purification and characterization of carbamoyl-phosphate synthetase from the deep-sea...
TRANSCRIPT

Eur. J . Biochem. 236, 189-199 (1996) 0 FEBS 1996
Purification and characterization of carbamoyl-phosphate synthetase froin the deep-sea hyperthermophilic archaebacterium Pyrococcus abyssi Cristina PURCAREA I, Valtrie SIMON ’, Daniel PRIEUR’ and Guy HERVE’
’ Laboratoire de Biochimie des Signaux RCgulateurs Cellulaires et MolCculaires, URA CNRS 1682, UniversitC Pierre et Marie Curie, Paris, France Laboratoire de BactCriologie Marine, UPR CNRS 4601, Station Biologique, Roscoff, France
(Received 3 October 1995) - EJB 95 1610/4
Carbamoyl-phosphate synthetase was purified from the deep-sea hyperthermophilic archaebacterium Pyrococcus abyssi. This enzyme appears to be monomeric and uses ammonium salts as nitrogen donor. Tts activity is inhibited by some nucleotides that compete with ATP. In contrast with the carbamoyl- phosphate synthetases investigated so far, this enzyme is very resistant to high temperature. Its low molecular mass (46.6 kDa) and its catalytic properties suggest that the gene coding for this enzyme is a previously postulated ancestor whose duplication gave the genes coding for carbamoyl-phosphate synthe- tases and carbamate kinases.
Keywords : carbamoyl-phosphate synthetase; hyperthermophile; deep-sea vents ; Archae; Pyrococcus.
Pyrococcus ahyssi is a hyperthermophilic archaebacterium recently isolated from a deep-sea hydrothermal vent located 2000 m deep in the North-Fiji Basin [I , 21. This microorganism has been characterized as a strict anaerobic, sulfur-metabolizing archaeum, whose optimum temperature for growth is 96°C [3] . Although R abyssi grows at atmospheric pressure, its generation time decreases at high pressure and its maximal temperature for growth is increased when cultured under hydrostatic pressure ; this microorganism is therefore classified as barotolerant to bar- ophilic [3] . Cell growth at such a high temperature raises physio- logical problems, e.g. the instability of some metabolites, cellu- lar components, and enzymes. For instance, carbamoyl phos- phate, the common precursor for the biosynthesis of arginine and pyrimidine nucleotides, is a very unstable compound. At 96”C, its half-life is only a few seconds 14, 51. In all organisms, carbamoyl phosphate is synthesized by carbamoyl-phosphate synthetases (CPS), enzymes which are known for their instabil- ity. Most of the eubacteria, plants like pea [6, 71, and mung bean [ X I possess a single CPS that provides carbamoyl phosphate for both the arginine and pyrimidine pathways. Eucaryotic organ- isms, except for plants, possess two CPS enzymes, each specific for one of these two pathways (CPS-A and CPS-P or CPS I and CPS 11, respectively). Some CPS enzymes use NH, as the donor of the amino group of carbamoyl phosphate while others use glutamine. The latter enzymes contain a subunit or domain (glu- tamine amidotransferase) that binds glutamine, catalyses its de- amination, and transfers the NH, to the CPS catalytic site. For instance, Escherichia coli CPS is composed of two subunits, a
Correspondence to G. HervC, Laboratoire de Biochimie des Signaux Regulateurs Cellulaires et Moltculaires, URA CNRS 1682, Universitt Pierre et Marie Curie, 96 Boulevard Raspail, F-75006 Paris, France
Ahbreviarions. ATC, aspartate transcarbamoylase; CPS, carbamoyl- phosphate synthetase: PPRibP, 5-phosphoribosyl diphosphate; Ap.A, adenosine (5’)triphospho(S’)adenosine: Ap.A, adenosine (5’)pentaphos- pho(5’)adenosine.
Enzymes. Carbamoyl-phosphate synthetase (EC 6.3.5.5) ; aspartate carbamoyltransferase (aspartate transcarbamylase) (EC 2.1.3.2): carba- mate kinase (ATP:carbamate phosphotransferase) (EC 2.7.2.2).
small subunit (42 kDa), which is the glutamine amidotransfer- ase, and a large subunit (117 kDa) which is CPS [9]. In the absence of the small subunit, the large subunit of CPS, is active and uses NH: as substrate.
In a previous study, aspartate transcarbamoylase (ATC), the first enzyme of the pyrimidine pathway, which uses carbamoyl phosphate as substrate, was partially purified from the cell-free extracts of P. abyssi and its catalytic and regulatory properties were determined [lo]. In the present work, the biosynthesis of carbamoyl phosphate was investigated in this hyperthermophilic archaebacterium. The results obtained show that this microorga- nism possesses a single CPS enzyme that uses NH,‘ but not glutamine as substrate. This enzyme has been purified to homo- geneity and its catalytic and regulatory properties were deter- mined. In contrast with the CPS enzymes known so far, this enzyme is extremely thermostable. Its activity is inhibited by some nucleotides. Its molecular mass and properties suggest that the gene coding for this enzyme is an ancestral form whose du- plication and fusion provided the presently known genes of CPS enzymes and carbamate kinases.
MATERIALS AND METHODS
Chemicals. Nucleotides, adenosine (5’)triphospho(5’)ade- nosine (AplA), adenosine (5’)pentaphospho(5’)adenosine (Ap5A) pyruvate kinase, rabbit muscle lactate dehydrogenase, catalase from bovine liver, ribonuclease A (type I-AS from bo- vine pancreas), ovalbumin (grade V), myoglobin (type I1 from whale skeletal muscle), acivicin (a-amino-3-chloro-4,5-dihydro- 5-isoxazole acetic acid) and phosphoenolpyruvate were purchased from Sigma; blue dextran and bovine serum albumin (BSA) were from Serva; 5-phosphoribosyl diphosphate (PPRibP) was from P-L Biochemicals Inc. ; antipyrine (13- dimethyl-2-phenyl-3-pyrazolone) was from Kodak; diacetyl mo- noxime (2,3-butanedione monoxime) was from Aldrich; DEAE Affi-Gel Blue and hydroxyapatite (Bio-Gel HTP) were from Bio-Rad. Sodium [’4C]bicarbonate was from NEN (2.1 GBq/

190 Purcarea et al. (Eul: J. Riochem. 236)
mmol ; 57.30 mCi/mmol). N-(Phosphonacety1)-L-aspartate was a generous gift from Drs V. Narayanan and L. Kedda of the Drug Synthesis and Chemistry Branch, Division of Cancer Treatment, NIH, Silver Spring, MD.
Growth of bacteria. I? abyssi strain GE5 (CNCM 1-1302) was grown under anaerobic conditions at 95°C in an artificial sea water medium containing elemental sulfur supplemented with bactopeptone and yeast extract (YPS) as previously re- ported [3]. This procedure was adapted to large volumes by G. Raguenes and G. Barbier (Institut Francais de Recherche et Ex- ploitation des Mers, Brest) and A. Escaut, S. Cortial, and D. Joseph (Centre National de la Recherche Scientifique, Gif-sur- Yvette). A 20-1 culture was used as inoculum for a 200-1 fer- menter. Anoxic conditions were obtained by bubbling N, (16.7 I/ rnin). The medium was stirred at 220 rpm. The cell growth was stopped after 5 h in the exponential phase and the cells were col- lected by continuous centrifugation at 10000 g for 10 rnin and stored at -80°C. Such a culture provided about 200 g bacteria of which about 20 g were used in each purification procedure.
The cell-free extracts were prepared by resuspending the bacteria in S O mM Tris/HCI, pH 8, 1 mM dithiothreitol and 0.1 mM EDTA. These cells were disrupted by sonication six times for 30 s using a Biosonik I11 sonicator at 20 kilocycles/s.
Enzyme assay. The carbamoyl-phosphate synthetase activity was determined by the radioactive test [I11 using NaH['4C]C0,, which is transformed into the stable compound carbamoyl aspar- tate in the coupled reaction with an excess of ATC. Since the two substrates ammonium chloride and sodium bicarbonate, as well as the intermediary product carbamoyl phosphate, are ther- molabile compounds, the activity was measured at 37°C and, when possible, at 70°C. Depending on the reaction temperature, the coupling enzyme used was either the catalytic subunits of E. coli ATC (3.7 pg at 37°C and 37 pg at 70°C) or F! abyssi partially purified ATC (6 pg at 70°C). At 37"C, the standard incubation contained 50 mM Tris/HCl, pH 8, 120 mM ammo- nium chloride, 60 mM NaH['"C]CO,, 0.75 mM Mg ATP (from a stoichiometric mixture of ATP and magnesium chloride) and 20 mM aspartate. At 70°C and pH 7.5, the concentrations of the substrates were 150, 100, 1.5, and 30 mM, respectively, in a total volume of 300 pl. The samples were incubated in glass tubes for hemolysis and covered with glass beads. After 30 rnin of incubation at 37°C or 2 min at 70"C, the reaction was stopped by the addition of 0.7 ml 10% trichloracetic acid. After elimina- tion of the precipitate by centrifugation at 10000 g for 10 min, the supernatant was evaporated in an oven at 95°C. The solid residue was redissolved in 4 ml water and the radioactivity of the samples was measured after addition of 8 ml Aquasol scintil- lation mixture. 1 U CPS activity catalyzes the formation of 1 pmol carbamoyl aspartate/h. The specific enzyme activity is defined as U . mg-' and was determined at 37°C under the standard conditions described above.
