since results from
grafting experiments suggested that thalamic patterning was
under the control of the ZLI, we implanted SHH-expressing
cells to explore molecular changes in this area. We observed
that Shh induced its own expression in the contacting host
epithelium, suggesting an autoregulative mechanism of
induction and maintenance of its own expression in ZLI
cells. However, the mechanisms which restrict Shh expres-
sion to the ZLI are still unknown. A good candidate in this
regard is Otx2, which has previously been considered as a
negative regulator of Shh expression in the alar plate. In
addition to this auto-inductive effect, Shh (from exogenous
SHH-expressing cells and endogenous Shh+-host cells)
induced Nkx2.2 expression in flanking neuroepithelial cells.
Other neuroepithelial genes whose expression was regulated
by Shh included: Six3, which was induced in the thalamus
and Pax6 and Mab21 which were downregulated in the
thalamus and the pretectum-mesencephalon, respectively. In
the mantle layer of the thalamus, Gbx2 expression was
activated, whereas Dlx2 was ectopically induced in the
prethalamic mantle layer. These data strongly suggest that
Shh is the signaling molecule, which mediates the organizer
activity of the ZLI. The graded repression of Pax6
expression in the thalamus, close to the ZLI, and the
recapitulation of specific gene expression patterns in ectopic
sites suggest that the morphogenetic properties of the ZLI
are indeed generated by Shh. The fact that Dlx2 was only
activated anterior to the ZLI and Gbx2 posterior to the ZLI
is indicative of a differential potential of anterior and
posterior neuroepithelia (Fig. 7A).
ZLI control of cell fate and growth in the thalamus
An ectopic source of physiological levels of Shh induced
reproducible anatomical alterations in the neuroepithelium.
This is the first time that structural effects in the brain due to
ectopic expression of Shh have been explored. Results from
previous studies are restricted to the first few days after
experimentation and the effects on brain regionalization and
structure have been exclusively speculations, based on
altered gene expression patterns. In the present study, we
report two long-term structural effects: (1) overgrowth of the
area where SHH-expressing cells were implanted, and (2)
changes in brain cyto- and myelo-architecture. It is known
that Shh can act as a mitogen in the expansion of granule
cell precursors in the cerebellum (Dahmane and Ruiz-i-
Altaba, 1999; Wallace, 1999; Wechsler-Reya and Scott,
1999), in addition to regulating dorsal brain growth by
controlling precursor proliferation (Dahmane et al., 2001).
These effects could in part be achieved via the regulation of
cyclin D1 (Ishibashi and McMahon, 2002). In addition,
these authors reported that the lack of Fgf15 expression in
the thalamus, regulated by Shh, is possibly involved in the
diencephalic alterations seen in the Shh knockout. However,
we did not observe any modifications in the expression of
Fgf8 or Fgf19 (Wright et al., 2004), as well as other
candidate genes, such as Wnt3a or Wnt8b, in response to
SHH-expressing cell implantation (Figs. 5G–I; data not
shown).
The epithalamic area was chosen as the region of the
diencephalon in which SHH-expressing cells were
implanted for some long-term survival experiments. We
have demonstrated that supernumerary dorsal thalamic
nuclei developed in place of the habenular region (a typical
structure of the epithalamus). These structures expressed
Gbx2, a marker of thalamic cells (Martinez-de-la-Torre et
al., 2002). The cytoarchitecture of the mesencephalic cortex
flanking the induced area was found in other experiments to
be substantially altered in that normal cell layering did not
develop, suggesting a change in the cytogenetic program
towards a more nuclear-like, rather than cortical structure.
A number of genes, such as Pax6 (Warren and Price,
1997), Emx2, and Otx2 (Suda et al., 2001), have been
shown to influence diencephalic regionalization. However,
members of other molecular families including Dlx, Wnts,
Bmps, and Fgfs may also participate in this process.
Multiple and varied upstream and downstream interactions
between Shh and these molecules may underlie the
generation of the different nuclear areas which make up
the complex anatomy of the diencephalon. The induction of
ectopic structures which recapitulate normal topologic
relations supports the importance of topology in gene
expression domains and its relation to a morphogenetic
activity of Shh as a signaling molecule.
