ret/ptc rearrangement occurring in primary peritoneal carcinoma
TRANSCRIPT

http://ijs.sagepub.com/International Journal of Surgical Pathology
http://ijs.sagepub.com/content/17/3/187The online version of this article can be found at:
DOI: 10.1177/1066896908329593
2009 17: 187 originally published online 14 January 2009INT J SURG PATHOLSheils
Karen Denning, Jinghuan Li, Sinead Aherne, Giovanni Tallini, Eoin Gaffney, J.J. O'Leary, Horst Zitzelsberger and Orla Richard Flavin, Gerhard Jackl, Stephen Finn, Paul Smyth, Martina Ring, Esther O'Regan, Susanne Cahill, Kristian Unger,
RET/PTC Rearrangement Occurring in Primary Peritoneal Carcinoma
Published by:
http://www.sagepublications.com
can be found at:International Journal of Surgical PathologyAdditional services and information for
http://ijs.sagepub.com/cgi/alertsEmail Alerts:
http://ijs.sagepub.com/subscriptionsSubscriptions:
http://www.sagepub.com/journalsReprints.navReprints:
http://www.sagepub.com/journalsPermissions.navPermissions:
http://ijs.sagepub.com/content/17/3/187.refs.htmlCitations:
What is This?
- Jan 14, 2009 OnlineFirst Version of Record
- May 14, 2009Version of Record >>
at Leabharlann TCD / Trinity College Dublin Library on May 2, 2013ijs.sagepub.comDownloaded from

RET/PTC Rearrangement Occurring in Primary Peritoneal Carcinoma
Richard Flavin, FRCPath, Gerhard Jackl, PhD, Stephen Finn, PhD, FRCPath, Paul Smyth, PhD, Martina Ring, Esther O’Regan, PhD, BDS, Susanne Cahill, PhD, Kristian Unger, PhD, Karen Denning, MSc, Jinghuan Li, MSc, Sinead Aherne, BSc, Giovanni Tallini, MD, Eoin Gaffney, MD, J. J. O’Leary, MD, DPhil, Horst Zitzelsberger, PhD, and Orla Sheils, PhD
187
through a multicomponent complex consisting of glycosyl-phoshatidylinositol and RET tyrosine kinase.5 The GDNF/RET signaling pathway has an important role in regulating the development of the peripheral nervous system and kidney together with germ cell differentiation.5,6 Germline mutations in c-RET have been implicated in multiple endocrine neoplasia types 2A and 2B, familial medullary thyroid carcinoma,7,8 and Hirschsprung’s disease.9 Sporadic mutations in RET are associated with the develop-ment of papillary thyroid carcinoma (PTC).10,11 At least 16 varieties of RET rearrangement have been described involving 11 different genes,8,12,13 the com-monest being RET/PTC1 and RET/PTC 3. The oncogene is created by the fusion of the TK domain of RET to a ubiquitously expressed donor.14 PTC1 consists of the fusion of the RET TK domain with the 5′ end of the H4 gene.15,16 The chimeric protein
R ET (rearranged during transfection) proto-oncogene is a tyrosine kinase receptor, coded for on chromosome 10q11.2, which consists
of an extracellular cadherin motif, a cysteine rich domain, and an intracellular tyrosine kinase (TK) domain.1-4 Glial cell line–derived neurotrophic factor (GDNF) family ligands (a subclass of the transform-ing-growth factor-β [TGF-β] superfamily) signal
RET/PTC rearrangements are initiating events in the development of a significant proportion of papillary thyroid carcinomas. Activated RET/PTC mutations are thought to be restricted to thyroid disease, but this study proposes that these events may also occur in nonthyroid tumors. A total of 57 nonthyroid papillary tumors were examined for RET/PTC rearrangements using interphase fluorescence in situ hybridization, Taqman reverse transcriptase polymerase chain reac-tion, and immunohistochemistry. Taqman single nucle-otide polymorphism detection was used to analyze for expression of mutated BRAF T1799A. In all, 20% (3/15) of primary peritoneal carcinoma had detectable RET/PTC1 rearrangements by all 3 methodologies. A
further case of similar histotype had an alternate RET/PTC rearrangement. No RET/PTC1 rearrangements were detected in the remaining tumor cohort. All 57 tumors were homozygous for wild-type BRAF. The results indicate that RET/PTC rearrangements occur in a small subset of nonthyroid papillary tumors. These rearrangements may not be directly implicated in tumor growth; rather representing “passenger” muta-tions reflecting RET instability in secondary tumor subclones.
Keywords: RET; RET/PTC1; peritoneal serous carci-noma; fluorescence in situ hybridization (FISH); reverse transcriptase polymerase chain reaction (RT-PCR)
From the Department of Histopathology, Trinity College Medical School, Dublin, Ireland (RF, SF, PS, MR, EO’R, SC, KD, JL, SA, EG, JJO’L, OS); Institute of Molecular Radiobiology, GSF-National Research Centre for Environment and Health, Neuherberg, Germany (GJ, KU, HZ); and the Department of Pathology, Bologna University School of Medicine, Bologna, Italy (GT).
Address correspondence to: Richard Flavin, FRCPath, Molecular Histopathology, IMM, St. James’s Hospital, Dublin 8, Ireland; e-mail: [email protected].
International Journal of Surgical Pathology
Volume 17 Number 3June 2009 187-197
© 2009 The Author(s)10.1177/1066896908329593
http://ijsp.sagepub.com
at Leabharlann TCD / Trinity College Dublin Library on May 2, 2013ijs.sagepub.comDownloaded from

188 International Journal of Surgical Pathology / Vol. 17, No. 3, June 2009
localizes to the cytosol where it is constitutively phosphorylated. H4 is a 55-kD nuclear and cytosolic protein, whose loss of function might have a role in thyroid carcinogenesis by impairing apoptosis.17
Despite the proven association of RET/PTC with papillary carcinoma, identification of RET/PTC in other thyroid tumor histotypes, such as oncocytic adenomas and carcinomas,18 and nonneoplastic enti-ties such as hyperplastic nodules19,20 and Hashimoto’s thyroiditis21-23 seems to challenge the validity of RET/PTC as a tumor marker and its specificity for papillary carcinoma.
The primary objective of this study was to investi-gate the possibility that RET/PTC rearrangements may be expressed in nonthyroid papillary tumors using 3 independent techniques. To complete the molecular profile, these tumors were further analyzed for BRAF (T1799A) mutations.
Materials and Methods
Ethical approval for the study in accordance with the Helsinki Declaration was obtained from the St. James’s and Federation of Dublin Voluntary Hospitals Ethics Committee.
