molecular and comparative analysis of the hyperthermostable pyrococcus furiosus glutamate...
TRANSCRIPT

Biocata/rsis, 1994, Vol. 11, pp. 131—141 © 1994 Harwood Academic Publishets GmbHReprints availabte directly from the publisher Printed in MalavsiaPhotocopying permitted by license only
MOLECULAR AND COMPARATIVE ANALYSISOF THE HYPERTHERMOSTABLE PYROCOCCUS
FURIOSUS GLUTAMATE DEHYDROGENASE ANDITS GENE
RIK 1. L. EGGEN*, ANS C. M. GEERLING,WILFRIED G. B. VOORHORST, REMCO KORT and WILLEM M. de VOS
Departmen t of Microbiology, Wagen ingen Agrictiltural University, Hesselink vanSuchtelen weg 4, 6703 CT Wageningen, The Netherlands
The gene for glutamate dehydrogenase (GDH) from Pi’rococcusfuriostis has been cloned, sequencedand expressed in Escherichia coti. Significant GDH activity could be detectcd in this host, allowingthe furthcr structure-function analysis of this hyperthermostable hexameric enzyrne. The deduccdprimary sequence of the P.furiosus GDH was homotogous to various bacteriat, archacal and eukaryalGDHs. Detailed comparativc analysis of the primary sequences of these GDHs suggest that a decreascin Gly residues can be a general stabilizing feature of proteins functional under extreme conditionssuch as high temperatures or high salt concentrations whereas an increasc in 11e residucs and adecrease in Cys residues is typical for hyperthermostable enzymcs.
KEY WORDS Archaea, Hyperthermophile
INTRODUCTION
Recent years have shown a rapidty expanding interest in the hyperthermophulicprokaryotes, that are able to grow at or above the boiling point of water. Mostinterest is focused on understanding the adaptation mechanisms that a]lowmetabolism to function and biomolecules, such as DNA, proteins or membranes,to remain intact at such extreme temperatures. In addition, enzymes fromhyperthermophiles might be exploited in industrial applications, since theseenzymes are thermoactive and show a high thermal and chemical stability.
In the research aimed at understanding the hyperthermostability and hyperthermoactivity of organisms and their biomolecules, continuous efforts areundertaken to isolate and identify new hyperthermophulic species (Stetter er al.,1990). Up to now, these species have all been phylogenetically grouped withinthe archaeal domain (Woese et al., 1990). from the hyperthermophulic organisms, two very similar species of the genus Pyrococeus, P. furiosus (Fiata andStetter, 1986) and P. ivoesei (Zillig et al., 1987) have been used as models fordetailed biochemical and molecular biological studies on hyperthermophily. This
* To whom correspondcncc should be addressed.Present address: Swiss Fcderal Institute for Environmental Science and Technology (EAWAG).CH-$600, Dübendorf, Switzerland.
131
Kopie für Lizenzkunden von subito eV., geliefert und ausgedruckt für TNO (SLIO6XOO2O5E)

132 RIKI.L. EGGEN ETAL
work bas led to the isolation and characterization of their hydrolytic enzymes(Constantino et al., 1990; Koch et al., 1991; Kengen et aL, 1993) and enzymesin the central metabolism (Bryant and Adams, 1989; Mukund and Adams, 1991;Consatvi ei al., 1991; Robb et al., 1 992), which proved to be highly thermostable.In addition, remarkable adaptive metabolic features to resist high temperatureshave been described in Pyrococci such as a non-phosphorylated EntnerDouderoif route (Mukund and Adams, 1991; Schafer and Schönheit, 1991, 1992,1993), the use of a hyperthermostable ferredoxin as electron carrier (Aono et al,1989) and the presence of high guantities of phospho-inositol-phosphate asstabilizing agent (Scholz et al., 1992). The understanding of structural adaptations of hyperthermostable enzymes requires detailed structure-stability analysesthat can be greatly facilitated by protein engineering. For this approach it isessential to perform genetic studies aimed at the characterization and functionalexpression of the gene of interest. The first gene of an archaeal hyperthermophileto be isolated was the P. woesci glyceraldehyde-3-phosphate dehydrogenase gene(GAPDH, Zwicki et al., 1990). Upon expression of this gene in Escherictzia coli,activity could be measured at high temperatures. Currently, primary sequencesderived from cloned genes are available for the enzymes DNA polymerase(PFUPOL, Uemori et al., 1993), glutamine synthetase (GS, Tiboni et al., 1993)and glutamate dehydrogenase (GDH, Eggen et al., 1993).
To further understand the functioning of enzymes at high temperatures, wehave focused our research on GDH as a model enzyme since GDH from otherorganisms is a weil-studied enzyme (Smith et al., 1975), GDH is highly conservedin evolution (Benachenhou-Lahfa et al., 1993), primary sequences are availableof GDHs with different thermostabilities and thermoactivities (BenachenhouLahfa ei al., 1993) and a 3-dimensional structure of the Ctostridium svmbiosumGDH has been solved (Baker et al., 1992). In P. furiosus, GDH represents up to20% of the total protein (Consalvi et al., 1991; Robb et al., 1992). It was recentlyshown in this departrnent that GDH plays an important role in the centralmetabolism of P. furiosus and is involved in the production of alanine whichconstitutes a sink for reducing equivalents (Kengen and Stams, 1994).
In this report we will discuss the comparative analysis of the primary sequencesof functionally homologous GDHs from f. furiosus and other organisms. Trendsin amino acid changes, possibly related with hyperthermostability, will bediscussed.
METHODS
Bacterial strains and growth conditions. F. furiosus (DSM 3638) was cultured at98C in a synthetic seawater medium as described before (Eggen ei al., 1990).Escherichia coti TG1 [F’(traD35 proAB 1acZAM]5) supE hsdA59 lacproAB (Gibson, 1984)], and E. coli K12zHlttrp (Remaut et al., 1981) cultivatedin LB-based medium were used for routine cloning and expression experiments.For complementation experiments, two E. coli strains that were auxotrophic forglutamate were used; strain PA340 [thr-1 leuB6 gdh-1 hisGi g1t331 argHl thi-]ara-14 lacYl gal-6 malAl xvl-7 mtt-2 rpsL9 to A2L supE44 (Berberich, 1972)],kindly provided by 8. Bachmann(Department of Biology, Yale University, NewHaven) and the restriction-deficient host Q100 [thr-] leuB6 gdh-1 hisG] gltB3l
Kopie für Lizenzkunden von subito e.V., geliefert und ausgedruckt für TNO (SLIO6XOO2O5E)

PYROCOCCL’S FLR lOSt’S GDH 133
argHl thi-] ara-14 /acYI gat-6 malAl xi’t-7 rntl-2 rpsL9 tonA2).’)J supË’44 hsUR2(Snedecor eI al., 1991)], kindly supplied by 13. Snedecor (Gcnentech, mc., SanFrancisco). Both auxotrophic mutants were cultivated in M9 minimal medium(Sambrook et al., 1991), suppiemented with 0.4% glucose, 1 mM threonine,arginine, leucine, histidine and thiamine.