In conclusion, we present evidence that neuroepithelial
interaction between the prechordal/epichordal epithelia,
which constitute two differently specified territories, is the
cellular process which underlies the specification of the ZLI,
in much the same way as Otx2/Gbx2 expressing territories
participate in the development of the IsO. The organizer
properties are driven by signaling molecules secreted by
organizer cells, i.e., Shh in the ZLI and Fgf8 in the IsO,
which control the development of the neighbouring regions
by means of morphogenetic properties (Fig. 7B). The results
of the present study indicate that the ZLI is the organizer of
the diencephalon, regulating the gene expression patterns
which determine prethalamic and thalamic regionalization.
These events are necessary to generate nuclear and cellular
diversity, which is the basis of the complex functional
properties of this brain region.
Acknowledgments
We thank R. Garcia-Lopez for cresyl violet staining; M.
Rodenas, F. Almagro, and M. Bonete for technical

C. Vieira et al. / Developmental Biology 284 (2005) 351–363362
assistance. Work supported by the grants: UE QLRT-1999-
31625; QLRT-2000-02310; FCT/MCES; DIGESIC-MEC
PM98-0056; BFI2002-02979; GV CTDIA/2002/91. The
authors would like to express their thanks to the agency
ACTS (Academic Consulting and Translating Services;
http://www.euskalnet.net/acts) for having corrected the
English of this paper.
References
Agarwala, S., Sanders, T.A., Ragsdale, C.W., 2001. Sonic hedgehog
control of size and shape in midbrain pattern formation. Science 291,
2147–2150.
Braun, M.M., Etheridge, A., Bernard, A., Robertson, C.P., Roelink, H.,
2003. Wnt signaling is required at distinct stages of development for the
induction of the posterior forebrain. Development 130, 5579–5587.
Briscoe, J., Pierani, A., Jessel, T.M., Ericson, J., 2000. A homeodomain
protein code specifies progenitor cell identity and neuronal fate in the
ventral neural tube. Cell 101, 435–445.
Bulfone, A., Puelles, L., Porteus, M.H., Frohman, M.A., Martin, G.R.,
Rubenstein, J.L., 1993. Spatially restricted expression of Dlx-1, Dlx-2
(Tes-1), Gbx-2, and Wnt-3 in the embryonic day 12.5 mouse
forebrain defines potential transverse and longitudinal segmental
boundaries. J. Neurosci. 13, 3155–3172.
Chiang, C., Litingtung, Y., Lee, E., Young, K.E., Corden, J.L., Westphal,
H., Beachy, P.A., 1996. Cyclopia and defective axial patterning in mice
lacking Sonic hedgehog gene function. Nature 383, 407–413.
Cobos, I., Shimamura, K., Rubenstein, J.L., Martinez, S., Puelles, L., 2001.
Fate map of the avian anterior forebrain at the four-somite stage, based
on the analysis of quail-chick chimeras. Dev. Biol. 239, 46–67.
Crossley, P.H., Martinez, S., Ohkubo, Y., Rubenstein, J.L., 2001.
Coordinate expression of Fgf8, Otx2, Bmp4, and Shh in the rostral
prosencephalon during development of the telencephalic and optic
vesicles. Neuroscience 108, 183–206.
Dahmane, N., Ruiz-i-Altaba, A., 1999. Sonic hedgehog regulates the growth
and patterning of the cerebellum. Development 126, 3089–3100.
Dahmane, N., Sanchez, P., Gitton, Y., Palma, V., Sun, T., Beyna, M.,
Weiner, H., Ruiz-i-Altaba, A., 2001. The Sonic Hedgehog–Gli pathway
regulates dorsal brain growth and tumorigenesis. Development 128,
5201–5212.
Duprez, D., Fournier-Thibault, C., Le Douarin, N., 1998. Sonic hedgehog
induces proliferation of committed skeletal muscle cells in the chick
limb. Development 125, 495–505.
Echevarria, D., Vieira, C., Gimeno, L., Martinez, S., 2003. Neuroepithelial
secondary organizers and cell fate specification in the developing brain.
Brain Res. Rev. 43, 179–191.
Ericson, J., Rashbass, P., Schedl, A., Brenner-Morton, S., Kawakami, A.,
van Heyningen, V., Jessel, T.M., Briscoe, J., 1997. Pax6 controls
progenitor cell identity and neuronal fate in response to graded Shh
signaling. Cell 90, 169–180.
Fan, C.M., Kuwana, E., Bulfone, A., Fletcher, C.F., Copeland, N.G.,
Jenkins, N.A., Crews, S., Martinez, S., Puelles, L., Rubenstein, J.L.,
Tessier-Lavigne, M., 1996. Expression patterns of two murine homo-
logs of Drosophila single-minded suggest possible roles in embryonic
patterning and in the pathogenesis of Down syndrome. Mol. Cell.