Case Selection
Papillary tumors were selected from archival form-alin-fixed, paraffin-embedded tissue (N = 57), between the years 1991 and 2007 from St. James’s Hospital and The Adelaide and Meath National Children’s Hospital (Dublin, Ireland). The study cohort included 15 primary peritoneal carcinomas, 10 serous tumors of the ovary (including 2 border-line tumors), 10 papillary renal cell carcinomas, 10 urothelial cell carcinomas, 5 serous carcinomas of the endometrium, 2 endometrioid carcinomas of the endometrium, 1 adenoacanthoma of the endometrium, and 4 carcinomas with mixed phe-notypes. Slides were reviewed by a histopathologist (RF), original diagnoses confirmed and classified according to World Health Organisation guide-lines. Primary peritoneal carcinoma was identified according to Gynaecological Oncology Group cri-teria. Patient data are summarized in Table 1.
Interphase Fluorescence In Situ Hybridization Analysis for RET/PTC Rearrangement Detection
Interphase fluorescence in situ hybridization (FISH) was performed on 10-µm paraffin-embedded sec-tions (62 samples, including 57 tumors and 5 refer-ence cases) following previous validated protocols.24 All sections had been microdissected to ensure that only tumor tissue was present on the slide for subse-quent laser scanning microscopy scoring.
DNA probe generation from 3 yeast artificial chromosome clones (313F4, 214H10, 344H4) cov-ering the RET locus and probe hybridization were performed as previously described.24 Microscopic analyses were carried out using a confocal laser scanning microscope (Zeiss LSM 510, Zeiss, Jena, Germany). Cell nuclei exhibiting rearranged RET gene show a split FISH signal in red (distal to RET) and green (proximal to RET) in addition to an over-lapping signal.
Normal cells show 2 overlapping nuclei. A total of 100 nuclei with strong and well-delineated signals were scored. Only cells with 2 overlapping signals or 1 split and 1 overlapping signal were counted to ensure only complete cell nuclei had been scored. Images of tumor cell nuclei were captured as previously described.24 The TPC-1 cell line carrying a RET/PTC1 rearrangement was used as control, with rearrange-ment positive cells present in each cell scored.
To determine a meaningful cut-off level indicating a significantly increased proportion of cells with split FISH signals compared with baseline frequency, epi-thelial cells from 5 normal samples, including normal kidney, ovary, and endometrium were scored after careful review of the histology sections to exclude samples with any significant alteration. The mean value (x) and standard error (SE) of cells with split signals was determined. The mean value (x) ± SE value was 0.8 ± 0.8. Data from a previously analyzed cohort of normal thyroids (n = 4) selected for inter-phase FISH were used to validate the FISH scores from the above cohort.21,24 The resulting x ± SE was 1.5 ± 0.7. No statistically significant difference in FISH score difference between normal thyroids and reference cases was observed (P = .41). For calcula-tion of a cut-off level, a stringent value of 12.75 times
at Leabharlann TCD / Trinity College Dublin Library on May 2, 2013ijs.sagepub.comDownloaded from

RET/PTC in Primary Peritoneal Carcinoma / Flavin et al 189
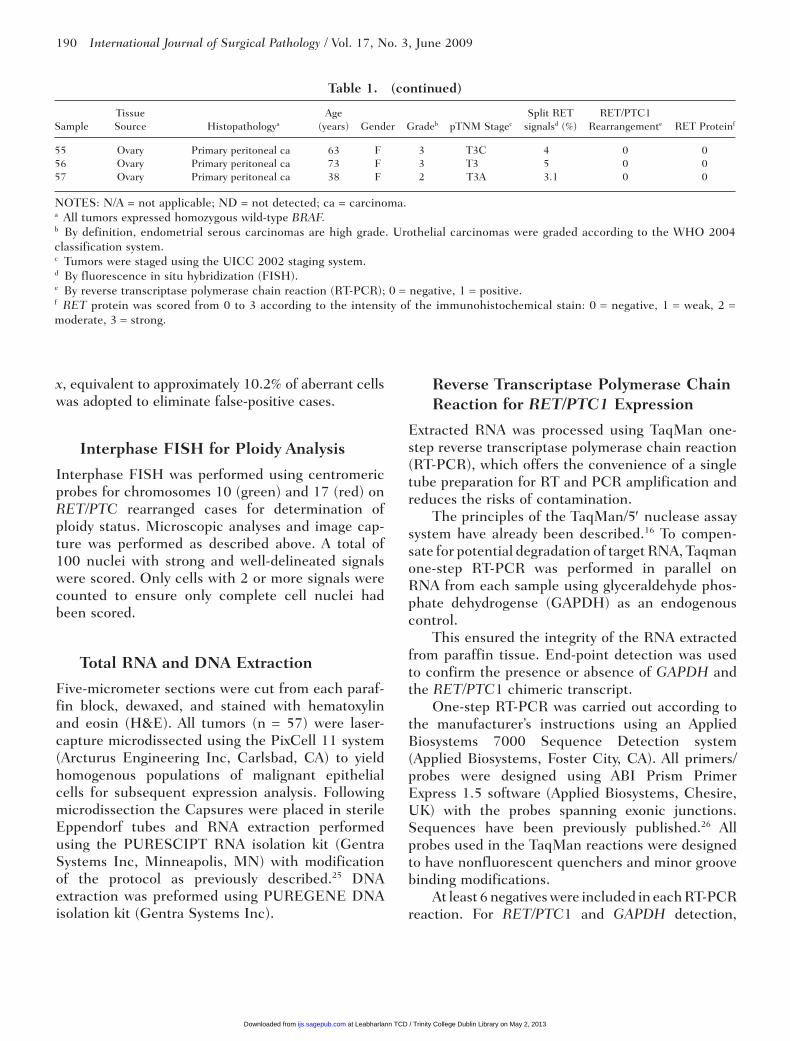
Table 1. Summary of Patient Clinicopathological Data
Tissue Age Split RET RET/PTC1
Sample Source Histopathologya (years) Gender Gradeb pTNM Stagec signalsd (%) Rearrangemente RET Proteinf
1 Bladder Urothelial ca 73 M High T1 at least 3 0 0 2 Bladder Urothelial ca 70 F Low TA 3.3 0 0 3 Bladder Urothelial ca 71 M Low TA 10.1 0 0 4 Bladder Urothelial ca 55 F High T1 5.6 0 0 5 Bladder Urothelial ca 66 F High TA 3 0 2 6 Bladder Urothelial ca 79 F High TA 1 0 0 7 Endometrium Adenoacanthoma 56 F 2 T3A 8.6 0 0 8 Endometrium Serous ca (squamous 91 F 2 T3A 2 0 3 differentiation) 9 Endometrium Serous ca 57 F 3 T1B 0 0 010 Endometrium Serous ca 52 F 3 T1A at least 3 0 011 Endometrium Endometrioid ca 77 F 1 T1C 3.6 0 012 Endometrium Serous ca 57 F 3 T2A 6.1 0 013 Endometrium Serous ca 49 F 3 T3B 1.8 0 014 Endometrium Serous ca 63 F 3 T4 0 0 015 Endometrium Endometrioid ca 57 F 3 T1B 0 0 016 Kidney Urothelial ca 57 M High T3 0 0 017 Kidney Urothelial ca 57 M High T3 ND 0 018 Kidney Papillary renal cell ca type 2 66 F 3 T2 0 0 019 Kidney Papillary renal cell ca type 2 52 F 3 T1A 