microarray analysis of in vitro pericyte differentiation reveals an angiogenic program of gene...
TRANSCRIPT

©2004 FASEB
The FASEB Journal express article10.1096/fj.04-1604fje. Published online December 3, 2004.
Microarray analysis of in vitro pericyte differentiation reveals an angiogenic program of gene expression Sujata Kale,*,† Jun-ichi Hanai,* Barden Chan,* Anil Karihaloo,† Gary Grotendorst,‡, 1 Lloyd G. Cantley,† and Vikas P. Sukhatme* *Divisions of Nephrology and Hematology-Oncology and Center for Study of the Tumor Microenvironment, Beth Israel Deaconess Medical Center and Harvard Medical School, Boston, MA; †Division of Nephrology Yale University School of Medicine, New Haven, CT; and ‡Department of Cell Biology and Anatomy, University of Miami, Florida.
1Present address for G. Grotendorst: Lovelace Respiratory Research Institute, 2425 Ridgecrest Drive SE, Albquerque, NM 87108 Corresponding author: Vikas P. Sukhatme, 330 Brookline Ave., RW 563, Beth Israel Deaconess Medical Center, Boston, MA 02215. E-mail: [email protected]
ABSTRACT
The vasculature consists of endothelial cells (ECs) lined by pericyte/vascular smooth muscle cells (vSMCs). Pericyte/vSMCs provide support to the mature vasculature but are also essential for normal blood vessel development. To determine how pericyte-EC communication influences vascular development, we used the well-established in vitro model of TGFβ-stimulated differentiation of 10T1/2 cells into pericyte/vSMCs. Microarray analysis was performed to identify genes that were differentially expressed by induced vs. uninduced 10T1/2 cells. We discovered that these cells show an angiogenic program of gene expression, with up-regulation of several genes previously implicated in angiogenesis, including VEGF, IL-6, VEGF-C, HB-EGF, CTGF, tenascin C, integrin α5, and Eph receptor A2. Up-regulation of some genes was validated by Western blots and immunocytochemistry. We also examined the functional significance of these gene expression changes. VEGF and IL-6 alone and in combination were important in 10T1/2 cell differentiation. Furthermore, we used a coculture system of 10T1/2 and human umbilical vein ECs (HUVECs), resulting in the formation of cordlike structures by the HUVECs. This cordlike structure formation was disrupted when neutralizing antibodies to VEGF or IL-6 were added to the coculture system. The results of these studies show that factors produced by pericytes may be responsible for recruiting ECs and promoting angiogenesis. Therefore, a further understanding of the genes involved in pericyte differentiation could provide a novel approach for developing anti-angiogenic therapies.
Key words: VEGF • angiogenesis • endothelial cells • vasculogenesis
Page 1 of 30(page number not for citation purposes)

ngiogenesis is the process in which new blood vessels develop from a preexisting vascular network (1). This process is of fundamental importance for several physiological and pathological events such as organ development and tumor growth (2, 3). Blood vessels are relatively simple structures composed of three cell types.
Endothelial cells (ECs) form the inner luminal lining, mural cells (pericytes/vascular smooth muscle cells [vSMCs]) constitute the medial layer, and fibroblasts make up the outer adventitial layer (4). In contrast to angiogenesis, vasculogenesis begins with the de novo differentiation of angioblasts to ECs, which subsequently form tubelike structures (1, 5).
In the vasculature, ECs and pericyte/vSMCs are closely apposed to each other and therefore interact with and influence each other (6). The nature of such interactions has largely been studied in vitro. Such analysis suggests a role for the chemoattractant and mitogenic properties of several soluble factors known to be produced by ECs such as platelet-derived growth factor (PDGF) (7), basic fibroblast growth factor (bFGF) (8), and heparin binding-epidermal growth factor (HB-EGF) (9). The ECs of the developing vessels produce PDGF-BB, whereas PDGF receptor β is expressed by developing SMCs and the surrounding mesenchyme (10). Pericyte/vSMCs also produce factors that may have an effect on the proliferation and survival of ECs. Heterotypic cell-cell contact as well as soluble factors produced have been shown to influence EC survival and proliferation (11). To date, little information is available regarding the genes expressed when mesenchymal cells differentiate to pericytes. To better understand the role of pericytes in the process of angiogenesis, we used a well-established system of differentiation of 10T1/2 cells to pericytes (12). Treatment with TGFβ induces the 10T1/2 cells to differentiate into pericytes as evidenced by the expression of α smooth muscle actin (αSMA), tropomyosin, desmin, and calponin. Using microarray analysis, we have identified genes induced during the process of pericyte differentiation that regulate subsequent angiogenesis.
MATERIALS AND METHODS
Cell culture
C3H10T1/2 cells (abbreviated as 10T1/2 cells) were purchased from ATCC and maintained in Dulbecco’s modified Eagle’s medium (DMEM), supplemented with 10% heat-inactivated FBS. Cells were maintained at a confluence of 80–90% and were used only up to passage 15. HUVECs were purchased from Clonetics (San Diego, CA). Cells were maintained in EGM2-MV complete medium (Clonetics) containing endothelial basal medium (EBM-2), supplemented with 5% fetal bovine serum, gentamycin, amphotericin B, hydrocortisone, ascorbic acid, and the following growth factors: VEGF, bFGF, EGF, and IGF-1. All cell lines were grown at 37°C in a 100% humidified incubator with 5% CO2. To study the differentiation of 10T1/2 cells to pericyte/vSMCs, cells were cultured in the presence of 1 ng/ml rTGFβ1 for various time points. Cells were cultured in eight-well chamber slides at a density of 8000 cells/well/250 µl medium with or without TGF β, 1 ng/ml from day 1 through day 7.
Immunostaining
Cells were stained for the various pericyte/SMC markers using the ABC immunostaining procedure. The primary antibodies used were anti-αSMA (DAKO, Carpinteria, CA]; 1:50
A
Page 2 of 30(page number not for citation purposes)

dilution), anti-desmin (DAKO; 1:50 dilution), anti-tropomyosin (Sigma, St. Louis, MO; 1:100), anti-calponin (Sigma; 1:40), and anti-APN (PharMingen, San Diego, CA). In addition, an isotype control was used for each antibody. For gene validation, we used antibodies to EphA2 (Upstate Biotechnology, Waltham, MA; 10 µg/ml), anti-CTGF (from G. Grotendorst; 1:50), anti-VEGF (Research Diagnostics, Minneapolis, MN; 10 µg/ml), anti-mouse integrin α5 (PharMingen; 10 µg/ml), anti-mouse tenascin (Sigma; 1:200), and anti-HB-EGF (M. Klagsbrun, Children’s Hospital, Harvard Medical School, Boston, MA). This was followed by the secondary antibody (Sigma; anti-mouse IgG, biotin conjugated, 1:300) and then use of the ABC-HRP (DAKO). The substrate used was diaminobenzidine (DAKO) reagent, resulting in a brown reaction product. Each experiment was performed at least three times.
Cocultures
HUVECs and 10T1/2 cells were initially cultured in their respective medium. For cocultures, HUVECs and 10T1/2 cells were mixed in varying ratios (1:1, 2:1, 3:1, 10:1, 20:1, 40:1) in EGM-2 MV medium. Cells were cultured in eight-well chamber slides, fixed after 72 h, and sequentially stained for EC and pericyte markers. Cells were initially stained using the mouse anti-human P1H12 antibody (Chemicon, Temecula, CA) for ECs. Biotin-tyramide was used to amplify the signal, followed by ABC-AP. The substrate used was new fuchsin, and the reaction product was red. Alpha SMA staining for pericytes followed this. A biotinylated secondary antibody was used followed by ABC-HRP and diaminobenzidine as the substrate. The reaction product was brown.
RNA extraction
10T1/2 cells were cultured in the presence of TGFβ1 for 48 h. RNA was extracted from these cells using the RNA-STAT reagent (Tel-Test Inc., Friendswood, Texas). RNA was extracted from uninduced cells (t=0) after 48 h of culture in the presence of TGFβ1, as per the manufacturer’s instructions. The extracted RNA was then cleaned using RNeasy (Qiagen, Valencia, CA). RNA was determined to be of sufficient quality by gel analysis and if the OD260/OD280 ratio was between 1.9 and 2.1. RNA was subjected to microarray analysis using the murine U74A chips from Affymetrix.
Functional significance of genes analyzed by DNA microarray
To determine the importance of VEGF, IL-6, and integrin α5 on pericyte differentiation and on pericyte-HUVEC cord formation, neutralizing antibodies to murine VEGF, IL-6, and integrin α5 were added either individually or in combination to TGFβ1-induced 10T1/2 cell cultures, as well as cocultures of HUVEC and 10T1/2 cells. GS was used as the control for anti-VEGF and anti-IL-6. Rat IgG2a was the isotype control for anti-integrin α5. Each of the following were added each day to the culture: 4 µg/ml anti-VEGF, 2 µg/ml of anti-IL-6, and 20 µg/ml of anti-integrin α5 antibodies.
Page 3 of 30(page number not for citation purposes)

SDS-PAGE and Western blot
Cell lysates were prepared and separated by PAGE (precast gels, Bio-Rad, Hercules, CA). This was followed by electroblotting onto a polyvinylidenedifluoride (PVDF) membrane. After blocking with 5% BSA in Tris-buffered saline/Tween-20 (TTBS) for 1 h, the membrane was incubated overnight with either anti-SMA antibody, anti-EphA2, anti-CTGF, or anti-VEGF antibody. After washing with TTBS, the membrane was incubated with a horseradish peroxidase-conjugated secondary antibody (Amersham, Arlington Heights, IL; 1:5000 dilution) for 1 h. Protein bands were detected using SuperSignal West Pico Chemiluminescent Substrate (Pierce, Rockford, IL).
RESULTS
Differentiation of 10T1/2 cells
The 10T/1/2 cell system makes use of a multipotent mouse embryonic cell line as a presumptive precursor to pericyte/SMCs (12). These cells can be induced to undergo differentiation to pericyte/SMCs by TGFβ1. 10T1/2 cells were cultured in eight-well chamber slides in the presence of TGFβ1. As described by D’Amore et al. (12), these cells changed their shape from polygonal to spindle within 24 h, with up-regulation of αSMA expression. In our experiments, maximum αSMA expression was seen at 48 h of culture (Fig. 1A). Since αSMA alone cannot be used as a marker of pericyte/SMC differentiation, we also assessed expression of other pericyte/SMC markers: calponin (Fig. 1B), desmin (Fig. 1C), and tropomyosin (Fig. 1D). Expression of all of these markers was noted after 48 h in culture. Earlier studies have suggested that “activated pericytes,” that is, pericytes at the sites of active angiogenesis, express aminopeptidase A (APA) (13), aminopeptidase N (APN) (14), and N-cadherin (15). We assessed expression of APA and APN in induced 10T1/2 cell cultures using immunocytochemistry. We did not observe any expression of APA. However, APN first appeared at 24 h of culture and peaked at 48 h of culture (Fig. 1E), with 60–70% positive cells.
The importance of TGFβ1 in maintenance of the differentiated state of the induced pericytes was examined next. 10T1/2 cells were cultured in the presence of TGFβ1 for 48 h in duplicate wells. At this time point, one set was stained to confirm that cells had differentiated to pericyte/SMCs. For the second set of cultures, the medium was removed and cells were refed without TGFβ1 for 3 days. The differentiated state of the cells was then determined both microscopically and by αSMA expression. In the absence of TGFβ1, cells reversed their differentiated phenotype (Fig. 1), as seen by a change in the cell shape from spindle back to polygonal, and a decrease in αSMA expression back to that of undifferentiated cells.
Microarray analysis of genes expressed during the differentiation of 10T1/2 cells
Microarray analysis of genes expressed by 10T1/2 cells at 48 h of culture in the presence of TGFβ was performed using the murine Affymetrix chips. Genes that were either up-regulated or down-regulated fourfold or more were determined. This gene group is as shown in Table 1. Of these genes, those shown in bold were selected for further analysis on the basis of reagent availability and a known role in angiogenesis; other genes involved in angiogenesis as
Page 4 of 30(page number not for citation purposes)

ascertained by Pubmed search (on words “angiogenesis” and “gene of interest’) are indicated with a asterisk (see Table 1).
VEGF-A is one of the most important growth and survival factors for endothelium (3). It causes proliferation of ECs, induces angiogenesis, regulates vasculogenesis, is induced by hypoxia, and can stimulate cell migration and inhibit apoptosis (16). It is also a mitogen for pericytes under hypoxic conditions (17).
IL-6 is a proinflammatory cytokine produced by macrophages (18). It induces secretion of VEGF in cervical cancer cells (19). Substantially high microenvironmental IL-6 levels promote tumor angiogenesis and the development of cervical cancer (19). Overexpression of IL-6 in basal cell carcinoma cell lines decreases apoptosis and increases tumorigenic potency (20).
Heparin binding epidermal growth factor-like growth factor (HB-EGF) is an EGF-related peptide with prominent effects on cell growth and migration (21). Cultured mesangial cells synthesize HB-EGF and is a potent mitogen for SMCs (22).
VEGF-C can be stimulated by serum, PDGF, EGF, and TGFβ (23) and may play a role in tumor progression via lymphangiogenesis (24) and angiogenesis in human esophageal carcinoma.
EphA2 receptor tyrosine kinases/Eck are the Eph family of receptors, the ligands for which are ephrins. EphA2 is particularly involved in tubular network formation by aggressive melanoma cells in vitro (25). Recent studies have localized EphA receptors to the vasculature (26).
Integrin α5 has recently been shown to play a significant role in vessel formation, both in embryonic stem cell cultures and in teratocarcinomas (27).
Tenascin C is an extracellular matrix protein that may be involved in EC activation and may also play a role in tumor angiogenesis (22). Its increased expression in tumor vessels correlates with a high proliferative index of the tumor (28, 29). Sprouting and cord-forming ECs specifically express tenascin C in vitro (30).
In addition, we elected to study another gene, CTGF, an ECM-associated heparin binding protein known to be specifically induced by TGFβ. VEGF also induces CTGF expression in ECs and pericytes (31) and is involved in their proliferation and migration (32, 33). It also plays a role in EC survival (34). CTGF is a potent angiogenesis factor inducing tube formation of vECs in vitro and neovascularization in vivo (35).
Validation of gene regulation
Up-regulation of several of the modulated genes was validated using immunocytochemistry as well as Western analysis. 10T1/2 cells were cultured with or without TGFβ1 for 48 h. By immunocytochemical staining, we confirmed up-regulation of the following proteins: connective tissue growth factor (CTGF) (Fig. 2A), HB-EGF (Fig. 2B), VEGF-A (Fig. 2C), EphA2 (Fig. 2D), integrin α5 (Fig. 2E), tenascin (Fig. 2F). Integrin α5 (Fig. 2E) and tenascin (Fig. 2F) were expressed only in the presence of TGFβ1, with no detectable expression in uninduced cells. Cells showed a basal low-level expression of VEGF (Fig. 2C) that was enhanced in the presence of
Page 5 of 30(page number not for citation purposes)

