molecular characterization of glucose6phosphate dehydrogenase deficiency in brazil
TRANSCRIPT

1993 81: 2150-2154
DT Chiu, L Zuo, L Chao, E Chen, E Louie, B Lubin, TZ Liu and CS Du new base substitutions in the human G6PD gene(G6PD) deficiency in patients of Chinese descent and identification of Molecular characterization of glucose-6-phosphate dehydrogenase
http://bloodjournal.hematologylibrary.org/site/misc/rights.xhtml#repub_requestsInformation about reproducing this article in parts or in its entirety may be found online at:
http://bloodjournal.hematologylibrary.org/site/misc/rights.xhtml#reprintsInformation about ordering reprints may be found online at:
http://bloodjournal.hematologylibrary.org/site/subscriptions/index.xhtmlInformation about subscriptions and ASH membership may be found online at:
reserved.Copyright 2011 by The American Society of Hematology; all rights900, Washington DC 20036.weekly by the American Society of Hematology, 2021 L St, NW, Suite Blood (print ISSN 0006-4971, online ISSN 1528-0020), is published
For personal use only. by guest on July 21, 2011. bloodjournal.hematologylibrary.orgFrom

Molecular Characterization of Glucose-6-Phosphate Dehydrogenase (G6PD) Deficiency in Patients of Chinese Descent and Identification of New Base
Substitutions in the Human G6PD Gene By D.T.Y. Chiu, L. Zuo, L. Chao, E. Chen, E. Louie, B. Lubin, T.Z. Liu, and C.S. Du
The underlying DNA changes associated with glucose-6- phosphate dehydrogenase (G6PD)-deficient Asians have not been extensively investigated. To fill this gap, we se- quenced the GGPD gene of 43 GGPD-deficient Chinese whose GGPD was well characterized biochemically. DNA samples were obtained from peripheral blood of these in- dividuals for sequencing using a direct polymerase chain reaction (PCR) sequencing procedure. From these 43 sam- ples, we have identified five different types of nucleotide substitutions in the G6PD gene: at cDNA 1388 from G to A (Arg to His); at cDNA 1376 from G to T (Arg to Leu); at cDNA 1024 from C to T (Leu to Phe); at cDNA 392 from G
LUCOSE-6-PHOSPHATE dehydrogenase (G6PD) de- G ficiency is one of the most common enzymopathies, affecting over 200 million people worldwide.',' The deficiency predisposes subjects to neonatal jaundice, drug- or infection- mediated hemolytic crisis, favism, and, less commonly, to chronic nonspherocytic hemolytic anemia.3.4 Clinical and biochemical analyses have identified nearly 400 putative variants of G6PD.'x4 Recent advances in the molecular bi- ology of G6PD suggest that diverse point mutations may cause phenotypic heterogeneity."' Most of the genetic studies of G6PD have been done on samples from African Americans4** and people of Mediterranean ancestry.' The DNA sequence of Asian G6PD variants has not been exten- sively investigated.
There is a regional high frequency (2% to 16%) of G6PD deficiency in Taiwan and southern China.'-" The clinical presentations of the G6PD-deficient subjects of Asian origin appear to be different from those of African Americans with G6PD deficiency. Approximately 20% to 40% of neonatal
From the Chang Gung Medical College, Tao- Yuan, Taiwan, China: Children's Hospital Oakland, Oakland, CA; Genentech. Inc, South San Francisco, CA; San Francisco State University, San Francisco, CA: and Sun Yat Sen University of Medical Sciences, Guangzhou, China.
Submitted July 16, 1992: accepted November 25, 1992. Supported in part by National Institutes of Health Grants No. HL
36255, 20985, and DK 32094 as well as grants from Chang Gung Medical College (NMRP 161 and 197) and the National Science Council of the Republic of China (NSC81-0412-B182-19 and NSC81-
Presented in part at the 33rd Annual Meeting of the American Society of Hematology, Denver, CO, and, in part, as a thesis toward the completion of an advanced degree for L.C. in the Center for Ad- vanced Medical Technology at San Francisco State University, CA.
Address reprint requests to D.T. Y. Chiu, Professor, School ofMed- ical Technology, Chang Gung Medical College, 259 Wen-Hwa 1 Rd, Kwei-San, Tao- Yuan, Taiwan, Republic of China.