7 0 020 Kidney Papillary renal cell ca type 2 52 M 2 T2 1 0 021 Kidney Papillary renal cell ca type 1 52 M 2 T1 2.6 0 022 Kidney Papillary renal cell ca type 1 56 M 2 T2 7 0 023 Kidney Papillary renal cell ca type 2 56 M 4 T2 6.3 0 024 Kidney Papillary renal cell ca type 1 48 M 2 T1B 7.9 0 125 Kidney Papillary renal cell ca type 1 62 M 2 T1 12.8 0 026 Kidney Papillary renal cell ca type 1 49 M 3 T1B 6.6 0 027 Kidney Papillary renal cell ca type 2 49 M 3 T2 4.7 0 028 Kidney Urothelial ca 63 M High T3 3 0 329 Kidney Urothelial ca 67 F High TA 6 0 030 Ovary Serous ca 54 F 1 T1A 3 0 131 Ovary Serous ca 86 F 1 T1A 3 0 032 Ovary Serous ca 49 F 2 T3C 10 0 033 Ovary Mixed serous and 59 F 2 T1A 6.9 0 0 endometrioid ca34 Ovary Serous ca 66 F 3 T3C 1.3 0 035 Ovary Serous ca 76 F 3 T3C 1.2 0 036 Ovary Borderline serous tumor 47 F N/A N/A 7.4 0 037 Ovary Borderline serous tumor 58 F N/A N/A 3 0 038 Ovary Primary peritoneal ca 68 F 3 T3C 5 0 039 Ovary Primary peritoneal ca 76 F 2 T3C 1.2 0 0 (<15% clear cell ca)40 Ovary Primary peritoneal ca 50 F 3 T3C 21.1 1 141 Ovary Serous ca 59 F 2 T3C ND 0 042 Ovary Primary peritoneal ca 84 F 2 T3C 1.1 0 043 Ovary Primary peritoneal ca 64 F 3 T3C 5 0 144 Ovary Primary peritoneal ca 54 F 3 T3C 18.3 1 245 Ovary Primary peritoneal ca 50 F 3 T3C 3 0 046 Ovary Primary peritoneal ca 72 F 3 T3C 0 0 047 Ovary Primary peritoneal ca 71 F 2 T3C 2 0 048 Ovary Seromucinous ca 77 F 2 T3 4.9 0 149 Ovary Primary peritoneal ca 81 F 2 T3B 14.1 1 150 Ovary Serous ca 64 F 3 T2C 3.6 0 051 Ovary Primary peritoneal ca 69 F 3 T3C 1.9 0 052 Ovary Primary peritoneal ca 64 F 3 T3C 7 0 353 Ovary Primary peritoneal ca 71 F 3 yT3a 10.6 0 154 Ovary Serous ca 65 F 3 T3C 1.9 0 0
(continued)
at Leabharlann TCD / Trinity College Dublin Library on May 2, 2013ijs.sagepub.comDownloaded from
190 International Journal of Surgical Pathology / Vol. 17, No. 3, June 2009
x, equivalent to approximately 10.2% of aberrant cells was adopted to eliminate false-positive cases.
Interphase FISH for Ploidy Analysis
Interphase FISH was performed using centromeric probes for chromosomes 10 (green) and 17 (red) on RET/PTC rearranged cases for determination of ploidy status. Microscopic analyses and image cap-ture was performed as described above. A total of 100 nuclei with strong and well-delineated signals were scored. Only cells with 2 or more signals were counted to ensure only complete cell nuclei had been scored.
Total RNA and DNA Extraction
Five-micrometer sections were cut from each paraf-fin block, dewaxed, and stained with hematoxylin and eosin (H&E). All tumors (n = 57) were laser-capture microdissected using the PixCell 11 system (Arcturus Engineering Inc, Carlsbad, CA) to yield homogenous populations of malignant epithelial cells for subsequent expression analysis. Following microdissection the Capsures were placed in sterile Eppendorf tubes and RNA extraction performed using the PURESCIPT RNA isolation kit (Gentra Systems Inc, Minneapolis, MN) with modification of the protocol as previously described.25 DNA extraction was preformed using PUREGENE DNA isolation kit (Gentra Systems Inc).
Reverse Transcriptase Polymerase Chain Reaction for RET/PTC1 Expression
Extracted RNA was processed using TaqMan one-step reverse transcriptase polymerase chain reaction (RT-PCR), which offers the convenience of a single tube preparation for RT and PCR amplification and reduces the risks of contamination.
The principles of the TaqMan/5′ nuclease assay system have already been described.16 To compen-sate for potential degradation of target RNA, Taqman one-step RT-PCR was performed in parallel on RNA from each sample using glyceraldehyde phos-phate dehydrogense (GAPDH) as an endogenous control.
This ensured the integrity of the RNA extracted from paraffin tissue. End-point detection was used to confirm the presence or absence of GAPDH and the RET/PTC1 chimeric transcript.
One-step RT-PCR was carried out according to the manufacturer’s instructions using an Applied Biosystems 7000 Sequence Detection system (Applied Biosystems, Foster City, CA). All primers/probes were designed using ABI Prism Primer Express 1.5 software (Applied Biosystems, Chesire, UK) with the probes spanning exonic junctions. Sequences have been previously published.26 All probes used in the TaqMan reactions were designed to have nonfluorescent quenchers and minor groove binding modifications.
