recent advances in commercial production of astaxanthin from microalgae
TRANSCRIPT

Miguel Olaizola and Mark E. Huntley
Recent advances in commercial production of astaxanthin from microalgae
In
Recent Advances in Marine Biotechnology. Volume 9. Biomaterials and Bioprocessing
(Eds. M. Fingerman and R. Nagabhushaman)
Science Publishers, New Hampshire, pp: 143-164. 2003.

Recent Advances in Commercial Production ofAstaxanthin from Microalgae
Miguel Olaizola and Mark E. Huntley
Mera Pharmaceuticals (formerly Aquasearch, Inc.) Kailua-Kona, &&Hawau 96740, U.S.A.
What is Astaxanthin
Astaxanthin (C40H52O4, FW 596.8) is a xanthophyll, an oxidative deriva-tive of carotenes (Figure 1). It is the main carotenoid pigment found inaquatic animals (Miki 1991). This red-orange pigment is closely related toother well-known carotenoids such as b-carotene or lutein. In many ofthe aquatic animals in which it is found, astaxanthin has a number ofessential biological functions, ranging from protection against oxidationof essential polyunsaturated fatty acids, protection against ultraviolet (UV)light effects, immune response, pigmentation, and communication, to re-productive behavior and improved reproduction (Meyers 1993). In spe-cies such as salmon or shrimp, astaxanthin is considered essential tonormal growth and survival, and has been attributed vitamin-like prop-erties (Torissen et al. 1989).
For Correspondence: Miguel Olaizola, [email protected]
HO
OH
O
Astaxanthin 3S,3’S
3
3’
HO
O
OH
O
Astaxanthin 3R,3’S
3
3’
OOH3¢
3HO
OO
OH3¢
HO3
HO
O
OH
O
Astaxanthin 3R,3’R
3
3’
3
OOH3¢
Astaxanthin 3R, 3�SO
HO
Figure 1. Astaxanthin molecules.
Astaxanthin 3R, 3�SO
Astaxanthin 3S, 3�S(3, 3�-dihydroxy-4, 4�-diketo-b-carotene)

144 MARINE BIOTECHNOLOGY
Astaxanthin in Nature
Astaxanthin can be found in many of our favorite seafoods such as salmon,trout, red seabream, shrimp, lobster and fish eggs (Torissen et al. 1989). Itis also found in birds like flamingo, quail, and other species (Egeland1993, Inborr 1998). Astaxanthin, like other carotenoids, cannot be synthe-sized by animals and must be provided in the diet. While mammals andfish such as salmon or trout are unable to convert other dietary carotenoidsinto astaxanthin, crustaceans like shrimp and some fish species (e.g., koicarp) have a limited capacity to convert closely related dietary carotenoidsinto astaxanthin, although they benefit strongly from being fed astaxanthindirectly (Meyers 1993). Mammals lack the ability to synthesize astaxanthin,or to convert dietary astaxanthin into vitamin A: unlike b-carotene, astax-anthin has no pro-vitamin A activity in these animals (Jyonouchi et al.1995). Some micro-organisms can be quite rich in astaxanthin.Haematococcus pluvialis, a chlorophyte alga, is believed to be the organismwhich can accumulate the highest levels of astaxanthin in nature. Thefunction of astaxanthin appears to be to protect the algae from adverseenvironment changes, such as increased UV-light photoxidation andevaporation of the water pools in which it lives (Droop 1953, 1961,Almgren 1966). Commercially grown Haematococcus can accumulate asmuch as 30 g of astaxanthin per kg of dry biomass (Olaizola 2000), a levelthat is 1,000 to 3,000 fold higher than that found in salmon fillets.
Chemistry of Astaxanthin
The structure of the astaxanthin molecule is at the base of its uniqueproperties (Figure 1). The presence of the hydroxyl and keto endings oneach ionone ring allows the molecule the ability to be esterified and havehigh anti-oxidant activity and a polar configuration. Free astaxanthin isparticularly sensitive to oxidation. Consequently, it is usually foundeither conjugated to proteins, like in salmon muscle or lobster exoskeleton,or esterified with one or two fatty acids (monoester and diester forms),which stabilizes the molecule (Meyers 1993). In H. pluvialis, the esterifiedform predominates, mostly as astaxanthin monoester (Lorenz andCysewski 2000). Various astaxanthin isomers are found in nature. Theall-E form, characterized by an alignment of the isoprene groups in astraight line, tends to be more bio-available in salmon than the Z-forms.Three forms of the all-E isomer can be found. They are the 3S,3¢S, 3R,3¢S,and 3R,3¢R stereoisomers, characterized by the configuration of the twohydroxyl groups on the molecule.
Form and level of deposition of astaxanthin differ among tissues andspecies. In salmon, esterified astaxanthin predominates in the skin,

RECENT ADVANCES IN COMMERCIAL PRODUCTION 145
teguments, and eggs, while free astaxanthin is the main form in the flesh,serum and other internal organs (Torrisen et al. 1989). In shrimp, esterifiedastaxanthin predominates, except in the ovaries and eggs (Menasvetaet al. 1993, Dall 1995). In red seabream, mostly esterified astaxanthin isfound in the skin (Fujita et al. 1983, Ito et al. 1986). The more stableesterified form is believed to be an adaptive feature to be able to storeastaxanthin in tissues without excessive oxidation (Miki 1991).
Furr and Clark (1997) reviewed the various steps of digestion, absorp-tion, and plasma transport of dietary carotenoids in mammals. In theplasma, they noted that non-polar carotenoids like b-carotene, a-carotene,or lycopene, are mostly transported by VLDL (very low densitylipoproteins) and LDL (low density lipoproteins) and the polar carotenoidslike zeaxanthin or lutein are more likely to be transported by LDL andHDL (high density lipoproteins). To date, we are aware of only one pub-lished study that studied in detail absorption and plasma kinetics ofastaxanthin in humans. Østerlie et al. (1999) confirmed the bio-availabil-ity of astaxanthin supplied in a single high dosage of 100 mg and con-firmed its transport in the plasma by lipoproteins. Maximum levels ofastaxanthin (1.3 +/- 0.1 mg/L) were reached 6.7 +/- 1.2 hr after adminis-tration, and the plasma astaxanthin elimination half-life was 21 +/- 11 hr.In the plasma, astaxanthin was present mainly in VLDL (36-64% of totalastaxanthin), whereas LDL and HDL contained 29% and 24% of totalastaxanthin, respectively. The astaxanthin isomer distribution in theseVLDL, LDL, and HDL was not affected by time.
Astaxanthin Applications in Animal Husbandry
Consumer acceptance of farmed animal products depends to somedegree on appearance (e.g., coloration). Astaxanthin cannot be synthe-sized by animals and it must be provided in the diet. Lorenz and Cysewski(2000) have recently reviewed the applications of astaxanthin in animalhusbandry. The main applications of astaxanthin enriched feeds are tocolor the flesh, skin and even eggs of salmon, trout, red sea bream, shrimpand chicken. However, beyond the coloring effect, astaxanthin has beenfound to offer numerous benefits in these organisms such as
· Increased reproductive performance· Increased rate of growth (weight gain)· Enhanced immune response· Provitamin-A activity in fishes.New applications for astaxanthin are being worked on. For example,
according to Astacarotene AB, astaxanthin supplementation may increasethe fertility of horses, pigs and fur animals (http://www.astacarotene.se/).

