sinking of heterosigma akashiwo results in increased toxicity of this harmful algal bloom species
TRANSCRIPT

Harmful Algae 13 (2012) 95–104
Sinking of Heterosigma akashiwo results in increased toxicity of this harmful algalbloom species
Lucas Powers a, Irena F. Creed a, Charles G. Trick a,b,*a Department of Biology, University of Western Ontario, London, Ontario N6A 5B7, Canadab Department of Microbiology and Immunology, Schulich School of Medicine and Dentistry, University of Western Ontario, London, Ontario N6A 5B7, Canada
A R T I C L E I N F O
Article history:
Received 7 June 2010
Received in revised form 21 September 2011
Accepted 12 October 2011
Available online 22 October 2011
Keywords:
Heterosigma akashiwo
Sinking
Bioassays
Toxicity
Bloom propagation
A B S T R A C T
Notable physiological responses such as toxicity and sinking rates of the red tide forming raphidophyte
Heterosigma akashiwo are correlated with high levels of macronutrient stress. Individual cells of this
species are also capable of forming benthic vegetative cysts that overwinter in marine sediment and
contribute to bloom propagation in subsequent seasons. It was hypothesized that there is variability in
the rates of sinking within cell cultures and that sinking cells are more toxic than the neutrally buoyant or
floating cells. Using laboratory-based settling columns, various isolates of H. akashiwo were allowed to
separate, and the toxicities of sinking and floating populations were analyzed. Sinking and floating rates
were significantly higher during the late stationary growth phase for all isolates. For two H. akashiwo
isolates, sinking populations were significantly more toxic than those that were positively buoyant. A
similar trend was observed in a third strain, however the relationship was not significant. Differences in
adaptive ecophysiology among the different strain likely caused the variation. It is suggested that the
most toxic cells within a bloom are those found at the lower depths, potentially interacting with the
benthic community or ensuring that subsequent bloom propagation contains cells with the potential for
toxicity.
� 2011 Elsevier B.V. All rights reserved.
Contents lists available at SciVerse ScienceDirect
Harmful Algae
jo u rn al h om epag e: ww w.els evier .c o m/lo cat e/ha l
1. Introduction
The global prevalence and severity of harmful algal blooms(HABs) appears to be increasing in many marine ecosystems(Smayda, 1989; Horner et al., 1997; Anderson et al., 2002; Heisleret al., 2008). HABs are characterized by the dominance of a singlephytoplankton species, eventually forming dense concentrationsof algal biomass that threaten the health of the ecosystem by anumber of mechanisms. It remains unclear, however, preciselywhat environmental parameters mediate bloom formation andwhat toxicological mechanisms HAB species employ that result inthese harmful effects (Anderson et al., 2002; Hallegraeff and Hara,2003), with considerable variation among the relatively smallpercentage of algal genera capable of forming HABs (Morris, 1999;Landsberg, 2002).
Generally, potentially harmful species can affect a communityin one of the following ways (Smayda, 1997a,b; Hallegraeff andHara, 2003): (i) causing significant water discoloration due to
* Corresponding author at: Department of Biology, University of Western
Ontario, London, Ontario N6A 5B7, Canada. Tel.: +1 519 661 3899;
fax: +1 519 850 2343.
E-mail addresses: [email protected] (L. Powers), [email protected] (I.F. Creed),
[email protected] (C.G. Trick).
1568-9883/$ – see front matter � 2011 Elsevier B.V. All rights reserved.
doi:10.1016/j.hal.2011.10.007
extremely dense cellular aggregations, which creates a physicalbarrier to light penetration that subsequently results in anoxicconditions as blooms dissipate; (ii) producing potent toxins thatbioaccumulate and are known to be directly harmful to humans;and (iii) demonstrating allelopathic and anti-biological strategiesthat can induce high mortality in a wide range of aquaticorganisms, particularly fish, with devastating effects on aquacul-ture (Honjo, 1993). Considerable debate surrounds species in thisthird category, as no single process or mechanism has beenconclusively identified to explain how these species achieveichthyotoxicity.
A phytoplankton species of particular concern within this thirdcategory is the red-tide forming alga, Heterosigma akashiwo (Hada)(Hara and Chihara, 1987). Coastal blooms of this fish-killingraphidophyte have been observed in both the Atlantic and PacificOceans and have been implicated in fish-kills in aquacultureoperations in Canada, Chile, Japan and New Zealand (Hallegraeff andHara, 2003). Additionally, severe economic losses associated withfin-fish mortality attributed to H. akashiwo have been reported inJapan (Honjo, 1993), British Colombia, Canada (Taylor, 1991; Taylorand Haigh, 1993), Washington State, USA (Connell and Cattolico,1996), and New Zealand (Chang et al., 1990). More recently, bloomsof H. akashiwo have been reported in San Francisco Bay, USA(Herndon and Cochlan, 2006) and the inner waters of BritishColombia and Washington State (Horner et al., 1997).

L. Powers et al. / Harmful Algae 13 (2012) 95–10496
Although a global problem, the mechanism of H. akashiwo
toxicity has yet to be determined. Because H. akashiwo is capable ofharming a wide spectrum of marine organisms such as zooplank-ton, copepods, benthic larvae and fish, the toxin(s) or mechanismsresponsible for its potent toxicity remain elusive. Studies haveattributed the toxicity associated with raphidophytes to theproduction of two reactive oxygen species (ROS), superoxide (O2
��)(Oda et al., 1998; Marshall et al., 2005) and hydrogen peroxide(H2O2) (Twiner and Trick, 2000; Twiner, 2002); haemolyticcompounds, which lyse red blood cells (de Boer et al., 2004; Fuet al., 2004; Ling and Trick, 2010); and a brevetoxin-like organicneurotoxin (Khan et al., 2008). However, none of these putativetoxic agents have been independently linked to the ichthyotoxicityof H. akashiwo. Finfish may not be the target of H. akashiwo toxicity,but rather they may be negatively affected by compoundsproduced by H. akashiwo as a means of bloom formation,continuance or maintenance.
Blooms of raphidophyte HAB species, such as Heterosigma, butalso Chattonella, often appear to be ephemeral, because the cellsdisperse or concentrate to the highly visible surface bloom(Hershberger et al., 1997). The dynamics of this verticalconcentration is often critical to the energetic and nutrient needsof the migrating cells and these dynamics have drawn considerableresearch attention. From early studies (Smayda, 1970, 1997a) it hasbecome established that flagellated cells such as H. akashiwo canregulate its relative position via both active and passive locomo-tion processes to optimize access to resources such as nutrientsand light, a critical feature in photoreceptive strategy.