At least 6 negatives were included in each RT-PCR reaction. For RET/PTC1 and GAPDH detection,
Table 1. (continued)
Tissue Age Split RET RET/PTC1
Sample Source Histopathologya (years) Gender Gradeb pTNM Stagec signalsd (%) Rearrangemente RET Proteinf
55 Ovary Primary peritoneal ca 63 F 3 T3C 4 0 056 Ovary Primary peritoneal ca 73 F 3 T3 5 0 057 Ovary Primary peritoneal ca 38 F 2 T3A 3.1 0 0
NOTES: N/A = not applicable; ND = not detected; ca = carcinoma.a All tumors expressed homozygous wild-type BRAF.b By definition, endometrial serous carcinomas are high grade. Urothelial carcinomas were graded according to the WHO 2004 classification system.c Tumors were staged using the UICC 2002 staging system.d By fluorescence in situ hybridization (FISH).e By reverse transcriptase polymerase chain reaction (RT-PCR); 0 = negative, 1 = positive.f RET protein was scored from 0 to 3 according to the intensity of the immunohistochemical stain: 0 = negative, 1 = weak, 2 = moderate, 3 = strong.
at Leabharlann TCD / Trinity College Dublin Library on May 2, 2013ijs.sagepub.comDownloaded from

cDNA from the cell line TPC-1 was included as a positive control. All samples were run in triplicate.
Immunohistochemistry
Mouse monoclonal IgG antibody directed against the C-terminal (containing the TK domain) of the human RET oncoprotein (Labvision/Neomarkers Corp, Fremont, CA) was used to detect wild-type and rear-ranged RET. Antibody was standardized to a 1:15 dilu-tion. Four-micrometer tissue sections were cut, dewaxed, and incubated in absolute methanol solu-tion with 0.3 mL of hydrogen peroxide for 30 minutes. Antigen retrieval was carried out by boiling the slides in 0.01 mM citrate buffer, pH 6.0, for 90 seconds.
Slides were then treated with blocking serum for 10 minutes after which they were incubated with pri-mary antibody at 25°C for 60 minutes, followed by biotinylated anti-rabbit IgG (Vectostain ABC kits, Vector Laboratories, Inc, Burlingame, CA) and premixed ABC reagent (Vector Laboratories, Inc). Negative controls replaced primary antiserum with nonimmune bovine serum. Chromogen detection was performed with diaminobenzdine (DAB; DAKO Corp, Carpinteria, CA) solution (0.5 mL of stock DAB in 4.5 mL of Tris buffer with 20 µL of hydrogen peroxide).
Slides were counterstained with hematoxylin. Two pathologists (RF and SF, blinded to the original diagnosis) scored the sections independently. A score of 0 to 3 (0 = absent; 1 = weak; 2 = moderate; 3 = strong) was assigned to the intensity of staining. Only slides with >10% quantity of tumor staining were deemed positive.
Tumors positive for RET/PTC1 by RT-PCR were also stained for TTF-1 and thyroglobulin (both DAKO Corp), at dilutions of 1:1400 and 1:40 using the standard ABC technique, to rule out the possi-bility of thyroid metastasis.
BRAF Mutation (T1799A) Detection
TaqMan single nucleotide polymorphism (SNP) detection was used for mutation detection as previ-ously described by one of the authors.27 Primers and probes were used and designed according to the Applied Biosystems Assays-by-Design Service and have been previously published.27 Amplification and analysis was performed on ABI Prism 7000 Sequence Detection System (Applied Biosystems) for 40 cycles (92°C for 15 seconds, 60°C for 1 minute).
Six of each; no template controls, homozygous con-trols (B-CPAP(MUT) and TPC-1(WT) cell lines), and heterozygous mutation controls (K-2 cell line) were included in each run for control and allele calling pur-poses. All unknown samples were analyzed in duplicate. Samples displaying neither allele call were deemed to have insufficient DNA and were reextracted.
Statistical Analysis
The associations between RET/PTC rearrangement and clinicopathological characteristics were examined by means of Fisher’s exact test (Analyse-It Software Ltd, Leeds, UK). Comparison of FISH scores was examined by means of a Mann–Whitney test. All tests were 2-tailed, and the significance level was set at P < .05.
Results
See Table 1 for a summary of the results. Of the patients, 13 were men of whom 8 had papillary renal cell carcinoma and 5 had urothelial cell carcinoma. The remaining 44 patients were women. The median age of all the patients was 63 years (men 56 years, women 64 years).
Interphase FISH Analysis
FISH has the potential to detect RET rearrange-ments regardless of the specific fusion partner involved and allows examination of the rearrange-ment at a single-cell level (see Figure 1). Five of 57(9%) papillary tumors, corresponding to 4 of 15 (27%) primary peritoneal carcinoma and 1 of 10 pap-illary renal cell carcinoma (10%) had a proportion of cells with split RET signals above the cut-off level. (A further 2 of 57 [3.5%] papillary tumors, correspond-ing to 1 of 10 [10%] urothelial carcinoma and 1 of 10 [10%] serous tumors of the ovary had a proportion of cells with split signals that were borderline with respect to significant split RET signals.)
The highest frequency of rearranged cells after FISH interphase analysis was 21.1 (case 40). No extra chromosome 10 copies were detected in these 5 positive cases.
RT-PCR for RET/PTC1 Expression
Three of 57 (5%) papillary tumors had detectable RET/PTC1 mRNA, corresponding to 3 of 15 (20%)
RET/PTC in Primary Peritoneal Carcinoma / Flavin et al 191
at Leabharlann TCD / Trinity College Dublin Library on May 2, 2013ijs.sagepub.comDownloaded from

192 International Journal of Surgical Pathology / Vol. 17, No. 3, June 2009
primary peritoneal carcinoma. The remaining 54 of 57 (95%) papillary tumors had no detectable RET/PTC1 chimeric transcript.
Immunohistochemistry for RET Protein
Twelve of 57 (21%) papillary tumors had detect-able wild-type or RET/PTC chimeric protein. This corresponds to 6 of 15 (40%) primary peritoneal carcinomas, 2 of 10 (20%) urothelial carcinomas, 3 of 17 (18%) serous tumors (endometrial and ovary), and 1 of 10 (10%) papillary renal cell carcinoma (see Figure 1).
