natural igm prevents autoimmunity by enforcing b cell central tolerance induction
TRANSCRIPT

of June 12, 2016.This information is current as
InductionEnforcing B Cell Central Tolerance Natural IgM Prevents Autoimmunity by
BaumgarthTrang T. T. Nguyen, Rebecca A. Elsner and Nicole
http://www.jimmunol.org/content/194/4/1489doi: 10.4049/jimmunol.1401880January 2015;
2015; 194:1489-1502; Prepublished online 16J Immunol
MaterialSupplementary
0.DCSupplemental.htmlhttp://www.jimmunol.org/content/suppl/2015/01/16/jimmunol.140188
Referenceshttp://www.jimmunol.org/content/194/4/1489.full#ref-list-1
, 26 of which you can access for free at: cites 58 articlesThis article
Subscriptionshttp://jimmunol.org/subscriptions
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/ji/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/cgi/alerts/etocReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists, Inc. All rights reserved.Copyright © 2015 by The American Association of9650 Rockville Pike, Bethesda, MD 20814-3994.The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on June 12, 2016http://w
ww
.jimm
unol.org/D
ownloaded from
by guest on June 12, 2016
http://ww
w.jim
munol.org/
Dow
nloaded from

The Journal of Immunology
Natural IgM Prevents Autoimmunity by Enforcing B CellCentral Tolerance Induction
Trang T. T. Nguyen,*,† Rebecca A. Elsner,*,‡ and Nicole Baumgarth*,†,‡,x
It is unclear why selective deficiency in secreted (s)IgM causes Ab-mediated autoimmunity. We demonstrate that sIgM is required
for normal B cell development and selection. The CD5+ B cells that were previously shown to accumulate in body cavities of sIgM2/2
mice are not B-1a cells, but CD19int, CD432, short-lived, BCR signaling–unresponsive anergic B-2 cells. Body cavity B-1 cells were
>10-fold reduced, including VH11+ and phosphotidylcholine-specific B-1a cells, whereas splenic B-1 cells were unaffected and
marginal zone B cells increased. Follicular B cells had higher turnover rates, survived poorly after adoptive transfer, and were
unresponsiveness to BCR stimulation in vitro. sIgM bound to B cell precursors and provided a positive signal to overcome a block
at the pro/pre–B stage and during IgVH repertoire selection. Polyclonal IgM rescued B cell development and returned autoan-
tibody levels to near normal. Thus, natural IgM deficiency causes primary autoimmune disease by altering B cell development,
selection, and central tolerance induction. The Journal of Immunology, 2015, 194: 1489–1502.
Immunoglobulin M is produced by all jawed vertebrates. It isthe first isotype produced in ontogeny and the first Ig producedin response to an insult. Its pentameric structure is also unique
among the other Ig isotypes, indicating its unique contributionsto immunity and the host’s interactions with its environment (1).Spontaneous “natural” IgM secretion occurs without external mi-crobial stimulation (2, 3). Major sources of natural IgM in miceare B-1 cells situated in spleen and bone marrow, producingat least 80% of the circulating IgM (4, 5). Natural IgM-producingB-1 cells appear to be selected on self-antigens (6, 7) and exhibitdual reactivity to both self and common microbial Ags (1, 8, 9).This selection process might ensure the generation of evolu-tionary “useful” specificities (8). Indeed, natural Abs appear tobind particularly to “altered” self-antigens, such as Ags expressedon dead and dying cells, which is thought to allow the efficientremoval of tissue debris and thereby the removal of potentialautoantigens (1, 9–12).Rapid T-independent IgM responses to systemic application of
microbial components, such as LPS of Gram-negative bacteria, orpolysaccharide Ags are induced by both B-1 (13, 14) and bymarginal zone (MZ) B cells (15), which have a high propensity forrapid differentiation to IgM-secreting cells. Finally, most con-ventional B cell responses result in the initial production of IgMby early activated B cells, prior to class-switch recombination to
IgG, IgA, or IgE (16). Early low-affinity IgM may facilitate Agdeposition in the developing germinal centers (17).Selective IgM deficiency is a little studied, relatively rare pri-
mary immunodeficiency of humans, reported to occur at a preva-lence rate of 0.03% (18). Selective IgM deficiency is oftenassociated with recurrent infections (18), consistent with findingsin sIgM-deficient mice (ms2/2), which showed increased mor-bidity and mortality from various bacterial and viral infections(19–22). The data highlight the importance of both natural andAg-induced IgM in immune protection from pathogen encounter.Mechanistically less well understood is the observed develop-
ment of autoantibodies against dsDNA (12, 23) and the increasedrisk of autoimmune diseases such as arthritis and systemic lupuserythematosus in a subset of humans with selective IgM deficiencyand in ms2/2 mice (11, 12, 18). It has been argued that this isattributable to a break of peripheral B cell tolerance due to inef-fective removal of cell debris in the absence of natural Abs (1, 11,12). This is consistent with the repertoire of self-specificities thatpreferentially bind to dead and dying self and other components ofthe altered self (24, 25). Yet, no studies to date have demonstratedsuch lack of self-antigen removal. Moreover, various BCR trans-genic and knock-in mice have been generated during the last twodecades, which express highly restricted oligoclonal or evenmonoclonal B cells, and often lack B-1 cells and/or B-1 cell–de-rived IgM (26–29). These mice do not appear to suffer from au-toimmune disease, indicating that autoantibody production in IgMdeficiency may have other underlying causes.Positive and negative selection events during B cell development
are critical for the elimination of self-reactive B cells. The fate ofthe developing B cells is strongly dependent on the strength of BCRinteraction with self-antigens (30, 31). Autoreactive immatureB cells may either 1) undergo L chain rerearrangement, that is,change their Ag specificity, 2) become anergic, that is, unre-sponsive, and express the BCR inhibitory surface molecule CD5,or 3) die via apoptosis (31, 32). Overall strengths of the selectingsignals appear to determine also B cell subset selection. Relativelystrong signals seem to favor development of B-1 and follicular(FO) B cells, whereas weaker signals drive MZ B cell develop-ment (33, 34).Lack of sIgM may affect B cell development, possibly via ex-
pression of the recently identified FcmR (35–38). However, re-
*Center for Comparative Medicine, University of California, Davis, Davis, CA95616; †Graduate Group in Immunology, University of California, Davis, Davis,CA 95616; ‡Microbiology Graduate Group, University of California, Davis, Davis,CA 95616; xDepartment of Pathology, Microbiology, and Immunology, School ofVeterinary Medicine, University of California, Davis, Davis, CA 95616
Received for publication July 23, 2014. Accepted for publication December 18, 2014.
This work was supported by National Institutes of Health Grants AI51354, AI85568,and U19AI109962, by the Graduate Group in Immunology, University of California,Davis, and by a Vietnamese Education Fellowship to T.T.T.N.
Address correspondence and reprint requests to Dr. Nicole Baumgarth, Center forComparative Medicine, University of California, Davis, County Road 98 and Hutch-ison Drive, Davis, CA 95616. E-mail address: [email protected]
The online version of this article contains supplemental material.
Abbreviations used in this article: 7-AAD, 7-aminoactinomycin D; ANA, anti-nuclear Ab; FO, follicular; MZ, marginal zone; PtC, phophatidylcholine; s, secreted;SA, streptavidin; WT, wild-type.
Copyright� 2015 by The American Association of Immunologists, Inc. 0022-1767/15/$25.00
www.jimmunol.org/cgi/doi/10.4049/jimmunol.1401880
by guest on June 12, 2016http://w
ww
.jimm
unol.org/D
ownloaded from

ported alterations appeared not only subtle but also difficult toreconcile: two independently generated strains of ms2/2 micewere reported to have increased numbers of MZ B and B-1 cells,but a normal FO B cell compartment (39, 40). The increased MZB cell development in ms2/2 mice may indicate that sIgM mayaffect B cell subset selection during development, and that in itsabsence overall strengths of the selecting signals is reduced,resulting in increases in the MZ B cell compartment. However,this is not consistent with the reported expansion of their B-1 cellcompartment (40), which one would expect to be reduced andwith a lack of effects on FO B cells.To determine the causes of autoantibody production in selective
IgM deficiency we re-examined the previously generated ms2/2
mice (40). Our findings demonstrate that the accumulation ofautoantibodies is due to nonredundant effects of sIgM on B celldevelopment and repertoire selection. Escape from central toler-ance induction in the absence of sIgM explains the accumulationof anergic CD5+CD432 B-2 cells and the generation of auto-antibodies and identifies sIgM as a nonredundant regulator ofB cell selection.
Materials and MethodsMice
Eight- to 12-wk-old female and male mice, C57BL/6J (wild-type [WT];CD45.2, Igh-b), B6.SJL-Ptprca Pepcb/BoyJ (CD45.1), B6.129S7-Rag1tm1Mom/J, and B6.Cg-Igha Thy1a Gpi1a/J (Igh-a) mice were obtainedfrom The Jackson Laboratory. Breeding pairs of B6.129S-sIgM2/2 (ms2/2
CD45.2, Igh-a) mice were a gift from Dr. Frances Lund (University ofAlabama at Birmingham). Heterozygous ms+/2 mice were created byintercrossing ms2/2 and C57BL/6J mice. Heterozygous CD45.13 CD45.2F1 mice were created by intercrossing B6.SJL-Ptprca Pepcb/BoyJ withC57BL/6J mice. Igh-a 3 Igh-b mice were created by intercrossing B6.Cg-Igha Thy1a Gpi1a/J (Igh-a) with C57BL/6J mice. All mice were keptin specific pathogen-free housing, placed in filtered-air ventilated racks,screened for the absence of 17 common mouse pathogens (list availableupon request). Mice were euthanized by overexposure to carbon dioxide.
To generate mixed bone marrow irradiation chimeras, single-cell sus-pensions of bone marrow from the femur and tibia of WT (C57BL/6J) andms2/2 mice were injected i.v. at a 1:1 ratio into CD45.13 CD45.2 F1 micelethally irradiated by exposure to a gamma irradiation source (800 rad).Similarly, WT and FcmR2/2 (CD45.2; provided by Dr. Hiromi Kubagawa[University of Alabama at Birmingham]) mixed bone marrow chimeraswere generated by injecting WT or FcmR2/2 bone marrow into lethallyirradiated CD45.1 mice. Chimeras were allowed to reconstitute for at least7 wk before use in experiments. All procedures and experiments involvinganimals were approved by the Animal Use and Care Committee of theUniversity of California, Davis.
B cell reconstitution after sublethal irradiation
Groups of four WT and ms2/2 mice were sublethally irradiated by whole-body exposure to a gamma irradiation source (350 rad). The mice wereallowed to reconstitute peripheral B cell compartments for 12 d. Fre-quencies of transitional B cells (CD93+) in bone marrow and spleen weredetermined by flow cytometry.
Transfer of sIgM
Groups of three or four ms2/2 mice were injected i.p. with either 50 ml WTserum, containing ∼200 mg IgM as assessed by ELISA, ms2/2 serum, or200 mg monoclonal IgM (Sigma-Aldrich, clone MOPC-104E, specific fora-1,3-glucose) per mouse three times per week for 3 wk to reconstitutesIgM. Control mice received PBS only.
