pitx3 microarray pub
TRANSCRIPT

a PATTERNS & PHENOTYPES
Microarray-Based Identification of Pitx3Targets During Xenopus EmbryogenesisLara Hooker, Cristine Smoczer, Farhad KhosrowShahian, Marian Wolanski,and Michael J. Crawford*
Background: Unexpected phenotypes resulting from morpholino-mediated translational knockdown ofPitx3 in Xenopus laevis required further investigation regarding the genetic networks in which the genemight play a role. Microarray analysis was, therefore, used to assess global transcriptional changes down-stream of Pitx3. Results: From the large data set generated, selected candidate genes were confirmed byreverse transcriptase-polymerase chain reaction (RT-PCR) and in situ hybridization. Conclusions: Wehave identified four genes as likely direct targets of Pitx3 action: Pax6, b Crystallin-b1 (Crybb1), Hes7.1,and Hes4. Four others show equivocal promise worthy of consideration: Vent2, and Ripply2 (aka Ledger-line or Stripy), eFGF and RXRa. We also describe the expression pattern of additional and novel genesthat are Pitx3-sensitive but that are unlikely to be direct targets. Developmental Dynamics 241:1487–1505,2012. VC 2012 Wiley Periodicals, Inc.
Key words: microarray; Pitx3; morpholino; Xenopus laevis; eye; lens; somite; Pax6; segmentation clock; retinoid;mutant
Key findings:� Amicroarray assay and secondary confirmation of Pitx3 morphants indicates that Pax6, b Crystallin-b1 (Crybb1),Hes7.1, and Hes4 are good candidates for direct targets.
� Four other genes show equivocal promise worthy of consideration: Vent2, and Ripply2 (aka Ledgerline or Stripy),eFGF and RXRa.
� Novel genes described (but that are likely indirectly affected) are described: Rbp4l, Galectin IX, Baz2b, andRdh16.
� In addition to previously described interactions in lens and brain, Pitx3 also intersects the segmentation pathwayand retinoid regulation.
Accepted 9 July 2012
INTRODUCTION
Pitx3 encodes a bicoid-like transcrip-tion factor that is characterized by alysine residue at position 50 of thehomeodomain. The aphakia (ak)mouse represents a natural Pitx3 mu-tant model that is the result of twodeletions in its regulatory region thatabolish eye and brain expression,
but leave muscle expression intact(Semina et al., 2000; Rieger et al.,2001; Coulon et al., 2007). This geno-type displays microphthalmic eyesthat lack developed lenses. They alsodisplay impaired differentiation of do-paminergic neurons in the substantianigra: mutants mimic the symptomsof Parkinson’s disease (PD; Varnumand Stevens, 1968; van den Munckhof
et al., 2003). In humans, PITX3 dis-ruption can lead to congenital cata-racts, anterior segment mesenchymaldysgenesis (ASMD), Peter’s anomaly,and/ or microphthalmia (Sakazumeet al., 2007). This implicates PITX3 asa major player in the control of genetranscription in lens fibers. In theventral tegmental area (VTA) andsubstantia nigra compacta (SNc)
Dev
elop
men
tal D
ynam
ics
Department of Biological Sciences, University of Windsor, Windsor, Ontario, CanadaGrant sponsor: Natural Science and Engineering Research Council of Canada; Grant number: 203459.*Correspondence to: Michael J. Crawford, Department of Biological Sciences, University of Windsor, 401 Sunset Avenue,Windsor, Ontario, N9B 3P4, Canada. E-mail: [email protected]
DOI 10.1002/dvdy.23836Published online 10 August 2012 in Wiley Online Library (wileyonlinelibrary.com).
DEVELOPMENTAL DYNAMICS 241:1487–1505, 2012
VC 2012 Wiley Periodicals, Inc.

regions of the midbrain, PITX3 is nec-essary for the terminal differentiationand survival of mesencephalic dopa-minergic neurons (mDA; van denMunckhof et al., 2003; Hwang et al.,2009). Zebrafish pitx3 morphants alsoexhibit small eyes with lens degenera-tion, along with misshapen heads, abent dorsal axis, and reduced jawsand fins (Shi et al., 2005). Disruptionof Pitx3 in Xenopus laevis impedes de-velopment of lens and retina, andrecent evidence suggests an addi-tional role in dorsal axis segmenta-tion and in laterality (Khosrowsha-hian et al., 2005; Shi et al., 2005;Smoczer et al., In Press). In zebrafish,Pitx3 expresses in the hypoblast ofgastrulating embryos (Dutta et al.,2005), and the transcript is detectableby reverse transcriptase-polymerasechain reaction (RT-PCR) in pregas-trula Xenopus (Khosrowshahianet al., 2005). These two studies
Dev
elop
men
tal D
ynam
ics
Fig. 1. Microarray data represented according to putative gene function. The 100 most up-and down-regulated transcripts affected by Pitx3-morpholino-mediated knockdown were cate-gorized by sequence analysis for stages 19 and 27 of X. laevis embryonic development. Colorscorrespond to functional groups in the legend (right).