Resource availability also regulates the formation of cysts, a lifecycle stage that is relatively rare among HAB species (Smayda,1997b). Cysts generally form after the dissipation of high biomassblooms, when nutrient stress becomes physiologically over-whelming for the cells or cooler temperatures signal the imminentonset of winter conditions. Vegetative cysts are extremely hardyand are capable of overwintering in the benthic sediment andreemerging as reproductive cells during the following spring andsummer seasons (Tomas, 1978). H. akashiwo cysts sink rapidly tothe sediment and are thus less likely to senesce due toenvironmental stresses or are consumed by an algal grazer beforedepositing into the benthic ecosystem (Smayda, 1990, 1998).Furthermore, cells that do re-emerge after overwintering can alsobe toxic towards benthic algal grazers, and thus the potentialeffects of H. akashiwo are not limited to the pelagic environment(Tomas and Deason, 1981).
Not all blooms of H. akashiwo are toxic and not all toxic bloomsremain toxic at all times; blooms may exhibit traits of potenttoxicity or remain relatively harmless in a given year (Black,2000). H. akashiwo grown in nutrient-limited conditions generallyexhibits greater toxicity due to changes in physiological dynamicscaused by the stress of severe nutrient depletion over time. Thechange in toxicity is often related to the changing nutrientenvironment, with nutrient stressed cells being more toxic thanthe nutrient replete cells in a population (Honjo, 1993). In the lifecycle of the bloom there are complex simultaneous activities(Smayda, 1997a). For example, the accumulation of biomasseventually becomes self-shading and/or nutrient limiting, andthus individuals may then employ variable strategies to cope withthese stresses. The result is an accumulation of cells in the surfacewaters with other individuals sinking to the benthos. How anindividual cell behaves will depend on the greatest stress to thatparticular cell and where nutrients are within the water column.These are not independent because nutrient-limited cells oftensink within the water column (Bienfang et al., 1982; Lecourt et al.,1996; Muggli et al., 1996). This adds a spatial element whenconsidering the dynamics of cells and toxins in a harmful algalbloom.
If resource conditions, in particular the availability of macro-nutrients, positively affect both the sinking rates and potentialtoxicity of a cell, then it is likely that the most highly toxic cells willbe found lower in the water column because both are commonphytoplankton responses to macronutrient stress. There arenotable ecological and practical consequences associated withthis possibility. If more highly toxic cells migrate to lower depths, itincreases the possibility that these cells will aggregate andoverwinter in sediment in a vegetative state (Kiorboe et al.,1994). Individuals that survive in marine sediment seed thefollowing year’s potential bloom, and thus if highly toxic cells sinkand overwinter preferentially, then regions may become moreprone to toxic blooms over time. The purpose of this research wasto establish the association between cellular nutrient status,sinking and toxicity. Specifically, under what conditions and atwhat point in its cellular life cycle is Heterosigma most prone tosinking? Additionally, what is the toxic potential of cells undervarious nutrient stresses, and how is toxicity related to thephysiological responses of sinking and floating? This knowledge isvital in establishing an increased predictive power of toxic bloomformation and also in understanding the spatial dynamics of toxicblooms in natural environments. In a broad sense, the ultimategoal of this HAB research is to develop methodologies foreffectively mediating the harmful effects of H. akashiwo andpossibly preventing their proliferation in marine coastal waters.
2. Materials and methods
2.1. Phytoplankton cultures and nutrient conditions
All phytoplankton strains were non-axenic and grown in batchcultures in autoclave-sterilized, modified ESAW medium (Harrisonet al., 1980). A 10% (v/v) inoculum of cells was added to 45 mL ofmedium to a final volume of 50 mL in autoclaved 250 mLErlenmeyer flasks. Cells were acclimated to the experimentalN:P ratio for two generations under constant temperature (18 8C)and irradiance (160 mmol photons m�1 s�1) prior to the start of theexperiment (experimental T0). Since light:dark cycles could inducea diel migration cycle, all cultures were maintained under aconstant light flux of 160 mmol photons m�1 s�1. Balanced growthconditions, representative of nitrate to phosphate ratios found innatural marine ecosystems not under environmental stress, werecharacterized by a 15:1 nitrate to phosphate molar ratio in themodified ESAW media. Phosphate-limited media were character-ized by a 50:1 nitrate to phosphate molar ratio, achieved byincreasing the concentration of nitrate in ESAW media. Nitrate-limited media contained a molar nitrate to phosphate ratio of 5:1.Three strains of H. akashiwo, strain Can 764R (Northeast PacificCulture Collection), strain ‘‘Can Ed’’ (from coastal British Columbiawaters, isolated by Dr. Ed Black, Department of Fisheries andOceans, Canada) and strain ‘‘Spain’’ (isolated by A. Butron fromwaters of the Basque Region, Spain in 2002), were compared in thisstudy. Tetraselmis apiculata (UTEX 2562), a strain commonly usedas a food source in aquaculture facilities and considered to be non-toxic, was used as a control species, given that it is the sameapproximate size and shape of the Heterosigma isolates.
2.2. Culture growth and growth phases
Beginning on the day of inoculation, all cultures were sampledevery other day to monitor cell growth. A 0.5 mL subsample ofculture sample was pipetted into 8 mL FloTubesTM in a sterilelaminar flow hood and analyzed for cell counts on a FACSCaliburflow cytometer [B-D Biosciences]. Samples were run for oneminute using a previously formatted marine phytoplanktontemplate. Monitoring continued until cell counts ceased to

L. Powers et al. / Harmful Algae 13 (2012) 95–104 97
increase for at least two or at most three samplings. After at leasttwo samplings with no observed increase, the culture wasconsidered to be in late stationary growth phase (after approxi-mately 14–16 days). Early stationary phase was delineated as thefirst day that exponential growth of cultures was observed to stop(after approximately 10–12 days). Exponential growth phase wascharacterized by exponential increases in cell numbers (approxi-mately 8–10 days).
2.3. Column fractionations
Settling column experiments were conducted during theexponential growth phase, early stationary growth phase, andlate stationary growth phase. Once in the desired growth phase,quadruplicate flasks of 50 mL were pooled together into a single250 mL Erlenmeyer flask for each culture. Culture (150 mL) wasthen poured into a corresponding borosilicate settling column,each 53 cm high, 3.5 cm in diameter with a 250 mL total capacity.Each column was covered in industrial grade aluminum foil so thatno light penetrated the columns. Separations were conducted for a6-h period, in darkness and at a constant temperature of 18 8C.
After each 6-h-settling period, three distinct fractions of thecolumn sample were removed. The top 25 mL was removed usinggentle aspiration from above, collecting all media to a prescribedmark. The top sample was representative of the floating cellfraction. The bottom 25 mL of the undisturbed column, represen-tative of the sinking cell fraction, was slowly decanted from abottom drain. The intermediate fraction consisted of the remaining100 mL and was representative of cells without a tendency to floator sink. Once all fractions were separated, 0.5 mL subsamples wereremoved from each fraction for cell counts. Additionally, 15 mLsubsamples were collected from each respective fraction to befiltered for chlorophyll-a based biomass analysis.