Flow cytometry
Spleen and lymph node cells were obtained by grinding the organ betweenfrosted ends of glass slides. Bone marrow was obtained by flushing fibulaand tibia with medium using a 23-gauge needle. Single-cell suspensionswere stained as previously described (41). Briefly, after Fc receptorblocking (5 mg/ml anti-CD16/32) for 20 min on ice, cells were stainedwith the following fluorochome conjugates: streptavidin (SA)–Qdot 605,SA-allophycocyanin, CD93-PE, CD80-PE, CD86-allophycocyanin, CD45.1–(PerCP, Cy5.5-PE), CD45R–Cy7-allophycocyanin, CD3e–Cy55-allophyco-
cyanin, BrdU-FITC (all BD Pharmingen), CD45.2–(Cy7-allophycocyanin,Cy7-PE) (BioLegend), CD5–(biotin, Cy5-PE), CD45R–(FITC, Cy7-allophycocyanin), FITC-labeled, phophatidylcholine (PtC)-containingliposomes–FITC, CD21-(FITC, biotin), CD23-(FITC, allophycocyanin),CD43–(PE, Cy7-allophycocyanin), CD19–(Cy5-PE, allophycocyanin), CD21–Cy5.5-PE, CD24–Cy5.5-PE, BP-1–allophycocyanin, IgD–Cy7-PE, and IgM–(Cy7-allophycocyanin, Cy55-allophycocyanin) (all in-house generated). Deadcells were excluded by Live/Dead Pacific Blue staining (Invitrogen). Stainingfor BrdU was done according to the manufacturer’s protocols using a BDPharmigen BrdU flow kit. To accurately set gates to identify CD5- and CD43-expressing cells, we used “fluorescence minus one” controls in which cellswere stained with all reagents except anti-CD5 and anti-CD43, respectively.
For in vitro sIgM binding, sIgM (SouthernBiotech, clone 11E10, specificfor LPS) was conjugated to biotin and added to bone marrow and spleencells for 1 h at 4˚C. Staining was revealed with SA-allophycocyanin. Deadcells were excluded by Live/Dead Pacific Blue staining.
For cell cycle analysis, surface-stained splenocytes were fixed with 2.5%paraformaldehyde, washed in PBS plus 1% BSA, and stained with anti–Ki67-FITC and 7-aminoactinomycin D (7-AAD) (BD Pharmingen) overnight at4˚C. FACS analysis was done using a 15-parameter FACSAria (41) or anLSRFortessa (BD Biosciences) equipped with four lasers and optics for 22-paramenter analysis. Analysis was done using FlowJo (gift of Adam Treister).
FACS sorting of live “non-dump,” non–autofluorescent CD19+ B cellsubsets was done using a FACSAria (BD Biosciences). Sort gates weredefined as follows: FO B cells, CD21intCD23+; MZ B cells, CD21hiCD232;anergic B cells, CD21intCD232. Sort gates for bone marrow CD45R+CD432
B cell precursors were defined as follows: fraction D, IgD2IgM2; fraction E,IgD2IgM+; fraction F, IgD+IgM+. Sorting purities were .95%.
Magnetic B cell enrichment
Splenic B cells were enriched using a mixture of depleting Abs (CD90.2-biotin, Dx5-biotin, Gr-1–biotin) and anti-biotin microbeads (MiltenyiBiotec) after Fc blocking. CD232 (non-FO) B cells in the peritoneal cavitywere enriched by adding anti–CD23-biotin to the above Ab mixture.Nylon-filtered splenocytes were separated using autoMACS (MiltenyiBiotec). Purities of enriched B cells were .90% as determined by sub-sequent FACS analysis.
In vitro B cell proliferation assay
FACS-purified MZ, FO, and anergic CD21intCD232 B cells from WT andms2/2 mice were labeled with 0.5 mM CFSE in PBS at a concentration of107 cells/ml for 20 min at 37˚C, washed twice with PBS, and cultured at2.5 3 105 cells/well in the presence or absence of 20 mg/ml anti-IgMmedium (RPMI 1640, 292 mg/ml L-glutamine, 100 mg/ml penicillin/streptomycin, 10% heat-inactivated FCS, 0.03 M 2-ME) in 96-wellround-bottom plates for 3 d at 37˚C in 5% CO2. FACS analysis was done toidentify the number of dividing cells.
Testing for serum anti-nuclear Ab
The method of anti-nuclear Ab (ANA) staining was adapted from previouspublications (42). 3T3 cells (A31, provided by Dr. Peter Barry, Universityof California, Davis) were grown on glass slides overnight and fixed withacetone for 10 min. The slides were blocked with Fc block in 10% FCS/PBS buffer for 1 h, washed twice with PBS and once with washing buffers(PBS plus 0.1% BSA plus 1% normal calf serum), and then incubated withmouse serum in PBS for 2 h (1:100 dilution for 2.5-mo-old mice and1:1000 dilution for 8-mo-old mice). Staining was revealed using goat anti-mouse IgG-biotin and SA–Alexa Fluor 594, each incubated for 1 h at roomtemperature. The 3T3-stained slides were washed with PBS for 5 min atleast three times and mounted with Fluoromount-G (SouthernBiotech).Images were collected with an Olympus BX61 microscope with anOlympus DP72 color camera and were processed by using MetaMorph(Molecular Devices) and ImageJ (National Institutes of Health) software.Differential interference contrast images were taken for outlining the cells.Compared images were collected on the same day and analyzed in thesame way. The scale bars were from two images (original magnification320 and 340) and applied to others.
ELISA
DNA-specific IgG was measured by ELISAs. Ninety-six–well plates(Maxisorb; Thermo Fisher Scientific) were coated with 10 mg/ml dsDNAfrom calf thymus or 10 mg/ml ssDNA (degraded dsDNA after heating at94˚C for 14 min in 1 N NaOH) in PBS overnight. After blocking with PBS/1% heat-inactivated calf serum/0.1% milk powder/0.05% Tween 20 for1 h, 2-fold serially diluted serum in PBS was added for 2 h. Plates werewashed and Ab binding was revealed using biotin-conjugated anti-IgG
1490 SECRETED IgM REGULATES B CELL SELECTION
by guest on June 12, 2016http://w
ww
.jimm
unol.org/D
ownloaded from

(SouthernBiotech) followed by a SA-HRP (Vector Laboratories, Burlin-game, CA) incubation for 1 h and then substrate (10 mg/ml 3,39,5,59-tet-ramethylbenzidine in 0.05 mM citric acid, 3% hydrogen peroxide) for 20min. Reactions were stopped with 1 N sulfuric acid, and absorbance wasread at 450 nm and reference wavelength of 595 nm using a SpectraMaxM5 (Molecular Devices). Relative unit Ab levels were calculated bycomparison with NZB mouse serum.
Quantitative PCR and quantitative RT-PCR
DNAwas isolated from sorted cells by a DNeasy kit (Qiagen) and stored inthe DNA storage buffer at 220˚C until processed. IgHV regions wereamplified by quantitative PCR as described previously (43) in SYBRGreen buffer (Applied Biosystems). For quantitative RT-PCR analysis ofVH11 mRNA expression, total RNA from peritoneal cavity B cells wasisolated with RNeasy (Qiagen) and stored in the RNA storage buffer(Ambion) at 280˚C until processed. cDNAwas prepared by using randomhexamers (Promega) with SuperScript II reverse transcriptase (Invitrogen).Primers and probe for IGVH11 were designed using IDT software: forwardprimer, 59-GAAGTGCAGCTGTTGGAGA-39; reverse primer, 59-GTA-CAGGGTGCTCTTGTCATT-39. Relative expression was normalized toGAPDH (IDT).
Statistical analysis
Statistical analysis was done using a two-tailed Student t test. A p value,0.05 was considered statistically significant.
ResultsAn unusual population of CD5+/2CD21intCD232 B cellsdevelops in ms2/2 mice
Measurements of autoantibodies by ELISA in 10-wk-old ms2/2
and age-matched WT controls confirmed earlier reports on thegeneration of high titers of IgG autoantibodies against dsDNA andssDNA in ms2/2 mice (Fig. 1A, 1B) (12, 23). We also found in-creased anti-nuclear IgG Abs, which further increased with age(Fig. 1C, 1D). Thus, selective IgM deficiency causes the devel-opment of autoantibodies against multiple self-antigens, whichincrease with age. Comparison of total serum IgG levels in WTand ms2/2 mice as assessed by ELISA revealed no significantdifferences between the mouse strains at 3 mo of age and onlyvery modest increases at 8 mo (Supplemental Fig. 1A). Similarly,the frequencies of IgG-secreting cells in the spleen as measured byELISPOT and the frequencies of spleen CD19+CD138+ plasmacells were not significantly different between these two mousestrains (Supplemental Fig. 1B, 1C). Thus, the difference in auto-antibody production in ms2/2 mice cannot be explained by dif-ferences in total IgG levels and instead suggest B cell repertoiredifferences.Modest increases in MZ (39) and peritoneal cavity B-1 cells in
ms2/2 mice (40) were previously reported. We confirmed the in-creased frequencies of MZ B cells and a concomitant decrease inFO B cells in ms2/2 mice (Fig. 2A, 2B), but we identified multipleadditional defects. First, ms2/2 mice had significantly reducednumbers of CD19+ B cells, especially in lymph nodes (Fig. 2A,2B). Moreover, we noted an unusual population of CD21intCD232
CD19+ B cells in spleens and lymph nodes of ms2/2 mice that wasnot present in WT mice and blurred the differentiation of MZ andFO B cells somewhat (Fig. 2A). Further analysis identified themas CD5+/2CD21loCD232CD432CD45RhiCD932 (Fig. 2C). De-spite expression of CD5 on some of these cells, they were notclassical B-1a cells, as they were mostly CD432, and CD5 ex-pression levels of those expressing CD5 were lower and their ex-pression of CD45R was higher compared with those of B-1a cells(Fig. 2C). They were not transitional B cells either, as they lackedexpression of CD93. Based on their expression of CD80 and CD86they appeared to be Ag experienced. In WT mice, very few B cellsfell into the CD21intCD232 B cell gate. Those that did were eitherB-1 (CD5+CD21loCD232CD43+CD45Rlo) or transitional B cells
(CD52CD21loCD232CD432CD45RhiCD93+) (Fig. 2C, 2D). FOcells also showed phenotypic alterations in ms2/2 mice. Theyexpressed reduced levels of IgD and some seemed to express verylow levels of CD5 based on comparison with control stains that didnot include CD5 (Fig. 2D and data not shown). These dramaticchanges of the peripheral B cell compartments suggested that sIgMis a nonredundant component required for normal B cell develop-ment and B cell subset selection and/or maintenance. We thereforeexplored the relationship between the effects of IgM on the B cellcompartment and the regulation of autoantibody production.