Fig. 2.
1488 HOOKER ET AL.

suggest an earlier involvement forthe gene in dorso-anterior patterningthan is generally understood.
Pitx3 binds target DNA to regulatetranscription of downstream genes bymeans of bicoid binding elements(BBE; TAATCC; Lamonerie et al.,1996; Amendt et al., 1998). Pitx3directly regulates MIP/Aquaporin O,which encodes an abundant protein inthe lens that functions as an osmoticregulator and cell adhesion molecule(Chepelinsky, 2009; Huang and He,2010; Sorokina et al., 2011). In zebra-fish, pitx3 acts upstream of the tran-scription factor foxe3, which is neces-sary for the transition of lensepithelial cells into differentiated sec-ondary lens fibers by means of nucleardegradation (Shi et al., 2005). Pitx3 isalso thought to regulate the balancebetween mitosis and terminal differ-entiation in the equatorial region ofthe lens: here, it operates upstream ofcell cycle inhibitors p27Kip1 andp57Kip2 (Ho et al., 2009). Withinmidbrain regions, it directly regulatestyrosine hydroxylase (TH) expression,the rate-limiting enzyme in dopamineproduction (Landis et al., 1988; Lebelet al., 2001; Messmer et al., 2007). Italso controls the neurotransmission ofdopamine in mDA neurons by meansof regulation of vesicular monoaminetransporter 2 (VMAT2) and dopaminetransporter (DAT; Hwang et al., 2009).Direct regulation of Adh2 in mDAneurons affects the production of reti-noic acid that is necessary for properneuron development (Jacobs et al.,2007). To complicate matters, Pitx3 isa versatile transcription factor:depending upon signaling context, itcan act as either a transcriptional ac-
tivator or as a repressor (Cazorlaet al., 2000; Messmer et al., 2007).
We performed a microarray analysisto compare the transcriptomes of Pitx3-and control-morphants at stages 19(when eye development is commencing)and 27 (when lens differentiationbegins; Nieuwkoop and Faber, 1967).We elected to use morpholinos becauseectopic expression and dominant nega-tive approaches could affect theresponse elements of other Pitx familymembers: the ectopic expressionapproach is impossible to restrict solelyto Pitx3 expression domains, and thehomeodomain sequences of Pitx2 and 3,for example, are identical. Pitx2 and 3differ from Pitx1 by a single amino acidin the turn between helices I and II.
Although the preponderance of liter-ature regarding the gene relates tolens and mDA neurons, Pitx3 alsoexpresses broadly throughout gastru-lation, and later in somites, and lat-eral plate mesoderm (Pommereitet al., 2001; Khosrowshahian et al.,2005; Smoczer et al., In Press). Inzebrafish, pitx3 expresses in the de-marcation of the mesendoderm-derived polster (Dutta et al., 2005).Ectodermal explants have been usefulas source material for Xenopus micro-array experiments in the past, but thisrestriction to a single germinal layerwould miss some likely Pitx3 targets,and in addition would require the com-plicating necessity of neural inducingagents. That said, the interpretationof results can also be confounded bythe feature that morpholino-mediatedtranslational knockdown, unlike RNAiapproaches, solely affects translationand does not appear to affect mRNAdegradation rates. Indeed, some
embryos are suspected to compensatefor morpholino-mediated knockdownby releasing more transcript into cir-culation (Eisen and Smith, 2008).We designed our search for Pitx3 tar-
gets to be as broad as possible, and con-sequently we sampled from wholeembryos. The results generated a longlist of genes that are affected by Pitx3mis-regulation. We characterized noveltranscripts that represent putative tar-gets of Pitx3 and report plausiblegenetic pathways that are regulated bythis multifaceted transcription factor.
