multinutrient undernutrition dysregulates the resident macrophage proinflammatory cytokine network,...
TRANSCRIPT

Multinutrient undernutrition dysregulates the residentmacrophage proinflammatory cytokine network, nuclearfactor-�B activation, and nitric oxide production
Gregory M. Anstead,*,†,1 Bysani Chandrasekar,† Qiong Zhang,† and Peter C. Melby*,†
*Medical Service, Department of Veterans Affairs Medical Center, South Texas Veterans Health Care System and†Department of Medicine, University of Texas Health Science Center at San Antonio San Antonio, TX
Abstract: We have described previously a murinemodel of multinutrient undernutrition that repro-duced the features of moderate human malnutri-tion and led to increased early dissemination ofLeishmania donovani. Peritoneal cells from thesemalnourished mice produced decreased NO afterstimulation with IFN-�/LPS. We hypothesized thatmalnutrition may cause a deficit in NF-�B activa-tion, a principal transcription pathway for induc-ible NO synthase and proinflammatory cytokines.Macrophages from malnourished mice, stimulatedwith IFN-�/LPS, showed increased IL-6 productionand decreased IL-10 and TNF-� production. Neu-tralization of TNF-� in macrophage cultures fromthe control mice mimicked the effect of malnutri-tion on NO and IL-10 production, whereas supple-mental TNF-� added to cultures of macrophagesfrom malnourished mice increased NO secretion.NF-�B nuclear binding activity in macrophagesfrom the malnourished mice was reduced earlyafter stimulation, but increased to supranormalvalues by 16- or 24-h poststimulation. BlockingNO production in the macrophages from the con-trol mice reproduced the effect of malnutrition onthe late activation of NF-�B, whereas supplementalNO decreased the late NF-�B activation in themalnourished mice. Thus, in macrophages fromthe malnourished mice, initial deficits in NF-�Bactivity probably lead to decreased TNF-�, whichresults in decreased NO; however, IL-6 is regu-lated independently from NF-�B and TNF-�. Thelate activation of NF-�B in the macrophages frommalnourished mice is due to absence of negativefeedback from NO. J. Leukoc. Biol. 74: 982–991;2003.
Key words: transcription factor � immunodeficiency � tumor necro-sis factor-� � interleukin-6 � interleukin-10
INTRODUCTION
In synergy with infection, malnutrition contributes to 56% ofall childhood deaths worldwide [1]. However, the molecularand cellular bases for the immunosuppression produced by
malnutrition are not well understood [2]. In particular, theimpact of malnutrition on innate immunity has not been ade-quately addressed [3]. Innate immunity refers to host defensemediated by macrophages, monocytes, neutrophils, and NKcells, which recognize pathogens using receptors that bind tobroadly expressed microbial products, such as LPS [4]. Mac-rophages are central to the innate and acquired immune re-sponses; they are activated as part of the innate immuneresponse by a variety of stimuli. IFN-� is the most potentactivating cytokine of macrophages [5] and LPS is the best-studied microbial stimulus [6]. After treatment with IFN-� andLPS, macrophages release TNF-�, IL-1�, and IL-6, the proin-flammatory cytokines [7]. Preceding proinflammatory cytokineproduction is the activation of the transcription factor NF-�B,which promotes the expression of these cytokines, chemokines,adhesion molecules, and inducible nitric oxide synthase(NOS2) [8, 9]. The effects of malnutrition on macrophagefunction have been addressed in several studies [10–13].However, these papers have not assessed the influence ofmalnutrition on the proinflammatory cytokine network andNF-�B regulation.
Previously, we developed a mouse model of multinutrientundernutrition (protein-energy, zinc, and iron deficiency) thatmimicked the growth characteristics of human weanling mal-nutrition and predisposed to increased dissemination of theparasite Leishmania donovani [3]. In this study, we use thismodel to demonstrate that multinutrient undernutrition (hence-forth referred to as malnutrition) leads to extensive dysfunctionof the macrophage proinflammatory cytokine network andNF-�B regulation.
MATERIALS AND METHODS
Mice
Weanling (3-wk old) female nu/� BALB/c mice (heterozygous at the nu locus,functionally normal) were obtained from the breeding colony of the South TexasVeterans Health Care System, San Antonio, TX.
1 Correspondence: Division of Infectious Diseases, Department of Medicine,University of Texas Health Science Center at San Antonio 7703 Floyd CurlDrive San Antonio, TX 78229-3900. E-mail: [email protected]
Received February 10, 2003; revised June 18, 2003; accepted June 19,2003; doi: 10.1189/jlb.0203064.
982 Journal of Leukocyte Biology Volume 74, December 2003 http://www.jleukbio.org

Diets and Feeding
Mice received a 3-day acclimation on standard chow after weaning and priorto the change to the two experimental diets. The control group (well-nourished(WN)) mice received a diet that contained 17% protein, 100 ppm iron, 30 ppmzinc, which was provided ad libitum. The mice in the malnourished (MN) groupreceived a diet with 3% protein, 10 ppm iron, 1 ppm zinc; these mice received12% less food by weight per day compared with the mice in the control group[3]. Both diets provided 3.9 kcal/g of chow. Prior to the experimental feeding,the mice were weight-matched and both groups had free access to water. Themice were housed in groups of four with low trace element bedding (Apha-Dri;Shepard Specialty Papers, Kalamazoo, MI).
Culture of resident peritoneal macrophages
Peritoneal cells were collected from mice in both diet groups by lavage usingDMEM with 2% FBS. Typically, 8�12 mice from each diet group were usedin each experiment. The cells were plated at 1 � 106 cells per mL in 10% FBSin DMEM with 1% (vol/vol) 1 M HEPES, 1% (vol/vol) penicillin-streptomycin(each at 10,000 IU/mL), and 0.1 mM 2-mercaptomethanol. The cells wereallowed to adhere for 3 h and then washed 3 times with PBS to removenonadherent cells. Fresh media (DMEM with 10% FBS) was added. Theadherent cells were treated with IFN-� (PharMingen, San Diego, CA; 80U/mL). After 1 h, LPS (Escherichia coli 0111:B4; Sigma, St. Louis, MO) (20ng/mL)) was added. Supernatants and/or cells were collected at the timesindicated in the results section. In specific experiments (see below), thestimulated macrophages were treated with a NOS2 inhibitor, an NO donor, amAb against TNF-�, or supplemental TNF-�.
NOS2 was inhibited in macrophage cultures using L-N6-(1-iminoethyl)lysine (L-NIL) (Calbiochem Novabopchem Corp., San Diego, CA); L-NIL wasadded to the cultures from the WN mice 15-min after the addition of LPS to aconcentration of 1 mM. After 24 h, the supernatants were analyzed for TNF-�,IL-6, IL-10, and NO and the cells were harvested for NF-�B determination.
Supplemental NO was provided to the IFN-�/LPS-stimulated macrophagecultures from the MN mice by adding sodium nitroprusside (SNP; Calbiochem)15-min after addition of LPS to a concentration of 0.1 mM. After 24 h, thesupernatants were assayed for TNF-�, IL-6, IL-10, and NO, and the cells wereharvested for NF-�B determination.
TNF-� was neutralized in the macrophage cultures using a rat anti-mouseIgG1 mAb (PharMingen). The mAb was added to macrophage cultures from theWN mice 15-min after the addition of LPS, to give concentrations of anti-TNF-� mAb of either 20 �g/mL or 50 ng/mL. After 24 h, the supernatants wereanalyzed for TNF-�, IL-6, IL-10, and NO.
Supplemental TNF-� was provided to the IFN-�/LPS-stimulated macro-phage cultures from the MN mice by adding murine TNF-� (PharMingen)60-min after addition of LPS to a concentration of 500 U/mL. After 24 h, thesupernatants were assayed for TNF-�, IL-6, IL-10, and NO.
NO production by macrophages
Supernatants from the macrophage cultures were tested for nitrite by the Griessreaction, after conversion of nitrate to nitrite with nitrate reductase (colorimet-ric assay; Cayman Chemical Co., Ann Arbor, MI).
Cytokine assays
Cytokine levels were determined by ELISA, using the following kits: IL-6,IL-10, TNF-� (eBioscience, San Diego, CA), and IL-1� (R&D Systems,Minneapolis, MN).
NF-�B DNA binding activity and dimercomposition
Isolation of nuclear protein extracts and EMSA were performed as describedpreviously [14]. A ds oligonucleotide (Santa Cruz Biotechnology, Santa Cruz, CA)containing a tandem repeat of the decameric consensus sequence (5-GGGACTT-TCC-3) was used as a probe. For the competition assay, the protein extract (20 �g)was preincubated with homologous unlabeled oligonucleotide for 5 min on ice,followed by the addition of labeled probe. Absence of protein extract, competitionwith 100-fold molar excess unlabeled NF-�B, and mutant NF-�B oligo (5-AGTTGA GGC GAC TTT CCC AGG C-3; Santa Cruz Biotechnology) served ascontrols. In the gel supershift assay to determine the dimeric composition of
NF-�B, the protein extract (20 �g) was preincubated for 40 min on ice with eitheranti-p50 or -p65 subunit-specific polyclonal antibodies (1 �g; Santa Cruz Bio-technology) prior to the addition of 32P-labeled NF-�B consensus oligo.
