microbial-related biogenic structures from the middle - lneg
TRANSCRIPT

Abstract: A review with the description of the low diverse ichnoassembla -ges from the slates of Valongo Formation is made for the first time. Amongthe organic-rich mining, simple trace fossils dominated by Planolites-likeworm burrows and very rare Chondrites developed in short-termed dysoxicintervals, we describe Phycodes canelensis nov. isp. as an adaptation to storefaecal pellets poorer in nutrients away from the feeding area or forcultivating bacteria. The most remarkable are very large-size, concentricallyringed, pyritized structures, developed in relation with the decompositionof orthoconic cephalopods and dissolution of their shells, which arereinterpreted now as microbialites.This study documents a dysoxic-dominated ecosystem in dependence ofbacterial metabolism, still with a relative abundance of Ediacaran holdoversthat was developed in the wider period of tiering complexification anddiversification of ecospaces, feeding habits and optimization, with thegeneralized appearance of deeper deposit-feeding patterns in the offshoreduring the Ordovician radiation.
Keywords: slates; dysoxic/anoxic fluctuations; pyritization; Ediacaranholdovers; Ordovician radiation.
Resumo: Pela primeira vez é feita uma revisão com descrição formal dasassociações de icnofósseis pouco diversificadas das ardósias da Formaçãode Valongo. Entre as estruturas de bioturbação simples resultantes daexplora ção de ambientes ricos em matéria orgânica dominadas por galeriasvermiformes do tipo Planolites, e com raras Chondrites desenvolvidas nosintervalos disóxicos de curta duração, descrevemos Phycodes canelensis
nov. isp. como uma adaptação comportamental ara armazenar pelóidesfecais, mais pobres em nutrientes, para mantê-los afastados da área dealimentação, ou como meio de cultura bacteriana para revisitação posterior.Mas as mais surpreendentes são estruturas piritizadas de origem orgânica,com disposição em anéis concêntricos e de grandes dimensões, que seformaram em relação com a decomposição de cefalópodes ortocónicos, bemcomo da dissolução das suas conchas, e que se reinterpretam aqui comomicrobialitos. Este trabalho documenta um ecossistema disóxico na dependência dometabolismo bacteriano, com uma abundância relativa de relíquias compor -ta men tais ediacáricas, que se desenvolveu num período de complexificaçãoda estratificação ecológica e de diversificação dos ecospaços e dos hábitosalimentares, com a optimização dos recursos, que aconteceu com ageneralização no offshore de estratégias mais complexas e profundas deobtenção de alimento pelos deposítivoros endobentónicos durante o grandeEvento de Biodiversificação no Ordovícico.
Palavras-chave: ardósias; flutuações disóxicas/anóxicas; piritização;radiação ordovícica; relíquias ediacáricas.
1. Introduction
Slates may be characterized by concentrations of organic matter.High organic inputs occur from areas of primary productivity withdensity stratification within the water column, which preventsmixing of surface and bottom waters, and allows rapid oxygendepletion or complete anoxia in benthic environments (Oschmann,2000). Oxygen is a major limiting factor for metazoan life. Forinstance, decomposers use large amounts of oxygen for respiration,which when combined with a stratified water column can result inanoxia at and below sea floor (Weissberger et al., 2009). This willbasically impede all metazoan benthic life from existing. In the newcircumstances, sulfate-reducing bacteria carry most of the anaerobicrespiration and thrive in such inhospitable environments. Hydrogensulfide released by these organisms is, in part, trapped by ferrousiron causing black coloration of the mud. Iron reduction is causedby a variety of Archaeobacteria and Eubacteria, and is particularlyimportant in association with sulfate reduction. The activity of ironreducing bacteria, together with sulfate reducing bacteria, can bedetected in the fossil record by their metabolic waste precipitates,such as pyrite special aggregates, macroscale burrows and pyritizedfossils (shells and spaces left by skeletal dissolution), and evenconcretions and concretionary layers (Oschmann, 2000; see alsoRetallack, 2015). Pyritization is, thus, a form of permineralizationof organic structures.
Microbialites are trace fossils that record the interactionbetween microbial activity and sediments (Shapiro, 2007). At a slowrate of re-oxygenation, long-term dysaerobic conditions may exist,favoring the development of microbial mats filling the interstitialvoids in the sediment (Wetzel and Uchman, 1998). Microbial matshave been always thriving in dysoxic and even anoxigenicenvironments since the dawn of life on Earth. However, along theirevolution, some metazoans found fruitful ways to establish trophicrelationships with microbial mats in both oxyx and dysoxic
Artigo original
Original Article
Microbial-related biogenic structures from the Middle
Ordovician slates of Canelas (northern Portugal)
Estruturas biogénicas com relação microbiana nas ardósias
do Ordovícico Médio de Canelas (Norte de Portugal)
C. Neto de Carvalho1*, H. Couto2, M. V. Figueiredo3, A. Baucon1,4
© LNEG – Laboratório Nacional de Geologia e Energia IP
Versão online: http://www.lneg.pt/iedt/unidades/16/paginas/26/30/209 Comunicações Geológicas (2016) 103, Especial I, 23-38ISSN: 0873-948X; e-ISSN: 1647-581X
1 UNESCO Global Naturtejo Geopark, Geology and Paleontology Office, CentroCultural Raiano, Av. Joaquim Morão, 6060-101 Idanha-a-Nova, Portugal. 2 ICT- Instituto de Ciências da Terra, Universidade do Porto. Rua do Campo Alegre687, 4169-007 Porto, Portugal 3 Centro de Interpretação Geológica de Canelas, Canelas de Cima Cx 213, 4540-252Arouca, Portugal/Ardósias Valério and Figueiredo, Lda., 4540-252 Arouca, Portugal. 4 Dipartimento di Scienze Chimiche e Geologiche, University of Modena. Via Campi,103 - 41125 Modena , Italy *Corresponding author/Autor correspondente: [email protected]
24 C. Neto de Carvalho et al. / Comunicações Geológicas (2016) 103, Especial I, 23-38
environments. As a result, endobenthos are shallower tiered underhypoxic periods (Weissberger et al., 2009). In this paper, we presenta medium diverse ichnoassemblage from the dark slates from theMiddle Ordovician. These shallow tiered, oxygen-related ichnocoe -noses (sensu Savrda and Botjer, 1986) seem to be developed in closeassociation with microbial-related pyrite rich sediments and locallyabundant shelly epifauna.
2. Geological setting
2.1 Stratigraphy
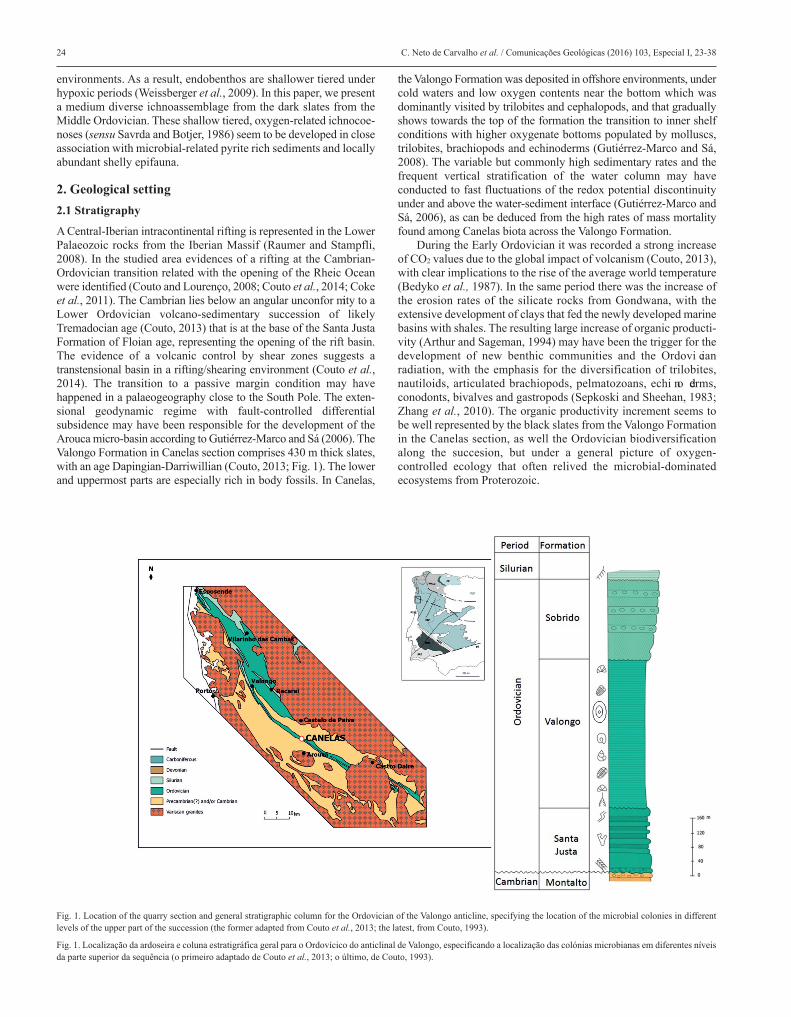
A Central-Iberian intracontinental rifting is represented in the LowerPalaeozoic rocks from the Iberian Massif (Raumer and Stampfli,2008). In the studied area evidences of a rifting at the Cambrian-Ordovician transition related with the opening of the Rheic Oceanwere identified (Couto and Lourenço, 2008; Couto et al., 2014; Cokeet al., 2011). The Cambrian lies below an angular unconfor mity to aLower Ordovician volcano-sedimentary succession of likelyTremadocian age (Couto, 2013) that is at the base of the Santa JustaFormation of Floian age, representing the opening of the rift basin.The evidence of a volcanic control by shear zones suggests atranstensional basin in a rifting/shearing environment (Couto et al.,2014). The transition to a passive margin condition may havehappened in a palaeogeography close to the South Pole. The exten -sional geodynamic regime with fault-controlled differentialsubsidence may have been responsible for the development of theArouca micro-basin according to Gutiérrez-Marco and Sá (2006). TheValongo Formation in Canelas section comprises 430 m thick slates,with an age Dapingian-Darriwillian (Couto, 2013; Fig. 1). The lowerand uppermost parts are especially rich in body fossils. In Canelas,
the Valongo Formation was deposited in offshore environments, undercold waters and low oxygen contents near the bottom which wasdominantly visited by trilobites and cephalopods, and that graduallyshows towards the top of the formation the transition to inner shelfconditions with higher oxygenate bottoms populated by molluscs,trilobites, brachiopods and echinoderms (Gutiérrez-Marco and Sá,2008). The variable but commonly high sedimentary rates and thefrequent vertical stratification of the water column may haveconducted to fast fluctuations of the redox potential discontinuityunder and above the water-sediment interface (Gutiérrez-Marco andSá, 2006), as can be deduced from the high rates of mass mortalityfound among Canelas biota across the Valongo Formation.
During the Early Ordovician it was recorded a strong increaseof CO2 values due to the global impact of volcanism (Couto, 2013),with clear implications to the rise of the average world temperature(Bedyko et al., 1987). In the same period there was the increase ofthe erosion rates of the silicate rocks from Gondwana, with theextensive development of clays that fed the newly developed marinebasins with shales. The resulting large increase of organic producti -vity (Arthur and Sageman, 1994) may have been the trigger for thedevelopment of new benthic communities and the Ordovi cianradiation, with the emphasis for the diversification of trilobites,nautiloids, articulated brachiopods, pelmatozoans, echi no derms,conodonts, bivalves and gastropods (Sepkoski and Sheehan, 1983;Zhang et al., 2010). The organic productivity increment seems tobe well represented by the black slates from the Valongo Formationin the Canelas section, as well the Ordovician biodiversificationalong the succesion, but under a general picture of oxygen-controlled ecology that often relived the microbial-dominatedecosystems from Proterozoic.
