structural and ultrastructural evidence of neurotoxic effects of fried potato chips on rat postnatal...
TRANSCRIPT

lable at ScienceDirect
Nutrition 27 (2011) 1066–1075
Contents lists avai
Nutrition
journal homepage: www.nutr i t ionjrnl .com
Basic nutritional investigation
Structural and ultrastructural evidence of neurotoxic effects of fried potatochips on rat postnatal development
Hassan I. El-Sayyad Ph.D. a, Hekmat L. El-Gammal Ph.D. a, Lotfy A. Habak Ph.D. a,Heba M. Abdel-Galil Ph.D. a, Augusta Fernando Ph.D. b, Rajiv L. Gaur Ph.D. b, Allal Ouhtit M.Ph., Ph.D. b,*aDepartment of Zoology, Faculty of Science, Mansoura University, Mansoura, EgyptbDepartment of Genetics, College of Medicine and Health Sciences, Sultan Qaboos University, Muscat, Sultanate of Oman
a r t i c l e i n f o
Article history:Received 2 March 2011Accepted 16 June 2011
Keywords:AcrylamideFried potato chipsCerebellar cortexMuscle developmentPregnant ratsPostnatal development
* Corresponding author. Tel.: þ968-2414-3526; faxE-mail address: [email protected] (A. Ouhtit).
0899-9007/$ - see front matter � 2011 Published bydoi:10.1016/j.nut.2011.06.008
a b s t r a c t
Objective: Acrylamide (ACR), a proved rodent carcinogen and neurotoxic agent, is present insignificant quantities in commonly consumed foods such as fried potato chips (FPC) and Frenchfries, raising a health concern worldwide. We investigated and compared the neurotoxic effects ofACR and FPC on postnatal development.Methods: Female rats were treated with ACR (30 mg/kg of body weight), fed a diet containingapproximately 30% of FPC during pregnancy, or fed a standard diet (control) and their offspringwere examined.Results: Female rats treated with ACR or fed a diet containing FPC during pregnancy gave birth tolitters with delayed growth and decreased body and brain weights. Light microscopic studies of thecerebellar cortex of treated animals revealed drastic decreases in Purkinje cells and internalgranular layers. Different patterns of cell death were detected in Purkinje cells and neurons in thebrains of pups born to treated mothers. Ultrastructural analysis of Purkinje cells revealed changesin the endoplasmic reticulum, loss of the normal arrangement of polyribosomes, swollen mito-chondria with abnormally differentiated cristae, and an abnormal Golgi apparatus. The gastroc-nemius muscle in the ACR and FPC groups showed extensive degeneration of myofibrils asevidenced by poorly differentiated A, H, and Z bands.Conclusion: The present study reveals for the first time that rat fetal exposure to ACR, as a purecompound or from a maternal diet of FPC, causes cerebellar cortical defects and myodegenerationof the gastrocnemius muscle during the postnatal development of pups. These results warranta systematic study of the health effects of the consumption of FPC and French fries in the generalpopulation.
� 2011 Published by Elsevier Inc.
Introduction
Acrylamide (ACR) is a neurotoxic agent known to produceseveral pathologic effects in animals and humans at high dosesgiven repeatedly [1], but little is known of its potential effectsduring pre- and postnatal development. Humans are continu-ously exposed to low doses of ACR found in significant amountsin carbohydrate-containing food items cooked at high temper-atures [2,3]. Cigarette smoke is an additional and importantsource of ACR exposure [4]. According to recent data from theWorld Health Organization, the human intake of ACR is 1 to4 mg/kg of body weight per day [5].
: þ968-2414-3331.
Elsevier Inc.
Humans are continuously exposed to ACR through the diet,beginning with in utero exposures, because ACR can cross theplacenta [6–8]. The exposure continues through breast-feeding[8,9] and the consumption of baby food [10]. In pregnantwomen, 10% to 50% of dietary ACR has been found to coursethrough the placental blood to the fetus [8]. Breast milk has beenfound to contain ACR up to 18.8 mg/L [8]. The solubility of ACR inwater enables its passage through maternal–placental andblood–brain barriers [8]. ACR has half-lives of approximately 2 hinbloodand5h to8d in tissue [11,12]. ACRhasbeenshowntobindto neuronal DNA [13], which may result in the production ofabnormal structural proteins essential for axonal function [14,15].Currentevidencehas suggested that the cumulativeneurotoxicityproduced by ACR exposure is linked to nerve terminal damage inthe central and peripheral nervous systems [16–19].

H. I. El-Sayyad et al. / Nutrition 27 (2011) 1066–1075 1067
In addition, studies of ACR neuropathology using differentdoses and conditions have provided evidence that ACR toxicity isassociated with initial nerve terminal degeneration and cere-bellar Purkinje cell injury (reviewed by LoPachin et al. [20]). Earlymorphologic studies have indicated distal axonopathy because oftechnical limitations and a difficulty to detect nerve terminaldegeneration by standard neuropathologic examinations [16,17].The research on the effects of ACR in the brain has been focusedon nerve terminals and Purkinje cells as primary sites of directACR action (reviewed by LoPachin et al. [20]). The underlyingmechanisms of ACR-mediated neurotoxicity have been linked tooxidative stress [21]. Studies on experimental models haveshown that ACR exposure results in skeletal muscle weakness,particularly in the gastrocnemius muscle [20,22–24].