Proinflammatory cytokine and NOS2 mRNAexpression by Northern blot analysisRNA extraction, Northern blotting, autoradiography, and densitometry were per-formed as described previously [15]. Total RNA was extracted from the culturedmacrophages using acid-guanidium isothiocyanate-phenol-chloroform. The RNAwas denatured in 2.2 M formaldehyde, size fractionated on 0.8% agarose gelscontaining 0.5 g/ml ethidium bromide to check RNA integrity and loading equiv-alency, and electroblotted at 4°C onto a nitrocellulose membrane (Schleicher andSchuell, Keene, NH) in 0.025 M phosphate buffer, pH 6.5. Ribonucleic acids wereUV cross-linked to the membrane. The blot was prehybridized for 4 h at 42°C ina prehybridization buffer that contained 50% formamide, 0.1% SDS, 5 � SSC,2.5 � Denhardt’s, 250 �g/ml denatured sonicated salmon sperm DNA (Stratagene,La Jolla, CA), 50 mM Na2PO4, pH 6.5. The blots were then hybridized at 42°C for16 h with 32P-labeled probes (6 � 105 cpm/ml), washed twice at 23°C in 6 �SSC/0.5% SDS, twice at 37°C in 1 � SSC/0.5% SDS, and once at 57°C in 0.l �SSC/0.5% SDS. All blots were then exposed at �80°C to Kodak XAR-5 film withKodak-intensifying screens, and the intensities of the autoradiographic bands werequantified by videoimaging. The same membrane was reprobed after stripping offits previous label. mRNA sizes were determined in relation to the mobility of 28Sand 18S rRNA and an mRNA ladder (GibcoBRL, Grand Island, NY). The cDNAprobes (NOS2, IL-6, and TNF-�; American Type Culture Collection, Manassas,VA) were labeled with [�-32P]dCTP (3000 Ci/mmol; Amersham, Piscataway, NJ)using random hexanucleotide primers (Boehringer-Mannheim, Indianapolis, IN),and the oligonucleotide probe (h28S rRNA; 40 base ss oligo; Oncogene Science,Uniondale, NY) was 5 end-labeled with (�-32P)ATP using T4 polynucleotidekinase.
StatisticsResults are expressed as the mean and standard error of the mean. Compar-isons between group means were performed using Student’s two-tailed t test.
RESULTS
Malnutrition produced decreased weight-for-age
Mice on the control diet received chow ad libitum and con-sumed an average of 3.2 g/d. Mice in the deficient diet groupwere provided 2.8 g/d. At the end of 6 wks of feeding, mice inthe deficient diet group showed decreased weight-for-age andan altered growth curve (Fig. 1). The MN mice had a slowlydeclining growth curve (12.5% weight loss) analogous to hu-man weanling malnutrition and were an average of 69% ofweight-for-age compared with the controls (P 0.001), whichcorresponds to moderate malnutrition [3].
Macrophages from malnourished mice producedless NO after stimulation
As described previously with resident peritoneal cells [3], residentperitoneal macrophages (isolated by adherence to plastic) from theMN mice also produced less NO after stimulation with IFN-� andLPS (Fig. 2A). At 1 h after stimulation, there were equivalent lowlevels of NO produced by both groups of macrophages. However,at 16 and 24 h, NO production by macrophages from the MN micewas reduced 54 %and 49%, respectively, compared with thosefrom the WN mice (P � 0.04).
Macrophages from malnourished mice had analtered proinflammatory response to IFN-�/LPS
At four time points (1-, 4-, 16-, and 24 h), resident peritonealmacrophages from the MN mice produced decreased levels of
Anstead et al. Undernutrition and macrophage cytokine network 983
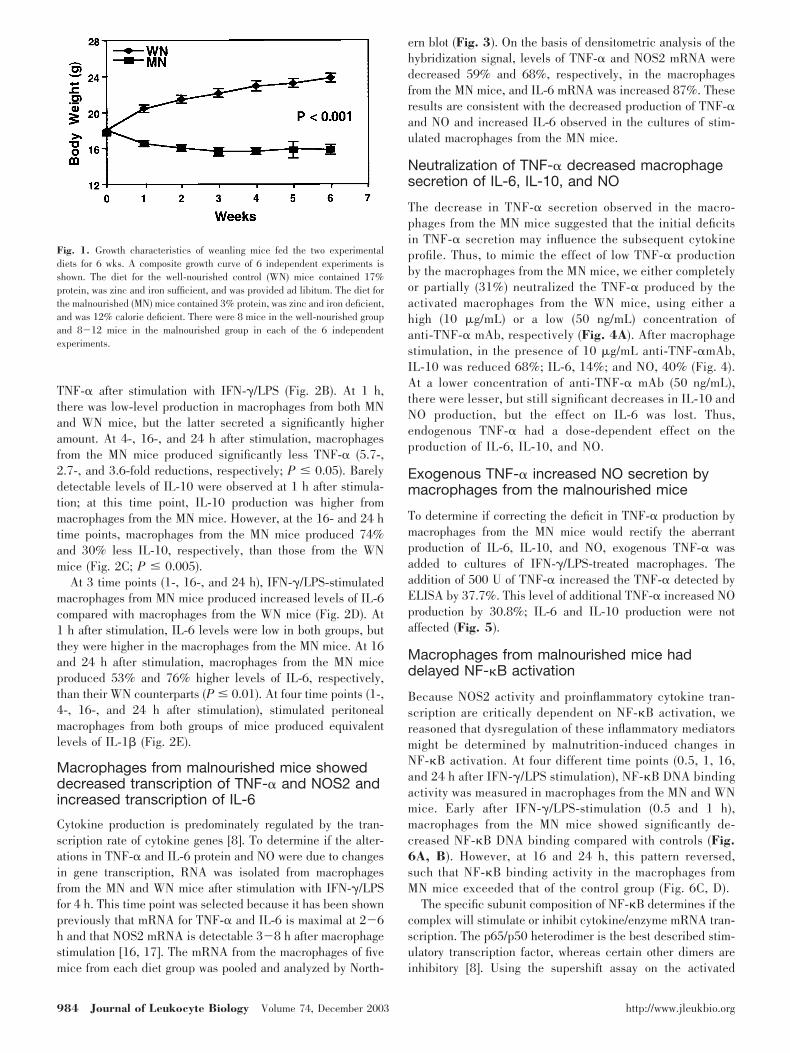
TNF-� after stimulation with IFN-�/LPS (Fig. 2B). At 1 h,there was low-level production in macrophages from both MNand WN mice, but the latter secreted a significantly higheramount. At 4-, 16-, and 24 h after stimulation, macrophagesfrom the MN mice produced significantly less TNF-� (5.7-,2.7-, and 3.6-fold reductions, respectively; P � 0.05). Barelydetectable levels of IL-10 were observed at 1 h after stimula-tion; at this time point, IL-10 production was higher frommacrophages from the MN mice. However, at the 16- and 24 htime points, macrophages from the MN mice produced 74%and 30% less IL-10, respectively, than those from the WNmice (Fig. 2C; P � 0.005).
At 3 time points (1-, 16-, and 24 h), IFN-�/LPS-stimulatedmacrophages from MN mice produced increased levels of IL-6compared with macrophages from the WN mice (Fig. 2D). At1 h after stimulation, IL-6 levels were low in both groups, butthey were higher in the macrophages from the MN mice. At 16and 24 h after stimulation, macrophages from the MN miceproduced 53% and 76% higher levels of IL-6, respectively,than their WN counterparts (P � 0.01). At four time points (1-,4-, 16-, and 24 h after stimulation), stimulated peritonealmacrophages from both groups of mice produced equivalentlevels of IL-1� (Fig. 2E).