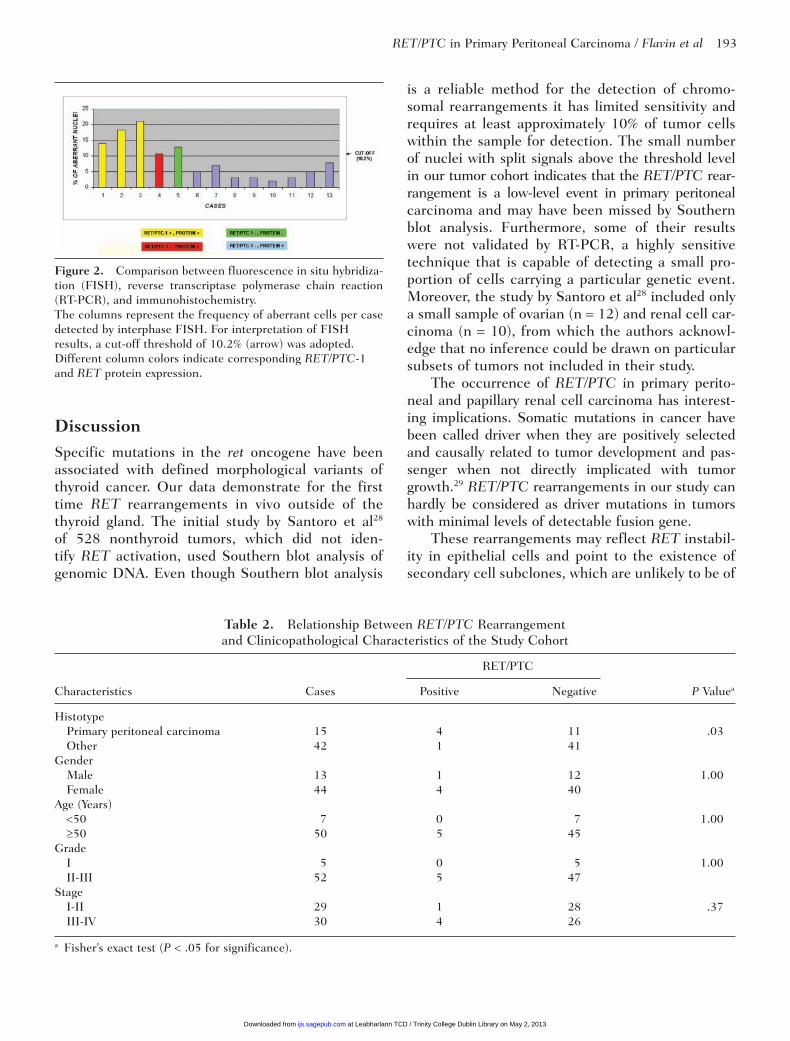
Comparison Between FISH, RT-PCR, and Immunohistochemistry
A comparison between the different approaches used in this study is shown in Figure 2. All 3 cases (cases 40, 44, and 49) of primary peritoneal carci-noma with detectable RET/PTC1 mRNA by RT-PCR exhibited rearranged cells in FISH analysis above the cut-off level and demonstrated RET/PTC chi-meric protein expression.
A single case of primary peritoneal carcinoma (case 53) had split FISH signals in more than 10.2% of cells analyzed and demonstrated RET/
PTC chimeric protein expression. However, no RET/PTC-1 mRNA was detected by RT-PCR Therefore, it is likely that this case harbors a RET rearrangement other than H4/RET (RET/PTC-1). A single case of papillary renal cell carcinoma (case 25) showed neither chimeric RET/PTC protein expression nor demonstrable RET/PTC1 mRNA by RT-PCR. However, this case exhibited split FISH signals in more than 10.2% of cells analyzed. It is likely that this case harbors a RET rearrangement other than H4/RET (RET/PTC1) that is not expressed at the protein level, possibly as a result of posttranscrip-tional regulation. Alternatively, the sensitivities of both the RT-PCR and immunohistochemistry were not high enough to detect a few RET/PTC1 rear-ranged cells, which may also be a reflection of tumor heterogeneity.24
The last subgroup consists of 2 cases of pri-mary peritoneal carcinoma (cases 43 and 52), 2 cases of urothelial cell carcinoma (cases 5 and 28), 3 cases of serous tumors (endometrial and ovary: cases 8, 30, and 48), and 1 case of papillary renal cell carcinoma (case 24). None of these cases had demonstrable RET/PTC1 mRNA by RT-PCR or split FISH signals greater than the cut-off level. However, they all demonstrated RET protein expression, which must logically be the wild-type protein. The remaining 44 cases did not demon-strate RET rearrangements by any of the 3 meth-odologies.
BRAF Mutation (T1799A) Detection
BRAF T1799A mutation was not detected in any of the 57 tumors—all were homozygous for wild type BRAF.
Correlation of RET/PTC Rearrangement With Clinicopathological Features
Our next investigation was to consider whether the presence of RET/PTC rearrangement had any rela-tionship with the clinicopathological characteris-tics of the study cohort. We found that there was a statistically significant relationship between the presence of RET/PTC rearrangement and tumor histotype (see Table 2). Cases with a positive RET/PTC rearrangement showed a significant associa-tion with primary peritoneal carcinoma histotype (P = .03).
Figure 1. Histological appearance of primary peritoneal carci-noma with corresponding fluorescence in situ hybridization (FISH) images.Split FISH signals are highlighted with an arrow. A, Hematoxylin and eosin section of primary peritoneal carcinoma (case 44, Table 1). Original magnification: ×100. B, RET immunohisto-chemical section of corresponding primary peritoneal carcinoma. Original magnification: ×200. C, FISH image of corresponding primary peritoneal carcinoma. Original magnification: ×400.
at Leabharlann TCD / Trinity College Dublin Library on May 2, 2013ijs.sagepub.comDownloaded from
DiscussionSpecific mutations in the ret oncogene have been associated with defined morphological variants of thyroid cancer. Our data demonstrate for the first time RET rearrangements in vivo outside of the thyroid gland. The initial study by Santoro et al28 of 528 nonthyroid tumors, which did not iden-tify RET activation, used Southern blot analysis of genomic DNA. Even though Southern blot analysis
is a reliable method for the detection of chromo-somal rearrangements it has limited sensitivity and requires at least approximately 10% of tumor cells within the sample for detection. The small number of nuclei with split signals above the threshold level in our tumor cohort indicates that the RET/PTC rear-rangement is a low-level event in primary peritoneal carcinoma and may have been missed by Southern blot analysis. Furthermore, some of their results were not validated by RT-PCR, a highly sensitive technique that is capable of detecting a small pro-portion of cells carrying a particular genetic event. Moreover, the study by Santoro et al28 included only a small sample of ovarian (n = 12) and renal cell car-cinoma (n = 10), from which the authors acknowl-edge that no inference could be drawn on particular subsets of tumors not included in their study.
The occurrence of RET/PTC in primary perito-neal and papillary renal cell carcinoma has interest-ing implications. Somatic mutations in cancer have been called driver when they are positively selected and causally related to tumor development and pas-senger when not directly implicated with tumor growth.29 RET/PTC rearrangements in our study can hardly be considered as driver mutations in tumors with minimal levels of detectable fusion gene.