RESULTS AND DISCUSSION
Microarray Analysis
Morpholino specificity has been previ-ously published and reported to selec-tively reduce Pitx3 transcript and pro-tein levels, with the control-morpholinohaving none of these effects (Khosrow-shahian et al., 2005). This specificityhas subsequently been confirmed usinga second Pitx3 morpholino and mis-sense control (Smoczer et al., In Press).Xenopus microarray GeneChips (Affy-metrix) were used, and the data wereanalyzed comparing control-morpho-lino treatments to Pitx3-morpholinotreatments. The threshold for consider-ation was set at a two-fold cutoff with aP value of < 0.05. We categorized thetop 100 up- and down-regulated tran-scripts at each stage, with regard tofunction, and generated pie charts toshow their distribution (Fig. 1).Among gene categories, the largest
group affected consists of transcriptswith unknown function (expressedsequence tags; ESTs). Other tran-scripts encoded secreted factors andligands, transport and binding pro-teins, and modifying enzymes. Insummary, changes in expression pro-files for these genes implicate Pitx3 insome of the indirect controls uponmorphogenesis such as those exertingan effect by means of regulation ofsecreted morphogens.When assessed in broad strokes, the
secreted factors and ligands are nota-bly less up-regulated in morphants atstage 27 than at stage 19; however, bycontrast, transcription factors aremore up-regulated at stage 27. Atstage 19, structural proteins weremore profoundly affected (both up- ordown-regulated) as a consequence of
Dev
elop
men
tal D
ynam
ics
Fig. 2. In situ hybridization analysis for putative targets of Pitx3 involved in eye development. Vis-ual comparisons of gene expression patterns between right-side injected control-morpholino (Cmo)or Pitx3-morpholino (Pmo) embryos and their untreated contralateral control. A–E: Pitx3 expressionpatterns are presented for comparison (adapted from Khosrowshahian et al., 2005; Smoczer et al.,In Press). A: demonstrates faint but detectable signal throughout the ectoderm and in agreementwith reverse transcriptase-polymerase chain reaction (RT-PCR) results. B: Expression is detectablethroughout neural ridge, while at stage 22, the gene is expressed in a cleared specimen where anarrow indicates presomitic mesoderm. By stage 27 (D), Pitx3 is detectable throughout much of thehead ectoderm, as well as in branchial arches and somites. This pattern restricts later to somites,otic vesicle, lens, and brain (D). F–G0: Vent2 expression is reduced in the developing eye field atstage 19 for the Pitx3-morpholino (Pmo) injected side (A0 white arrow) and at stage 27 (B0 blackarrow), when compared with control-morpholino (Cmo) injected embryos (A,B). H–I0: Pax6 showsreduced expression in eye field on Pmo side of embryos at stage 19 (C0 black arrow) and 27 (D0
white arrow). J–J0: Crybb1 shows drastic loss of expression in the eye vesicle on the Pmo-treatedside of stage 27 embryo (E0) and no difference caused by Cmo treatment (E). Dotted line representsthe midline of the embryo, separating injected right side from contralateral left side control.
MICROARRAY OF PITX3 TARGETS 1489

Dev
elop
men
tal D
ynam
ics
TABLE
1.Data
SummaryforGenesAnalyzedforMicroarrayConfirmationa
Gen
eID
UniG
ene
ID
Gen
eHighestBLASTn
Hit
(Xen
opuslaevis)
Microarray
Ratio
Coinciden
tPathway
Inductive
Secon
dary
RT-PCR
Con
firm
s
Change
inIS
H
expression
pattern
Putative
BBE
sites
in5kbof
5’U
TR
Baz2
bXl.19899
EST-Mod
erately
similar
tobromod
omain
adjacentto
zinc
finger
dom
ain,2B
0.400(19)
HH
HNo
Yes
12
Bix4
Xl.399
hom
eobox
protein
BIX
4(bix4)
0.363(27)
HNo
N/A
N/A
Crybb1
Xl.21502
BetaB1-crystallin
(Crybb1)
0.325(27)
HYes
(27)
Yes
9
eFGF
Xl.11
81
fibroblast
growth
factor
4B
(fgf4-b)
0.406(19)
HNo
N/A
4
XeF
GF(ii)em
bryon
icfibroblast
growth
factor
Galectin
IXXl.15089
EST-Fish-egg
lectin-likeisoform
10.154(19)
HH
No(19)
N/A
N/A
0.246(27)
Yes
(27)
Gsc
Xl.801
Goosecoid
(gsc)
2.328(19)
HNo
N/A
14
4.743(27)*
2.267(27)*
Hes4
(Hairy2b)
Xl.25977
basic-helix-loop-helix
transcription
factor
hairy2b(hairy2)
0.299(27)
HH
HYes
(27)
Yes
(27)
14
Hes7†(E
sr4)
Xl.15142
EST-Highly
similarto
Xen
opuslaevis
Esr-4
0.381(27)
HH
No(27)
Yes
10
Hes7.1
†
(Xhr1)
Xl.12126
EST-Mod
erately
similar
totranscription
factor
HES-7.1-B
(XHR1)
0.211
(27)
HH
Yes
Yes
11
Hox
A11
Xl.266
Hom
eobox
A11
(Hox
A11
)0.423(27)
HNo
Yes
9Lim
1Xl.21652
EST-LIM
class
hom
eodom
ain
protein
(Lim
5/Lhx5)
(lhx1)(Xlim-2B)
2.046(27)
HH
No
Yes
5
L-M
af
Xl.767
neu
ralretinaleucine
zipper
(nrl)
0.357(19)
HNo
N/A
12
bZIP
transcription
factor
L-M
af(m
af)
Obscnl
Xl.13958
EST-Wea
kly
similarto
obscurin,cytosk
eletal
calm
odulinand
titin-interacting
RhoG
EF
6.211
(19)
HNo(19)
Yes
(27)