TGFβ1. CTGF (Fig. 2A), HB-EGF (Fig. 2B), and EphA2 (Fig. 2D) expression was also increased in the presence of TGFβ1. Western analysis confirmed TGFβ-induced expression of the following genes: CTGF, VEGF-A, and EphA2 (Fig. 3). CTGF was expressed at a very low level in uninduced cells with a two- to threefold increase in the presence of TGFβ1. VEGF-A and EphA2 were increased several fold by TGFβ1 treatment.
Evidence for functional significance of modulated genes
The importance of several of these factors to act in a paracrine or autocrine manner during pericyte differentiation was examined. To assess a potential autocrine effect of VEGF, IL-6, or integrin α5 on 10T1/2 cell differentiation, we cultured the cells in the presence of TGFβ1 with or without neutralizing antibodies to VEGF-A, IL-6, or integrin α5. Differentiation was assessed by Western blotting for αSMA. In the presence of anti-VEGF-A, there was a 25–30% decrease in αSMA expression (Fig. 4), whereas neutralizing antibodies to IL-6 and integrin α5 failed to prevent 10T1/2 cell differentiation. These results suggest that VEGF may play a role in 10T1/2 cell differentiation to pericytes.
To examine whether factors produced by differentiating pericytes have a paracrine effect on adjacent ECs, we used a 2-D coculture system. In this system, human umbilical vein ECs (HUVECs) were cultured with murine10T1/2 cells, leading to the formation of cordlike structures. The addition of either anti-VEGF antibody (Fig. 5A) or anti-IL-6 antibody (Fig. 5B) to the coculture inhibited in vitro angiogenesis by ~50–60% when compared with goat serum (GS) control. While anti-VEGF-A and anti-IL-6 could each individually affect the formation of cordlike structures, the largest inhibitory effect was seen when anti-VEGF and anti-IL-6 were added together (Fig. 5C) Anti-integrin α5 did not have any inhibitory effect (Fig. 5D) when compared with isotype control antibody. To address the possibility that the neutralizing antibodies affected survival of ECs, we treated isolated cultures of HUVECs with these antibodies (Fig. 5E) and noted no deleterious effects on the EC survival.
Effects on 10T1/2 cell differentiation in cocultures of HUVECs and 10T1/2 cells
The results of our HUVEC and 10T1/2 cell coculture experiments indicated that pericytes induce ECs to form cordlike structures. To determine whether ECs reciprocally influence 10T1/2 cells in these cocultures, we examined expression of αSMA by immunocytochemistry. 10T1/2 cells in contact with HUVECs showed a change in cell shape as well as increased expression of αSMA (Fig. 6), indicating that EC-10T1/2 cell contact can induce pericyte differentiation. Neutralizing antibodies to murine VEGF-A, IL-6, and integrin α5 resulted in a 50–70% decrease in αSMA expression (Fig. 6). These results demonstrate that factors produced by pericytes function in an autocrine as well as a paracrine manner. DISCUSSION Angiogenesis is a complex process that involves several sequential events. These include invasion of ECs into connective tissue, migration, proliferation, cell-cell association, and lumen formation. The ECs then associate with pericyte/SMCs, thereby leading to the formation of mature, stable vasculature (36). Several studies have suggested the importance of pericytes in
Page 6 of 30(page number not for citation purposes)

angiogenesis (6, 37–41). In this study, we evaluated the gene expression pattern for differentiated pericytes from precursor 10T1/2 cells.
The in vitro model used to study this process was that of 10T1/2 cells differentiating to pericytes in the presence of TGFβ1 (12). Cells changed from hexagonal to spindle shape with an increase in expression of αSMA as well as other pericyte markers. This was consistent with previous results of D’Amore et al. (12).
ECs and pericytes closely interact with each other (42). A number of autocrine and paracrine factors are produced that have been implicated in vessel assembly and growth. Cell-cell interactions have been shown to mediate differentiation and stabilization of developing tissue in several other model systems as well (43–45). Interestingly, in the developing nervous system, heterotypic interactions between the innervating neuron and the target cell affect the survival and differentiation of both cell types (46). Among the factors produced in the vasculature, VEGF is a potent mitogen as well as permeability factor for ECs. We now find that this factor is produced not only by ECs but also by pericytes.
As discussed in the results, several genes were up-regulated during the differentiation of 10T1/2 cells to pericytes. Some of the genes that were up-regulated in addition to VEGF-A include VEGF-C, tenascin-C, HB-EGF, EphA2, IL-6, and integrin α5. Using various experimental models, each of these genes has been previously implicated in angiogenesis. However, our results demonstrate for the first time that perciytes alone express all these genes during their differentiation process.
The regulation of pericyte growth in vivo has been poorly understood. However, genetic studies using knockout mice have indicated several genes to be important, namely, dHAND (47), PDGF-BB as well as PDGF-R-β (48, 49), and LKLF (50, 51). A G protein-coupled receptor, Edg-1, is also important for recruiting mural cells to the vessel wall (52).
Autocrine and paracrine mechanisms in vascular development
Several autocrine and paracrine factors have been shown to be important in angiogenesis (3, 32, 53). Addition of TGFβ to 10T1/2 cells promotes their differentiation to pericytes (12). This effect of TGFβ on 10T1/2 cell differentiation is reversed if this factor is removed. This would suggest that once the angiogenic program is turned on by TGFβ, removal of TGFβ not only turns off the angiogenic program but reverts the pericytes to their original undifferentiated state. It is conceivable that modulation of TGFβ in a tumor microenvironment could explain the net pro-/anti-angiogenic effect of this molecule (54, 55).
Our microarray analysis results suggest that VEGF-A might act in an autocrine fashion, partly promoting the differentiation of 10T1/2 cells. Blocking VEGF-A activity using neutralizing antibodies resulted in decreased αSMA expression in 10T1/2 cells despite the presence of TGFβ1. This indicates that VEGF-A plays an important role in the differentiation of pericytes. This further suggests that VEGF-A produced by pericytes, acts in an autocrine as well as paracrine manner.
Page 7 of 30(page number not for citation purposes)

Cell-cell interactions are critical in the formation of a stable mature vessel
We used a 2-D coculture system to assess the effects of heterotypic cell-cell interactions between ECs and pericytes. In this system, TGFβ produced by ECs causes differentiation of 10T1/2 cells (12). In the current study, when HUVECs were cultured alone, they did not form cordlike structures. Also, 10T1/2 cells cultured alone did not differentiate to pericytes. When cultured together, HUVECs formed cordlike structures and 10T1/2 cells differentiated to pericytes. The optimum ratio of HUVECs:10T1/2 cells in these cocultures was 2:1 or 3:1. At higher concentrations of 10T1/2 cells, no such cordlike structures were observed, indicating that 10T1/2 cells might secrete an inhibitory factor that probably reaches a critical concentration as the number of 10T1/2 cells is increased. However, we have not attempted to isolate this factor. Also, when the number of HUVECs was increased, we did not see any cordlike structure formation, indicating that the ratio of ECs to 10T1/2 cells is critical to promote cordlike structure formation in this system. This data might be extrapolated to the in vivo situation to suggest that a critical balance between pro- and anti-angiogenic factors is important in promoting angiogenesis.
We assessed the role of several of the gene products up-regulated in pericyte differentiation (VEGF-A, IL-6, and integrin α5) in the process of cord formation by ECs. We used EGM2-MV complete medium that was supplemented with human growth factors. The 10T1/2 cells are murine cells, and therefore for studying the role of factors important in cord formation by ECs, we used neutralizing antibodies to the murine VEGF, IL-6, and integrin α5. Our results indicate that the inhibition of cord formation is not due to the neutralization of VEGF, which serves as a survival factor for ECs. HUVECs cultured alone and treated with the neutralizing antibodies at the same concentrations did not affect the proliferation of HUVEC cells, suggesting that these factors might act indirectly inducing secretion of others factors and/or that cell-cell interaction between ECs and pericytes was important for this process. This suggestion was strengthened by the results of our coculture experiments in which addition of neutralizing antibodies to VEGF-A and IL-6 decreased the percentage of αSMA-expressing cells and further caused inhibition of cordlike structure formation by ECs. The results of coculture experiments suggest that pericytes influence ECs either directly via cell-cell contact and/or indirectly via secreted factors and might “lead” in the angiogenic sprout. This finding is further substantiated by recent work of Darland et al. They show that differentiated pericytes produce VEGF that may act in a juxtacrine/paracrine manner and as a survival and/or stabilizing factor for ECs in microvessels. (56). Further, it was recently shown that pericytes form a sheath around the ECs and that this sheath extended beyond the EC. Interestingly, pericytes regularly accompany endothelial sprouts on growing blood vessels and appear to be involved in sprout formation (41) in determining the location of sprouts as well as in guiding the outgrowth of sprouts (57, 58). Pericytes are located on the growing front of the tumor vasculature where active angiogenesis is occurring (41). Pericytes are among the first cells to invade the newly vascularized corpus luteum (59) and are also the ones to proliferate early on in tumor angiogenesis (60). However, pericyte coverage is variable in tumors. Based on our microarray data as well as these observations by others, pericytes might play a significant role in blood vessel growth in some tumors in which there is substantial pericyte coverage, and would therefore be a potential target in anti-angiogenic therapy in these cases (61, 62). Indeed, very recent data targeting pericytes (via blockade of PDGF signaling) in conjunction with anti-
Page 8 of 30(page number not for citation purposes)

endothelial therapy suggest that such combination therapy may be particularly efficacious (56, 63). A limitation of this study is that our in vitro data will need to be verified in vivo. ACKNOWLEDGMENTS We thank the members of the Sukhatme Laboratory for valuable discussions. This work was supported in part from seed funds from the Department of Medicine at Beth Israel Deaconess Medical Center and a research grant from Gambro to VPS. REFERENCES
1. Risau, W. (1997) Mechanisms of angiogenesis. Nature 386, 671–674
2. Holmgren, L., O'Reilly, M. S., and Folkman, J. (1995) Dormancy of micrometastases: balanced proliferation and apoptosis in the presence of angiogenesis suppression. Nat. Med. 1, 149–153
3. Ferrara, N. (2000) Vascular endothelial growth factor and the regulation of angiogenesis. Recent Prog. Horm. Res. 55, 15–35
4. Darland, D. C., and D'Amore, P. A. (1999) Blood vessel maturation: vascular development comes of age. J. Clin. Invest. 103, 157–158
5. Yancopoulos, G. D., Klagsbrun, M., and Folkman, J. (1998) Vasculogenesis, angiogenesis, and growth factors: ephrins enter the fray at the border. Cell 93, 661–664
6. Hirschi, K. K., and D'Amore, P. A. (1997) Control of angiogenesis by the pericyte: molecular mechanisms and significance. EXS 79, 419–428
7. Westermark, B., Siegbahn, A., Heldin, C. H., and Claesson-Welsh, L. (1990) B-type receptor for platelet-derived growth factor mediates a chemotactic response by means of ligand-induced activation of the receptor protein-tyrosine kinase. Proc. Natl. Acad. Sci. USA 87, 128–132
8. Montesano, R., Vassalli, J. D., Baird, A., Guillemin, R., and Orci, L. (1986) Basic fibroblast growth factor induces angiogenesis in vitro. Proc. Natl. Acad. Sci. USA 83, 7297–7301
9. Higashiyama, S., Abraham, J. A., and Klagsbrun, M. (1993) Heparin-binding EGF-like growth factor stimulation of smooth muscle cell migration: dependence on interactions with cell surface heparan sulfate. J. Cell Biol. 122, 933–940
10. Holmgren, L., Claesson-Welsh, L., Heldin, C. H., and Ohlsson, R. (1992) The expression of PDGF alpha- and beta-receptors in subpopulations of PDGF-producing cells implicates autocrine stimulatory loops in the control of proliferation in cytotrophoblasts that have invaded the maternal endometrium. Growth Factors 6, 219–231
Page 9 of 30(page number not for citation purposes)