Macrophages from malnourished mice showeddecreased transcription of TNF-� and NOS2 andincreased transcription of IL-6
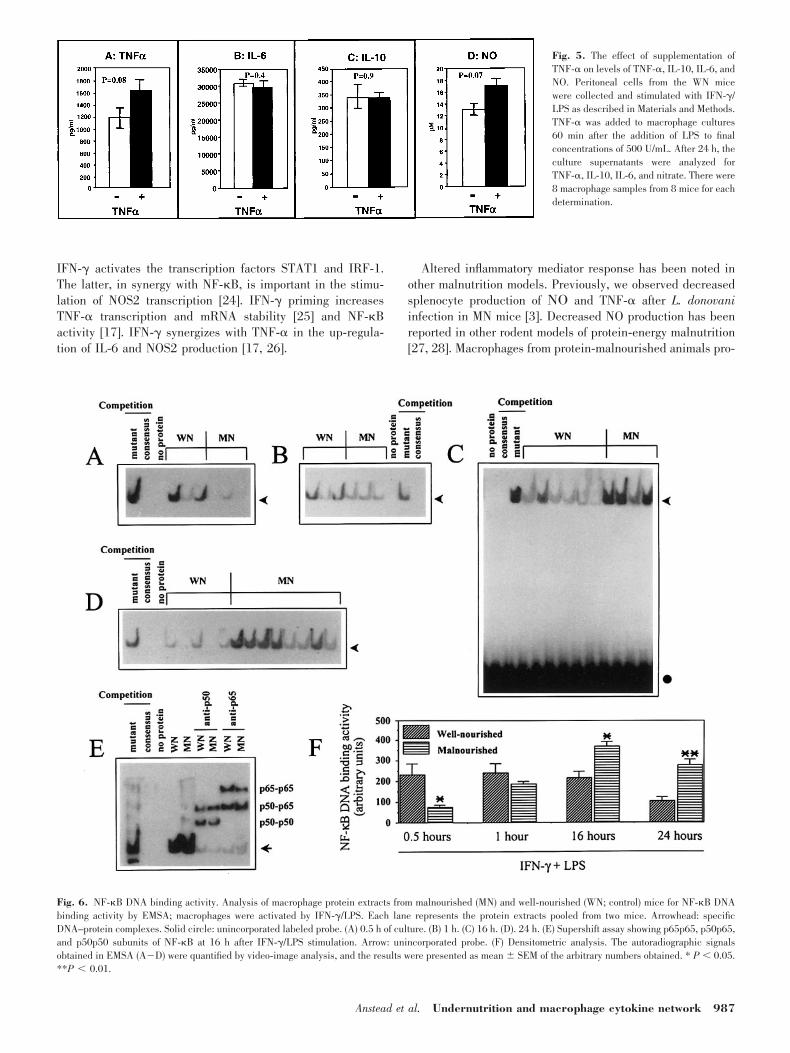
Cytokine production is predominately regulated by the tran-scription rate of cytokine genes [8]. To determine if the alter-ations in TNF-� and IL-6 protein and NO were due to changesin gene transcription, RNA was isolated from macrophagesfrom the MN and WN mice after stimulation with IFN-�/LPSfor 4 h. This time point was selected because it has been shownpreviously that mRNA for TNF-� and IL-6 is maximal at 2�6h and that NOS2 mRNA is detectable 3�8 h after macrophagestimulation [16, 17]. The mRNA from the macrophages of fivemice from each diet group was pooled and analyzed by North-
ern blot (Fig. 3). On the basis of densitometric analysis of thehybridization signal, levels of TNF-� and NOS2 mRNA weredecreased 59% and 68%, respectively, in the macrophagesfrom the MN mice, and IL-6 mRNA was increased 87%. Theseresults are consistent with the decreased production of TNF-�and NO and increased IL-6 observed in the cultures of stim-ulated macrophages from the MN mice.
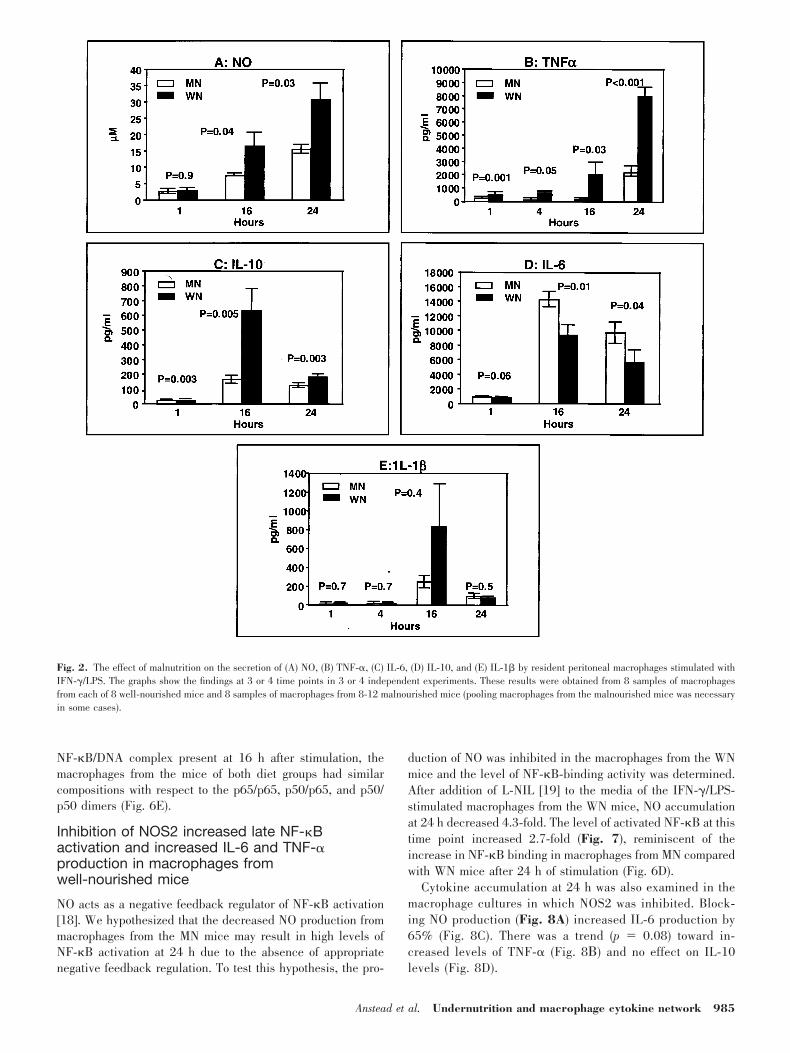
Neutralization of TNF-� decreased macrophagesecretion of IL-6, IL-10, and NO
The decrease in TNF-� secretion observed in the macro-phages from the MN mice suggested that the initial deficitsin TNF-� secretion may influence the subsequent cytokineprofile. Thus, to mimic the effect of low TNF-� productionby the macrophages from the MN mice, we either completelyor partially (31%) neutralized the TNF-� produced by theactivated macrophages from the WN mice, using either ahigh (10 �g/mL) or a low (50 ng/mL) concentration ofanti-TNF-� mAb, respectively (Fig. 4A). After macrophagestimulation, in the presence of 10 �g/mL anti-TNF-�mAb,IL-10 was reduced 68%; IL-6, 14%; and NO, 40% (Fig. 4).At a lower concentration of anti-TNF-� mAb (50 ng/mL),there were lesser, but still significant decreases in IL-10 andNO production, but the effect on IL-6 was lost. Thus,endogenous TNF-� had a dose-dependent effect on theproduction of IL-6, IL-10, and NO.
Exogenous TNF-� increased NO secretion bymacrophages from the malnourished mice
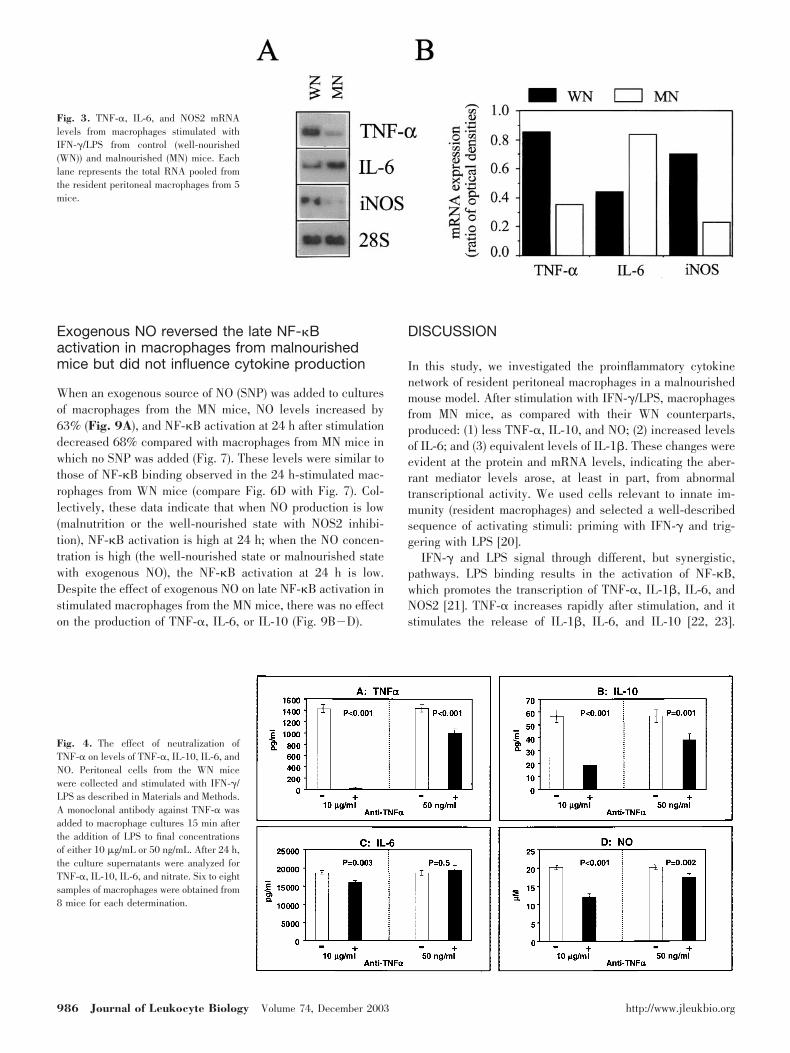
To determine if correcting the deficit in TNF-� production bymacrophages from the MN mice would rectify the aberrantproduction of IL-6, IL-10, and NO, exogenous TNF-� wasadded to cultures of IFN-�/LPS-treated macrophages. Theaddition of 500 U of TNF-� increased the TNF-� detected byELISA by 37.7%. This level of additional TNF-� increased NOproduction by 30.8%; IL-6 and IL-10 production were notaffected (Fig. 5).