146 MARINE BIOTECHNOLOGY
Astaxanthin may also serve to enhance the effect of anti-stress agentsadministered to farm animals (Ito et al. 1999).
Astaxanthin Applications in Human Health
Electron-stealing molecules known as free radicals (e.g., hydroxyl andperoxyl radicals) as well as highly reactive forms of oxygen (e.g., singletoxygen) are produced in the body during various normal metabolic reac-tions and processes. Physiological stress, air pollution, tobacco smoke,exposure to chemicals, or exposure to UV light, can enhance the produc-tion of these unwanted oxidizing agents (Papas 1999). Phagocytesinvolved in the immune response against microorganisms can also gener-ate an excess of free radicals to aid in their defensive degradation of aninvader. Within cells, free radicals can damage DNA, proteins, and lipidmembranes. This oxidative damage has been linked to ageing (Harman1981, Ames et al. 1993), atherogenesis (Steinberg et al. 1989, Francis 2000),ischemia-reperfusion injury (Simpson and Lucchesi 1987, Takayama et al.1992), infant retinopathy (Phelps 1987), age-related macular degeneration(Gerster 1991), and carcinogenesis (Moody and Hassan 1982, Marnett1987, Breimer 1990). Dietary antioxidants such as vitamin C or E andcarotenoids have been shown to help fight this oxidative damage both inin vitro and in vivo studies (Kurashige et al. 1990, Tinkler et al. 1994,Anderson et al. 1999). A direct consequence of these antioxidant proper-ties is the role of dietary antioxidants in helping prevent and fight anumber of human diseases, as shown in epidemiological studies as wellas in a number of animal model and human clinical studies (Seddon et al.1994, Ames 1998, Anderson et al. 1999, Biesalski 1999, Cooper et al. 1999,Hennekens 1999, Lyle et al. 1999).
Carotenoids are potent biological antioxidants that quench singletoxygen and other reactive species by absorbing the excited energy ofsinglet oxygen onto the carotenoid chain, leading to the degradation ofthe carotenoid molecule, but preventing other molecules or tissues frombeing damaged (Tinkler et al. 1994, Mortensen et al. 1997). Astaxanthin�sability to quench singlet oxygen and scavenge free radicals has beendemonstrated by a number of in-vitro and in-vivo studies (Reviewed inGuerin et al. 2003). Astaxanthin shows very good capability at protectingmembranous phospholipids and other lipids against peroxidation (Terao1989, Miki 1991, Palozza and Krinsky 1992, Lim et al. 1992, Naguib 2000).Most studies show that astaxanthin is a stronger antioxidant than vita-min E and other carotenoids like b-carotene or lutein. In fact, astaxanthinhas been shown to have up to 500-fold stronger free radical antioxidantactivity than vitamin E and 38 times more than b-carotene (Kurashige

RECENT ADVANCES IN COMMERCIAL PRODUCTION 147
et al. 1990, Shimidzu et al. 1996). The antioxidant properties of astaxanthinare believed to play a key role in a number or other properties such asprotection against UV-light photooxidation, inflammation, cancer, ulcer-ous Helicobacter pylori infection, ageing, and age-related diseases, or thepromotion of the immune response, heart health, eye health, liver func-tion, joint health, and prostate health. For a recent review of astaxanthin�sapplications in human health see Guerin et al. (2003).
Development of Microalgal (and Haematococcus)Cultivation Technology
Humans have consumed microalgae from natural populations through-out history. For example, Nostoc has long been consumed in Asia. Also,historical records indicate that when the Spanish arrived in Tenochtitlan,today�s Mexico City, in the 16th century they found the local populationharvesting an ooze (Spirulina) from the shores of Lake Texcoco fromwhich they produced small cakes for human consumption. Similarly, thetribes from Lake Rombou (Republic of Chad) in Northern Africa wereknown to harvest wild populations of Spirulina (Ciferri 1983). One canalso imagine collateral microalgal consumption in coastal communitiesvia consumption of filter feeders (e.g., clams and mussels) or epiphyteson macroalgae and other marine organisms.
On the other hand, scientists have conducted laboratory research onthe cultivation of microalgae for little over a hundred years whileattempts at large-scale cultivation are even more recent. Here, we willnote some of the significant historical events that have led to the develop-ment of commercial mass cultures without which commercial cultivationof Haematococcus would not have been possible.
In the 17th century, Anton van Leeuwenhoek was probably the firstindividual to observe and describe microalgae using simple one-lensmicroscopes. During the 17th, 18th and 19th centuries the first experimentsthat would establish the basis for plant growth and nutrition were beingcarried out by the likes of J.B. van Helmont, S. Hales, J. Priestley,J. Ingenhouszm, J. Senebier, T. de Saussure, and J.R. Mayer.
Early research on microalgae suggested that nitrogen and phosphoruswere necessary nutrients for their growth (Famintzin 1871) and that theyuse the same nutrients as plants. In 1890, Beijerink was able to establishbacteria-free cultures of Chlorella. By eliminating other microorganismsfrom the algal cultures, studies specifically designed to understand thenutrition, growth and metabolism of microalgae could be carried out(Chick 1903). Once these basic concepts were understood it became anissue of scaling up the cultivation systems.