Stoichiometry of the reaction. To determine the stoichiome- try of the reaction catalyzed by the F! abyssi enzyme, the rates of ATP consumption and carbamoyl phosphate production were compared as follows. In the overall reaction, the amount of ATP used was determined on the basis of the production of ADP in a coupled assay with pyruvate kinase and lactate dehydrogenase [12]. The incubations were performed in the presence of 6 pg F! abyssi enzyme, 20 U pyruvate kinase, 30 U lactate dehydroge- nase, 80 mM Tris/HCI, pH 8,200 mM NH,CI, 66 mM NaHCO,, 0.75 mM Mg ATP, 100 mM KCI, 150 yM NADH and 1 mM phosphoenolpyruvate. The reaction was monitored by following the absorbance at 340 nm using a Shimadzu spectrophotometer. In the same overall CPS reaction, the carbamoyl phosphate pro- duction was followed using the CPS-ATC assay described above, in the presence of 20 mM aspartate.
In the bicarbonate-dependent adenosinetriphosphatase partial reaction, followed in the absence of ammonium chloride. the amount of ATP consumed was determined on the basis of the production of ADP as described above. These adenosinetriphos- phatase reactions were performed at 27°C.
All nucleoside phosphate derivatives, PPRibP and ornithine as well as Nacetyl-L-glutamine, AplA, and Ap,A solutions were adjusted to pH 8. Their influence on the rate of the CPS reaction was measured in the presence of 60 pM MgATP at both 3 7 "C and 70°C. The inhibition is expressed as follows:
x100, 0 0 - 0, inhibition (%) = ~
011
where u,, is the reaction rate in the absence of nucleotide u hile u, is the reaction rate in the presence of inhibitor.
The pH dependence of the activity was determined using a three-buffer system composed of 51 mM diethanolamine, 51 mM N-ethylmorpholine, and 0.1 M Mes adjusted at different pH values between 6 and 10 [13] at both 37°C and 70°C. The pH measurements were performed using a Knick 655 pH nieter and an Ingold microelectrode.
Pressure assay. Hydrostatic pressure tests were performed using the previously described high-pressure reactor that allows injection, mixing, and sampling without release of the pressure [14]. When the apparatus was used to follow the rate of reaction under pressure, 300-yl samples were collected in 0.7 ml 0.2 M acetic acid to stop the enzymatic reaction. The amount of car- bamoyl aspartate formed was then determined by the colorimet- ric assay [15].
Electrophoretic analysis. SDS/PAGE was carried out ac- cording to Laemmli [16], using 12% polyacrylamide. The molecular-mass markers used (Bio-Rad prestained low-range mixture) were phosphorylase b (106 kDa), BSA (80 kDa), (will- bumin (49.5 kDa), carbonic anhydrase (32.5 kDa), soybean trypsin inhibitor (27.5 kDa) and lysozyme (18.5 kDa). These are the apparent molecular mass values given by the supplier. for prestaining alters the migration of the markers in SDS/PAGE. The electrophoretic analyses under non-denaturing conditions were performed according to Jovin [17]. Proteins were detected using the silver-stain method [18]. CPS activity was detected in a 7% acrylamide non-denaturing gel as follows: after protein migration, the gel was cut into 1-mm slices, which were incu- bated in the standard assay mixture for 40 rnin at 37°C. The isoelectric point was determined using a Phast System isoelectric focusing apparatus (Pharmacia).
Molecular mass determination. The CPS molecular mass was estimated using size-exclusion chromatography [19----211 and SDS/PAGE. A Sepharose 6B column (1.6 cmX90 cm) was equilibrated with buffer A (50 mM Tris/HCl, pH 8, 1 mM dithio- threitol, 0.1 mM EDTA) at a flow rate of 7.2 ml . h-'. The void volume (0,) and the maximum elution volume were determined using blue dextran ( 5 mg) and dithiothreitol (10 mg), respective- ly, in a final volume of 1 ml adding buffer A and two drops of 99.5% glycerol. The proteins used for the calibration of the col- umn were ribonuclease, ovalbumin, BSA, and myoglobin. 10 mg of each protein in a final volume of 1 ml in the presence of two drops of 99.5% glycerol were loaded two at a time and eluted at the same flow rate; 1.5-ml fractions were collected. The elution volume (N,) of each standard marker was calculated in grams. Proteins were identified either by their absorbance at 280 nm or by their enzymatic activity.
Protein assay. Protein was determined according to Lnwry [22], using BSA as standard.
Data processing. All the kinetic data were treated with the KaleidaGraph program on a Macintosh computer. The substrate saturation curves were fitted either to the Hill or to the Micha-

Purcarea et al. ( E m J . Biorhem. 236) 191
Table 1. Requirements for the P. abyssi CPS reaction. The activity was measured at 37°C as described in the Materials and Methods sec- tion.
Substrateshnhibitors Residual activity
Complete system - NH,CI - NaHCO, - ATP - MgC1, - MgC1, + EDTA (2 mM) - NH4C1 + Gln (50 mM) - aspartate + Ap,A (0.1 mM) + acivicine (2 mM) + N-acetylglutamate (5 mM) + N-(Phosphonoacety1)-L-aspartate
%
100.0 1.6
31.0 0.6
35.3 1.2
13.0 1.2
8.0 97.0
100.0 (0.01 mM) 15.8
elis-Menten equation. The pK, values were calculated from the Dixon equation [23] fitted to the V,,, versus pH curve.
RESULTS
Characterization of CPS activity in l? abyssi cell-free ex- tracts. When a dialyzed cell-free extract of 19 abyssi is chro- matographed on a Sephacryl S 300 HR column, at either 23°C or 70”C, a single peak of ammonium-dependent carbamoyl- phosphate synthetase activity is detected. The nature of this CPS reaction was verified by measuring its dependence on each of the substrates of the CPS and ATC enzymes used in the coupled assay. The results are shown in Table 1. A strict dependence on NH,C1 and ATP is observed. The dependence of the reaction on aspartate, Mg”, and NaHCO, is less strict. In the absence of sodium bicarbonate, a 31 % residual rate of activity is observed due to bicarbonate contamination of the solution by the atmo- spheric CO,. In the absence of added MgCl,, a 35% residual rate of activity is observed, due to the contaminating traces of Mg” present in the other components of the incubation medium (NaHCO,, NH,Cl). Accordingly, in the presence of 2 mM EDTA the reaction is completely inhibited. In the presence of 50 mM glutamine as the amino group donor, only 13 % of the residual rate of reaction is observed, a result that can be explained by the NH, content of the reagents used, especially glutamine. This lack of significant glutamine-dependent activity was verified un- der various conditions of substrate concentrations both in the crude extract and after complete purification of the enzyme. As far as aspartate is concerned, slight proteolysis during the incu- bation is most probably responsible for the 7 % residual activity observed. A similar steady-state rate of reaction was observed whether the amount of carbamoyl phosphate formed was mea- sured using the CPS-ATC coupled assay or by reaction with hydroxylamine in the absence of ATC [24] at both 37°C and 70°C. This CPS reaction was further identified through the use of specific inhibitors of either ammonium- or glutamine-depen- dent CPS and ATC. Ap,A is a potent and specific inhibitor of E. coli CPS since it binds simultaneously to the two ATP sites involved in this enzyme reaction [25]. As shown in Table 1, at a concentration of 0.1 mM this compound inhibits the investigated reaction by 92 %. Acivicine, a strong inhibitor of glutamine-de- pendent CPS [26], does not affect the activity present in I? abyssi extracts, which is consistent with the lack of utilisation of glutamine by this enzyme (Table 1).
It was shown previously that under some conditions, reduc- ing agents such as dithiothreitol can inhibit the glutaminase ac- tivity of E. coli [27]. It was verified that the cell-free extract of I? ubyssi does not exhibit glutamine-dependent CPS activity wether or not dithiothreitol is added in the extraction buffer at a concentration of 1 mM.