11. Hirschi, K. K., Rohovsky, S. A., Beck, L. H., Smith, S. R., and D'Amore, P. A. (1999) Endothelial cells modulate the proliferation of mural cell precursors via platelet-derived growth factor-BB and heterotypic cell contact. Circ. Res. 84, 298–305
12. Hirschi, K. K., Rohovsky, S. A., and D'Amore, P. A. (1998) PDGF, TGF-beta, and heterotypic cell-cell interactions mediate endothelial cell-induced recruitment of 10T1/2 cells and their differentiation to a smooth muscle fate. J. Cell Biol. 141, 805–814
13. Schlingemann, R. O., Oosterwijk, E., Wesseling, P., Rietveld, F. J., and Ruiter, D. J. (1996) Aminopeptidase a is a constituent of activated pericytes in angiogenesis. J. Pathol. 179, 436–442
14. Alliot, F., Rutin, J., Leenen, P. J., and Pessac, B. (1999) Pericytes and periendothelial cells of brain parenchyma vessels co- express aminopeptidase N, aminopeptidase A, and nestin. J. Neurosci. Res. 58, 367–378
15. Gerhardt, H., Wolburg, H., and Redies, C. (2000) N-cadherin mediates pericytic-endothelial interaction during brain angiogenesis in the chicken. Dev. Dyn. 218, 472–479
16. Liu, W., Ahmad, S. A., Reinmuth, N., Shaheen, R. M., Jung, Y. D., Fan, F., and Ellis, L. M. (2000) Endothelial cell survival and apoptosis in the tumor vasculature. Apoptosis 5, 323–328
17. Reinmuth, N., Liu, W., Jung, Y. D., Ahmad, S. A., Shaheen, R. M., Fan, F., Bucana, C. D., McMahon, G., Gallick, G. E., and Ellis, L. M. (2001) Induction of VEGF in perivascular cells defines a potential paracrine mechanism for endothelial cell survival. FASEB J. 15, 1239–1241
18. Breviario, F., d'Aniello, E. M., Golay, J., Peri, G., Bottazzi, B., Bairoch, A., Saccone, S., Marzella, R., Predazzi, V., Rocchi, M., et al. (1992) Interleukin-1-inducible genes in endothelial cells. Cloning of a new gene related to C-reactive protein and serum amyloid P component. J. Biol. Chem. 267, 22190–22197
19. Wei, L. H., Kuo, M. L., Chen, C. A., Cheng, W. F., Cheng, S. P., Hsieh, F. J., and Hsieh, C. Y. (2001) Interleukin-6 in cervical cancer: the relationship with vascular endothelial growth factor. Gynecol. Oncol. 82, 49–56
20. Jee, S. H., Shen, S. C., Chiu, H. C., Tsai, W. L., and Kuo, M. L. (2001) Overexpression of interleukin-6 in human basal cell carcinoma cell lines increases anti-apoptotic activity and tumorigenic potency. Oncogene 20, 198–208
21. Swinscoe, J. C., and Carlson, E. C. (1992) Capillary endothelial cells secrete a heparin-binding mitogen for pericytes. J. Cell Sci. 103, 453–461
22. Polihronis, M., Murphy, B. F., Pearse, M. J., and Power, D. A. (1996) Heparin-binding epidermal growth factor-like growth factor, an immediate-early gene for mesangial cells, is up-regulated in the Thy- 1.1 model. Exp. Nephrol. 4, 271–278
Page 10 of 30(page number not for citation purposes)

23. Enholm, B., Paavonen, K., Ristimaki, A., Kumar, V., Gunji, Y., Klefstrom, J., Kivinen, L., Laiho, M., Olofsson, B., Joukov, V., et al. (1997) Comparison of VEGF, VEGF-B, VEGF-C and Ang-1 mRNA regulation by serum, growth factors, oncoproteins and hypoxia. Oncogene 14, 2475–2483
24. Valtola, R., Salven, P., Heikkila, P., Taipale, J., Joensuu, H., Rehn, M., Pihlajaniemi, T., Weich, H., deWaal, R., and Alitalo, K. (1999) VEGFR-3 and its ligand VEGF-C are associated with angiogenesis in breast cancer. Am. J. Pathol. 154, 1381–1390
25. Hess, A. R., Seftor, E. A., Gardner, L. M., Carles-Kinch, K., Schneider, G. B., Seftor, R. E., Kinch, M. S., and Hendrix, M. J. (2001) Molecular regulation of tumor cell vasculogenic mimicry by tyrosine phosphorylation: role of epithelial cell kinase (Eck/EphA2). Cancer Res. 61, 3250–3255
26. Bovenkamp, D. E., and Greer, P. A. (2001) Degenerate PCR-based cloning method for Eph receptors and analysis of their expression in the developing murine central nervous system and vasculature. DNA Cell Biol. 20, 203–213
27. Taverna, D., and Hynes, R. O. (2001) Reduced blood vessel formation and tumor growth in alpha5-integrin- negative teratocarcinomas and embryoid bodies. Cancer Res. 61, 5255–5261
28. Tokes, A. M., Hortovanyi, E., Kulka, J., Jackel, M., Kerenyi, T., and Kadar, A. (1999) Tenascin expression and angiogenesis in breast cancers. Pathol. Res. Pract. 195, 821–828
29. Vacca, A., Ribatti, D., Fanelli, M., Costantino, F., Nico, B., Di Stefano, R., Serio, G., and Dammacco, F. (1996) Expression of tenascin is related to histologic malignancy and angiogenesis in b-cell non-Hodgkin's lymphomas. Leuk. Lymphoma 22, 473–481
30. Schenk, S., Chiquet-Ehrismann, R., and Battegay, E. J. (1999) The fibrinogen globe of tenascin-C promotes basic fibroblast growth factor-induced endothelial cell elongation. Mol. Biol. Cell 10, 2933–2943
31. Suzuma, K., Naruse, K., Suzuma, I., Takahara, N., Ueki, K., Aiello, L. P., and King, G. L. (2000) Vascular endothelial growth factor induces expression of connective tissue growth factor via KDR, Flt1, and phosphatidylinositol 3-kinase- akt-dependent pathways in retinal vascular cells. J. Biol. Chem. 275, 40725–40731
32. Fan, W. H., Pech, M., and Karnovsky, M. J. (2000) Connective tissue growth factor (CTGF) stimulates vascular smooth muscle cell growth and migration in vitro. Eur. J. Cell Biol. 79, 915–923
33. Moussad, E. E., and Brigstock, D. R. (2000) Connective tissue growth factor: what's in a name? Mol. Genet. Metab. 71, 276–292
34. Babic, A. M., Chen, C. C., and Lau, L. F. (1999) Fisp12/mouse connective tissue growth factor mediates endothelial cell adhesion and migration through integrin alphavbeta3,
Page 11 of 30(page number not for citation purposes)

promotes endothelial cell survival, and induces angiogenesis in vivo. Mol. Cell. Biol. 19, 2958–2966
35. Shimo, T., Nakanishi, T., Nishida, T., Asano, M., Kanyama, M., Kuboki, T., Tamatani, T., Tezuka, K., Takemura, M., Matsumura, T., et al. (1999) Connective tissue growth factor induces the proliferation, migration, and tube formation of vascular endothelial cells in vitro, and angiogenesis in vivo. J. Biochem. (Tokyo) 126, 137–145
36. Gerhardt, H., and Betsholtz, C. (2003) Endothelial-pericyte interactions in angiogenesis. Cell Tissue Res. 314, 15–23
37. Schlingemann, R. O., Rietveld, F. J., de Waal, R. M., Bradley, N. J., Skene, A. I., Davies, A. J., Greaves, M. F., Denekamp, J., and Ruiter, D. J. (1990) Leukocyte antigen CD34 is expressed by a subset of cultured endothelial cells and on endothelial abluminal microprocesses in the tumor stroma. Lab. Invest. 62, 690–696
38. Schlingemann, R. O., Rietveld, F. J., de Waal, R. M., Ferrone, S., and Ruiter, D. J. (1990) Expression of the high molecular weight melanoma-associated antigen by pericytes during angiogenesis in tumors and in healing wounds. Am. J. Pathol. 136, 1393–1405
39. Schlingemann, R. O., Rietveld, F. J., Kwaspen, F., van de Kerkhof, P. C., de Waal, R. M., and Ruiter, D. J. (1991) Differential expression of markers for endothelial cells, pericytes, and basal lamina in the microvasculature of tumors and granulation tissue. Am. J. Pathol. 138, 1335–1347
40. Wesseling, P., Schlingemann, R. O., Rietveld, F. J., Link, M., Burger, P. C., and Ruiter, D. J. (1995) Early and extensive contribution of pericytes/vascular smooth muscle cells to microvascular proliferation in glioblastoma multiforme: an immuno-light and immuno-electron microscopic study. J. Neuropathol. Exp. Neurol. 54, 304–310
41. Morikawa, S., Baluk, P., Kaidoh, T., Haskell, A., Jain, R. K., and McDonald, D. M. (2002) Abnormalities in pericytes on blood vessels and endothelial sprouts in tumors. Am. J. Pathol. 160, 985–1000
42. Diaz-Flores, L., Gutierrez, R., Varela, H., Rancel, N., and Valladares, F. (1991) Microvascular pericytes: a review of their morphological and functional characteristics. Histol. Histopathol. 6, 269–286
43. Cunha, G. R. (1994) Role of mesenchymal-epithelial interactions in normal and abnormal development of the mammary gland and prostate. Cancer 74, 1030–1044
44. O'Reilly, M. A., Stripp, B. R., and Pryhuber, G. S. (1997) Epithelial-mesenchymal interactions in the alteration of gene expression and morphology following lung injury. Microsc. Res. Tech. 38, 473–479
Page 12 of 30(page number not for citation purposes)

45. Olson, E. N., Sternberg, E., Hu, J. S., Spizz, G., and Wilcox, C. (1986) Regulation of myogenic differentiation by type beta transforming growth factor. J. Cell Biol. 103, 1799–1805
46. Chao, M., Casaccia-Bonnefil, P., Carter, B., Chittka, A., Kong, H., and Yoon, S. O. (1998) Neurotrophin receptors: mediators of life and death. Brain Res. Rev. 26, 295–301
47. Yamagishi, H., Olson, E. N., and Srivastava, D. (2000) The basic helix-loop-helix transcription factor, dHAND, is required for vascular development. J. Clin. Invest. 105, 261–270
48. Hellstrom, M., Gerhardt, H., Kalen, M., Li, X., Eriksson, U., Wolburg, H., and Betsholtz, C. (2001) Lack of pericytes leads to endothelial hyperplasia and abnormal vascular morphogenesis. J. Cell Biol. 153, 543–553
49. Hellstrom, M., Kaln, M., Lindahl, P., Abramsson, A., and Betsholtz, C. (1999) Role of PDGF-B and PDGFR-beta in recruitment of vascular smooth muscle cells and pericytes during embryonic blood vessel formation in the mouse. Development 126, 3047–3055
50. Kuo, C. T., Veselits, M. L., and Leiden, J. M. (1997) LKLF: A transcriptional regulator of single-positive T cell quiescence and survival. Science 277, 1986–1990
51. Wani, M. A., Means, R. T., Jr., and Lingrel, J. B. (1998) Loss of LKLF function results in embryonic lethality in mice. Transgenic Res. 7, 229–238
52. Liu, Y., Wada, R., Yamashita, T., Mi, Y., Deng, C. X., Hobson, J. P., Rosenfeldt, H. M., Nava, V. E., Chae, S. S., Lee, M. J., et al. (2000) Edg-1, the G protein-coupled receptor for sphingosine-1-phosphate, is essential for vascular maturation. J. Clin. Invest. 106, 951–961
53. Gupta, S., Clarkson, M. R., Duggan, J., and Brady, H. R. (2000) Connective tissue growth factor: potential role in glomerulosclerosis and tubulointerstitial fibrosis. Kidney Int. 58, 1389–1399
54. Benckert, C., Jonas, S., Cramer, T., Von Marschall, Z., Schafer, G., Peters, M., Wagner, K., Radke, C., Wiedenmann, B., Neuhaus, P., et al. (2003) Transforming growth factor beta 1 stimulates vascular endothelial growth factor gene transcription in human cholangiocellular carcinoma cells. Cancer Res. 63, 1083–1092
55. Gohongi, T., Fukumura, D., Boucher, Y., Yun, C. O., Soff, G. A., Compton, C., Todoroki, T., and Jain, R. K. (1999) Tumor-host interactions in the gallbladder suppress distal angiogenesis and tumor growth: involvement of transforming growth factor beta1. Nat. Med. 5, 1203–1208
56. Darland, D. C., Massingham, L. J., Smith, S. R., Piek, E., Saint-Geniez, M., and D'Amore, P. A. (2003) Pericyte production of cell-associated VEGF is differentiation-dependent and is associated with endothelial survival. Dev. Biol. 264, 275–288
Page 13 of 30(page number not for citation purposes)