2.4. Biomass measurements
The total biomass of each fraction for all isolates duringexperiments was quantified using an extracted chlorophyllmethod (Parsons et al., 1984). Exactly 15 mL of fraction sampleswere filtered over 25 mm diameter, 0.4 mm pore polycarbonatefilters using a vacuum filtering apparatus. Filters were then placedin 90% acetone and transferred to a �4 8C freezer for 24 h. After thatperiod, filters were removed from the acetone using forceps andthe remaining solution was analyzed on a Turner-10AU Fluorom-eter according to the protocol outlined in Welschmeyer (1994).Raw fluorescence output was transformed into chlorophyll-a
concentrations (mg mL�1) using a standard curve generated byanalyzing spinach extracts of known concentrations on aspectrophotometer at wavelengths of 630 nm, 647 nm, 664 nmand 750 nm.
2.5. Floating and sinking rates
Floating and sinking rates of populations are mechanisticallydefined and cannot be directly related to the physiologicalmotivation of the cells. For example, cells expressing ascendingvertical migration would be pooled with cells that are non-committal to migration but more buoyant than the medium.Similarly, cells expressing descending vertical migration would bepooled with dense cells accumulated through passive sinking. Forthese reasons, the relative distribution differences in the stableexperimental column was used to define the floating and sinkingrates of the populations within each treatment. Relative floatingand sinking rates for each culture were calculated using a modifiedSETCOL method (Bienfang et al., 1982). Once the biomass of eachfraction was known, floating (A) and sinking (c) rates were
generated using the following equation:
A or c ¼BfracðobsÞ � BfracðpredÞ
Btot
h
t
� �(1)
where Bfrac(obs) is the observed biomass in the top (floating) orbottom (sinking) fractions, Bfrac(pred) is the predicted biomass of thefraction based on the sum of biomass in the column multiplied bythe fraction volume relative to the total volume; h is the height ofthe column of water, and t is the incubation time in hours.
2.6. Toxicity bioassay
Each sinking cell and floating cell fraction from each columnwas used in a standard bioassay to assess the relative toxicity ofcells found within the fractions. A sample of cells from eachfraction was standardized to a concentration of 15,000 cells mL�1.A standardized sample (1 mL) was added to each well on a 24-welltray and left to incubate with ten young (1-day old) or ten mature(2-days old) brine shrimp per well (Artemia salina) (Vanhaeckeet al., 1981). Every fraction was assayed with both young andmature brine shrimp independently. Addition of HCl (20%, v/v) wasused as a positive control, and these samples were incubated withbrine shrimp under identical conditions to culture samples.Autoclave-sterilized modified ESAW media (salt solution only,no macronutrients or trace elements added) were used as negativecontrols and were treated identically to other treatments. After theinitial 24-h incubation, brine shrimp were observed under amicroscope and classified as dead, struggling, or alive. Followingthe initial count, brine shrimp were incubated for a subsequent 24-h period and dead, struggling and healthy brine shrimp weretallied again. Struggling individuals were readily distinguishablefrom dead and healthy brine shrimp; struggling individuals lackedcoordination in movement and/or feeding. Extended observations(72 h) indicated that struggling individuals regained a normalmotility behavior, rather than succumb to the toxicity of theassessment.
2.7. Statistical analysis
All data were analyzed using SPSS 17.0 statistical software.Floating and sinking rates were compared across growth phasesand between nutrient treatments using one-way ANOVAs. Allbrine shrimp bioassay mortality data were analyzed using one-and two-way ANOVAs and were checked for homogeneity andindependent factor interactions. An alpha of 0.05 was used in allstatistical analyses. Toxicity bioassay results were computed andcompared between isolates using values that combined the totalnumber of brine shrimp observed to be dead and the total numberobserved to be struggling. These ‘‘dead + struggling’’ valuespermitted greater statistical power and increased relative toxicitydifferences among isolates. Means are reported �1 SD.
3. Results
3.1. Floating and sinking rates
The fractionation of the culture into sinking and floating sub-populations occurred in all strains studied. Even though H.
akashiwo is recognized as a surface forming bloom species thatforms sinking cysts (Honjo, 1993), the sinking and floating rates ofthe comparison flagellate (T. apiculata strain 2562.) equaled orexceeded that of H. akashiwo strains. Under these conditions, thephysiological or ecological importance of sinking and/or floating toH. akashiwo was not distinctive.
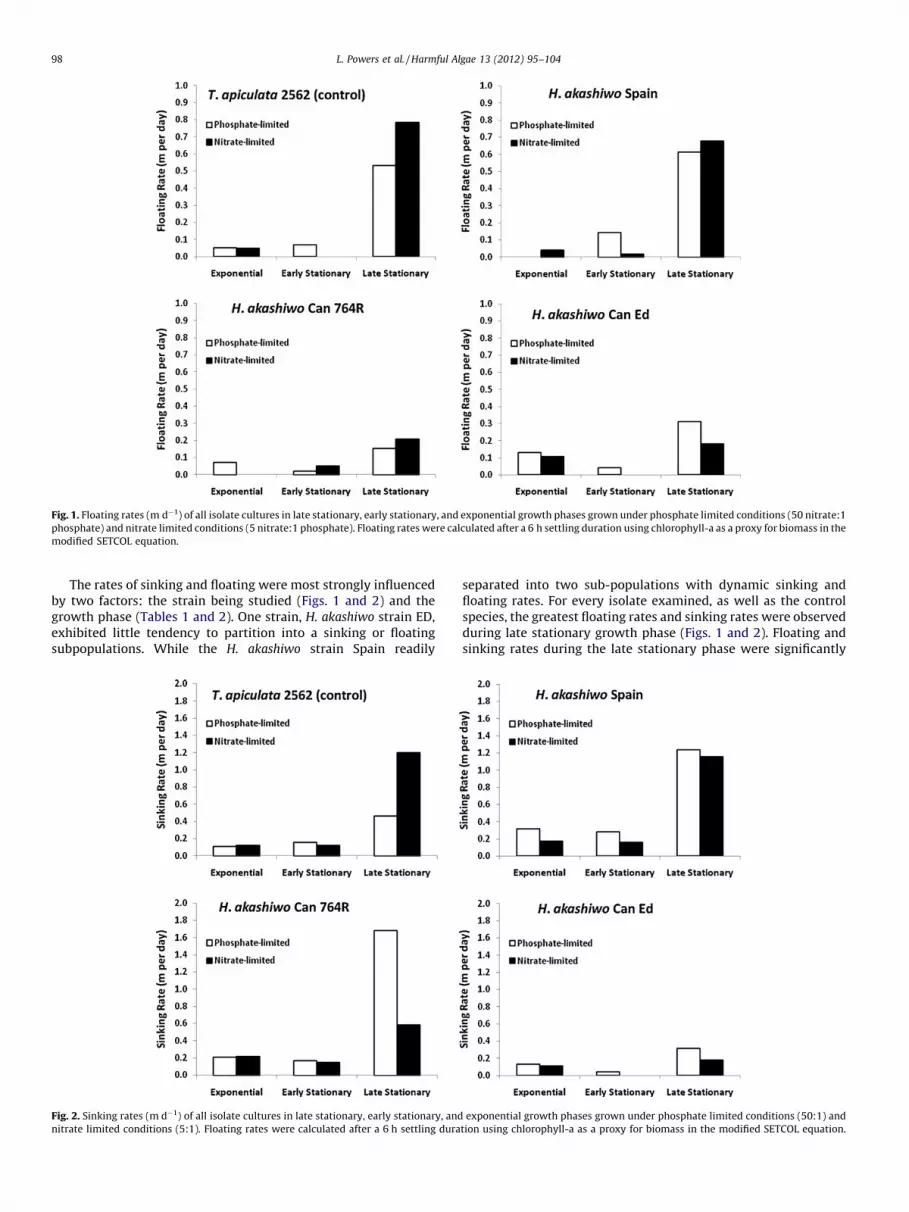
Fig. 1. Floating rates (m d�1) of all isolate cultures in late stationary, early stationary, and exponential growth phases grown under phosphate limited conditions (50 nitrate:1
phosphate) and nitrate limited conditions (5 nitrate:1 phosphate). Floating rates were calculated after a 6 h settling duration using chlorophyll-a as a proxy for biomass in the
modified SETCOL equation.