N-Acetyl-L-glutamate was shown to stimulate the activity of the ammonium-dependent CPS while this compound does not influence the reaction catalyzed by the glutamine-dependent CPS enzyme [28]. At a concentration up to 5 mM, this com- pound has no influence on the rate of the reaction catalyzed by the I? abyssi enzyme (Table 1). The dependence on ATC of the coupled assay used is confirmed by the 85 ?k inhibition observed in the presence of N-(phosphonoacety1)-L-aspartate, a specific inhibitor of ATC [29-311.
Enzyme purification. All operations, unless indicated, were carried out at room temperature. After each chromatography step, the absorbance at 280 nm and the CPS activity profiles were determined in order to select the appropriate fractions for further treatment.
Preparation of cell-jree extract. About 20 g GE5 cells frozen at -80°C were thawed and suspended in 20 ml degassed buffer A and disrupted by sonication as indicated in the Materials and Methods section. The extracts obtained were centrifuged at 1OOOOg for 20 min. The supernatant was then collected as the cell-free extract.
Q Sepharose Fast Flow chromatography. A 15-ml sample of cell-free extract was applied to a column of Q Sepharose Fast Flow (1.6 cmX25 cm) previously equilibrated with buffer A and run at 1.28 ml . min-I. The column was washed with 90 ml of the same buffer, and 3-ml fractions were collected. Elution was performed first with 90 ml 0.1 M NaCl and subsequently with a 500-ml linear gradient of increasing NaCl concentration (0.1 - 0.3 M) in buffer A. 2-ml fractions were collected. CPS activity was eluted between 0.20 M and 0.22 M NaCl. The active frac- tions were pooled and dialyzed at 4°C overnight against the same buffer.
DEAE Affi-Gel Blue chromatography. The dialyzed prepara- tion (24 ml) from the Q-Sepharose Fast Flow step was loaded on top of a 1.4 cmX13 cm DEAE Affi-Gel Blue column equili- brated with buffer A. After washing with 56 ml of the same buffer at 0.5 ml . min-I, the column was eluted at the same flow rate with a 120-ml linear gradient of 0-0.3 M NaCl and 1-ml fractions were collected. The CPS peak was eluted between 0.17 M and 0.20 M NaCI. The fractions with the highest CPS activity were pooled and dialyzed overnight at 4°C against buffer A.
Hydroxyapatite chromatography. A 5.13-ml sample of the CPS preparation after DEAE Affi-Gel Blue chromatography was applied to a 0.9 cmX 17 cm hydroxyapatite column previously equilibrated with buffer A followed by 30 ml of the same buffer using a flow rate of 0.21 ml . min--l. The elution was performed at the same flow rate in 1-ml fractions using a linear gradient (0-0.15 M) of potassium phosphate adjusted at pH 8. The CPS peak was eluted between 0.1 15 M and 0.125 M potassium phos- phate. The fractions with the highest activity were pooled.
Sephacryl S 300 high-resolution Chromatography. A 1.9 cm X 9 0 cm Sephacryl S 300 HR column previously equilibrated with buffer A was loaded with 4.9 ml concentrated CPS obtained from the hydroxyapatite column. The elution was performed with 140 ml of the same buffer and 1-ml fractions were collected at a flow rate of 0.17 ml . min-’. The enzyme was eluted in fractions 56-64.
This purification procedure is summarized in Table 2. The overall purification was 166-fold. The specific activity of the final preparation was 31.5 U . mg-’ at 37°C.

192 Purcarea et al. ( E m J. Biochem. 236)
Table 2. Purification of R abyssi CPS. At each step the CPS activity was measured at 37°C under the conditions indicated in the Materials and Methods section.
Step Vol- Pro- Activ- Specific Yield Purifi- ume tein ity activity cation
ml mg U U mg I % -fold
Cell-free extract 15.0 167.25 32.42 0.19 100 -
Q Sepharose Fast Flow 24.0 7.49 22.96 3.06 70.8 16 DEAE Aff-Gel Blue 5.1 1.31 8.92 6.80 27.5 36 Hydroxyapatite 4.9 0.32 8.87 27.63 27.3 145 Sephacryl S 300 HR 8.8 0.27 8.65 31.45 26.7 166
101, . . . ' . " ' ' . " ' I ' ' , " " ' . I " " ' 0 10 20 30 40 50 60
migration (mm)
100 , , , , I . , ' . I , " ' I " ' ' I ' . . ' I , ' ' .
BSA C
Ovdburnin
10 80 90 100 110 120 130 140
Elution volume (g)
Fig. Analysis of P abyssi CPS by SDSPAGE and chromatogra y...y. SDS/PAGE analysis of the purified CPS was performed as described i n the Materials and Methods section in the presence of molecular mass markers. (A) Photograph of the gel. (B) Plot of molecular mass against the migration distance. (C) The same preparation was analysed by size- exclusion chromatography as described in the Materials and Methods section.
Molecular mass and polypeptide composition. The final CPS preparation showed a single protein band when analyzed by gel electrophoresis under non-denaturing conditions. It was verified (see Materials and Methods section) that the CPS activity is as- sociated with this band. The SDS/PAGE analysis shows also a single band (Fig. 1A). The comparison of its rate of migration with that of marker proteins indicates a molecular mass of 46 600 i 1500 Da (Fig. 1 B).
1.2 1 ' ' """I ' """'I ' ' """I . ' "'7 t j
103 104 0 10 10' 102
LAP, A/AP, A1 (PM) Fig. 2. Influence of Ap5A and Ap,A on R abyssi CPS. The CPS activity was measured at 37°C as described in the Materials and Methods scction in the presence of 65 pM Mg ATP and in the presence or abseiice of increasing concentrations of ApsA (A) or Ap,A (0).
Size-exclusion chromatography on a Sepharose 6B column calibrated with proteins of 13.4-240 kDa was used to estimate the molecular mass of the native form of the enzyme in an at- tempt to determine its oligomeric structure [19-211. The result is shown in Fig. 1C. On the assumption that P abyssi CPS i s a globular protein, its estimated molecular mass is 50700 2 800 Da. The presence of saturating concentrations of the three substrates of the enzyme in both the sample and the elution buffer did not alter this approximated value. Taken together, these results suggest that CPS from P. abyssi is a monomer.
Carbamoyl-phosphate synthetase or carbamate kiriase? Some organisms like Pseudomonas aeruginosa [32, 331, Myo- plasma lzorninis [34], Streptococcus ,faecium [35, 361, and Strep- tococcus faecalis [37] are able to use arginine as an energy source through the arginine deiminase pathway [38] whose third step is catalyzed by carbamate kinase, an enzyme which pro- duces ATP and carbamate using carbamoyl phosphate and ADP. The equilibrium constant of this reaction is such that it can pro- ceed towards the production of carbamoyl phosphate in the pres- ence of ammonium, bicarbonate, and ATP. Since the enzyme that catalyzes the production of carbamoyl phosphate in F! abyssi shows properties that are common to CPS and carbamate kinases, some experiments were specifically devised to further identify this P. abyssi enzyme.
hhibition by Ap.4. Ap,A is a potent inhibitor of E. coli CPS that binds simultaneously to the two ATP sites of this enzyme. The potency of the inhibition of this compound is probahly due to the distance between the two adenosine rings equaling the distance between the two ATP-binding sites of the enzyme. In contrast, AplA has no significant effect [25]. Since carhamate kinases have only one ATP site, such a large difference in the influence of these two compounds should not he observed in this case. Consequently, the activity of the P abyssi enzyme was measured in the presence of increasing concentrations of the two chemicals. Ap,A produces 50% inhibition at a concentration of 3 pM (Fig. 2). At 0.1 mM, this compound inhibits the I? abyssi enzyme activity by 92% whereas, at the same concentration, the inhibition by Ap,A is only 5 %.
Preincubation of ammonium chloride and bicarboniite for the production of carbamate. At 37"C, ammonium chloride and bicarbonate produce spontaneously carbamate with a chemical equilibrium constant of [NH,][HCO;]/[NH2CO0 1 = 0.33 [37]. Consequently, the initial rate of the carbamate kinase reaction should be enhanced by preincubation of ammonium chloride and bicarbonate through accumulation of the substrate carbarnate. To test this possibility, these two substrates were incubated together

Purcarea et al. ( E m J. Biochem. 236) 193
0.10 -
0.05
0.00 L
0 10 20 30 40 SO 60 70 Incubation time (min)
Fig. 3. Stoichiometry of ATP consumption and carbamoyl phosphate production in the reaction catalyzed by the P. abyssi enzyme. Under identical conditions of enzyme and substrate concentrations, the rates of ATP consumption and carbamoyl phosphate (CP) production wttre measured as indicated in the Materials and Methods section. (0) ATP consumption in the overall reaction; (0) carbamoyl phosphate produc- tion in the overall reaction; (A) ATP consumption in the bicarbonate- dependent adenosine triphosphate reaction performed in the absence of ammonium chloride.
for 30 min at 37 "C before starting the enzymatic reaction. This procedure did not change the initial velocity of the reactiom.