57. Nehls, V., Denzer, K., and Drenckhahn, D. (1992) Pericyte involvement in capillary sprouting during angiogenesis in situ. Cell Tissue Res. 270, 469–474
58. Tsuzuki, H., and Sasa, S. (1994) Ultrastructural observation of capillary sprouts in the dental organs of rat molars. Kaibogaku Zasshi 69, 684–696
59. Reynolds, L. P., Grazul-Bilska, A. T., and Redmer, D. A. (2000) Angiogenesis in the corpus luteum. Endocrine 12, 1–9
60. Nehls, V., and Drenckhahn, D. (1993) The versatility of microvascular pericytes: from mesenchyme to smooth muscle? Histochemistry 99, 1–12
61. Dome, B., Paku, S., Somlai, B., and Timar, J. (2002) Vascularization of cutaneous melanoma involves vessel co-option and has clinical significance. J. Pathol. 197, 355–362
62. Eberhard, A., Kahlert, S., Goede, V., Hemmerlein, B., Plate, K. H., and Augustin, H. G. (2000) Heterogeneity of angiogenesis and blood vessel maturation in human tumors: implications for antiangiogenic tumor therapies. Cancer Res. 60, 1388–1393
63. Bergers, G., Song, S., Myer-Morse, N., Bergsland, E., and Hanahan, D. (2003) Benefits of targeting both pericytes and endothelial cells in the tumor vasculature with kinase inhibitors. Journal of Clinical Investigation 111, 1287–1295
Received April 13, 2004; accepted October 13, 2004.
Page 14 of 30(page number not for citation purposes)
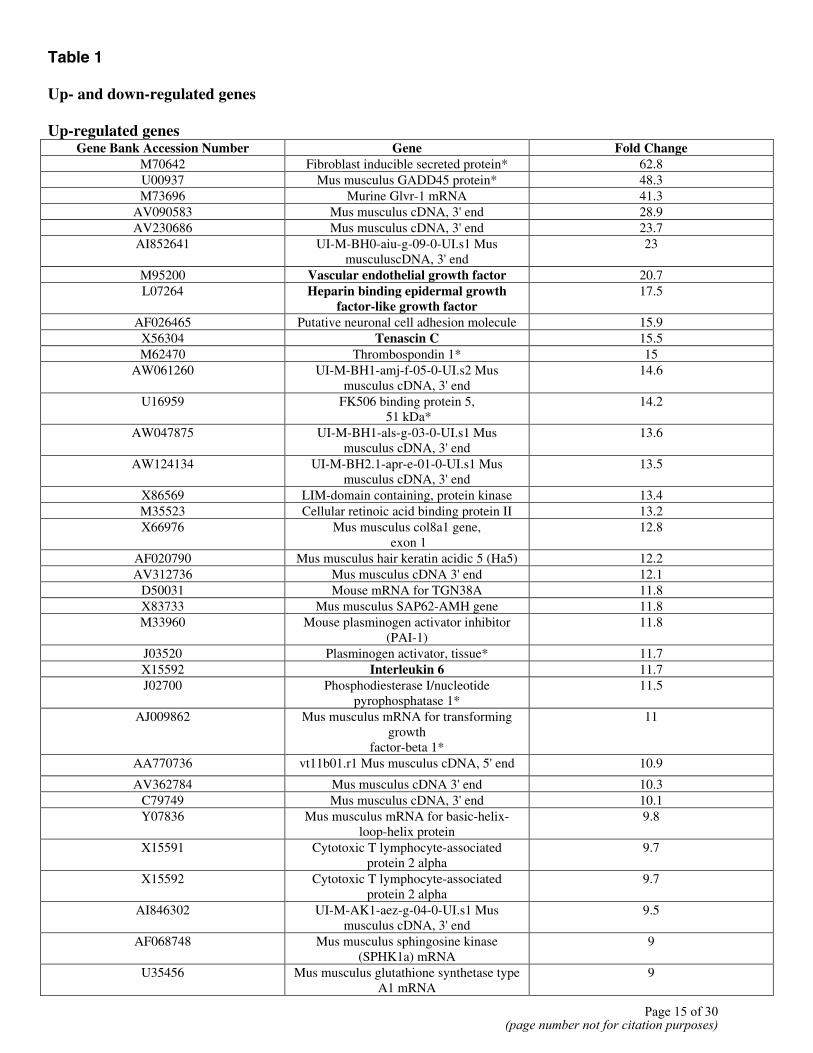
Table 1 Up- and down-regulated genes Up-regulated genes
Gene Bank Accession Number Gene Fold Change M70642 Fibroblast inducible secreted protein* 62.8 U00937 Mus musculus GADD45 protein* 48.3 M73696 Murine Glvr-1 mRNA 41.3
AV090583 Mus musculus cDNA, 3' end 28.9 AV230686 Mus musculus cDNA, 3' end 23.7 AI852641 UI-M-BH0-aiu-g-09-0-UI.s1 Mus
musculuscDNA, 3' end 23
M95200 Vascular endothelial growth factor 20.7 L07264 Heparin binding epidermal growth
factor-like growth factor 17.5
AF026465 Putative neuronal cell adhesion molecule 15.9 X56304 Tenascin C 15.5 M62470 Thrombospondin 1* 15
AW061260 UI-M-BH1-amj-f-05-0-UI.s2 Mus musculus cDNA, 3' end
14.6
U16959 FK506 binding protein 5, 51 kDa*
14.2
AW047875 UI-M-BH1-als-g-03-0-UI.s1 Mus musculus cDNA, 3' end
13.6
AW124134 UI-M-BH2.1-apr-e-01-0-UI.s1 Mus musculus cDNA, 3' end
13.5
X86569 LIM-domain containing, protein kinase 13.4 M35523 Cellular retinoic acid binding protein II 13.2 X66976 Mus musculus col8a1 gene,
exon 1 12.8
AF020790 Mus musculus hair keratin acidic 5 (Ha5) 12.2 AV312736 Mus musculus cDNA 3' end 12.1
D50031 Mouse mRNA for TGN38A 11.8 X83733 Mus musculus SAP62-AMH gene 11.8 M33960 Mouse plasminogen activator inhibitor
(PAI-1) 11.8
J03520 Plasminogen activator, tissue* 11.7 X15592 Interleukin 6 11.7 J02700 Phosphodiesterase I/nucleotide
pyrophosphatase 1* 11.5
AJ009862 Mus musculus mRNA for transforming growth
factor-beta 1*
11
AA770736 vt11b01.r1 Mus musculus cDNA, 5' end 10.9
AV362784 Mus musculus cDNA 3' end 10.3 C79749 Mus musculus cDNA, 3' end 10.1 Y07836 Mus musculus mRNA for basic-helix-
loop-helix protein 9.8
X15591 Cytotoxic T lymphocyte-associated protein 2 alpha
9.7
X15592 Cytotoxic T lymphocyte-associated protein 2 alpha
9.7
AI846302 UI-M-AK1-aez-g-04-0-UI.s1 Mus musculus cDNA, 3' end
9.5
AF068748 Mus musculus sphingosine kinase (SPHK1a) mRNA
9
U35456 Mus musculus glutathione synthetase type A1 mRNA
9
Page 15 of 30(page number not for citation purposes)
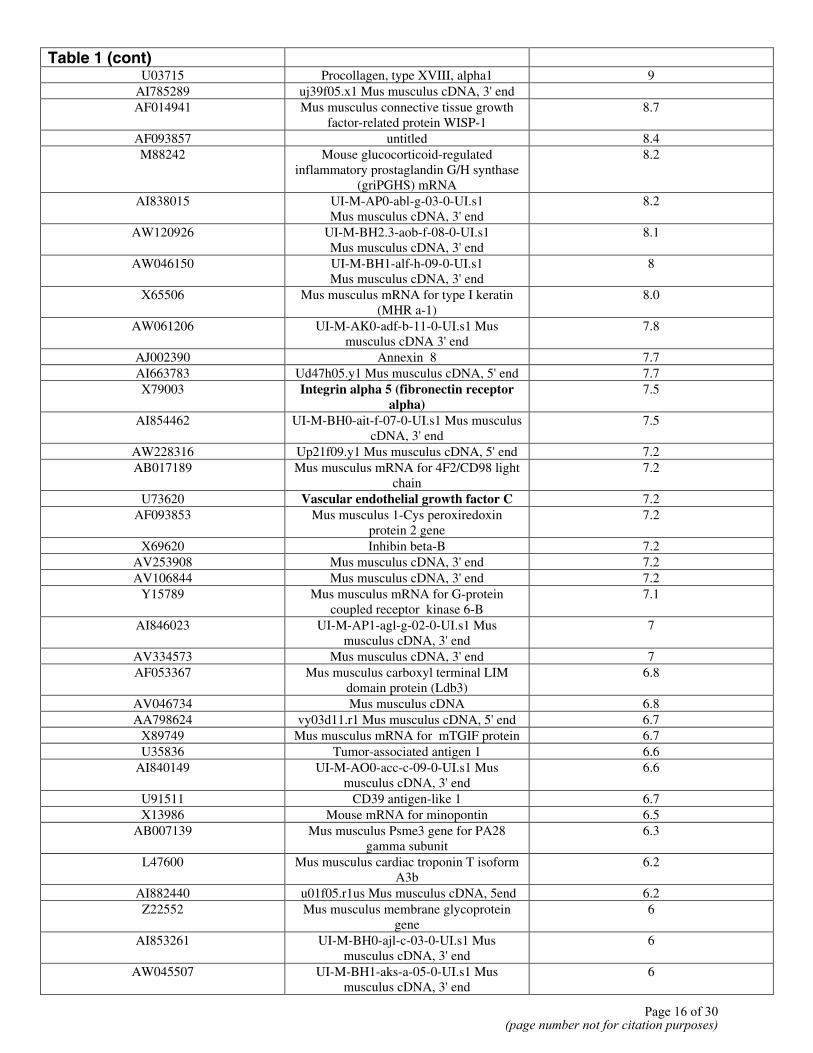
Table 1 (cont)
U03715 Procollagen, type XVIII, alpha1 9 AI785289 uj39f05.x1 Mus musculus cDNA, 3' end AF014941 Mus musculus connective tissue growth
factor-related protein WISP-1 8.7
AF093857 untitled 8.4 M88242 Mouse glucocorticoid-regulated
inflammatory prostaglandin G/H synthase (griPGHS) mRNA
8.2
AI838015 UI-M-AP0-abl-g-03-0-UI.s1 Mus musculus cDNA, 3' end
8.2
AW120926 UI-M-BH2.3-aob-f-08-0-UI.s1 Mus musculus cDNA, 3' end
8.1
AW046150 UI-M-BH1-alf-h-09-0-UI.s1 Mus musculus cDNA, 3' end
8
X65506 Mus musculus mRNA for type I keratin (MHR a-1)
8.0
AW061206 UI-M-AK0-adf-b-11-0-UI.s1 Mus musculus cDNA 3' end
7.8
AJ002390 Annexin 8 7.7 AI663783 Ud47h05.y1 Mus musculus cDNA, 5' end 7.7 X79003 Integrin alpha 5 (fibronectin receptor
alpha) 7.5
AI854462 UI-M-BH0-ait-f-07-0-UI.s1 Mus musculus cDNA, 3' end
7.5
AW228316 Up21f09.y1 Mus musculus cDNA, 5' end 7.2 AB017189 Mus musculus mRNA for 4F2/CD98 light