L. Powers et al. / Harmful Algae 13 (2012) 95–10498
The rates of sinking and floating were most strongly influencedby two factors: the strain being studied (Figs. 1 and 2) and thegrowth phase (Tables 1 and 2). One strain, H. akashiwo strain ED,exhibited little tendency to partition into a sinking or floatingsubpopulations. While the H. akashiwo strain Spain readily
Fig. 2. Sinking rates (m d�1) of all isolate cultures in late stationary, early stationary, and
nitrate limited conditions (5:1). Floating rates were calculated after a 6 h settling dura
separated into two sub-populations with dynamic sinking andfloating rates. For every isolate examined, as well as the controlspecies, the greatest floating rates and sinking rates were observedduring late stationary growth phase (Figs. 1 and 2). Floating andsinking rates during the late stationary phase were significantly
exponential growth phases grown under phosphate limited conditions (50:1) and
tion using chlorophyll-a as a proxy for biomass in the modified SETCOL equation.

Table 1Median floating rates of all three H. akashiwo isolates and T. apiculata 2562 (m d�1) (�25% quartile values). Sinking rates denoted with the same lower case letter were not
significantly different from each other. Median sinking rates were calculated by determining the median values for all isolates during each respective growth phase.
Growth phase Median floating rate (m d�1) 25% quartile value 75% quartile value p-Value
Late stationary 0.422 a 0.195 0.645 p < 0.05
Early stationary 0.033 b 0.008 0.060 p > 0.05
Exponential 0.049 b 0.019 0.090
Mean values shown with the same lower-case letter were not significantly different.
Table 2Median sinking rates of all three H. akashiwo isolates and T. apiculata 2562 (m d�1) (�25% quartile values). Sinking rates denoted with the same lower case letter were not
significantly different from each other. Median sinking rates were calculated by determining the median values for all isolates during each respective growth phase.
Growth phase Median Sinking rate (m d�1) 25% quartile value 75% quartile value p-Value
Late stationary 0.871 a 0.553 1.218 p < 0.05
Early stationary 0.159 b 0.140 0.215 p > 0.05
Exponential 0.174 b 0.174 0.137
Mean values shown with the same lower-case letter were not significantly different.
L. Powers et al. / Harmful Algae 13 (2012) 95–104 99
different from those observed in both the exponential growthphase and early stationary growth phase for each isolate of H.
akashiwo and T. apiculata 2562. Within the late stationary growthphase, nutrient regime (phosphorus (P)-limited, 50 nitrate:1phosphate; and nitrogen (N)-limited, 5 nitrate:1 phosphate) didnot significantly affect floating and sinking rates for most of the H.
akashiwo isolates or the control species (p > 0.05). The oneexception was H. akashiwo strain Can 764R where the sinkingrate of P-limited, late stationary cells was about 3� more rapidthan similarly aged N-limited cells. Even with the large variation inthe strain dynamics, within any one isolate the sinking rate wasconsistently in the order of 2� faster than the floating rates.
3.2. Toxicity bioassays
Each strain of H. akashiwo was significantly more toxic to A.
salina than the control strain T. apiculata 2562 at all nutrientconditions and during all growth phases (p < 0.05) (Table 3). T.
apiculata 2562 showed no indication of toxicity towards A. salina.Additionally, there was no significant difference between therelative toxicities of each H. akashiwo strain (p > 0.05). Likewise, nosignificant difference in toxicity was observed between P-limitedand N-limited conditions for all three H. akashiwo strains(p > 0.05).
Table 3Mean number of affected brine shrimp (Artemia salina) for floating and sinking fractions
stationary growth phase (�SD).
Isolate Nitrate to phosphate ratio Fraction Me
Can Ed 50:1 Floating 0.8
Sinking 2.3
5:1 Floating 2.1
Sinking 3.6
Spain 50:1 Floating 3.1
Sinking 4.4
5:1 Floating 2.9
Sinking 4.3
Can 764R 50:1 Floating 3.5
Sinking 3.5
5:1 Floating 2.9
Sinking 4.1
T. apiculata 2562 50:1 Floating 0.0
Sinking 0.1
5:1 Floating 0.0
Sinking 0.0
Mean values shown with the same lower-case letter within each N:P ratio were not si
The sinking population of H. akashiwo strain Can Ed wassignificantly more toxic than floating population under bothnutrient conditions in late stationary growth phase (p < 0.001).There was also a significant difference in the toxicity of H. akashiwo
strain Can Ed between respective nutrient conditions (p < 0.001).Floating populations of H. akashiwo strain Can Ed grown under P-limited conditions (50 nitrate:1 phosphate) displayed the leasttoxicity of any isolate and fraction during late stationary growthphase, negatively affecting (mortality + struggling individuals)only 8.8(�7.2)% of brine shrimp.