148 MARINE BIOTECHNOLOGY
In 1905, the first cultures of marine microalgae were established inPlymouth, UK. Culture scale was 60 mL. In the 1930s 26-liter scale cul-ture of a marine diatom was accomplished at the Woods Hole Oceano-graphic Institution (Ketchum and Redfield 1938). They were able to growenough material to make chemical measurements on the biomass. Duringthe 1940s the first automated and continuous photobioreactors at labora-tory scale were developed by Myers and Clark (1944). Photobioreactorscale increased to 70 liters (Retovsky 1946). By 1949 continuous culturewas achieved in the laboratory. Building on their earlier techniques,Ketchum and Redfield (1949) demonstrated how to achieve optimumyields by continuously adding fresh nutrient medium and harvesting.
At about this time the first economic analysis of large-scale culturewas carried out at the Stanford Research Institute with funding from theCarnegie Institution. It was concluded that Chlorella could be grown as acheap source of protein. This work stimulated interest worldwide, andled to numerous pilot plants (see Table 1 in Goldman 1979) utilizingopen systems of, mainly, two types: paddlewheel-mixed raceway ponds(Dortmund, Germany) or round ponds (Tamiya 1957). In the ensuingdecades, open-pond systems have become larger and more successful atculturing Chlorella (supported by frequent inoculation from indoor grownmaterial) and other species that can withstand extreme culture conditions(Spirulina and Dunaliella) which inhibit competition from contaminatingorganisms. Efforts to grow other commercially interesting species failedbecause of contamination. In the 1960s, 70s and 80s the first commercialproducers of Chlorella, Spirulina and Dunaliella were established. Thus,after three decades of research on a wide variety of microalgae, onlythese three have been successfully produced at commercial scale. A fewother microalgae�e.g., Isochrysis, Chaetoceros�used in aquacultureoperations are grown in a similar way, using massive inoculations fromindoor grown material. Contamination and control are frequently citedin the literature as the main barriers to commercialization of microalgalproduction. As far as we know, large-scale single-phase open pond sys-tems have proved unsatisfactory for production of Haematococcus (Bubrick1991, Margalith 1999). A variety of authors have agreed on the limits ofopen pond systems and suggested that enclosed outdoor photobioreactortechnology is the key to increase the number of species commerciallycultivated (e.g., Vonshak 1992, Chaumont 1993, Borowitzka 1995). It wasnot until the 1990s when the first commercial production of a new algalspecies would become established using enclosed photobioreactor tech-nology (Olaizola 2000).
As mentioned earlier, enclosed photobioreactors (EPB) of up to tens ofliters capacity had been built by the 1940s (Retovsky 1946). The develop-ment of EPB technology has recently been summarized by Tredici (1999).
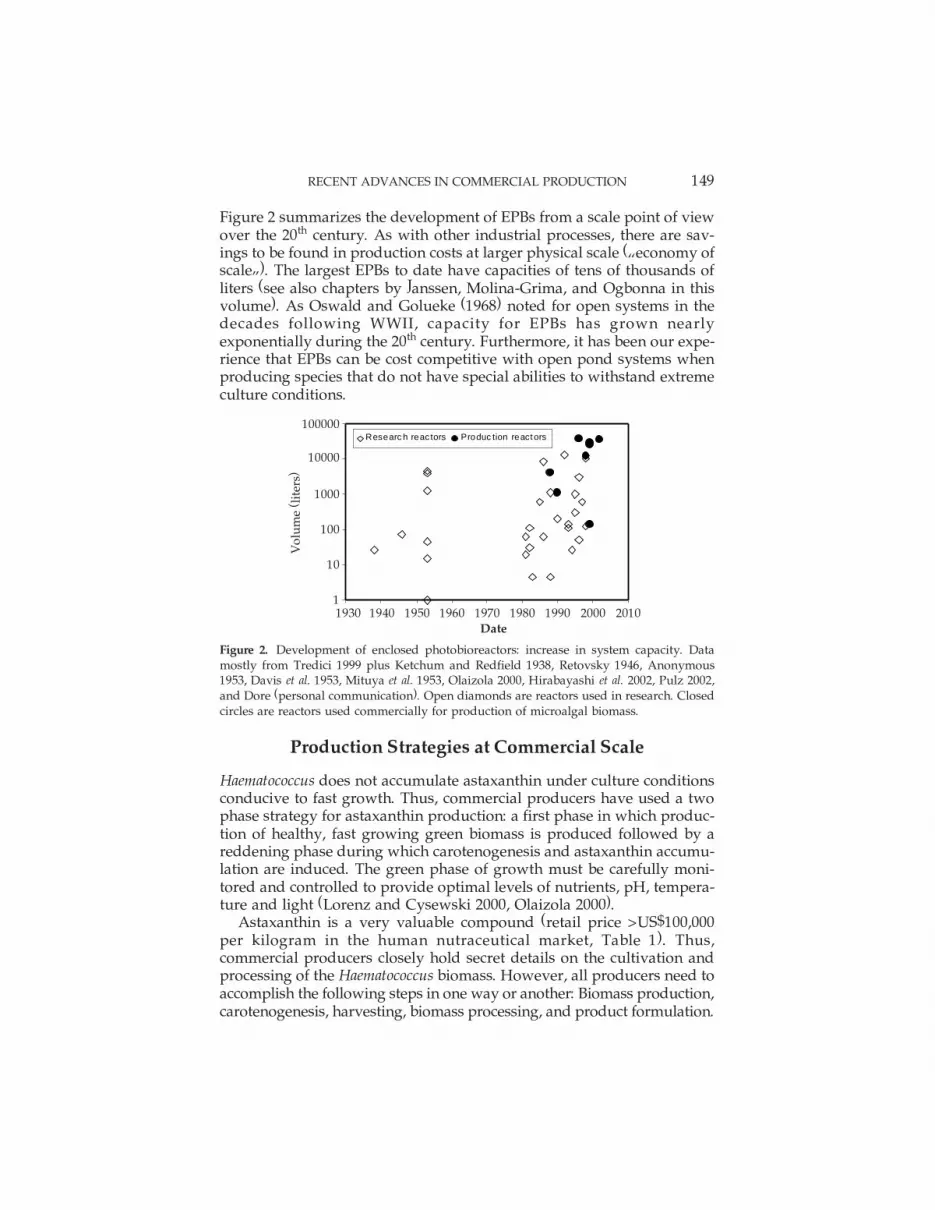
RECENT ADVANCES IN COMMERCIAL PRODUCTION 149
Figure 2 summarizes the development of EPBs from a scale point of viewover the 20th century. As with other industrial processes, there are sav-ings to be found in production costs at larger physical scale (�economy ofscale�). The largest EPBs to date have capacities of tens of thousands ofliters (see also chapters by Janssen, Molina-Grima, and Ogbonna in thisvolume). As Oswald and Golueke (1968) noted for open systems in thedecades following WWII, capacity for EPBs has grown nearlyexponentially during the 20th century. Furthermore, it has been our expe-rience that EPBs can be cost competitive with open pond systems whenproducing species that do not have special abilities to withstand extremeculture conditions.
Figure 2. Development of enclosed photobioreactors: increase in system capacity. Datamostly from Tredici 1999 plus Ketchum and Redfield 1938, Retovsky 1946, Anonymous1953, Davis et al. 1953, Mituya et al. 1953, Olaizola 2000, Hirabayashi et al. 2002, Pulz 2002,and Dore (personal communication). Open diamonds are reactors used in research. Closedcircles are reactors used commercially for production of microalgal biomass.
Researc h reac tors Produc tion react ors100000
10000
1000
100
10
11930 1940 1950 1960 1970 1980 1990 2000 2010
Date
Vol
ume
(lit
ers)
Production Strategies at Commercial Scale
Haematococcus does not accumulate astaxanthin under culture conditionsconducive to fast growth. Thus, commercial producers have used a twophase strategy for astaxanthin production: a first phase in which produc-tion of healthy, fast growing green biomass is produced followed by areddening phase during which carotenogenesis and astaxanthin accumu-lation are induced. The green phase of growth must be carefully moni-tored and controlled to provide optimal levels of nutrients, pH, tempera-ture and light (Lorenz and Cysewski 2000, Olaizola 2000).
Astaxanthin is a very valuable compound (retail price >US$100,000per kilogram in the human nutraceutical market, Table 1). Thus,commercial producers closely hold secret details on the cultivation andprocessing of the Haematococcus biomass. However, all producers need toaccomplish the following steps in one way or another: Biomass production,carotenogenesis, harvesting, biomass processing, and product formulation.