Fig. 1. Location of the quarry section and general stratigraphic column for the Ordovician of the Valongo anticline, specifying the location of the microbial colonies in differentlevels of the upper part of the succession (the former adapted from Couto et al., 2013; the latest, from Couto, 1993).
Fig. 1. Localização da ardoseira e coluna estratigráfica geral para o Ordovícico do anticlinal de Valongo, especificando a localização das colónias microbianas em diferentes níveisda parte superior da sequência (o primeiro adaptado de Couto et al., 2013; o último, de Couto, 1993).

Microbial colonies and ichnofossils in slates 25
2.2 Paleontology
The record of trace fossils from Middle and Late Ordovician mudstonesuccessions in the Iberian Peninsula remains poorly developed (afterAceñolaza and Gutiérrez-Marco, 1999). Trace fossils from ValongoFormation in the North of Portugal has been identified or onlysummarily described and partially figured (Couto and Gutiérrez-Marco, 2000; Gutiérrez-Marco and Sá, 2004, 2006, 2008; Sá et al.,2009), usually under biostratigraphic studies. Trace fossils inmudstones are often barely recognizable due to rarity, compactionor rapid weathering (Wetzel and Uchman, 1998). The ichnologicalassemblages known from the Valongo Formation are low diversifiedand occur in shallow tiers, due to the almost permanent anoxicconditions under the interface water-sediment (Gutiérrez-Marco andSá, 2006). Perhaps the most remarkable biogenic structure is theone described as Rotundusichnium arouquensis in Gutiérrez-Marcoand Sá (2006), and later Rotundusichnium isp. in Gutiérrez-Marcoand Sá (2008) and Sá et al. (2009). The ichnological recordidentified and/or summarily described so far includes Taenidium cf.planicostatum, Phycodes noha, Chondrites, Cladichnus?,Tomaculum problematicum, Palaeophycus, Sericichnus?, Trichichnus,Arachnostega gastro chaenae, bioerosion structures in shells suchas Terebripora perplexa made by endolithic bryozoans andtriangular-to-irregular bite marks in molluscs and trilobitesattributed to nautiloids (Couto and Gutiérrez-Marco, 2000;Gutiérrez-Marco and Sá, 2004, 2006, 2008; Sá et al., 2009).However, the Praedichnion identified and figured by Gutiérrez-Marco and Sá (2006) and Gutiérrez-Marco et al. (2008, Fig. 1F)hardly can be distinguished from scavenging bite marks. As a matterof fact, the “bite mark” in the figured example affecting a trilobiteis rather a split chip of slate. Palaeophycus stuffed with Tomaculum
described by Gutíérrez-Marco and Sá (2004) in the lower part ofthe Valongo Formation may be better assigned by definition toAlcyonidiopsis Massalongo, 1856. Trichichnus is mostly a verticalburrow filled with iron oxides most difficult to distinguish from thevertical shafts of Chondrites.
3. Methods
The present contribution is based on the collection of Centro deInterpretação Geológica de Canelas, organized and managed by
Manuel Valério Figueiredo. From a larger collection resulting frommore than 20 years of quarrying operation and research, 1118 fossilswere analyzed, measured and inventoried so far. Measurementswere taken from the most relevant morphological features, alsobecause tectonics has distorted significantly the fossils, andbehavioral interactions, such as possible bite marks and bioturbationwithin the internal moulds, besides taphonomic aspects withpaleobiological implications (e.g., exuvia, spatial patchiness) wereespecially scrutinized. The collection is famous by the remarkablenumber of giant trilobites (Delgado, 1892; Thadeu, 1956; Gutiérrez-Marco et al., 2009; Figueiredo, 2011), the commonest ones beingNeseuretus avus (21.76%), Ogyginus forteyi (16.75%) and Ectillaenus
giganteus (11.45%). Asaphid, dikelocephalid, illaenid, lichid,bathycheilid, calimenid, pliomerid, cheirurid and dalmanitidtrilobites are dominant in the Canelas biota, but the shelly bedsinclude also epiplanktonic graptolites, bivalves, brachiopods,rostroconchs, gastropods, orthoceratid and endoceratid cephalopods,cystoids, crinoids, ophiuroids, hyolithids, conularids and ostracods(Sá and Gutíérrez-Marco, 2006). 77.5% of the fossils may representcorpses that show the low impact of hydrodynamic remobilizationand scavenging in Valongo Formation. Fossils are preserved mostlyas internal moulds (78.2%) that are partially pyritised or showingpyrite lining (sensu Canfield and Raiswell, 1991).
4. “Rotundusichnium” as discoid microbial colonies
4.1 Description of the structures
Perhaps the most remarkable trace fossils ever found among theCanelas biota are three dimensional, dome-shaped and large-sizedstructures that were attributed before to Rotundusichnium Plička(1989); see discussion below (Fig. 3). 10 discoid structures werefound in specific layers from the upper part of the succession. Theyhave an ellipsoidal shape showing variable size, from 600 to >1250mm (CIGC97), and are formed by a variable number of up to 20rings independent from, but placed next to another. The presentthickness varies between 6mm and several cm, but suffered fromflattening and elongation by tectonic strain. In thin sections, thesestructures are shown to be made of chlorite, sericite (sometimesrecrystallized in muscovite), quartz, chloritoid, and opaques. Amongthese, pyrite is abundantly aligned along the foliation planes,
Fig. 2. Elliptical ringed, pyritized structures observed under metallographic microscope. A – Pyrite aligned along the lamination planes and scattered in the matrix. B – Pyrite(yellow) in framboids and individual euhedral crystals; graphitoids (brownish pink) driven along foliation (photomicrograph, thin section, reflected light, parallel nichols).
Fig. 2. Estruturas piritizadas em anéis elípticos analisados no microscópio metalo gráfico. A – Pirite alinhada ao longo dos planos de laminação sedimentar e dispersa pela matriz.B – Pirite (amarelo) sob a forma de frambóides e de cristais euédricos individualizados; grafitóides (rosa acastanhado) presente ao longo da foliação (microfotografia de lâminadelgada observada em luz reflectida e nicóis paralelos).

26 C. Neto de Carvalho et al. / Comunicações Geológicas (2016) 103, Especial I, 23-38
lenticular or scattered in the matrix, occurring in framboids orindividual euhedral crystals. Rare graphitoids and ilmenite occuralso driven along foliation (Fig. 2).
The size of the structures is not related with the number of rings,as we can understand by comparing specimens in Fig. 3. Theellipsoidal shape is obviously controlled by tectonic strain (Figs.3B, 3D). According to Gutiérrez-Marco and Sá (2006) the strongVariscan sin-D1 tectonic deformation, from which resulted theslates, is responsible for the generalized dilatation of the fossils upto 30%. Moreover, originally concentric fossils became ellipticalaccording with the direction of maximum compression. Theelongation and flattening are parallel to the regional lineation andfoliation, respectively. Due to that the rings are imbricateincreasingly dipping towards the core (Fig. 3F). The external ringis always much larger, horizontal and thinner (Fig. 3C). Usuallythese discoid structures show a lower planar side related with thepresence of large endoceratid cephalopods or their “phantoms”(Gutiérrez-Marco and Sá, 2006; Figs. 3C, 3E). The cephalopod maybe located or not in the core of the structure and be orientedaccording to the longer dimension of the ellipse or the shorter one.In the specimen CIGC95, the structure envelops the endoceratidshell, above and below it (Fig. 3E). The annulated discs arecommonly smaller than the related cephalopods.
4.2 Previous interpretation of “Rotundusichnium”
Some papers and abstracts deal briefly with these forms as animaltrace fossils (Gutiérrez-Marco and Sá, 2006; Gutiérrez-Marco andSá, 2008; Sá et al., 2009). Identified as Rotundusichnium
arouquensis due exclusively to their age (Gutiérrez-Marco and Sá,2006), much older than the oldest forms of this ichnogenus known,which are dated from Maastrichtian (Uchman, 2004), they neverhad, however, formal taxonomic review (referred as Rotundusichnium
isp. by Gutiérrez-Marco and Sá, 2008; Sá et al., 2009).These forms from Canelas were initially described as
Rotudusichnium, an ichnogenus considered nomem dubium byUchman (1998). Its interpretation was associated to the bacterialdecay of the cephalopods and systematic grazing by an unknownanimal, starting from the maximum concentration of organic matterand exploring the surrounding sediments by making concentricfurrows with a proboscis. This behavior would evidence tigmotaxyfor a maximal efficiency in the exploration of the limited nutritionalresource (Gutiérrez-Marco and Sá, 2006). The animal would bepositioned in the furrow developed immediately before and producea furrow next to it, by using a bulldozing proboscis, jumping fromthe latest furrow to the newly one and starting again.
Plička (1989) described the ichnogenus Rotundusichnium
(Llarena, 1946), in which includes as type ichnospeciesHelminthoida zumayensis described by Llarena (1946) as a thinfurrow coiled over itself making a planar spiral. Plička defines theichnogenus, not as a spiral, but as a circular impression with aconcentric internal division developed on a sandstone or mudstonebed, with a variable size between 2.7-26 cm (Plička, 1989). Thisimpression is interpreted as being produced by an organism rotatingon the sea floor, or as cubichnion.
The form Rotundusichnium magnum Plička would representexceptionally large individual circles with areas dipping towardsthe core of the structure. The same pattern was identified asSpirorhaphe zumayensis (Llarena) by Książkiewicz (1970).Książkiewicz (1977) describes Spirorhaphe zumayensis from thePolish Carpathians as exichnial structures with very narrow-gaugedfurrows. The bottom of the furrows is inclined towards the centreof the “spiral”. For this author, in most cases these traces occur indark, almost black shales. Seilacher (1977) synonymised Helminthoida
zumayensis with the ichnogenus Scolicia. However, features of
Scolicia are not found in these structures and the problem ofichnogeneric affinity still persists (Uchman, 1999). It is considereda typical deep-sea trace fossil (Uchman, 2004), a graphoglyptid thatoccurs in fine-grained turbidites (Uchman, 2001). Developing anemended diagnosis, Uchman (1998) describes “Rotundusichnium”as a spiral epichnion, with ridges partially superposed and dippingtowards the core of the spiral. These ridges could be perpendicularlystriated.
The “Rotundusichnium” from Canelas would correspond to theoldest representatives of this behavior worldwide, as an earlyadaptation to efficiently explore nutritional hotspots in low-oxygenshallow marine environments that became successful in theoccupation of abyssal niches during the Cretaceous (Gutiérrez-Marco and Sá, 2006). Among the problems that this interpretationrise is the fact that, a) they are elliptical concentric, and not spiralstructures, b) they have a pyrite-rich composition which differs inproportion from the surrounding rock, c) there is no connectionbetween rings, d) the structures are characterized by a central corewith similar composition to the rings and not a first ring surroundinga non-bioturbated area, e) they present an external ring, much largerand flat than the remaining ones, and f) rings are intersected byorthid brachiopods.
The mentioned problematic features are more compatible withmicrobial growth (problematic features a, d, e, f), metabolism(problematic feature b) and growth (problematic feature c) ratherthan an animal trace.
4.3 Microbial-related discoid/concretionary structures
This section discusses in detail the previously mentioned problematicfeatures and relates them with microbial growth, metabolism andlocomotion.