Macrophages from malnourished mice haddelayed NF-�B activation
Because NOS2 activity and proinflammatory cytokine tran-scription are critically dependent on NF-�B activation, wereasoned that dysregulation of these inflammatory mediatorsmight be determined by malnutrition-induced changes inNF-�B activation. At four different time points (0.5, 1, 16,and 24 h after IFN-�/LPS stimulation), NF-�B DNA bindingactivity was measured in macrophages from the MN and WNmice. Early after IFN-�/LPS-stimulation (0.5 and 1 h),macrophages from the MN mice showed significantly de-creased NF-�B DNA binding compared with controls (Fig.6A, B). However, at 16 and 24 h, this pattern reversed,such that NF-�B binding activity in the macrophages fromMN mice exceeded that of the control group (Fig. 6C, D).
The specific subunit composition of NF-�B determines if thecomplex will stimulate or inhibit cytokine/enzyme mRNA tran-scription. The p65/p50 heterodimer is the best described stim-ulatory transcription factor, whereas certain other dimers areinhibitory [8]. Using the supershift assay on the activated
Fig. 1. Growth characteristics of weanling mice fed the two experimentaldiets for 6 wks. A composite growth curve of 6 independent experiments isshown. The diet for the well-nourished control (WN) mice contained 17%protein, was zinc and iron sufficient, and was provided ad libitum. The diet forthe malnourished (MN) mice contained 3% protein, was zinc and iron deficient,and was 12% calorie deficient. There were 8 mice in the well-nourished groupand 8�12 mice in the malnourished group in each of the 6 independentexperiments.
984 Journal of Leukocyte Biology Volume 74, December 2003 http://www.jleukbio.org
NF-�B/DNA complex present at 16 h after stimulation, themacrophages from the mice of both diet groups had similarcompositions with respect to the p65/p65, p50/p65, and p50/p50 dimers (Fig. 6E).
Inhibition of NOS2 increased late NF-�Bactivation and increased IL-6 and TNF-�production in macrophages fromwell-nourished mice
NO acts as a negative feedback regulator of NF-�B activation[18]. We hypothesized that the decreased NO production frommacrophages from the MN mice may result in high levels ofNF-�B activation at 24 h due to the absence of appropriatenegative feedback regulation. To test this hypothesis, the pro-
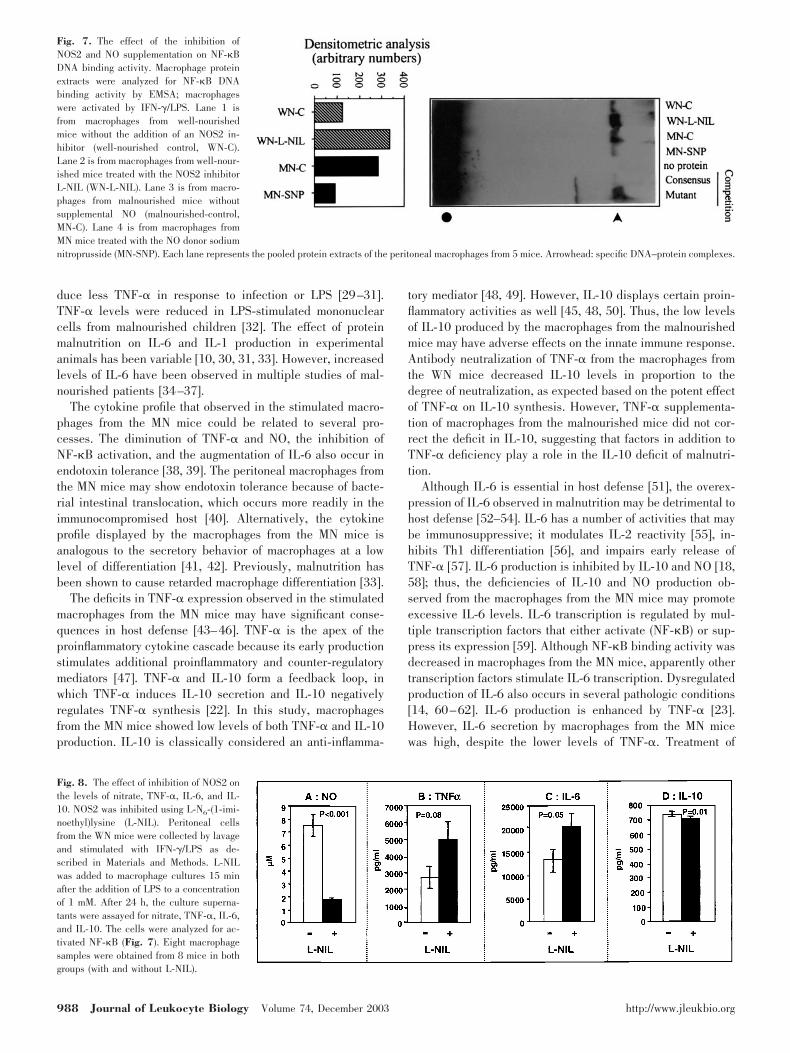
duction of NO was inhibited in the macrophages from the WNmice and the level of NF-�B-binding activity was determined.After addition of L-NIL [19] to the media of the IFN-�/LPS-stimulated macrophages from the WN mice, NO accumulationat 24 h decreased 4.3-fold. The level of activated NF-�B at thistime point increased 2.7-fold (Fig. 7), reminiscent of theincrease in NF-�B binding in macrophages from MN comparedwith WN mice after 24 h of stimulation (Fig. 6D).
Cytokine accumulation at 24 h was also examined in themacrophage cultures in which NOS2 was inhibited. Block-ing NO production (Fig. 8A) increased IL-6 production by65% (Fig. 8C). There was a trend (p � 0.08) toward in-creased levels of TNF-� (Fig. 8�) and no effect on IL-10levels (Fig. 8D).
Fig. 2. The effect of malnutrition on the secretion of (A) NO, (B) TNF-�, (C) IL-6, (D) IL-10, and (E) IL-1� by resident peritoneal macrophages stimulated withIFN-�/LPS. The graphs show the findings at 3 or 4 time points in 3 or 4 independent experiments. These results were obtained from 8 samples of macrophagesfrom each of 8 well-nourished mice and 8 samples of macrophages from 8-12 malnourished mice (pooling macrophages from the malnourished mice was necessaryin some cases).