150 MARINE BIOTECHNOLOGY
Table 1. A sampling of nutraceutical astaxanthin retailers. The average retail price forastaxanthin, based on information at these websites, is >US$100,000 kg�1.
Retailer Web address
Aina Hawaiian Tropical http://www.hawaiitropicals.com/astazain.htmlProductsAquasearch, AstaFactor http://www.store.yahoo.net/astafactor/Biorejuvenate http://www.biorejuvenate.com/prod.htmlCyanotech, Bioastin http://www.buynutrex.com/cgi-bin/shop.cgi?key=10Source Naturals http://www.nutrimart.com/exe-bin/queries/detail.idc?ItemNo=SN1277Swanson Health http://www.swansonvitamins.com/Products, SolgarSwanson Health http://www.swansonvitamins.com/Products, Swanson UltraVision Works, Bioastin http://www.visionworksusa.com/products.asp?p_num=49Vitamin ProShop, Solgar http://www.vitaminproshop.com/solascom4mg6.htmlZanaMaxx http://www.answertocancer.com/zanamaxx.htm
Biomass Production
Haematococcus biomass can be produced photoautotrophically orheterotrophically at commercial scale. So far, there has been one successfulproducer that uses a mixotrophic-based indoor system (Astacarotene AB,www.astacarotene.se, Sweden). Cultivation of the algae takes place in asemi-closed system comprising stainless steel tanks of 2 to 30 m3 capacitywith light tubes submerged in the cultivation media. The advantage ofthe system over those run outdoors is the possibility to maintain highhygiene standards and to closely control the cultivation parameters andproduct quality (J. Inborr, personal communication).
The other three successful producers to date use autotrophic-basedoutdoor systems:
Cyanotech Corp. (www.cyanotech.com, Hawaii, USA). Cyanotech usesdouble channel enclosed photobioreactors (Phytodome) of 28,000 litercapacity (J. Dore, personal communication, Figure 3) and open ponds toproduce green biomass. Production scale-up of green Haematococcusbiomass is accomplished by placing more Phytodomes and open pondsinto operation. It is not known how many Phytodomes and ponds areused at any one time . However, we can assume similar area productivitiesto other outdoor Haematococcus photobioreactors (Olaizola 2000). If weconsider that Cyanotech�s production plant is about 30 times larger thanMera Pharmaceuticals� (see below), but that only a fraction of it is dedi-cated to astaxanthin production, the total production of green biomasscould be as high as 13-15 tons yr�1. Cyanotech is the largest producer inthis industry. The green biomass produced is used to inoculate openponds for carotenogenesis (Lorenz and Cysewski 2000).

RECENT ADVANCES IN COMMERCIAL PRODUCTION 151
Mera Phamaceuticals, Inc. (www.merapharma.com, Hawaii, USA, formelyAquasearch, Inc.). Mera Pharmaceuticals uses computer controlledserpentine type tubular photobiorreactors (Mera Growth Module or MGM,Figure 4) of 25,000 liter capacity and occupying an area of about 100 m2
each. Production scale-up is accomplished by placing more MGMs intooperation. The average productivity of this system is nearly 15 g m�2 d�1
of healthy green (near zero astaxanthin content) Haematococcus biomass(Olaizola 2000). The production plant has infrastructure to support asmany as 12 MGMs for a total capacity of up to 18 kg d�1 (6.6 tons yr�1).The green biomass produced is used to inoculate open ponds wherestressing and carotenogenesis takes place.
Micro Gaia, Inc. (www.microgaia.com, Hawaii, USA). Micro Gaia usessmaller enclosed photobioreactors (Biodome) of 140 liter capacity andoccupying 1 m2 surface area (Figure 5). The biodomes are built by boltingtogether 2 hemispherical transparent surfaces. The result is a hemispheri-cal layer of culture, 5 cm thick, which enhances light capture (Hirabayashiet al. 2002). As of early 2002, Micro Gaia has 300 Biodomes in operationdedicated to green biomass production. The productivity of each Biodomeis not known but, assuming similar area productivities to those of the
Figure 3. Cyanotech�s Phytodome (Courtesy of J. Dore, Cyanotech Corp.).

152 MARINE BIOTECHNOLOGY
Figure 4. The Mera Growth Module.
Figure 5. Micro Gaia�s Biodome with green cells (Courtesy of A. Diffley, Micro Gaia, Inc.).
previous systems, we can estimate a total production of about 1.64 tonsyr�1. The green biomass produced is used to inoculate other Biodomeswhere carotenogenesis is induced.

RECENT ADVANCES IN COMMERCIAL PRODUCTION 153
Minapro Ltd. (www.minapro.com, Israel). Minapro uses smaller yet closedphotobioreactors (hanging sleeves) according to information on theirwebsite. No details are available on the capacity or productivity of theirsystem.
There are two other producers, that we are aware of, nearingcommercialization:
Algatechnologies Ltd. (Israel) will use a two step photoautotrophicsystem to produce green Haematococcus biomass. The first step will consistof vertical flat panel reactors in greenhouses followed by scale up inoutdoor tubular EPBs (Boussiba 2002). No data on the productivity orcapacity of this system is known at this time.
BioProcess Island hf (www.bioprocess.is, Iceland) intends to utilize amixotrophic approach by providing both fixed carbon and light to theircultures. Their prototype indoor reactors have a capacity of 10,000 liters.No details are available on the productivity of this system. They intendto take advantage of inexpensive electric power (geothermal, hydro)available in Iceland (N.-H. Norsker, personal communication).
Carotenogenesis Induction
Carotenogenesis and astaxanthin accumulation are induced inHaematococcus by changing the culture conditions from growth-promotingto stress-promoting: e.g., nutrient stress, light stress, salt stress, temperaturestress, etc. (see references in Olaizola 2000). It is not clear how mixotrophi-cally grown Haematococcus is induced to accumulate astaxanthin atcommercial scale. However, Astacarotene�s output averages 200 g d�1 m�3
biomass with a 3% astaxanthin content (J. Inborr, personal communication).For autotrophically grown Haematococcus the easiest way to accomplish
a combination of stresses is to simply dilute the green culture withfreshwater in combination with any additional stress/inducement factors(such as acetate, iron, salt, heat, etc). Dilution quickly accomplishes twothings: it immediately lowers the nutrient concentration in the mediumand increases the amount of irradiance that each cell receives. This can bedone either in EPBs or open ponds. Micro Gaia uses their Biodome systemfor the reddening phase (Figure 6). For the 300 Biodomes used for greenbiomass production, they use 700 reddening Biodomes whereHaematococcus accumulates astaxanthin at up to 3.3% of the dry weight(Hirabayashi et al. 2002). Algatechnologies is expected to also use EPBsfor the reddening phase (Boussiba 2002) but no details are available as tocapacity or productivity.
Both Cyanotech and Mera Pharmaceuticals use open ponds for thereddening phase. While open pond technology is not adequate forvegetative production of Haematococcus (green phase) because of the