N/A
1490 HOOKER ET AL.

Dev
elop
men
tal D
ynam
ics
TABLE
1.(C
ontinued)
Gen
eID
UniG
ene
ID
Gen
eHighestBLASTn
Hit
(Xen
opuslaevis)
Microarray
Ratio
Coinciden
tPathway
Inductive
Secon
dary
RT-PCR
Con
firm
s
Change
inIS
H
expression
pattern
Putative
BBE
sites
in5kbof
5’U
TR
Pax6
Xl.647
Paired
box
6(pax6-b)
0.226(19)
HH
No(19)
Yes
13
0.477(27)
Yes
(27)
Rbp4l
Xl.17576
EST
6.164(19)
HYes
N/A
7Wea
kly
similarto
RET_B
Humanplasm
aretinol-bindingprotein
precu
rsor
(PRBP)
4.429(27)
Rdh16
Xl.5553
EST-retinol
deh
ydrogen
ase
16
(all-trans)
(rdh16)
6.288(19)
HH
No
N/A
15
2.758(27)
Ripply2
Xl.9206
Led
gerline(Stripy)
2.014(19)
HH
HYes
(19)
Ambiguou
s20
RXRa
Xl.877
retinoidX
receptor,
alpha(rxra)
0.319(19)
HNo
N/A
1
Spr1
Xl.17379
Sp5transcription
factor
(sp5)
0.441(19)
HNo
Ambiguou
sN/A
Sp1-likezinc-finger
protein
XSPR-1
Spr2
Xl.2755.1
Sp1-likezinc-finger
protein
XSPR-2
0.396(27)
HNo
Yes
N/A
GLIfamilyzincfinger
1,
gen
e2(gli1.2)
Ven
t2Xl.37
VENThom
eobox
2,
gen
e2(ven
tx2.2;Xom
)0.406(27)
HH
HH
Yes
Ambiguou
s17
Wnt1
Xl.21471
Wnt1
related(W
nt7c)
0.274(19)
N/A
N/A
N/A
80.172(27)
aCom
bined
insitu
hybridization
resu
ltswith
reverse
transcriptase-polymerase
chain
reaction
(RT-PCR)ou
tcom
e,compared
tothemicroarrayprediction
ofgen
etranscript
beh
aviorin
resp
onse
toxP
itx3
knockdow
n.Highlightedgen
esrepresentthebest-fitcandidatesforputativedirecttargetsof
Pitx3since
insitu
hybridization
andRT-PCR
con-
firm
themicroarraydata.Only
thegen
esthathadstatisticallysignificantRT-PCR
resu
ltsacrossthreereplicateswereindicatedon
table
as‘‘Y
es’’confirm
edbyRT-PCR.For
promoter
analysis,
putativePitx3andbicoid-bindingelem
ents
(BBE)weresearched
inthe5000bpupstream
region
from
ATG
ofX.trop
icalishom
olog
swhereavailable
at
Ensembl.org(TAATCC,TAATCT,TAATGG,TAATCA,andputativebindingsitesforPitx3;[Leb
elet
al.,2001]).
*Designate
multiple
Affymetrixprobesets
thatiden
tify
tothesa
megen
etranscript.
y Hes7andHes7.1
are
discreteproductsarisingfrom
separate
gen
esandthatsh
are
only
40%
aminoacididen
tity.Hes7sh
ares90%
iden
tity
withmurineHes7.