Anstead et al. Undernutrition and macrophage cytokine network 985
Exogenous NO reversed the late NF-�Bactivation in macrophages from malnourishedmice but did not influence cytokine production
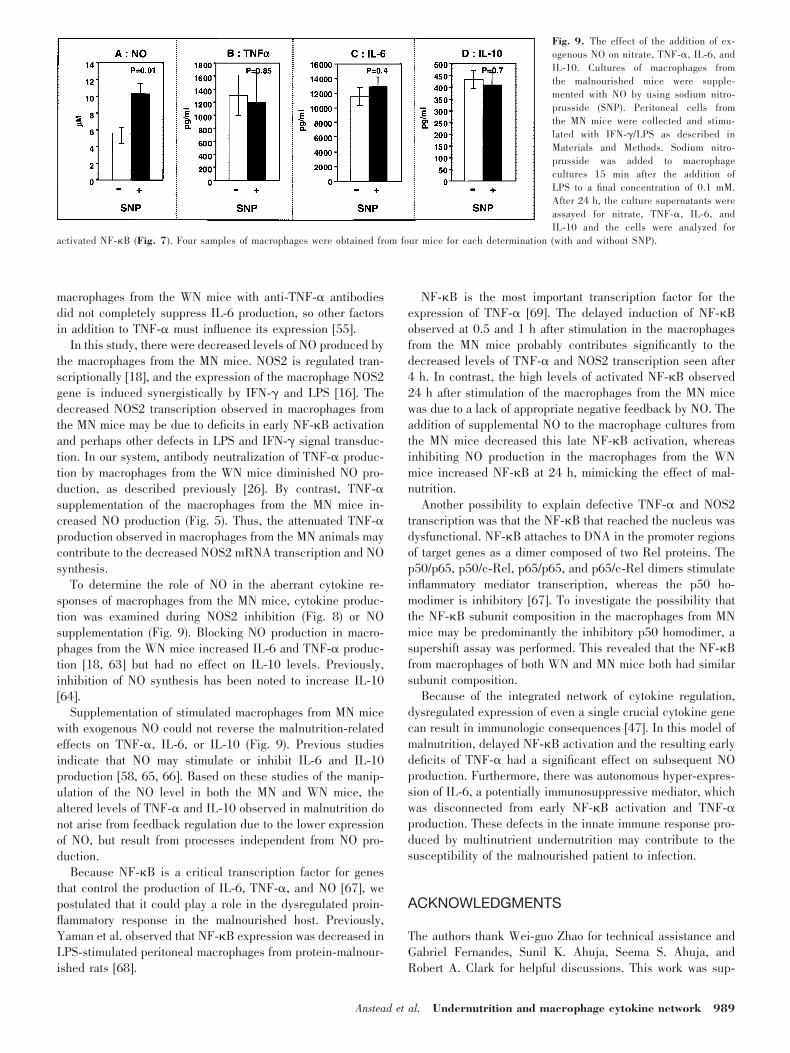
When an exogenous source of NO (SNP) was added to culturesof macrophages from the MN mice, NO levels increased by63% (Fig. 9A), and NF-�B activation at 24 h after stimulationdecreased 68% compared with macrophages from MN mice inwhich no SNP was added (Fig. 7). These levels were similar tothose of NF-�B binding observed in the 24 h-stimulated mac-rophages from WN mice (compare Fig. 6D with Fig. 7). Col-lectively, these data indicate that when NO production is low(malnutrition or the well-nourished state with NOS2 inhibi-tion), NF-�B activation is high at 24 h; when the NO concen-tration is high (the well-nourished state or malnourished statewith exogenous NO), the NF-�B activation at 24 h is low.Despite the effect of exogenous NO on late NF-�B activation instimulated macrophages from the MN mice, there was no effecton the production of TNF-�, IL-6, or IL-10 (Fig. 9B�D).
DISCUSSION
In this study, we investigated the proinflammatory cytokinenetwork of resident peritoneal macrophages in a malnourishedmouse model. After stimulation with IFN-�/LPS, macrophagesfrom MN mice, as compared with their WN counterparts,produced: (1) less TNF-�, IL-10, and NO; (2) increased levelsof IL-6; and (3) equivalent levels of IL-1�. These changes wereevident at the protein and mRNA levels, indicating the aber-rant mediator levels arose, at least in part, from abnormaltranscriptional activity. We used cells relevant to innate im-munity (resident macrophages) and selected a well-describedsequence of activating stimuli: priming with IFN-� and trig-gering with LPS [20].
IFN-� and LPS signal through different, but synergistic,pathways. LPS binding results in the activation of NF-�B,which promotes the transcription of TNF-�, IL-1�, IL-6, andNOS2 [21]. TNF-� increases rapidly after stimulation, and itstimulates the release of IL-1�, IL-6, and IL-10 [22, 23].
Fig. 4. The effect of neutralization ofTNF-� on levels of TNF-�, IL-10, IL-6, andNO. Peritoneal cells from the WN micewere collected and stimulated with IFN-�/LPS as described in Materials and Methods.A monoclonal antibody against TNF-� wasadded to macrophage cultures 15 min afterthe addition of LPS to final concentrationsof either 10 �g/mL or 50 ng/mL. After 24 h,the culture supernatants were analyzed forTNF-�, IL-10, IL-6, and nitrate. Six to eightsamples of macrophages were obtained from8 mice for each determination.
Fig. 3. TNF-�, IL-6, and NOS2 mRNAlevels from macrophages stimulated withIFN-�/LPS from control (well-nourished(WN)) and malnourished (MN) mice. Eachlane represents the total RNA pooled fromthe resident peritoneal macrophages from 5mice.
986 Journal of Leukocyte Biology Volume 74, December 2003 http://www.jleukbio.org
IFN-� activates the transcription factors STAT1 and IRF-1.The latter, in synergy with NF-�B, is important in the stimu-lation of NOS2 transcription [24]. IFN-� priming increasesTNF-� transcription and mRNA stability [25] and NF-�Bactivity [17]. IFN-� synergizes with TNF-� in the up-regula-tion of IL-6 and NOS2 production [17, 26].
Altered inflammatory mediator response has been noted inother malnutrition models. Previously, we observed decreasedsplenocyte production of � and TNF-� after L. donovaniinfection in MN mice [3]. Decreased NO production has beenreported in other rodent models of protein-energy malnutrition[27, 28]. Macrophages from protein-malnourished animals pro-
Fig. 5. The effect of supplementation ofTNF-� on levels of TNF-�, IL-10, IL-6, andNO. Peritoneal cells from the WN micewere collected and stimulated with IFN-�/LPS as described in Materials and Methods.TNF-� was added to macrophage cultures60 min after the addition of LPS to finalconcentrations of 500 U/mL. After 24 h, theculture supernatants were analyzed forTNF-�, IL-10, IL-6, and nitrate. There were8 macrophage samples from 8 mice for eachdetermination.
Fig. 6. NF-�B DNA binding activity. Analysis of macrophage protein extracts from malnourished (MN) and well-nourished (WN; control) mice for NF-�B DNAbinding activity by EMSA; macrophages were activated by IFN-�/LPS. Each lane represents the protein extracts pooled from two mice. Arrowhead: specificDNA–protein complexes. Solid circle: unincorporated labeled probe. (A) 0.5 h of culture. (B) 1 h. (C) 16 h. (D). 24 h. (E) Supershift assay showing p65p65, p50p65,and p50p50 subunits of NF-�B at 16 h after IFN-�/LPS stimulation. Arrow: unincorporated probe. (F) Densitometric analysis. The autoradiographic signalsobtained in EMSA (A�D) were quantified by video-image analysis, and the results were presented as mean � SEM of the arbitrary numbers obtained. * P 0.05.**P 0.01.
Anstead et al. Undernutrition and macrophage cytokine network 987
duce less TNF-� in response to infection or LPS [29–31].TNF-� levels were reduced in LPS-stimulated mononuclearcells from malnourished children [32]. The effect of proteinmalnutrition on IL-6 and IL-1 production in experimentalanimals has been variable [10, 30, 31, 33]. However, increasedlevels of IL-6 have been observed in multiple studies of mal-nourished patients [34–37].
The cytokine profile that observed in the stimulated macro-phages from the MN mice could be related to several pro-cesses. The diminution of TNF-� and NO, the inhibition ofNF-�B activation, and the augmentation of IL-6 also occur inendotoxin tolerance [38, 39]. The peritoneal macrophages fromthe MN mice may show endotoxin tolerance because of bacte-rial intestinal translocation, which occurs more readily in theimmunocompromised host [40]. Alternatively, the cytokineprofile displayed by the macrophages from the MN mice isanalogous to the secretory behavior of macrophages at a lowlevel of differentiation [41, 42]. Previously, malnutrition hasbeen shown to cause retarded macrophage differentiation [33].
The deficits in TNF-� expression observed in the stimulatedmacrophages from the MN mice may have significant conse-quences in host defense [43–46]. TNF-� is the apex of theproinflammatory cytokine cascade because its early productionstimulates additional proinflammatory and counter-regulatorymediators [47]. TNF-� and IL-10 form a feedback loop, inwhich TNF-� induces IL-10 secretion and IL-10 negativelyregulates TNF-� synthesis [22]. In this study, macrophagesfrom the MN mice showed low levels of both TNF-� and IL-10production. IL-10 is classically considered an anti-inflamma-
tory mediator [48, 49]. However, IL-10 displays certain proin-flammatory activities as well [45, 48, 50]. Thus, the low levelsof IL-10 produced by the macrophages from the malnourishedmice may have adverse effects on the innate immune response.Antibody neutralization of TNF-� from the macrophages fromthe WN mice decreased IL-10 levels in proportion to thedegree of neutralization, as expected based on the potent effectof TNF-� on IL-10 synthesis. However, TNF-� supplementa-tion of macrophages from the malnourished mice did not cor-rect the deficit in IL-10, suggesting that factors in addition toTNF-� deficiency play a role in the IL-10 deficit of malnutri-tion.