4.3.1 Concentric, non-connected rings (problematic features a, c)
Concentric disconnected rings are explained by the growth andgliding behavior of microbial colonies, a process that findscorrespondence with the modern and fossil record. Modern oxygen-depleted environments typically contain anaerobic heterotrophic,aerobic chemolithoautotrophic and anaerobic photoautotrophicbacteria (e.g., Oschmann, 2000). Bacterial colonies are self-organized biological structures that show collective behavior. Amongothers, bacterial colonies look for biogeochemical differentiation ofmicroenvironments at the sediment-water interface. The increase ofthe colony strongly depends on the nutrient concentration andsedimentation vs. nutrient diffusion rates. The organisms are ableto glide in order to position themselves as close as possible tooptimal food source. Extensive sulfate-reducing microbial mats areknown from the shelf to the deep sea. According to Stolz (2000)microbial communities may be divided in biofilms (microbialcolonies) and microbial mats (stratification of the microbialpopulations into distinct layers). Stromatolites, as microbialites, aremicrobial mat buildups resulting of trapping, binding and precipitationof sediment by the extracellular polymeric matrix produced by thebacteria (Stolz, 2000).
Microbial colonies typically occur as discoidal mounds, amongother forms (see Grazhdankin and Gerdes, 2007). No constrains onmaximum size, concentrically ringed and very large forms as foundfor the discoidal structures from Canelas are evidences for thepresence of microbial colonies (Grazhdankin, 2001). In the discoidalstructures there is the division of the colonies into concentric zonesand wedge-shaped radial oriented sectors. Both characterizemicrobial colonies growing in highly-concentrated nutritionallyheterogeneous environments. Thus, the concentric rings are a resultof non-linear synchronized growth associated with recurrentchanges in the quality, intensity and duration of chemical fields in

Microbial colonies and ichnofossils in slates 27
Fig. 3. Microbial colonies in the Dapingian-Darriwilian slates of Valongo Formation, at Canelas. A – Structure associated to the endoceratid cephalopod shell (CIGC109). B –Largest form preserving the original ellipsoidal 3 D shape. C – Flatten structure showing part of the dome and the lower part with a “phantom” of the cephalopod responsible forits development (CIGC98). D – Phantom and remain of cephalopod oriented perpendicularly to the main structure axis; the outer ring is flat and much larger than the inner ones,like in C (CIGC92). E – Idem; the cephalopod is covered by both upper and lower-ring sides (CIGC95). F – Cross section showing centripetal-oriented rings and the core with thesame pyrite-rich composition of the rings; compare with the model idealized in Fig. 5 (CIGC91). Scales are 5 cm long, except for B, which is 10 cm.
Fig. 3. Colónias microbianas nas ardósias dapingiano-darriwilianas da Formação de Valongo, em Canelas. A – Estrutura associada a concha de cefalópode endoceratídeo (CIGC109).B – A forma de maiores dimensões preservando a estrutura original em elipsóide tridimensional. C – Estrutura achatada mostrando parte do domo superior e a face inferior comum “fantasma” do cefalópode responsável pelo seu desenvolvimento (CIGC98). D – Impressão e restos de cefalópode orientado perpendicularmente ao eixo maior da estrutura; oanel externo é plano e muito mais largo que aqueles mais internos, tal como em C (CIGC92). E – Idem; o cefalópode encontra-se coberto pelas faces superior e inferior dos anéis(CIGC95). F – Secção mostrando os anéis com inclinação centrípeta e o núcleo com a mesma composição sedimentar, rica em pirite, dos anéis; compare-se com o modelo idealizadona Fig. 5 (CIGC91). As escalas gráficas têm 5 cm, excepto em B, que possui 10 cm.

28 C. Neto de Carvalho et al. / Comunicações Geológicas (2016) 103, Especial I, 23-38
the substrate, as in epigenetic Liesegang rings, where concretionarygrowth was rhythmical in anisotropic sediments, such as in stiff mud(Seilacher, 2001), resulting in discoid forms. The probability ofpreservation of microbial relicts increases with the inhospitabilityof the depositional environment (Oschmann, 2000). Withcontinuous venting of H2S, concentration is highest in the border,where nutrients are abundant but minimal in the colony, wherenutrients are consumed, taking as principle that the process ofdiffusion is faster than the process of mat growth. In this way, bysurface translocation behaviour of bacterial population, there is ringaccretion to the colony showing weak, irresilient connectionsbetween adjacent rings (see Fig. 5 for graphical interpretation of themicrobial colonies).
Discoidal structures may be found from Mesoproterozoic andeven earlier and continued into the early Cambrian, according toGrazhdankin and Gerdes (2007). Three-dimensional disk-shapedcasts and composite discoidal moulds are mostly interpreted as basalholdfast structures of unidentified organisms (Gehling et al., 2000).The discoid structures were terraced bacterial colonies, in broad andflat concentric rings. The coherent ring pattern in the discoidalfossils is interpreted as accretionary growth rings. Ions diffusethrough mucus very slowly. Under the low laminar flows the cellclusters tend to be approximately circular in shape (Stoodley et al.,1999). Ellipsoidality is related with the tectonic main Variscanorogenic strain direction and intensity.
The discoid structures from Canelas may be compared with themorphotype “Cyclomedusa” of Grazhdanski and Gerdes (2007, Fig.1D), corresponding to microbial colonies exhibiting a dense ringstructure. Their distribution seems to be random, but stronglycontrolled by the presence of cephalopods. The levels showingmicrobial colonies are not bioturbated. They are relatively uncommonin the Valongo Formation because cephalopds are free swimminganimals and the colonies developed from their death and sinking inanoxic bottoms, with the rare presence of epibenthic organisms anddevoided of endobenthic bioturbation.
4.3.2 Pyrite (problematic feature b)
Microbial mats are regarded to have been commoner in thePrecambrian than in the Phanerozoic, although recent studiessuggest that there is a false perception of an existing Precambriandominance of microbially-induced sedimentary structures (Davieset al., 2015). Occurrences of post-Palaeozoic microbial mats in deepsea and/or oxygen-depleted environments were described by severalauthors (in Hagadorn and Bottjer, 1999). Microbial mats are oftenidentified in the stratigraphic record by concentrations of heavymetals, such as pyrite, magnetite or ilmenite, and the surfacefilaments tend to baffle and trap finer-grained micaceous sediment(Hagadorn and Bottjer, 1999). Biostabilization of sediments isprovided by sticky extracellular polymeric substances in additionto organic meshwork of bacterial filaments. The activity of ironreducing, and sulfate reducing bacteria, can be detected by theirmetabolic waste products, disseminated solid precipitates such aspyrite, burrow fillings and linings, pyritized fossils, evenconcretions and concretionary layers (Oschmann, 2000). Raiswell(1987) among other authors describe the precipitation of pyrite ondecaying organic matter in reducing chemical environments. Pyriteis found replicating fine anatomical details, or lining internalmoulds, in many body fossils or as in the filling of most trace fossilsof Canelas. In fact, deposit-feeding mucus may have served both asfood source and as a “culture” medium for the sulfate reducers(Schieber, 2002). Fine-grained pyrite formation involving sulfate-reducing heterotrophic bacteria in natural systems occurs rapidlyvia biogeochemical diagenetic processes (Donald and Southam,1999). Localized iron sulfide growth in sediments requires that the
sediment is anoxic but also non-sulfidic (Brett and Allison, 1998).Metal precipitation is induced in response to metabolic wasteproduction of reactive inorganic ligands such as sulfide, or throughenzyme mediated changes in metal redox state (Ferris, 2000).Dissolving of Endoceratid cephalopod carbonate shells duringdecaying in an acidic substrate may have facilitated growth ofsulfate reducing bacteria and contributed for the development of thecolony. Pyrite growth in a microbial mat matrix includes anabnormally loose grain fabric with “floating” grains and pyrite thatis free of mineral inclusions (Schieber, 2002).
According to Wilkin and Barnes (1996), framboids are denselypacked, internally spherical aggregates of pyrite. Framboid sizedistribution is a sediment property fixed very early during anoxicdiagenesis and preserved through diagenesis and lithification,meaning that framboidal pyrite wouldn’t grow with successiveburial. Because spherulization cannot be produced by some physicalproperty of the iron-sulfur water system, it must be imposed bysome preexisting, e.g., a sulfate reducing bacterial microaggregate(Raiswell et al., 1993; Southam et al., 2011).
4.3.3 Central core (problematic feature d)
The ellipsoidal structures from Canelas are normally associated withlarge-size cephalopods, showing different dimensions andorientations with relation to their remains. After the death of acephalopod, gradual decay would lead to the deposition on theanoxic sea bottom. Due to the collapse of the dwelling chamber byslow decaying of the soft body and sediment accumulation, therewould be the release of fluids and gases rich in H2S that woulddiffuse from the venting area in all directions around the shell(described by Zangeil et al., 1969 for the goniatite shells and thelarge Rayonnoceras found in the Fayaetteville Shale; Gutiérrez-Marco and Sá, 2006), providing conditions for the growth ofanaerobic heterotrophs or aerobic chemoautotrophs.
4.3.4 External ring (problematic feature e)
The inoculation of the microbial colonies may have been triggeredby the localized nutritional burst, during the brief time required forbiogenic decomposition. This movement towards chemicalattractants is commonly known as chemotaxis. The same processwas described by Zangeil et al. (1969) for nucleation of concretionsin the Upper Mississipian Fayetteville black shale. The coherentring pattern found in concretions may be interpreted as accretionarygrowth rings (Seilacher et al., 2003) or nucleation halos of Southamet al. (2011). Growth is compact at high concentrations of nutrients.Isotropy in the concentric rings results from low nutrient-richdiffusivity and solubility in mud sediments (Chadan, 1991;Ortoleva, 1994). The Black Island Member of the Winnipeg Forma -tion (Ordovician) contains a variety of early diagenetic iron sulfidemorphologies, including elongate concretions associated to burrows(Schieber, 2002). According to Zangeil et al. (1969) pyritic nodulesare associated with small goniatite shells and large shells ofRayonnoceras, with the collapse of the living chambers due tooverpressure following escape of gas from the decomposing carcass.Pyrite occupies all the chamber space. The interaction betweenorganic degradation products from the cephalopods and metallicions from the mud lead to onion-skin concretions made by cone-shaped concentrations of sulfides in the unconsolidated sedimentabove the venting holes (Zangeil et al., 1969). In the MiddleOrdovician of Sweden there is concentric bedding-parallel stromato -litic growth around shells (Zessin, 2011). The Dalby Limestoneshows orthoconic cephalopods enveloped by laminated encrusta -tions, disc-shaped concretions that vary in size, reaching up to 60cm diameter and 10 cm thickness. This results in various large disc-shape structures, once called Medusina silurica von Huene, 1904

Microbial colonies and ichnofossils in slates 29
(Holmer, 1989). In the Dapingian-Darriwillian formations from theArmorican Massif there are fossiliferous concretions that resultedfrom local physical-chemical modifications in the vicinity ofdecaying organisms, prior to lithification (Becq-Giraudon et al.,1992; Dabard and Loi, 2012). These are just some examples ofeodiagenetic development of certain concretions coincident with thedecomposition process of enclosed organisms that has beenrepeatedly described since almost one century ago (e.g., Tarr, 1921;Zangeil et al., 1969; Brett and Allison, 1998).
The number of rings is variable in all the discoidal structuresfound in Canelas, being the last ring significantly larger than theremaining ones. The wrinkles parallel to the longer dimension ofthe discoidal structures, similar to those found in the deformedtrilobites, are evidence for a common taphonomical history. On thecontrary to what happens with undisturbed fossil remains insidenodules, linked to anaerobic eodiagenesis around the decayingorganism (Becq-Giraudon et al., 1992), biofilm viscoelasticityallows the biofilm to have been structurally deformed together withthe large cephalopod when exposed to varying shear stresses(Stoodley et al., 1999), such as tectonic deformation during theVariscan Orogeny. This clearly points to a bacterial colony originof the remarkable large annulated discoids of Canelas instead of justa bacterial-induced precipitation as concretion.