Neurosci. 7, 1–16.
Figdor, M.C., Stern, C.D., 1993. Segmental organization of embryonic
diencephalon. Nature 363, 630–634.
Garcia-Lopez, R., Vieira, C., Echevarria, D., Martinez, S., 2004. Fate map
of the diencephalon and the zona limitans at the 10-somites stage in
chick embryos. Dev. Biol. 268, 514–530.
Garda, A.L., Echevarria, D., Martinez, S., 2001. Neuroepithelial co-
expression of Gbx2 and Otx2 precedes Fgf8 expression in the isthmic
organizer. Mech. Dev. 101, 111–118.
Gonzalez, G., Puelles, L., Medina, L., 2002. Organization of the mouse
dorsal thalamus based on topology, calretinin immunostaining and gene
expression. Brain Res. Bull. 57, 439–442.
Hamburger, V., Hamilton, H.L., 1951. A series of normal stages in the
development of the chick embryo. J. Morphol. 88, 49–92.
Hashimoto-Torii, K., Motoyama, J., Hui, C., Kuroiwa, A., Nakafuku, M.,
Shimamura, K., 2003. Differential activities of Sonic hedgehog
mediated by Gli transcription factors define distinct neuronal subtypes
in the dorsal thalamus. Mech. Dev. 120, 1097–1111.
Hidalgo-Sanchez, M., Simeone, A., Alvarado-Mallart, R.M., 1999. Fgf8
and Gbx2 induction concomitant with Otx2 repression is correlated with
midbrain–hindbrain fate of caudal prosencephalon. Development 126,
3191–3203.
Ingham, P.W., McMahon, A.P., 2001. Hedgehog signaling in animal
development: paradigms and principles. Genes Dev. 15, 3059–3087.
Ishibashi, M., McMahon, A.P., 2002. A sonic hedgehog-dependent signal-
ing relay regulates growth of diencephalic and mesencephalic primor-
dial in the early mouse embryo. Development 129, 4807–4819.
Joyner, A.L., Liu, A., Millet, S., 2000. Otx2, Gbx2 and Fgf8 interact to
position and maintain a mid–hindbrain organizer. Curr. Opin. Cell Biol.
12, 736–741.
Kiecker, C., Lumsden, A., 2004. Hedgehog signaling from the ZLI
regulates diencephalic regional identity. Nat. Neurosci. 7, 1242–1249.
Kobayashi, D., Kobayashi, M., Matsumoto, K., Ogura, T., Nakafuku, M.,
Shimamura, K., 2002. Early subdivisions in the neural plate define
distinct competence for inductive signals. Development 129, 83–93.
Larsen, C.W., Zeltser, L.M., Lumsden, A., 2001. Boundary formation and
compartition in the avian diencephalon. J. Neurosci. 21, 4699–4711.
Mariani, M., Corradi, A., Baldessari, D., Malgaretti, N., Pozzoli, O.,
Fesce, R., Martinez, S., Boncinelli, E., Consalez, G.G., 1998. Mab21,
the mouse homolog of a C. elegans cell-fate specification gene,
participates in cerebellar, midbrain and eye development. Mech. Dev.
79, 131–135.
Mariani, M., Baldessari, D., Francisconi, S., Viggiano, L., Rocchi, M.,
Zappavigna, V., Malgaretti, N., Consalez, G.G., 1999. Two murine and
human homologs of mab-21, a cell fate determination gene involved in
Caenorhabditis elegans neural development. Hum. Mol. Genet. 8,
2397–2406.
Marti, E., Bovolenta, P., 2002. Sonic hedgehog in CNS development: one
signal, multiple outputs. Trends Neurosci. 25, 89–96.
Martinez, S., Puelles, L., 2000. Neurogenetic compartments of the mouse
diencephalon and some characteristic gene expression patterns. Results
Probl. Cell Differ. 30, 91–106.
Martinez, S., Geijo, E., Sanchez-Vives, M.V., Puelles, L., Gallego, R.,
1992. Reduced junctional permeability at inter-rhombomeric bounda-
ries. Development 116, 1069–1076.
Martinez-Barbera, J.P., Signore, M., Boyl, P.P., Puelles, E., Acampora,
D., Gogoi, R., Schubert, F., Lumsden, A., Simeone, A., 2001.