154 MARINE BIOTECHNOLOGY
impossibility to maintain unialgal cultures, it is quite appropriate for thecarotenogenesis phase. This is so for two reasons. First, commercialproducers have come up with stress strategies that will develop fullyreddened Haematococcus aplanospores within 5-6 days (too short for acontaminant to impact the productivity of the system). Second, thereddening phase is characterized by culture conditions unsuitable for fastgrowth, including that of any possible contaminant.
Cyanotech utilizes 500,000 liter capacity ponds (Figure 7) to, after 5-6days, obtain Haematococcus biomass that has an astaxanthin content of >1.5% (Cysewski and Thomason 2002). Mera Pharmaceuticals utilizes 90,000liter capacity ponds for the reddening phase. After 5-6 days (Figure 8),the Haemtococcus biomass has an astaxanthin content of >3.0% of the dryweight (Olaizola 2000).
Harvesting
The Haematococcus aplanospores are significantly denser than the me-dium (settling velocity is about 1 cm min�1, unpublished observation) soharvesting is easily accomplished by gravitational settling either in thecultivation units or in special tanks. Further concentrating is usually donewith the aid of centrifuges (Lorenz and Cysewski 2000, Olaizola 2000).
Figure 6. Micro Gaia�s Biodome with red aplanospores (Courtesy of A. Diffley, Micro Gaia,Inc.).

RECENT ADVANCES IN COMMERCIAL PRODUCTION 155
Figure 7. 500,000 liter ponds used by Cyanotech for reddening of Haematococcus biomass(Courtesy of J. Dore, Cyanotech Corp.).
Biomass Processing
One of the characteristic responses of Haematococcus to stressing condi-tions is, besides astaxanthin accumulation, the generation of a very toughcell wall (i.e., a cyst, Mendes-Pinto et al. 2001 and references therein).This wall resists digestion by either animals (in feed applications) or
Figure 8. 90,000 liter ponds used by Mera Pharmaceuticals for reddening of Haematococcusbiomass. From left to right, ponds are just inoculated (day 0) or 1, 2, 3, and 4 days old (i.e.,ready for harvest).

156 MARINE BIOTECHNOLOGY
humans (in nutraceutial applications). Thus, for the consumers to obtainthe benefit of astaxanthin supplementation, the cysts must be broken up.
Mera Pharmaceuticals uses a homogenizer that pressurizes the har-vested biomass to >700 bars as the material is passed through a cell-disrupting homogenizing valve (Olaizola 2000). This method thoroughlyruptures >90% of the cells.
It is not known what techniques other commercial producers use todisrupt the Haematococcus cells. Ball mills can be used to disruptHaematococcus cells at commercial scale but we have found them to beless efficient than the homogenizer (unpublished observations). Our ownexperience indicates that enzyme and solvent pre-treatments may behelpful (unpublished data). However, one must be careful regarding whataids are used especially if the product is intended for human consumption.
The next step in processing the Haematococcus biomass is to reduce thewater content of the now broken cell biomass. At this point in the process,the astaxanthin in the product is very sensitive to degradation. Theastaxanthin molecules are no longer protected by the cyst structure.Because astaxanthin is such a good antioxidant it is also very reactive socare must be taken to reduce exposure to light, oxygen and hightemperature as much as possible. Our own experience indicates that useof hot spray dryers (nearly 200°C) can reduce the yield of the operationsignificantly (20-40%, unpublished observation). Cyanotech uses apatented technique to decrease the partial pressure of oxygen in a spraydryer to limit losses (Cysewski 1994). Mera Pharmaceuticals usesproprietary technology that allows for very gentle drying of the biomassat <100°C. With this new technology, less than 1% of the astaxanthin islost during drying. We are not aware of what drying technology otherproducers use.
End Products and Formulation
After drying, the Haematococcus biomass is in the form of small particles.For animal husbandry applications the dried product can be mixed withapproved preservatives such as ethoxyquin. The product can be directlyshipped to the feed manufacturer where it is incorporated into their for-mulations.
The dried product can also be used directly in the human nutraceuticalfield by pressing it into tablets (much like Spirulina and Chlorella tabletsare produced). However, because of exposure of astaxanthin to theatmosphere in such tablets, and consequent degradation, this is not thepreferred formulation. Both Cyanotech and Mera Pharmaceuticals haveattempted to produce suspensions of dried Haematococcus in oil for thehuman market since oil would create a barrier between atmospheric

RECENT ADVANCES IN COMMERCIAL PRODUCTION 157
oxygen and astaxanthin. Cyanotech tried suspensions in rosemary oil butfound that astaxanthin was very unstable in this formulation (Cysewskiand Thomason 2002). Our own observations are that dried particlessuspended in oil can cause leaks in gelatin capsules resulting in a productunacceptable to the consumer.
The solution to these formulation problems has been the developmentof extraction methodologies using non-petrochemical solvents. Forhuman applications, use of petrochemical solvents could create healthand/or acceptability concerns because of possible residues in the finalproduct. Mera Phamaceuticals has developed a proprietary oil extractionmethod in which edible oils are used as the extraction solvent. Alterna-tively, super-critical CO2 extraction can be used to produce an astaxanthin-rich oleoresin that is then diluted with edible oils to the appropriateconcentration for encapsulation (Cysewski and Thomason 2002). Most ofthe available retail astaxanthin products for human use in the markettoday are formulated with astaxanthin extracts (Table 1).
Astaxanthin Markets, Competition fromSynthetic Astaxanthin
The largest producers of astaxanthin in the world market today do notproduce astaxanthin from Haematococcus but via chemical synthesis (BASF,Hoffman-La Roche) and have established the world market price forastaxanthin at about US$2000-2500 kg�1. We suspect that the actual pro-duction cost for synthetic astaxanthin is <US$1000 kg�1. For naturalHaematococcus astaxanthin to be produced at such cost, and assuming 3%astaxanthin content of the biomass, it would be necessary to produceHaematococcus biomass at significantly less than US$30 kg�1. Considering,as opposed to, e.g., Spirulina, the added cost of processing theHaematococcus biomass we feel that this low cost cannot be achieved byany of the commercial producers listed in the text. However, as the pro-duction technology is optimized and transferred to lower-cost localesHaematococcus astaxanthin will become cost competitive.
Feed Astaxanthin
The largest consumer of astaxanthin today is the salmon feed industry. Arecent study by the FDA found that 3S,3¢S is the main stereoisomer foundin wild Pacific and Atlantic salmon species. The study concluded that, inorder to achieve the same astaxanthin stereoisomer profile as their wildcounterparts, farmed salmon should be fed a diet containing the sameastaxanthin stereoisomers as in the natural diet of wild salmon (Turujman