Although IL-6 is essential in host defense [51], the overex-pression of IL-6 observed in malnutrition may be detrimental tohost defense [52–54]. IL-6 has a number of activities that maybe immunosuppressive; it modulates IL-2 reactivity [55], in-hibits Th1 differentiation [56], and impairs early release ofTNF-� [57]. IL-6 production is inhibited by IL-10 and NO [18,58]; thus, the deficiencies of IL-10 and NO production ob-served from the macrophages from the MN mice may promoteexcessive IL-6 levels. IL-6 transcription is regulated by mul-tiple transcription factors that either activate (NF-�B) or sup-press its expression [59]. Although NF-�B binding activity wasdecreased in macrophages from the MN mice, apparently othertranscription factors stimulate IL-6 transcription. Dysregulatedproduction of IL-6 also occurs in several pathologic conditions[14, 60–62]. IL-6 production is enhanced by TNF-� [23].However, IL-6 secretion by macrophages from the MN micewas high, despite the lower levels of TNF-�. Treatment of
Fig. 7. The effect of the inhibition ofNOS2 and NO supplementation on NF-�BDNA binding activity. Macrophage proteinextracts were analyzed for NF-�B DNAbinding activity by EMSA; macrophageswere activated by IFN-�/LPS. Lane 1 isfrom macrophages from well-nourishedmice without the addition of an NOS2 in-hibitor (well-nourished control, WN-C).Lane 2 is from macrophages from well-nour-ished mice treated with the NOS2 inhibitorL-NIL (WN-L-NIL). Lane 3 is from macro-phages from malnourished mice withoutsupplemental NO (malnourished-control,MN-C). Lane 4 is from macrophages fromMN mice treated with the NO donor sodiumnitroprusside (MN-SNP). Each lane represents the pooled protein extracts of the peritoneal macrophages from 5 mice. Arrowhead: specific DNA–protein complexes.
Fig. 8. The effect of inhibition of NOS2 onthe levels of nitrate, TNF-�, IL-6, and IL-10. NOS2 was inhibited using L-N6-(1-imi-noethyl)lysine (L-NIL). Peritoneal cellsfrom the WN mice were collected by lavageand stimulated with IFN-�/LPS as de-scribed in Materials and Methods. L-NILwas added to macrophage cultures 15 minafter the addition of LPS to a concentrationof 1 mM. After 24 h, the culture superna-tants were assayed for nitrate, TNF-�, IL-6,and IL-10. The cells were analyzed for ac-tivated NF-�B (Fig. 7). Eight macrophagesamples were obtained from 8 mice in bothgroups (with and without L-NIL).
988 Journal of Leukocyte Biology Volume 74, December 2003 http://www.jleukbio.org
macrophages from the WN mice with anti-TNF-� antibodiesdid not completely suppress IL-6 production, so other factorsin addition to TNF-� must influence its expression [55].
In this study, there were decreased levels of NO produced bythe macrophages from the MN mice. NOS2 is regulated tran-scriptionally [18], and the expression of the macrophage NOS2gene is induced synergistically by IFN-� and LPS [16]. Thedecreased NOS2 transcription observed in macrophages fromthe MN mice may be due to deficits in early NF-�B activationand perhaps other defects in LPS and IFN-� signal transduc-tion. In our system, antibody neutralization of TNF-� produc-tion by macrophages from the WN mice diminished NO pro-duction, as described previously [26]. By contrast, TNF-�supplementation of the macrophages from the MN mice in-creased NO production (Fig. 5). Thus, the attenuated TNF-�production observed in macrophages from the MN animals maycontribute to the decreased NOS2 mRNA transcription and NOsynthesis.
To determine the role of NO in the aberrant cytokine re-sponses of macrophages from the MN mice, cytokine produc-tion was examined during NOS2 inhibition (Fig. 8) or NOsupplementation (Fig. 9). Blocking NO production in macro-phages from the WN mice increased IL-6 and TNF-� produc-tion [18, 63] but had no effect on IL-10 levels. Previously,inhibition of NO synthesis has been noted to increase IL-10[64].
Supplementation of stimulated macrophages from MN micewith exogenous NO could not reverse the malnutrition-relatedeffects on TNF-�, IL-6, or IL-10 (Fig. 9). Previous studiesindicate that NO may stimulate or inhibit IL-6 and IL-10production [58, 65, 66]. Based on these studies of the manip-ulation of the NO level in both the MN and WN mice, thealtered levels of TNF-� and IL-10 observed in malnutrition donot arise from feedback regulation due to the lower expressionof NO, but result from processes independent from NO pro-duction.
Because NF-�B is a critical transcription factor for genesthat control the production of IL-6, TNF-�, and NO [67], wepostulated that it could play a role in the dysregulated proin-flammatory response in the malnourished host. Previously,Yaman et al. observed that NF-�B expression was decreased inLPS-stimulated peritoneal macrophages from protein-malnour-ished rats [68].
NF-�B is the most important transcription factor for theexpression of TNF-� [69]. The delayed induction of NF-�Bobserved at 0.5 and 1 h after stimulation in the macrophagesfrom the MN mice probably contributes significantly to thedecreased levels of TNF-� and NOS2 transcription seen after4 h. In contrast, the high levels of activated NF-�B observed24 h after stimulation of the macrophages from the MN micewas due to a lack of appropriate negative feedback by NO. Theaddition of supplemental NO to the macrophage cultures fromthe MN mice decreased this late NF-�B activation, whereasinhibiting NO production in the macrophages from the WNmice increased NF-�B at 24 h, mimicking the effect of mal-nutrition.
Another possibility to explain defective TNF-� and NOS2transcription was that the NF-�B that reached the nucleus wasdysfunctional. NF-�B attaches to DNA in the promoter regionsof target genes as a dimer composed of two Rel proteins. Thep50/p65, p50/c-Rel, p65/p65, and p65/c-Rel dimers stimulateinflammatory mediator transcription, whereas the p50 ho-modimer is inhibitory [67]. To investigate the possibility thatthe NF-�B subunit composition in the macrophages from MNmice may be predominantly the inhibitory p50 homodimer, asupershift assay was performed. This revealed that the NF-�Bfrom macrophages of both WN and MN mice both had similarsubunit composition.
Because of the integrated network of cytokine regulation,dysregulated expression of even a single crucial cytokine genecan result in immunologic consequences [47]. In this model ofmalnutrition, delayed NF-�B activation and the resulting earlydeficits of TNF-� had a significant effect on subsequent NOproduction. Furthermore, there was autonomous hyper-expres-sion of IL-6, a potentially immunosuppressive mediator, whichwas disconnected from early NF-�B activation and TNF-�production. These defects in the innate immune response pro-duced by multinutrient undernutrition may contribute to thesusceptibility of the malnourished patient to infection.
ACKNOWLEDGMENTS
The authors thank Wei-guo Zhao for technical assistance andGabriel Fernandes, Sunil K. Ahuja, Seema S. Ahuja, andRobert A. Clark for helpful discussions. This work was sup-
Fig. 9. The effect of the addition of ex-ogenous NO on nitrate, TNF-�, IL-6, andIL-10. Cultures of macrophages fromthe malnourished mice were supple-mented with NO by using sodium nitro-prusside (SNP). Peritoneal cells fromthe MN mice were collected and stimu-lated with IFN-�/LPS as described inMaterials and Methods. Sodium nitro-prusside was added to macrophagecultures 15 min after the addition ofLPS to a final concentration of 0.1 mM.After 24 h, the culture supernatants wereassayed for nitrate, TNF-�, IL-6, andIL-10 and the cells were analyzed for
activated NF-�B (Fig. 7). Four samples of macrophages were obtained from four mice for each determination (with and without SNP).
Anstead et al. Undernutrition and macrophage cytokine network 989

ported by funding from the U.S. Department of Veterans Affairs(GMA, PCM), the Howard Hughes Medical Institute (GMA), aGrant-in-Aid (0150105N) from the American Heart Associa-tion (BC), and the National Institutes of Health (NIH HL-68020) (BC).
REFERENCES
1. Pelletier, D. L. (1994) The potentiating effects of malnutrition on childmortality: epidemiologic evidence and policy implications. Nutr. Rev. 52,409–415.
2. Woodward, B. (1998) Protein, calories, and immune defenses. Nutr. Rev.56, S84–S92.
3. Anstead, G. M., Chandrasekar, B., Zhao, W., Yang, J., Perez, L. E., Melby,P. C. (2001) Malnutrition alters the innate immune response and increasesearly visceralization following Leishmania donovani infection. Infect. Im-mun. 69, 4709–4718.