These rearrangements may reflect RET instabil-ity in epithelial cells and point to the existence of secondary cell subclones, which are unlikely to be of
RET/PTC in Primary Peritoneal Carcinoma / Flavin et al 193
Table 2. Relationship Between RET/PTC Rearrangement and Clinicopathological Characteristics of the Study Cohort
RET/PTC
Characteristics Cases Positive Negative P Valuea
Histotype Primary peritoneal carcinoma 15 4 11 .03 Other 42 1 41 Gender Male 13 1 12 1.00 Female 44 4 40 Age (Years) <50 7 0 7 1.00 ≥50 50 5 45 Grade I 5 0 5 1.00 II-III 52 5 47 Stage I-II 29 1 28 .37 III-IV 30 4 26
a Fisher’s exact test (P < .05 for significance).
Figure 2. Comparison between fluorescence in situ hybridiza-tion (FISH), reverse transcriptase polymerase chain reaction (RT-PCR), and immunohistochemistry.The columns represent the frequency of aberrant cells per case detected by interphase FISH. For interpretation of FISH results, a cut-off threshold of 10.2% (arrow) was adopted. Different column colors indicate corresponding RET/PTC-1 and RET protein expression.
at Leabharlann TCD / Trinity College Dublin Library on May 2, 2013ijs.sagepub.comDownloaded from

194 International Journal of Surgical Pathology / Vol. 17, No. 3, June 2009
any pathological consequence in determining bio-logical behavior.
It is interesting to hypothesize on the association between RET/PTC rearrangements and primary peritoneal carcinoma. One hypothesis is that inflam-mation favors the occurrence of the rearrangement as a secondary phenomenon. The propensity of thyrocytes to undergo RET recombination has been explained by the peculiar arrangement of chromatin that juxtaposes RET and its fusion partners in inter-phase nuclei. It is possible that free radical produc-tion, cytokine secretion, cellular proliferation, and other events related to inflammation trigger the occurrence of the rearrangement predisposed to it by an unstable chromatin confirmation.30 Of note, a proinflammatory tumor microenvironment has been described in ovarian cancer which contributes to immune cell recruitment and differentiation (“tumor-immune cell education model”).31 Conversely, the occurrence of RET/PTC may directly influence the inflammation within these tumors. Of note, RET/PTC protein expression produces a strong inflam-matory response in experimental animal models32 and activates numerous inflammatory mediators and molecules within thyroid epithelial cells.33-36 RET/PTC transformed cells can modify their microenvi-ronment to promote autonomous cell proliferation in neighboring nonneoplastic thyrocytes.33 Similarly, ovarian cancer cells can “educate” immune infil-trates to produce the type of cytokines that will facilitate tumor growth and metastases as well as acquiring immune tolerance.31
Another hypothesis is that a putative link may exist between X-ray radiation and RET/PTC1 activa-tion analogous to the known association in PTC. Increased risk of epithelial ovarian carcinoma has been described following diagnostic X-rays37 and high frequency of RET rearrangements following radiation have been observed in PTC.38,39
More specifically, preferential RET/PTC1 induc-tion has been associated with X-rays,40 allowed by spatial juxtaposition of the RET and H4 proto-oncogenes,41 whereas RET/PTC3 rearrangements were more prevalently associated with PTCs arising in children of Belarus and the Ukraine post-Cherno-byl. Interestingly, one of the patients with a demon-strable RET/PTC rearrangement in this study (case 53) had preoperative chemotherapy; however, no association between RET/PTC induction and this form of therapy has been previously described.
It is interesting to speculate that RET/PTC1 acti-vation maybe contributing in part to the adaptation
of a papillary growth pattern. Yap et al42 demon-strated that tyrosine phosphorylation alters thyroid epithelial organization by interfering with actin-associated adhesive junctions. Under the influence of tyrosine phosphorylation, thyroid cells lose their capacity to form follicles, spread and migrate into confluent monolayers and hence the capacity to form follicles in vivo. Fischer et al43 cultured thyroid cells infected with a retroviral vector expressing activated RET/PTC1 and demonstrated alterations in the nuclear envelope and chromatin structure, found in later work to be induced by RET/PTC dur-ing interphase.44 It is plausible therefore that acti-vated RET/PTC1 may influence the growth pattern in papillary tumors. Interestingly, the tumors with activated RET, while having papillary morphology did not have the classical nuclear clearing (“Orphan Annie”) of PTC.
However, RET/PTC rearrangements have been found in tumors (ie, a subset of Hurthle cell tumors)45 that may lack both papillary architecture and/or classic nuclear features. As an adjunct all RET rear-ranged cases were negative for mutated BRAF, sug-gesting distinct alternative pathways in the epipathogenesis of these tumors akin to PTC.46
This study had some technical limitations. Degraded RNA isolated from formalin-fixed par-affin embedded tissue was used. Highly sensitive RT-PCR carries a higher risk of false-positive ampli-fication compared with other techniques. To mini-mize the effects of these limitations, 3 methodologies were employed to identify RET rearrangements. Specifically, interphase FISH and RT-PCR analysis were employed on laser-microdissected cells (reduc-ing the propensity for contamination by RET express-ing macrophages) after careful review of the samples before molecular analysis. The number of tumor nuclei with split FISH signals above background was also limited; a stringent cut-off level of 10.2% posi-tive cells was employed to detect the chromosomal rearrangement. This cut-off level parallels previous studies, which used a cut-off level of between 5% and 10% to separate cases from false-positives.24,47 Immunohistochemical detection of RET protein expression was used to further corroborate our find-ings. The chimeric RET/PTC gene encodes a protein product that contains the cytoplasmic portion of ret.48 Therefore, immunohistochemical detection of the carboxy terminus of the RET protein should serve as a reliable marker to detect rearranged RET.49,50 However, one cannot entirely exclude the possibility of the RET antibody detecting only wild-type RET as
at Leabharlann TCD / Trinity College Dublin Library on May 2, 2013ijs.sagepub.comDownloaded from

it is feasible that a steric change of the epitope may occur after RET rearrangement.
Indeed, according to Rebelo et al,51 positive stain-ing may correspond to the expression of the wild-type RET, RET rearrangement, or both. Moreover, prob-lems with interpreting the specificity of weak focal immunostaining for RET have been reported.52,53 To overcome this problem we only deemed cases with dif-fuse RET immunostaining as being positive as recom-mended by previous authors.52 Any possibility that the tumors that expressed rearranged RET in our series, represented metastases from the thyroid is diminished given the fact no protein expression of TTF-1 or thy-roglobulin was detected in any of these tumors.