158 MARINE BIOTECHNOLOGY
et al. 1997). The 3S,3¢S stereoisomer is the main form found inHaematococcus, while synthetic astaxanthin contains primarily the 3R,3¢Sstereoisomer. Salmon appear unable to convert the 3R,3¢S stereoisomerinto the more common 3S,3¢S form. In fact, the FDA study showed thatfarmed salmon could be easily distinguished from the wild salmon be-cause the farmed salmon are fed synthetic astaxanthin and accumulateastaxanthin isomers in the flesh in the same ratio as is found in their diet.Why does the salmon feed industry, then, use synthetic astaxanthin? Theanswer is that the public, at large, does not demand and is not willing topay a higher price for naturally pigmented salmon. We feel this is sobecause of lack of awareness by the consumer. Most consumers probablydo not realize that most of the salmon consumed today is farmed andpigment is added to their diets. Furthermore, consumers may not beaware that the pigment added to the diet is a synthetic product. As longas the consumer is uninformed there will be very little demand for natu-ral astaxanthin-fed salmon and astaxanthin producers will have to com-pete on price alone. Once the consumer is educated, or regulations favorthe use of natural products, we would expect to see a premium price fornatural versus synthetic astaxanthin as has occurred in the vitamin E andb-carotene market (Bahner 1993a,b).
We are aware of a few feed applications where natural astaxanthin ispreferred over synthetic astaxanthin (koi, chicken, red seabream diets)because of enhanced deposition of pigment in the tissues or regulatoryrequirements. However, these markets, at the present time, appear quitesmall and will not be discussed further here.
Nutraceutical Astaxanthin
Because of the possible functions of astaxanthin in the human body (Guerinet al. 2003) a market for nutraceutical astaxanthin is starting to develop.As far as we know, all Haematococcus astaxanthin producers haveintroduced astaxanthin products for human consumption (e.g., Table 1).Because of the much higher price obtainable for nutraceutical astaxanthin,the higher costs of producing Haematococcus astaxanthin are justified.While the size of the nutraceutical astaxanthin market is closely guardedby commercial producers we expect that, as the public learns of the healthbenefits of astaxanthin supplementation (Guerin et al. 2003), the marketcould reach a size of several hundred million US$ within 5-10 years.Because of the perceived negatives of consuming a synthetic product inthe human nutraceutical market we also expect no competition from syn-thetic astaxanthin in this market.

RECENT ADVANCES IN COMMERCIAL PRODUCTION 159
4.0%
3.0%
2.0%
1.0%
0.0%
Ast
axan
thin
% o
f bi
omas
s
1998 1999 2000 2001 2002Year
Future of Haematococcus Astaxanthin
We believe that through consumer education and lowering of the pro-duction costs the future of Haematococcus astaxanthin is bright indeed.We are pursuing two strategies to lower our production costs.
First, through our own internal (unpublished) research, we arecontinuously improving our production technology to produceHaematococcus biomass with a higher final astaxanthin content (Figure 9).As noted elsewhere (Olaizola 2000 and references therein) Haematococcushas been shown to accumulate >6% of its dry weight as astaxanthin inlaboratory scale cultures. Thus, we suggest that commercial scale cultureshave the potential to reach 50-100% higher astaxanthin content that thatachieved so far at commercial scale. Any improvement in astaxanthincontent of the final product translates directly into lower costs since thecosts are proportional to the amount of biomass produced and processed.
Second, we plan to lower our production costs by expanding ourproduction capacity into locales with lower land, labor, and energy costssuch as China. We suspect that other producers of Haematococcusastaxanthin will need to accomplish similar changes to remain competitive.
Summary
Astaxanthin, a carotenoid pigment, has important applications in thenutraceutical, cosmetics, food and feed industries. Astaxanthin is both astrong coloring agent and a potent antioxidant. Astaxanthin�s strong
Figure 9. Changes in astaxanthin content of Haematococcus biomass produced at MeraPharmaceutical�s Hawaii facility. The bars represent the yearly averages. The error bars areone standard deviation wide.

160 MARINE BIOTECHNOLOGY
antioxidant activity points to its potential to target a number of healthconditions.
Haematococcus has been identified as the richest source of naturalastaxanthin and is now being cultivated at industrial scale by a handfulof companies. Production of Haematococcus biomass requires enclosedphotobioreactors for at least part of the growth cycle. Because of therequirements for benign weather and ample sunlight, most production iscarried out in the tropics (e.g., Hawaii). Haematococcus accumulatesastaxanthin mostly under conditions that are not conducive to growth,thus commercial producers have adopted a two phase strategy: greenbiomass production under conditions of nutrient sufficiency and lightmanagement to avoid photoinhibition followed by a stress phase toinduce carotenogenesis. Alternatively, a few producers have devised pro-duction systems based on heterotrophic growth and artificial light, thusindependent of the ambient light field.
Cost of production are higher than for other whole algal products(e.g., Spirulina) since the biomass needs to be further processed to increasebioavailability of astaxanthin. Producers of synthetic astaxanthin nowsupply most of the world market, especially in feed coloring applications.Since natural astaxanthin cannot compete with the synthetic version onprice yet, producers are establishing themselves in specialized marketswhere the natural product has advantages such as in human nutraceuticalsand specialty animal feeds.
Acknowledgements
The authors thank Dr. John Dore for providing the photographs ofCyanotech�s Phytodome and reddening ponds and Mr. Alex Diffley forproviding photographs of Micro Gaia�s Biodomes. We also thankDr. J. Dore and Dr. M. Lopez for reviewing an earlier version of thismanuscript.
REFERENCES
Ames, B.N. 1998. Micronutrients prevent cancer and delay aging. Toxicol. Lett. 102-103: 5-18.
Ames, B.N., M.K. Shigenaga and T.M. Hagen. 1993. Oxidants, antioxidants, and thedegenerative diseases of ageing. Proc. Natl. Acad. Sci. USA. 90: 7915-22.
Anonymous. 1953. Pilot-plant studies in the production of Chlorella. Pages 235-272 in AlgalCulture from Laboratory to Pilot Plant, J.S. Burlew, ed., Carnegie Institution of Wash-ington Publication No. 600, Carnegie Institution, Washington, D.C.
Almgren, K. 1966. Ecology and distribution in Sweden of algae belonging toHaematococaceae. Svensk. Botanisk. Tidskrift. BD.60, H.1, 49-73.