4.3.5 Intersection with brachiopods (problematic feature f)
Some of the rings are intersected by orthid brachiopods, and crossup to two rings, without any evidence of turning around the bodyfossil obstacle as would be expected from an animal producing afeeding burrow/crawling trail.
5. Animal-microbial interactions
Phycodes canelensis isp. nov.Figs. 4A, 4B
Etymology: After the Canelas village where the quarry is located.Type section: Valério and Figueiredo Quarry (Canelas, Arouca,
Portugal), Valongo Formation, Dapingian to Darriwilian (MiddleOrdovician).
Type Material: Several specimens found isolated or together.Holotype – CIGC100 (Fig. 4A); paratypes CIGC82 and CIGC108.
Diagnosis: Flabellate-shaped Phycodes filled with faecal pellets.Description: Flabellate-shaped burrow system starting from a
broad area connected upwards by a single burrow. From this sac-likearea up to 170 mm long, 4 to 9 upward-directed burrows diverge as afan with different, sometimes large angles between them (Fig. 4A).These projections show an elliptical cross section due to com paction,and an irregular shape and size. Filling is composed of faecal pelletsthat may be aligned with the border of the burrows (Fig. 4B).
Remarks: Phycodes noha was described for the first time byMikuláš (1995) from the shales of the Klabava Formation (Floian),Central Bohemia. It was separated from the somewhat similarPhycodes palmatus (Hall, 1852) in having some burrows withpestle-shaped terminations (Mikuláš, 1995). P. canelensis hasfanlike projections with more irregular terminations. Gutiérrez-Marco and Sá (2008: Figs. 1B and 1C) and Sá et al. (2009) attributedto P. noha the forms from Canelas. However, as P. noha is filledwith fine-grained clay material usually differing in color from thesurrounding rock (Mikuláš, 1995), and the forms from ValongoFormation are stuffed with faecal pellets, a different feeding strategyis here advocated. Simple burrows stuffed with faecal pellets areattributed to Alcyonidiopsis Massalongo, 1856. Syncoprulus Richterand Richter was considered synonym of Alcyonidiopsis byChamberlain (1977). Phymatoderma granulata (Schlotheim) showssimilarities with P. canelensis concerning overall shape and type of
filling with faecal pellets (Fu, 1991: Text-fig. 16; Izumi, 2012), butthis ichnogenus displays a non-ramose branching by definition.CIGC 100 shows 5 structures in slightly different stratinomic levels,without any interaction (Fig. 4A). This fact may open the questionwhether their producers were strategically avoiding the contact witheach other, if living at the same time period. The filling of theburrows with faecal pellets enabled the producers of Phycodes
canelensis, i.e., annelids or arthropods, to keep their metabolic wasteaway from their feeding territory, as recorded for Phymatoderma
granulata (Izumi, 2012). A cache strategy for the pellet-filledPhycodes as described for Tubotomaculum mediterranensis byGarcía-Ramos et al. (2014) may be another alternative. Theseauthors hypothesize the recurrent use of the burrows as wastedisposal storage places for cultivate chemoautotrophic sulfideoxidizing bacteria that could be used as food supplies duringstarving periods of low organic input/oxic events.
Planolites beverleyensis (Billings, 1862)Figs. 4C, 4D
Description: Mostly horizontal flattened cylindrical forms with sizevarying in the same burrow from 9 to 52 mm. Burrow width variesfrom 5 mm to 22 mm and can be as long and continuous as 1820mm (Fig. 4C). Burrows follow a gently curving to strong winding,mostly horizontal but sometimes oblique path, sometimesdeveloping irregular meanders, and may show rare branching;branching makes 90º with the main burrow and usually showssmaller tunnels. Filling has a composition different from the hostrock. Sometimes a central cylindrical core has more relief for thepresence of pyrite (Fig. 4D). Pyritised forms of gently curvingPlanolites isp. can also be found in the internal moulds of large andgiant trilobites.
Remarks: Irregular width and flattening are evidences for an activebackfill in a temporary, unwalled burrow developed in a softground.Absence of burrow wall or lining and active backfilling makePlanolites distinct from Palaeophycus (Pemberton and Frey, 1982).Faint menisci and the filling composition may relate Planolites withTaenidium as behaviors of a same soft-bodied vermiform producer. Infact, incipient backfilling may be found in Planolites (Fillion andPickerill, 1990). The same happens when we compare Planolites withHelminthopsis abeli. The sudden change of width along the burrowmay be also the result of diagenesis, differential compaction, shearstress parallel to bedding and changing of stratinomic level. Differentburrow widths in the same tier testimony distinct ontogenetic sizes oftheir producers. Branching is not a common characteristic in Planolites
(Pemberton and Frey, 1982) and could be related with thesuperposition of different burrows “fused” by the strong compactionand tectonics that affected the slates. Pyrite may occur filling thecentral core of the burrow or as secondary deposition developing ahalo, sometimes 3mm thick, around the burrow. The pyrite may beevidence for bacterial activity in the slimy mucus secreted by theproducer to facilitate burrowing. Deposit-feeder mucus may haveserved both as food source and as a “culture” medium for the sulfatereducers (Schieber, 2002). CIGC 90 shows several forms associatedto a large cephalopod shell and pyrite-rich nodules.
Taenidium barretti (Bradshaw, 1981)Fig. 4E
Description: Unwalled, unbranched cylindrical burrows showingmeniscate backfill and a pyritised core, in a winding path. Burrowwidth is 7-21 mm. Meniscate backfill is packeted in portions 9 mmlong or, in other cases, homogeneous, and non-compartimentalized.Menisci are slightly arcuate to semispherical.
Remarks: Taenidium cf. planicostatum was firstly identified byGutiérrez-Marco and Sá, 2006; 2008: Fig. 1D; Sá et al., 2009).

30 C. Neto de Carvalho et al. / Comunicações Geológicas (2016) 103, Especial I, 23-38
Burrow fill may be similar to Planolites. Meniscate burrows reflectthe activity of vagile organisms moving into the substrate in searchfor food, by combining ingestion and bypassing.
Helminthopsis abeli (Książkiewicz, 1970)Fig. 4F
Description: Unwalled and unbranched, irregularly windingcylindrical burrows, mainly horizontal, with a tendency to meandering.Burrow diameter is about 7mm, preserved in semi-relief. Burrow fillshows different composition from the surrounding sediment. Theburrow diameter may not be constant along the length.
Remarks: Burrow fill is biogenic massive and similar toPlanolites and may have shared a common producer.
Cladichnus radiatum (Schröter, 1894)Fig. 4G
Description: C. radiatum consists of an inclined to vertical tube andradiating horizontal, sometimes bifurcate tunnels, and the center ofradiation is within the plan of the branches. The branches are 4-12mm wide and few cm long. Filling is different from the host rock.Backfill seems to be retrusive.
Remarks: So far there are 4 ichnospecies included in theichnogenus Cladichnus, namely C. fischeri (Heer, 1876/77), C.
radiatum (Schröter, 1894), C. aragonensis Uchman, 2001 and C.
parallelum Wetzel and Uchman, 2013 (revised by Wetzel andUchman, 2013). C. radiatum consists of an inclined to vertical tubeand radiating horizontal, sometimes bifurcate tunnels, and the centerof radiation is within the plan of the branches. Bioturbation wasdeveloped in sandy softgrounds by the active redeposition ofsediment during the forward burrowing of the animal (D’Alessandroand Bromley, 1987). Backfill shows grain and mineral sorting whichis typical from deposit feeding.
Arachnostega gastrochaenae (Bertling, 1992)Fig. 4H
Description: Irregular, elongate and net-like burrow systems insediment fills of shells. The size of the whole structure is highlyvariable and ultimately depends on the volume of the internalmould. Ramifying, simple branching made of straight, curved orangular tunnels, <1-to-1 mm wide, on the surface and within theinternal moulds, that are remained open with a circular to ellipticalcross section.
Remarks: This monoichnospecific ichnogenus described byBertling (1992) was revised by Rodrigues et al. (2005) and Fatkaet al. (2011). These bioturbation structures developed in firmsubstrates cannot be misinterpreted with bioerosion because theyare preserved as negative and full relieves, only in the internal castsand not in the surrounding substrate, without evidences of post-diagenetical reworking as rocky substrate (Rodrigues et al., 2005).Arachnostega gastrochaenae is distinguished from Korymbichnus
conflabellatus Damborenea and Manceñido (1996), also occurringin internal moulds, by the more irregular branching and shorterbranches that can be braided or reticulated. Among the collectionof 1118 fossils from the CIGC collection, A. gastrochaenae wasfound only in 15 specimens, including trilobites (Ectillaneus
giganteus mainly, but also Ogyginus forteyi, Neseuretus avus,Bathycheilus castilianus and Hungioides bohemicus), gastropodsand cephalopods. Before, they were known generally only frommollusks and trilobites (Gutiérrez-Marco and Sá, 2006; 2008). Wemay conclude that the common anoxic events most of the timesprecluded the exploitation of the organic-rich sediments inside
shells by the specialized producer of Arachnostega, possibly apolychaete (Bertling, 1992). Its exploratory tunnels on the concealedmicro-habitats were kept open in the firmground that may haveresulted from exopolymer-binding bacteria growing from decayingorganism.
Arachnostega gastrochaenae was described in rocks from thesame age of Canelas, in Montes de Toledo (Spain), by Rodrigues et
al. (2005), harvesting internal moulds of mollusks (bivalves, gastro -pods, rostroconchs and cephalopods), trilobites and echinoderms.Arachnostega is part of the so called “Cambrian substrate revo -lution” (Fatka et al., 2011).
Chondrites intricatus (Brongniart, 1823)Fig. 4I
Description: Small tree-like tunnel systems branching from a central(vertical) shaft. The burrows are irregularly projected downward,but usually short and markedly flattened, forming acute angles.Burrow fill is different from the host sediment, usually light colored.Burrow width between 1-3 mm.
Remarks: Chondritid burrows are interpreted either as deposit-feeding burrows (Phymatoderma, Chondrites, Pilichnus) orchemosynthetic traces (Chondrites) (Bromley, 1996; Hertweck et
al., 2007; Uchmann and Gazdzicki, 2010; Izumi, 2012;). Chondrites
intricatus is a feeding system related to infaunal deposit feeders.According to Fu (1991), the tracemaker of Chondrites may be ableto live under dysaerobic conditions as chemosymbiontic organism;this hypothesis is supported by the incipient Chondrites producedby chemosymbiotic thyasirid bivalves (Bromley, 1996). Chondrites
occurs only in the upper part of the Valongo Formation, and neverdevelops mottled ichnofabrics as in the Middle Ordoviciananalogous facies of the upper Plattin rocks of Eastern Missouri(Shourd and Levin, 1976).
Tomaculum problematicum (Groom, 1902)Fig. 4J
Description: Clusters of pellets with a cylindrical shape, <1-3mmlong and <1mm wide. Usually they show the same composition ofthe host rock.
Remarks: CIGC1986 The ichnogenus Tomaculum was revised byEiserhardt et al. (2001). Tomaculum problematicum pelletedaggregates are relatively common and expected fossils in the slates ofthe Valongo Formation (Gutiérrez-Marco and Sá, 2004, 2006; 2008).
Mandibulichnus serratus (Zamora et al., 2011)
Description: V or W-shaped serrated brakes in shelly fossils.Breakage shape ranges from straight to wavy and show differentsizes (Zamora et al., 2011).