B cell development from ms2/2 bone marrow precursors isrescued in the presence of sIgM
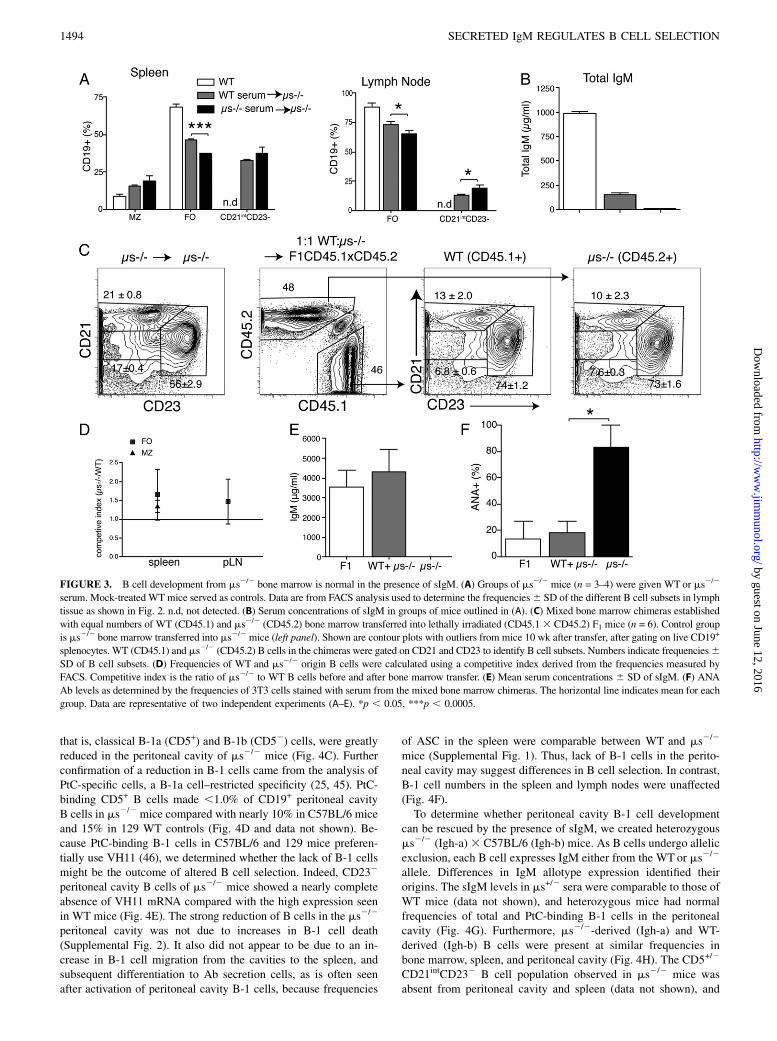
First, we determined whether the B cell developmental changes inms2/2 mice could be normalized by the administration of sIgM.For that we injected polyclonal IgM purified from WT serum intoms2/2 mice three times per week for 3 wk. Transfer of sIgMresulted in significant increases in the frequencies of FO B cellsand reduced frequencies of CD21intCD232 B cells in spleen andlymph nodes compared with mice receiving serum from ms2/2
mice, a decrease that reached statistical significance in the lymphnodes (Fig. 3A). Owing to the very short half-live of IgM (2 d)(44), the levels of circulating serum sIgM could not be normalizedwith this approach (Fig. 3B), which may explain why a morecomplete rescue of B cells was not achieved.Therefore, to test for the development of B cells from precursors
of ms2/2 mice in the presence of normal levels of serum IgMwe generated mixed bone marrow chimeras, transferring WT(CD45.1) and ms2/2 (CD45.2) bone marrow at a 1:1 ratio intolethally irradiated CD45.1 3 CD45.2 F1 recipients. The CD45.1/CD45.2 double-positive radio-resistant recipient cells wereeasily distinguished from the single-positive WT and ms2/2 donorB cells in the host (Fig. 3C, second panel). Recipients of WT ormixed ms2/2 WT bone marrow had similar serum IgM levels,whereas recipients of ms2/2 bone marrow lacked sIgM (Fig. 3E).Precursors from ms2/2 mice reconstituted roughly half of eachB cell compartment in the mixed bone marrow chimeras (Fig. 3C,3D). Frequencies and phenotypes of all major B cell subsets werenormal (Fig. 3C, 3D, and not shown). Furthermore, CD21int
CD232 B cells did not develop in the mixed chimeras, but theydid develop in controls that received ms2/2 bone marrow only andthus lacked IgM (Fig. 3C, 3E). Importantly, ANA developmentwas greatly reduced in mixed bone marrow chimeras, comparedwith mice receiving ms2/2 bone marrow only. The latter generatedmeasurable levels of ANA within 10 wk after adoptive transfer(Fig. 3F). Thus, precursors of ms2/2 mice have no inherent de-velopmental defect in B cell development when raised in thepresence of sIgM.
B-1 cell frequencies are strongly reduced in the peritonealcavity of ms2/2 mice but are maintained at normal numbers inthe spleen
Increases in peritoneal cavity CD5+CD45R+ B-1 cells have beenreported for ms2/2 mice (40). Although our results confirmedincreases in frequencies of CD5+ B cells (Fig. 4A), these cellswere mostly CD432CD45Rhi and CD19int, that is, they did notrepresent classical CD43+CD45RloCD19hi B-1a cells of WT mice(Fig. 4A, 4B). The difference in CD43 staining shown (Fig. 4A,4B) is highly significant (mean fluorescence intensity of 2131 6556 on WT CD19+IgM+ cells versus 195 6 34 for ms2/2 cells,p , 0.005). Also, the difference for CD45R expression was greatlysignificant (1,758 6 259 versus 3,727 6 115, p , 0.0001), aswas the reduction in expression of CD19 (12,720 6 1,036 versus8,920 6 320, p , 0.005). Therefore, CD19hiCD45RloCD43+ cells,
The Journal of Immunology 1491
by guest on June 12, 2016http://w
ww
.jimm
unol.org/D
ownloaded from
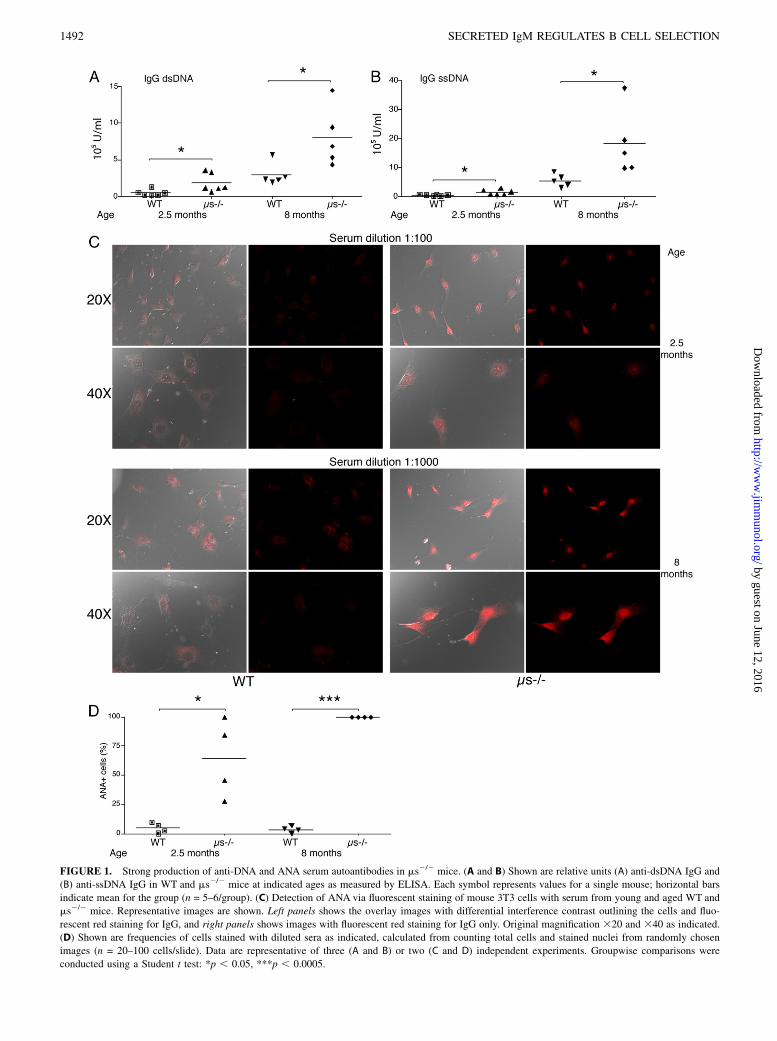
FIGURE 1. Strong production of anti-DNA and ANA serum autoantibodies in ms2/2 mice. (A and B) Shown are relative units (A) anti-dsDNA IgG and
(B) anti-ssDNA IgG in WT and ms2/2 mice at indicated ages as measured by ELISA. Each symbol represents values for a single mouse; horizontal bars
indicate mean for the group (n = 5–6/group). (C) Detection of ANA via fluorescent staining of mouse 3T3 cells with serum from young and aged WT and
ms2/2 mice. Representative images are shown. Left panels shows the overlay images with differential interference contrast outlining the cells and fluo-
rescent red staining for IgG, and right panels shows images with fluorescent red staining for IgG only. Original magnification 320 and 340 as indicated.
(D) Shown are frequencies of cells stained with diluted sera as indicated, calculated from counting total cells and stained nuclei from randomly chosen
images (n = 20–100 cells/slide). Data are representative of three (A and B) or two (C and D) independent experiments. Groupwise comparisons were
conducted using a Student t test: *p , 0.05, ***p , 0.0005.