MICROARRAY OF PITX3 TARGETS 1491

Pitx3 knockdown than at stage 27. Asimilar picture developed for signaltransduction. The disruptions are con-sistent with embryos experiencingimpaired movement, signaling andmorphological changes during neuru-lation at stage 19, when the body planis arguably at its most ductile phase.Overall, chromatin modifying geneswere up-regulated more than down-regulated at both stages.
Our aim was to use the microarrayexperiments to deduce novel Pitx3pathways, so we first focused uponthe transcripts that were most up-and down-regulated in response tomorpholino-mediated knockdown ofPitx3. In published studies involvingsamples from rapidly developing sys-tems, microarray and RT-PCR resultshave occasionally been at odds. More-over, microarrays are likely to be sen-sitive to subtle differences in the stag-ing of developmental samples:quantitative data might not be fairlyinterpreted in absolute terms. Weelected to categorize on the basis oftrend: if gene expression levels werealtered two-fold or more relative tocontrols, and this was repeated in asecond experiment, we pursued thegene for further analysis using semi-quantitative RT-PCR analysis andriboprobe in situ hybridization. Genes
that expressed in expression patternsthat overlapped with Pitx3 weredeemed possible direct target genes ofPitx3. Of this subset, we focused uponthose that also possessed putativePitx3 binding motifs in the 50 untrans-lated region (UTR) of X. tropicalissequences. These were used for thereason that they were uniformlyavailable, and all of the Xenopus lae-vis ESTs and genes that we haveexamined to date enjoy near perfecthomology (Table 1). We then lookeddeeper into the data set to see if genesin the same signaling pathway or de-velopmental process were similarlyaffected (Table 2). If the behaviors ofthe expanded set grouped in a logicalmanner, and if the behaviors wereconsistent with the Pitx3 knockdownphenotypes, these genes were furtheranalyzed by RT-PCR or in situhybridization.
The affected genes can be classifiedas: potential direct targets of Pitx3;genes that operate within a Pitx3regulated pathway; or genes that areaffected indirectly and outside of thedomain of Pitx3 expression as aresult of grossly perturbed patternsof organ differentiation. Only fourgenes with putative Pitx3 bindingmotifs displayed both RT-PCR andriboprobe in situ hybridization pat-
terns that were unequivocally con-sistent with the microarray trend:Pax6, bb1Crystallin (Crybb1), Hes7.1,and Hes4. Two others, Vent2, andRipply2 (aka Ledgerline or Stripy)displayed altered in situ hybridiza-tion patterns that were difficult tointerpret with respect to expressionlevel, because their respective pat-terns were affected differently in dis-parate domains (Table 1). For exam-ple, although Vent2 expression isobliterated in the optic region consist-ent with the microarray trend, thegene is up-regulated in the posteriorendoderm. Similarly, the banded pat-tern of Ripply2 expression is anteri-orized and delayed by morpholino atearly stages, but appears to recoverto some extent by stage 27.In X. laevis, Pitx3 expresses in the
developing lens, the otic vesicle, andhead mesenchyme, as well as in thebranchial arches and along the ante-roposterior axis in the developingsomites (Pommereit et al., 2001;Khosrowshahian et al., 2005). Insofaras Pitx3 is critical to lens placodefunction, it plays a critical role in frogretina induction (Khosrowshahianet al., 2005), so one might expect geneexpression in retina to be indirectlyaffected as well. Eye pathway genesPax6, L-Maf, and Crybb1, express in
Dev
elop
men
tal D
ynam
ics
TABLE 2. Additional Genes Identified in the Microarray Data That Pertain to Genetic Pathways Implicated in this
Study
Gene ID UniGene ID Gene Highest BLASTn Hit (Xenopus laevis) Microarray ratio
Rax1 Xl.186 Retina and anterior neural fold homeobox (Rax-a) 2.148 (19)bB3-crystallin Xl.26355 Crystallin, beta B3 (crybb3) 0.366 (27)c-crystallin-like Xl.23710 Transcribed locus, strongly similar to NP_001087320.1
crystallin, gamma A (X. laevis)0.140 (19)
cB-crystallin Xl.21441 Crystallin, gamma B (crygb) 0.298 (19)bB3-crystallin-like Xl.26349 69% similar to beta-crystallin B3 (H. sapiens) 3.800 (27)R>bA4-crystallin Xl.19126 (retired)
replaced Xl.67080Crystallin, beta 4 (cryba4) 0.223 (19)
2.741 (27)bB1-crystallin-like Xl.1337 Transcribed locus, strongly similar to XP_002938264.1
predicted: beta-crystallin B1-like (X. tropicalis)2.122 (27)
Tbx4 Xl.21543 T-box 4 (tbx4) 0.436 (27)*0.237 (27)*
Tbx5 Xl.529 T-box 5 (tbx5-b) 0.432 (27)HoxA10 Xl.21639 Homeobox A10 (hoxa10) 0.373 (27)HoxA13 Xl.21581 Homeobox A13 (hoxa13) 2.337 (27)Galectin I Xl.747 Lectin, galactoside-binding, soluble, 1 (lgals1) 2.264 (19)
0.436 (27)Galectin IIa Xl.17371 Galectin family xgalectin-IIa (xgalectin-IIa) 0.291 (19)Galectin IIb Xl.21879 Galectin 4 (lgals4-a) 2.367 (19)Galectin IIIa Xl.15364 Lectin, galactoside-binding, soluble, 9c (lgals9c-a) 0.414 (27)Galectin IIIb Xl.21878 Lectin, galactoside-binding, soluble, 9c (lgals9c-b) 0.304 (19)