Remarks: Although some of the examples figured in Gutiérrez-Marco and Sá (2006) fit in the description of Mandibulichnus
serratus others are just mechanical breaks affecting flattened fossils(see Gutiérrez-Marco and Sá, 2008: Fig. 1F). Only 8 probablepredation/scavenging scars where recognized in the CIGCcollection so far. We can conclude based on 79.3% completespecimens studied in that collection, of a total of 1118 specimensinventoried so far, that predation/scavenging bites are very rare andthat most of the fauna died instead from sudden changes of the redoxpotential discontinuity. Although scavenging clearly wasresponsible for disarticulation of skeletons in Canelas bottoms it israther difficult otherwise to separate from molting behaviours andbacterial decomposition of organic tissues.

Microbial colonies and ichnofossils in slates 31
Fig. 4. Microbially-related, organic-rich derived feeding patterns from the Valongo Formation. A – Phycodes canelensis nov. isp. (Paratype: CIGC108). B – Detail of one specimenof five included in the same slab filled with faecal pellets (Holotype: CIGC100). C – Planolites beverleyensis, large forms filled with iron oxydes (CIGC90). D – Small form of P.
beverleyensis (CIGC2340). E – Taenidium barretti (CIGC83). F – Helminthopsis abeli (CIGC1976). G – Cladichnus radiatum (CIGC81). H – Arachnostega gastrochaenae, withclear branching (CIGC105). I – Chondrites intricatus (CIGC104). J – Tomaculum problematicum (CIGC84) as burrow fill. Scale as the white bar with 2 cm, except for C, whichmeasures 10 cm.
Fig. 4. Padrões de alimentação tipicamente relacionados com a presença microbiana e abundância de matéria orgânica não degradada da Formação de Valongo. A – Phycodes
canelensis nov. isp. (Paratipo: CIGC108). B – Detalhe de um de cinco icnofósseis presentes no mesmo nível e preenchidos por pelóides fecais (Holótipo: CIGC100). C – Planolites
beverleyensis, formas de grandes dimensões preenchidas por óxidos de ferro (CIGC90). D – Pequena forma de P. beverleyensis (CIGC2340). E – Taenidium barretti (CIGC83). F– Helminthopsis abeli (CIGC1976). G – Cladichnus radiatum (CIGC81). H – Arachnostega gastrochaenae, com ramificações claras e ausência de preenchimento (CIGC105). I– Chondrites intricatus (CIGC104). J – Tomaculum problematicum (CIGC84) preenchendo uma galeria. Escala gráfica com 2 cm, excepto em C, onde esta mede 10 cm.

32 C. Neto de Carvalho et al. / Comunicações Geológicas (2016) 103, Especial I, 23-38
6. Proterozoic lifestyles in an oxygen-depleted environmentduring the onset of the Ordovician biodiversificationevent
6.1 Paleoenvironment of Canelas during the Middle Ordovician
The ichnosite of Canelas is characterized by the followingcharacteristics:
Low bioturbation intensity. Bioturbation intensity is primarilyrelated to sedimentation rates; population density and behaviour arealso important drivers of bioturbation intensity (Gingras et al., 2011).
Sporadically heterogeneous trace-fossil distribution. Followingprevious studies (Gingras et al., 2011), the sporadically heterogeneousdistribution of trace fossils of Canelas indicate the persistent, spatio-temporal variability in physico-chemical conditions. Fluctuation inoxygen concentration is supported by tiering and body fossilcontent. In fact Canelas is an Ordovician Lagersttätte dominated bytrilobites (Gutiérrez-Marco et al., 2009) showing shallow, simpletiering. In the oxic short intervals epibenthic communities wererelatively diversified and frequently abundant but endobenthicfaunas are only occasionally present.
Low ichnodiversity. Overall, the study site shows a moderateichnodiversity, although trace fossils occur in predominantlymonoichnospecific assemblages and very scattered. Low-diversitysuites indicate environmental stress (Gingras et al., 2011).
Dysoxic ichnotaxa. Chondrites and Cladichnus are typical dysoxicichnotaxa (Bromley, 1996; Buatois and Mángano, 2011; Monaco et
al., 2012), indicating that low oxygen was the source of environmentalstress. The trace fossils from Valongo Formation represent a typicaloxygen-related ichnocoenosis of Savrda and Bottjer (1986), dominatedby simple winding feeding burrows and few chemosymbiont-specialized behaviours in shallow tiers. It is included in the Zoophycosichnofacies, in the sense of stressed, quiet-water environmentsexperiencing anoxia and thus preventing highly diverse ichnofaunas(Martin, 2004). Planolites tier in the ichnofabrics may reflect theposition of the redox boundary (Wetzel and Uchman, 1998).
Sedimentological features (lamination; abundance of pyrite)support this hypothesis.
Unlined burrows. Trace fossils of the study site do not presentburrow linings, which is a feature related to non-shifting substratessuch as cohesive muds and/or microbially bound sediments(Gingras et al., 2011).
Microbial-dominated substrates. In the Valongo Formationprevails a Helminthopsis ichnoguild, composed by very simplefeeding burrows and non-specialized grazing trails. They mostlyrepresent transitory, near-surface to very shallow tiered tracesproduced by vagile, Planolites-like vermiform animals (Mánganoand Buatois, 2007). The ichnoguild of the study site may reflect thefeeding exploitation of microbial-rich sediments. Deposit feederswere found to digest and assimilate microbial organisms from thesediment with much greater efficiency than the more refractoryparticulate organic matter (see Levinton and Kelaher, 2004).
For these reasons, the studied ichnosite was characterized by: (a)sedimentation exceeding bioturbation; (b) variability in physico-chemical conditions; (c) environmental stress; (d) dysoxic settings;(e) firm substrates (f) microbial-bound sediments. These characteristicsare typically associated to dysoxic offshore conditions.
6.2 Comparison with Proterozoic biomat-dominated ichnofaunas
The prevailing view is that, until the Phanerozoic, Earth was a mat-dominated world. An increase in diversity and complexity of benthicactivity across the Neoproterozoic-Cambrian boundary lead to theso called “Cambrian substrate revolution “(e.g., Droser and Bottjer,1988; Droser et al., 2002; Marenco and Bottjer, 2008).Nevertheless,recent studies suggest that claims for the stratigraphic restriction of
microbially-induced sedimentary structures to intervals prior to theevolution of grazing organisms result from sampling bias (Davieset al., 2016).
The late Neoproterozoic and Cambrian was a period offundamental change on Earth following the breakup of Rodinia andthe early stages in the formation of the Gondwana, global“snowball” glaciations and major changes in ocean and atmosphericchemistry (Mapstone and Mcillroy, 2006). It was proposed that theNeoproterozoic rise in oxygen after at least three global glaciations(“the Snowball Earth”) was a trigger of the “Cambrian explosion”(see Jensen et al., 2006). Bioturbation in Early Cambrian subtidalmarine substrates was largely a nearly bedding-surface processdominated by Planolites-like simple feeding burrows (Marenco andBottjer, 2008).
The Ordovician radiation was responsible by the replacementof the Cambrian Evolutionary Fauna by the Palaeozoic EvolutionaryFauna that dominated the marine realm until the end-Permian massextinction (Sepkoski, 1995; Van Roy et al., 2010). While Middleand Late Cambrian oceans were dominated by trilobites andphosphatic brachiopods, the Ordovician radiation that may havestarted in the Floian-Dapingian was a faunal turnover responsiblefor a rapidly increasing biodiversity, especially vertical burrowingorganisms (Zhang et al., 2010). Diversification of phytoplanktonwas suggested as a primary trigger for Ordovician biodiversification(Servais et al., 2010). During the Ordovician there was theintensification of the ecospace use in the offshore (Bottjer andDroser, 1994; Orr, 2001). The Ediacaran biotope came to berestricted to deeper or confined environments after Cambrian,possibly related with the increase of bioturbation and benthic tieringcomplexification, which disturbed most of the facies dominated bymicrobial mats and contributed for generalizing softgrounds.
Rare exceptions that evidence the ongoing Ordovician radiationin the marine environments are Arachnostega gastrochaenae
produced by specialized metazoans adapted to explore the verylimited ecospace of the animal shell casts, as well as Phycodes
canelensis or Chondrites, well adapted to dysoxic environments.Chondrites only occurs in the upper part of the Valongo Formation,despite hypoxic and anoxic conditions dominate across the wholesuccession. These tunnel systems are usually attributed tochemosymbiontic producers such as bivalves and polychaetes (Fu,1991).
The advent of burrowing greatly increased the spatial complexityof the sea floor (e.g., Seilacher, 1999). Resource distributionsbecame increasingly patchy with an increase in the range of resourceconcentrations (Plotnick, 2007). According to Orr (2001), thesignificant restructuring of deeper-marine communities at, or near,the Cambrian-Ordovician boundary is attributed to competition forecospace and/or resources within shallow-marine environments, asresult of which pascichnia and agrichnia were displaced offshore.Ediacaran-type palaeoichnocoenoses started to be emplaced undervariable, but low oxygen levels during the deposition of dark-colored layers (Orr, 2001). These anoxic, intertidal, supratidal,hypersaline or thermal kinds of extreme ecological refuges havehosted diverse relict matground communities until the present(Pflüger, 1999). The microbial discoid structures from ValongoFormation occur in black slate facies with basically no bioturbationand clearly represents one example in the fossil record of suchextreme anoxigenic refuges. As it was shown before, enigmaticdiscoidal fossils are common in Proterozoic (e.g., Narbonne, 2005;Mapstone and Mcillroy, 2006; Bengston et al., 2007; Retallack,2007; 2015). Most of these “medusoid” forms were more recentlyinterpreted as colonial microbes (e.g., Bengtson et al., 2007;Grazhdankin and Gerdes, 2007). The simplicity of feeding patternsand simple tiering, with low ichnofabric indices, in the ValongoFormation at Canelas are reminiscent of the type of bioturbation

Microbial colonies and ichnofossils in slates 33
reported from the exceptionally preserved Cambrian faunas of SiriusPasset, Chengjiang and Kaili, and the Early Ordovician deepermarine faunas of Fezouata and Afom Gam (Van Roy et al., 2010;Botting et al., 2015).
Benthic population density depends on the benthic foodavailability and oxygen rates. However, the sedimentary rate is alsoa limiting environmental parameter; in low rates of deposition thepatchiness and swarming effect can be lost to the time-averagingeffect (Wetzel and Uchman, 1998), with several generations ofbenthic communities adapted to the changing environmentalconditions and mixing together in an ecological palimpsest. Theburrowing depth and complexity of an animal depends on suchenvironmental conditions as food availability, pore- and bottom-water oxygenation, sedimentary rates, and substrate consistency.The permeability of muds is low, and also diffusion and convectionof oxygen into the sediment is low (Wetzel and Uchman, 1998).Endogenic pascichnia, such as Planolites, Taenidium orHelminthopsis found in the Canelas section were produced byinfaunal vagile deposit feeders dependent on pore water as a sourceof oxygen, and then less tolerant to benthic oxygen deficiency.Decomposers use large amounts of oxygen for respiration which,combined with a stratified water column, can result in hypoxia atthe seafloor (Weissberger et al., 2009). Aerobic short-term eventsat the sea floor also supported a variety of feeding behaviors oftrilobites, brachiopods, echinoderms, molluscs and hyolithids.However, long-term anaerobic to dysaerobic events that characterizethe black slates in Canelas typically support sparsely populatedcommunities of small, mostly soft bodied, deposit-feedingorganisms (Ekdale and Mason, 1988). Being benthic food contentvery high, behavioral specialization documented by the mostlysimple and winding burrows, with only some few more complexbranching systems developed by dysaerobic specialists, in theValongo Formation, was not required.