Enzymes and chemicals. All restriction enzymes and DNA modifying enzymeswere purchased from Life Technologies mc. (Gaithersburg, Md.), and PharmaciaLKB Biotechnologies (Uppsala, Sweden). Biochemicals were obtained fromBoehringer (Mannheim, Germany). Hybond N and [-32P]dATP (110 TBq/mmoÏ) were from Amersham.
DNA isolation, manipulation and sequences analysis. Chromosomal DNA wasisolated as described before (Eggen et al., 1991). Plasmid DNA isolations. DNAligations, transformations and other DNA maniputations were done by established procedures (Sambrook et al., 1991). Plasmids pUCI$ or pUCI9 (YanischPerron et al., 1985) and pPLC2833, a derivative of pPLC28 (Remaut et aL,1981). and bacteriophages M13mp18 or M13mp19 (Yanisch-Pecron et al., 1985)were used for cloning, expression and sequencing experiments. Computeranalysis of the sequence was done with the PC/GENE program version 5.01(Genofit, Geneva, Switzerland) and the GCG package version 7.0 (Devereuxet al., 1984) at the CAOS/CAMM Center, University of Nijmegen, TheNetherlands.
RESULTS AND DISCUSSION
Expression and codon usage of the P. furiosus gdh gene
Using a PCR protocol, with oligonucleotides designed from the NH,-terminalamino acid sequence of purified GDH, the gdh gene from P. utiosus has beenidentified, cloned and sequenced (Eggen eI al., 1993). The nucleotide sequence ofthe gdh gene revealed a gene of 1260 base pairs coding for a product of 420residues (Fig. 1). Cloning and expression of the P. furiosus gdh gene in Escheriehiacoti was demonstrated by the following observations: (i) The NH2-terminal aminoacid sequence of the purified pyrococcal GDH was identical to that found in thededuced amino acid sequence (Fig. 1); (ii) Antiserum raised against the purifiedGDH reacted with GDH, produced in E. coli from the cloned P. furiosus gdhgene; (iii) The deduced primary sequence of the F. furioszis GDH was very similar
1 MVEQDPYEIV IKQLERMQY MEI$EEALEF LKRPQRIVEV TIPVEM000S VKVFTGFRVQ
61 HNWARGPTKG GIRWHPEETL STVKALMWM TWKTAVMDLP YGGGKGGIIV DPKKLSDREK
121 ERLARGYIRA IYDVISPYED IPAPDVYTNP QIMAWMMDEY ETISRRKTPA FGIITGKPLS
181 IGGSLGRIEA TARGASYTIR EMKVLGWDT LKGKTIAIQG YGNAGYVLAK IMSEDFGMKV
241 VAVSDSKGGI YNPDGLNADE VLKWKNEHGS VKDFPGATNI TKEELLELEV DVLAPMIEE
301 VITKKNADNI KAKIVAEVAN GPVTPEADEI LFEKGILQIP DFLCNAGGVT VSYFEWVQNI
361 TGYYWTIEEV RERLDKKMTK AFYDVYNIAK EKNIHMRDM YVVAVQRVYQ AMLDRGNKH
figure 1 Deduced amino acid sequence of the P. furiosus GDH in the universal single letter code.Amino acid residucs corresponding with the NH,-tcrrninal residues of the purificd enzyme areunderlined.
Kopie kit Lizenzkunden von subito e.V., geliefert und ausgedruckt für TNO (SL106X00205E)

134 REK 1. L. EGGEN ET AL
(>35%) to GDH sequences from other sources. However, although GDH wasproduced in E. coti, no significant actÏvity at 37°C or 90°C could be detected,possibly as a result of a low-level expression (less than 0.5% of total proteincontent). As an altemative procedure for the identification of GDH activity weselected for complementation of E. coli gdh mutants PA340 or Ql00. AgainGDH was produced in these hosts but no complementation of the glutarnateauxotrophy was detectable in both mutant strains expressing the P. furiosus gdhge ne.
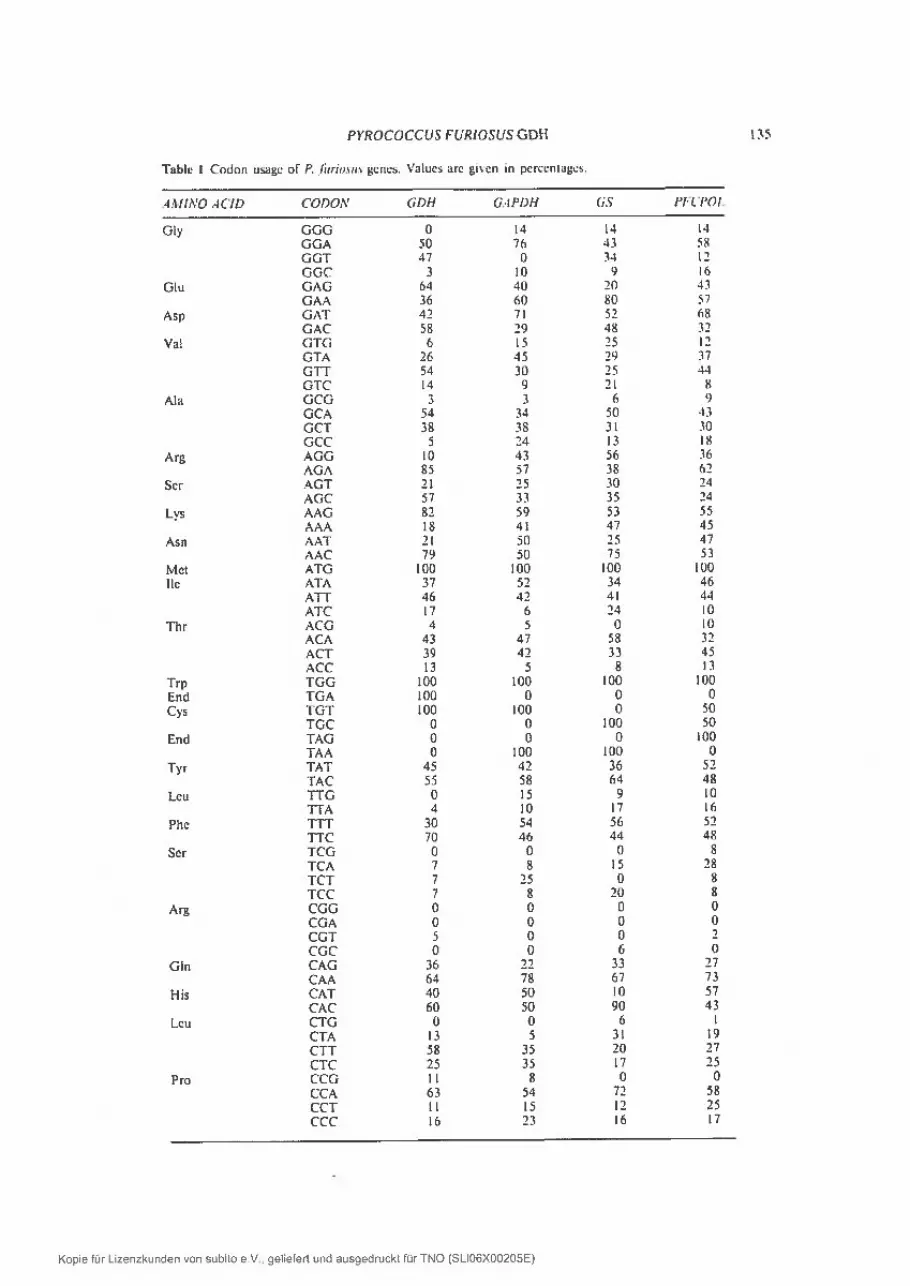
Analysis of the codon usage of the gdh gene in comparison with codon usagesof currently described P. furiosus and P. woesei genes (Table 1) shows that moreA or T residues are present in the wobble position than might be expected fromthe G + C content of the genome (38%; Fiala and Stetter, 1986). In the codons forserine, there is a strong discrimination against the TC dinucleotide. In addition,as was described before for the GAPDH gene by Zwicki eI al. (1990), a dear biasagainst CG dinucleotides is observed in the genes described so far. This isreflected in the preferentially used codons for alanine, arginine, proline andthreonine. In E. coli however, CG codons are frequently used (Gouy and Gautier,1982), which could result in a reduced expression of genes from hyperthermophilic organisms in this host.