RECENT ADVANCES IN COMMERCIAL PRODUCTION 161
Anderson, J., M. Gowri, J. Turner, L. Nichols, V.A. Diwadkar, C.K. Chow and P.R. Oeltgen.1999. Antioxidant supplementation effects on low-density lipoprotein oxidation forindividuals with Type 2 diabetes mellitus. J. Am. Coll. Nutr. 18: 451-461.
Beijerinck, M.W. 1890. Kulturversuche mit Zoochlorellen, Lichengonidien und anderenniederen Algen. Bot. Zeit. 48: 725-739, 741-754, 757-768, 781-785.
Biesalski, H.K. 1999. The role of antioxidative vitamins in primary and secondary preven-tion of coronary heart disease. Int. J. Vitam. Nutr. Res. 69: 179-86.
Borowitzka, M.A. 1995. Microalgae as sources of pharmaceuticals and other biologicallyactive compounds. J. Appl. Phycol. 7: 3-15.
Boussiba, S. 2002. Production of carotenoids by microalgae with special emphasis onastaxanthin from the green alga Haematococcus pluvialis. 13th International Caro-tenoid Symposium, Honolulu, Hawaii, January 6th January 11th, 2002.
Bubrick, P. 1991. Production of astaxanthin from Haematococcus. Bioresource Technol. 38:237-239.
Breimer, L.H. 1990. Molecular mechanisms of oxygen radical carcinogenesis and mutagen-esis: the role of DNA base damage. Mol. Carcinog. 3: 188-197.
Chaumont, D. 1993. Biotechnology of algal biomass production: a review of systems foroutdoor mass culture. J. Appl. Phycol. 5: 593-604.
Chemical Marketing Reporter. 1993. Schnell Publishing Co., Inc., NY. Aug. 30, 1993 andOct. 11, 1993.
Chick, H. 1903. A study of a unicellular green alga, occurring in polluted water, withspecial reference to its nitrogenous metabolism. Proc. Roy. Soc. Lond. 71: 458-476.
Ciferri, O. 1983. Spirulina, the edible microorganism. Microbiol. Rev. 47: 551-578.Cooper, D., A. Eldridge and J.C. Peters. 1999. Dietary carotenoids and certain cancers,
heart disease, and age-related macular degeneration: a review of recent research.Nutr. Rev. 57: 201-14.
Cysewsky, G.R. 1994. Ocean-chill drying of microalgae and microalgal products. US Patent#5,276,977. Cyanotech Corp., USA.
Cysewski, G.R. and M.A. Thomason. 2002. Commercial potential for Haematococcusmicroalgae as a natural source of astaxanthin. 13th International Carotenoid Sym-posium, Honolulu, Hawaii, January 6th January 11th, 2002.
Dall W. 1995. Carotenoids versus retinoids (Vitamin A) as essential growth factors inpenaeid prawns (Penaeus semisulcatus). Mar. Biol. 124: 209-213.
Davis, E.E., J. Dedrick, C.S. French, H.W. Milner, J. Myers, J.H.C. Smith and H.A. Spoehr.1953. Laboratory experiments on Chlorella culture at the Carnegie Institution ofWashington Department of Plant Biology. Pages 105-153 in Algal Culture from Labo-ratory to Pilot Plant, J.S. Burlew, ed., Carnegie Institution of Washington PublicationNo. 600, Carnegie Institution, Washington, D.C.
Droop, M.R. 1953. On the ecology of flagellates from some brackish and fresh waterrockpools of Finland. Acta Bot. Fenn. 51: 1-52.
Droop, M.R. 1961. Haematococcus pluvialis and its allies. III. Organic nutrition. Rev. Algol.N.S.5: 247-259.
Egeland E.S. 1993. Carotenoids in combs of capercaillie (Tetrao urogallus) fed defined diets.Poult. Sci. 72: 747-751.
Famintzin, A. 1871. Die anorganische Salze als ausgereichnetes Hülfsmittel zum Studiumder Entwicklung niederer chlorophyllhaltiger Organismen. Bull. Acad. Sci. St. Peters-burg 17 (S31).
Francis, G.A. 2000. High density lipoprotein oxidation: in vitro susceptibility and potentialin vivo consequences. Biochim. Biophys. Acta 1483: 217-235.
Fujita T., M. Satake, T. Watanabe, C. Kitajima, W. Miki, K. Yamaguchi and S. Konosu.1983. Pigmentation of cultured red seabream with astaxanthin diester purified fromKrill oil. Bull. Jap. Soc. Sci. Fish. 49: 1855-1861.
Furr, H.C. and R.M. Clark. 1997. Intestinal absorption and tissue distribution of caro-tenoids. Nutr. Biochem. 8: 364-377.

162 MARINE BIOTECHNOLOGY
Gerster, H. 1991. Review: antioxidant protection of the ageing macula. Age Ageing 20: 60-69.
Goldman, J.C. 1979. Outdoor algal mass cultures. Water Res. 13: 1-19.Guerin, M., M. Huntley and M. Olaizola. 2003. Haematococcus astaxanthin: applications for
human health and nutrition. Trends Biotechnol. 21: 210-216.Harman, D. 1981. The ageing process. Proc. Natl. Acad. Sci. USA. 78: 7124-7128.Hennekens, C.H. 1999. Antioxidant vitamins and cardiovascular disease. Pages 463-477 in
Antioxidant Status, Diet, Nutrition, and Health, C. H. Hennekens, ed., CRC Press, BocaRaton, Florida.
Hirabayashi, S., H.T. Gevarra, T. Sato, N. Nakatsuka, S. Sakurai and Y. Tsuchiya. 2002.Production of astaxanthin from the green alga Haematococcus pluvialis in closedphoto bioreactors. 13th International Carotenoid Symposium, Honolulu, Hawaii,January 6th January 11th, 2002.
Inborr, J. 1998. Haematococcus the poultry pigmentor. Feed Mix 6: 31-34.Ito, Y., T. Kamata, Y. Tanaka and M. Sameshima. 1986. Studies on the improvement of
body color of red seabream Pagrus major by astaxanthin and astaxanthin dipalmitate.Aquaculture 34: 77-80.
Ito, S., E. Ogata and M. Yamada. 1999. Anti-stress agent for animals and a method ofreducing stress in animals, United States Patent # 5,937,790. Showa Denko KabushikiKaisha (Tokyo, Japan), United States.
Jyonouchi, H., S. Sun and M. Gross. 1995. Effect of carotenoids on in vitro immunoglobulinproduction by human peripheral blood mononuclear cells: Astaxanthin, a caro-tenoid without vitamin A activity, enhances in vitro immunoglobulin production inresponse to a T-dependent stimulant and antigen. Nutr. Cancer 23: 171-183.
Ketchum, B.H., L. Lillick and A.C. Redfield. 1949. The growth and optimum yields ofunicellular algae in mass culture. J. Cell. Comp. Physiol. 33: 267-280.
Ketchum, B.H. and A.C. Redfield. 1938. A method for maintaining a continuous supply ofmarine diatoms by culture. Biol. Bull. 75: 165-169.
Kurashige, M., E. Okimasu, M. Inoue and K. Utsumi. 1990. Inhibition of oxidative injury ofbiological membranes by astaxanthin. Physiol. Chem. Phys. Med. NMR 22: 27-38.
Lim, B., A. Nagao, J. Terao, K. Tanaka, T. Suzuki and K. Takama. 1992. Antioxidantactivity of xanthophylls on peroxyl radical-mediated phospholipid peroxidation.Biochim. Biophys. Acta 1126: 178-84.
Lorenz, R.T. and G.R. Cysewski. 2000. Commercial potential for Haematococcus microalgaeas a natural source of astaxanthin. Trends Biotechnol. 18: 160-167.
Lyle, B.J., J.A. Mares-Perlman, B.E. Klein, R. Klein and J.L. Greger. 1999. Antioxidantintake and risk of incident age-related nuclear cataracts in the Beaver Dam EyeStudy. Am. J. Epidemiol. 149: 801-809.
Margalith, P.Z. 1999. Production of ketocarotenoids by microalgae. Appl. Microbiol. Biotechnol.51: 431-438.
Marnett, L.J. 1987. Peroxyl free radicals: potential mediators of tumor initiation and pro-motion. Carcinogenesis 8: 1365-1373.
Menasveta P., W. Worawattanamateekul, T. Latscha and J.S. Clark. 1993. Correction ofBlack Tiger Prawn (Penaeus monodon Fabricius) coloration by astaxanthin. Aquac.Engin. 12: 203-213.
Mendes-Pinto, M.M., M.F.J. Raposo, J. Bowen, A.J. Young and R. Morais. 2001. Evaluationof different cell disruption processes on encysted cells of Haematococcus pluvialis:effects on astaxanthin recovery and implications for bioavailability. J. Appl. Phycol.13: 19-24.