A recent study has shown that ACR levels of potato chipsprepared by frying increasewith frying temperature (19.6 ng/g at170�C, 39 ng/g at 180�C, and 95 ng/g at 190�C) [25]. Furthermore,humans are continuously exposed to ACR at low levels throughthe diet (and potentially smoking), which raises the possibility oflong-term neurologic degeneration caused by ACR during the lifespan [26]. Concerns of the potential health issues associatedwitha long-term dietary intake of ACR prompted us to investigate andcompare the effects of ACR with those of fried potato chips (FPC)given to rats during pregnancy and for 4 wk postnatally on thecerebellar cortical development of young pups and its effect onthe histopathology of the gastrocnemius muscle.
Materials and methods
ACR treatment
Acrylamide of the highest purity (Sigma-Aldrich, St. Louis, MO, USA) wasused in the present work. The dose of 30mg/kg of body weight [17] was dissolvedin 0.2 mL of saline solution and administered daily by stomach tube to pregnantrats from day 6 of gestation until parturition and throughout the lactation period(4 wk).
FPC supplementation
Fried potato chips were purchased from the market. A standard diet con-taining approximately 30% of FPC was fed to pregnant rats from the sixth day ofgestation until parturition and 4 wk postpartum.
Animals and treatment
Eighty fertile virgin female and fertile male albino rats (Rattus novegius),caged at a ratio of three females to one1 male and weighing approximately 120 g,were bred at the Mansoura University experimental animal core facility. Animalsubjects were maintained in the animal care facility throughout the duration ofthe experiment. The ambient temperature was kept at 22�C to 24�C with a 12-hlight/dark cycle. Breeding was encouraged by exposing female rats to healthyfertile male rats for 12 h from 20:00 to 08:00. In the morning, vaginal smearswere obtained to identify the onset of gestation by confirming the presence ofsperm in the smear. All experiments were carried out in accord with protocolsapproved by the local experimental animal ethics committee.
The pregnant rats were then divided into three groups of 20 each. The controlgroupwasmaintained on a standard diet (Table 1). The second group consisted ofACR-treated pregnant rats who each received a daily oral gavage of ACR 30mg/kg
Table 1Description of different groups of rats and their respective diets
Animal group Food supplemented during study
Control (n ¼ 20) standard diet*
ACR (n ¼ 40) standard diet* þ ACR (30 mg/kg body weight)in 0.2 mL of saline
FCP (n ¼ 20) standard diet* þ w30% FPCs
ACR, acrylamide; FCP, fried potato chip* The standard diet consisted of 50% grind barley, 10% grind yellowmaize, 20%
milk, and 10% vegetable.
starting from the sixth day of gestation until parturition and at 1, 2, 3, and 4 wkpostpartum. The subchronic dose of ACR was given as previously reported [27].The third group consisted of FPC-fed pregnant rats that were given a standarddiet consisting of approximately 30% FPC starting from the sixth day of pregnancyuntil lactation.
Histopathology and ultrastructural analyses
Newborns and young pups of the control and experimental groups weresacrificed and the following parameters were investigated. Growth patterns ofpups at parturition and 1, 2, 3, and 4 wk postpartum were determined.
Gross morphology and brain weight of young rats
The whole brain was obtained and weighed (data not shown) at the statedintervals during the experimental procedures.
Light microscopic investigations
The cerebellum and gastrocnemius muscle of the sacrificed young pups ofthe control and treated dams were fixed in 10% paraformaldehyde solution,dehydrated in ascending grades of ethyl alcohol, cleared in xylol, and embeddedin paraplast. Five-micrometer sections were stained with hematoxylin and eosin.Extrawhole brains from other specimens of the control and experimental groupswere stained with the Golgi–Cox technique and examined under a bright-fieldlight microscope.
Transmission electron microscopic investigations
The specimens were dehydrated, subjected to critical point drying, andcoated by platinum-palladium ion sputtering. The specimens were then evalu-ated by a USM-5400LV Joel scanning electron microscope (Peabody, MA, USA).Differentiated cerebellum sections and gastrocnemius muscles of 3-wk-old ratswere collected, fixed in 2.5% glutaraldehyde and 2% paraformaldehyde in 0.1 Mcacodylate buffer (pH 7.4), rinsed, and postfixed in a buffered solution of 1%osmium tetroxide at 4�C for 1.5 h. Dehydration in ascending grades of ethylalcohol was performed and the specimens were embedded in epoxy resin.Ultrathin sections were obtained using a diamond knife on an LKB microtomeand mounted on formvar-coated grids, stained with uranyl acetate and leadcitrate, and examined by a Joel transmission electron microscope.
Extra whole brains from other specimens of the control and experimentalgroups were processed for staining with the Golgi–Cox technique according tothe method of Kiernan [27] and examined under a bright-field light microscope.
Statistical analysis
Statistical analysis was carried out between the control and the experimentalgroups at each developmental stage to compare the different values of bodyweight. Means and standard errors were calculated for each group, and theStudent t test was applied. The differences were considered statistically signifi-cant from the control at P < 0.05.