Since GDH is an enzyme with an hexameric structure, formation of thisstructure might be dependent on the concentration of monomers. Therefore, wetried to increase the expression level of GDH in E. coli. Using an expressionsystem under control of the lambda L promoter (Fig. 2), we recently succeededin obtaining significant expression of the P. fliriosus gdh gene in E. coti up to 5%of the total protein content (Lebbink et aÏ., manuscript in preparation). Inextracts from these E. coli ceils, GDH activity at 90°C was observed. Preliminarydata indicate that GDH from P. furiosus and GDH produced in E. coti have asimilar pH optimum and similar activities at high temperatures. These resuitscreate the possibility to study the effect of knowledge-based modelling on enzymeactivity and stability.
Analysis f the predieted arnin.o acid sequence of the F. fitriosus GDH
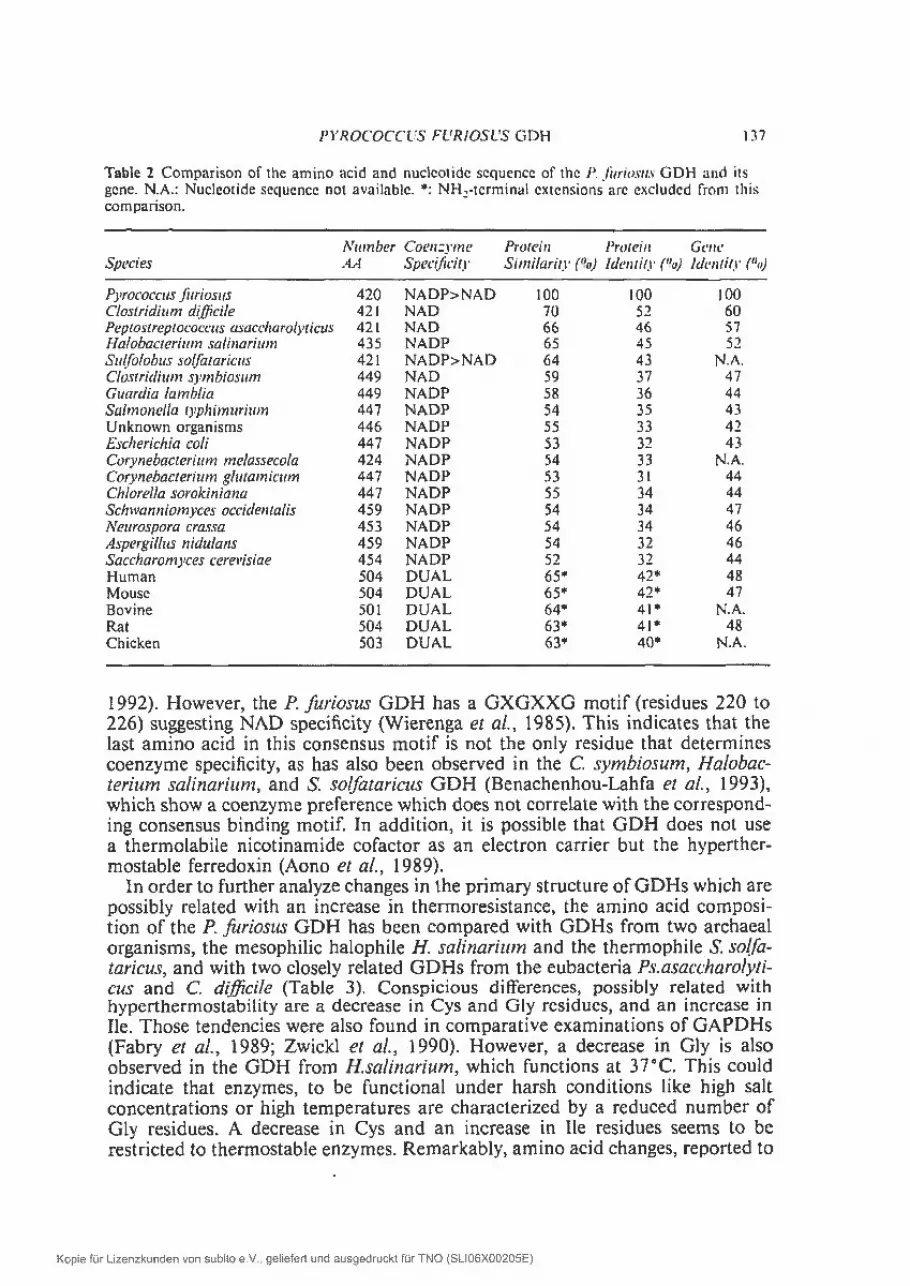
The deduced amino acid sequence has been used to examine the amino acidcomposition of the F. ftriosus GDH and to search for amino acid sequencespossibly related with hyperthermostability of the GDH (Consalvi et al., Robbet al., 1992; Eggen et aL, 1993). Computer comparisons of the deduced aminoacid sequence of the P. furiosus GDH with the SwissProt data base (release 21)revealed dear similarities with GDHs from a variety of organisms (Table 2,Benachenhou-Lahfa ei al., 1993). In addition, similarities were observed witham ino acid sequences of phenylalanine dehydrogenases from Thermoactinomveesintermedius (27,4% identity in 179 residues; Takada et al., 1991) and Baciltussphaeticus (27. 1% identity in 266 residues; Okazaki et aL, 1988) and with leucinedehydrogenase (27.1% identity in 262 residues) from Bacithts stearothermophitus(Nagata et al., 1988). It is remarkable that the F. furiosus GDH has the highestoverall identity with two eubacterial sequences, CÏostrïdium dijjciÏe (52%) andPeptostreptococcus asaccharolvtieus (46%); two archaeal sequences, Hatobacteritun saÏinariuni (45tYo) and Stilfolobus so/fataricus (43%); and witli vertebrateGDHs (40—42%). The similarity with a second group, composed of enzymes from
Kopie für Lizenzkunden von subito e.V., geliefert und ausgedruckt für TNO (SLIO6XOO2O5E)
PYROCOCCUS FURIOSUS GDH
Tablc t Codon usuge of P. fiiriou gdnes. Valucs are gt en rn percentages.