Staichiometry and mechanism of the reaction catulyzed by the P. abyssi enzyme. The reaction catalyzed by CPS comprises three steps [39]:
MgATP + HCO, * -0COOPO:- + MgADP, (1)
-0COOPO;- + Gln/NH,I + H,O 5 NH,COO- + Pi + Glu, (2)
NH,COO- + MgATP * NH,COOPO:- + MgADP. (3)
The stoichiometry of the overall reaction is 2 mol ATP con- sumed for 1 mol carbamoyl phosphate synthesized. In contrast, carbamate kinase catalyzed only the phosphorylation of carba- mate, a reaction similar to the third step of the CPS reaction. Consequently, in this case the stoichiometry is 1 mol ATP con- sumed for 1 mol carbamoyl phosphate synthesized.
To further identify the nature of the enzyme that catalyzes the synthesis of carbamoyl phosphate in P. abyssi, the stoichiom- etry of this reaction was determined by comparing the rates of production of ADP and carbamoyl phosphate. Under the same conditions of CPS and substrate concentrations, ADP production was measured using a coupled assay with pyruvate kinase and lactate dehydrogenase, and carbamoyl phosphate production was measured using a coupled assay with aspartate transcarbamo- ylase, as described in the Materials and Methods section.
The results of the kinetic experiments presented in Fig. 3 show unambiguously that the stoichiometry of the overall reac- tion is 2 mol ATP consumed/mol carbamoyl phosphate formed.
Furthermore, Fig. 3 shows directly the existence of the bicar- bonate-dependent ATPase reaction (first partial reaction) in the absence of ammonium chloride, a reaction specific of CPS en- zymes. Under the same conditions of enzyme and substrate con- centrations, the rate of ATP consumption corresponds to about 50% that of the overall reaction. It was verified that this ATPase reaction is bicarbonate dependent; in the absence of this com- pound, the reaction was reduced by 94%. In the case of the E. coli CPS, this partial reaction is specifically inhibited by adeno- sine 5'-[a$-methyleneltriphosphate [40]. At a concentration of 5 mM, this compound decreases the rate of reaction by 78%.
Under the same conditions and in the presence of a normalized concentration of ATP, the l? abyssi enzyme is inhibited by 75 %.
Taken together, these results lead to the conclusion that the single enLyme that catalyzes the production of carbamoyl phos- phate in l? abyssi is a carbamoyl-phosphate synthetase and not a carbamate kinase.
Substrate saturation curves. The l? abyssi optimal temperature for growth is 96°C [3]. Such a high temperature could not be used for the determination of the kinetic parameters of the puri- fied CPS enzyme of this organism, due to the thermal instability of its substrates and of the isolated catalytic subunits of E. coli ATC used in the coupled assay. Furthermore, at temperatures higher than 70 "C the unstable product carbamoyl phosphate is decomposed more rapidly than it is used by ATC. One of its degradation products, cyanate [4], reacts with the amino group of aspartate leading to very high control values in the absence of ATC. Consequently, the highest temperature that could be used is 70°C. At this temperature, the reaction rate is constant for only 2 min whereas it is constant for more than 40 min at 37 "C. Moreover, previous work showed that the regulatory properties of l? abyssi ATC [lo] vary when determined at low and high temperature, a phenomenon which was also observed in the case of CPS from E. coli and phosphofructokinase from Bacillus steurothermophilus [41]. Consequently, in the present work most of the kinetic experiments were performed at both 37°C and 70°C.
Ammonium chloride. The results reported above showed that t? abyssi CPS uses ammonium salts as the donor of the amino group. Fig. 4A shows the NH,CI saturation curve of the enzyme at 70°C. The corresponding Eadie plot (Fig. 4A, insert) indi- cates the existence of apparent negative cooperativity in the utilization of this substrate, a phenomenon which was also ob- served at 37°C. The fit of this saturation curve to the Hill equa- tion provides values of So, and n,, of 33 -C 4 mM and 0.53 -C 0.08, respectively. Similar values were obtained at 37°C (Table 3). Such a low affinity implies that the saturation curves for the other two substrates could not be established in the presence of saturating concentrations of NH,Cl. Therefore, the saturation curves for sodium bicarbonate and Mg ATP were determined at three different non-saturating concentrations of NH,CI. 30, 80, and 150 mM, and their K,, or S,,, values were estimated through intercept replots.
Sodium bicarbonate. The sodium bicarbonate saturation curves of the enzyme at 70°C are shown in Fig.4B. The K,,, values obtained at this temperature and at 37°C are 7.2 2 0.6 mM and 2.3 2 0.1 mM, respectively (Table 3).
MgAT?! Previously studied CPSs from other organisms re- quire MgATP as substrate [11, 421. Consequently, the saturation of P. abyssi CPS by this complex was established. Fig. 4 C shows the result obtained at 70°C. This saturation curve shows apparent cooperativity in the utilization of MgATP. This inter- pretation is confirmed by the Eadie plot (Fig. 4C, insert) which provides a nH value of 1.420.1 and a S,,, of 8 8 2 1 pM. At 37"C, the values of these parameters are 1.1 2 0 . 1 and 84 ? 12 pM, respectively (Table 3). An excess of either ATP or Mg2+ over the stoichiometric presence of these two compounds provokes a decrease in the rate of the reaction.
The maximal velocity of the enzyme in the presence of satu- rating concentrations of the three substrates was 34.8 2 0.2 U . mg-l and 643 -C 11 U . mg-' at 37°C and 70°C, respectively.
Lack of potussium chloride requirement for inuximul activity. In several organisms, the reaction catalyzed by CPS requires the presence of K'. Such is the case for the E. coli enzyme [43], S. cerevisiue CPS-P [44], and the mouse spleen enzyme [45], which require a high concentration of KCI for maximal activity.
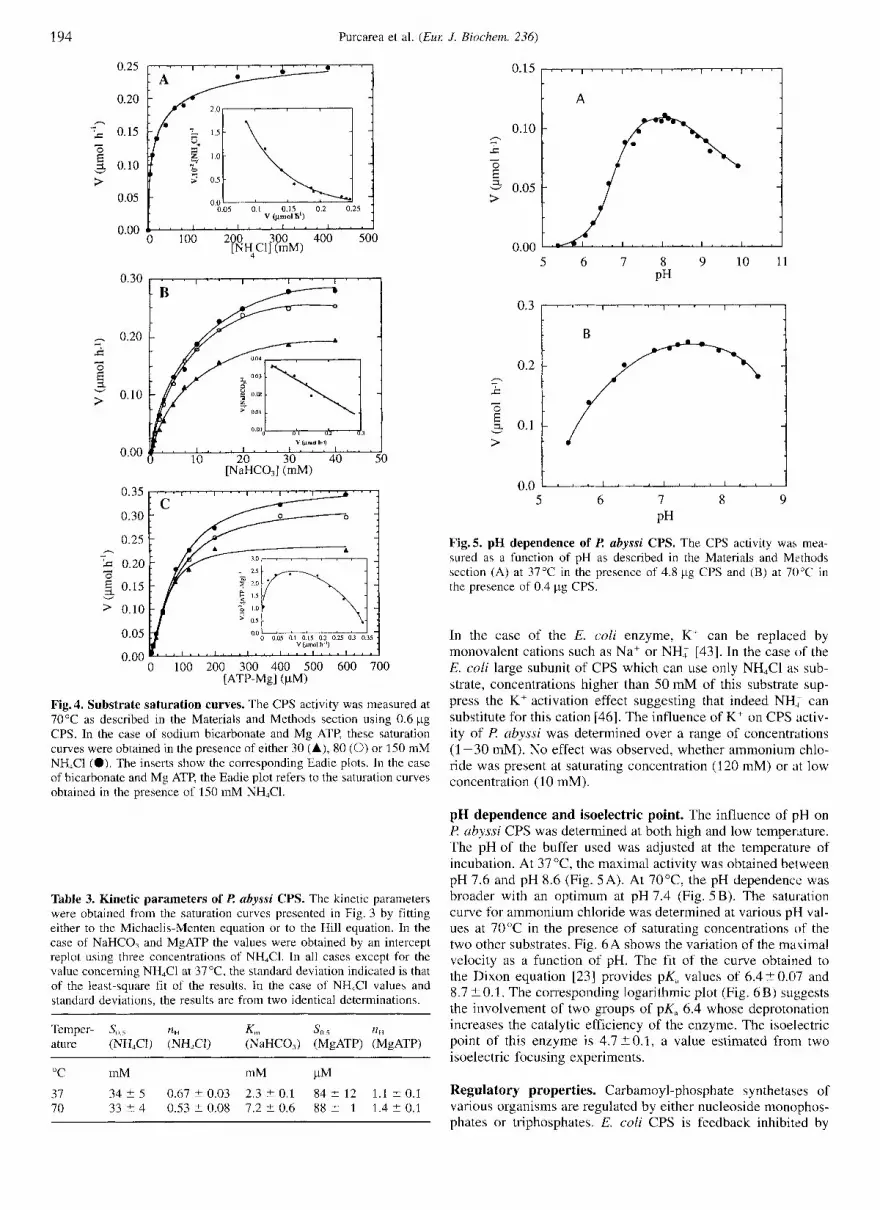
194 Purcarea et al. ( E m J. Biochem. 236)
0.25
0.20
7z 0.15
3. 0.10 >
0.05
0.00
h
c
5 v
0 100 200 300 400 500 [NH4C11 (mM)
0.20 - c 0
E, ; 0.10
0.00
0.35
0.30
0.25 h - 5 0.20 0
0.15
> 0.10 v
V.""