4. Beutler, B. A. (1999) The role of tumor necrosis factor in health anddisease. J. Rheumatol. 26, Suppl 57, 16–21.
5. Boehm, U., Klamp, T., Groot, M., Howard, J. C. (1997) Cellular responsesto interferon-gamma. Annu. Rev. Immunol. 15, 749–795.
6. Guha, M., Mackman, N. (2001) LPS induction of gene expression inhuman monocytes. Cell. Signal. 13, 85–94.
7. Dinarello, C. A., Mier, J. W. (1987) Lymphokines. N. Engl. J. Med. 317,940–945.
8. Blackwell, T. S., Christman, J. W. (1997) The role of nuclear factor-kappaB in cytokine gene regulation. Am. J. Respir. Cell Mol. Biol. 17, 3–9.
9. Chan, J., Flynn, J. 1999. Nitric Oxide in Mycobacterium Infection. InNitric Oxide and Infection. F. Fang, ed. Kluwer Academic/Plenum Pub-lishers, New York.
10. Redmond, H. P., Leon, P., Lieberman, M. D., Hofmann, K., Shou, J.,Reynolds, J. V., Goldfine, J., Johnston, R. B., Jr., Daly, J. M. (1991)Impaired macrophage function in severe protein-energy malnutrition.Arch. Surg. 126, 192–196.
11. Redmond, H. P., Shou, J., Kelly, C. J., Schreiber, S., Miller, E., Leon, P.,Daly, J. M. (1991) Immunosuppressive mechanisms in protein-caloriemalnutrition. Surgery 110, 311–317.
12. Reynolds, J. V., Redmond, H. P., Ueno, N., Steigman, C., Ziegler, M. M.,Daly, J. M., Johnston, R. B., Jr. (1992) Impairment of macrophage acti-vation and granuloma formation by protein deprivation in mice. Cell.Immunol. 139, 493–504.
13. Rivadeneira, D. E., Grobmyer, S. R., Naama, H. A., Mackrell, P. J.,Mestre, J. R., Stapleton, P. P., Daly, J. M. (2001) Malnutrition-inducedmacrophage apoptosis. Surgery 129, 617–625.
14. Chandrasekar, B., Mitchell, D. H., Colston, J. T., Freeman, G. L. (1999)Regulation of CCAAT/Enhancer binding protein, interleukin-6, interleu-kin-6 receptor, and gp130 expression during myocardial ischemia/reper-fusion. Circulation 99, 427–433.
15. Chandrasekar, B., Freeman, G. L. (1997) Induction of nuclear factorkappaB and activation protein 1 in postischemic myocardium. FEBS Lett.401, 30–34.
16. Fujihara, M., Ito, N., Pace, J. L., Watanabe, Y., Russell, S. W., Suzuki, T.(1994) Role of endogenous interferon-beta in lipopolysaccharide-triggeredactivation of the inducible nitric-oxide synthase gene in a mouse macro-phage cell line, J774. J. Biol. Chem. 269, 12773–12778.
17. Donnelly, R. P., Freeman, S. L., Hayes, M. P. (1995) Inhibition of IL-10expression by IFN-gamma up-regulates transcription of TNF-alpha inhuman monocytes. J. Immunol. 155, 1420–1427.
18. Connelly, L., Palacios-Callender, M., Ameixa, C., Moncada, S., Hobbs,A. J. (2001) Biphasic regulation of NF-kappa B activity underlies the pro-and anti-inflammatory actions of nitric oxide. J. Immunol. 166, 3873–3881.
19. Moore, W. M., Webber, R. K., Jerome, G. M., Tjoeng, F. S., Misko, T. P.,Currie, M. G. (1994) L-N6-(1-iminoethyl)lysine: a selective inhibitor ofinducible nitric oxide synthase. J. Med. Chem. 37, 3886–3888.
20. Adams, D. O., Hamilton, T. A. (1987) Molecular transductional mecha-nisms by which IFN gamma and other signals regulate macrophagedevelopment. Immunol. Rev. 97, 5–27.
21. Kawai, T., Takeuchi, O., Fujita, T., Inoue, J., Muhlradt, P. F., Sato, S.,Hoshino, K., Akira, S. (2001) Lipopolysaccharide stimulates the MyD88-independent pathway and results in activation of IFN-regulatory factor 3and the expression of a subset of lipopolysaccharide-inducible genes.J. Immunol. 167, 5887–5894.
22. Wanidworanun, C., Strober, W. (1993) Predominant role of tumor necrosisfactor-alpha in human monocyte IL-10 synthesis. J. Immunol. 151,6853–6861.
23. Tracey, K. J., Cerami, A. (1993) Tumor necrosis factor, other cytokinesand disease. Annu. Rev. Cell Biol. 9, 317–343.
24. Saura, M., Zaragoza, C., Bao, C., McMillan, A., Lowenstein, C. J. (1999)Interaction of interferon regulatory factor-1 and nuclear factor kappa Bduring activation of inducible nitric oxide synthase transcription. J. Mol.Biol. 289, 459–471.
25. Hayes, M. P., Freeman, S. L., Donnelly, R. P. (1995) IFN-gamma primingof monocytes enhances LPS-induced TNF production by augmenting bothtranscription and MRNA stability. Cytokine 7, 427–435.
26. Drapier, J. C., Wietzerbin, J., Hibbs, J. B., Jr. (1988) Interferon-gammaand tumor necrosis factor induce the L-arginine-dependent cytotoxiceffector mechanism in murine macrophages. Eur. J. Immunol. 18, 1587–1592.
27. Schaffer, M. R., Tantry, U., Ahrendt, G. M., Wasserkrug, H. L., Barbul, A.(1997) Acute protein-calorie malnutrition impairs wound healing: a pos-sible role of decreased wound nitric oxide synthesis. J. Am. Coll. Surg.184, 37–43.
28. Chan, J., Tian, Y., Tanaka, K. E., Tsang, M. S., Yu, K., Salgame, P.,Carroll, D., Kress, Y., Teitelbaum, R., Bloom, B. R. (1996) Effects ofprotein calorie malnutrition on tuberculosis in mice. Proc. Natl. Acad. Sci.USA 93, 14857–14861.
29. Dai, G., McMurray, D. N. (1998) Altered cytokine production and im-paired antimycobacterial immunity in protein-malnourished guinea pigs.Infect. Immun. 66, 3562–3568.
30. Skerrett, S. J., Henderson, W. R., Martin, T. R. (1990) Alveolar macro-phage function in rats with severe protein calorie malnutrition. Arachi-donic acid metabolism, cytokine release, and antimicrobial activity. J. Im-munol. 144, 1052–1061.
31. Ikeda, S., Saito, H., Fukatsu, K., Inoue, T., Han, I., Furukawa, S.,Matsuda, T., Hidemura, A. (2001) Dietary restriction impairs neutrophilexudation by reducing CD11b/CD18 expression and chemokine produc-tion. Arch. Surg. 136, 297–304.
32. Doherty, J. F., Golden, M. H., Remick, D. G., Griffin, G. E. (1994)Production of interleukin-6 and tumour necrosis factor-alpha in vitro isreduced in whole blood of severely malnourished children. Clin. Sci.(Lond.) 86, 347–351.
33. Honda, Y., Takahashi, K., Naito, M., Fujiyama, S. (1995) The role ofmacrophage colony-stimulating factor in the differentiation and prolifera-tion of Kupffer cells in the liver of protein-deprived mice. Lab. Invest. 72,696–706.
34. Johann-Liang, R., O'Neill, L., Cervia, J., Haller, I., Giunta, Y., Licholai,T., Noel, G. J. (2000) Energy balance, viral burden, insulin-like growthfactor-1, interleukin-6 and growth impairment in children infected withhuman immunodeficiency virus. AIDS 14, 683–690.
35. de Martino, M., Galli, L., Chiarelli, F., Verrotti, A., Rossi, M. E., Bindi, G.,Galluzzi, F., Salti, R., Vierucci, A. (2000) Interleukin-6 release by cul-tured peripheral blood mononuclear cells inversely correlates with heightvelocity, bone age, insulin-like growth factor-I, and insulin-like growthfactor binding protein-3 serum levels in children with perinatal HIV-1infection. Clin. Immunol. 94, 212–218.
36. Sauerwein, R. W., Mulder, J. A., Mulder, L., Lowe, B., Peshu, N.,Demacker, P. N., van der Meer, J. W., Marsh, K. (1997) Inflammatorymediators in children with protein-energy malnutrition. Am. J. Clin. Nutr.65, 1534–1539.
37. Malave, I., Vethencourt, M. A., Chacon, R., Quinones, D., Rebrij, C.,Bolivar, G. (1998) Production of interleukin-6 in cultures of peripheralblood mononuclear cells from children with primary protein-calorie mal-nutrition and from eutrophic controls. Ann. Nutr. Metab. 42, 266–273.