Importantly, our results have broad implications for molecular diagnostics. When trying to diagnose metastatic PTC, the existence of minor cell sub-clones with RET rearrangements in nonthyroid pap-illary tumors highlights the importance of using quantitative methods to detect RET/PTC rearrange-ments. In conclusion, this study demonstrates that RET/PTC rearrangement is present in a small number of nonthyroid papillary tumors, specifically primary peritoneal and papillary renal cell carci-noma, therefore RET/PTC detection should not be equated per se with PTC. It indicates that in some primary peritoneal and papillary renal cell carci-noma, RET/PTC represents a passenger mutation occurring in a small number of tumor cells.
References 1. Ishizaka Y, Itoh F, Tahira T, et al. Human ret proto-on-
cogene mapped to chromosome 10q11.2. Oncogene. 1989; 4:1519-1521.
2. Iwamoto T, Taniguchi M, Asai N, Ohkusu K, Nakashima I, Takahashi M. cDNA cloning of mouse ret proto-oncogene and its sequence similarity to the cadherin superfamily. Oncogene. 1993;8:1087-1091.
3. Takahashi M, Buma Y, Hiai H. Isolation of ret proto-oncogene cDNA with an amino-terminal signal sequence. Oncogene. 1989;4:805-806.
4. Takahashi M, Buma Y, Iwamoto T, Inaguma Y, Ikeda H, Hiai H. Cloning and expression of the ret proto-onco-gene encoding a tyrosine kinase with two potential trans-membrane domains. Oncogene. 1988;3:571-578.
5. Takahashi M. The GDNF/RET signaling pathway and human diseases. Cytokine Growth Factor Rev. 2001;12: 361-373.
6. Manie S, Santoro M, Fusco A, Billaud M. The RET recep-tor: function in development and dysfunction in congeni-tal malformation. Trends Genet. 2001;17:580-589.
7. Asa SL. How familial cancer genes and environmentally induced oncogenes have changed the endocrine land-scape. Mod Pathol. 2001;14:246-253.
8. Jhiang SM. The RET proto-oncogene in human cancers. Oncogene. 2000;19:5590-5597.
9. Romeo G, Ronchetto P, Luo Y, et al. Point muta-tions affecting the tyrosine kinase domain of the RET proto-oncogene in Hirschsprung’s disease. Nature. 1994;367:377-378.
10. Grieco M, Santoro M, Berlingieri MT, et al. PTC is a novel rearranged form of the ret proto-oncogene and is frequently detected in vivo in human thyroid papillary carcinomas. Cell. 1990;60: 557-563.
11. Santoro M, Carlomagno F, Hay ID, et al. Ret oncogene activation in human thyroid neoplasms is restricted to the papillary cancer subtype. J Clin Invest. 1992;89: 1517-1522.
12. Klugbauer S, Jauch A, Lengfelder E, Demidchik E, Rabes HM. A novel type of RET rearrangement (PTC8) in childhood papillary thyroid carcinomas and charac-terization of the involved gene (RFG8). Cancer Res. 2000;60:7028-7032.
13. Salassidis K, Bruch J, Zitzelsberger H, Lengfelder E, Kellerer AM, Bauchinger M. Translocation t(10;14) (q11.2:q22.1) fusing the kinetin to the RET gene creates a novel rearranged form (PTC8) of the RET proto-onco-gene in radiation-induced childhood papillary thyroid carcinoma. Cancer Res. 2000;60:2786-2789.
14. Williams GH, Williams ED. Identification of tumour-specific translocations in archival material. J Pathol. 1995;175:279-281.
15. Grieco M, Cerrato A, Santoro M, Fusco A, Melillo RM, Vecchio G. Cloning and characterization of H4 (D10S170), a gene involved in RET rearrangements in vivo. Oncogene. 1994;9:2531-2535.
16. Pasini B, Hofstra RM, Yin L, et al. The physical map of the human RET proto-oncogene. Oncogene. 1995;11: 1737-1743.
17. Celetti A, Cerrato A, Merolla F, Vitagliano D, Vecchio G, Grieco M. H4(D10S170), a gene frequently rearranged with RET in papillary thyroid carcinomas: functional characterization. Oncogene. 2004;23:109-121.
18. Cheung CC, Ezzat S, Ramyar L, Freeman JL, Asa SL. Molecular basis of Hurthle cell papillary thyroid carci-noma. J Clin Endocrinol Metab. 2000;85:878-882.
19. Elisei R, Romei C, Vorontsova T, et al. RET/PTC rear-rangements in thyroid nodules: studies in irradiated and not irradiated, malignant and benign thyroid lesions in children and adults. J Clin Endocrinol Metab. 2001;86:3211-3216.
20. Ishizaka Y, Kobayashi S, Ushijima T, Hirohashi S, Sugimura T, Nagao M. Detection of ret/PTC transcripts in thyroid adenomas and adenomatous goiter by an RT-PCR method. Oncogene. 1991;6:1667-1672.
RET/PTC in Primary Peritoneal Carcinoma / Flavin et al 195
at Leabharlann TCD / Trinity College Dublin Library on May 2, 2013ijs.sagepub.comDownloaded from

196 International Journal of Surgical Pathology / Vol. 17, No. 3, June 2009
21. Rhoden KJ, Unger K, Salvatore G, et al. RET/papil-lary thyroid cancer rearrangement in nonneoplastic thyrocytes: follicular cells of Hashimoto’s thyroidi-tis share low-level recombination events with a sub-set of papillary carcinoma. J Clin Endocrinol Metab. 2006;91:2414-2423.
22. Sheils OM, O’Eary JJ, Uhlmann V, Lättich K, Sweeney EC. ret/PTC-1 Activation in Hashimoto thyroiditis. Int J Surg Pathol. 2000;8:185-189.
23. Wirtschafter A, Schmidt R, Rosen D, et al. Expression of the RET/PTC fusion gene as a marker for papillary carcinoma in Hashimoto’s thyroiditis. Laryngoscope. 1997;107:95-100.
24. Unger K, Zitzelsberger H, Salvatore G, et al. Heterogeneity in the distribution of RET/PTC rearrangements within individual post-Chernobyl papillary thyroid carcinomas. J Clin Endocrinol Metab. 2004;89:4272-4279.
25. Sheils OM, Sweeney EC. TSH receptor status of thyroid neoplasms: TaqMan RT-PCR analysis of archival mate-rial. J Pathol. 1999;188:87-92.