1492 SECRETED IgM REGULATES B CELL SELECTION
by guest on June 12, 2016http://w
ww
.jimm
unol.org/D
ownloaded from
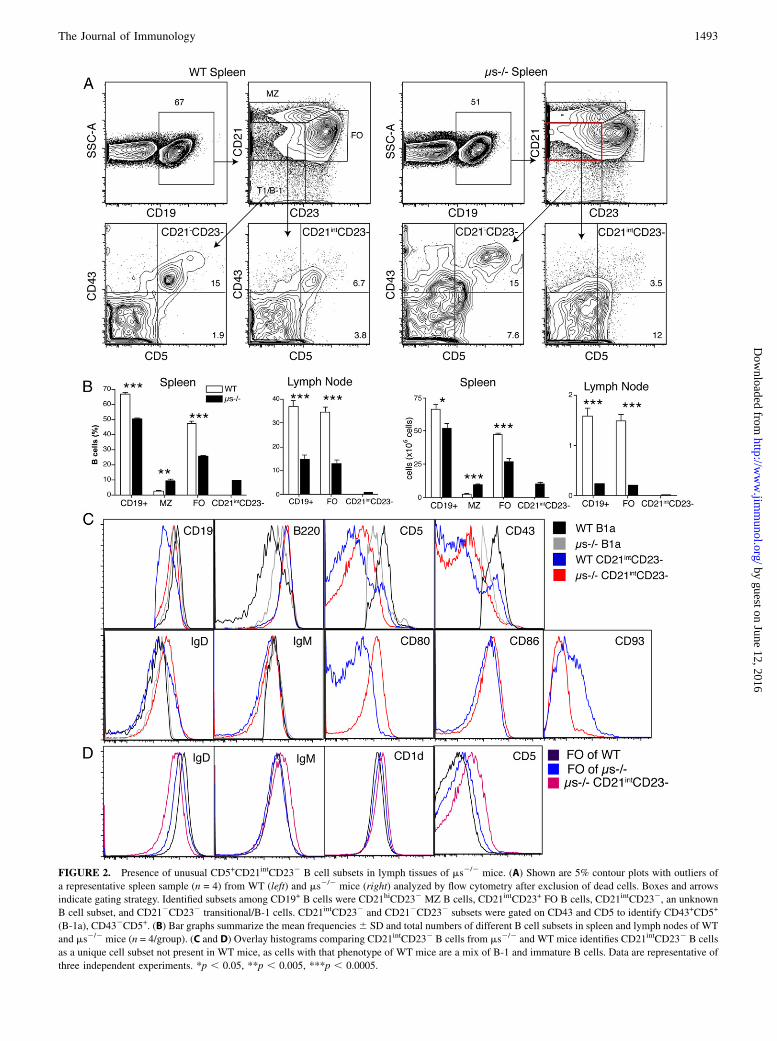
FIGURE 2. Presence of unusual CD5+CD21intCD232 B cell subsets in lymph tissues of ms2/2 mice. (A) Shown are 5% contour plots with outliers of
a representative spleen sample (n = 4) from WT (left) and ms2/2 mice (right) analyzed by flow cytometry after exclusion of dead cells. Boxes and arrows
indicate gating strategy. Identified subsets among CD19+ B cells were CD21hiCD232 MZ B cells, CD21intCD23+ FO B cells, CD21intCD232, an unknown
B cell subset, and CD212CD232 transitional/B-1 cells. CD21intCD232 and CD212CD232 subsets were gated on CD43 and CD5 to identify CD43+CD5+
(B-1a), CD432CD5+. (B) Bar graphs summarize the mean frequencies6 SD and total numbers of different B cell subsets in spleen and lymph nodes of WT
and ms2/2 mice (n = 4/group). (C and D) Overlay histograms comparing CD21intCD232 B cells from ms2/2 and WT mice identifies CD21intCD232 B cells
as a unique cell subset not present in WT mice, as cells with that phenotype of WT mice are a mix of B-1 and immature B cells. Data are representative of
three independent experiments. *p , 0.05, **p , 0.005, ***p , 0.0005.
The Journal of Immunology 1493
by guest on June 12, 2016http://w
ww
.jimm
unol.org/D
ownloaded from
that is, classical B-1a (CD5+) and B-1b (CD52) cells, were greatlyreduced in the peritoneal cavity of ms2/2 mice (Fig. 4C). Furtherconfirmation of a reduction in B-1 cells came from the analysis ofPtC-specific cells, a B-1a cell–restricted specificity (25, 45). PtC-binding CD5+ B cells made ,1.0% of CD19+ peritoneal cavityB cells in ms2/2 mice compared with nearly 10% in C57BL/6 miceand 15% in 129 WT controls (Fig. 4D and data not shown). Be-cause PtC-binding B-1 cells in C57BL/6 and 129 mice preferen-tially use VH11 (46), we determined whether the lack of B-1 cellsmight be the outcome of altered B cell selection. Indeed, CD232
peritoneal cavity B cells of ms2/2 mice showed a nearly completeabsence of VH11 mRNA compared with the high expression seenin WT mice (Fig. 4E). The strong reduction of B cells in the ms2/2
peritoneal cavity was not due to increases in B-1 cell death(Supplemental Fig. 2). It also did not appear to be due to an in-crease in B-1 cell migration from the cavities to the spleen, andsubsequent differentiation to Ab secretion cells, as is often seenafter activation of peritoneal cavity B-1 cells, because frequencies
of ASC in the spleen were comparable between WT and ms2/2
mice (Supplemental Fig. 1). Thus, lack of B-1 cells in the perito-neal cavity may suggest differences in B cell selection. In contrast,B-1 cell numbers in the spleen and lymph nodes were unaffected(Fig. 4F).To determine whether peritoneal cavity B-1 cell development
can be rescued by the presence of sIgM, we created heterozygousms2/2 (Igh-a)3 C57BL/6 (Igh-b) mice. As B cells undergo allelicexclusion, each B cell expresses IgM either from the WT or ms2/2
allele. Differences in IgM allotype expression identified theirorigins. The sIgM levels in ms+/2 sera were comparable to those ofWT mice (data not shown), and heterozygous mice had normalfrequencies of total and PtC-binding B-1 cells in the peritonealcavity (Fig. 4G). Furthermore, ms2/2-derived (Igh-a) and WT-derived (Igh-b) B cells were present at similar frequencies inbone marrow, spleen, and peritoneal cavity (Fig. 4H). The CD5+/2
CD21intCD232 B cell population observed in ms2/2 mice wasabsent from peritoneal cavity and spleen (data not shown), and
FIGURE 3. B cell development from ms2/2 bone marrow is normal in the presence of sIgM. (A) Groups of ms2/2 mice (n = 3–4) were given WT or ms2/2
serum. Mock-treated WT mice served as controls. Data are from FACS analysis used to determine the frequencies6 SD of the different B cell subsets in lymph
tissue as shown in Fig. 2. n.d, not detected. (B) Serum concentrations of sIgM in groups of mice outlined in (A). (C) Mixed bone marrow chimeras established
with equal numbers of WT (CD45.1) and ms2/2 (CD45.2) bone marrow transferred into lethally irradiated (CD45.13 CD45.2) F1 mice (n = 6). Control group
is ms2/2 bone marrow transferred into ms2/2 mice (left panel). Shown are contour plots with outliers from mice 10 wk after transfer, after gating on live CD19+
splenocytes. WT (CD45.1) and ms2/2 (CD45.2) B cells in the chimeras were gated on CD21 and CD23 to identify B cell subsets. Numbers indicate frequencies6SD of B cell subsets. (D) Frequencies of WT and ms2/2 origin B cells were calculated using a competitive index derived from the frequencies measured by
FACS. Competitive index is the ratio of ms2/2 to WT B cells before and after bone marrow transfer. (E) Mean serum concentrations 6 SD of sIgM. (F) ANA
Ab levels as determined by the frequencies of 3T3 cells stained with serum from the mixed bone marrow chimeras. The horizontal line indicates mean for each
group. Data are representative of two independent experiments (A–E). *p , 0.05, ***p , 0.0005.
1494 SECRETED IgM REGULATES B CELL SELECTION
by guest on June 12, 2016http://w
ww
.jimm
unol.org/D
ownloaded from
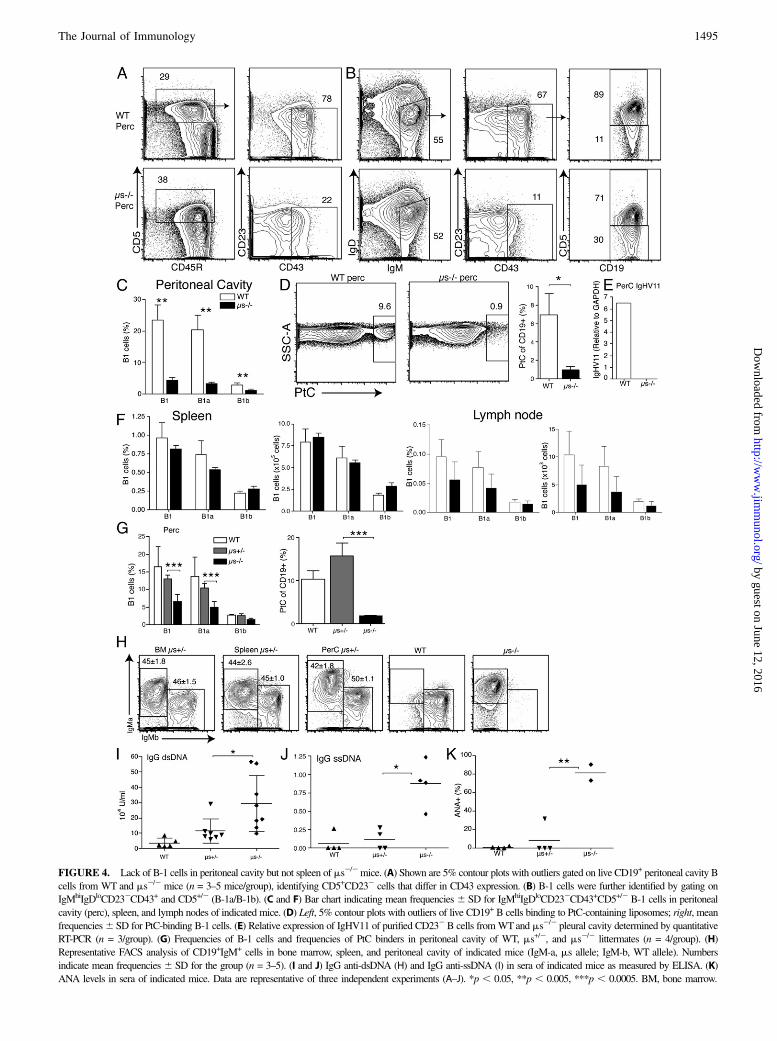
FIGURE 4. Lack of B-1 cells in peritoneal cavity but not spleen of ms2/2mice. (A) Shown are 5% contour plots with outliers gated on live CD19+ peritoneal cavity B
cells from WT and ms2/2 mice (n = 3–5 mice/group), identifying CD5+CD232 cells that differ in CD43 expression. (B) B-1 cells were further identified by gating on
IgMhiIgDloCD232CD43+ and CD5+/2 (B-1a/B-1b). (C and F) Bar chart indicating mean frequencies 6 SD for IgMhiIgDloCD232CD43+CD5+/2 B-1 cells in peritoneal
cavity (perc), spleen, and lymph nodes of indicated mice. (D) Left, 5% contour plots with outliers of live CD19+ B cells binding to PtC-containing liposomes; right, mean
frequencies6 SD for PtC-binding B-1 cells. (E) Relative expression of IgHV11 of purified CD232B cells fromWTand ms2/2 pleural cavity determined by quantitative
RT-PCR (n = 3/group). (G) Frequencies of B-1 cells and frequencies of PtC binders in peritoneal cavity of WT, ms+/2, and ms2/2 littermates (n = 4/group). (H)
Representative FACS analysis of CD19+IgM+ cells in bone marrow, spleen, and peritoneal cavity of indicated mice (IgM-a, ms allele; IgM-b, WT allele). Numbers
indicate mean frequencies 6 SD for the group (n = 3–5). (I and J) IgG anti-dsDNA (H) and IgG anti-ssDNA (I) in sera of indicated mice as measured by ELISA. (K)
ANA levels in sera of indicated mice. Data are representative of three independent experiments (A–J). *p , 0.05, **p , 0.005, ***p , 0.0005. BM, bone marrow.