6.3 Comparison with ecologically similar Phanerozoic ichnofaunas
The studied ichnosite is characterized by very specific ecologicalfeatures: (a) low-oxygen settings; (b) low hydrodynamic energy(wave base not interacting with the seafloor); (c) biomat-dominatedmuddy substrate. Similar ecosystems are not rare in the Phanerozoicgeological record (Davies et al., 2016), therefore it is interesting todelineate the general ichnological features of low-oxygen, low-energy, biomat-dominated marine ecosystems. However, thecomparison of the studied ichnofauna with environmentally similarPhanerozoic ichnofaunas is challenging because metazoan andmicrobial biogenic structures are rarely discussed contemporaneously.The following ichnosites present comparable features to the Canelasichnosite:
The Atuel Canyon (herein referred as AC) is characterized by aLate Silurian-earliest Devonian sedimentary succession with bothmetazoan trace fossils and microbial mats. Deposition wasinfluenced by gravity flows. In addition, the palaeoenvironmentpresented low-oxygen settings, high sedimentation rates and muddysubstrates (Pazos et al., 2015).
The stream SW of Casera For (herein referred as SC) ischaracterized by dark shales, that have received ichnologicalattention in the study of the Pramollo ichnolagerstätte. The presenceof the ichnotaxon Dictyodora liebeana probably indicates a LowerCarboniferous age (Baucon and Carvalho, 2008). Microbially-induced structures and metazoan trace fossils are reported, low-oxygen conditions are likely because of sedimentological(laminated, dark sediments), geochemical (pyrite) and ichnologicalfeatures (low bioturbation intensity).
The aforementioned sites are characterized by low to moderateichnodiversity (Canelas, AC, SC), low bioturbation and sporadically
heterogeneous trace-fossil distribution (Canelas, SC; similarinformation is not available for AC), unlined traces (Canelas, AC,SC). These features are explained by the inherent features of low-oxygen, quiet, biomat-dominated ecosystems. Low ichnodiversityis likely to reflect the stressful dysoxic conditions; higher(moderate) ichnodiversities may indicate the role played by biomat-dominated niches. Non-shifting substrates such as cohesive mudsand/or microbially bound sediments explain unlined traces.Sporadically heterogeneous trace fossil distribution may indicatepersistent, spatio-temporal variability, such as patchy distributionof biomat-dominated sediments.
The ichnofauna of Canelas differs from both the AC and SCichnofaunas for most of its metazoan ichnogenera (Phycodes,
Planolites, Taenidium, Helminthopsis, Cladichnus, Chondrites) aswell as for the presence of pellets and bioerosional ichnotaxa. Incontrast, the AC and SC ichnofaunas are remarkably similar, beingcharacterized by abundant Nereites and Dictyodora. The onlydifferences concern the presence of graphoglyptids (SC), Zoophycos
(AC) and Diplichnites (AC).These differences are explained by the different geometry of
biomats. The microbial mats of Canelas are dome-shaped, therefore– at the centrimetric scale - they offered a highly heterogeneouslandscape of nutrients and geotechnical properties. The planarmicrobial mats of AC and SC provided a relatively homogeneousnutrient landscape, highly favourable for undermat miners such asthe Nereites tracemaker. In this regard, further studies are requiredto evaluate the behavioural relationship between Dictyodora andmicrobial mats. Similarly, the association between Nereites,
Dictyodora and planar microbial mats is a possible ichnologicalregularity that could be tested by considering ecologically similarichnosites.
7. Conclusions
A review with the description of the low diverse ichnoassemblagesfrom the Ordovician (Dapingian-Darriwillian) slates of the ValongoFormation are made for the first time. This study documents adysoxic-dominated ecosystem in dependence of bacterialmethabolism, based on the collection of the Centro de InterpretaçãoGeológica de Canelas. This collection is famous by the remarkablenumber of giant trilobites that occur in Canelas section with a patchydistribution of thousands of carcasses and exuvia in a limitednumber of beds. Much less common are the trace fossils found inthis section. Body fossil moulds and trace fossils fills are total orpartially permineralized by pyrite. The presence of pyrite aggregatesand framboids may represent anaerobic heterotrophs (sulfatereducers) as mat microbes.
The most remarkable trace fossils ever found among theCanelas biota are three dimensional, dome-shaped and large-sizedstructures that were attributed before to Rotundusichnium Plička(1989). The ellipsoidal structures from Canelas are normallyassociated with large-size cephalopods, showing differentdimensions and orientations with relation to their remains. After thedeath of a cephalopod, gradual decay led to their deposition on theanoxic sea bottom. Due to the collapse of the dwelling chamber byslow decaying of the soft body and sediment accumulation, therehas been the release of fluids and gases rich in H2S that was diffusedfrom the venting area in all directions around the dissolving shell,providing conditions for the growth of anaerobic heterotrophs oraerobic chemoautotrophs by chemotropism. Therefore, the discoidstructures are now interpreted as terraced bacterial colonies, withbroad and flat concentric rings (Fig. 5). The coherent ring patternin the discoidal fossils is interpreted as accretionary growth ringsresulting from swarming. These microbial colonies are function ofecological and taphonomic windows.

34 C. Neto de Carvalho et al. / Comunicações Geológicas (2016) 103, Especial I, 23-38
The variable but commonly high sedimentary rates and thefrequent vertical stratification of the water column may haveconducted to fast fluctuations of the redox potential discontinuityunder and above the water-sediment interface. The organicproductivity increment seems to be well represented by the blackslates from the Valongo Formation in the Canelas section, as wellthe Ordovician biodiversification along the succession, but under ageneral picture of oxygen-controlled ecology that often relived themicrobial-dominated ecosystems from Proterozoic.
Among the microbial-rich deposit feeding strategies found inthe Valongo Formation, Phycodes canelensis is a new ichnospeciesdescribed as flabellate-shaped Phycodes filled with faecal pellets.The filling of the burrows with faecal pellets enabled the producersof Phycodes canelensis, i.e., annelids or arthropods, to keep theirmetabolic waste away from their feeding territory. A cache strategymay be another alternative to explain the new behavioral pattern.
Canelas section is an Ordovician concentration lagersttättedominated by trilobites that typically shows shallow, simple tiering.In the oxic short intervals epibenthic communities were relativelydiversified and frequently abundant but endobenthic faunas are onlyoccasionally present. The Helminthopsis ichnoguild is composedby very simple feeding burrows and non-specialized grazing trails.They mostly represent transitory, near-surface to very shallow tieredtraces produced by vagile, Planolites-like vermiform animalsdominated by simple winding feeding burrows and fewchemosymbiont-specialized behaviors. Rare exceptions thatevidence the ongoing Ordovician radiation in the marine environmentsare Arachnostega gastrochaenae produced by specializedmetazoans adapted to explore the very limited ecospace of theanimal shell casts, as well as Phycodes canelensis or Chondrites,the latest trace fossil usually present in dysoxic substrates.
Persistence of relict microbial colonies and microbial-relatedfeeding communities in the Canelas section after the onset of the“Agronomic Revolution” in the basal Cambrian and during theOrdovician radiation of burrowers, that pervaded shallow-marineecosystems, clearly represents a fine example in the fossil record ofsuch extreme, mostly anoxigenic refuges where “Proterozoicholdovers” may have found temporary conditions to survive.
Acknowledgments
We highlight the contribution of Sören Jensen (Univ. Extremadura)for the initial discussions about the origin of Canelas discoidalstructures. Joana Rodrigues and Mariana Vilas Boas (UNESCONaturtejo Global Geopark) are greatly appreciated for their help inthe development of Figs. 1, 3 and 4. The research of Andrea Bauconis supported by the ROSAE project.
References
Aceñolaza, G. F. and Gutiérrez-Marco, J. C., 1999. Icnofósiles del Ordoví -cico terminal (Pizarras Chavera, Pizarras de Orea: Hirnantiense) dealguns localidades españolas. Boletín Geológico y Minero, 110(2): 123-134.
Arthur, M. A. and Sageman, B. B., 1994. Marine black shales: Depositionalmechanisms and environments of ancient deposits. Annual Review of
Earth and Planetary Sciences, 22: 499-551.Baucon, A. and Neto de Carvalho, C., 2008. From the river to the sea:
Pramollo, a new ichnolagerstätte from the Carnic Alps. Studi Trent. Sci.
Nat. Acta Geol., 83: 87–114.Baucon, A. and Felletti, F., 2013a. Neoichnology of a barrier-island system:
The Mula di Muggia (Grado lagoon, Italy): Palaeogeography,
Palaeoclimatology, Palaeoecology, 375: 112–124.Baucon, A. and Felletti, F., 2013b. The IchnoGIS method: Network science
and geostatistics in ichnology. Theory and application (Grado lagoon,Italy): Palaeogeography, Palaeoclimatology, Palaeoecology, 375: 83–111.
Becq-Giraudon, J.-F., Bouilli, S. and Chauvel, J. -J., 1992. Genesis andsignificance of the silico-aluminous nodules in the Ordovician of theMontagne Noire and the Massif Armoricain (France). Sedimentary
Geology, 77: 77-87.Bengtson, S., Rasmussen, B. and Krapež, B., 2007. The Palaeoproterozoic
megascopic Stirling biota. Paleobiology, 33: 71-120.Bertling, M., 1992. Arachnostega n. ichnog. – burrowing traces in internal
moulds of boring bivalves (Late Jurassic, Northern Germany). Paläont.
Z., 66(1/2): 177-185.Botting, J. P., Muir, L. A., Jordan, N. and Upton, C., 2015. An Ordovician
variation on Burgess Shale-type biotas. Scientific Reports, 5 (9947): 1-11.
Bottjer, D. J. and Droser, M. L., 1994. The history of Phanerozoicbioturbation. In: Donovan S.K. (ed.), The Paleobiology of Trace Fossils.Wiley, Chichester: 155-176.
Brett, C. E. and Allison, P. A., 1998. Paleontological approaches to theenvironmental interpretation of marine mudrocks. In: J. Schieber, W.Zimmerle and P. Sethi (eds), Shales and Mudstones. Schweizerbart,Stuttgart: 301-349.
Bromley, R. G., 1996. Trace Fossils. Biology, Taphonomy and Applications.Chapman and Hall, London, 361.
Buatois, L. A. and Mángano, M. G., 2011. Ichnology: Organism-Substrate
Interactions in Space and Time. Cambridge University Press,Cambridge, New York, 358.
Canfield, D.E. and Raiswell, R., 1991. Pyrite formation and fossilpreservation. In: P.A. Allison and D.E.G. Briggs (Eds.), Taphonomy:releasing the data locked up in the fossil record. Plenum Press, NewYork: 338-388.
Chamberlain, C. K., 1977. Ordovician and Devonian trace fossils fromNevada. Nevada Bureau of Mines and Geology, 90: 1-24.
Fig. 5. “Onion-skin” or concretionary model for the development of microbial coloniesfrom decaying endoceratid cephalopods in low-oxygen bottoms. A – Deposition of thecephalopod on the anoxic bottom. B – Decaying of the animal in the dwelling chamberand progressive dissolution of the carbonate shell trigger chemical and nutritionalconditions for culture of the microbial colony, whose growth in concentric ringsfollowed the fluid dispersion in all spatial dimensions of the substrate (dark and whitecolors used for defining independent rings). C – Section view of the microbial colonygrowth within the substrate.