1492 HOOKER ET AL.
the developing lens, and thus are goodcandidates for Pitx3 targets. Vent2,Rbp4l (purpurin), Galectin IX, andRax1 express in early retina, and are
all affected in morphants. They likelyrepresent examples of the indirectconsequences of Pitx3 perturbation.Moreover, a microarray survey of
Aphakia mice revealed a linkbetween Pitx3 perturbation and reg-ulation of Pax6 and Rbp4 (Munster,2005). All of the aforementioned
Dev
elop
men
tal D
ynam
ics
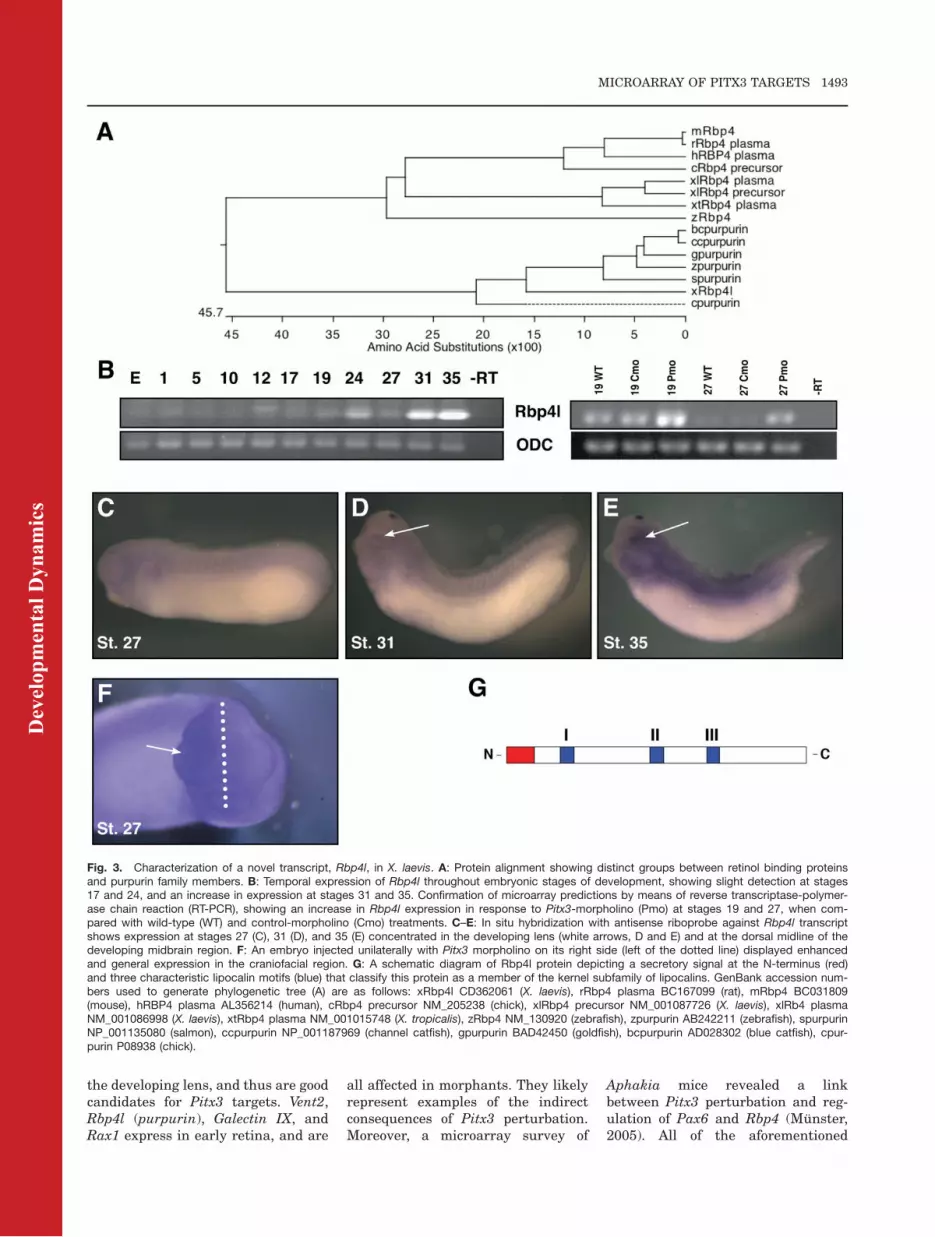
Fig. 3. Characterization of a novel transcript, Rbp4l, in X. laevis. A: Protein alignment showing distinct groups between retinol binding proteinsand purpurin family members. B: Temporal expression of Rbp4l throughout embryonic stages of development, showing slight detection at stages17 and 24, and an increase in expression at stages 31 and 35. Confirmation of microarray predictions by means of reverse transcriptase-polymer-ase chain reaction (RT-PCR), showing an increase in Rbp4l expression in response to Pitx3-morpholino (Pmo) at stages 19 and 27, when com-pared with wild-type (WT) and control-morpholino (Cmo) treatments. C–E: In situ hybridization with antisense riboprobe against Rbp4l transcriptshows expression at stages 27 (C), 31 (D), and 35 (E) concentrated in the developing lens (white arrows, D and E) and at the dorsal midline of thedeveloping midbrain region. F: An embryo injected unilaterally with Pitx3 morpholino on its right side (left of the dotted line) displayed enhancedand general expression in the craniofacial region. G: A schematic diagram of Rbp4l protein depicting a secretory signal at the N-terminus (red)and three characteristic lipocalin motifs (blue) that classify this protein as a member of the kernel subfamily of lipocalins. GenBank accession num-bers used