The Journal of Immunology 1495
by guest on June 12, 2016http://w
ww
.jimm
unol.org/D
ownloaded from

IgG autoantibody levels against dsDNA (Fig. 4I), ssDNA (Fig. 4J),and ANA-specific autoantibodies (Fig. 4K) were strongly reducedin sera from 10-wk-old ms+/2 mice compared with age-matchedms2/2 controls. The slight but not statistically significant increasesof autoantibodies in the heterozygous compared with the WT micemight be due to local concentrations of sIgM in the immediate vi-cinity of B cells in the bone marrow. In summary, sIgM is a nonre-dundant factor required for the normal development of B-2 cells andperitoneal cavity, but not splenic B-1 cells.
ms2/2 CD21intCD232 B cells are anergic
CD5 is a negative regulator of BCR signaling, and its expressionon conventional B cells has been previously linked to anergy (32).The expression of CD5 on some FO and CD21intCD232 B cellssuggested the emergence of anergic B cells in ms2/2 mice. AnergicB cells are relatively short-lived and are unresponsive to BCR-mediated stimulation (47). Indeed, B cell subsets in ms2/2 mice,including MZ and FO B cells, showed enhanced turnover com-pared with their WT counterparts as measured by enhanced BrDUincorporation (Fig. 5A), as noted by others (48). The turnover rateof CD21intCD232 B cells was similar to that of FO B cells in ms2/2
mice, but it was enhanced compared with those in WT mice(Fig. 5A), and their rate of cell death, as measured by 7-AAD-incorporation, was highest among all B cell populations (Fig. 5B).To determine whether the higher rate of BrDU incorporation was
due to reduced B cell survival in ms2/2 mice, we transferred equalnumbers of CFSE-labeled splenic B cells from WT (CD45.1) andms2/2 (CD45.2) mice into WT mice. All B cell subsets from ms2/2
mice, including the CD5+/2CD21intCD232 cells, survived poorlycompared with WT B cells at both 2 and 25 d after transfer(Fig. 5C, 5D). Additionally, ms2/2 mice had increased frequenciesof B cells expressing the proliferation marker Ki67, as well as in-creased frequencies of CD19+ cells in the G1 phase of the cell cycle,compared with WT cells (Fig. 5E). Transfer of either sIgM con-taining WT serum or polyclonal IgM could reverse these increases,whereas monoclonal IgM could not (Fig. 5E, 5F).We tested whether the reduced ability of ms2/2 B cells to
survive in vivo could be due to a lack of BCR-mediated signaling,as unresponsiveness to BCR signaling is a hallmark of anergicB cells, and continuous “tonic” signaling through the BCR is re-quired for B cell survival. Indeed, all B cell subsets from ms2/2
mice showed greatly reduced proliferation in response to anti-IgMstimulation compared with their WT counterparts. The CD21int
CD232CD5+/2 B cells appeared least responsive (Fig. 5G). Thus,B cells from ms2 /2 mice have a shortened lifespan and arehyporesponsive to BCR stimulation, suggesting that a lack ofsIgM leads to the development of anergic B cells.
Lack of sIgM causes alterations in B cell development
Given that the transfer of mature ms2/2 B cells into a sIgM-complete environment could not rescue ms2/2 B cell survival(Fig. 5D), sIgM might affect very early stages of B cell develop-ment. Analysis of bone marrow precursors showed that early pro–B cells (Hardy scheme, fraction A) and pro–B cells (fraction B)were increased in ms2/2 compared with WT mice, but they werereduced after the pre–B cell stage (Fig. 6A, Supplemental Fig. 3A).Staining for CD93 expression on Hardy fraction E further demon-strated a strong reduction of immature B cells in the bone marrow(Fig. 6A). Because reduced BCR expression might affect identifi-cation of immature B cells (usually done via IgM/IgD staining), weadded staining for CD93 to the analysis, which confirmed the de-crease of immature B cells in ms2/2 mice (Fig. 6A).The rate of proliferation among pre-pro– and late pre–B cells
was similar between ms2/2 and WT mice (Fig. 6B). Thus, the
increases of fractions A and B were not due to their increasedexpansion, but were likely attributable to increased accumulationdue to reduced progression through the pre–B cell stage. Con-sistent with the presence of reduced B cell precursors in the bonemarrow, frequencies of splenic transitional B cells were reduced attransitional stage T1, but not at T2, in ms2/2 compared with WTmice (Fig. 6C, Supplemental Fig. 3B). Finally, to directly comparebone marrow output, we assessed peripheral B cell reconstitutionon day 12 after sublethal irradiation of ms2/2 and WT mice. Theresults showed significant reductions in the frequencies (Fig. 6D)and total numbers (not shown) of immature/transitional CD19+
CD93+ B cells in the bone marrow and spleen of ms2/2 micecompared with their WT controls. We conclude that the presenceof secreted IgM is required for normal bone marrow B cell de-velopment.
sIgM binds to bone marrow B cells
The data thus suggest that bone marrow B cell development isdependent on the presence of sIgM. To determine whether sIgMcan bind to bone marrow B cells, and thus to assess whether sIgM/Bcell precursor direct interaction may be responsible for the effectof sIgM on B cell development, we measured the binding of sIgMto B cells in bone marrow and periphery.Staining for total surface IgM, which stains for bound sIgM as
well as for membrane-bound BCR, showed that immature B cellsfrom ms2/2 mice had reduced IgM staining compared with WTcontrols (Fig. 6E). To distinguish surface IgM expression frombinding of sIgM to B cells via Fc or other receptors, which wouldnot occur in ms2/2 mice, we created heterozygous Igh-a 3 Igh-bmice, in which the expressed IgM allele and surface-bound IgMfrom the nonexpressed allele were differentiated by surfacestaining with allotype-specific anti-IgMa and anti-IgMb. AllB cells stained significantly for the nonexpressed Igh allotype,with stronger surface binding on peripheral B cells compared withbone marrow precursors (Fig. 6F), which explained the reducedstaining for total surface IgM in ms2/2 compared with WT mice(Fig. 6E). Also, in vitro exposure of spleen and bone marrow cellsto sIgM showed strong sIgM binding to B cells, particularlyB cells from ms2/2 mice (Fig. 6G), including immature and ma-ture B cells, but little to other cell types (Fig. 6G, SupplementalFig. 4). We conclude that sIgM binds to B cells and their pre-cursors in vivo.
FcmR2/2 B cells show no defect in B cell development
Of the known surface IgM receptors, only the FcmR is expressed byearly B cell precursors (35, 37), and we aimed to assess its effectson B cell development. First, we tested for its contribution to sIgMbinding to B cells and non–B cells. For that we exposed in vitroIgM labeled with biotin to bone marrow and spleen cells frombone marrow irradiation chimeras generated by reconstituting WTC57BL/6 (CD45.1) mice with bone marrow from either WTC57BL/6 mice or FcmR2/2mice. WT and FcmR2/2 B cells wereable to bind sIgM on their cell surface. The lack of the FcmR ledto a measurable but only very modest reduction in sIgM binding toB cells of bone marrow and spleen, whereas non–B cells showedno discernable IgM binding (Fig. 7A, 7B).To determine whether direct binding of IgM via the FcmR
might regulate B cell development, we studied the effects of B celldevelopment in these bone marrow irradiation chimeras. Bonemarrow B cell development appeared normal (Fig. 7C), and pe-ripheral B cell subset composition in the spleen was similar to thatof WT but not ms2/2 mice. An exception was the significant re-duction in MZ B cells (Fig. 7D, 7E), whereas total serum IgMlevels were increased in chimeras reconstituted with FcmR2/2
1496 SECRETED IgM REGULATES B CELL SELECTION
by guest on June 12, 2016http://w
ww
.jimm
unol.org/D
ownloaded from
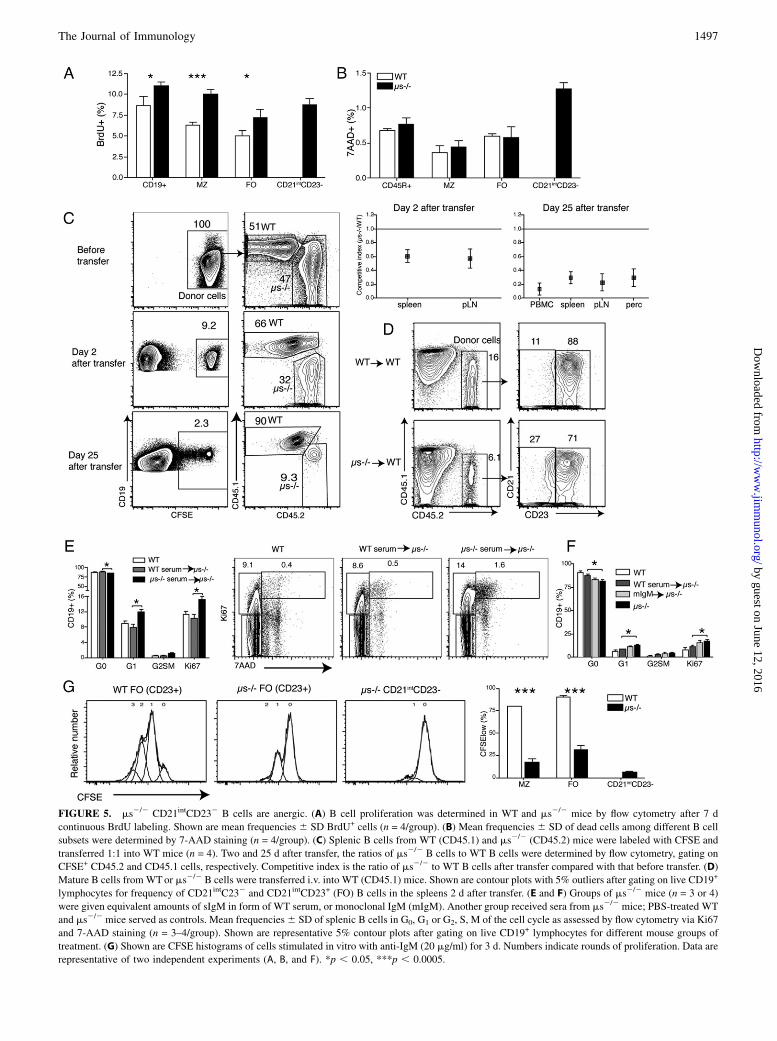
FIGURE 5. ms2/2 CD21intCD232 B cells are anergic. (A) B cell proliferation was determined in WT and ms2/2 mice by flow cytometry after 7 d
continuous BrdU labeling. Shown are mean frequencies 6 SD BrdU+ cells (n = 4/group). (B) Mean frequencies 6 SD of dead cells among different B cell
subsets were determined by 7-AAD staining (n = 4/group). (C) Splenic B cells from WT (CD45.1) and ms2/2 (CD45.2) mice were labeled with CFSE and
transferred 1:1 into WT mice (n = 4). Two and 25 d after transfer, the ratios of ms2/2 B cells to WT B cells were determined by flow cytometry, gating on
CFSE+ CD45.2 and CD45.1 cells, respectively. Competitive index is the ratio of ms2/2 to WT B cells after transfer compared with that before transfer. (D)
Mature B cells fromWTor ms2/2 B cells were transferred i.v. into WT (CD45.1) mice. Shown are contour plots with 5% outliers after gating on live CD19+
lymphocytes for frequency of CD21intC232 and CD21intCD23+ (FO) B cells in the spleens 2 d after transfer. (E and F) Groups of ms2/2 mice (n = 3 or 4)
were given equivalent amounts of sIgM in form of WT serum, or monoclonal IgM (mIgM). Another group received sera from ms2/2 mice; PBS-treated WT
and ms2/2 mice served as controls. Mean frequencies6 SD of splenic B cells in G0, G1 or G2, S, M of the cell cycle as assessed by flow cytometry via Ki67
and 7-AAD staining (n = 3–4/group). Shown are representative 5% contour plots after gating on live CD19+ lymphocytes for different mouse groups of
treatment. (G) Shown are CFSE histograms of cells stimulated in vitro with anti-IgM (20 mg/ml) for 3 d. Numbers indicate rounds of proliferation. Data are
representative of two independent experiments (A, B, and F). *p , 0.05, ***p , 0.0005.