Results
Gross morphology of young rats
Morphologic analysis of postnatal young pups of pregnantrats that were exposed to ACR (30 mg/kg of body weight) orreceived food supplementation containing FPC showed signifi-cant decreases in body (Fig. 1) and brain (Fig. 2) sizes and bodyweight (Fig. 3).
Ultrastructural analysis of cerebellar cortex
The cerebellar cortical surface of the 3-wk-old postnatal ratsexposed to ACR or FPC showed fewer immature cerebellar foldsmanifested by decreased invagination and the differentiation ofsuperficially emerging cerebellar cortical nerves (Fig. 4A–C).

Fig. 1. Effects of ACR and FPC on the appearance of young rats and their postnatal growth at day 1 and at weeks 1, 2, 3, and 4. (A–A4) Control; (B–B4) maternally treated withACR; (C–C4) maternally fed FPC. ACR, acrylamide; FPC, fried potato chips.
H. I. El-Sayyad et al. / Nutrition 27 (2011) 1066–10751068
Light microscopic observations of cerebellar cortex duringpostnatal development
Newborns of control subjects exhibited normal differentiationof the four main cell layers, namely the external granular layer,molecular layer, Purkinje cell layer, and internal granular layer(Fig. 5A), whereas the cerebellar cortex of delivered newbornoffspring from groups exposed to ACR or fed FPC showed alteredhistologic structures and an abnormal architecture characterizedby disrupted external and internal granular cell layers that man-ifested massive increases of pyknotic neuronal cells separated bywidened spaces (Fig. 5A1).
At 1 wk, compared with controls (Fig. 5B), the internalgranular cell layer of young pups of rats exposed to ACR or fedFPC (Fig. 5B1, B2) showed a smaller number of neuronal cells andless dense grouping, with the neuronal cells widely separated byisland spaces. There was a marked decrease in Purkinje cellnumber compared with the control group, and many of thesecells had karyolytic or pyknotic nuclei (Fig. 5B1, B2).
In 3-wk-old (Fig. 5C) and 4-wk-old (Fig. 5D) control postnatalrats, the external granular layer was markedly decreased, theneuronal cells appeared collapsed, and the Purkinje cells attainedmore growth and differentiation and were regularly arranged atthe periphery of the internal granular layer. However, the youngpups of rats treated with ACR or fed the FPC-containing dietshowed marked disruption of the internal granular cell layer,including a numerical decrease of neuronal cells and widenedglomerular spaces (Fig. 5C1, C2, D1, D2). Few Purkinje cells weredetected as solitary or aggregated in clusters and most appearedatrophied with pyknotic or karyolytic nuclei. Interestingly, thesepathologic alterationsweremore prominent in those pups of ratsfed the FPC-containing diet (Fig. 5C1, C2, D1, D2).
Transmission electron microscopic analysis
The most prominent abnormalities were observed in the 3-wk-old postnatal pups of rats treatedwith ACR (Fig. 6A1–C1) orfed the diet containing FPC (Fig. 6A2–C2) compared withcontrols (Fig. 6A–C). The Purkinje cells were shrunken andpossessed abnormal ultrastructural features. Some cellspossessed atrophied nuclei with a pattern of a dense distribu-tion of heterochromatin with or without irregular folded andruptured nuclear membranes associated with karyolysis anddegeneration of nuclear chromatin. The cytoplasmic organelleswere abnormally altered in structure and distribution. Therough endoplasmic reticulum appeared vesiculated and dis-rupted throughout the cytoplasm. The mitochondria showedconsiderable swelling and lack of differentiation of theirinternal compartments. TheGolgi apparatuswas elongated andhypertrophied. The polyribosomes appeared to be missing orlacked a regular arrangement pattern. The abnormal charac-teristic pattern of cell organelles manifested the disruption ofNissl body formation. The neuronal axon showed a markeddepletion of a myelin coating with an irregular distributionpattern along the neuronal axons. Some neuronal axonsshowed a complete absence of myelin.
Morphology of Purkinje cells
Purkinje cells of 3-wk-old control subjects stained witha Golgi–Cox stain exhibited well-defined branching with largecell bodies (Fig. 7A). However, the Purkinje cells of young pups ofrats treated with ACR or fed with FPC exhibited cell bodies withabnormal main axons devoid of dendritic formations (Fig. 7B, C).

Fig. 2. Effects of ACR and FPC on the shape and size of neonate brains at day 1 and weeks 1, 2, 3, and 4 postnatally. (A–A4) Control; (B–B4) treated with ACR; (C–C4) treatedwith FCP. ACR, acrylamide; Ce, cerebellum; CH, cerebrum; FPC, fried potato chips; MO, medulla oblongata; OB, olfactory bulb; PG, pineal gland.