The response of H. akashiwo strain Spain was more variable,with a significant difference in relative toxicity between floatingpopulations and sinking populations within both nutrient condi-tions (p < 0.001), but no significant difference between P-limitedand N-limited conditions (p = 0.678). Sinking populations of H.
akashiwo strain Spain grown under N-limited conditions displayedthe greatest toxicity of any isolate and fraction during latestationary growth phase, negatively affecting 43.1(�16.7)%.
Further, no significant differences in relative toxicity betweenfloating and sinking populations were found for H. akashiwo strainCan 764R within each nutrient condition (p = 0.134), nor betweenrespective nutrient conditions (p = 0.996). Although differenceswere not significant, there was a clear trend of sinking populationsbeing more toxic than floating populations grown at N-limited
of all isolates of H. akashiwo and the biological control T. apiculata 2562 during late
an dead + struggling A. salina per 10 individuals �SD p-Value
8 a 0.72 <0.001
1 b 1.45
2 a 1.21 <0.001
0 b 1.40
3 a 1.54 <0.001
0 b 1.54
0 a 1.21 <0.001
1 b 1.67
0 a 1.60 0.134
0 a 1.31
4 a 1.39 0.134
3 a 1.54
6 a 0.12 0.557
3 a 0.34
0 a 0.00 0.557
0 a 0.00
gnificantly different.

Fig. 3. Toxicity bioassay results for H. akashiwo Ed during exponential, early, and
late stationary growth phases shown as a percentage of total brine shrimp (�SD).
‘‘Upper’’ denotes floating cell fraction, while ‘Lower’ denotes sinking cell fraction.
Mortalities denoted with the same lower case letter were not significantly different.
Brine shrimp were tallied as dead, struggling or alive after 24 h, and again after 48 h.
Results shown here represent a 48-h incubation, and SD was calculated by combining
‘Dead’ and ‘Struggling’ into a single value.
Fig. 4. Toxicity bioassay results for H. akashiwo Spain 2002 during exponential,
early, and late stationary growth phases shown as a percentage of total brine shrimp
(�SD). ‘‘Upper’’ denotes floating cell fraction, while ‘Lower’ denotes sinking cell
fraction. Mortalities denoted with the same lower case letter were not significantly
different. Brine shrimp were tallied as dead, struggling or alive after 24 h, and again
after 48 h. Results shown here represent a 48-h incubation, and SD was calculated by
combining ‘Dead’ and ‘Struggling’ into a single value.
L. Powers et al. / Harmful Algae 13 (2012) 95–104100
conditions; however, this trend was not clear at P-limitedconditions (Figs. 3–6).
No significant interactions between nutrient condition andfraction (sinking/floating populations) were observed for anyisolate of H. akashiwo or T. apiculata 2562 during the late stationarygrowth phase (p < 0.05). Further, no significant differences werefound in the number of affected brine shrimp between young (48-hold) and juvenile (24-h old) (p < 0.05). Likewise, length ofincubation time of brine shrimp with H. akashiwo isolates hadno significant effect on the number of affected brine shrimp forboth juvenile and mature brine shrimp bioassays.
All three strains of H. akashiwo displayed their highest toxicityduring the transition period between nutrient-replete exponential
growth and the early stage of stationary growth. In none of theisolates did toxicity increase with prolonged nutrient starvation. H.
akashiwo strain Can Ed was most toxic during the early stationaryphase under N-limiting conditions. H. akashiwo strain Spainachieved its most potent toxicity during the exponential growthphase, also grown under nitrate-limited conditions. Finally, H.
akashiwo strain Can 764R displayed its highest toxicity during theexponential phase under N-limited conditions.
4. Discussion
The fundamental purpose of this research was to examine howgrowth phase and nutrient conditions affect the floating and

Fig. 5. Toxicity bioassay results for H. akashiwo 764R during exponential, early, and
late stationary growth phases shown as a percentage of total brine shrimp (�SD).
‘‘Upper’’ denotes floating cell fraction, while ‘Lower’ denotes sinking cell fraction.
Mortalities denoted with the same lower case letter were not significantly different.
Brine shrimp were tallied as dead, struggling or alive after 24 h, and again after 48 h.
Results shown here represent a 48-h incubation, and SD was calculated by combining
‘Dead’ and ‘Struggling’ into a single value.
Fig. 6. Toxicity bioassay results for T. apiculata 2562 during exponential, early, and
late stationary growth phases shown as a percentage of total brine shrimp (�SD).
‘‘Upper’’ denotes floating cell fraction, while ‘Lower’ denotes sinking cell fraction.
Mortalities denoted with the same lower case letter were not significantly different.
Brine shrimp were tallied as dead, struggling or alive after 24 h, and again after 48 h.
Results shown here represent a 48-h incubation, and SD was calculated by combining
‘Dead’ and ‘Struggling’ into a single value.
L. Powers et al. / Harmful Algae 13 (2012) 95–104 101
sinking behavior of H. akashiwo. It is congruent with the H.
akashiwo cellular life cycle that floating and sinking rates weregenerally greatest during late stationary growth phase. Under latestationary conditions the sinking rate predominated at about 2-times the floating rate. Yet, ascending cells were also enhancedduring this nutrient-stressed phase. Motility exhibited in flagellateHAB species is a measurable response that results from a complexinteraction between physiological processes intended to maximizelight receptivity and nutrient uptake and external stresses appliedby the physical environment (Watanabe et al., 1988; Kamykowskiet al., 1992). An increase in motility has been correlated withincreasing nutrient depletion as resources become more limited.In the late stationary growth phase, natural phytoplankton
communities and laboratory cultures have metabolized all ofthe nutrients that facilitated exponential growth. Thus cells at thisphase have (i) been exposed to a nutrient-limited environment fora notable duration of time and (ii) stopped allocating metabolicenergy resources to cellular growth and will subsequently allocatenutrient reserves to cell maintenance. Severe physiological stressand the potential to allocate remaining energetic complexes tomotility in order to escape that physiological stress in the watercolumn can explain why sinking rates were greatest and about 2�floating rates in the late stationary phase (Graneli and Johansson,2003), but floating rates were also significantly higher in the latestationary phase when compared to either of the two previousgrowth phases (Smayda, 1997b).