The Journal of Immunology 1497
by guest on June 12, 2016http://w
ww
.jimm
unol.org/D
ownloaded from

bone marrow compared with those of WT-reconstituted mice(Fig. 7F). These findings were similar to the original reports withthis strain of FcmR2/2 mice (49).In vitro responses to anti-IgM stimulation were comparable
between FcmR2/2 B cells and WT B cells, whereas the ms2/2
B cells showed significant reduced B cell proliferation (Fig. 7G,7H). Thus, sIgM direct binding to B cells via the FcmR cannotexplain the significant effects of sIgM on bone marrow B celldevelopment. Because the lack of FcmR expression reduced, butdid not abrogate, sIgM binding to B cells (Fig. 7A), the presenceof other IgM-binding proteins on the surface of the developingB cells may regulate B cell development.
sIgM affects B cell repertoire and selection
The observed binding of sIgM onto B cell precursors may alterB cell selection at the immature B cell stage and with it the B cellrepertoire. Comparison of the BCR repertoire of ms2/2 and WTmice using quantitative PCR on FACS-purified B cell fractionstaken at different checkpoints during bone marrow B cell devel-opment showed significant differences of VH usage among bonemarrow fractions D and E from ms2/2 and WT mice (Fig. 8A,8B). Mature B cells in the bone marrow and FO B cells in theperiphery showed similar repertoire differences (Fig. 8C). Com-parison of IgHV usage among FO B cells between WT and ms2/2
mice and the CD21intCD232 B cells in ms2/2 mice also revealed
repertoire differences demonstrating the strong effects of sIgM ondevelopment of all B cell compartments (Fig. 8D). Combined withthe findings of increased autoantibody generation in ms2/2 mice,as well as reduced VH11 gene usage among B-1 cells, the datademonstrate that the lack of sIgM causes repertoire changes inboth the B-1 and the B-2 cell compartments.
DiscussionThe present study revealed profound effects of sIgM on the de-velopment of most B cells and B cell subsets, thereby providinga mechanism through which sIgM prevents autoantibody forma-tion, namely by facilitating normal B cell development and en-forcing negative selection of autoreactive B cells. Although thea lack of autoantigen clearance may contribute to autoantibodydevelopment in ms2/2 mice (1, 9–12), the observed effects ofsIgM on B cell development in bone marrow and periphery, aswell as the reversal of autoantibody production following the re-establishment of normal B cell development, suggest that theseeffects are the major underlying cause for the development ofautoreactive B cells in IgM deficiency.The direct binding of sIgM to B cells, which we observed at all
stages of B cell development from the pre–B cell stage onward(Fig. 6E, 6F, and data not shown), may provide the nonredundantsignal for B cell development and selection. Among the threeknown B cell sIgM receptors, neither the Fca/mR nor the com-
FIGURE 6. Lack of sIgM changes B cell development in bone marrow and periphery. (A) Bar graph shows their mean frequencies 6 SD of B cell
precursors according to Hardy and Hayakawa (30): A, pre-pro; B, pro; C, late pro; C9, early pre; D, late pre; E, immature; F, mature B cells. (B) Frequencies6SD for BrdU+ cells after continuous labeling for 7 d (n = 4/group). (C) Bar graphs showing the frequencies and total numbers of CD93+ transitional (T1/2)
B cells in spleens of WTand ms2/2 mice. (D) Bar graph shows the frequencies6 SD of transitional B cells (CD93+) in bone marrows and spleens 12 d after
sublethal irradiation of WT and ms2/2 mice. (E) Overlay histogram shows total surface IgM expression of bone marrow fraction E from ms2/2 and WT
mice. (F) Shown are 5% contour plots of FACS staining for surface IgM-a and IgM-b of B cells from Igh-a 3 Igh-b heterozygous mice (n = 2). Overlay
histograms of sIgM-a staining, that is, staining for sIgM bound to the cell surface of IgM-b BCR expressing CD19+ B cells from bone marrow fraction E
and spleen, respectively, are shown. MFI, mean fluorescence intensity. (G) In vitro binding of sIgM to bone marrow B cells (left panel) and non–B cells
(right panel) as measured by staining for sIgM using IgM allotype-specific to exclude staining for BCR. Data are representative of at least two independent
experiments. *p , 0.05, **p , 0.005, ***p , 0.0005. BM, bone marrow.
1498 SECRETED IgM REGULATES B CELL SELECTION
by guest on June 12, 2016http://w
ww
.jimm
unol.org/D
ownloaded from
plement receptors CR1/2 are expressed on pro–B cells, pre–B cells, and immature B cells (50–52), and thus they cannot beresponsible for the observed effects of IgM deficiency. FcmR isexpressed early in B cell development and was a potential can-didate receptor for IgM (37, 53). Additionally, mice lacking theFcmR2/2 develop autoantibodies despite harboring normal levelsserum sIgM (37, 53), further suggesting a central role for sIgMin regulation of B cell development rather than removal ofautoantigens for the enforcement of central tolerance induction.However, studies with bone marrow FcmR2/2 chimeras (Fig. 7)failed to reveal B cell developmental defects similar to thoseobserved in the ms2/2 mice. The phenotype of the bone marrowirradiation chimera is consistent with the original description of
the FcmR2/2 mice from which we obtained the bone marrow (49).In those mice, overall B cell development was found to be unaf-fected, except for the reduction in MZ B cells and an increase insplenic B-1 cells, as well as increases in serum IgM. A secondreport with independently generated FcmR2/2 mice reported noB cell abnormalities (50). However, a third group who generatedFcmR2/2 mice reported reduction in all B cell precursors in thebone marrow, except mature fraction F B cells cells. In contrast toour findings with the sIgM2/2 mice, they found increased num-bers of peritoneal cavity B-1 cells in the FcmR2/2 mice (38).Thus, based on published work with three different FcmR2/2
mice, the lack of this receptor appears to have some effects onB cell development. Those reported changes are more subtle and/
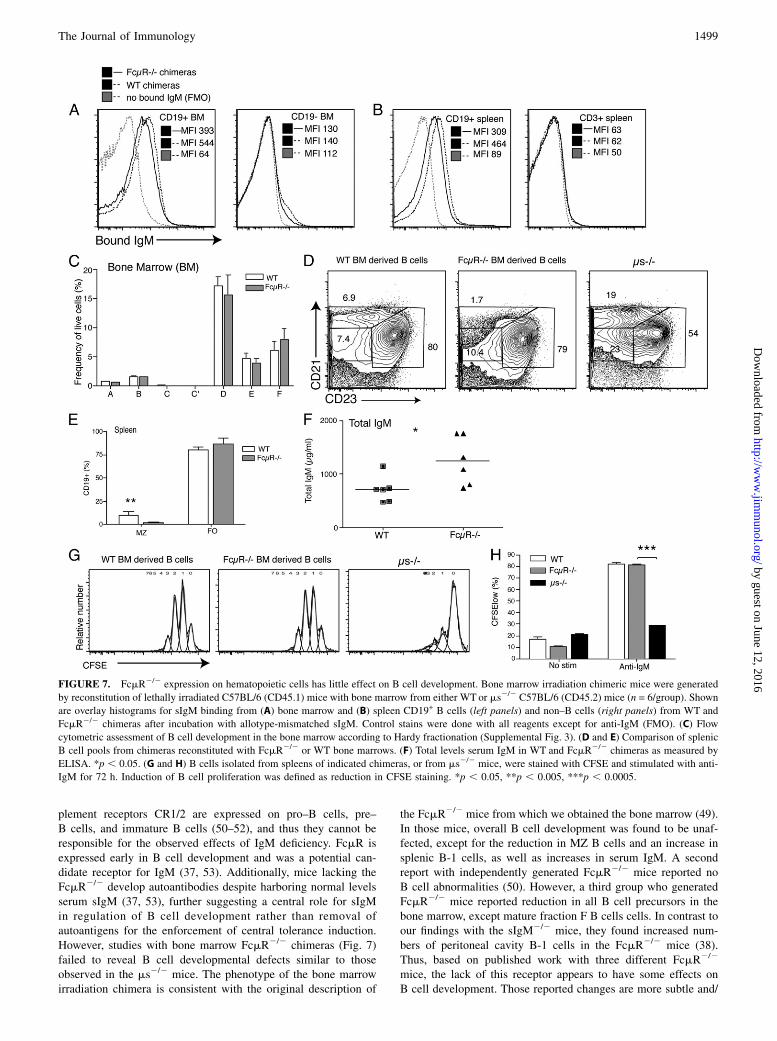
FIGURE 7. FcmR2/2 expression on hematopoietic cells has little effect on B cell development. Bone marrow irradiation chimeric mice were generated
by reconstitution of lethally irradiated C57BL/6 (CD45.1) mice with bone marrow from either WT or ms2/2 C57BL/6 (CD45.2) mice (n = 6/group). Shown
are overlay histograms for sIgM binding from (A) bone marrow and (B) spleen CD19+ B cells (left panels) and non–B cells (right panels) from WT and
FcmR2/2 chimeras after incubation with allotype-mismatched sIgM. Control stains were done with all reagents except for anti-IgM (FMO). (C) Flow
cytometric assessment of B cell development in the bone marrow according to Hardy fractionation (Supplemental Fig. 3). (D and E) Comparison of splenic
B cell pools from chimeras reconstituted with FcmR2/2 or WT bone marrows. (F) Total levels serum IgM in WT and FcmR2/2 chimeras as measured by
ELISA. *p , 0.05. (G and H) B cells isolated from spleens of indicated chimeras, or from ms2/2 mice, were stained with CFSE and stimulated with anti-
IgM for 72 h. Induction of B cell proliferation was defined as reduction in CFSE staining. *p , 0.05, **p , 0.005, ***p , 0.0005.