The publication costs ofthis article were defrayed in part by page charge payment. This article must therefore be hereby marked "advertisement" in accordance with 18 U.S.C. section 1734 solely to indicate this fact.
0412-Bl82-509).
0 1993 by The American Society of Hematology. 0006-49 71/93/81 08-000S$3.00/0
to T (Gly to Val); at cDNA 95 from A to G (His to Arg). These five nucleotide substitutions account for over 83% of our 43 GGPD-deficient samples and these substitutions have not been reported in non-Asians. The substitutions found at cDNA 392 and cDNA 1024 are new findings. The sub- stitutions at cDNA 1376 and 1388 account for over 50% of the 43 samples examined indicating a high prevalence of these two alleles among GGPD-deficient Chinese. Our findings add support to the notion that diverse point mu- tations may account largely for much of the phenotypic heterogeneity of GGPD deficiency. 0 1993 by The American Society of Hematology.
jaundice is related to G6PD deficiency in Taiwan and Guangzhou (unpublished observations). In contrast, neonatal jaundice has rarely been attributed to G6PD deficiency in the United States. This may be due in part to the fact that most African Americans with G6PD deficiency are asymp- tomatic. To further delineate the relationship between the exact genetic defect and phenotypic expression of G6PD de- ficiency, we sequenced the G6PD gene from 43 G6PD-de- ficient Chinese subjects whose G6PD has been well defined biochemically.
MATERIALS AND METHODS
Samples and other materials. After obtaining informed consent, blood specimens were collected from normal and G6PD-deficient individuals of Chinese origin residing in the United States, Taiwan, or China. Most of our samples were from Guangdong province. Unless stated otherwise, all biochemicals were purchased from Sigma (St. Louis, MO). Taq polymerase and the polymerase chain reaction (PCR) buffer were obtained from Promega (Madison, WI). The DNA Thermal Cycler used was a Perkin Elmer-Cetus (Norwalk, CT) model.
The G6PD activity in fresh red blood cells (RBCs) was quantitatively measured using the kit No. 345-B supplied by Sigma. Physicochemical properties of semipurified RBC G6PD were determined in accordance with the recommenda- tions of the WHO Scientific Group." G6PD variants were defined as previously described by Du et aL9
Genomic DNA was extracted from lymphocyte nuclei by the method of Kunkel et al." Direct PCR sequencing of the G6PD gene in the genomic DNA was carried out as previously described.6 The final volume of the PCR mixture was 50 pL containing 50 mmol/L Tris, pH 9.0,20 mmol/L (NH&S04, 1.5 mmol/L MgQ, 0.2 mmol/L of all four dNTPs, 0.5 to I .O pmol/L of both amplification primers, I U of Taq polymerase and 1 pg of DNA template. From the genomic DNA, a DNA fragment of 4.5 kb covering exons 3 through 13 was amplified by a first-round PCR using primers, P1 L(5'- GCAAGACAGACATGCTTGTGGC) and P 1 R( 5'-GCTGGGCTC- GGGTAGTAGCAGCA). For a higher quality and better yield, a second-round PCR was performed with 2% of the first-round PCR products using internal primers P2L(S'-AGGATGATGTATGT- AGGTCG) and P2R( 5'-AATGTGCAGCTGAGGTCAAT) to gen- erate a 4.2-kb fragment. After 10 minutes of predenaturation at 95"C, 35 cycles ofdenaturation (94"C, 1 minute), annealing (55"C, 1 min- ute) and extension (72"C, 5 minutes) were used for both rounds of PCR. Using unequal amounts of PCR primers (0.25 pmol/L and 25 fimol/L, respectively), asymmetric PCR (APCR) was camed out from a mixture containing 2% of the second-round PCR products to gen-
G6PD activity and characterization.
DNA analysis.
2150 Blood, Vol 8 1, No 8 (April 15). 1993: pp 2 150-21 54
For personal use only. by guest on July 21, 2011. bloodjournal.hematologylibrary.orgFrom
MOLECULAR DEFECTS IN CHINESE G6PD VARIANTS 2151
Table 1. DNA “Variants” of 43 Chinese GGPD-Deficient Samples
DNA “Variant” (cDNA no.) (exon no.)