chain 7.2
U73620 Vascular endothelial growth factor C 7.2 AF093853 Mus musculus 1-Cys peroxiredoxin
protein 2 gene 7.2
X69620 Inhibin beta-B 7.2 AV253908 Mus musculus cDNA, 3' end 7.2 AV106844 Mus musculus cDNA, 3' end 7.2
Y15789 Mus musculus mRNA for G-protein coupled receptor kinase 6-B
7.1
AI846023 UI-M-AP1-agl-g-02-0-UI.s1 Mus musculus cDNA, 3' end
7
AV334573 Mus musculus cDNA, 3' end 7 AF053367 Mus musculus carboxyl terminal LIM
domain protein (Ldb3) 6.8
AV046734 Mus musculus cDNA 6.8 AA798624 vy03d11.r1 Mus musculus cDNA, 5' end 6.7
X89749 Mus musculus mRNA for mTGIF protein 6.7 U35836 Tumor-associated antigen 1 6.6
AI840149 UI-M-AO0-acc-c-09-0-UI.s1 Mus musculus cDNA, 3' end
6.6
U91511 CD39 antigen-like 1 6.7 X13986 Mouse mRNA for minopontin 6.5
AB007139 Mus musculus Psme3 gene for PA28 gamma subunit
6.3
L47600 Mus musculus cardiac troponin T isoform A3b
6.2
AI882440 u01f05.r1us Mus musculus cDNA, 5end 6.2 Z22552 Mus musculus membrane glycoprotein
gene 6
AI853261 UI-M-BH0-ajl-c-03-0-UI.s1 Mus musculus cDNA, 3' end
6
AW045507 UI-M-BH1-aks-a-05-0-UI.s1 Mus musculus cDNA, 3' end
6
Page 16 of 30(page number not for citation purposes)
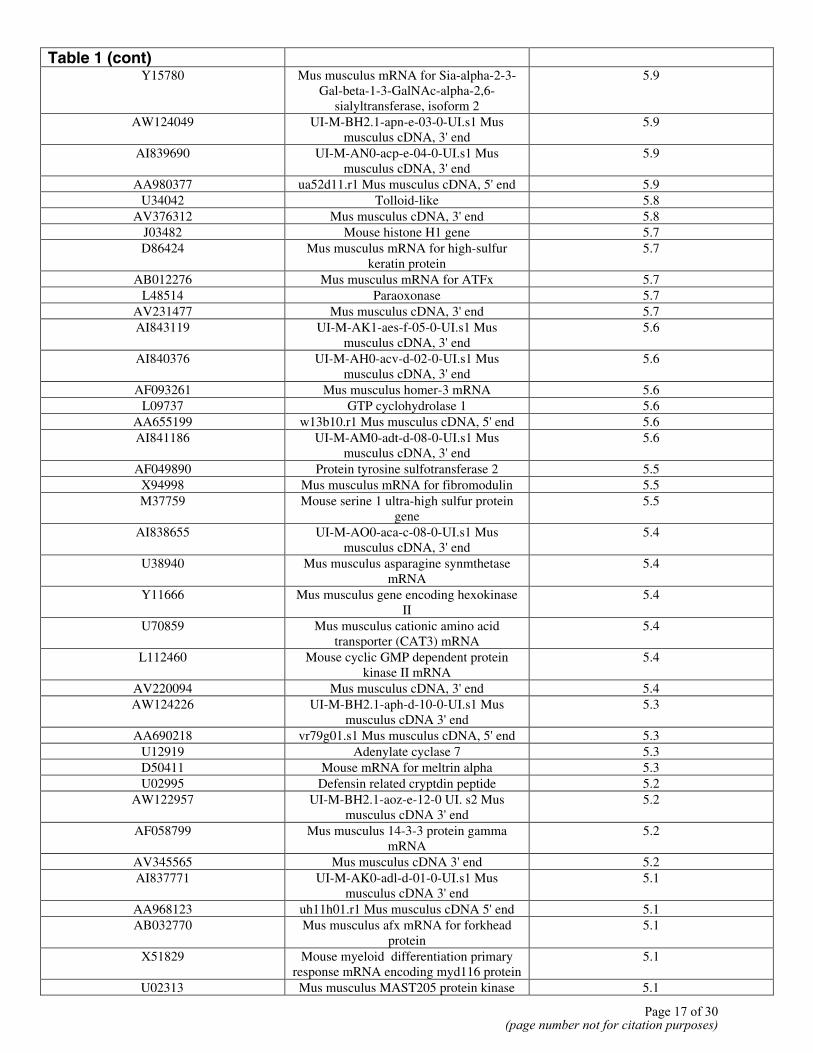
Table 1 (cont)
Y15780 Mus musculus mRNA for Sia-alpha-2-3-Gal-beta-1-3-GalNAc-alpha-2,6-
sialyltransferase, isoform 2
5.9
AW124049 UI-M-BH2.1-apn-e-03-0-UI.s1 Mus musculus cDNA, 3' end
5.9
AI839690 UI-M-AN0-acp-e-04-0-UI.s1 Mus musculus cDNA, 3' end
5.9
AA980377 ua52d11.r1 Mus musculus cDNA, 5' end 5.9 U34042 Tolloid-like 5.8
AV376312 Mus musculus cDNA, 3' end 5.8 J03482 Mouse histone H1 gene 5.7 D86424 Mus musculus mRNA for high-sulfur
keratin protein 5.7
AB012276 Mus musculus mRNA for ATFx 5.7 L48514 Paraoxonase 5.7
AV231477 Mus musculus cDNA, 3' end 5.7 AI843119 UI-M-AK1-aes-f-05-0-UI.s1 Mus
musculus cDNA, 3' end 5.6
AI840376 UI-M-AH0-acv-d-02-0-UI.s1 Mus musculus cDNA, 3' end
5.6
AF093261 Mus musculus homer-3 mRNA 5.6 L09737 GTP cyclohydrolase 1 5.6
AA655199 w13b10.r1 Mus musculus cDNA, 5' end 5.6 AI841186 UI-M-AM0-adt-d-08-0-UI.s1 Mus
musculus cDNA, 3' end 5.6
AF049890 Protein tyrosine sulfotransferase 2 5.5 X94998 Mus musculus mRNA for fibromodulin 5.5 M37759 Mouse serine 1 ultra-high sulfur protein
gene 5.5
AI838655 UI-M-AO0-aca-c-08-0-UI.s1 Mus musculus cDNA, 3' end
5.4
U38940 Mus musculus asparagine synmthetase mRNA
5.4
Y11666 Mus musculus gene encoding hexokinase II
5.4
U70859 Mus musculus cationic amino acid transporter (CAT3) mRNA
5.4
L112460 Mouse cyclic GMP dependent protein kinase II mRNA
5.4
AV220094 Mus musculus cDNA, 3' end 5.4 AW124226 UI-M-BH2.1-aph-d-10-0-UI.s1 Mus
musculus cDNA 3' end 5.3
AA690218 vr79g01.s1 Mus musculus cDNA, 5' end 5.3 U12919 Adenylate cyclase 7 5.3 D50411 Mouse mRNA for meltrin alpha 5.3 U02995 Defensin related cryptdin peptide 5.2
AW122957 UI-M-BH2.1-aoz-e-12-0 UI. s2 Mus musculus cDNA 3' end
5.2
AF058799 Mus musculus 14-3-3 protein gamma mRNA
5.2
AV345565 Mus musculus cDNA 3' end 5.2 AI837771 UI-M-AK0-adl-d-01-0-UI.s1 Mus
musculus cDNA 3' end 5.1
AA968123 uh11h01.r1 Mus musculus cDNA 5' end 5.1 AB032770 Mus musculus afx mRNA for forkhead
protein 5.1
X51829 Mouse myeloid differentiation primary response mRNA encoding myd116 protein
5.1
U02313 Mus musculus MAST205 protein kinase 5.1
Page 17 of 30(page number not for citation purposes)
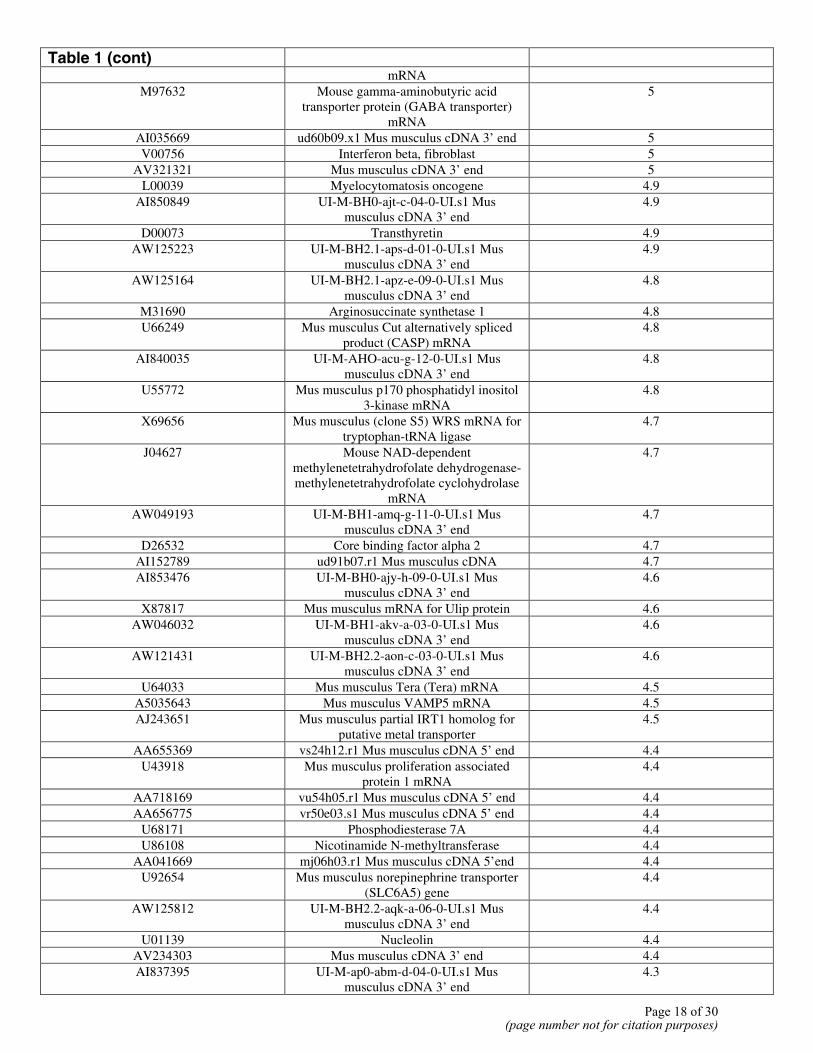
Table 1 (cont)
mRNA M97632 Mouse gamma-aminobutyric acid
transporter protein (GABA transporter) mRNA
5
AI035669 ud60b09.x1 Mus musculus cDNA 3’ end 5 V00756 Interferon beta, fibroblast 5
AV321321 Mus musculus cDNA 3’ end 5 L00039 Myelocytomatosis oncogene 4.9
AI850849 UI-M-BH0-ajt-c-04-0-UI.s1 Mus musculus cDNA 3’ end
4.9
D00073 Transthyretin 4.9 AW125223 UI-M-BH2.1-aps-d-01-0-UI.s1 Mus
musculus cDNA 3’ end 4.9
AW125164 UI-M-BH2.1-apz-e-09-0-UI.s1 Mus musculus cDNA 3’ end
4.8
M31690 Arginosuccinate synthetase 1 4.8 U66249 Mus musculus Cut alternatively spliced
product (CASP) mRNA 4.8
AI840035 UI-M-AHO-acu-g-12-0-UI.s1 Mus musculus cDNA 3’ end
4.8
U55772 Mus musculus p170 phosphatidyl inositol 3-kinase mRNA
4.8
X69656 Mus musculus (clone S5) WRS mRNA for tryptophan-tRNA ligase
4.7
J04627 Mouse NAD-dependent methylenetetrahydrofolate dehydrogenase- methylenetetrahydrofolate cyclohydrolase