The Journal of Immunology 1499
by guest on June 12, 2016http://w
ww
.jimm
unol.org/D
ownloaded from
or different from those shown in the present study with thesIgM2/2 mice. Most prominently, the accumulation of CD5+/2
CD432CD232CD21int anergic B cell population in the ms2/2 miceis not noted in any reports of the FcmR2/2 mice, nor was it found inthe FcmR2/2 bone marrow chimeras we created (Fig. 7). Given thesomewhat contradictory reports on the phenotype of the three in-dependently generated FcmR2/2 mice, compensatory mechanismsmay make any differences on B cell development less obvious thana complete lack of sIgM. Alternatively, the various approaches thatwere taken in targeting the FcmR might have led to the differencesin the effects. A direct comparison between the different gene-targeted mice could help to resolve some of these issues.Because natural polyclonal, but not monoclonal, IgM rescued
B cell development (Fig. 5F), our data indicate that natural IgMeither directly or indirectly enhances BCR signaling strengths andthereby alters B cell central tolerance induction at the pre–B andimmature B cell stage, possibly by facilitating autoantigen pre-sentation, as natural IgM-secreting cells appear selected on anddirected against self-antigens (6). However, a BCR ligand re-quirement for pre–B cell selection has not been documented (54).Alternatively, sIgM may provide a costimulatory signal more akinto that provided by pattern recognition receptors, increasing re-sponsiveness of the developing B cell to self-antigens. In supportof a model in which sIgM affects BCR signaling strengths, thelack of sIgM altered the peripheral B cell VH repertoire (Fig. 8),and thus B cell selection, and it led to the accumulation ofautoantibody-secreting cells as well as to the presence of CD5+/2
anergic and autoreactive B cells in the periphery, possibly in anattempt to silence these autoreactive cells. Taken together, the dataprovide evidence that B cell signaling and negative B cell selec-tion at the immature B cell stage are defective in the absence ofsIgM. The increases in MZ and decreases in FO and body cavityB-1 cells are also all consistent with reduced BCR-mediatedsignaling in ms2/2 mice (33, 34).Our findings of reduced peritoneal cavity B-1 cell frequencies
in the ms2/2 mice are opposite to conclusions drawn previously
by others (40), but these differences can be explained by thepresence of CD5+B220+ anergic B cells in the peritoneal cavityand spleen, which were likely misidentified as B-1a cells. Theirlack of CD43 expression, high expression of B220, intermedi-ate expression of CD19, lack of VH11 expression, and of PtCbinding are not consistent with emergence from the B-1 celllineage. Furthermore, their turnover was among the highest ofany B cells studied (Fig. 5), whereas B-1 cell turnover in theperitoneal cavity is very low (55, 56). Multiple mouse modelshave demonstrated that reduction in BCR signaling strengthreduces peritoneal cavity B-1 cell development (7). Thus, thedemonstration of curtailed B-1 cell development in the ms2/2
mice further supports a model in which sIgM acts as a positivecostimulatory signal during B cell development.A “chicken and egg” problem emerges from the fact that sIgM
is required for normal B cell development but that it is notusually transmitted from the maternal circulation to the fetus.Thus, the development of fetal IgM-secreting cells must notrequire IgM. Because B-1 cell numbers in the spleen werelargely unaffected in ms2/2 mice (Fig. 4), they might be theearliest source of natural IgM supporting further sIgM-dependentB-1 and B-2 cell development. This is consistent with studiesshowing profound reductions in peritoneal cavity B-1 cells aftersplenectomy (57) and with studies indicating that B-1 cell de-velopment arises in waves (58). Splenic B-1 cells may arise asa first wave of “IgM-independent” B-1 cells during fetal devel-opment, whereas body cavity B-1 cells emerge later and developin an sIgM-dependent manner. The data indicate a new, nonre-dundant role for splenic B-1 cells in the regulation of B-2 celldevelopment.In summary, the present study demonstrates that B-1 cells
produce natural IgM to regulate B cell development and selection,thereby suppressing the development of autoreactive B cells.Identification of natural IgM as a critical enforcer of central se-lection suggests use of these proteins as potential therapeuticsagainst Ab-mediated autoimmune diseases.
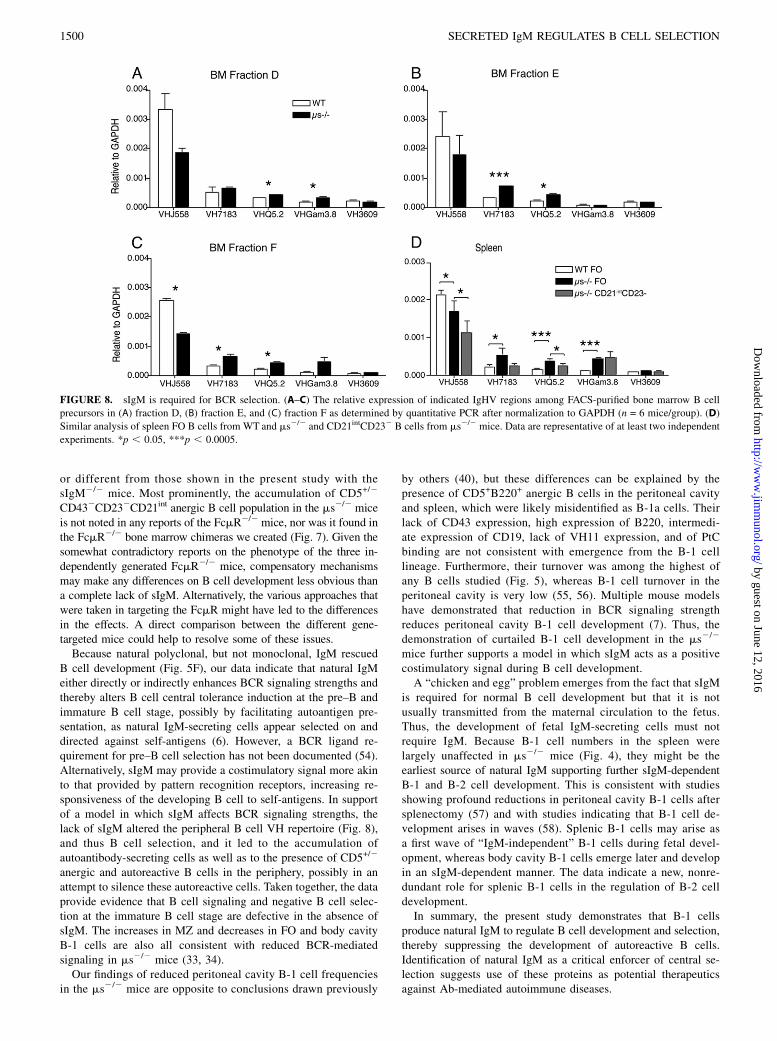
FIGURE 8. sIgM is required for BCR selection. (A–C) The relative expression of indicated IgHV regions among FACS-purified bone marrow B cell
precursors in (A) fraction D, (B) fraction E, and (C) fraction F as determined by quantitative PCR after normalization to GAPDH (n = 6 mice/group). (D)
Similar analysis of spleen FO B cells from WT and ms2/2 and CD21intCD232 B cells from ms2/2 mice. Data are representative of at least two independent
experiments. *p , 0.05, ***p , 0.0005.
1500 SECRETED IgM REGULATES B CELL SELECTION
by guest on June 12, 2016http://w
ww
.jimm
unol.org/D
ownloaded from

AcknowledgmentsWe thank Abigail Spinner and Frank Ventimiglia (California National Pri-
mate Research Center, University of California, Davis) for help with flow
cytometry and assistancewith cell imaging, respectively, and Adam Treister
for FlowJo software. Special thanks to Dr. Frances Lund and Hiromi
Kubagawa (University of Alabama at Birmingham) for sharing the
sIgM2/2 mice and FcmR2/2 bone marrow, respectively, for these studies.
DisclosuresThe authors have no financial conflicts of interest.
References1. Ehrenstein, M. R., and C. A. Notley. 2010. The importance of natural IgM:
scavenger, protector and regulator. Nat. Rev. Immunol. 10: 778–786.2. Hooijkaas, H., A. A. van der Linde-Preesman, S. Benne, and R. Benner. 1985.
Frequency analysis of the antibody specificity repertoire of mitogen-reactiveB cells and “spontaneously” occurring “background” plaque-forming cells innude mice. Cell. Immunol. 92: 154–162.
3. Van Oudenaren, A., J. J. Haaijman, and R. Benner. 1984. Frequencies ofbackground cytoplasmic Ig-containing cells in various lymphoid organs ofathymic and euthymic mice as a function of age and immune status. Immunology51: 735–742.
4. Choi, Y. S., J. A. Dieter, K. Rothaeusler, Z. Luo, and N. Baumgarth. 2012. B-1cells in the bone marrow are a significant source of natural IgM. Eur. J. Immunol.42: 120–129.
5. Benner, R., W. Hijmans, and J. J. Haaijman. 1981. The bone marrow: the majorsource of serum immunoglobulins, but still a neglected site of antibody forma-tion. Clin. Exp. Immunol. 46: 1–8.
6. Hayakawa, K., M. Asano, S. A. Shinton, M. Gui, D. Allman, C. L. Stewart,J. Silver, and R. R. Hardy. 1999. Positive selection of natural autoreactiveB cells. Science 285: 113–116.
7. Berland, R., and H. H. Wortis. 2002. Origins and functions of B-1 cells withnotes on the role of CD5. Annu. Rev. Immunol. 20: 253–300.
8. Baumgarth, N., J. W. Tung, and L. A. Herzenberg. 2005. Inherent specificities innatural antibodies: a key to immune defense against pathogen invasion. SpringerSemin. Immunopathol. 26: 347–362.
9. Vas, J., C. Gronwall, A. Marshak-Rothstein, and G. J. Silverman. 2012. Naturalantibody to apoptotic cell membranes inhibits the proinflammatory properties oflupus autoantibody immune complexes. Arthritis Rheum. 64: 3388–3398.
10. Ogden, C. A., R. Kowalewski, Y. Peng, V. Montenegro, and K. B. Elkon. 2005.IGM is required for efficient complement mediated phagocytosis of apoptoticcells in vivo. Autoimmunity 38: 259–264.
11. Notley, C. A., M. A. Brown, G. P. Wright, and M. R. Ehrenstein. 2011. NaturalIgM is required for suppression of inflammatory arthritis by apoptotic cells. J.Immunol. 186: 4967–4972.
12. Boes, M., T. Schmidt, K. Linkemann, B. C. Beaudette, A. Marshak-Rothstein, andJ. Chen. 2000. Accelerated development of IgG autoantibodies and autoimmunedisease in the absence of secreted IgM. Proc. Natl. Acad. Sci. USA 97: 1184–1189.
13. Alugupalli, K. R., R. M. Gerstein, J. Chen, E. Szomolanyi-Tsuda,R. T. Woodland, and J. M. Leong. 2003. The resolution of relapsing fever bor-reliosis requires IgM and is concurrent with expansion of B1b lymphocytes. J.Immunol. 170: 3819–3827.
14. Haas, K. M., J. C. Poe, D. A. Steeber, and T. F. Tedder. 2005. B-1a and B-1bcells exhibit distinct developmental requirements and have unique functionalroles in innate and adaptive immunity to S. pneumoniae. Immunity 23: 7–18.
15. Martin, F., and J. F. Kearney. 2000. B-cell subsets and the mature preimmunerepertoire. Marginal zone and B1 B cells as part of a “natural immune memory”.Immunol. Rev. 175: 70–79.