1388 (exon 12) 1376 (exon 12) 1024 (exon 9) 392 (exon 5)
95 (exon 2) Undefined’
Base Change
G to A G to T C to T G to T A to G -
Predicted Amino Acid Substitution
Arg to His Arg to Leu Leu to Phe Gly to Val His to Arg -
n
11 12 2 3 8 7
%
25.6 27.9
4.6 7.0
18.6 16.2
-
* The exons of these samples could not be completely sequenced.
erate an excess of single-stranded DNA. Three pairs of APCR primers were predominantly used to cover exons 3 through 13 of the G6PD gene. The sequences of these primers have been previously described by Beutler et al.’4
Routinely, 5 pL of the APCR products were electrophoresed on agarose gel for both quality and quantity analysis. APCR products were cleaned by the following steps: After the volume was adjusted to 100 pL with dH20, 50 pL of 7.5 mol/L ammonium acetate and 150 pL of cold 100% ethanol were added, mixed, and incubated at room temperature for 5 minutes; the mixture was centrifuged in an Eppendorf centrifuge for 5 minutes at room temperature. The su- pernatant was removed by aspiration and the pellet was washed with I50 pL Of 70% ethanol. It was then centrifuged for 3 minutes, followed by removal of the ethanol by aspiration. The DNA pellet was dried in a speed vacuum and then resuspended in water to a concentration of 0.5 to I .O pglpL. Usually, 2 pL of the above samples was used for
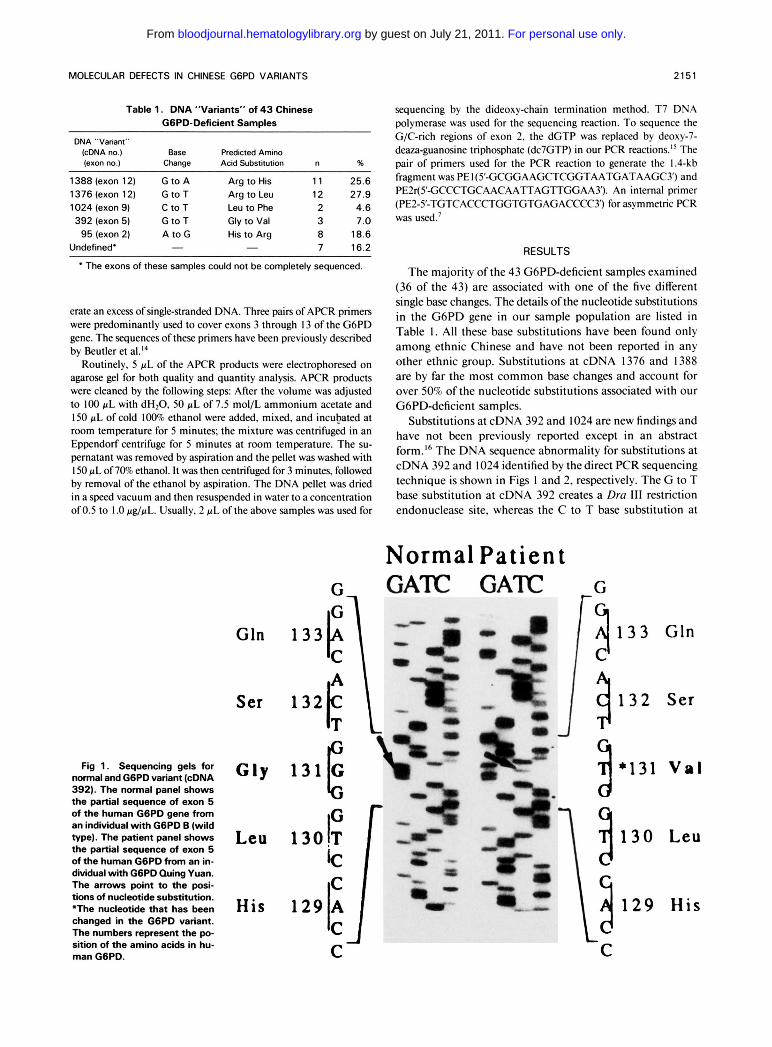
Fig 1. Sequencing gels for m a l and GGPD variant (cDNA 392). The normal panel shows t h e partial sequence of exon 5 of the human G6PD gene from an individual with GGPD B (wild type). The patient panel shows t h e partial sequence of exon 5 of the human G6PD from an in- dividual with GGPD Quing Yuan. The arrows point to the posi- tions of nucleotide substitution. *The nucleotide that has been changed in t h e GGPD variant. The numbers represent the po- sition of the amino acids in hu- man GGPD.