to generate phylogenetic tree (A) are as follows: xRbp4l CD362061 (X. laevis), rRbp4 plasma BC167099 (rat), mRbp4 BC031809(mouse), hRBP4 plasma AL356214 (human), cRbp4 precursor NM_205238 (chick), xlRbp4 precursor NM_001087726 (X. laevis), xlRb4 plasmaNM_001086998 (X. laevis), xtRbp4 plasma NM_001015748 (X. tropicalis), zRbp4 NM_130920 (zebrafish), zpurpurin AB242211 (zebrafish), spurpurinNP_001135080 (salmon), ccpurpurin NP_001187969 (channel catfish), gpurpurin BAD42450 (goldfish), bcpurpurin AD028302 (blue catfish), cpur-purin P08938 (chick).
MICROARRAY OF PITX3 TARGETS 1493
Dev
elop
men
tal D
ynam
ics
Fig. 5.
Fig. 4.

provide validation for the efficacy ofthe microarray. Unfortunately, noneof the previously published and char-acterized targets of Pitx3 are repre-sented on the microarray; however,one of the probe sets is to an ESTthat has homology to MIP/Aqua-porin O, and it is down-regulatedconsistent with expectation.
Riboprobe In Situ
Hybridization
We assessed the effect of Pitx3 pertur-bation by injecting embryos at thetwo-cell stage such that the left andright sides of the developing embryocould be compared as embryogenesisensued: morphant phenotypes weremonitored on the ‘‘mutant’’ side rel-ative to the contralateral control.Candidate gene expression patternswere assessed for perturbation inmorphants and for a role in devel-oping eye (Figs. 2–7), brain (Fig. 8),somite (Fig. 9), and tailbud(Fig. 10).
Eye development.
Among other domains, Vent2 (a.k.a.Ventx2) is expressed in the dorsal ret-ina (Fig. 2F,G) and it shows structuraland functional homology to two Dro-
sophila proteins, Om1D and BarH1,which are necessary for the differen-tiation of photoreceptor cells in the eye(Ladher et al., 1996). Along withVent2, Pax6, and Crybb1 are perturbedin Pitx3morphants (Fig. 2H–J).