26. Smyth P, Sheils O, Finn S, Martin C, O’Leary J, Sweeney EC. Real-time quantitative analysis of E-cadherin expression in ret/PTC-1-activated thyroid neoplasms. Int J Surg Pathol. 2001;9:265-272.
27. Smyth P, Finn S, Cahill S, et al. ret/PTC and BRAF act as distinct molecular, time-dependant triggers in a spo-radic Irish cohort of papillary thyroid carcinoma. Int J Surg Pathol. 2005;13:1-8.
28. Santoro M, Sabino N, Ishizaka Y, et al. Involvement of RET oncogene in human tumours: specificity of RET acti-vation to thyroid tumours. Br J Cancer. 1993;68:460-464.
29. Davies H, Hunter C, Smith R, et al. Somatic mutations of the protein kinase gene family in human lung cancer. Cancer Res. 2005;65:7591-7595.
30. Gandhi M, Medvedovic M, Stringer JR, Nikiforov YE. Interphase chromosome folding determines spatial prox-imity of genes participating in carcinogenic RET/PTC rearrangements. Oncogene. 2006;25:2360-2366.
31. Chen R, Alvero AB, Silasi DA, Steffensen KD, Mor G. Cancers take their Toll: the function and regulation of Toll-like receptors in cancer cells. Oncogene. 2008;27:225-233.
32. Powell DJ Jr, Eisenlohr LC, Rothstein JL. A thyroid tumor-specific antigen formed by the fusion of two self proteins. J Immunol. 2003;170:861-869.
33. Melillo RM, Castellone MD, Guarino V, et al. The RET/PTC-RAS-BRAF linear signaling cascade mediates the motile and mitogenic phenotype of thyroid cancer cells. J Clin Invest. 2005;115:1068-1081.
34. Puxeddu E, Knauf JA, Sartor MA, et al. RET/PTC-induced gene expression in thyroid PCCL3 cells reveals early acti-vation of genes involved in regulation of the immune response. Endocr Relat Cancer. 2005;12:319-334.
35. Russell JP, Engiles JB, Rothstein JL. Proinflammatory mediators and genetic background in oncogene mediated tumor progression. J Immunol. 2004;172:4059-4067.
36. Shinohara S, Rothstein JL. Interleukin 24 is induced by the RET/PTC3 oncoprotein and is an autocrine growth factor for epithelial cells. Oncogene. 2004;23:7571-7579.
37. Harlap S, Olson SH, Barakat RR, et al. Diagnostic x-rays and risk of epithelial ovarian carcinoma in Jews. Ann Epidemiol. 2002;12:426-434.
38. Klugbauer S, Lengfelder E, Demidchik EP, Rabes HM. High prevalence of RET rearrangement in thyroid tumors of children from Belarus after the Chernobyl reactor accident. Oncogene. 1995;11:2459-2467.
39. Rabes HM, Demidchik EP, Sidorow JD, et al. Pattern of radiation-induced RET and NTRK1 rearrangements in 191 post-Chernobyl papillary thyroid carcinomas: bio-logical, phenotypic, and clinical implications. Clin Cancer Res. 2000;6:1093-1103.
40. Mizuno T, Iwamoto KS, Kyoizumi S, et al. Preferential induction of RET/PTC1 rearrangement by X-ray irradia-tion. Oncogene. 2000;19:438-443.
41. Nikiforova MN, Stringer JR, Blough R, Medvedovic M, Fagin JA, Nikiforov YE. Proximity of chromosomal loci that participate in radiation-induced rearrangements in human cells. Science. 2000;290:138-141.
42. Yap AS, Stevenson BR, Cooper V, Manley SW. Protein tyrosine phosphorylation influences adhesive junction assembly and follicular organization of cultured thy-roid epithelial cells. Endocrinology. 1997;138: 2315-2324.
43. Fischer AH, Bond JA, Taysavang P, Battles OE, Wynford-Thomas D. Papillary thyroid carcinoma oncogene (RET/PTC) alters the nuclear envelope and chromatin struc-ture. Am J Pathol. 1998;153:1443-1450.
44. Fischer AH, Taysavang P, Jhiang SM. Nuclear envelope irregularity is induced by RET/PTC during interphase. Am J Pathol. 2003;163:1091-1100.
45. Belchetz G, Cheung CC, Freeman J, Rosen IB, Witterick IJ, Asa SL. Hurthle cell tumors: using molecular techniques to define a novel classification system. Arch Otolaryngol Head Neck Surg. 2002;128:237-240.
46. Soares P, Trovisco V, Rocha AS, et al. BRAF mutations and RET/PTC rearrangements are alternative events in the etiopathogenesis of PTC. Oncogene. 2003;22:4578-4580.
47. Barr FG, Qualman SJ, Macris MH, et al. Genetic het-erogeneity in the alveolar rhabdomyosarcoma subset without typical gene fusions. Cancer Res. 2002;62: 4704-4710.
48. Fusco A, Grieco M, Santoro M, et al. A new oncogene in human thyroid papillary carcinomas and their lymph-nodal metastases. Nature. 1987;328:170-172.
49. Cheung CC, Ezzat S, Freeman JL, Rosen IB, Asa SL. Immunohistochemical diagnosis of papillary thyroid car-cinoma. Mod Pathol. 2001;14:338-342.
50. Sugg SL, Ezzat S, Rosen IB, Freeman JL, Asa SL. Distinct multiple RET/PTC gene rearrangements in
at Leabharlann TCD / Trinity College Dublin Library on May 2, 2013ijs.sagepub.comDownloaded from

RET/PTC in Primary Peritoneal Carcinoma / Flavin et al 197
multifocal papillary thyroid neoplasia. J Clin Endocrinol Metab. 1998;83:4116-4122.
51. Rebelo S, Domingues R, Catarino AL, et al. Immunostaining and RT-PCR: different approaches to search for RET rearrangements in patients with papillary thyroid carcinoma. Int J Oncol. 2003; 23:1025-1032.
52. Cerilli LA, Mills SE, Rumpel CA, Dudley TH, Moskaluk CA. Interpretation of RET immunostaining in follicular lesions of the thyroid. Am J Clin Pathol. 2002;118:186-193.
53. Guyetant S, Michalak S, Valo I, Saint-Andre JP. Diagnosis of the follicular variant of papillary thyroid carcinoma. Significance of immunohistochemistry. Ann Pathol. 2003;23:11-20.
For reprints and permissions queries, please visit SAGE’s Web site at http://www.sagepub.com/journalsPermissions.nav
at Leabharlann TCD / Trinity College Dublin Library on May 2, 2013ijs.sagepub.comDownloaded from