sequencing by the dideoxy-chain termination method. T7 DNA polymerase was used for the sequencing reaction. To sequence the G/C-rich regions of exon 2. the dGTP was replaced by deoxy-7- deaza-guanosine triphosphate (dc7GTP) in our PCR reaction^.'^ The pair of primers used for the PCR reaction to generate the 1.4-kb fragment was PE I (5‘-GCGGAAGCTCGGTAATGATAAGCY) and PE2flS-GCCCTGCAACAATTAGTTGGAA3‘). An internal primer (PE2-S-TGTCACCCTGGTGTGAGACCCC3’) for asymmetric PCR was used.’
RESULTS
T h e majority of the 43 G6PD-deficient samples examined (36 of the 43) are associated with one of the five different single base changes. The details of the nucleotide substitutions in the G6PD gene in our sample population are listed in Table 1. All these base substitutions have been found only among ethnic Chinese and have not been reported in any other ethnic group. Substitutions at cDNA I376 and I388 are by far the most common base changes and account for over 50% of the nucleotide substitutions associated with our G6PD-deficient samples.
Substitutions at cDNA 392 and 1024 are new findings and have not been previously reported except in an abstract form.I6 The DNA sequence abnormality for substitutions at cDNA 392 and 1024 identified by the direct PCR sequencing technique is shown in Figs 1 and 2, respectively. The G to T base substitution a t cDNA 392 creates a Dra 111 restriction endonuclease site, whereas the C to T base substitution a t
For personal use only. by guest on July 21, 2011. bloodjournal.hematologylibrary.orgFrom
2152
Val
TYr
Leu
Val
Val
CHlU ET AL
Normal Patient G rq GATC GATC 1 .)
344[
343 1
3 4 0 E
341
C
\
I
I 3344 1 7 3 4 3 Tyr
T(
+342 P h e 9 7 j 3 4 1 Val
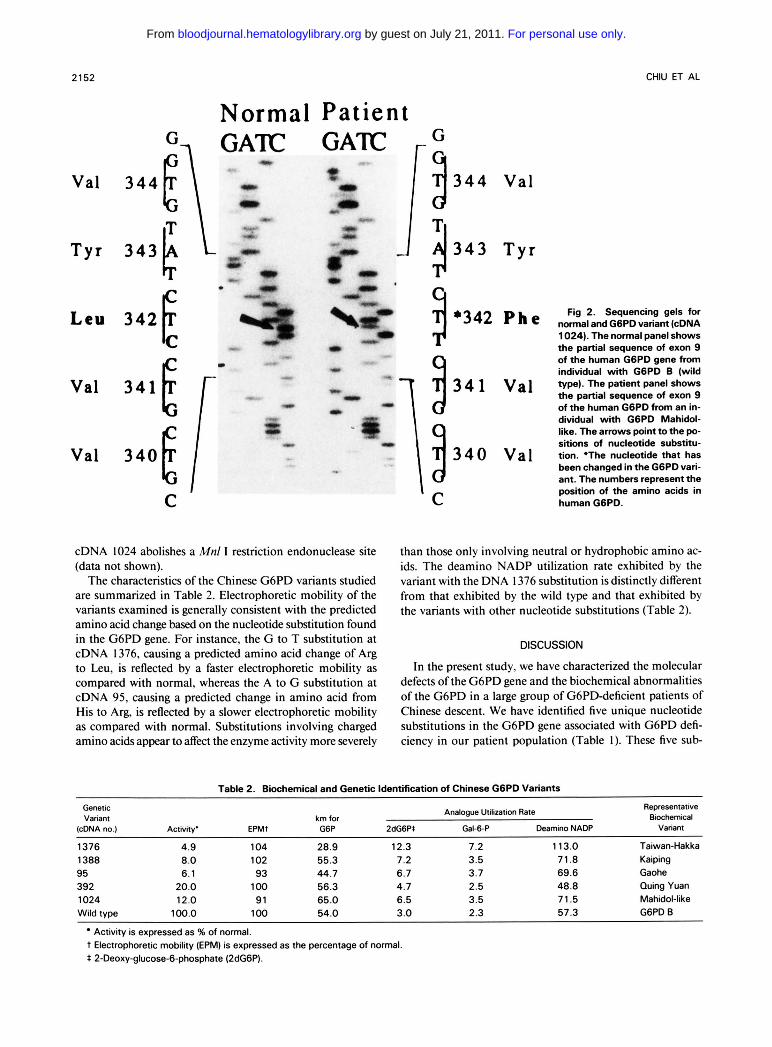
cDNA 1024 abolishes a Mnl I restriction endonuclease site (data not shown).