RECENT ADVANCES IN COMMERCIAL PRODUCTION 163
Meyers, S.P. 1993. The biological role of astaxanthin in salmonids and other aquatic spe-cies. First Int. Symp. on Nat. Colors and Foods, Nutrac., Bever. and Confect., Amherst,USA, Nov. 7-10.
Miki, W. 1991. Biological functions and activities of animal carotenoids. Appl. Chem. 63:141-146.
Mituya, A., T. Nyunoya and H. Tamiya, H. 1953. Pre-pilotplant experiments on algal massculture. Pages 273-281 in Algal culture from Laboratory to Pilot Plant, J.S. Burlew, ed.,Carnegie Institution of Washington Publication No. 600, Carnegie Institution,Washington, D.C.
Moody, C.S. and H.M. Hassan. 1982. Mutagenicity of oxygen free radicals. Proc. Natl. Acad.Sci. U.S.A. 79: 2855-2859.
Mortensen, A., L.H. Skibsted, J. Sampson, C. Rice-Evans and S.A. Everett. 1997. Compara-tive mechanisms and rates of free radical scavenging by carotenoid antioxidants.FEBS Lett. 418: 91-97.
Myers, J. and L.B. Clark. 1944. Culture conditions and the development of the photosyn-thetic mechanism. II. An apparatus for the continuous culture of Chlorella. J. Gen.Physiol. 28: 103-112.
Naguib, Y.M.A. 2000. Antioxidant acitivities of astaxanthin and related carotenoids.J. Agric. Food Chem. 48: 1158-1154.
Olaizola, M. 2000. Commercial production of astaxanthin from Haematococcus pluvialisusing 25,000 liter outdoor photobioreactors. J. Appl. Phycol. 12: 499-506.
Østerlie, M., B. Bjerkeng and S. Liaaen-Jensen. 1999. Blood appearance and distribution ofastaxanthin E/Z isomers among plasma lipoproteins in humans administered asingle meal with astaxanthin. 12th International Carotenoid Symposium, Cairns,Australia.
Oswald, W.J. and C.G. Golueke. 1968. Large-scale production of algae. Pages 271-305 inSingle-Cell Protein, R.I. Matales and S.R. Tennenbaum, eds., MIT Press, Cambridge,MA.
Palozza, P. and N.I. Krinsky. 1992. Astaxanthin and canthaxanthin are potent antioxidantsin a membrane model. Arch. Biochem. Biophys. 297: 291-295.
Papas, A.M. 1999. Determinants of antioxidant status in humans. Pages 21-36 in Antioxi-dant Status, Diet, Nutrition and Health, A.M. Papas, ed., CRC Press, Boca Raton,Florida.
Phelps, D.L. 1987. Current perspectives on vitamin E in infant nutrition. Am. J. Clin. Nutr.46(suppl.): 187-191.
Pulz, O. 2002. Industrial scale photobioreactors. 1st Congress of the International Societyfor Applied Phycology, Almeria, Spain, May 26-May 30, 2002.
Retovsky, R. 1946. Mass cultures of some unicellular algae. Stud. Bot. Cechoslovaca 7: 38-48.Seddon, J.M., U.A. Ajani, R.D. Sperduto, R. Hiller, N. Blair, T.C. Burton, M.D. Farber,
E.S. Gragoudas, J. Haller, D.T. Miller, L.A. Yannuzzi and W. Willett. 1994. Dietarycarotenoids, vitamins A, C, and E, and advanced age-related macular degeneration.J. Am. Med. Assoc. 272:1413-1420.
Shimidzu, N., M. Goto and W. Miki. 1996. Carotenoids as singlet oxygen quenchers inmarine organisms. Fish. Sci. 62: 134-137.
Simpson, P.J. and B.R. Lucchesi. 1987. Free radicals and myocardial ischemia and reperfusioninjury. J. Lab. Clin. Med. 110: 13-30.
Steinberg, D., S. Parthsarathy, T.E. Carew, J.C. Khoo and J.L. Witztum. 1989. Beyondcholesterol. Modifications of low-density lipoprotein that increase its atherogenicity.New Engl. J. Med. 320: 915-924.
Takayama, F., T. Egashira, Y. Kudo and Y. Yamanaka. 1992. Chemiluminescence-HPLCassay of phosphatidylcholine hydroperoxide generated by ischemia-reperfusion inthe liver of rats. Biochem. Pharmacol. 44: 2412-2414.
Tamiya, H. 1957. Mass culture of algae. Ann. Rev. Plant Physiol. 8: 309-334.

164 MARINE BIOTECHNOLOGY
Terao, J. 1989. Antioxidant activity of beta-carotene-related carotenoids in solution. Lipids24: 659-61.
Tinkler, J.H., F. Böhm, W. Schalch and T.G. Truscott. 1994. Dietary carotenoids protecthuman cells from damage. J. Photochem. Photobiol. 26: 283-285.
Torissen O.J., R.W. Hardy and K. Shearer. 1989. Pigmentation of salmonids-carotenoiddeposition and metabolism. CRC Crit. Rev. Aquat. Sci. 1: 209-225.
Tredici, M. 1999. Bioreactors, Photo. Pages 395-419 in Encyclopaedia of Bioprocess Technology:Fermentation, Biocatalysis and Bioseparation, Vol 1, M.C. Flickinger and S.W. Drew,eds., John Wiley & Sons, New York.
Turujman S.A., W.G. Wamer, R.R. Wei and R.H. Albert. 1997. Rapid liquid chromato-graphic method to distinguish wild salmon from aquacultured salmon fed syntheticastaxanthin. J. AOAC Int. 80: 622-632.
Vonshak, A. 1992. Tubular photobioreactors for algal mass production; prospects andachievements. Isr. J. Aquacult./Bamidgeh. 44: 151.