mRNA
4.7
AW049193 UI-M-BH1-amq-g-11-0-UI.s1 Mus musculus cDNA 3’ end
4.7
D26532 Core binding factor alpha 2 4.7 AI152789 ud91b07.r1 Mus musculus cDNA 4.7 AI853476 UI-M-BH0-ajy-h-09-0-UI.s1 Mus
musculus cDNA 3’ end 4.6
X87817 Mus musculus mRNA for Ulip protein 4.6 AW046032 UI-M-BH1-akv-a-03-0-UI.s1 Mus
musculus cDNA 3’ end 4.6
AW121431 UI-M-BH2.2-aon-c-03-0-UI.s1 Mus musculus cDNA 3’ end
4.6
U64033 Mus musculus Tera (Tera) mRNA 4.5 A5035643 Mus musculus VAMP5 mRNA 4.5 AJ243651 Mus musculus partial IRT1 homolog for
putative metal transporter 4.5
AA655369 vs24h12.r1 Mus musculus cDNA 5’ end 4.4 U43918 Mus musculus proliferation associated
protein 1 mRNA 4.4
AA718169 vu54h05.r1 Mus musculus cDNA 5’ end 4.4 AA656775 vr50e03.s1 Mus musculus cDNA 5’ end 4.4
U68171 Phosphodiesterase 7A 4.4 U86108 Nicotinamide N-methyltransferase 4.4
AA041669 mj06h03.r1 Mus musculus cDNA 5’end 4.4 U92654 Mus musculus norepinephrine transporter
(SLC6A5) gene 4.4
AW125812 UI-M-BH2.2-aqk-a-06-0-UI.s1 Mus musculus cDNA 3’ end
4.4
U01139 Nucleolin 4.4 AV234303 Mus musculus cDNA 3’ end 4.4 AI837395 UI-M-ap0-abm-d-04-0-UI.s1 Mus
musculus cDNA 3’ end 4.3
Page 18 of 30(page number not for citation purposes)
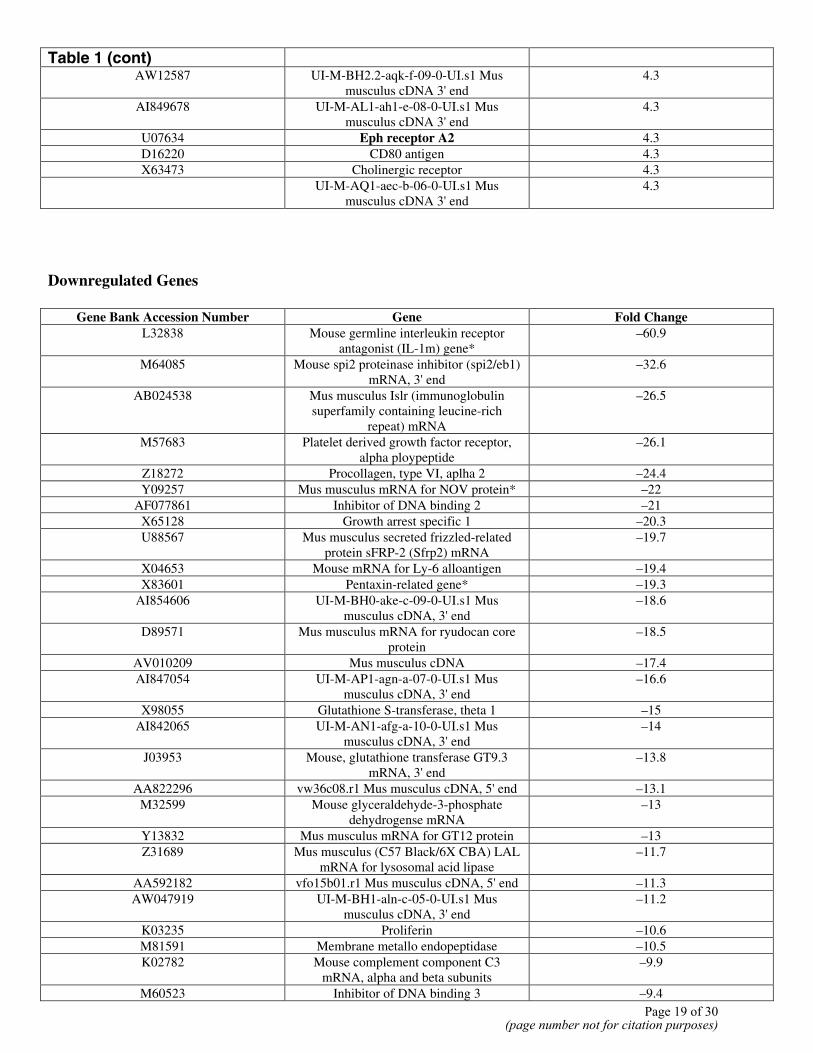
Table 1 (cont)
AW12587 UI-M-BH2.2-aqk-f-09-0-UI.s1 Mus musculus cDNA 3' end
4.3
AI849678 UI-M-AL1-ah1-e-08-0-UI.s1 Mus musculus cDNA 3' end
4.3
U07634 Eph receptor A2 4.3 D16220 CD80 antigen 4.3 X63473 Cholinergic receptor 4.3
UI-M-AQ1-aec-b-06-0-UI.s1 Mus musculus cDNA 3' end
4.3
Downregulated Genes
Gene Bank Accession Number Gene Fold Change L32838 Mouse germline interleukin receptor
antagonist (IL-1m) gene* –60.9
M64085 Mouse spi2 proteinase inhibitor (spi2/eb1) mRNA, 3' end
–32.6
AB024538 Mus musculus Islr (immunoglobulin superfamily containing leucine-rich
repeat) mRNA
–26.5
M57683 Platelet derived growth factor receptor, alpha ploypeptide
–26.1
Z18272 Procollagen, type VI, aplha 2 –24.4 Y09257 Mus musculus mRNA for NOV protein* –22
AF077861 Inhibitor of DNA binding 2 –21 X65128 Growth arrest specific 1 –20.3 U88567 Mus musculus secreted frizzled-related
protein sFRP-2 (Sfrp2) mRNA –19.7
X04653 Mouse mRNA for Ly-6 alloantigen –19.4 X83601 Pentaxin-related gene* –19.3
AI854606 UI-M-BH0-ake-c-09-0-UI.s1 Mus musculus cDNA, 3' end
–18.6
D89571 Mus musculus mRNA for ryudocan core protein
–18.5
AV010209 Mus musculus cDNA –17.4 AI847054 UI-M-AP1-agn-a-07-0-UI.s1 Mus
musculus cDNA, 3' end –16.6
X98055 Glutathione S-transferase, theta 1 –15 AI842065 UI-M-AN1-afg-a-10-0-UI.s1 Mus
musculus cDNA, 3' end –14
J03953 Mouse, glutathione transferase GT9.3 mRNA, 3' end
–13.8
AA822296 vw36c08.r1 Mus musculus cDNA, 5' end –13.1 M32599 Mouse glyceraldehyde-3-phosphate
dehydrogense mRNA –13
Y13832 Mus musculus mRNA for GT12 protein –13 Z31689 Mus musculus (C57 Black/6X CBA) LAL
mRNA for lysosomal acid lipase –11.7
AA592182 vfo15b01.r1 Mus musculus cDNA, 5' end –11.3 AW047919 UI-M-BH1-aln-c-05-0-UI.s1 Mus
musculus cDNA, 3' end –11.2
K03235 Proliferin –10.6 M81591 Membrane metallo endopeptidase –10.5 K02782 Mouse complement component C3
mRNA, alpha and beta subunits –9.9
M60523 Inhibitor of DNA binding 3 –9.4 Page 19 of 30
(page number not for citation purposes)
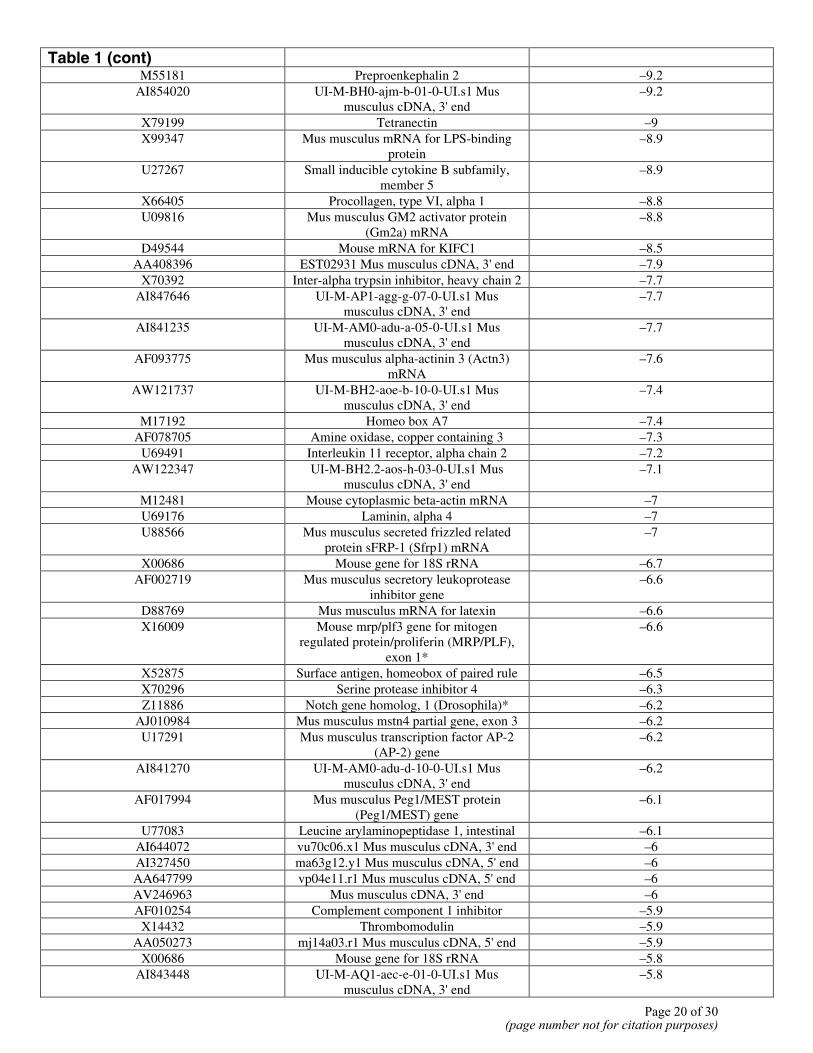
Table 1 (cont)
M55181 Preproenkephalin 2 –9.2 AI854020 UI-M-BH0-ajm-b-01-0-UI.s1 Mus
musculus cDNA, 3' end –9.2
X79199 Tetranectin –9 X99347 Mus musculus mRNA for LPS-binding
protein –8.9
U27267 Small inducible cytokine B subfamily, member 5
–8.9
X66405 Procollagen, type VI, alpha 1 –8.8 U09816 Mus musculus GM2 activator protein
(Gm2a) mRNA –8.8
D49544 Mouse mRNA for KIFC1 –8.5 AA408396 EST02931 Mus musculus cDNA, 3' end –7.9
X70392 Inter-alpha trypsin inhibitor, heavy chain 2 –7.7 AI847646 UI-M-AP1-agg-g-07-0-UI.s1 Mus
musculus cDNA, 3' end –7.7
AI841235 UI-M-AM0-adu-a-05-0-UI.s1 Mus musculus cDNA, 3' end
–7.7
AF093775 Mus musculus alpha-actinin 3 (Actn3) mRNA
–7.6
AW121737 UI-M-BH2-aoe-b-10-0-UI.s1 Mus musculus cDNA, 3' end
–7.4
M17192 Homeo box A7 –7.4 AF078705 Amine oxidase, copper containing 3 –7.3
U69491 Interleukin 11 receptor, alpha chain 2 –7.2 AW122347 UI-M-BH2.2-aos-h-03-0-UI.s1 Mus
musculus cDNA, 3' end –7.1
M12481 Mouse cytoplasmic beta-actin mRNA –7 U69176 Laminin, alpha 4 –7 U88566 Mus musculus secreted frizzled related