.-1.iIliVO .1CJD CODON GDH GiPDH (IS Pit POl.
Gly GGG 0 14 14 14GGA 50 76 43 58OGT 47 0 34 12GGC 3 10 9 16
Glu GAG 64 40 20 43GAA 36 60 80 57
Asp GAT 42 71 52 68GAC 58 29 48 32
Val GTG 6 15 25 12GTA 26 45 29 37GIT 54 30 25 44GTC 14 9 21 8
Ma GCG 3 3 6 9GCA 54 34 50 43GCT 38 38 31 30GCC 5 24 13 1%
Arg AGG 10 43 56 36AGA 85 57 38 62
Ser AGT 21 25 30 24AGC 57 33 35 24
Lys AAG 82 59 53 55AAA 18 41 47 45
Asn AAT 21 50 25 47AAC 79 50 75 53
Met ATG 100 100 100 100Iie ATA 37 52 34 46
ATT 46 42 41 44ATC 17 6 24 10
Thr ACG 4 5 0 10ACA 43 47 58 32ACT 39 42 33 45ACC 13 5 8 13
Trp TGG 100 100 100 100End TGA 100 0 0 0Cys IGT 100 100 0 50
TGC 0 0 100 50End lAG 0 0 0 100
TAA 0 100 100 0Tyr TAT 45 42 36 52
TAC 55 58 64 48Leu TTG 0 15 9 10
TTA 4 10 17 16Phc TTT 30 54 56 52
T[C 70 46 44 4$
Ser TCG 0 0 t) $TCA 7 8 is 28TCT 7 25 0 $TCC 7 8 20 8
Arg CGG 0 0 0 0CGA 0 0 0 0CGT 5 0 0 2CGC 0 0 6 0
Gin CAG 36 22 33 27CAA 64 78 67 73
His CAT 40 50 10 57CAC 60 50 90 43
Lcu CTG 0 0 6 1CIA 13 5 31 19CTT 58 35 20 27CIC 25 35 17 25
Pro CCG II 8 0 0CCA 63 54 72 58CCI 11 IS 12 25CCC 16 23 16 17
Kopie für Lizenzkunden von subito eV., geliefert und ausgedruckt für TNO (SLIO6XOO2O5E)

136 PiK 1. L. EGGEN ETAL
eubacterial and lower eukaryal origin is sliglitly less (32—34% identity). On thegene level, the similarity of the P. furiosus gdh gene with other gdh genes isremarkably high as well, varying between 42 and 60% (Table 2). These similarityvalues show that GDH is a weli-conserved enzyme and hence is a good modelenzyrne to study evolutionary and structure-function relationships. Based upondeduced prirnary GDH sequences, Benachenhou-Lahfa et al. (1993) haverecently described a phylogenetic tree which did not correspond with theuniversal tree of life based on 16S rRNA sequences (Woese et al., 1990; Lake,
1 99 1). It was proposed that there could exist two families of GDHs, each ofwhich is characterized by specific sequence motifs. Our observation that the F.Iiirjosus GDH has a higher similarity with two GDHs from bacterial organismsthan with two archaeal GDHs substantiates this tree.
The GDH sequence of P, furiostis was aligned with amino acid sequences ofother GDHs using the clustal V multiple sequence alignment method (Devereuxet al., 1984) and some modifications by hand (Fig. 3). Inspection of the alignedsequences revealed that 34 amino acids (out of 420 residues) of the P. furiosusGDH are conserved in all sequences. These inciude 1 3 glycines, despite thedrastic reduction of glycine residues in this enzyme (Table 3). The conservedamino acids are clustered within regions of the GDH that are involved insubstrate binding (residues 50 to 145), containing the active-site lysine and thesite for coenzyme binding (residues 200 to 335). Within the latter domain wehave searched the R fitriosus GDH for the presence of the coenzyme bindingmotif for NADP (GXGXXA, Wierenga et al., 1985) that should correlate withthe observed preference for this coenzyme (Consalvi et aL, 1991; Robb et al.,
pPLc28332.skb
Figure 2 Schematic presentation of the pPLc2833bascd construction, used to express the P.furiositsgdh gene (open arrow) in Escherichia coil, under control of the L promoter. Location and orientationof the P1 promoter, ampicillin resistance gene (ApR) and origin of replication (ori) are indicated withan arrow. mcs: multipic cloning site used to introduce the gdh gcne containing HindIII-HindIIfragment.
Kopie für Lizenzkunden von subito eV., geliefert und ausgedruckt für TNO (SLIO6XOO2O5E)
PYROCOCCL!S ELRIOSUS cDH 13?
Table 2 Comparison of the amino acid and nuclcoiide sequence of the P. hiriosu. GDH and itsgene. N.A.: Nucleotide sequence not available. : NH2-tcrminal cxtcnsions are excluded from thiscomparison.
ATU?fll)L,r (‘oen: me Protein PrO (Ciii Gen t’
Species AA Speelficitr SinillariR () Identity (°o, ldcntiti (
Pyrococcus/iiriosus 420 NADP>NAD 100 100 lOOC’tostridium diflicite 421 NAD 70 52 60Pepiostreptococcus asaceharolyticos 42 t NAD 66 46 57Halobacterium sallnariu,n 435 NADP 65 45 52Sulfolobus sotfataricus 421 NADP>NAD 64 43 NA.Ctostridium srmbiosum 449 NAD 59 37 47Guardia lambila 449 NADP 58 36 44Salmonella typhirnurium 447 NADP 54 35 43Unknown ofganisms 446 NADP 55 33 42Escherichia coli 447 NADP 53 32 43Corynebacterium metassecota 424 NAD? 54 33 N.A.Cor nehaclerium glutamicitin 447 NADP 53 3 1 44Ch/ore/la sorokiniana 447 NADP 55 34 44Schwanniomyces occiden la/is 459 NAD? 54 34 47Nertrospo;a crassa 453 NADP 54 34 46Aspergillus nidutans 459 NADP 54 32 46Saccharomyces cerevîsiae 454 NADP 52 32 44Human 504 DUAL 65* 42* 48Mouse 504 DUAL 65* 42* 47Bovine 501 DUAL 64* 41* N.A.Rat 504 DLIAL 63* 4J* 48Chicken 503 DUAL 63* 4Q* N.A.
1992). However, the P. furiosus GDH bas a GXGXXG motif (residues 220 to226) suggesting NAD specificity (Wierenga et al., 1985). This indicates that thelast amino acid in this consensus motif is not the only residue that determinescoenzyme specificity, as bas also been observed in the C. symbiosurn, Halobacteriurn salinatium, and S. solfataricus GDH (Benachenhou-Lahfa et al., 1 993),which show a coenzyme preference which does not correlate with the corresponding consensus binding motif. In addition, it is possible that GDH does not usea thermotabile nicotinamide cofactor as an electron carrier but the hyperthermostable ferredoxin (Aono et al., 1 989).