L. Powers et al. / Harmful Algae 13 (2012) 95–104102
Floating and sinking rates observed during column fractiona-tions for H. akashiwo isolates ranged from about 0.2 to 1.9 m d�1.There is scarce literature directly reporting settling rates in thespecies and isolates examined. Some studies have focused on thesettling dynamics of diatom species, where reported settling ratesare generally higher, likely due to the fact that individual cells inmany diatom species are often quite large, ranging from 2 to200 mm, and have highly dense silica frustules. In comparison, the‘‘cell wall-free’’ H. akashiwo reaches a maximum size of �15–25 mm (Smayda, 1970). Bienfang (1980) found that sinking rates inopen ocean phytoplankton communities were significantly higherwhen those communities consisted of individuals >20 mm inlength, as opposed to communities consisting of small individuals3–20 mm in length. Based on these previous studies, the floatingand sinking rates seen during this experiment appear to representrealistic rates for natural H. akashiwo communities. However,maximum floating and sinking rates, however, did not correlatewith the periods of greatest relative toxicity. In most cases, thecells were most toxic against brine shrimp during the activelygrowing or early stationary phases.
During these earlier life cycle phases, the characteristics ofH. akashiwo toxicity were revealed in the A. salina assay. Incontrast to other marine biotoxins [e.g., domoic acid, (Bates andTrainer, 2006)], cell toxicity decreased as cells underwentnutrient starvation. This may indicate that the toxicity isnot an accumulated biomolecule, but rather a physiologicalmechanism.
There was no significant difference between the mortalityexperienced by young versus mature brine shrimp. Furthermore,the lack of significant difference between the 24 and 48 hexposures suggests several characteristics of the toxic mechanismin H. akashiwo. First, the young brine shrimp are still physicallyattached to a nutrient-rich yolk that sustains cysts duringdormancy, thus the brine shrimp at this age are not actuallyingesting food particles from the external environment. Maturebrine shrimp, however, have shed this yolk and will actively grazeon algae adjacent in the water column (Vanhaecke et al., 1981).Ingestion may not result in increased mortality, and thus the toxiccompounds being produced by H. akashiwo may be actingexternally on affected organisms (Honjo, 1993; Khan et al.,2008). Second, mortality did not significantly increase after asecond 24 h period (48 h total), supporting the hypothesis that anypotential H. akashiwo toxins are highly labile and will not continueto affect other organisms for a notable period of time after initialexposure (Graneli and Hansen, 2006). Third, the struggling brineshrimp recovered after 72 h. These results contrast with HABspecies that produce known toxic compounds, such as thedinoflagellate Karenia mikimotoi, which has been shown to causemortality for several days in bioassay experiments.
All H. akashiwo isolates were most toxic to A. salina at somepoint prior to the late stationary growth phase, and there was aclear trend towards greater toxicity under N-limited conditions.At first this seems counter-intuitive because it was previouslystated that: (i) cellular physiological stress is induced by nutrientlimitation, (ii) increased stress likely increases the toxic proper-ties of individual cells, and (iii) stress is highest in the latestationary phase. Thus, it was originally hypothesized thattoxicity would likely be highest in the late stationary growthphase as well. However, recent insights into the potentialmechanisms of toxicity in H. akashiwo suggest that toxicitymay be an indirect byproduct of hemolytic iron scavenging (Lingand Trick, 2010). Iron is an essential cofactor in cellular enzymesthat regulate the uptake of nitrogen from the environment, andthus nitrogen limitation for phytoplankton communities may bethe result of iron depletion (Utkilen and Gjolme, 1995). Hemolyticscavenging may be indicative of increased scavenging for peptides
to increase the potential N-pool in environments with limitedsupply of inorganic-N.
This model of toxicity is supported by the finding that the N-limited condition (5 nitrate:1 phosphate) was significantly moretoxic than the phosphate (P)-limited condition (50 nitrate:1phosphate) during the growth phase of highest overall toxicity forH. akashiwo strains Can Ed and Can 764R (early stationary andexponential, respectively). In contrast, the toxicity of H. akashiwo
strain Spain was more affected by P-limitation, illustrating thatclonal variation is not uncommon in H. akashiwo studies, and thisresult is consistent with results reported by Furnas (1990) andMartinez et al. (2010).
If hemolytic scavenging is the primary mechanism of toxicity inH. akashiwo, it is likely that relative toxicities were greatest duringexponential and early stationary growth phases because cells areallocating the majority of energy resources to cell growth andreproduction during the exponential phase. These processes haveonly recently slowed or ceased during the early stationary phase,with some cells still reproducing, thus individuals will attempt tomaximize nutrient uptake and also have the cellular resources toactively compete for limited nutrients. Additionally, cells may beoptimizing the intracellular storage of nutrients before nutrientavailability becomes so scarce that such processes becomeenergetically inefficient (Watanabe et al., 1988). Further, whencells enter the late stationary growth phase as nutrients become soreplete that energetically expensive processes like hemolyticscavenging are no longer worth the metabolic requirements,individual cells will yield metabolic activity and use stored energymolecules to actively pursue nutrients and light elsewhere in thewater column; in other words they increase motility (Smayda,1997b).
The variability between H. akashiwo isolates observed shouldserve as a cautionary note. The plasticity of toxic responses meansthat attempting to generalize the ecophysiology of H. akashiwo inorder to construct a more comprehensive framework of bloomdynamics may be an impractical task. The behavior and toxicity ofindividuals within a particular bloom will be highly dependentupon the isolate(s) that dominate within the bloom. Subsequently,research should focus on common regional H. akashiwo isolates todevelop an understanding at more local scales, and researchersshould be hesitant to apply the lessons learned from a particularisolate to the bloom dynamics of another.
There was a significant difference between floating and sinkingfractions for H. akashiwo strains Can Ed and Spain, and although thedifference was not significant for strain Can 764R, there was a cleartrend towards greater toxicity in sinking populations than in thefloating or ascending population. This demonstrates that whencellular toxicity is greatest, cells that tend to sink are likely going tobe the most toxic individuals within a population during thatspecific timeframe.
As the population metabolizes surplus nutrients and moves intothe late stationary growth phase, the most toxic cells will already beat deeper depths in the water column. Nutrient deplete conditions(possibly coupled with the corresponding decrease in irradiancewith depth) could drive these cells towards the benthic environmentin search of new nutrients or induce vegetative resting cystformation (Yamochi, 1984). And as cells sink to nutrient-rich deeperwaters, cell toxicities may change with the altered physiologicalstate. The potential effects of H. akashiwo on members of the benthiccommunity in marine coastal waters are not well understood, but itis likely that benthic organisms are as susceptible to the toxicity of H.
akashiwo as pelagic organisms (Shikata et al., 2007). Cysts arecapable of rapid descent towards the benthos with sinking rates ofabout 5–11 m d�1, and thus it is these individuals that have thehighest probability of acting as ‘‘seeds’’ during subsequent bloomsand seasons (Imai and Itakura, 1999).