38. Takasuka, N., Tokunaga, T., Akagawa, K. S. (1991) Preexposure of mac-rophages to low doses of lipopolysaccharide inhibits the expression oftumor necrosis factor-alpha mRNA but not of IL-1 beta mRNA. J. Immu-nol. 146, 3824–3830.
39. Takasuka, N., Matsuura, K., Yamamoto, S., Akagawa, K. S. (1995) Sup-pression of TNF-alpha mRNA expression in LPS-primed macrophagesoccurs at the level of nuclear factor-kappa B activation, but not at the levelof protein kinase C or CD14 expression. J. Immunol. 154, 4803–4812.
40. Shou, J., Redmond, H. P., Leon, P., Hofmann, K. P., Daly, J. M. (1991)Elemental diet alters macrophage function in mice. J. Surg. Res. 51,192–196.
41. Scheibenbogen, C., Andreesen, R. (1991) Developmental regulation of thecytokine repertoire in human macrophages: IL-1, IL-6, TNF-alpha, andM-CSF. J. Leukoc. Biol. 50, 35–42.
42. Takashiba, S., Van Dyke, T. E., Amar, S., Murayama, Y., Soskolne, A. W.,Shapira, L. (1999) Differentiation of monocytes to macrophages primes
990 Journal of Leukocyte Biology Volume 74, December 2003 http://www.jleukbio.org

cells for lipopolysaccharide stimulation via accumulation of cytoplasmicnuclear factor kappaB. Infect. Immun. 67, 5573–5578.
43. Green, S. J., Nacy, C. A., Meltzer, M. S. (1991) Cytokine-induced syn-thesis of nitrogen oxides in macrophages: a protective host response toLeishmania and other intracellular pathogens. J. Leukoc. Biol. 50, 93–108.
44. Herring, A. C., Lee, J., McDonald, R. A., Toews, G. B., Huffnagle, G. B.(2002) Induction of interleukin-12 and gamma interferon requires tumornecrosis factor alpha for protective T-cell-mediated immunity to pulmo-nary Cryptococcus neoformans infection. Infect. Immun. 70, 2959–2964.
45. Appelberg, R. (1994) Protective role of interferon gamma, tumor necrosisfactor alpha and interleukin-6 in Mycobacterium tuberculosis and M.avium infections. Immunobiology 191, 520–525.
46. Yap, G. S., Scharton-Kersten, T., Charest, H., Sher, A. (1998) Decreasedresistance of TNF receptor p55- and p75-deficient mice to chronic toxo-plasmosis despite normal activation of inducible nitric oxide synthase invivo. J. Immunol. 160, 1340–1345.
47. Brennan, F. M., Feldmann, M. 1999. Cytokine networks. In The CytokineNetworks. Balkwill, F., ed. Oxford University Press, Oxford.
48. Xu, K., Yen, T., Geczy, C. L. (2001) IL-10 up-regulates macrophageexpression of the S100 protein S100A8. J. Immunol. 166, 6358–6366.
49. Thivierge, M., Parent, J. L., Stankova, J., Rola-Pleszczynski, M. (1999)Modulation of formyl peptide receptor expression by IL-10 in humanmonocytes and neutrophils. J. Immunol. 162, 3590–3595.
50. Zidek, Z., Frankova, D. (1999) Interleukin-10 in combination with inter-feron-gamma and tumor necrosis factor-alpha enhances in vitro productionof nitric oxide by murine resident peritoneal macrophage. Eur. CytokineNetw. 10, 25–32.
51. van der Poll, T., Keogh, C. V., Guirao, X., Buurman, W. A., Kopf, M.,Lowry, S. F. (1997) Interleukin-6 gene-deficient mice show impaireddefense against pneumococcal pneumonia. J. Infect. Dis. 176, 439–444.
52. Suzuki, Y., Rani, S., Liesenfeld, O., Kojima, T., Lim, S., Nguyen, T. A.,Dalrymple, S. A., Murray, R., Remington, J. S. (1997) Impaired resistanceto the development of toxoplasmic encephalitis in interleukin-6-deficientmice. Infect. Immun. 65, 2339–2345.
53. Tilg, H., Dinarello, C. A., Mier, J. W. (1997) IL-6 and APPs: anti-inflammatory and immunosuppressive mediators. Immunol. Today 18,428–432.
54. Hatzigeorgiou, D. E., He, S., Sobel, J., Grabstein, K. H., Hafner, A., Ho,J. L. (1993) IL-6 down-modulates the cytokine-enhanced antileishmanialactivity in human macrophages. J. Immunol. 151, 3682–3692.
55. Holzheimer, R. G., Molloy, R., Mendez, M. V., O'Riordain, D., Curley, P.,Nestor, M., Collins, K., Saproschetz, I., Mannick, J. A., Rodrick, M. L.(1995) Multiple system organ failure may be influenced by macrophagehypoactivation as well as hyperactivation–importance of the double chal-lenge. Eur. J. Surg. 161, 795–803.
56. Diehl, S., Anguita, J., Hoffmeyer, A., Zapton, T., Ihle, J. N., Fikrig, E.,Rincon, M. (2000) Inhibition of Th1 differentiation by IL-6 is mediated bySOCS1. Immunity 13, 805–815.
57. Tanaka, T., Kanda, T., McManus, B. M., Kanai, H., Akiyama, H., Sekigu-chi, K., Yokoyama, T., Kurabayashi, M. (2001) Overexpression of inter-leukin-6 aggravates viral myocarditis: impaired increase in tumor necrosisfactor-alpha. J. Mol. Cell. Cardiol. 33, 1627–1635.
58. Stadler, J., Harbrecht, B. G., Di Silvio, M., Curran, R. D., Jordan, M. L.,Simmons, R. L., Billiar, T. R. (1993) Endogenous nitric oxide inhibits thesynthesis of cyclooxygenase products and interleukin-6 by rat Kupffercells. J. Leukoc. Biol. 53, 165–172.
59. Sanceau, J., Kaisho, T., Hirano, T., Wietzerbin, J. (1995) Triggering of thehuman interleukin-6 gene by interferon-gamma and tumor necrosis factor-alpha in monocytic cells involves cooperation between interferon regula-tory factor-1, NF kappa B, and Sp1 transcription factors. J. Biol. Chem.270, 27920–27931.
60. Chouaib, S., Asselin-Paturel, C., Mami-Chouaib, F., Caignard, A., Blay,J. Y. (1997) The host-tumor immune conflict: from immunosuppression toresistance and destruction. Immunol. Today 18, 493–497.
61. Libermann, T. A., Baltimore, D. (1990) Activation of interleukin-6 geneexpression through the NF-kappa B transcription factor. Mol. Cell. Biol.10, 2327–2334.
62. Daynes, R. A., Araneo, B. A., Ershler, W. B., Maloney, C., Li, G. Z., Ryu,S. Y. (1993) Altered regulation of IL-6 production with normal aging.Possible linkage to the age-associated decline in dehydroepiandrosteroneand its sulfated derivative. J. Immunol. 150, 5219–5230.
63. Borges, M. M., Kloetzel, J. K., Andrade, H. F., Jr., Tadokoro, C. E.,Pinge-Filho, P., Abrahamsohn, I. (1998) Prostaglandin and nitric oxideregulate TNF-alpha production during Trypanosoma cruzi infection. Im-munol. Lett. 63, 1–8.
64. Hogaboam, C. M., Steinhauser, M. L., Schock, H., Lukacs, N., Strieter,R. M., Standiford, T., Kunkel, S. L. (1998) Therapeutic effects of nitricoxide inhibition during experimental fecal peritonitis: role of interleu-kin-10 and monocyte chemoattractant protein 1. Infect. Immun. 66,650–655.
65. Marcinkiewicz, J., Grabowska, A., Chain, B. (1995) Nitric oxide up-regulates the release of inflammatory mediators by mouse macrophages.Eur. J. Immunol. 25, 947–951.
66. Kallmann, B. A., Malzkorn, R., Kolb, H. (1999) Exogenous nitric oxidemodulates cytokine production in human leukocytes. Life Sci. 65, 1787–1794.
67. Ghosh, S., May, M. J., Kopp, E. B. (1998) NF-kappa B and Rel proteins:evolutionarily conserved mediators of immune responses. Annu. Rev.Immunol. 16, 225–260.
68. Yaman, M., Fogler, R., Abadir, A. R., Gintautas, J. (1999) Protein caloriemalnutrition induces decreased macrophage NF-kappa B expression.Proc. West. Pharmacol. Soc. 42, 97–98.
69. Jongneel, C. V. 1992. The TNF and lymphotoxin promoters. In Tumornecrosis factors. the molecules and their emerging role in medicine. B. A.Beutler, ed. Raven Press, New York.
Anstead et al. Undernutrition and macrophage cytokine network 991