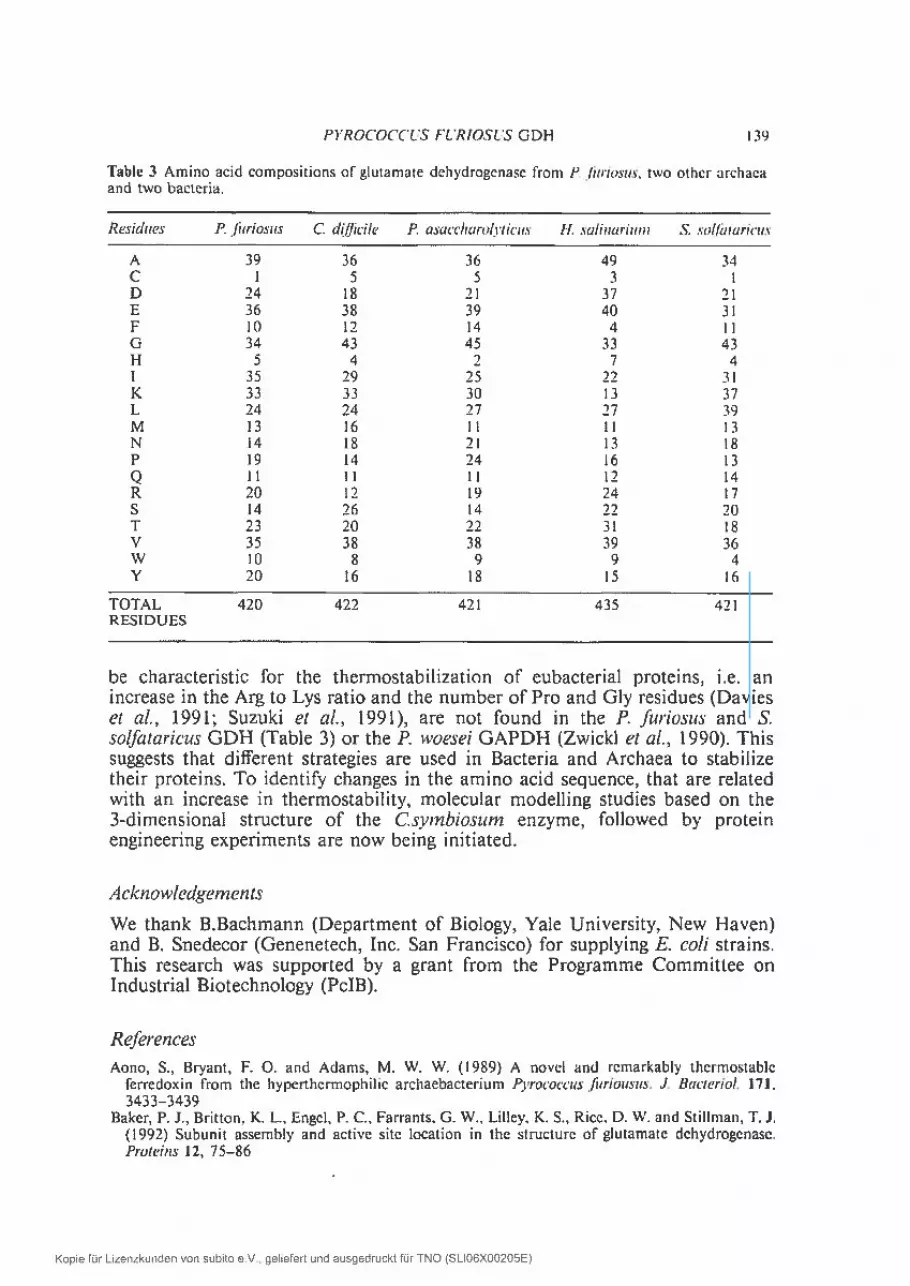
In order to further analyze changes in the primary structure of GDHs which arepossibly related with an increase in thermoresistance, the amino acid composition of the P. furiosus GDH has been compared with GDHs from two archaealorganisms, the mesophilic halophile H. satinariu,n and the thermophile S. solfataricus, and with two c]osely related GDHs from the eubacteria Rv. asaccharolyticus and C. dtfflci/e (Table 3). Conspicious ditTerences, possibly retated withhyperthermostability are a decrease in Cys and Gly residues, and an increase inIie. Those tendencies were also found in comparative examinations of GAPDHs(Fabry et al., 1989; Zwicki et aL, 1990). However, a decrease in Gly is alsoobserved in the GDH from H.saÏinariurn, which functions at 37°C. This couldindicate that enzymes, to be functional under harsh conditions like high saltconcentrations or high temperatures are characterized by a reduced number ofGly residues. A decrease in Cys and an increase in Iie residues seems to berestricted to thermostable enzymes. Remarkably, amino acid changes, reported to
Kopie für Lizenzkunden von subito e.V., geliefert und ausgedruckt für TNO fSLJO6XOO2O5E)

138 lUK 1. L. EGGEN EI AL.
, . , . , . .B
10 O 30 40 5Q 60 7V 80 CP.fur -
c. d ‘t -
— -
u. 1-—————
vripS .sa
—c . i,—-4;. 1.-S . typ -
1-€ .co1
•I;o. T•C31lJ
—Chi $01•Sch cc
—H cr1AndSL .c.r
-HMnusBov JreRtCckh
. 0, a B. c. 0, a, O °.
100 HO 120 130 t40 150 160 7O 180 19 200 210 fl0 23P. tr LPr6 Gil VPXL5OREXEL GYIk4 1.DVYT1PQ GIlC.dif
U. ial LPFGGXGVAVNPELSPEEKEPLTRRFTQEIRDV_I NÇ1PAPOP GRSYAIS. iol IPPO SO .0EY1KETGXSF.MFTGP.V GG1G1jpLY5TGLGVATtM GVEEPRYI IFGWVSYYAG(FC5c. y. Lp soçopxsoREvbrcG*ç’rELTRN-S. IS’Stp
E colC.