H. I. El-Sayyad et al. / Nutrition 27 (2011) 1066–1075 1069
Histologic observations of the gastrocnemius muscle
Comparative light microscopic analysis of the gastrocnemiusmuscle of the pups of the control versus the ACR-treated andFPC-fed groups revealedmarked pathologic alterations that weremore prominent in the FPC group. Compared with newborncontrol rats (Fig. 8A), the newborn pups of ACR-treated and FPC-fed mothers exhibited muscular dystrophy manifested by a lossof muscular fiber arrangement integrity and a myodegeneration
Fig. 3. Effects of ACR and FPC on the body weight of newly born offspring at day 1and at weeks 1, 2, 3, and 4 postnatally. * P < 0.05 (Student t test) for treated groupsversus the control group. ACR, acrylamide; FPC, fried potato chips.
of muscle fibers (Fig. 8B1, B2). The gastrocnemius muscle ofyoung pups from rats treated with ACR or fed the FPC-containingdiet exhibitedmyopathologic alterations (Fig. 8B2). Transmissionelectron microscopic analysis of the gastrocnemius muscle of 3-wk-old control offspring exhibited a regular arrangement ofmuscle structures (Fig. 9A). In the ACR and FPC groups, clearmyodegeneration and dramatic pathologic alterations weredetected, which were more prominent in those pups of rats fedthe FPC-supplemented diet (Fig. 9B, C).
Discussion
Previous ACR neurotoxicity studies in rodents were con-ducted before the knowledge of the presence of ACR in friedfoods (reviewed by the World Health Organization [5]). Thisstudy provides the first evidence of the comparable neurotoxiceffects of ACR and FPC using structural and ultrastructuralapproaches. The present results indicate that the prenatalexposure of rat fetuses to ACR, as a pure compound or froma maternal diet of FPC, caused cerebellar cortical degenerationand myodegeneration of the gastrocnemius muscle during thepostnatal development of pups.
Acrylamide is generated during the heating of specific food-stuffs as a result of a Maillard reaction between amino acids andsugars [28–30]. In particular, ACR is formed when the amino acidasparagine in the presence of sugar is heated above 100�C. Largeamounts of asparagine have been found in potatoes and cereals,which were found to have the highest measured levels of ACR inthe Swedish National Food Administration survey [31].

Fig. 4. Scanning electron micrographs showing the effects of ACR and FPC on the cerebellum of a 3-wk-old fetus. (A, A1) Control; (B, B1) ACR treatment; (C, C1) FCP treatment.ACR, acrylamide; FPC, fried potato chips.
H. I. El-Sayyad et al. / Nutrition 27 (2011) 1066–10751070
Several lines of evidence from the literature support ourfindings. Rat litters born to mothers treated with ACR duringgestation have shown a pronounced decrease of mean bodyweight during postnatal growth [32,33]. The investigatorsattributed the decrease of body weight to a severe decrease infeeding associated with little or no milk in the stomach, whichmight be due to loss of appetite [32,34]. In the present study,postnatal young pups from rats treated with ACR and FPCshowed similar results. Furthermore, the brain weights of post-natal young pups of ACR-treated or FPC-fed dams differedmarkedly from controls, suggesting early signs of neurotoxicity.A previous study has reported an increase in the level of ACR inthe blood of treated mothers and the umbilical cord blood oftheir neonates, leading to the formation of hemoglobin adductsand decreased cell replication and neurotoxicity [35].
Previous studies have reported ACR-induced neurotoxicity inthe rat model [36,37]. The present study demonstrated thatmaternal treatment with ACR or feeding with FPC throughout
pregnancy and lactation leads tomassive histologic alterations inthe development of the cerebellar cortex, including increasedrates of apoptotic cell death of Purkinje cells and granularneuronal cells. These alterations suggest that maternal treatmentwith ACR or feeding with FPC can cause a significant inhibition inthe proliferation and maturation process of the cerebellar cortexin the newborn and during postnatal growth. At the transmissionelectron microscopic level, the Purkinje cells of the experimentalgroups exhibited cell changes manifested by the fragmentationand disruption of the rough endoplasmic reticulum, swolleninternal compartments of the mitochondria, and an abnormallyelongated and hypertrophied Golgi apparatus. The characteristicpattern of Nissl bodies disappeared. The neuronal axons showeda marked depletion of myelinated coating with an irregularlydistributed pattern along the neuronal axons. Some neuronalaxons showed a complete absence of myelin. The nuclei exhibi-ted clumped or karyolytic chromatin materials. Lopachin et al.[37] attributed the significant decrease of myelination to the