Regionalisation of anterior neuroectoderm and its competence in
responding to forebrain and midbrain inducing activities depend on
mutual antagonism between OTX2 and GBX2. Development 128,
4789–4800.
Martinez-de-la-Torre, M., Martinez, S., Puelles, L., 1990. Acetylcholines-
terase-histochemical differential staining of subdivisions within the
nucleus rotundus in the chick. Anat. Embryol. (Berl) 181, 129–135.
Martinez-de-la-Torre, M., Garda, A.L., Puelles, E., Puelles, L., 2002. Gbx2
expression in the late embryonic chick dorsal thalamus. Brain Res. Bull.
57, 435–438.
McConnell, S.K., Kaznowski, C.E., 1991. Cell cycle dependence of laminar
determination in developing neocortex. Science 254, 282–285.
Nieto, M.A., Patel, K., Wilkinson, D.G., 1996. In situ hybridization analysis
of chick embryos in whole mount and tissue sections. Methods Cell
Biol. 51, 219–235.
Oliver, T.G., Read, T.A., Kessler, J.D., Mehmeti, A., Wells, J.F., Huynh,
T.T., Lin, S.M., Wechsler-Reya, R.J., 2005. Loss of patched and
disruption of granule cell development in a preneoplastic stage of
medulloblastoma. Development 132, 2425–2439.

C. Vieira et al. / Developmental Biology 284 (2005) 351–363 363
Price, M., Lazzaro, D., Pohl, T., Mattei, M.G., Ruther, U., Olivo, J.C.,
Duboule, D., Di Lauro, R., 1992. Regional expression of the
homeobox gene Nkx-2.2 in the developing mammalian forebrain.
Neuron 8, 241–255.
Puelles, L., 2001. Brain segmentation and forebrain development in
amniotes. Brain Res. Bull. 55, 695–710.
Puelles, L., Rubenstein, J.L.R., 2003. Forebrain gene expression domains
and the evolving prosomeric model. Trends Neurosci. 26, 469–476.
Puelles, L., Amat, J.A., Martinez-de-la-Torre, M., 1987. Segment-related,
mosaic neurogenetic pattern in the forebrain and mesencephalon of
early chick embryos: I. Topography of AChE-positive neuroblasts up to
stage HH18. J. Comp. Neurol. 266, 247–268.
Puelles, E., Acampora, D., Lacroix, E., Signore, M., Annino, A., Tuorto, F.,
Filosa, S., Corte, G., Wurst, W., Ang, S.L., Simeone, A., 2003. Otx
dose-dependent integrated control of antero-posterior and dorso-ventral
patterning of midbrain. Nat. Neurosci. 6, 453–460.
Roelink, H., Porter, J.A., Chiang, C., Tanabe, Y., Chang, D.T., Beachy, P.A.,
Jessel, T.M., 1995. Floor plate and motor neuron induction by different
concentrations of the amino-terminal cleavage product of sonic hedge-
hog autoproteolysis. Cell 81, 445.
Shimamura, K., Rubenstein, J.L.R., 1997. Inductive interactions direct
early regionalization of the mouse forebrain. Development 124,
2709–2718.
Shimamura, K., Hartigan, D.J., Martinez, S., Puelles, L., Rubenstein, J.L.,
1995. Longitudinal organization of the anterior neural plate and neural
tube. Development 121, 3923–3933.
Suda, Y., Hossain, Z.M., Kobayashi, C., Hatano, O., Yoshida, M., Matsuo,
I., Aizawa, S., 2001. Emx2 directs the development of diencephalon in
cooperation with Otx2. Development 128, 2433–2450.
Wallace, V.A., 1999. Purkinje-cell-derived Sonic hedgehog regulates
granule neuron precursor cell proliferation in the developing mouse
cerebellum. Curr. Biol. 9, 445–448.
Warren, N., Price, D.J., 1997. Roles of Pax-6 in murine diencephalic
development. Development 124, 1573–1582.
Wechsler-Reya, R.J., Scott, M.P., 1999. Control of neuronal precursor pro-
liferation in the cerebellum by Sonic hedgehog. Neuron 22, 103–114.
Wright, T.J., Ladher, R., McWhirter, J., Murre, C., Schoenwolf, G.C.,
Mansour, S.L., 2004. Mouse FGF15 is the ortholog of human and chick
FGF19, but is not uniquely required for otic induction. Dev. Biol. 269,
264–275.