The characteristics of the Chinese G6PD variants studied are summarized in Table 2. Electrophoretic mobility of the variants examined is generally consistent with the predicted amino acid change based on the nucleotide substitution found in the G6PD gene. For instance, the G to T substitution at cDNA 1376, causing a predicted amino acid change of Arg to Leu, is reflected by a faster electrophoretic mobility as compared with normal, whereas the A to G substitution at cDNA 95, causing a predicted change in amino acid from His to Arg, is reflected by a slower electrophoretic mobility as compared with normal. Substitutions involving charged amino acids appear to affect the enzyme activity more severely
C I
Fig 2. Sequencing gels for normal and G6PD variant (cDNA 1024). The normal panel shows the partial sequence of exon 9 of the human G6PD gene from individual with G6PD B (wild type). The patient panel shows the partial sequence of exon 9 of the human G6PD from an in- dividual with G6PD Mahidol- like. The arrows point to the po- sitions of nucleotide substitu- tion. *The nucleotide that has been changed in the G6PD vari- ant. The numbers represent the position of the amino acids in human G6PD.
than those only involving neutral or hydrophobic amino ac- ids. The deamino NADP utilization rate exhibited by the variant with the DNA 1376 substitution is distinctly different from that exhibited by the wild type and that exhibited by the variants with other nucleotide substitutions (Table 2).
DISCUSSION
In the present study, we have characterized the molecular defects of the G6PD gene and the biochemical abnormalities of the G6PD in a large group of G6PDdeficient patients of Chinese descent. We have identified five unique nucleotide substitutions in the G6PD gene associated with G6PD defi- ciency in our patient population (Table 1). These five sub-
Table 2. Biochemical and Genetic Identification of Chinese G6PD Variants
Representative Analogue Utilization Rate Biochemical
Genetic Variant km for
(cDNA no.) Activity’ EPMt G6P 2dG6P* Gal-6-P Deamino NADP Variant
Taiwan-Hakka 1376 4.9 1 04 28.9 12.3 7.2 113.0 1388 8.0 102 55.3 7.2 3.5 71.8 Kaiping 95 6.1 93 44.7 6.7 3.7 69.6 Gaohe 392 20.0 100 56.3 4.7 2.5 48.8 Quing Yuan 1024 12.0 91 65.0 6.5 3.5 71.5 Mahidol-like Wild type 100.0 100 54.0 3.0 2.3 57.3 G6PD B
Activity is expressed as % of normal. t Electrophoretic mobility (EPM) is expressed as the percentage of normal. * 2-Deoxy-glucose-6-phosphate (2dG6P).
For personal use only. by guest on July 21, 2011. bloodjournal.hematologylibrary.orgFrom

MOLECULAR DEFECTS IN CHINESE GGPD VARIANTS 2153
stitutions have thus far only been found in ethnic Chinese and have not been reported in other ethnic groups.
Substitutions at cDNA 392 and at cDNA 1024 are new findings (Figs 1 and 2). It is interesting to note that both substitutions involve only neutral and hydrophobic amino acids (Table I ) . The substitution at cDNA 392 creates a Dra I11 restriction endonuclease site, whereas the substitution at cDNA 1024 abolishes a Mnl I restriction endonuclease site. These two substitutions account for about 1 1% of the G6PD- deficient samples examined in our study.