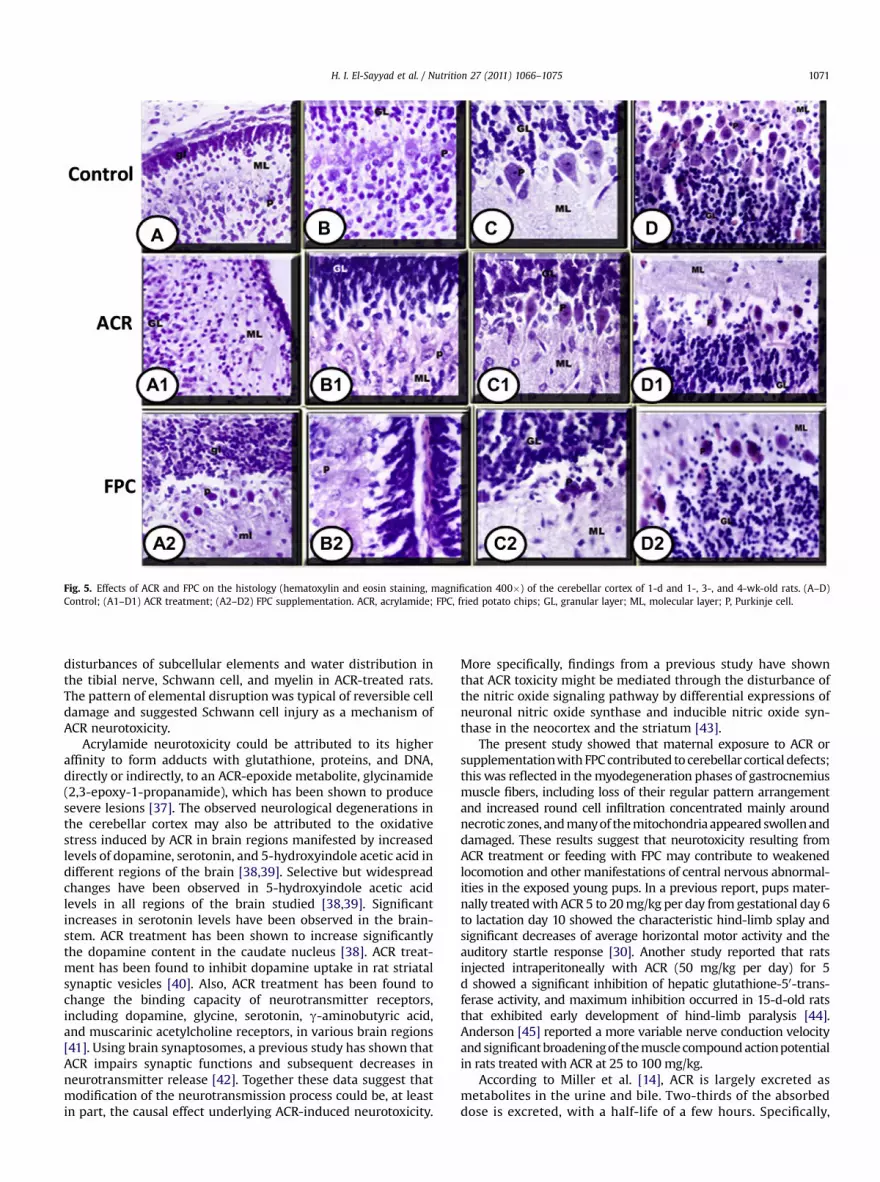
Fig. 5. Effects of ACR and FPC on the histology (hematoxylin and eosin staining, magnification 400�) of the cerebellar cortex of 1-d and 1-, 3-, and 4-wk-old rats. (A–D)Control; (A1–D1) ACR treatment; (A2–D2) FPC supplementation. ACR, acrylamide; FPC, fried potato chips; GL, granular layer; ML, molecular layer; P, Purkinje cell.
H. I. El-Sayyad et al. / Nutrition 27 (2011) 1066–1075 1071
disturbances of subcellular elements and water distribution inthe tibial nerve, Schwann cell, and myelin in ACR-treated rats.The pattern of elemental disruption was typical of reversible celldamage and suggested Schwann cell injury as a mechanism ofACR neurotoxicity.
Acrylamide neurotoxicity could be attributed to its higheraffinity to form adducts with glutathione, proteins, and DNA,directly or indirectly, to an ACR-epoxide metabolite, glycinamide(2,3-epoxy-1-propanamide), which has been shown to producesevere lesions [37]. The observed neurological degenerations inthe cerebellar cortex may also be attributed to the oxidativestress induced by ACR in brain regions manifested by increasedlevels of dopamine, serotonin, and 5-hydroxyindole acetic acid indifferent regions of the brain [38,39]. Selective but widespreadchanges have been observed in 5-hydroxyindole acetic acidlevels in all regions of the brain studied [38,39]. Significantincreases in serotonin levels have been observed in the brain-stem. ACR treatment has been shown to increase significantlythe dopamine content in the caudate nucleus [38]. ACR treat-ment has been found to inhibit dopamine uptake in rat striatalsynaptic vesicles [40]. Also, ACR treatment has been found tochange the binding capacity of neurotransmitter receptors,including dopamine, glycine, serotonin, g-aminobutyric acid,and muscarinic acetylcholine receptors, in various brain regions[41]. Using brain synaptosomes, a previous study has shown thatACR impairs synaptic functions and subsequent decreases inneurotransmitter release [42]. Together these data suggest thatmodification of the neurotransmission process could be, at leastin part, the causal effect underlying ACR-induced neurotoxicity.
More specifically, findings from a previous study have shownthat ACR toxicity might be mediated through the disturbance ofthe nitric oxide signaling pathway by differential expressions ofneuronal nitric oxide synthase and inducible nitric oxide syn-thase in the neocortex and the striatum [43].
The present study showed that maternal exposure to ACR orsupplementationwith FPCcontributed to cerebellar cortical defects;this was reflected in themyodegeneration phases of gastrocnemiusmuscle fibers, including loss of their regular pattern arrangementand increased round cell infiltration concentrated mainly aroundnecrotic zones, andmanyof themitochondria appearedswollenanddamaged. These results suggest that neurotoxicity resulting fromACR treatment or feeding with FPC may contribute to weakenedlocomotion and other manifestations of central nervous abnormal-ities in the exposed young pups. In a previous report, pups mater-nally treatedwith ACR 5 to 20mg/kg per day from gestational day 6to lactation day 10 showed the characteristic hind-limb splay andsignificant decreases of average horizontal motor activity and theauditory startle response [30]. Another study reported that ratsinjected intraperitoneally with ACR (50 mg/kg per day) for 5d showed a significant inhibition of hepatic glutathione-50-trans-ferase activity, and maximum inhibition occurred in 15-d-old ratsthat exhibited early development of hind-limb paralysis [44].Anderson [45] reported a more variable nerve conduction velocityandsignificantbroadeningof themuscle compoundactionpotentialin rats treated with ACR at 25 to 100 mg/kg.