L. Powers et al. / Harmful Algae 13 (2012) 95–104 103
Although toxicity in H. akashiwo is environmentally regulated,there may be a genetic component common to strains that arecapable of achieving high toxic levels (Imai and Itakura, 1999).Within individual communities or blooms, if the individualscapable of highest toxicity are also those forming cysts andsubsequently seeding future communities or blooms, then theseassemblages will consist of the progeny of the most toxic cells fromthe prior season or year. Thus, when environmental conditionsallow for high biomass bloom formation, then that bloom will begenerally more toxic than previous occurrences.
Future research should incorporate the possible effects of lightand temperature on both sinking and toxicity, as both arephysiological responses intended to optimize the capture anduptake of nutrients from the environment. For example, as bloombiomass increases so does the amount of self-shading endured byindividual cells. It is possible that low-levels of photosyntheticirradiance may encourage the formation of vegetative cysts ifcompetition for resources becomes overwhelming. Likewise,studies in Japan (Yamochi, 1984; Imai and Itakura, 1999) havedemonstrated the importance of both surface and bottom watertemperatures in the survival and re-emergence of vegetative cells.It would be valuable to examine the role temperature plays ininducing toxicity and the effect it may have on encouragingvegetative cysts in the benthos to ascend back to the pelagic zoneand re-emerge.
5. Conclusion
The physiology of H. akashiwo is highly responsive to resourceavailability the surrounding environment. In natural communities,various responses are regulated by the availability of nutrients andlight and are also intimately associated with the age and growthphase of individuals within a bloom. This study showed thatalthough there was variability among different isolates, thephysiological responses of sinking and floating to nutrient stressmay also be associated with the relative toxicity of individual cellsin that individuals prone to sinking are also generally more toxic. Ifcells with high sinking rates are also the most toxic, there may benegative impacts on benthic communities. Similarly, if there arevariations of toxicity potential within a population, vegetativeresting cysts are likely to be formed by particularly toxic cells, andthus subsequent blooms may be more conducive to becomingtoxic.
Acknowledgement
A NSERC Discovery Grant to C.G.T supports this work.[TS]
References
Anderson, D.M., Gilbert, P.M., Burkholder, J.M., 2002. Harmful algal blooms andeutrophication: nutrient sources, composition, and consequences. Estuaries 25,704–726.
Bates, S.S., Trainer, V.L., 2006. The ecology of harmful diatoms. In: Graneli, E., Turner,J.T. (Eds.), Ecology of Harmful Algae. Springer-Verlag, Berlin, pp. 81–88.
Bienfang, P.K., 1980. Phytoplankton sinking rates in oligotrophic waters off Hawaii,USA. Mar. Biol. 61, 69–77.
Bienfang, P.K., Harrison, P.J., Quarmby, L.M., 1982. Sinking rate response to deple-tion of nitrate, phosphate, and silicate in four marine diatoms. Mar. Biol. 67,295–302.
Black, E.A., 2000. Differentiation of morphology and toxicity in harmful algal bloomscaused by the raphidophyte alga Heterosigma akashiwo. Ph.D. Thesis, Universityof Victoria, Canada. 207 p.
Chang, F.H., Anderson, C., Boustead, N.C., 1990. First record of a Heterosigma(Raphidophyceae) bloom with associated mortality of cage-reared salmon inBig Glory Bay, New Zealand. New Zeal. J. Mar. Freshwat. Res. 24, 461–469.
Connell, L.B., Cattolico, R.A., 1996. Fragile algae: axenic culture of field-collectedsamples of Heterosigma carterae. Mar. Biol. 125, 421–426.
de Boer, K.M., Tyl, M.R., Vrieling, E.G., van Rijssel, M., 2004. Effects of salinity andnutrient conditions on growth and hemolytic activity of Fibrocapsa japonica(Raphidophyceae). Aquat. Microb. Ecol. 37, 171–181.
Fu, M., Koulman, A., van Rijssel, M., Lutzen, A., de Boer, K.M., Tyl, M.R., Liebezeit, G.,2004. Chemical characterization of three hemolytic compounds from themicroalgal species Fibrocapsa japonica (Raphidophyceae). Toxicon 43,355–363.
Furnas, M., 1990. In situ growth rates of marine phytoplankton: approachesto measurement, community and species growth rates. J. Plankton Res. 12,1117–1151.
Graneli, E., Hansen, P.J., 2006. Allelopathy in harmful algae: a way to compete forresources? In: Graneli, E., Turner, J.T. (Eds.), Ecology of Harmful Algae. Spring-er-Verlag, Berlin, Germany, pp. 189–201.
Graneli, E., Johansson, N., 2003. Increase in the production of allelopathic sub-stances by Prymnesium parvum cells grown under N-or P-deficient conditions.Harmful Algae 2, 135–145.
Hallegraeff, G.M., Hara, Y., 2003. Taxonomy of harmful marine raphidophytes. In:Hallegraeff, G.M., Anderson, D.M. (Eds.), Manual on Harmful Marine Microalgae.UNESCO Publishing, Paris, pp. 511–522.
Hara, Y., Chihara, C., 1987. Morphology, ultrastructure and taxonomy of theraphidophycean alga Heterosigma akashiwo. Bot. Mag. Tokyo 100, 151–163.
Harrison, P.J., Waters, R.E., Taylor, F.J.R., 1980. A broad spectrum artificial seawatermedium for coastal and open ocean phytoplankton. J. Phycol. 16, 28–35.
Heisler, J., Glibert, P.M., Burkholder, J.M., Anderson, D.M., Cochlan, W.P., Dennison,W.C., Dortch, Q., Gobler, C.J., Heil, C.A., Humphries, E., Lewitus, A., Magnien, R.,Marshall, H.G., Sellner, K., Stockwell, D.A., Stoecker, D.K., Suddleson, M., 2008.Eutrophication and harmful algal blooms: a scientific consensus. Harmful Algae8, 3–13.
Herndon, J., Cochlan, W.P., 2006. Nitrogen utilization by the raphidophyte Hetero-sigma akashiwo: growth and uptake kinetics in laboratory cultures. HarmfulAlgae 2, 260–270.
Hershberger, P.K., Rensel, J.E., Matter, A.L., Taub, F.B., 1997. Vertical distribution ofthe chloromonad flagellate Heterosigma carterae in columns: implications forbloom development. Can. J. Fish. Aquat. Sci. 54, 2228–2234.
Honjo, T., 1993. Overview of bloom dynamics and physiological ecology of Hetero-sigma akashiwo. In: Toxic Phytoplankton Blooms in the Sea. Proceedings of the5th International Conference on Toxic Marine Phytoplankton, Elsevier, NewYork, pp. 33–41.
Horner, R.A., Garrison, D.L., Plumley, F.G., 1997. Harmful algal blooms and red tideproblems on the US west coast. Limnol. Oceanogr. 42, 1076–1088.
Imai, I., Itakura, S., 1999. Importance of cysts in the population dynamics of the redtide flagellate Heterosigma akashiwo (Raphidophyceae). Mar. Biol. 133,755–762.