Of the nucleotide substitutions identified in our samples (Table I), substitutions at cDNA 1376 and 1388 are the two most common nucleotide changes associated with G6PD de- ficiency, and they have been previously reported by our group6,I7 and others." These two nucleotide substitutions ac- count for over 50% of our G6PD-deficient individuals, sug- gesting that these two alleles are common among Chinese with G6PD deficiency. It is rather interesting that both sub- stitutions are located in exon 12 of the G6PD gene and are only 12 nucleotides apart. Moreover, both substitutions result in a replacement of an arginine residue by other amino acids.
The high frequency of cDNA 1376 and cDNA 1388 sub- stitutions among Chinese with G6PD deficiency (Table 1) may explain the apparent difference in the clinical presen- tations between G6PD-deficient American blacks and G6PD- deficient Chinese. Beutler4 has recently stated that the more severe form of G6PD deficiency is due to mutations clustering near the putative NADP binding site of G6PD, whereas milder forms of G6PD deficiency are mostly caused by mu- tations near the amino end of the enzyme. Although both cDNA 1376 and cDNA 1388 substitutions are not in the immediate vicinity of the putative NADP binding domain,' their close proximity to this region may alter the kinetics of G6PD in such a way that individuals with either of these two variants would be more susceptible to chemical-induced he- molytic crisis. In contrast, the mutations giving rise to the A or the A- variants among African Americans are located far away from the putative NADP binding site toward the amino end. It also should be noted that although the nature of the mutations may be a major determinant of clinical seventy in G6PD deficiency, other factors such as nutrition, environ- ment, and additional genetic factors may play a role in the pathophysiology of G6PD deficiency.17
Aside from a previously reported three-base deletion in exon 2,19 the nucleotide substitution at cDNA 95 is the only base change that has been reported in this exon of the G6PD gene.7 This substitution accounts for almost one fifth of the G6PD-deficient samples examined in our study. Our findings suggest that this allele is quite common among Chinese with G6PD deficiency and that the amino end of G6PD may play some functional role for this enzyme.
Interestingly, we have also found that 4 of the 43 samples examined have a substitution at cDNA I3 1 1 from C to T causing no amino acid change (data not shown). This finding confirms the notion postulated by other i n v e s t i g a t ~ r s ~ @ ~ ~ that this third-position mutation is a worldwide polymorphism in the G6PD gene.
In conclusion, we have found that the nucleotide substi- tutions associated with G6PD deficiency in Chinese are dis-
tinctly different from those associated with G6PD deficiency in other ethnic groups. Most importantly, our findings support the postulate that G6PD deficiency is mainly caused by di- verse point mutation^',^'^ and that this wide spectrum of point mutations may contribute to the heterogeneity in the patho- physiology of this disease.
ACKNOWLEDGMENT
The authors are in debt to Dr T.V. Wang of Chang Gung Medical College, Tao-Yuan, Taiwan, ROC, for his critical review of this manuscript. The authors are also grateful to Jomei Tsai and Peter B.D. Chiu for their excellent preparation of this manuscript.
REFERENCES 1. Luzzatto L, Battistuzzi G: Glucose-6-phosphate dehydrogenase.
Adv Hum Genet 14:217, 1985 2. Beutler E: The genetics of glucose-6-phosphate dehydrogenase
deficiency. Semin Hematol 27:137, 1990 3. Luzzatto L, Afolayan A: Different types of human erythrocyte
glucose-6-phosphate dehydrogenase, with characterization of two new genetic variants. J Clin Invest 47:1833, 1968
4. Beutler E: Glucose-6-phosphate dehydrogenase deficiency. N Engl J Med 324:169, 1991