According to Miller et al. [14], ACR is largely excreted asmetabolites in the urine and bile. Two-thirds of the absorbeddose is excreted, with a half-life of a few hours. Specifically,

Fig. 6. Effects of ACR and FPC on the cerebellar cortex of 3-wk-old postnatal rat pups shown by transmission electron micrographs. (A–C) Control; (A1–C1) ACR-treatment;(A2–C2) FPC supplemented with food. Arrows indicate the differences, including atrophied nuclei, ruptured nuclear membrane, vesiculated rough endoplasmic, swelling ofmitochondria, elongated and hypertrophied Golgi apparatus. Magnifications 4500� in A–A2 and 10000� in B–C2. ACR, acrylamide; FPC, fried potato chips.
H. I. El-Sayyad et al. / Nutrition 27 (2011) 1066–10751072
protein-bound ACR or ACR metabolites in the blood andpossibly in the nervous systemhave a half-life of about 10 d. It isevident from these studies that regular consumption of ACR-containing FPC during pregnancy has adverse effects on fetalgrowth and early childhood and is deleterious to bone growth.Although much is known about relatively high-dose ACRexposure and its direct effects on the peripheral nervous
Fig. 7. Effects of acrylamide and fried potato chips on the gross morphology of Purkinacrylamide (B) or fed fried potatoes chips (C), the Purkinje cell exhibited a cell body with(A). Magnification 400�.
system, little is known about the developmental effects offeeding an ACR-containing diet. The main focus of this studywas to determine if ACR or FPC (containing ACR) affects thedevelopment of the cerebellar cortex and locomotion. Theresults of this study warrant a systematic study of the healtheffects of the consumption of FPC and French fries in the generalpopulation.
je cells in the cerebellar cortex of 3-wk-old rats. In pups maternally treated withan abnormal main axon devoid of dendrite formation compared with the controls

Fig. 8. Effects of ACR and FPC on the histology (hematoxylin and eosin staining, magnification 400�) of the gastrocnemius muscle of 1-d-old and 1-wk-old postnatal rats.(A, B) Control; (A1, B1) ACR treatment; (A2, B2) FPC supplementation. ACR, acrylamide; FPC, fried potato chips; NP, necrotic parts; RCI, round cell infiltration.
H. I. El-Sayyad et al. / Nutrition 27 (2011) 1066–1075 1073
Our findings clearly indicate that ACR administration andFPC feeding caused toxicity. ACR intoxicationmay be attributedto its greater solubility in water, facilitating ready absorptionthrough the oral cavity [28–30]. Also, in vitro studies haveshown that ACR can be absorbed through the skin [32].Absorbed ACR is then conjugated by glutathione-S-transferaseto N-acetyl-S-(3-amino-3-oxopropyl) cysteine or reacts withcytochrome P450 to produce glycinamide [28,33–35]. Glyci-namide formation occurs as a result of a marked inhibition ofglutathione-S-transferase by ACR [36,37]. Dixit et al. [46]reported that glutathione-S-transferase showed higher bin-ding capacity with ACR that was three times greater in the ratliver and kidney than in the brain; as a detoxifying enzyme, itsbinding to ACR or glycinamide may interfere with cell functionand promote cell death. The deleterious effects of ACR or itsmetabolite glycinamide seem to be attributed to their higherelectrophilic property owing to the presence of a- and b-un-saturated systems on its chemical structures, causing them toform adducts with sulfhydryl groups on hemoglobin and otherproteins [34,40]; thismay decrease the hemoglobin surface andthus its capacity to carry oxygen to the tissue, causing celldegeneration. Previous studies have detected DNA adducts in
the liver, lung, and kidney tissues in ACR-treated adult mice orrats [45,46].
In summary, as we recently reported, our experimentalmodel provides compelling evidence for the hazardous effectsof consumed food containing significant amounts of ACR,a proved rodent carcinogen, particularly FPC [47]. ACR islargely excreted as metabolites in the urine and bile, andtwo-thirds of the absorbed dose is excreted, with a half-lifeof a few hours [28,32]. However, ACR or ACR metabolitesare retained by proteins in the blood and possibly in thenervous system, extending its half-life to 10 d. The highestconcentrations of ACR have been identified in potatoes (ashigh as 3500 mg/kg in FPC and French fries) and grain-basedfoods that are cooked at very high temperatures [9]. In ourstudy, the hazardous effects were more pronounced with FPCcompared with ACR alone, suggesting that ACR might syn-ergize with other compound(s) from FPC to increase theeffects. Ongoing in vitro experiments aim to identify thesecompounds and better understand their mechanisms ofaction. Evidence from the present work and others suggestthat regular consumption of FPC during childhood could bedeleterious for regular bone growth. Similarly, regular

Fig. 9. Effects ofACRand FPCon thegastrocnemiusmuscleof a 3-wk-old rat revealedbytransmission electron microscopy. In the (B) ACR and (C) FPC groups, clear myodegen-eration anddramatic pathologic alterationswere detected,whichweremoreprominentinpups of rats fed FPC, comparedwith the (A) control group.Magnifications 4500� in A,10000� in B, 8000� in C. ACR, acrylamide; FPC, fried potato chips.