Fig. 5. Modelo em “casca de cebola” ou concreção, para o desenvolvimento de colóniasmicrobianas por decomposição de cefalópodes endoceratídeos em fundos próximo daanoxia. A – Deposição do cefalópode no fundo anóxico. B – À medida que asedimentação acaba por cobrir o animal, a decomposição das partes moles na câmarade habitação, com a dissolução da concha carbonatada, favorece as condições químicase nutricionais para a cultura da colónia, que cresceu sob a forma de anéis concêntricosacompanhando a dispersão do fluido nas três dimensões do substrato (as cores servemapenas para reforçar que cada anel é independente dos restantes). C – Perspectiva docrescimento da colónia a envolver progressivamente o cefalópode no interior dossedimentos.

Microbial colonies and ichnofossils in slates 35
Chadan, J., 1991. Geochemical self-organization: an application of nonlineardynamics. In: G.V. Middleton (ed.), Nonlinear dynamics, chaos and
fractals with applications to geological systems. Geological Associationof Canada, Short Course Notes, 9: 83-96.
Coke, C., Teixeira, R., Gomes, E., Corfu, F. and Rubio Ordoñez, A., 2011.Early Ordovician volcanism in Eucísia and Mateus areas, CentralIberian Zone, northern Portugal. Mineralogical Magazine, C -Goldschmidt Abstracts, 685.
Couto, H., 1993. As mineralizações de Sb-Au da região Dúrico-Beirã. Phdthesis. Faculty of Sciences of University of Porto, 607.
Couto, H., 2013. The Ordovician of Valongo Anticline (Northern Portugal):State of Art. 13th SGEM GeoConference on Science and Technologies
In Geology, Exploration and Mining, SGEM2013 ConferenceProceedings, 1: 203-208.
Couto, H. and Gutiérrez-Marco, J. C., 2000. Alguns fósseis raros ou poucoconhecidos na Formação de Valongo (Ordovícico Médio, norte dePortugal). In: J.B. Díez and A.C. Balbino (eds.), I Congresso Ibérico de
Paleontologia/XV Jornadas de la Sociedad Española de Paleontología,Évora: 191-192.
Couto, H., Knight, J. and Lourenço, A., 2013. Late Ordovician ice-marginalprocesses and sea-level change from the north Gondwana platform:Evidence from the Valongo Anticline (northern Portugal). Palaeogeo -
gra phy, Palaeoclimatology, Palaeoecology, 375: 1-15.Couto, H., Knight, J. and Lourenço, A., 2014. Rifting at the Cambrian–
Ordovician transition in northwestern Portugal. Comunicações
Geológicas, 101: 251-254.Couto H. and Lourenço, A., 2008. Cambrian-Ordovician transition in
Valongo Anticline: evidences of a continental rift in a Gondwanapassive margin. 33rd International Geological Congress Oslo, Norway.SES-07 Dynamics of sedimentary basins. Abstract CD-Rom.
Dabard, M.-P. and Loi, A., 2012. Environmental control on concretion-forming processes: examples from Palaeozoic terrigenous sediments ofthe North Gondwana margin, Armorican Massif (Middle Ordovicianand Middle Devonian) and SW Sardinia (Late Ordovician). Sedimentary
Geology, 267-268: 93-103.D’Alessandro, A. and Bromley, R.G., 1987. Meniscate trace fossils and the
Muensteria-Taenidium problem. Palaeontology, 30: 743-763.Damborenea, S. E. and Manceñido, M. O., 1996. Icnofósiles (Nucleocavia)
preservados sobre moldes internos de conchillas del Jurásico Medio deloeste argentino. Asociación Paleontológica Argentina, Publicación
Espe cial, 4: 111-120.Davies, N. S., Liu, A. G., Gibling, M. R. and Miller, R. F., 2016. Resolving
MISS conceptions and misconceptions: A geological approach tosedimentary surface textures generated by microbial and abioticprocesses. Earth-Science Review, 154: 210–246.
Delgado, J. F. N., 1892. Fauna Silurica de Portugal: descripção de uma
forma nova de trilobite Lichas (Uralichas) Ribeiroi. Typ. Acad. RealScienc., 31.
Donald, R. and Southam A. M. G., 1999. Low temperature anaerobic bacterialdiagenesis of ferrous monosulfide to pyrite. Geochim. Cosmochim. Acta,63: 2019-2023
Droser, M. L. and Bottjer, D. J., 1988. Trends in depth and extent ofbioturbation in Cambrian carbonate marine environments, westernUnited States. Geology, 16(3): 233-236
Droser, M. L., Jensen, S. and Gehling, J. G., 2002. Trace fossils and substratesof the terminal Proterozoic-Cambrian transition: implications for the recordof early bilaterians and sediment mixing. PNAS, 99(20): 12572-12576.
Eiserhardt, K. -H., Koch, L. and Eiserhardt, W. L., 2001. Revision desIchnotaxon Tomaculum Groom, 1902. Neues Jahrbuch für Geologie
und Paläontologie, Abhandlungen, 221(3): 325-358.Ekdale, A. A. and Mason, T. R., 1988. Characteristic trace-fossil associations
in oxygen-poor sedimentary environments. Geology, 16: 720-723.Engelsen, A., Sundbäck, K. and Hulth, S., 2010. Links between bottom-
water anoxia, the polychaete Nereis diversicolor, and the growth ofgreen-algal mats. Estuaries and Coasts, 33: 1365–1376.
Fatka, O., Mikuláš, R., Szabad, M., Micka, V. and Valent, M., 2011.Arachnostega Bertling, 1992 in the Drumian (Cambrian) sediments ofthe Teplá-Barrandian region (Czech Republic). Acta Geologica
Polonica, 61(4): 367-381.Ferris, F. G., 2000. Microbe-metal interactions in sediments. In: R.E. Riding
and S.M. Awramik (eds.), Microbial Sediments. Springer-Verlag, Berlin:121-126.
Figueiredo, M. V., 2011. The Trilobites of Canelas – Two decades living with
slate and giant trilobites. Centro de Interpretação Geológica de Canelas,85.
Fillion, D. and Pickerill, R. K., 1990. Ichnology of the Upper Cambrian? toLower Ordovician Bell Island and Wabana groups of easternNewfoundland, Canada. Palaeontographica Canadiana, 7: 1-119.
Fu, S., 1991. Funktion, verhalten und einteilung fucoider und lophocteniiderlebensspuren. Courier Forschungsinstitut Senckenberg, 135: 1-79.
García-Ramos, J. C., Mángano, M. G., Piñuela, L., Buatois, L. A. andRodríguez-Tovar, F. J., 2014. The ichnogenus Tubotomaculum: anenigmatic pellet-filled structure from Upper Cretaceous to Miocenedeep-marine deposits of southern Spain. Journal of Paleontology, 88(6):1189-1198.
Gehling, J. G., Narbonne, G. M. and Anderson, M. M., 2000. The first namedEdiacaran body fossil, Aspidella terranovica. Palaeontology, 43: 427-456.
Gingras, M., Hagadorn, J. W., Seilacher, A., Lalonde, S. V, Pecoits, E.,Petrash, D. and Konhauser, K. O., 2011. Possible evolution of mobileanimals in association with microbial mats. Nature Geoscience, 4(6):372-375.
Gingras, M. K., MacEachern, J. A. and Dashtgard, S. E., 2011b. Processichnology and the elucidation of physico-chemical stress. Sedimentary
Geology, 237: 115–134. Grazhdankin, D., 2001. Microbial origin of some of the Ediacaran fossils.
Geol. Soc. Amer., Abst. Progr., 33: 429.Grazhdankin, D. and Gerdes, G., 2007. Ediacaran microbial colonies.
Lethaia, 40: 201-210.Gutiérrez-Marco, J. C. and Sá, A. A., 2004. O enigma dos Tiges de végétaux
terrestres de idade ordovícica, encontrados por Nery Delgado (1908) naFormação Valongo (norte de Portugal). Comunicações Geológicas, 91:37-44.
Gutiérrez-Marco, J. C. and Sá, A. A., 2006. Icnofósseis. In: A.A. Sá, J.C.Gutiérrez-Marco (eds), Trilobites gigantes das ardósias de Canelas
(Arouca). Ardósias Valério and Figueiredo, Lda., Arouca: 163-180.Gutiérrez-Marco, J. C. and Sá, A. A., 2008. Nota preliminar sobre los
icnofosiles de la Formación Valongo (Ordovicico Medio) en la secciónde Canelas (Geoparque Arouca, norte de Portugal). In: J.I. Ruíz-Omeñaca, L. Piñuela and J.C. García-Ramos (eds), Libro de Resúmenes
de los XXIV Jornadas de la Sociedad Española de Paleontología.Museo del Jurásico de Asturias, Colunga: 37-38.
Gutiérrez-Marco, J. C., Sá, A. A., García-Bellido, D.C., Rábano, I. andValério, M., 2009. Giant trilobites and trilobite clusters from theOrdovician of Portugal. Geology, 37(5): 443-446.
Hagadorn, J. W. and Bottjer, D. J., 1999. Restriction of a LateNeoproterozoic Biotope: suspect-microbial structures and trace fossilsat the Vendian-Cambrian transition. Palaios, 14: 73-85.
Hertweck, G., Wehrmann, A. and Liebezeit, G., 2007. Bioturbationstructures of polychaetes in modern shallow marine environments andtheir analogues to Chondrites group traces. Palaeogeography,
Palaeoclimatology, Palaeoecology, 245: 382–389.Holmer, L. E., 1989. Middle Ordovician phosphatic inarticulate brachiopods
from Västergötland and Dalarna, Sweden. Fossils and Strata, 26: 1-172.Izumi, K., 2012. Formation process of the trace fossil Phymatoderma
granulata in the Lower Jurassic black shale (Posidonia Shale, southernGermany) and its paleoecological implications. Palaeog., Palaeocl.,
Palaeoecol., 353-355: 116-122.Jensen, S., Droser, M. L. and Gehling, J. G., 2006. A critical look at the
Ediacaran trace fossil record. In: S. Xiao and A.J. Kaufman (eds.),

36 C. Neto de Carvalho et al. / Comunicações Geológicas (2016) 103, Especial I, 23-38
Neoproterozoic Geobiology and Paleobiology. Springer Topics inGeobiology, 27: 115-157.
Kȩdzierski, M., Uchman, A., Sawlowicz, Z. and Briguglio, A., 2015. Fossilizedbioelectric wire - The trace fossil Trichichnus. Biogeosciences, 12:2301–2309.
Książkiewicz, M., 1970. Observations on the ichnofauna of the PolishCarpathians. In: T.P. Crimes and J.C. Harper (eds), Trace fossils.Liverpool: 283-322.
Książkiewicz, M., 1977. Trace fossils in the flysch of the Polish Carpathians.Paleont. Pol., P.A.N., Zaklad paleobiologii, Warszawa-Kraków: 1-208.
Levin, L., Rathburn, A., Gutierrez, D., Munoz, P. and Shankle, A., 2003.Bioturbation by symbiont-bearing annelids in near-anoxic sediments:Implications for biofacies models and paleo-oxygen assessments.Palaeogeography, Palaeoclimatology, Palaeoecology, 199: 129–140.
Levinton, J. and Kelaher, B., 2004. Opposing organizing forces of depositfeeding marine communities. Journal of Experimental Marine Biology
and Ecology, 300: 65-82.Llarena, J. G., 1946. Revision de algunos datos paleontologicos del flysch
Cretaceo y Numulitico de Guipuzcoa. Notas y Trabajos, IGME, 15: 113-163.Mángano, M. G. and Buatois, L. A., 2007. Trace fossils in evolutionary
palaeoecology. In: W. Miller III (ed.), Trace fossils: Concepts, Problems,
Prospects. Elsevier: 391-409.Mapstone, N. B. and Mcillroy, D., 2006. Ediacaran fossil preservation:
taphonomy and diagenesis of a discoid biota from the Amadeus Basin,central Australia. Precambrian Research, 149: 126-148.