pt5 tFGH 544LTG5Z5 -GE - lSGII5VEkA£0. gb CP tO0 KGGSGFOPXGK!CLE1I€FCQSFKIELII1HCh t .sor LP 60006SSFDPKGKS FCQSFHIECIO1H-- IS V30VP OI000M1EIOYCFG 01 T__-OVLtCG-Q S ATGYGA 040 GES- .LKOKRCCVSGMSVAQYCAOLCCESch .scc 0---GVLTGkG- S100PGAT tOKE .FKQflVECSGSG40AYMtKYIE0.cr LS)cGg AoFopgoI(soAuRRrc U4(t4--F.SId5 .cerlj.,,anthuh.8o.v 1ntot VPF00AG(IUPGhYTDNECEKICI, cken
0, a,, 8, o, 8 0•
240 250 200 270 200 203 300 310 320 330 340P.rurC dit IVLTPDILINp INLTPD ILTNII. sol t_GATIVAtS0__VNA._MYEPDGIDTAS0PSiLEt_PEAVTT0t YIS_HEESLCTL__0000LIPAA OIIVIOYEN GAlM 40StlA000 SAOIPO IL.Sh5. so $.GM1V50--—S6GG-—0t4EXG1DVCAIE tKE010SVINYPE .AOVPOILANC. sy. £i(PiGQ—-iWOt CATQt3YDt.EQM (CV EPM -fti30 PO40YAPSI(,OVhS. Ie. p_svYI, OP ECY EO CII CAT NO V5O00-R1RLV .01.020A00N 70CA 40.130 VIÇYOP500058S ,Cyp VLFAPGA.MN7 YLYAPO0MNErob F_GMP1TASO._5$GT_PVDE50.F7_0EYLRLiE VAVYMICF F_SLVYLEGOO,W5LpvoIALpcAToNotDyoA31LIAHsvxAvAoGANn1CATar00 VLFAPGKMNCO.b IRFOPOKMACs.gbu C-GAWISFSU-- -55GW- .VKT?tG_VD-VMLREIKEVRRMYSVYGEVE0CIII lar K-GPIRLSCSO- _SRGY__VYEPNG.FT0f0AYO0CH.4SMt5E sOTAVYVGn0RxPN4Et.0CaVnLRFPCA1iI lot LIK St Alt IIYCP(,*MSch.DCC LG0IVV5CSfl---5k0S-- 1050KG- I0PEOVLEIj (LXFGSLEEITKESVKI.FSGE) VEY1M- ‘V-40 AiPSA1 5400 4M 40M01511KN.cro L.GACVVSL5o--s0GNLVATGESA.sld cGGSYVS150---SKG5LSc.cer c_0005VSCSO---51(0--I tSETGHurn F_sMCtAIIGE_._S0G5_.IWtPDGmPKE1f0r010HSsIO rp.IPYEOSIIEA.___DC0tiIPMSEK0.TK5N_0P0.__0APCAth1t_t KINVIPOLVINiklus, p_GpVGE__SoGS__It0lP0G-IDPKaE0FKLOSILG FP_tÇYY5SILEA._._0CuIl.IPMSEX01T SK-AFRA4II GAJIC I.E-R NII4VIFOLOLNBoelne F-OXCi’AVGL- --5055-- IWtIP0G-IQPXftEOFKLQINGTILG pp_t11ittsicEV_0C01C1PM5E00(.TX5 --0 GA 05lFt€-ft N!HVIPOCYLMtot F_oexc0gvGE.__S05__lW30—I3PXtEtFU.0SILG FPPXVÏEG5ILEA___.0cOtLtPM50XI0500P00__0T40fltGADKIntE NIHVIPCLYLNCh ickon KINVIPttLYLH
0
____________________
350 350 370 300 390 400 410 420pjur MGVT055FEWQKITCYVW TIEEt040t1N 4DAAÏYAV-’OAtERG0YAtiC.dit AGGYTVSYFE40QKCYGOYW SE_EE__VE05EEI»40(7$ESIW(tKEE3H _IEM5)*1SIOX-VAEAi4-KLRGWYPaoo SGCYL0SYVEWt*QYG IEAE___V0EKQGAflt%(KGVFAVADEYHVT tRCAVYl0YAI-XSI0VAN-KIRGWYKIol AGGVÎY5YPE.t1I40RAU KEMIIOA.1R-SAEAX-EAIIGtWP5.jol AOGYYOSYVEWAJ0ILE1I SEEF.AUV00?FNDYØLWII DLITMj4OU000_WWWCMGILUsy. A650LVSGPENSONSERI.SW TAtE__’iDSKl.H04T0I)GGA»R G0_Kt.VAGANtVGFC_IA0A0tRW0. Iso A050SVSOCOM5I3IISSRLIM$.typ A0GVAT5GLEPAtl4A40LSW — NYVCG.ANlO(-VAD*$4fiGYL3 M0U,iSI,LI,MNMAII,l —
AGGVATS5LEPOq1MRLGW 00XASARtHNtStOZ4144CtEO0GE40QT NYGAN1AGFXJÂAV.lAG1U£0 .n.l I,GG405M.E)Ç39ASRDSI4 — DWVGANIA(K.VA801l41ICo.gts V5SVATSAL1ÇNASR5FLCOLior MOVAYSGLEPTQIOONSCIN TREE__VR0KiERIYi(31Y3SAJNPEY40 QAtIAGÇTX_VA0AVKP05MSCh.tCc £00 V%LEMQN$iR051iUcro CO00AYS0LES1LIAJ p_POJWPNCONFF9OltAXTWEMEGELP0LVAG5t1A0F0xVA0KI5tPsid ASG’IAVSSLEW’QNSARLSWSc.c4r L6yAY5SCt,QhI5tTWtKIs ASGTW5IFEI0.KNLK4VS350LSF0YER0SNYHLLHSVQ€SLE40FO0OT1P1 U00RISGAS000E0050LAYMERShR IKNTAJ4(YNL Gt.0I.RtMVONAIEK_VFKVY_NEALWTFTilool. PYVPtAEF00RISGA3EX IVIISGLAYTPORSdA IKOTNKYNL G1_OLKTMOV%EVFKVIIEAGVrFTKon me A0GVW$iPQILkN1iIHh500LTÇK5tR0510.C00C1CbT PIVPTAEFI3NI$6ASESSIVN$01.AYT)8pal PVYP1AtFR0SCASCX01V05G A RGA IÇTNii(Ytfl. 0tOLRTUWi lEK-OP Y00004TflCOIcken AGCV1YSAF.kHLKO4SYGRLT 000GSIYlii.IYS000SLERXFGO(000T P0VPTAEFR15GASEKIL405OI.AYT6RS 11ÇAJq(IL____GL_0tRIU30MIEX_YF11LTtT
Figure 3 A1inment of GDH sequences from different organismS. Pyrococcus furiosus (P. fur; thisvork), Qlostridiuni diftiei/e (C.diO, Peptostteptococcus asacctrarolyticus (Psasa), Halobacteriurnsalinctriuni (H.sal). Sulfblohu.ç solfataricus (Ssol). Clostridium symbiosum (Csym), Giardia lamblia(Glam), Salmonetla t3’phimuriu?n (S.typ), Unknoosn microorganisrfl (?), Escherichia coti (E. col),Corynebacteritmi metassecoÏa (ComeI); C’omwbacteriu,n gluia,nicuin (Co.glu): Chiorella sorokiniana(ChLsor); Schwanniomyces occidentalis fSchocc); Neurospora crassa (Ncra), Saccharom.3’ces cerevisiac fS.ccr), ,4spe,gilliis ,iidttlans (A.nid), Human, Mouse, Rat, Bovine and Chicken fBenachenhouLahfa er al.. 1993). Idcntieal residues in all GDHs are indicated by asterisks. Secondary structures,derived from the Csnibiosuin GDH (Tefler c’t al., 1992) are inciuded above the alignment
Kopie für Lizenzkunden von subito e.V., geliefert und ausgedruckt füt TNO (SLIO6XOO2O5E)
PYROCOCCUS FUR!OSL’S GDH 139
Table 3 Arnino acid compositions of glutamate dehydrogenasc from 1’ /hrtosus, two other archacaand two bacteria.