H. I. El-Sayyad et al. / Nutrition 27 (2011) 1066–10751074
consumption of FPC during pregnancy may cause fatalprenatal and/or postnatal abnormalities and malformation.
References
[1] Gold BG, Schaumberg S. Acrylamide. In: Spencer PS, Schaumberg HH,Ludolph AC, editors. Experimental and clinical neurotoxicology. 2nd ed.New York: Oxford University Press; 2000. p. 124–32.
[2] Tareke E, Rydberg P, Karlsson P. Analysis of acrylamide, a carcinogenformed in heated foodstuffs. J Agric Food Chem 2000;50:4998–5006.
[3] Tareke E, Rydberg P, Karlsson P, Eriksson S, Tornqvist M. ACR: a cookingcarcinogen? Chem Res Toxicol 2000;13:517–22.
[4] Smith CJ, Perfetti TA, Rumple MA, Rodman A, Doolittle DJ. IARC group 2Acarcinogens reported in cigarette mainstream smoke. Food Chem Toxicol2000;38:371–83.
[5] World Health Organization. Acrylamide, safety evaluation of certaincontaminants in food. WHO food additives series no. 55. Geneva: WorldHealth Organization; 2006. p. 1–156.
[6] Schettgen T, Kutting B, Hornig M, Beckmann MW, Weiss T, Drexler H,Angerer J. Trans-placental exposure of neonates to acrylamideda pilotstudy. Int Arch Occup Environ Health 2004;77:213–6.
[7] Dearfield KL, Abernathy CO, Ottley MS, Brantner JH, Hayes PF. Acrylamide:its metabolism, developmental and reproductive effects, genotoxicity, andcarcinogenicity. Mutat Res 1988;195:45–77.
[8] Sorgel F, Weissenbacher R, Kinzig-Schippers M, Hofmann A, Illauer M,Skott A, Landersdorfer C. Acrylamide: increased concentrations in home-made food and first evidence of its variable absorption from food, variablemetabolism and placental and breast milk transfer in humans. Chemo-therapy 2002;48:267–74.
[9] Takahashi M, Shibutani M, Nakahigashi J, Sakaguchi N, Inoue K,Morikawa T, et al. Limited lactational transfer of acrylamide to rat offspringon maternal oral administration during the gestation and lactation periods.Arch Toxicol 2009;83:785–93.
[10] Fohgelberg P, Rosen J, Hellenas KE, Abramsson-Zetterberg L. The acryl-amide intake via some common baby food for children in Sweden duringtheir first year of lifedan improved method for analysis of acrylamide.Food Chem Toxicol 2005;43:951–9.
[11] Erkeko�glu P, Baydar T. Toxicity of acrylamide and evaluation of its exposurein baby foods. Nutr Res Rev 2010;23:323–33.
[12] Garey J, Paule MG. Effects of chronic oral acrylamide exposure on incre-mental repeated acquisition (learning) task performance in Fischer 344rats. Neurotoxicol Teratol 2010;32:220–5.
[13] Edwards PM. The distribution and metabolism of ACR and its neurotoxicanalogues in rats. Biochem Pharmacol 1975;24:1277–82.
[14] Miller MJ, Carter DE, Sipes IG. Pharmacokinetics of ACR in Fisher-344 rats.Toxicol Appl Pharmacol 1982;63:36–44.
[15] Sickles DW, Sperry AO, Testino A, Friedman M. ACR effects on kinesin-related proteins of the mitotic/meiotic spindle. Toxicol Appl Pharmacol2007;222:111–21.
[16] LoPachin RM, Ross JF, Lehning EJ. Nerve terminals as the primary site ofACR action: a hypothesis. Neurotoxicology 2002;23:43–59.
[17] LoPachin RM, Ross JF, Reid ML, Das S, Mansukhani S, Lehning EJ. Neuro-logical evaluation of toxic axonopathies in rats: ACR and 2,5-hexanedione.Neurotoxicology 2002;23. 95–10.
[18] Lehning EJ, Balaban CD, Ross JF, LoPachi RM. ACR neuropathy. II. Spatio-temporal characteristics of nerve cell damage in brainstem and spinal cord.Neurotoxicology 2002;23:415–29.
[19] Lehning EJ, Balaban CD, Ross JF, Reid MA, LoPachin RM. ACR neuropathy. I.Spatiotemporal characteristics of nerve cell damage in rat cerebellum.Neurotoxicology 2002;23. 397–14.
[20] LoPachin RM, Balaban CD, Ross JF. Acrylamide axonopathy revisited. ToxicolAppl Pharmacol 2003;188:135–53.
[21] Sayre LM, Perry G, Smith MA. Oxidative stress and neurotoxicity. Chem ResToxicol 2008;21:172–88.
[22] Miller MS, Spencer PS. The mechanisms of acrylamide axonopathy. AnnuRev Pharmacol Toxicol 1985;25:643–66.