0 100 200 300 400 500 600 700 [ATP-Mgl (I.W
Fig.4. Substrate saturation curves. The CPS activity was measured at 70°C as described in the Materials and Methods section using 0.6 pg CPS. In the case of sodium bicarbonate and Mg ATP, these saturation curves were obtained in the presence of either 30 (A), 80 (0) or 150 mM NH,C1 (0). The inserts show the corresponding Eadie plots. In the case of bicarbonate and Mg ATP, the Eadie plot refers to the saturation curves obtained in the presence of 150 mM NH,Cl.
Table 3. Kinetic parameters of Z? abyssi CPS. The kinetic parameters were obtained from the saturation curves presented in Fig. 3 by fitting either to the Michaelis-Menten equation or to the Hill equation. In the case of NaHCO, and MgATP the values were obtained by an intercept replot using three concentrations of NH,Cl. In all cases except for the value concerning NH,CI at 3 7 T , the standard deviation indicated is that of the least-square fit of the results. In the case of NH,CI values and standard deviations, the results are from two identical determinations.
Temper- S,, , nH K,,, so 5 nH
"C mM mM PM
ature (NH,Cl) (NH,Cl) (NaHCO,) (MgATP) (MgATP)
37 34 t 5 0.67 ? 0.03 2 3 f 0.1 84 t 12 1.1 i 0.1 70 33 2 4 0.53 ? 0.08 7.2 2 0.6 88 5 1 1.4 f 0.1
0.10 h - b d
0
E, - 0.05 >
0.00 5 6 7 8 9 1 0 1 1
PH
0.0 5 6 7 8 9
PH
Fig.5. pH dependence of R abyssi CPS. The CPS activity was mea- sured as a function of pH as described in the Materials and Mcthods section (A) at 37°C in the presence of 4.8 pg CPS and (B) at 7(1°C in the presence of 0.4 pg CPS.
In the case of the E. coli enzyme, K' can be replaced by monovalent cations such as Na' or NH: [43]. In the case of the E. coli large subunit of CPS which can use only NH,C1 as sub- strate, concentrations higher than 50 mM of this substrate sup- press the K' activation effect suggesting that indeed NH, can substitute for this cation [46]. The influence of K' on CPS activ- ity o f f ? abyssi was determined over a range of concentrations (1 -30 mM). No effect was observed, whether ammonium chlo- ride was present at saturating concentration (120 mM) or at low concentration (10 mM).
pH dependence and isoelectric point. The influence of pH on f? abyssi CPS was determined at both high and low temperature. The pH of the buffer used was adjusted at the temperature of incubation. At 37"C, the maximal activity was obtained between pH 7.6 and pH 8.6 (Fig. 5A). At 70"C, the pH dependence was broader with an optimum at pH 7.4 (Fig. 5B). The saturation curve for ammonium chloride was determined at various pH val- ues at 70°C in the presence of saturating concentrations of the two other substrates. Fig. 6A shows the variation of the mavimal velocity as a function of pH. The fit of the curve obtained to the Dixon equation [23] provides pK, values of 6.420.07 and 8.7 i- 0.1. The corresponding logarithmic plot (Fig. 6B) suggests the involvement of two groups of pK, 6.4 whose deprotonation increases the catalytic efficiency of the enzyme. The isoelectric point of this enzyme is 4.720.1, a value estimated from two isoelectric focusing experiments.
Regulatory properties. Carbamoyl-phosphate synthetaszs of various organisms are regulated by either nucleoside monophos- phates or triphosphates. E. coli CPS is feedback inhibited by

Purcarea et al. (Eur: .I. Biochem. 236) 195
800
600
400
200
n
l " " " ' " " " " ~ A
5 6 7 8 9 PH
l B L
3 -
2 -
5 6 7 8 9 PH
Fig. 6. Effect of pH on the V,,, of R abyssi CPS. The saturation curves using ammonium chloride were determined at 70°C at different pH val- ues in the presence of saturating concentrations of the two complenien- tary substrates as described in the Materials and Methods section. (A) Variation of. V,,,, as a function of pH. (B) Variation of log(V,,,,) as a function of pH. The dotted lines indicate the tangents with the slopes of 0 and +2. Their intersection indicates a pK', value of 6.4.
UMP and stimulated by IMP and ornithine [47]. The Saccharo- myces cerevisiae CPS-P is feedback inhibited by UTP [44] while the CPS-A is unaffected by this compound [48]. The mammalian CPS-P, which is p&t of the multifunctional protein CAD, is feedback inhibited by UTP and stimulated by PPRibP [49]. In view of this diversity of regulatory properties, the influences on the activity of l? abyssi CPS of both nucleoside monophosphates and nucleoside triphosphates as well as ornithine and PPRibP were investigated.
The influence of a series of nucleotides on the enzyme activ- ity at a concentration of 1 mM at both 37°C and 70°C is shown
100
10-6 10-5 10-4 10-3 10-2 "TPI (MI
Fig. 8. Inhibition by nucleoside triphosphates. The CPS activity was measured at 37°C in the presence of 65 pM Mg ATP with increasing concentrations of CTP (O), UTP (O), TTP (A), ITP (+) and XTP (A). The inhibition was calculated as described in the Materials and Methods section.
in Fig. 7. Among the nucleoside monophosphates, AMP is the only one to have an inhibitory effect on the rate of the CPS reaction. No effect was observed in the cases of UMP, CMP, TMP, IMP, and XMP. As far as the ribonucleoside triphosphates are concerned, UTP, CTP, TTP, ITP, and XTP have a strong inhibitory effect. GTP has no effect on this activity. In general, this inribition is more pronounced at 37"C, an observation which is probably linked to the negative influence that high tem- perature has on ionic interactions. Among the deoxyribonucleo- side triphosphates, only dCTP and dGTP have a significant in- fluence on P. ahyssi CPS activity. It was also verified that ornith- ine does not affect the activity of P. abyssi CPS at both high and low temperatures. No activation by PPRibP could be detected up to a concentration of 1 mM. On the basis of these results, the inhibition by the more efficient ribonucleoside triphosphates was further investigated. Fig. 8 shows the variation of the enzyme inhibition as a function of nucleotide concentration at 37°C. The five ribonucleoside triphosphates tested, UTP, CTP, TTP, TTP, and XTP, show the same dosehesponse curve, 50% inhibition being obtained at a concentration of about 0.3 mM. At 70"C, the same result was obtained in the cases of CTP, TTP, ITP, and XTP except that the mid-point of the curve was shifted to about 1 mM. Under the same conditions, the inhibition by UTP is lower, with a midpoint at about 3 mM. To determine whether these inhibitors act in synergy or if their effects are additive or antagonistic, the influence of UTP in the presence of CTP on the CPS activity was investigated. Using the criteria of Webb for the calculation of the inhibitory effect of non-saturating con-
Fig.7. Influence of nucleotide effectors on the activity of P. abyssi CPS. The CPS activity was measured as described in the Materials and Methods section in the presence of 65 pM Mg ATP and of 1 mM various nucleotides at both 37°C (black) and 70°C (grey).