Marenco, K. N. and Bottjer, D. J., 2008. The importance of Planolites in theCambrian substrate revolution. Palaeogeography, Palaeoclimatology,
Palaeoecology, 258: 189-199.Martin, K. D., 2004. A re-evaluation of the relationship between trace fossils
and dysoxia. In: D. McIlroy (ed.), The Application of Ichnology to
Palaeoenvironmental and Stratigraphic Analysis. Geological Society,London, Special Publications, 228: 141-156.
Mikuláš, R., 1995. Trace fossils from the Klabava formation, Czechoslovakia.Acta Universitatis Carolinae Geologica, 0(3-4): 385-419.
Monaco, P., Trecci, T. and Uchman, A., 2012. Taphonomy and ichnofabricof the trace fossil Avetoichnus luisae Uchman and Rattazzi, 2011 inPaleogene deep-sea fine-grained turbidites : examples from Italy, Polandand Spain. Bollettino della Società Paleontologica Italiana, 51: 23–38.
Narbonne, G. M., 2005. The Ediacara biota: Neoproterozoic origin ofanimals and their ecosystems. Annu. Rev. Earth Sci., 33: 421-442.
Orr, P. J., 2001. Colonization of the deep-marine environment during the earlyPhanerozoic: the ichnofaunal record. Geological Journal, 36: 265-278.
Ortoleva, P., 1994. Agates, geodes, concretions and orbicules; self-organizedzoning and morphology. Fractals and Dynamic Systems in Geoscience,283.
Oschmann, W., 2000. Microbes and Black Shales. In: R.E. Riding and S.M.Awramik (eds.), Microbial Sediments. Springer-Verlag, Berlin: 137-148.
Pazos, P. J., Gutiérrez, C., Fernández, D. E., Heredia, A. M. and Comerio,M., 2015. The unusual record of Nereites, wrinkle marks and undermatmining trace fossils from the late Silurian-earliest Devonian of central-western margin of Gondwana (Argentina). Palaeogeography,
Palaeoclimatology, Palaeoecology, 439: 4-16.Pemberton, S. G. and Frey, R. W., 1982. Trace fossil nomenclature and the
Planolites-Palaeophycus dilema. Journal of Paleontology, 56: 843-881.Pflüger, F., 1999. Matground structures and Redox facies. Palaios, 14: 25-39.Plička, M., 1989. Rotundusichnium zumayensis ichnogen. n. a new trace
fossil from the Outer Carpathian Flysch of West Slovakia(Czechoslovakia). Západné Karpaty, sér. Paleont., 13: 69-79.
Plotnick, R., 2007. Chemoreception, odor landscapes, and foraging inancient marine landscapes. Palaeontologia Electronica, 10(1): 1-11.
Raiswell, R., 1987. Non-steady state microbiological diagenesis and theorigin of concretions and nodular limestones. Geological Society
Special Publication, 36: 41-54.Raiswell, R., Whaler, K., Dean, S., Coleman, M. L. and Briggs, D. E. G.,
1993. A simple three-dimensional model of diffusion-with-precipitation
applied to localized pyrite formation in framboids, fossils and detritaliron minerals. Marine Geology, 113: 89-100.
Raumer, J. F. and Stampfli, G. M., 2008. The birth of the Rheic Ocean –Early Palaeozoic subsidence patterns and subsequent tectonic platescenarios. Tectonophysics, 461: 9-20.
Retallack, G. J., 2007. Decay, growth, and burial compaction of Dickinsonia,an iconic Ediacaran fossil. Alcheringa, 31: 215-240.
Retallack, G. J., 2015. Reassessment of the Silurian problematicum Rutgersella
as another post-Ediacaran vendobiont. Alcheringa, 39: 573-588.Rodrigues, N. P. C., Gil Cid, M. D., Arroyo, F., Huineman, M., Lara, R. and
Torices, A., 2005. Arachnostega gastrochaenae Bertling, 1992 en fósilesdel Dobrotiviense (Ordovícico Médio) de los Montes de Toledo (SO deEspaña). Revista Espanõla de Paleontología, 20(1): 57-64.
Sá, A. A. and Gutiérrez-Marco, J. C., 2006. Trilobites gigantes das ardósias de
Canelas (Arouca). Ardósias Valério and Figueiredo, Lda., Arouca, 205.Sá, A. A., Gutiérrez-Marco, J. C., Rábano, I. and Valério, M., 2007.
Palaeontology and stratigraphy of the Ordovician in the Arouca region(central Portugal). Acta Palaeont. Sinica, 46 (suppl.): 434-439.
Sá, A. A., Gutiérrez-Marco, J. C., Rocha, D., Valério, M., Brilha, J. andRábano, I., 2009. Ordovician ichnofossils: a new scientific andeducational resource for the Arouca Geopark. In: C. Neto de Carvalhoand J.C. Rodrigues (eds.), New Challenges with Geotourism, VIIIEuropean Geoparks Conference. Geopark Naturtejo da MesetaMeridional, Idanha-a-Nova: 140-142.
Savrda, C. E. and Bottjer, D. J., 1986. Trace-fossil model for reconstructionof paleo-oxygenation in bottom waters. Geology, 14(1): 3-6
Schieber, J., 2002. The role of an organic slime matrix in the formation ofpyritized burrow trails and pyrite concretions. Palaios, 17: 104-109.
Seilacher, A., 1977. Evolution of trace fossil communities in the deep sea. In.A. Hallam (ed.), Patterns of Evolution. Elsevier, Amsterdam: 359-376.
Seilacher, A., 1999. Biomat-related lifestyles in the Precambrian. Palaios,14(1): 86-93.
Seilacher, A., 2001. Concretion morphologies reflecting diagenetic andepigenetic pathways. Sedimentary Geology, 143: 41-57.
Seilacher, A., Grazhdankin, D. and Legouta, A., 2003. Ediacaran Biota: thedawn of animal life in the shadow of giant protists. Palaeontological
Research, 7: 43-54.Sepkoski, J. J. Jr., 1995. The Ordovician Radiation: diversification and
extinction shown by global-genus level taxonomic data. In: Cooper,J.D., Droser, M.L. and Finney, S.C., (eds.), Ordovician Odyssey: Short
papers for the Seventh International Symposium on the Ordovician
System. SEPM, California: 393-396.Sepkoski, J. J. and Sheehan, P. M., 1983. Diversification, faunal change, and
community replacement during the Ordovician radiations. In: M.J.S.Tevesz and P.J. McCall (eds.), Biotic Interactions in Recent and Fossil
Benthic Communities. Plenum Press, New York: 673–717. Servais, T., Owen, A. W., Harper, D. A. T., Kröger, B. and Munnecke, A.,
2010. The Great Ordovician Biodiversification Event (GOBE): thepalaeoecological dimension. Palaeogeography, Palaeoclimatology,
Palaeoecology, 294: 99-119.Shapiro, R. S., 2007. Stromatolites: a 3.5-billion-year ichnologic record. In:
W. Miller III (ed.), Trace fossils: Concepts, Problems, Prospects.Elsevier: 382-390.
Shourd, M. L. and Levin, H. L., 1976. Chondrites in the upper PlattinSubgoup (Middle Ordovician) of Eastern Missouri). Journal of
Paleontology, 50(2): 260-268.Southam, G., Donald, R., Röstad, A. and Brock, C., 2011. Pyrite discs in
coal: evidence for fossilized bacterial colonies. Geology, 29(1): 47-50.Stolz, J.E. (2000) - Structure of microbial mats and biofilms. In: R.E. Riding
and S.M. Awramik (eds.), Microbial Sediments. Springer-Verlag, Berlin:1-8.
Stoodley, P., Boyle, J. D. and Lappin-Scott, H. M., 1999. Influence of flowon the structure of bacterial films. In: C.R. Bell, M. Brylinky, P.Johnson-Green (eds), Microbial Biosystems: new frontiers. AtlanticCanadian Society for Microbial Ecology, 7.

Microbial colonies and ichnofossils in slates 37
Tarr, W.A., 1921. Syngenetic origin of concretion in shale. Bulletin of the
Geological Society of America, 32: 373-384.Thadeu, D., 1956. Note sur le Silurien Beiro-Durien. Bol. Soc. Geol. de
Portugal, 12: 1-38.Uchman, A., 1998. Taxonomy and ethology of flysch trace fossils: revision
of the Marian Ksiażkiewickz collection and studies of complementarymaterial. Annales Societatis Geologorum Poloniae, 68: 105-218.
Uchman, A., 1999. Ichnology of the Rhenodanubian Flysch (LowerCretaceous-Eocene) in Austria and Germany. Beringeria, 25: 67-173.
Uchman, A., 2001. Eocene flysch trace fossils from the Hecho Group of thePyrenees, northern Spain. Beringeria, 28: 3-41.
Uchman, A., 2004. Phanerozoic history of deep-sea trace fossils. In: D.McIlroy (ed.), The Application of Ichnology to Palaeoenvironmental
and Stratigraphic Analysis. Geological Society, London, SpecialPublications, 228: 125-139.
Uchman, A. and Gaździcki, A., 2010. Phymatoderma melvillensis isp. nov. andother trace fossils from the Cape Melville Formation (Lower Miocene) ofKing George Island, Antarctica. Polish Polar Research, 31: 83–99.
Van Roy, P., Orr, P. J., Botting, J. P., Muir, L. A., Vinther, J., Lefebvre, B.,Hariri, K. and Briggs, D. E. G., 2010. Ordovician faunas of BurgessShale type. Nature, 465: 215-218.
Weissberger, E. J., Coiro, L. L. and Davey, E. A., 2009. Effects of hypoxiaon animal burrow construction and consequent effects on sedimentredox profiles. Journal of Experimental Marine Biology and Ecology,371: 60-67.
Wetzel, A. and Uchman, A., 1998. Biogenic sedimentary structures inmudstones an overview. In: J. Schieber, W. Zimmerle and P. Sethi (eds.),Shales and Mudstones. I. E. Schweizerbart’sche Verlagsbuchhandlung:351-369.
Wetzel, A. and Uchman, A., 2013. Cladichnus parallelum isp. nov. – a mid-to-deep-tier feeding burrow. Ichnos, 20(3): 120-128.
Wilkin, R. T. and Barnes, H. L., 1996. Pyrite formation by reactions of ironmonosulfides with dissolved inorganic and organic sulfur species.Geochimica Cosmochimica Acta, 60: 4167–4179.
Zamora, S., Mayoral, E., Esteve, J., Gámez Vintaned, J.A. and Santos, A.,2011. Exoskeletal abnormalities in paradoxidid trilobites from theCambrian of Spain, and a new type of bite trace. Bulletin of
Geosciences, 86(3): 665-673.Zangeil, R., Woodland, B. G., Richardson, E. S. and Zachary, D. L., 1969. Early
diagenetic phenomena in the Fayetteville black shale (Mississipian) ofArkansas. Sedimentary Geology, 3: 87-119.
Zessin, W., 2011. Ein rätselhaftes fossils Objekt (Laotina silurica Huene,1904) aus dem Ordovizium von Schweden und aus dem GeschiebeMecklenburgs. Mitteilungen der Naturforschenden Gesellschaft
Mecklenburg, 11(1): 53-59.Zhang, Y. D., Zhan, R. B., Fan, J. X., Cheng, J. F and Xiao; L., 2010.
Principal aspects of the Ordovician biotic radiation. Science China
Earth Sciences, 53: 382-394.