Kamykowski, D., Reed, R.E., Kirkpatrick, G.J., 1992. Comparison of sinking velocity,swimming velocity, rotation and path characteristics among six marine dino-flagellate species. Mar. Biol. 113, 319–328.
Khan, S., Arakawa, O., Onoue, Y., 2008. Neurotoxins in a toxic red tide of Heterosigmaakashiwo in Kagoshima Bay, Japan. Aquacult. Res. 28, 9–14.
Kiorboe, T., Lundsgaard, C., Olesen, M., Hansen, J.L.S., 1994. Aggregation andsedimentation processes during a spring phytoplankton bloom – a field experi-ment to test coagulation theory. J. Mar. Res. 52, 297–323.
Landsberg, J.H., 2002. The effects of harmful algal blooms on aquatic organisms. Rev.Fish. Sci. 10, 113–390.
Lecourt, M.D., Muggli, D.L., Harrison, P.J., 1996. Comparison of growth and sinkingrates of non-coccolith- and coccolith-forming strains of Emiliania huxleyi grownunder different irradiances and nitrogen sources. J. Phycol. 32, 17–21.
Ling, C., Trick, C.G., 2010. Expression and standardized measurement of hemolyticactivity in Heterosigma akashiwo. Harmful Algae 9, 522–529.
Marshall, J.A., Ross, T., Pyecroft, S., Hallegraeff, G., 2005. Superoxide production bymarine microalgae. Mar. Biol. 147, 541–549.
Martinez, R., Orive, E., Laza-Martinez, A., Seoane, S., 2010. Growth response of sixstrains of Heterosigma akashiwo to varying temperature, salinity and irradianceconditions. J. Plankton Res. 32, 529–538.
Morris, J.G., 1999. Harmful algal blooms: an emerging public health problem withpossible health links to human stress in the environment. Annu. Rev. EnergyEnviron. 24, 367–390.
Muggli, D.L., Lecourt, M.D., Harrison, P.J., 1996. Effects of iron and nitrogen source onthe sinking rate, physiology and metal composition of an oceanic diatom fromthe subarctic Pacific. Mar. Ecol. Prog. Series 132, 215–227.
Oda, T., Nakamura, A., Okamoto, T., Ishimatsu, A., Muramatsu, T., 1998. Lectin-induced enhancement of superoxide anion production by red tide phytoplank-ton. Mar. Biol. 131, 383–390.
Parsons, T.R., Maita, Y., Lalli, C.M., 1984. A Manual of Chemical and BiologicalMethods for Seawater Analysis. Permagon Press, Toronto.
Shikata, T., Nagasoe, S., Matsubara, T., Yamasaki, Y., Shimasaki, Y., Oshima, Y., Honjo,T., 2007. Effects of temperature and light on cyst germination and germinatedcell survival of the noxious raphidophyte Heterosigma akashiwo. Harmful Algae6, 700–706.
Smayda, T.J., 1970. The suspension and sinking of phytoplankton in the sea.Oceanogr. Mar. Biol. Ann. Rev. 8, 353–414.
Smayda, T.J., 1989. Primary production and the global epidemic of phytoplanktonbloom the sea: linkage? In: Cosper, E.M.,Bricelj, V.M.,Carpenter, E.J. (Eds.),NovelPhytoplankton, Blooms, Coastal and Estuarine Studies Number 35. Springer-Verlag, New York, pp. 449–484.
Smayda, T.J., 1990. Novel and nuisance phytoplankton blooms in the sea: evidencefor a global epidemic. In: Graneli, E., Sundstrom, B., Edler, L., Anderson, D.M.(Eds.), Toxic Marine Phytoplankton. Elsevier, New York, pp. 29–40.
Smayda, T.J., 1997a. Bloom dynamics: physiology, behavior, trophic effects. Limnol.Oceanogr. 42, 1132–1136.

L. Powers et al. / Harmful Algae 13 (2012) 95–104104
Smayda, T.J., 1997b. Harmful algal blooms: their ecophysiology and general rele-vance to phytoplankton blooms in the sea. Limnol. Oceanogr. 42, 1137–1153.
Smayda, T.J., 1998. Ecophysiology and bloom dynamics of Heterosigma akashiwo(Raphidophyceae). In: Anderson, D.M., Cembella, A.D., Hallegraeff, G.M.(Eds.), Physiological Ecology of Harmful Algal Blooms. Springer-Verlag,Berlin, pp. 113–131.
Taylor, F.J.R., 1991. Current problems with harmful phytoplankton blooms in BritishColumbia waters. In: Smayda, T.J., Shimizu, Y. (Eds.), Fifth International Con-ference on Toxic Marine Phytoplankton. Elsevier, Amsterdam, pp. 699–704.
Taylor, F.J.R., Haigh, N., 1993. The ecology of fish-killing blooms of the chloromonadflagellate Heterosigma in the Strait of Georgia and adjacent waters. Dev. Mar.Biol. 3, 705–710.
Tomas, C.R., 1978. Olisthodiscus luteus (Chrysophyceae) II. Formation and survivalof a benthic stage. J. Phycol. 14, 314–319.
Tomas, C.R., Deason, E., 1981. The influence of grazing by two Acartia species onOlisthodiscus luteus Carter. Mar. Ecol. 2, 215–223.
Twiner, M.J., 2002. Bioactive extracellular metabolites from the marine flagellate,Heterosigma akashiwo. Ph.D. Thesis, University of Western Ontario, Canada.127 pp.
Twiner, M.J., Trick, C.G., 2000. Toxic effects of Heterosigma do not appear to bemediated by hydrogen peroxide. Limnol. Oceanogr. 46, 1400–1405.
Utkilen, H., Gjolme, N., 1995. Iron stimulated toxin production in Microcystisaeruginosa. Appl. Environ. Biol. 61, 797–800.
Vanhaecke, P., Persoone, G., Claus, C., Sorgeloos, P., 1981. Proposal for a short-termtoxicity test with Artemia nauplii. Ecotoxicol. Environ. Saf. 5, 382–387.
Watanabe, M., Kohata, M., Kunugi, M., 1988. Phosphate accumulation and metabo-lism by Heterosigma akashiwo (Raphidophyceae) during diel vertical migrationin a stratified mesocosm. J. Phycol. 24, 244–248.
Welschmeyer, N.A., 1994. Fluorometric analysis of chlorophyll-a in the presence ofchlorophyll-b and pheopigments. Limnol. Oceanogr. 39, 295–303.
Yamochi, S., 1984. Mechanisms for outbreak of Heterosigma akashiwo red tide inOsaka Bay. Japan. J. Oceanogr. 40, 221–229.