196 Purcarea et al. ( E m J. Biochern. 236)
1.21 ' ' ' I " ' I " ' I ' " I " ' I
-0.01 0 0.01 0.02 0.03 0.04
l/[ATP-Mg] (pM-l)
Fig.9. Saturation cnrves for Mg ATP in the presence and absence of UTP (double reciprocal plot). The CPS activity was determined at 37°C as indicated in the Materials and Methods section in the presence (0) or absence (0) of 0.1 mM UTP using 0.5 pg of CPS.
centrations of these two effectors either alone or in combination [50], it was found that in the case of P. abyssi CPS the effects of CTP and UTP are simply additive.
In the case of CPS from E. coli and the hamster CAD pro- tein, the inhibition by UMP and UTP, respectively, is the result of a decrease of the enzyme affinity for its substrate Mg ATP [47, 51, 521. In the present case. the influence of 0.1 mM UTP on the Mg ATP saturation curve of the enzyme was determined. This nucleotide acts as an apparent competitive inhibitor towards Mg ATP, thus affecting only the affinity of the enzyme for this substrate (Fig. 9).
Influence of temperature. It is a general observation that car- bamoyl-phosphate synthetases are very heat-labile enzymes. Furthermore, their activity leads to the synthesis of carbamoyl phosphate which is an unstable metabolite. Some of the sub- strates of this enzyme are also unstable compounds. This en- zyme instability raises some questions as to its activity in P. abyssi, a microorganism whose optimal temperature for growth is 96°C. Consequently, in the present study, the influence of temperature on the stability and activity of this purified enzyme was investigated.
Thermal stability of P abyssi CPS. A P abyssi CPS sample was incubated at 95°C for 8 h. At different time intervals, sam- ples were taken and their activity was measured at 37°C under the standard conditions. For comparison, a parallel similar ex- periment was performed with an E. coli cell-free extract contain- ing the CPS of this mesophilic organism. The results presented in Fig. 10 show that although the activity of P. abyssi CPS decreases exponentially with time under these conditions it still retains 50% of its original activity after 3 h of preincubation at 95°C. Under these conditions, E. coli CPS is completely inacti- vated in less than 2 min. This impressive stability of P. abyssi CPS relates to the fact that this enzyme could be stored at 4°C for at least six months without decrease of its specific activity.
Influence of temperature on the reaction rate. The influence of temperature on the rate of the reaction catalyzed by this heat- stable CPS was investigated. For this purpose, ammonium chlo- ride saturation curves were determined at various temperatures i n the presence of saturating concentrations of the two other substrates, in order to obtain the maximal velocity. The values obtained over 70°C are less accurate due to the fact that above this temperature the rate of reaction was linear for only 1.5 min. Fig. 11 shows the remarkable increase of this maximal rate of reaction between 35 "C and 95 "C. The corresponding Arrhenius
0.8 -
0.6 -
0.4 -
0.2 -
0 . 0 L . 5 ' = " " " " " " " 0 2 4 6 8 10
Preincubation time (h)
Fig. 10. Thermostability off! abyssi CPS. F? nbyssi (0) CPS (16.7 pg in 250 p1) and E. coli (0) cell-free extract samples were preincubated at 95°C. At different time intervals, samples of 10 p1 and 100 pl, respec- tively, were taken and their activity was measured at 37°C under the standard conditions.
30 40 50 60 70 80 90 100 incubation temperature ("C)
Fig. 11. Influence of temperature on the maximal velocity of the re- action catalyzed by f! abyssi CPS. Ammonium chloride saturation curves of P ahyssi CPS were determined at different temperatures in the presence oi saturating concentrations of the two complementar) sub- strates under the standard conditions. The maximal velocities were cal- culated from the Hill equation. The insert shows the correspcdng Arrheniiis plot.
plot (Fig. I 1, insert) indicates an activation energy (Ed) of 77 kJ . mol-'. The values of the activation enthalpy ( A H " ) and activa- tion entropy ( A S " ) calculated from the plot of In(k/T) as a func- tion of 1/T are 74.5 kJ . mol-' and 19.1 kJ . mol-' . K-I, rcspec- tively. These values are of the same order of magnitude as those reported for different kinds of enzymes from mesophilic cirgan- isms.
Influence of hydrostatic pressure. Since P. abyssi is a barotol- erant to barophilic microorganism living under a hydrcistatic pressure of 20 MPa [3] , the influence of this parameter cm the stability and reaction rate of CPS from this organism was inves- tigated.
Enzyme stability under pressure. The enzyme stability was determined under pressure up to 300 MPa. After incubation of the enzyme for 15 min at increasing pressures at 36"C, samples were taken and their activity was measured under standard con- ditions at atmospheric pressure. Fig. 12A shows a progressive inactivation down to a constant value which corresponds to 50 7c of the initial CPS activity. This value is attained at 200 MPa. A complementary experiment showed that this residual activity is constant at 200 MPa for at least 50 min. This effect is irrevers-

Purcarea et al. ( E m J . Biochrrn. 236) 197
x 1.00 Y -rr
+ 0.50
2 0.25
0
KJ
2 .3 Y
2 .C Y
i
0.00 - 0 50 100 150 200 250 300 350
pressure (MPa)
" i o.2 t 1 0.0 ' I I I I I I I
0 20 40 60 80 100 120 140 Preincubation time (min)
Fig. 12. Stability of I? abyssi CPS under pressure. R ahyssi CPS sam- ples were preincubated (A) for 15 rnin at increasing pressures and (B) for increasing periods of time at 50 MPa and their activity was immediately measured under standard conditions, at atmospheric pressure.
ible even in the presence of a reducing agent, indicating that this partial inactivation is not due to pressure-induced oxidation. A similar effect was observed when the exposure to pressure was performed at 50 MPa for different periods of time, up to 135 min (Fig. 12B).
Influence of pressure on the CPS reaction rate. The influ- ence of pressure on the rate of the reaction catalyzed by F! ubyssi CPS was investigated. For this purpose, the rate of the catalytic reaction was determined as a function of pressure. The kinetics of the reaction were followed for 1 h at increasing pressure val- ues up to 200 MPa, as described in the Materials and Methods section. As in the case of the ATC from F! abyssi [lo], the pres- sure has a negative effect on the CPS reaction rate, indicating that this catalytic process involves a positive activation volume (Fig. 13). The calculation of this A V" from the slope of the In( V) versus pressure plot provides a value of 18.1 f O . l ml . mol-I.
DISCUSSION
The results presented above show that the recently discov- ered deep-sea hyperthermophilic archaebacterium I? abyssi pos- sesses a single carbamoyl-phosphate synthetase which utilizes ammonium salts but not glutamine as nitrogen donor. The stoi- chiometry of the reaction, the bicarbonate-dependent adenosine triphosphate activity of the enzyme, the inhibition of this partial reaction by adenosine 5'-[n,P-methylene] triphosphate, the spe- cific inhibition of the overall reaction by ApsA but not Ap3A, all indicate that this F! abyssi carbamoyl phosphate synthesizing enzyme is indeed a carbamoyl-phosphate synthetase and not a carbarnate kinase. Analysis of the purified enzyme by SDS/ PAGE indicates a molecular mass of 46 600 t 1500 Da. Under non-denaturating conditions, analysis by size-exclusion chroma- tography provides a molecular mass of about 50 kDa suggesting that this enzyme is monomeric. The presence of saturating con-
2.5 [ I I I I I
A W 2.0 18.1M.1 ml.mol-1
? v
5 1.5 -
1.0 -
5 0 50 100 150 200 250
pressure (MPa)
0.5
Fig. 13. Influence of pressure on the rate of the reaction catalyzed by I? abyssi CPS. The CPS reaction rate was measured as a function of pressure as indicated in the Materials and Methods section. At each pres- sure, the incubation medium contained 0.35 pg enzyme and saturating concentrations of the substrates (120 mM ammonium chloride, 100 mM sodium bicarbonate, 0.75 mM Mg ATP and 30 rnM aspartate) in the presence of 3.7 pg E. coli ATC catalytic subunits. The reaction was fol- lowed for 60 min taking a sample after 5 min and subsequently every other 5 rnin for the determination of the amount of carbamoyl aspartate formed. These incubations were performed at 33 "C.
centrations of the three substrates did not alter the elution profile in the size-exclusion chromatography experiments. In spite of this monomeric nature, CPS from F! abyssi shows slight appar- ent positive cooperativity for Mg ATP. This phenomenon most probably relates to the fact that in the case of the CPS known so far, two molecules of this substrate bind to different subsites at different steps of the reaction. Most probably, such a mecha- nism applies to the I? abyssi enzyme since its activity is also strongly inhibited by the bisubstrate analog Ap5A, but not by Ap.A. The NH,CI saturation curve indicates that the S,,, of the enzyme for this substrate is high (33 mM) and it shows some apparent negative cooperativity. This phenomenon was also re- ported in the case of the glutamine saturation curve of S. cue- visiae CPS-A [48] and its significance is not clear, although it could be related to the instability of this compound. In contrast with glutamine-dependent CPS enzymes, the FI abyssi enzyme does not require KCI. even in presence of a low concentration of NH,Cl. The pH dependence of the reaction catalyzed by I? ubyssi CPS shows an important increase of the maximal velocity between pH 6 and pH 7 at 70°C which suggests that the deproto- nation of two groups is responsible for this increase of catalytic efficiency [53].