16. Cebra, J. J., J. L. Komisar, and P. A. Schweitzer. 1984. CH isotype “switching”during normal B-lymphocyte development. Annu. Rev. Immunol. 2: 493–548.
17. Victora, G. D., and M. C. Nussenzweig. 2012. Germinal centers. Annu. Rev.Immunol. 30: 429–457.
18. Louis, A. G., and S. Gupta. 2014. Primary selective IgM deficiency: an ignoredimmunodeficiency. Clin. Rev. Allergy Immunol. 46: 104–111.
19. Baumgarth, N., O. C. Herman, G. C. Jager, L. E. Brown, L. A. Herzenberg, andJ. Chen. 2000. B-1 and B-2 cell-derived immunoglobulin M antibodies arenonredundant components of the protective response to influenza virus infection.J. Exp. Med. 192: 271–280.
20. Boes, M., A. P. Prodeus, T. Schmidt, M. C. Carroll, and J. Chen. 1998. A criticalrole of natural immunoglobulin M in immediate defense against systemic bac-terial infection. J. Exp. Med. 188: 2381–2386.
21. Choi, Y. S., and N. Baumgarth. 2008. Dual role for B-1a cells in immunity toinfluenza virus infection. J. Exp. Med. 205: 3053–3064.
22. Ochsenbein, A. F., T. Fehr, C. Lutz, M. Suter, F. Brombacher, H. Hengartner, andR. M. Zinkernagel. 1999. Control of early viral and bacterial distribution anddisease by natural antibodies. Science 286: 2156–2159.
23. Ehrenstein, M. R., H. T. Cook, and M. S. Neuberger. 2000. Deficiency in serumimmunoglobulin (Ig)M predisposes to development of IgG autoantibodies. J.Exp. Med. 191: 1253–1258.
24. Rowley, B., L. Tang, S. Shinton, K. Hayakawa, and R. R. Hardy. 2007. Auto-reactive B-1 B cells: constraints on natural autoantibody B cell antigen receptors.J. Autoimmun. 29: 236–245.
25. Mercolino, T. J., L. W. Arnold, L. A. Hawkins, and G. Haughton. 1988. Normalmouse peritoneum contains a large population of Ly-1+ (CD5) B cells thatrecognize phosphatidyl choline. Relationship to cells that secrete hemolyticantibody specific for autologous erythrocytes. J. Exp. Med. 168: 687–698.
26. Erikson, J., M. Z. Radic, S. A. Camper, R. R. Hardy, C. Carmack, andM. Weigert. 1991. Expression of anti-DNA immunoglobulin transgenes in non-autoimmune mice. Nature 349: 331–334.
27. Goodnow, C. C., J. Crosbie, S. Adelstein, T. B. Lavoie, S. J. Smith-Gill,R. A. Brink, H. Pritchard-Briscoe, J. S. Wotherspoon, R. H. Loblay, K. Raphael,et al. 1988. Altered immunoglobulin expression and functional silencing of self-reactive B lymphocytes in transgenic mice. Nature 334: 676–682.
28. Ota, T., C. Doyle-Cooper, A. B. Cooper, K. J. Doores, M. Aoki-Ota, K. Le,W. R. Schief, R. T. Wyatt, D. R. Burton, and D. Nemazee. 2013. B cells fromknock-in mice expressing broadly neutralizing HIV antibody b12 carry an in-nocuous B cell receptor responsive to HIV vaccine candidates. J. Immunol. 191:3179–3185.
29. Nemazee, D. A., and K. B€urki. 1989. Clonal deletion of B lymphocytes in a trans-genic mouse bearing anti-MHC class I antibody genes. Nature 337: 562–566.
30. Hardy, R. R., and K. Hayakawa. 2001. B cell development pathways. Annu. Rev.Immunol. 19: 595–621.
31. von Boehmer, H., and F. Melchers. 2010. Checkpoints in lymphocyte develop-ment and autoimmune disease. Nat. Immunol. 11: 14–20.
32. Hippen, K. L., L. E. Tze, and T. W. Behrens. 2000. CD5 maintains tolerance inanergic B cells. J. Exp. Med. 191: 883–890.
33. Pillai, S., and A. Cariappa. 2009. The follicular versus marginal zoneB lymphocyte cell fate decision. Nat. Rev. Immunol. 9: 767–777.
34. Casola, S., K. L. Otipoby, M. Alimzhanov, S. Humme, N. Uyttersprot,J. L. Kutok, M. C. Carroll, and K. Rajewsky. 2004. B cell receptor signalstrength determines B cell fate. Nat. Immunol. 5: 317–327.
35. Kubagawa, H., S. Oka, Y. Kubagawa, I. Torii, E. Takayama, D. W. Kang,G. L. Gartland, L. F. Bertoli, H. Mori, H. Takatsu, et al. 2009. Identity of theelusive IgM Fc receptor (FcmuR) in humans. J. Exp. Med. 206: 2779–2793.
36. Shima, H., H. Takatsu, S. Fukuda, M. Ohmae, K. Hase, H. Kubagawa,J. Y. Wang, and H. Ohno. 2010. Identification of TOSO/FAIM3 as an Fc receptorfor IgM. Int. Immunol. 22: 149–156.
37. Choi, S. C., H. Wang, L. Tian, Y. Murakami, D. M. Shin, F. Borrego,H. C. Morse, III, and J. E. Coligan. 2013. Mouse IgM Fc receptor, FCMR,promotes B cell development and modulates antigen-driven immune responses.J. Immunol. 190: 987–996.
38. Ouchida, R., H. Mori, K. Hase, H. Takatsu, T. Kurosaki, T. Tokuhisa, H. Ohno,and J. Y. Wang. 2012. Critical role of the IgM Fc receptor in IgM homeostasis,B-cell survival, and humoral immune responses. Proc. Natl. Acad. Sci. USA 109:E2699–E2706.
39. Baker, N., and M. R. Ehrenstein. 2002. Cutting edge: selection of B lymphocytesubsets is regulated by natural IgM. J. Immunol. 169: 6686–6690.
40. Boes, M., C. Esau, M. B. Fischer, T. Schmidt, M. Carroll, and J. Chen. 1998.Enhanced B-1 cell development, but impaired IgG antibody responses in micedeficient in secreted IgM. J. Immunol. 160: 4776–4787.
41. Rothaeusler, K., and N. Baumgarth. 2006. Evaluation of intranuclear BrdU de-tection procedures for use in multicolor flow cytometry. Cytometry A 69: 249–259.
42. Yamamoto, T., S. Takagawa, I. Katayama, K. Yamazaki, Y. Hamazaki,H. Shinkai, and K. Nishioka. 1999. Animal model of sclerotic skin. I: Localinjections of bleomycin induce sclerotic skin mimicking scleroderma. J. Invest.Dermatol. 112: 456–462.
43. Fuxa, M., J. Skok, A. Souabni, G. Salvagiotto, E. Roldan, and M. Busslinger.2004. Pax5 induces V-to-DJ rearrangements and locus contraction of the im-munoglobulin heavy-chain gene. Genes Dev. 18: 411–422.
44. Vieira, P., and K. Rajewsky. 1988. The half-lives of serum immunoglobulins inadult mice. Eur. J. Immunol. 18: 313–316.
45. Arnold, L. W., C. A. Pennell, S. K. McCray, and S. H. Clarke. 1994. Develop-ment of B-1 cells: segregation of phosphatidyl choline-specific B cells to the B-1population occurs after immunoglobulin gene expression. J. Exp. Med. 179:1585–1595.
46. Hardy, R. R., C. J. Wei, and K. Hayakawa. 2004. Selection during developmentof VH11+ B cells: a model for natural autoantibody-producing CD5+ B cells.Immunol. Rev. 197: 60–74.
47. Cambier, J. C., S. B. Gauld, K. T. Merrell, and B. J. Vilen. 2007. B-cell anergy:from transgenic models to naturally occurring anergic B cells? Nat. Rev.Immunol. 7: 633–643.
48. Notley, C. A., N. Baker, and M. R. Ehrenstein. 2010. Secreted IgM enhancesB cell receptor signaling and promotes splenic but impairs peritoneal B cellsurvival. J. Immunol. 184: 3386–3393.
49. Honjo, K., Y. Kubagawa, D. M. Jones, B. Dizon, Z. Zhu, H. Ohno, S. Izui,J. F. Kearney, and H. Kubagawa. 2012. Altered Ig levels and antibody responsesin mice deficient for the Fc receptor for IgM (FcmR). Proc. Natl. Acad. Sci. USA109: 15882–15887.
50. Nguyen, X. H., P. A. Lang, K. S. Lang, D. Adam, G. Fattakhova, N. Foger,M. A. Kamal, P. Prilla, S. Mathieu, C. Wagner, et al. 2011. Toso regulates thebalance between apoptotic and nonapoptotic death receptor signaling by facili-tating RIP1 ubiquitination. Blood 118: 598–608.
51. Molina, H., V. M. Holers, B. Li, Y. Fung, S. Mariathasan, J. Goellner, J. Strauss-Schoenberger, R. W. Karr, and D. D. Chaplin. 1996. Markedly impaired humoralimmune response in mice deficient in complement receptors 1 and 2. Proc. Natl.Acad. Sci. USA 93: 3357–3361.
52. Sakamoto, N., K. Shibuya, Y. Shimizu, K. Yotsumoto, T. Miyabayashi,S. Sakano, T. Tsuji, E. Nakayama, H. Nakauchi, and A. Shibuya. 2001. A novel
The Journal of Immunology 1501
by guest on June 12, 2016http://w
ww
.jimm
unol.org/D
ownloaded from

Fc receptor for IgA and IgM is expressed on both hematopoietic and non-hematopoietic tissues. Eur. J. Immunol. 31: 1310–1316.
53. Honda, S., N. Kurita, A. Miyamoto, Y. Cho, K. Usui, K. Takeshita, S. Takahashi,T. Yasui, H. Kikutani, T. Kinoshita, et al. 2009. Enhanced humoral immuneresponses against T-independent antigens in Fca/mR-deficient mice. Proc. Natl.Acad. Sci. USA 106: 11230–11235.
54. Herzog, S., M. Reth, and H. Jumaa. 2009. Regulation of B-cell proliferationand differentiation by pre-B-cell receptor signalling. Nat. Rev. Immunol. 9:195–205.
55. Kantor, A. B., and L. A. Herzenberg. 1993. Origin of murine B cell lineages.Annu. Rev. Immunol. 11: 501–538.
56. Deenen, G. J., and F. G. Kroese. 1992. Murine peritoneal Ly-1 B cells do not turnover rapidly. Ann. N. Y. Acad. Sci. 651: 70–71.
57. Wardemann, H., T. Boehm, N. Dear, and R. Carsetti. 2002. B-1a B cells that linkthe innate and adaptive immune responses are lacking in the absence of thespleen. J. Exp. Med. 195: 771–780.
58. Montecino-Rodriguez, E., and K. Dorshkind. 2012. B-1 B cell development inthe fetus and adult. Immunity 36: 13–21.
1502 SECRETED IgM REGULATES B CELL SELECTION
by guest on June 12, 2016http://w
ww
.jimm
unol.org/D
ownloaded from