Residues P. /iiriosus C difJkile P. asaccharuhyici.v FL.vu/iuariuin S..votfiiuricu.v
A 39 36 36 49 34C 1 5 5 3D 24 18 21 37 21E 36 38 39 40 31F 10 12 14 4 11G 34 43 45 33 43H 5 4 2 7 41 35 29 25 22 31K 33 33 30 13 37L 24 24 27 27 39M 13 16 11 II 13N 14 18 21 13 18P 19 14 24 16 13Q 11 II 11 12 14R 20 12 19 24 17S 14 26 14 22 20T 23 20 22 31 18Y 35 38 3$ 39 36W 10 8 9 9 4Y 20 16 18 IS 16
TOTAL 420 422 421 435 421RESIDUES
be characteristic for the thermostabilization of eubacterial proteins, i.e. anincrease in the Arg to Lys ratio and the number of Pro and Gly residues (Davieset al., 1991; Suzuki et al., 1991), are not found in the P. furioszts and S.sotfataricus GDH (Table 3) or the F. woesei QAPDH (Zwicki et al., 1990). Thissuggests that different strategies are used in Bacteria and Archaea to stabilizetheir proteins. To identify changes in the amino acid sequence, that are relatedwith an increase in thermostability, molecular modelling studies based on the3-dimensional structure of the C.svmbiosurn enzyme, followed by proteinengineering experiments are now being initiated.
Acknowtedgemen ts
We thank B.Bachmann (Department of Biology, Yale University, New Haven)and B. Snedecor (Genenetech, mc. San Francisco) for supplying E. coti strains.This research was supported by a grant from the Programme Committee onIndustrial Biotechnology (PcIB).
ReferencesAono, S., Btyant, F. 0. and Adams, M. W. W, (1989) A novcl and remarkably thcrmostablc
ferredoxin from the hyperthermophulic archaebacterium Pyrococcus furiousus. J. Bacteriol. 171.3433—3439
l3aker, P. 1., Britton. K. L., Engel, P. C., farranis. G. W., LiIley, K. S., Rice. D. W. and Stiliman, T. J.(1992) Subunit assembly and active site location in the structurc of glutamate dchydrogcnasc,Proteins 12, 75—86
Kopie für Lizenzkunden von subito e.V., geliefert und ausgedruckt für TNO (8L106X00205E)

140 RIK 1. L. EGGEN ET AL.
Bcnachenhou-Lahfa, N.. fortcrrc, P. and Labcdan, P. fl993) Evolution of Gtutamate dehydrogenasegcnes: Evidence for two paralogous protein families and unusual branching patterns of thearchacbacteria in the universal trec of life. J. 61e!. Evol. 36, 335—346
Berberich, M A. (1972) A glutamate dependent phenotypc in Escherichia coti K-12: the result of 2m utat iOflS. Blochein. BIOJJIO’S. Res. Cominun. 47, 1498—1 500
l3ryant, F. 0., and Adams, M. W. W. (1989) Characicrization of hydrogenasc from the hyperthermophilic archacbactcrium, P’rococctts /i’riosiis. J. Biot, Chem. 264, 5070—5079
Consalvi, V., Chiaraluce, R., Politi, L., Vaccaro, R., de Rosa, M. and Scandurra R. (1991) Extremelythermostable glutamatc dehydrogenase from the hyperthermophilic archaebacterium Pj’rococcztsfitriosus. Lor. .1. Bioche,n. 202, 1189—1196
Constantino. H. R.. Brown. S. t-t. and Kelly, R. M. (1990) Purihcation and characterization of anck-ghlcosidase from a hyperthermophilic archaebactcrium, Pi’rococcus furiosus, exhibiting aemperature optirnum of 105 to 115°C. J. Bac’ieriot. 172, 3654—3660
Davies, G. .1., Littiechild, J. A., Watson, H. C. and Halt, L. (1991) Sequence and expression of the gene°neoding 3-phosphoglvccratc kinasc from Bach/ns stearothermophitus. Gene 109, 39—45
Dcvcrcux, 1., Haehcrli, P. and Smithies, 0. (1984) A comprchensive set of sequence analysis programsfor the VAX. Nucleic Acicis Rc’s. 12, 387—395
Eggen, R., Gccrling, A., Waits, 3. and dc Vos, W. M. (1990) Characterization of pyrolysin, ahypcrthermoact ve seri ne proteasc from the archaebacteri um P.i’rococciis furiosus. ff41S MicrobiotLeus, 71, 17—20
Eggen R. t. L. Geerling, A. C. M.. Jetten, M. S. M. and dc Vos, W. M. (1991) Cloning, expressionand sequence analysis of the gcncs for carbon monoxide dehydrogenase of Meshanothrixsoehngenii. J. Bio!. Che,n. 266, 6883—6887
Eggen R. t. L., Geerling, A. C. M., Waldkötter, K., Antranikian, G. and de Vos, W. M. (1993) Theglutamate dehydrogenasc-encoding gene of the hyperthermophilic archaeon Pyrococcus furiosus:sequencc, transcription and anatysis of the deduced amino acid sequence. GL’ne 132, 143—148
Fabry, S., Lang, 3., Nicrmann, T., Vingron. M. and Hensel, R. (1989) Nucleotide scquence of theglyccraldchydc-3-phosphate dehydrogenase gene from the mesophilic methanogenic archaebacteriaitlelhanobacter!uin brj’antii and ?vfc’thanobacterium formicicum. Comparison with the respectivegene structure of the closely related extreme thermophile Methanothermus fervidus. Etu’. J.Bioc hem. 179, 405—413
Fiala, G., and Stetter, K, 0. (1986) Perococcus jitiosus sp. nov. represents a novet genus of marineheterotrophic archaebactcria growing optimally at l00°C.Arch. Microbiol. 145, 56—6 1
Gibson, T. 3. (1984) Studies on the Epstein-Barr virus genome. P1’I.D. Thesis Cambridge University,Cambridge, England
Gouy, Mand Gautier, C. (L982) Codon usage in bactcria: Corrclation with gene exprcssivity. NucleicAcicis Rc’s. 10, 7055—7074
Kengen, S. W. M., Luesink, E. J., Stams. A. J. M. and Zehnder, A. J. 8. (1993) Purification andcharacterization of an cxtremely therrnostab]e 3-g1ucosidase from the hyperthermophulic archaconPyrococcus furiosus. Lor. J. Riochem. 213, 305—3 12
Kcngcn, S. W. M. and Stams, A. J. M. (1994) Formation of L-alanine as a reduced end-product incarbohydrate fermentation by the hyper-thermophitic archaeon Pyrococcus Jiiriosus. Arch. Micro-bio!. 161, 168—175