I? abyssi CPS activity is inhibited by a series of nucleotides. However, the pattern of regulatory effects differs from that ob- served in previously described CPS. All the pyrimidine nucleo- side triphosphates as well as ITP and XTP, but not GTP, are strong inhibitors with very similar K, values (about 0.2 mM). It is known that eukaryotic CPS-P enzymes are feedback inhibited by UTP [44, 541. Nucleoside monophosphates are inactive to- ward F! abyssi CPS except for AMP. This inhibition pattern is also different from that of the carbamate kinase from FI uerugi- nma which is not sensitive to UTP, CTP, and AMP 1321. Con- trary to what is observed in the case of E. coli CPS and the mammalian CAD protein, no activator of the P. uby could be identified. For instance, IMP and ornithine activate E. coli CPS [46], PPRibP activates the mammalian CAD CPS [48], the CPS from Salmonella typhirnuriurn [24] and that from plants like Spinaciu oleraceu [55], Vinca rosea [56], Pisum sativum [7], Phaseolus aureus [8] and N-acetylglutamate activates the ammonium-dependent CPS [28]. None of these compounds has an effect on the activity of the I? abyssi enzyme. Contrary to

198 Purcarea et al. (Eur J . Biochem. 236)
what has been observed in the cases of ATC enzymes from the same organism [ lo] and of E. coli CPS and phosphofructokinase from Bacillus stearothermophilus [41], the nucleotide inhibitory effects are not altered at high temperature. As in the cases of known CPS enzymes in which nucleotides bind to specific allo- steric sites [57, 581, the feedback inhibitor UTP affects the S,,, for Mg ATP of the P ubyssi enzyme, suggesting that this nucleo- tide might bind to a specific regulatory site. However, since vari- ous nucleoside triphosphates inhibit the enzyme with about the same Kc, a direct competitive binding of these nucleotides to the catalytic site cannot be excluded.
The most striking property of F! abyssi CPS is its thermosta- bility as compared to the homologous enzymes known so far. After 3 h of incubation at 95"C, this enzyme still retains 50% of its original activity. Under the same conditions, E. coli CPS is completely inactivated in less than 2 min. Since the optimal temperature for growth of F! abyssi is 96°C 131, the enzyme might be even more stable in its natural intracellular environ- ment as the result of interactions with other proteins or cellular components. However, its slow inactivation at very high temper- ature might be compensated by continous biosynthesis. As far as the thermodynamics of the reaction are concerned, it is inter- esting to note that the value of the entropy of activation (AS') for this reaction increases with temperature. This observation indicates that the probability of formation of the transition state increases with temperature.
Is P. abyssi CPS one of the previously postulated ancestral forms of CPS? The CPS enzymes known so far contain either a glutamine amidotransferase domain or subunit, which transfers the amide nitrogen of glutamine to the CPS catalytic site. In E. coli, the 117-kDa large CPS subunit is non-covalently associated to a 42-kDa glutamine amidotransferase subunit [9]. The iso- lated large CPS subunit can use NH,' as substrate for the biosyn- thesis of carbamoyl phosphate [53]. In S. cerevisiae. the CPS of the arginine pathway (CPS-A) shows the same organization ; the molecular masses of the glutamine amidotransferase and CPS subunits are 45 kDa and 123 kDa, respectively [S9, 601. In the same microorganism, the two subunits of the pyrimidine path- way CPS enzymes (CPS-P) are covalently linked within a multi- functional protein 161) as they are in mammals [62]. In rat, CPS- A uses NH,' as substrate and is unable to use glutamine. How- ever, it was shown that this property does not result from the lack of the glutamine amidotransferase domain but from its inac- tivation as the result of substitution of an essential cysteine resi- due by serine 1631. It has been reported that the CPSs of two thermophilic eubacteria, Therinotoga muritima and Therinus uquaticus, use glutamine as substrate, suggesting that they also possess a glutamine amidotransferase subunit IS].
As far as the CPS subunit or domain is concerned, the amino acid sequence of all the enzymes analyzed so far shows a signifi- cant internal similarity between the N-terminal region and the C-terminal region. This is the case for E. coli CPS 1641, S. cere- visiae CPS specific for both the pyrimidine and arginine path- ways [59, 61 ] and the similar mammalian enzymes [62, 631. In E. coli, this similarity between the N and C regions concerns the sequences of residues at positions 1-400 and 553-933, respec- tively, which are separated by a 153-residue sequence, with the best score for internal similarity yet reported [64]. On the basis of these similarities it has been concluded that the present form of CPS genes results from duplication, and differentiation of an ancestral gene followed by fusion 159, 61 -641.
The results reported here show that the archaebacterium P abyss; possesses a single and monomeric CPS of molecular mass 46.6 kDa, which is strictly NH: dependent. It is thus very tempt- ing to speculate that this low-molecular-mass CPS is an ancestral
form whose gene duplication led to the presently known CPS. A gene duplication would produce a protein with a similar mo- lecular mass to the E. coli CPS large subunit when the molelxlar mass of the 153-residue linker is taken into account.
It is noteworthy that all these CPS enzymes from various organisms show significant degrees of similarity [62]. In addi- tion, some organisms such as Pseudomonas aeruginosa ~ 321, Streptococcus faeculis [37], Streptococcus fuecium 1361. and Mycoplasma hominis [34] possess a carbamate kinase m hich uses carbamoyl phosphate to synthesize ATP with formation of NH,' and NH,'. This reaction is reversible and can also lead to the synthesis of carbamoyl phosphate. These dimeric enzymes whose monomer molecular masses are in the range 33-37 kDa show sequence similarity with each half of the CPS [33. 621. Interestingly, this similarity is higher with the C-terminal rr: gion of these CPS enzymes [33]. On the basis of this similarity, i t has been proposed that CPS emerged from the fusion of an ancmral gene of carbamate kinase with genes coding for an ATP-binding site and for the allosteric domain [65]. Therefore, P. abyssi CPS might be the previously postulated ancestral form of CPS, de- rived from a carbamate kinase [33, 641. In the case of CPS, a further step of evolution was the association with the glutamine amidotransferase protein, allowing these enzymes to use gluta- mine as nitrogen donor instead of NH: .
The molecular mass of P ubyssi CPS is about 50% that of other identified CPS enzymes and, under the conditions used, it appears to be monomeric. The high sensitivity of this enzyme to ApsA as compared to Ap,A suggests that this monomeric protein possesses the two ATP-binding sites found in the CPS enzymes of higher molecular mass. Some implications of this observation are currently being investigated. The sequencing of the gene coding for the f? abyssi CPS is underway in order to test these hypotheses.
The authors are indebted to Dr G. Raguenes and Dr G. Birbier, IFREMER, Brest, and to Dr A. Escaut, S. Cortial and D. Joseph, ICSN, CNRS, Gif-sur-Yvette, for their help in the production of large arnounts of l? ahq'xsi cells and to Dr J. Vickrey for stimulating discussions and improvement of this manuscript. This work was supported by the i7eno-e Nationrcl de la Recherche Scientifique, UniversitC Pierre et Marie Curie, Park, and a fellowship from the MinistPre de.r Afaires ErmngZrc;) to C. Purcarea and is included in the doctorate thesis of C. Purcarea, presented on the 29th June, 1995, at the University Paris XI, Paris, France.
REFERENCES 1. Auzende, J. M., Urabe, T., and l'equipage scientifique (19891 c'. R.
Acad. Sri. Paris Serie 111 302, 1787-1795. 2. Erauso, G.. Charbonnier, F., Barbeyron, T., Forterre, P. & Prieur, D.
(1992) C. K . Acad. Sci. Paris Serie I l l 314, 387-393. 3. Erauso, G., Reysenbach, A. L., Godfroy, A,, Meunier, J. R., ('rump,
B., Partensky, F., Baross, J. A., Marteinsson, V., Barbier, G.. Pace. N. R. & Prieur, D. (1993) Arch. Microbiol. 160, 338-340.