protein sFRP-1 (Sfrp1) mRNA –7
X00686 Mouse gene for 18S rRNA –6.7 AF002719 Mus musculus secretory leukoprotease
inhibitor gene –6.6
D88769 Mus musculus mRNA for latexin –6.6 X16009 Mouse mrp/plf3 gene for mitogen
regulated protein/proliferin (MRP/PLF), exon 1*
–6.6
X52875 Surface antigen, homeobox of paired rule –6.5 X70296 Serine protease inhibitor 4 –6.3 Z11886 Notch gene homolog, 1 (Drosophila)* –6.2
AJ010984 Mus musculus mstn4 partial gene, exon 3 –6.2 U17291 Mus musculus transcription factor AP-2
(AP-2) gene –6.2
AI841270 UI-M-AM0-adu-d-10-0-UI.s1 Mus musculus cDNA, 3' end
–6.2
AF017994 Mus musculus Peg1/MEST protein (Peg1/MEST) gene
–6.1
U77083 Leucine arylaminopeptidase 1, intestinal –6.1 AI644072 vu70c06.x1 Mus musculus cDNA, 3' end –6 AI327450 ma63g12.y1 Mus musculus cDNA, 5' end –6 AA647799 vp04e11.r1 Mus musculus cDNA, 5' end –6 AV246963 Mus musculus cDNA, 3' end –6 AF010254 Complement component 1 inhibitor –5.9
X14432 Thrombomodulin –5.9 AA050273 mj14a03.r1 Mus musculus cDNA, 5' end –5.9
X00686 Mouse gene for 18S rRNA –5.8 AI843448 UI-M-AQ1-aec-e-01-0-UI.s1 Mus
musculus cDNA, 3' end –5.8
Page 20 of 30(page number not for citation purposes)
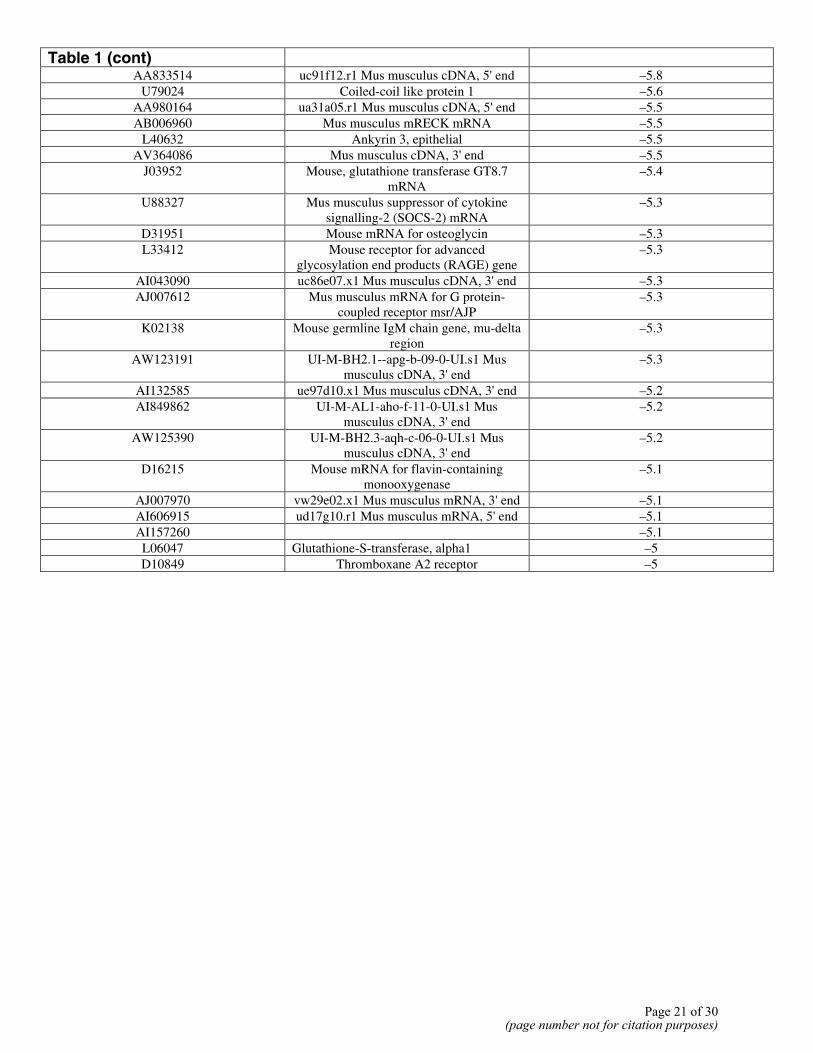
Table 1 (cont)
AA833514 uc91f12.r1 Mus musculus cDNA, 5' end –5.8 U79024 Coiled-coil like protein 1 –5.6
AA980164 ua31a05.r1 Mus musculus cDNA, 5' end –5.5 AB006960 Mus musculus mRECK mRNA –5.5
L40632 Ankyrin 3, epithelial –5.5 AV364086 Mus musculus cDNA, 3' end –5.5
J03952 Mouse, glutathione transferase GT8.7 mRNA
–5.4
U88327 Mus musculus suppressor of cytokine signalling-2 (SOCS-2) mRNA
–5.3
D31951 Mouse mRNA for osteoglycin –5.3 L33412 Mouse receptor for advanced
glycosylation end products (RAGE) gene –5.3
AI043090 uc86e07.x1 Mus musculus cDNA, 3' end –5.3 AJ007612 Mus musculus mRNA for G protein-
coupled receptor msr/AJP –5.3
K02138 Mouse germline IgM chain gene, mu-delta region
–5.3
AW123191 UI-M-BH2.1--apg-b-09-0-UI.s1 Mus musculus cDNA, 3' end
–5.3
AI132585 ue97d10.x1 Mus musculus cDNA, 3' end –5.2 AI849862 UI-M-AL1-aho-f-11-0-UI.s1 Mus
musculus cDNA, 3' end –5.2
AW125390 UI-M-BH2.3-aqh-c-06-0-UI.s1 Mus musculus cDNA, 3' end
–5.2
D16215 Mouse mRNA for flavin-containing monooxygenase
–5.1
AJ007970 vw29e02.x1 Mus musculus mRNA, 3' end –5.1 AI606915 ud17g10.r1 Mus musculus mRNA, 5' end –5.1 AI157260 –5.1 L06047 Glutathione-S-transferase, alpha1 –5 D10849 Thromboxane A2 receptor –5
Page 21 of 30(page number not for citation purposes)
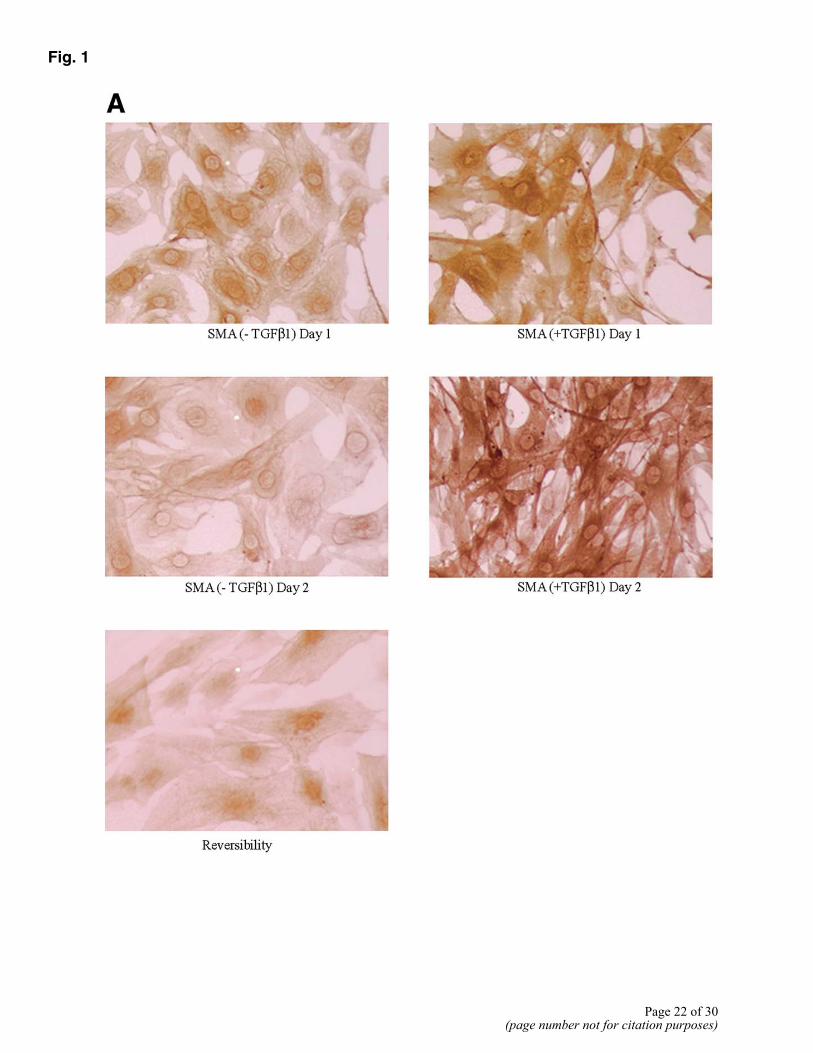
Fig. 1
Page 22 of 30(page number not for citation purposes)
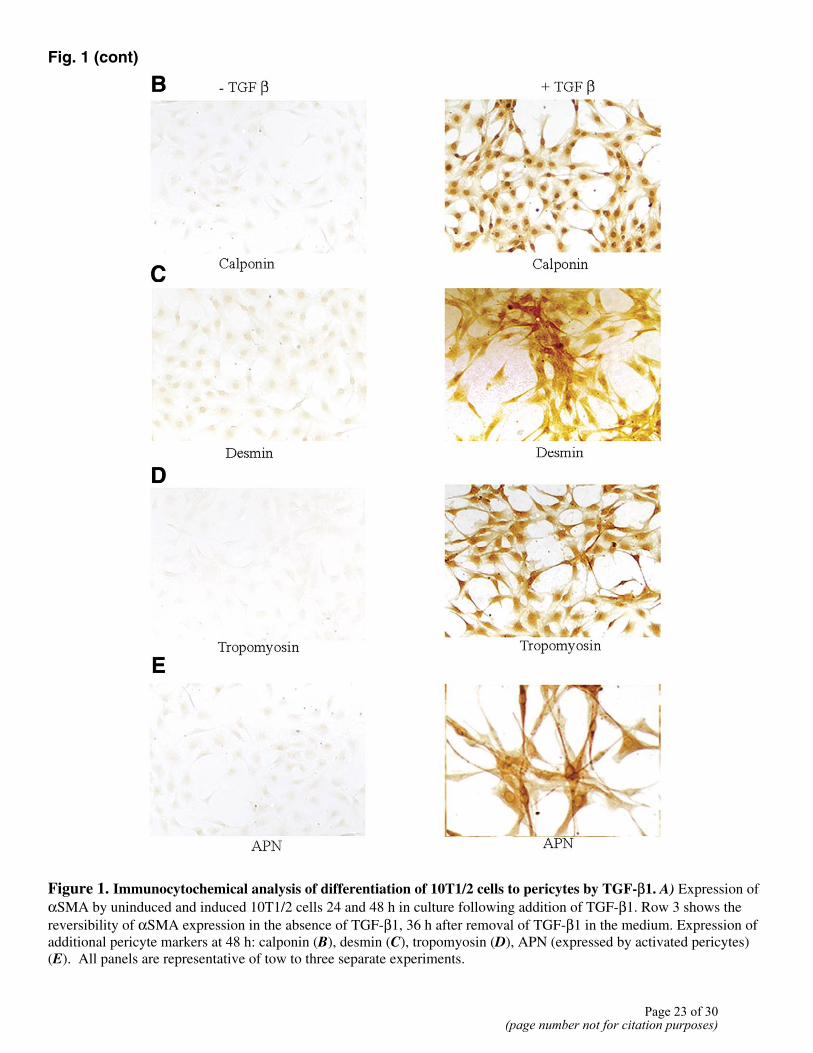
Fig. 1 (cont)
Figure 1. Immunocytochemical analysis of differentiation of 10T1/2 cells to pericytes by TGF-β1. A) Expression of αSMA by uninduced and induced 10T1/2 cells 24 and 48 h in culture following addition of TGF-β1. Row 3 shows the reversibility of αSMA expression in the absence of TGF-β1, 36 h after removal of TGF-β1 in the medium. Expression of additional pericyte markers at 48 h: calponin (B), desmin (C), tropomyosin (D), APN (expressed by activated pericytes) (E). All panels are representative of tow to three separate experiments.
Page 23 of 30(page number not for citation purposes)
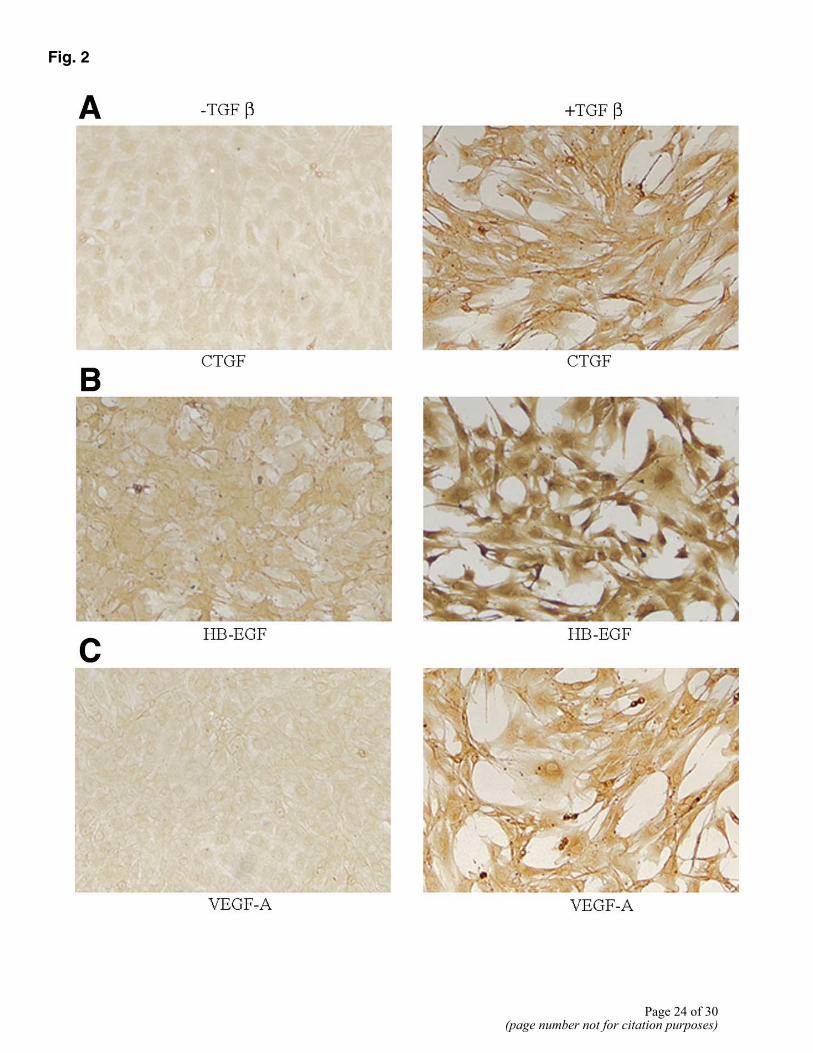
Fig. 2
Page 24 of 30(page number not for citation purposes)
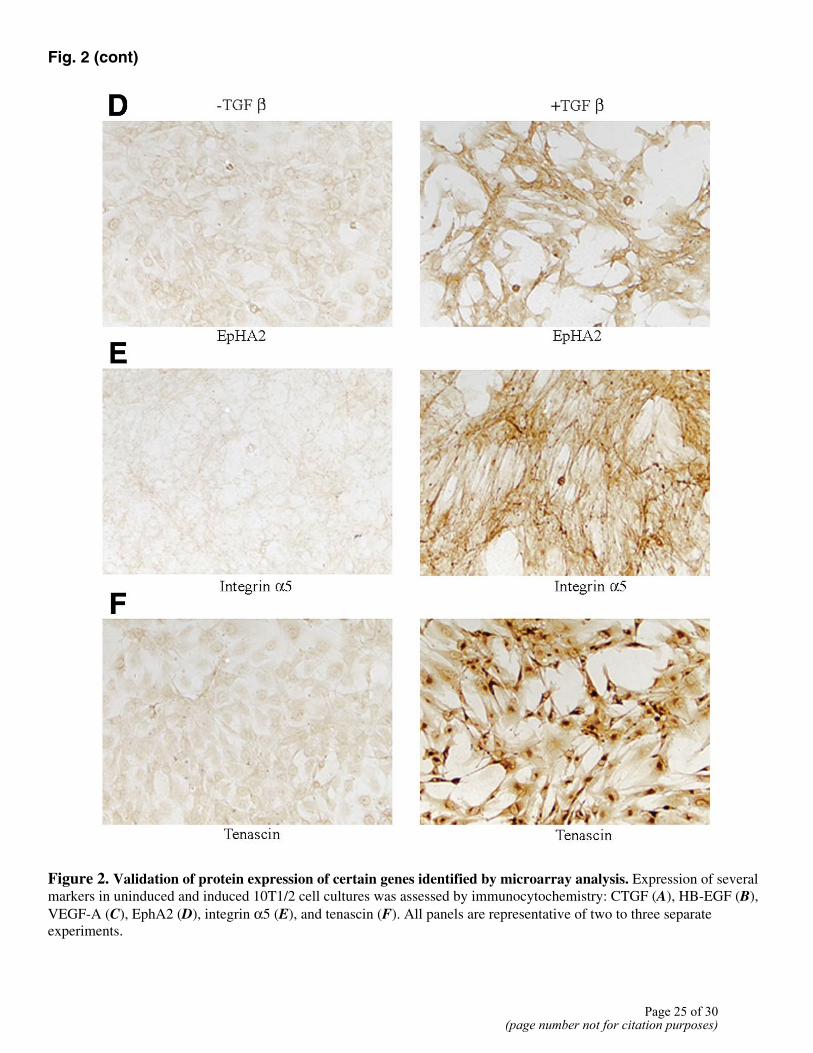
Fig. 2 (cont)
Figure 2. Validation of protein expression of certain genes identified by microarray analysis. Expression of several markers in uninduced and induced 10T1/2 cell cultures was assessed by immunocytochemistry: CTGF (A), HB-EGF (B), VEGF-A (C), EphA2 (D), integrin α5 (E), and tenascin (F). All panels are representative of two to three separate experiments.
Page 25 of 30(page number not for citation purposes)
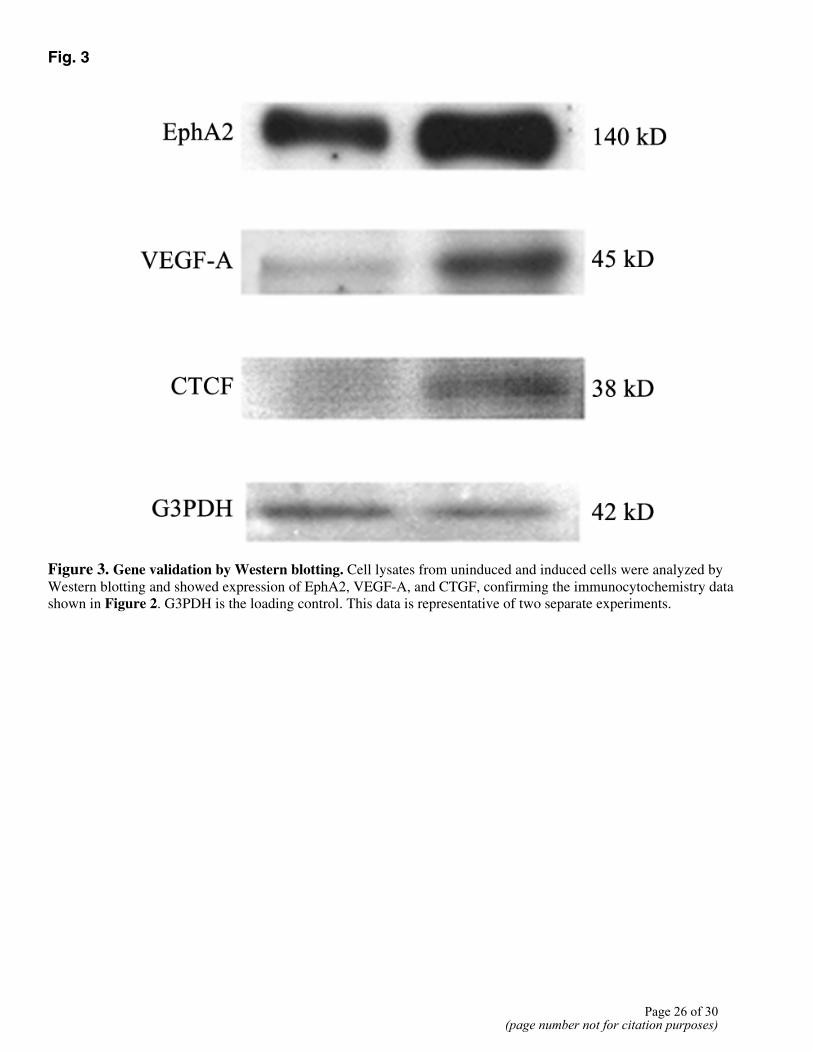
Fig. 3
Figure 3. Gene validation by Western blotting. Cell lysates from uninduced and induced cells were analyzed by Western blotting and showed expression of EphA2, VEGF-A, and CTGF, confirming the immunocytochemistry data shown in Figure 2. G3PDH is the loading control. This data is representative of two separate experiments.
Page 26 of 30(page number not for citation purposes)