Koch, R., Spreinat, A., Lemke, K. and Antranikian, G. (1991) Purification and properties of ahyperthermoactive c-amy1asc from the archacobacterium Pi’rococcus woesei. Arch. Microbiol. 155,572—578
Lake. 3. A. (199 t) Tracing origins with molecular sequences: metazoan and cukaryotic beginnings.i’renc/s Biochein. Sci. 16, 46—50
Mukund S.. and Adams, M. W. W. (199]) The nove] Lungsten-iron-sulfur protein of thehyperthermophilic arehaebacteriurn, Fyrococcusfuriosus, is an atdehyde ferredoxin oxidoreductase.J, Bio!. C’heni. 266, 14208—14216
Nagata, S. Tanizawa, K., Esaki, N., Sakamoto, Y., Ohshima, T., Tanaka, H. and Soda, K. (1988) Genecloning and sequencc determination of leucine dehydrogenasc from Bacitlus stearothermophilus andstructural comparison with other NAD(P)-dependent dehydrogcnases. Biochem. 27, 9056—9062
Okazaki. N.. Hibino, Y., Asano, Y., Ohmori. M., Numao. N. and Kondo, K. (1988) Cloning andnuctcotide scqucncing of phenylalaninc dehvdrogenase gene of Bad//os sphaerictts. Gene 63,33 7—34 1
Remaut. E.. Stanssens. P. and Wiers. W. (1981) Plasmid vectors for high-efliciency expressioncontrollcd hy the promoter of coliphage ]amhda. Gene 15, 8 1—93
Kopie füt Lizenzkunden von subito e.V., geliefert und ausgedruckt für TNO fSLIO6XOO2O5E)

PYROC’OCCLS FUR1OSLS GDH t4 t
Robb, F. T., Park. J.-B. and Adams. M. W. W. 91992) Charactcrization of an extrcmclv thermostableglutamate dehydrogenase: a key enzyme in the primarv mctabolism of the hypcrthcrmophilicarchacbactcrium, Pi’rococcus Juriovus. Biochini. Bioplns.Acia 1120. 26 7—272
Sambrook, J., Fritsch, E. F. and Maniatis. T. (1989) Molecular Cloning: a Laboratory Manual. (‘oldSpring Harbor Laboratory, Cold Spring Harbot, N.Y.
Schafer, T. and Schonheit, P. (1991) Pyruvatc mctabolism of the hyperthermophilic archacbactcriumPi’rococcus fliriOsus. Arch. ?fü’i’obio1. 155, 366—377
Schifer T., and Schonheit, P. t 1992) Maltose fermentation to acetatc, C02 and H-, in the anacrobichyperthermophilic archacon Fvrococciis fitrioszis: evidence for the operation of a novel sugarfermentalion pathway. Arch. Mierohiol. 15$, 188—202
Schfcr, T. and Schönhcit, P. (1993) Gluconcogenesis from pyruvatc in the hyperthcrmophilicarchaeon ?J’rococcus /riostts: involvement of reactions of the Embdcn-Mevcrhof pathway. :Irch.Microbiol. 159, 354—363
Schotz, S., Sonnenbichier, J., Schafer, W. and Hensel R. (1992) Di-myo-inositol- 1 , 1 ‘-phosphate: a newinositol phosphate isolated from Pi’rococcus wnesei. FEBS Lc’U. 306. 239—242
Smith. E. L., Austen, B. M., Blumenthal, K. M. and Nyc. J. F. (1975) Glutamate dchydrogcnases. In:Boycr PD (ed) The enzymes, vol XL Academic Press, New York, pp 293—367
Snedccor. 3., Chu, H. and Chen, 1. (1991) Selection, cxpression, and nucleotide sequencing of theglutamate dehydrogenasc gene of Peptosireptococcus asacharolidcus.J. Bacteriol. 173, 61 62—6167
Stetter, K. 0., Fiala, G.. Huber, G., Huber. R. and Segerer, A. (1990) Hyperthermophilicmicroorganisms. Fliul’IS Microbiologv Rc’views 75, 117—124
Suzuki, Y., Hatagaki, K. and Oda, 1-1. (1991) A hyperthermostabic puflulanase produced by anextreme thermophile, Badillus Jiavocaldaritis KP 1228, and evidcncc for the proline theory ofïncreasing protein thermostability. AppI. Microbiol. Biotechnol. 34, 707—7 14
Takada, H., Yoshimura, T.. Ohshima, T., Esaki, N. and Soda, K (1991) Thermostable phenylalaninedcli ydrogenase of Thermoacrinomeces interrnedïus: Cloni ng, expression. and scq uenci ng of i ts genef Biochem. 109, 371—376
Teller, J. K., Smith, R. J., McPherson, M. J., Engel, P. and Guest, J. R. (1992) The glutamatedehydrogenase gene of L’Iostridiuin s rnhiosurn: Cloning by polymerase chain reaction. sequcnceanalysis and over-expression in Escherichia cob. Eur. J. Biocheni. 206, 151—159
Tiboni, 0., Cammarano, P. and Sanangelantoni. A. M. (1993) Cloning and sequcncing of the gcneencoding glutamine synthetase 1 from the archaeon Pvrococcus woesci: Anomalous phylogeniesinlèrred from analysis of archaeal and bactcrial glutamine synthetasc 1 sequences. J. Bacieriol. 175,2961—2969
Uemori, T., Ishino, Y., Toh, 1-1., Asada, K. and Kato. 1. (1993) Otganization and nucleotide sequenccof the DNA polymerase gene from the archacon »vrococcus Jiiriosus. Nucleic Acids Research, 21,259—265
Wierenga, R. K., De Maeyer, M. C. H. and Hol, W. G. J. (1985) Interaction of pyrophosphatemoieties with a-helices in dinucicotide binding proteins. Biochern. 24, 1346—1357
Woesc, C. R., Kandler, 0. and Wheelis, M. (1990) Towards a natural systcm of organisms: Proposalfor the domains Archaea, Bacteria and Eucaiya. Proc. Nat!. Acad. Sci. USA 87, 4576—4579
Yanisch-Perron, C., Vieira, 3. and Messing, 3. fl985) lmproved M13 phagc cloning vectors and hoststrains: nucteotide scqucnces of the M13mp18 and pUCI9 vcctors. Gene 33, 103—t 19
Zillig, W., HoIz, 1., Klenk, H.-P., Trent, J., Wunderi, S., Janekovic, D., Imset, 1. and Haas, 8. (1987)Pyroeoccus woesei sp. nov., an ultrathermophihc marine archaebacterium, representing a novetorder, Ther,nococcates. Systein. Appi. Microhiol. 9, 62—70
Zwicki, P., Fabry, S., Bogedain, C., Haas, A. and Hcnsel, R. (1990) Glyceraldchydc-3-phosphatedehydrogenase from the hyperthermophilic archaebactcrium Pyrococcus wocsei: charactcrization ofthe enzyme, cloning and sequencing of the gene, and expression in Escherichia coli. ]. Bacieriol.172, 4329—4338
Kopie füt Lizenzkunden von subito e.V., geliefert und ausgedruckt für TNO (SLIO6XOO2O5E)