4. Allen, C. M. J. & Jones, M. E. (1964) Biochenzistn 3, 1238 -- 1247. 5. Van de Casteele, M., Demarez, M., Legrain, C., Glansdorfi N. &
6. O'Neal, T. D. & Naylor, A. W. (1969) Biochem. J. 113, 271 -279. 7. O'Neal, T. D. & Naylor, A. W. (1976) Plant. Physiol. (Brrhc,.da)
8. Ong, B. L. &Jackson, J. F. (1972) Biochem. J . 129, 583-503. 9. Trotta, P. P., Burt, M. F., Haschenineyer, R. W. & Meister, A (1971)
10. Purcarca, C., Erauso, G., Prieur, D. & HervC, G. (1994) Mic rohiol-
11. Robin, J.-P., Penverne, B. & HervC, G. (1989) ELM J . Biocheijz. 18.3.
12. Gaillard Miran. S., Chang, S . H. & Raushel. F. M. (1991) Biochern-
13. Leger, D. & I-Iervt, G. (1988) Biocherni.wy 27, 4293-4298
PiCrard, A. (1990) J. Gen. Microbiol. 136. 1177-1183.
57. 23-28.
Proc. Natl Acad. Sci. USA 68, 2599-2603.
ogY 140, 1967-1975.
519-528.
i s t ~ 30, 7901 -7907.

Purcarea et al. (Eur: J. Biochem. 236) 199
14. Hui Bon Hoa, G., Hamel, G., Else, A., Weill, G. & Hervt, G. (1990)
15. Prescott, L. M. & Jones, M. E. (1969) Anal. Biochern. 32, 408-
16. Laemmli, U. K. (1970) Nature 227, 680-685. 17. Jovin, T., Chrambach, A. & Naughton, M. A. (1964) Anal. Biochem.
18. Oakley, B. R., Kirsch, D. R. & Morris. N. R. (1980) Anal. Biochem.
19. Andrews, P. (1964) Biochem. J. 91, 222-233. 20. Andrews, P. (1965) Biochem. J. 96, 595-606. 21. Le Maire, M., Chabaud, R. & HervC, G. (1991) in Laboratory guide
to biochemistry, enzymology and protein physical chemistry: a study of aspartate trunscarbamylase, Plenum, New York.
22. Lowry, 0. H., Rosebrough, N. J., Farr, A. L. & Randall, R. J. (1951) J. Biol. Chem. 193, 265-275.
23. Dixon, M. (1953) Biochem. J . 55, 161-170. 24. Abdelal, A. & Ingraham, J. L. (1975) J . Biol. Chem. 250, 4410-
25. Powers, S. G., Griffith, 0. W. & Meister, A. (1977) J. Biol. Chem.
26. Earhart, R. H. & Neil, G. L. (1985) Adv. Enzyme Regul. 24, 179- 205.
27. Trotta, P. P., Pinkus, L. M., Haschemeyer, R. H. & Meister, A. (1974) J. B id . Chem. 249, 492-499.
28. Anderson, P. M. (1980) Science 208, 291 -293. 29. Collins, K. D. & Stark, G. R. (1971) J. Biol. Chem. 246, 6599-
30. Swyryd, E. A., Seaver, S. S. & Stark, G. R. (1974) J. B i d . Chem.
31. Penverne, B. & HervC, G. (1983) Arch. Biochem. Biophys. 225,
32. Abdelal, A. T., Bibb, W. E & Nainan, 0. (1982) J. Bacteriol. 151,
33. Baur, H., Luethi, E., Stalon, V., Mercenier, A. & Haas, D. (1989)
34. Schimke, R. T., Berlin, C. M., Sweeney, E. W. & Carroll, W. R.
35. Bishop, S. H. & Grisolia, S. (1966) Biochim. Biophys. Actu 118,
Anal. Biochern. 187, 258-261.
419.
9, 351 -369.
105, 361-363.
4417.
252, 3558-3560.
6605.
249, 6945-6950.
562-575.
141 1-1419.
Eur: J. Riochem. 179, 53-60.
(1966) J. B id . Chem. 241, 2228-2236.
211-218.
41.
42.
43. 44.
45.
46.
47. 48.
49.
50.
51.
52.
53.
54.
55.
56.
57.
58.
59.
60. 61.
36. Marina, A,, Bravo, J., Fita, I. & Rubio, V. (1994) J. Mol. Biol. 235,
37. Thorne, K. J. I. & Jones, M. E. (1963) J. Biol. Chem. 238, 2992-
38. Abdelal, A. (1979) Annu. Rev. Microbiol. 33, 139-168. 39. Anderson, P. M. & Meister, A. (1965) Biochemistry 4, 2803-2809. 40. Powers, S. G. & Meister, A. (1978a) J. Biol. Chem. 2.53, 1258-
62.
2998. 63.
64.
1265. 65.
1345 - 1347.
Note added in the proof. By the time this manuscript was submitted, it was reported by Legrain et al. (Legrain, C., Demarez, M., Glansdorff, N. & PiCrard, A. (1995) Microbiology 141; 1093-1099) that Pyrococcus furiosus also possesses an ammonium-dependent carbamoyl-phosphate synthetase. The estimated molecular mass of this enzyme (70 kDa) is significantly higher than that of P: abyssi CPSase. A detailed comparison of the structure and properties of these two enzymes will be of consider- able interest.
Braxton, B. L., Tlapak-Simmons, V. L. & Reinhart, G. D. (1994) J.
Raushel, F. M., Rawding, C. J., Anderson, P. M. & Villafranca, J. J.
Anderson, P. M. & Meister, A. (1966) Biochemistg 5, 3157-3163. Belkai'd, M., Penverne, B. & Herve, G. (1988) Arch. Biochem. Bio-
Levine, R. L., Hoogenraad, N. J. & Kretchmer, N. (1971) Biochem-
Rubino, S. D., Nyunoya, H. & Lusty, C. J. (1987) J. Biol. Chern.
Anderson, P. M. & Meister, A. (1966) Biochemistry 5, 3164-3169. Price, C. W., Holwell, J. H. & Abdelal, A. T. H. (1978) J . Gen.
Tatibana, M. & Shigesada, K. (1972) J. Biochem. (Tokyo) 72, 549-
Webb, J. L. (1963) Enzyme and metabolic inhibitors, Academic
Boettcher, B. & Meister, A. (1982) J . Bid. Chem. 257, 13971-
Shaw, S. M. & Carrey, E. A. (1992) Eul: J. Biochern. 207, 957-
Kalman, S. M., Duffield, P. H. & Brzozowski, T. (1966) J. B i d .
Mally, M. I., Grayson, D. R. & Evans, D. R. (1980) J. Biol. Chem.
Shibata, H., Ochiai, H., Sawa, Y. & Miyoshi, S. (1986) Plant Phy-
Kanamori, I., Ashihara, H. & Komamine, A. (1980) Z. Pflanzenphy-
Rubio, V., Cervera, J., Lusty, C. J., Bendala, E. & Britton, H. (1991)
Liu, X., Guy, H. I. & Evans, D. R. (1994) J . Biol. Chern. 269,
Lusty, C. J., Widgren, E. E., Broglie, K. E. & Nyunoya, H. (1983)
Nyunoya, H. & Lusty, C. J. (1984) J. Biol. Chem. 259,9790-9798. Souciet, J.-L., Potier, S., Hubert, J.-C. & Lacroute, F. (1987) Mol.
Simmer, J. P., Kelly, R. E., Rinker, J. A. G., Scully, J. L. & Evans,
Nyunoya, H., Broglie, K. E., Widgren, E. E. & Lusty, C. J. (1985)
Nyunoya, H. & Lusty, C. J. (1983) Proc. Natl Acad. Sci. USA 80,
Rubio, V. (1993) Biochem. SOC. Trans. 21, 198-202.
Bid . Chern. 269, 47-50.
(1979) Biochemistry 18, 5562-5566.
phys. 262, 171 -180.
istry 10, 3694-3699.
262, 4382-4386.
Microbiol. 106, 145-151.
560.
Press, New York.
13 976.
965.
Chem. 241, 1871-1877.
255, 11 372-1 1 380.
siol. (Bethesdu) 80, 126-129.
siol. 96, 7-16.
Biochemistry 30, 1068- 1075.
27 747-27 755.
J. Biol. Chem. 258, 14466-14412.
Gen. Genet. 207, 314-319.
D. R. (1990) J. Biol. Chem. 265, 10395-10402.
J. Biol. Chem. 260, 9346-9356.
4629 - 463 3.