5. Vulliam TJ, DUrso M, Battistuzzi G, Estrada M, Foulkes NS, Martini G, Calabro V, Poggi V, Giordano R, Town M, Luzzatto L, Persico MG: Diverse point mutations in the human glucose-6-phos- phate dehydrogenase gene cause enzyme deficiency and mild or severe hemolytic anemia. Proc Natl Acad Sci USA 855 17 1, 1988
6. Chiu DTY, Zuo L, Chen E, Chao L, Louie E, Lubin B, Liu TZ, Du CS: Two commonly occurring nucleotide base substitutions in Chinese G6PD variants. Biochem Biophys Res Commun 180: 988, 1991
7. Chao L, Du CS, Louie E, Zuo L, Chen E, Lubin B, Chiu DTY: A to G substitution identified in exon 2 of the G6PD gene among G6PD deficient Chinese. Nucleic Acids Res 19:6056, 1991
8. Hirono A, Kuhl W, Gelbart T, Forman L, Fairbanks VF, Beutler E Identification of the binding domain for NADP+ of human glucose- 6-phosphate dehydrogenase by sequence analysis of mutants. Proc Natl Acad Sci USA 86:10015, 1989
9. Du CS, Xu YK, Hua XY, Wu QL, Liu LB: Glucosed-phosphate dehydrogenase variants and their frequency in Guangdong, China. Hum Genet 80:385, 1988
10. Tang TK, Huang CS, Huang MJ, Tam KB, Tang CJC Iden- tification and diagnosis of glucose-6-phosphate dehydrogenase mu- tations in Chinese. Blood 78:86a, 1991 (abstr, suppl 1)
11. Chang JG, Perng I, Chiou SS, Chen TC, Liu TC, Lee LS, Chen PH, Tang T K Three mutations account for most of the affected Chinese with G6PD deficiency in Taiwan: Detection by natural and artificially created restriction recognition sites. Blood 78:87a, 199 1 (abstr, suppl 1)
12. WHO Scientific Group: Standardization of procedures for the study of glucose-6-phosphate dehydrogenase. WHO Tech Rep Ser 366:1, 1967
13. Kunkel LM, Smith KD, Boyer SH, Borganonkar DS, Wachtel SS, Miller OJ, Berg WR, Jones HS, Rary JM: Analysis of human Y- chromosome specific reiterated DNA in chromosome variants. Proc Natl Acad Sci USA 74:1245, 1977
14. Beutler E, Vives-Corrons JL, Hirono A, Prchal JT, Crader W: The molecular biology of variation in glucose-6-phosphate dehydro- genade. The Red Cell: Seventh Arbor Conference. New York, NY, Liss, 1989, p 39
15. Innis MA: PCR with 7-deaza-2'-deoxyguanosine triphosphate, in Innis MA, Gelfand DH, Sninsky JJ, White TJ (eds): PCR Protocols:
For personal use only. by guest on July 21, 2011. bloodjournal.hematologylibrary.orgFrom

2154 CHlU ET AL
A Guide to Methods and Applications. San Diego, CA, Academic, 1990, p 54
16. Chiu DTY, Zuo L, Chen E, Chao LT, Louie E, Lubin B, Du CS: DNA sequence abnormalities in Chinese glucose-6-phosphate dehydrogenase variants. Blood 78:252a, 1991 (abstr, suppl I )
17. Zuo L, Chen EY, Chang CN, Du CS, Liu TZ, Chiu DTY: A new mutation responsible for severe G6PD deficiency in two ethnic Chinese with different clinical presentations: Determination by a direct PCR sequencing technique. Int J Hematol 5939, 1992
18. Stevens DJ, Wanachiwanawin W, Mason PJ, Vulliamy TJ, Luzzatto L: G6PD Canton a common deficient variant in South East Asia is caused by a 459 Arg to Leu mutation. Nucleic Acids Res 18: 7 190, 1990
19. MacDonald D, Town M, Mason P, Vulliamy T, Luzzatto L, Goff D K Deficiency in red blood cells. Nature 350: I 15, 199 1
20. Kurdi-Haidar B, Mason PJ, Berrebi A, Ankra-badu G, ALAE A, Oppenheim A, Luzzatto L: Origin and spread of the G6PD variant (G6PD-Mediterranean) in the Middle East. Am J Hum Genet 47: 1013, 1990
2 I . Beutler E, Kuhl W The NT 13 I I polymorphism of G6PD: G6PD Mediterranean mutation may have originated independently in Europe and Asia. Am J Hum Genet 47: 1008, 1990
22. Kay AC, Kuhl W, Prchal JT, Beutler E: The origin of G6PD polymorphisms in Afro-Americans. Am J Hum Genet 50394, 1992
23. Vulliamy TJ, Othman A, Town M, Nathwani A, Falusi AG, Mason PJ, Luzzatto L: Polymorphic sites in the African population detected by sequence analysis of the G6PD gene outline the evo- lution ofthe variants A and A-. Proc Natl Acad Sci USA 88:8568, 1991
For personal use only. by guest on July 21, 2011. bloodjournal.hematologylibrary.orgFrom