Fig. 4
Figure 4. Role of various factors in the differentiation of 10T1/2 cells. Analysis of αSMA expression in induced 10T1/2 cells by Western analysis. Cells were cultured without TGF-β1 (lane 1) and with TGF-β1 alone (lane 2) or along with neutralizing antibodies to VEGF-A (lane 3), IL-6 (lane 4), integrin α5 (lane 6), and the corresponding isotype control antibodies (lanes 5 and 7). This data is representative of two separate experiments.
Page 27 of 30(page number not for citation purposes)
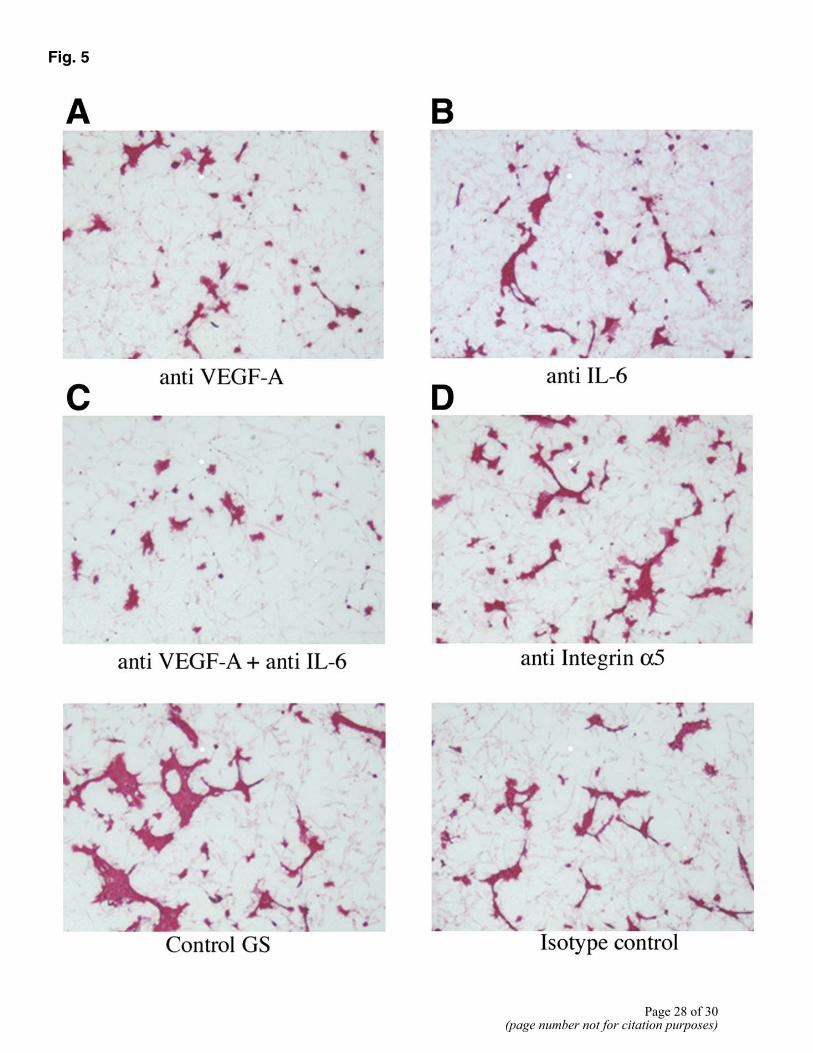
Fig. 5
Page 28 of 30(page number not for citation purposes)
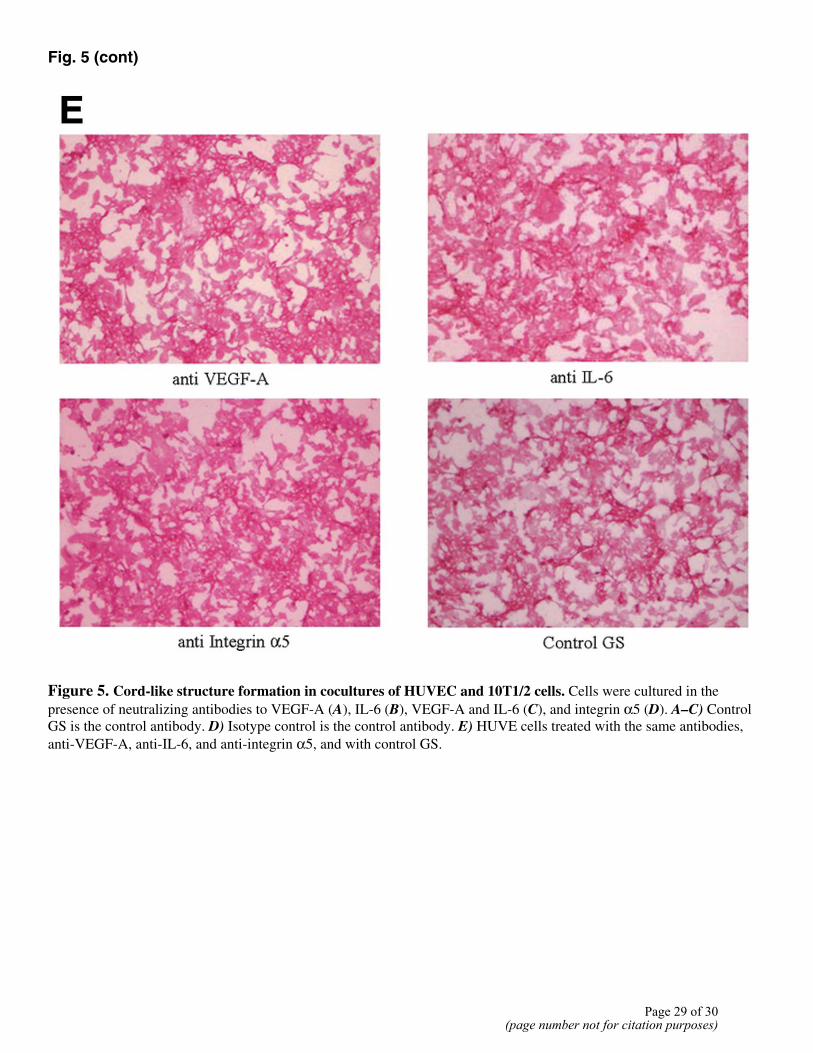
Fig. 5 (cont)
Figure 5. Cord-like structure formation in cocultures of HUVEC and 10T1/2 cells. Cells were cultured in the presence of neutralizing antibodies to VEGF-A (A), IL-6 (B), VEGF-A and IL-6 (C), and integrin α5 (D). A–C) Control GS is the control antibody. D) Isotype control is the control antibody. E) HUVE cells treated with the same antibodies, anti-VEGF-A, anti-IL-6, and anti-integrin α5, and with control GS.
Page 29 of 30(page number not for citation purposes)
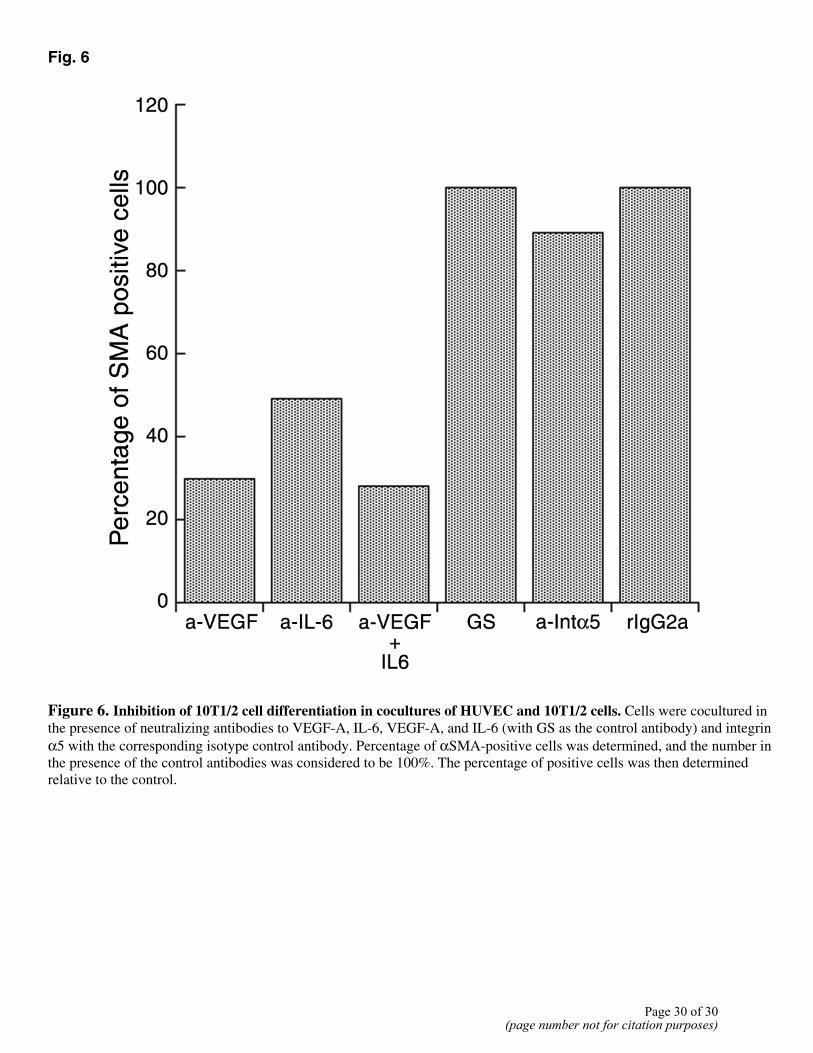
Fig. 6
Figure 6. Inhibition of 10T1/2 cell differentiation in cocultures of HUVEC and 10T1/2 cells. Cells were cocultured in the presence of neutralizing antibodies to VEGF-A, IL-6, VEGF-A, and IL-6 (with GS as the control antibody) and integrin α5 with the corresponding isotype control antibody. Percentage of αSMA-positive cells was determined, and the number in the presence of the control antibodies was considered to be 100%. The percentage of positive cells was then determined relative to the control.
Page 30 of 30(page number not for citation purposes)