[23] Couraud JY, Di Giamberardino L, Chretien M, Souyri F, Fardeau M. Acrylamideneuropathy and changes in the axonal transport and muscular content of themolecular forms of acetylcholinesterase. Muscle Nerve 1982;5:302–12.
[24] Richard DH. The etiology of acrylamide neuropathy: enolase, phospho-fructokinase, and glyceraldehyde-3- phosphate dehydrogenase activities inperipheral nerve, spinal cord, brain, and skeletal muscle of acrylamide-intoxicated cats. Toxicol Appl Pharmacol 1981;60:324–33.
[25] Lehning EJ, Balaban CD, Ross JF, LoPachin RM. ACR neuropathy. III.Spatiotemporal characteristics of nerve cell damage in forebrain. Neuro-toxicology 2003;24:125–36.
[26] Palazo�glu TK, Savran D, Gökmen V. Effect of cooking method (bakingcompared with frying) on acrylamide level of potato chips. J Food Sci2010;75:E25–9.
[27] Kiernan JA. Histological and histochemical methods, theory and practice.1st ed. Oxford: Pergamon Press; 1981. p. 262–3.
[28] Barber DS, Stevens S, LoPachin RM. Proteomic analysis of rat striatalsynaptosomes during acrylamide intoxication at a low dose rate. Toxicol Sci2007;100:156–67.
[29] Kim K. Effect of subchronic acrylamide exposure on the expression ofneuronal and inducible nitric oxide synthase in rat brain. J Biochem MolToxicol 2005;19:162–8.
[30] Mottram DS, Wedzicha BL, Dodson AT. ACR is formed in the Maillardreaction. Nature 2002;419:448–9.
[31] Stadler RH, Blank I, Varga N, Robert F, Hau J, Guy PA, et al. ACR fromMaillard reaction products. Nature 2002;419:449–50.
[32] Becalski A, Lau BP, Lewis D, Seaman SW. ACR in foods: occurrence, sources,and modeling. J Agric Food Chem 2003;51:802–8.
[33] Friedman MA, Tyl RW, Marr MC, Myers CB, Gerling FS, Ross WP. Effects oflactational administration of ACR on rat dams and offspring. Reprod Toxicol1999;13:511–20.
[34] Garey J, Ferguson SA, Paule MG. Developmental and behavioral effects ofACR in Fischer 344 rats. Neurotoxicol Teratol 2005;27:553–63.

H. I. El-Sayyad et al. / Nutrition 27 (2011) 1066–1075 1075
[35] Wise LD, Gordon LR, Soper KA, Duchai DM, Morrissey RE. Developmentalneurotoxicity evaluation of ACR in Sprague-Dawley rats. NeurotoxicolTeratol 1995;17:189–98.
[36] Schettgen T, Kutting B, Hornig M, Beckmann MW, Weiss T, Drexler H,Angerer J. Trans-placental exposure of neonates to ACRda pilot study. IntArch Occup Environ Health 2004;77:213–6.
[37] LoPachin RM, Castiglia CM, Saubermann AJ. ACR disrupts elementalcomposition and water content of rat tibial nerve. I. Myelinated axons.Toxicol Appl Pharmacol 1992;115:21–34.
[38] Abou-Donia MB, Ibrahim SM, Corcoran JJ, Lack L, Friedman MA,Lapadula DM. Neurotoxicity of glycinamide, an ACR metabolite, followingintraperitoneal injections in rats. J Toxicol Environ Health 1993;39:447–64.
[39] Park HR, Kim MS, Kim SJ, Park M, Kong KH, Kim HS, et al. Acrylamideinduces cell death in neural progenitor cells and impairs hippocampalneurogenesis. Toxicol Lett 2010;193:86–93.
[40] Ali SF. ACR-induced changes in the monoamines and their acid metabolitesin different regions of the rat brain. Toxicol Lett 1983;17:101–5.
[41] LoPachin RM, Barber DS, He D, Das S. Acrylamide inhibits dopamine uptakein rat striatal synaptic vesicles. Toxicol Sci 2006;89:224–34.
[42] Agrawal AK, Seth PK, Squibb RE, Tilson HA, Uphouse LL, Bondy SC.Neurotransmitter receptors in brain regions of acrylamide-treated rats. I:effects of a single exposure to acrylamide. Pharmacol Biochem Behav1981;14:527–31.
[43] Bondy SC, Tilson HA, Agrawal AK. Neurotransmitter receptors in brainregions of acrylamide-treated rats. II: effects of extended exposure toacrylamide. Pharmacol Biochem Behav 1981;14:533–7.
[44] Kim K. Effect of subchronic acrylamide exposure on the expression ofneuronal and inducible nitric oxide synthase in rat brain. J Biochem MolToxicol 2005;19:162–8.
[45] Anderson RJ. Alterations in nerve and muscle compound action poten-tials after acute ACR administration. Environ Health Perspect1982;44:153–7.
[46] Dixit R, Husain R, Mukhtar H, Seth PK. ACR induced inhibition ofhepatic glutathione-S-transferase activity in rats. Toxicol Lett1981;7:207–10.
[47] El-Sayyad HI, Abou-Egla MH, El-Sayyad FI, El-Ghawet HA, Gaur RL,Fernando A, et al. Effects of fried potato chip supplementation on mousepregnancy and fetal development. Nutrition 2011;27:343–50.