Pax6 is required and sufficient forthe initiation of eye developmentwhere it specifies the lens and retinalprimordia (Halder et al., 1995), and ittoo is perturbed in our assays. Themicroarray and RT-PCR data regard-ing L-Maf’s response to Pitx3 perturba-tion was ambiguous but is neverthelessworth following up: its relationship toPitx3 has not been directly assessed;however, Maf binding sites are deletedin the promoter of a naturally occur-ring mouse Pitx3 mutant (Seminaet al., 2000) and L-Maf itself appears toreciprocally possesses 12 putative Pitx3binding motifs in its 50-UTR. L-Maf isexpressed in the developing lens inresponse to inductive events from theoptic vesicle, and it is directly targetedby Pax6 in chicks (Reza et al., 2002).Maf acts specifically in the lens fibercells, where it can induce the expres-sion of structural proteins such as they- and bb1-crystallins (Crybb1; Ishiba-shi and Yasuda, 2001; Cui et al., 2004).Given the presence of numerous poten-tial Pitx3 binding sites in the Crybb1promoter, and the response of this gene
in our Pitx3 morphants, we speculatethat Maf and Pitx3 act in tandem toactivate the Cry genes. It is worth not-ing that other Cry genes representedon the microarray also underwent sig-nificant fractional change, albeit at lessspectacular levels, namely: g crystallin(0.14), g B crystallin (0.3), b B3 crystal-lin (3.8), b A3 crystallin (2.74), and spe-cies weakly similar to human b B1crystallin (2.12), and b B3 crystallin(0.37).
Novel Xenopus retinol-bindingprotein Rbp4l is expressed inlens. The microarray indicated thatan EST sequence encoding a 197amino acid protein (GenBankCD362061) was up-regulated atstages 19 and 27 by 6.2- and 4.4-fold,respectively. We obtained a clone fromNIBB (XL060f11) and after sequenc-ing it, we identified it as a member ofthe lipocalin protein family, namelyRBP4-like (Retinoid binding protein 4like -Rbp4l) or purpurin. These smallextracellular proteins characteristi-cally bind hydrophobic molecules andare typically known as transport pro-teins (Flower, 1996). Figure 3 showsthat Rbp4l shares 73% residue iden-tity with goldfish and salmon, 75%identity with zebrafish, and 78% simi-larity to chick Rbp4l. The similarity tohuman and murine retinoid-bindingprotein precursor is on 55 and 54%,respectively. Rbp4l consists of threeconserved motifs that create a cup-shaped cavity, enabling the protein tobind retinol, and the protein possessesa signal peptide for secretion (Bermanet al., 1987). In zebrafish, rbp4l istranscribed in photoreceptor cells,and the protein is diffusely detectablein all retinal layers (Tanaka et al.,2007). As a supplier of retinol, a pre-cursor of retinoic acid, this proteinactivates the retinoic acid and reti-noid receptor pathway (RAR andRXR, respectively; Nagy et al., 1996).Rbp4l functions as an extracellularmatrix protein in the inter-photore-ceptor matrix, and it appears to benecessary for cell adhesion and for thesurvival of photoreceptor cells in theneural retina (Berman et al., 1987;Nagy et al., 1996). Photoreceptor cellsrequire retinol for phototransductionand retinol is carried to them from thepigmented retinal layer, through thematrix, bound to Rbp4l. In contrast,
Dev
elop
men
tal D
ynam
ics
Fig. 4. Characterization of a novel transcript Galectin IX in X. laevis. A: Protein alignmentshowing amino acid similarities between Xenopus Galectin family members. B: Temporal expres-sion of Galectin IX throughout embryonic stages of development, shows expression beginningat gastrulation (stage 10), decreasing at stage 12, and expressing consistently at stages 17through 35. Confirmation of microarray predictions by means of reverse transcriptase-polymer-ase chain reaction (RT-PCR), detect an increase in expression at stage 19 and a decrease atstage 27 for Pitx3-morpholino (Pmo) -treated samples, compared with wild-type (WT) and con-trol-morpholino (Cmo). C–E: Galectin IX transcript expresses at stages 24 (C), 27 (D), and 31 (E)concentrated in the developing eye (white arrows) and presumptive pronephros, persisting in thenephric tubules and ducts. GenBank accession numbers used to generate phylogenetic tree (A)are as follows: xGalectinIa AB056478, xGalectinIb AB060969, xGalectinIIa AB060970, xGalecti-nIIb AB080016, xGalectinIIIa AB060971, xGalectinIIIb AB080017, xGalectinIVa AB060972, xGa-lectinVa M88105, xGalectinVb AB080018, xGalectinVIa AB080019, xGalectinVIIa AB080020,xGalectinVIIIa AB080021, xGalectinIX BJ056659.
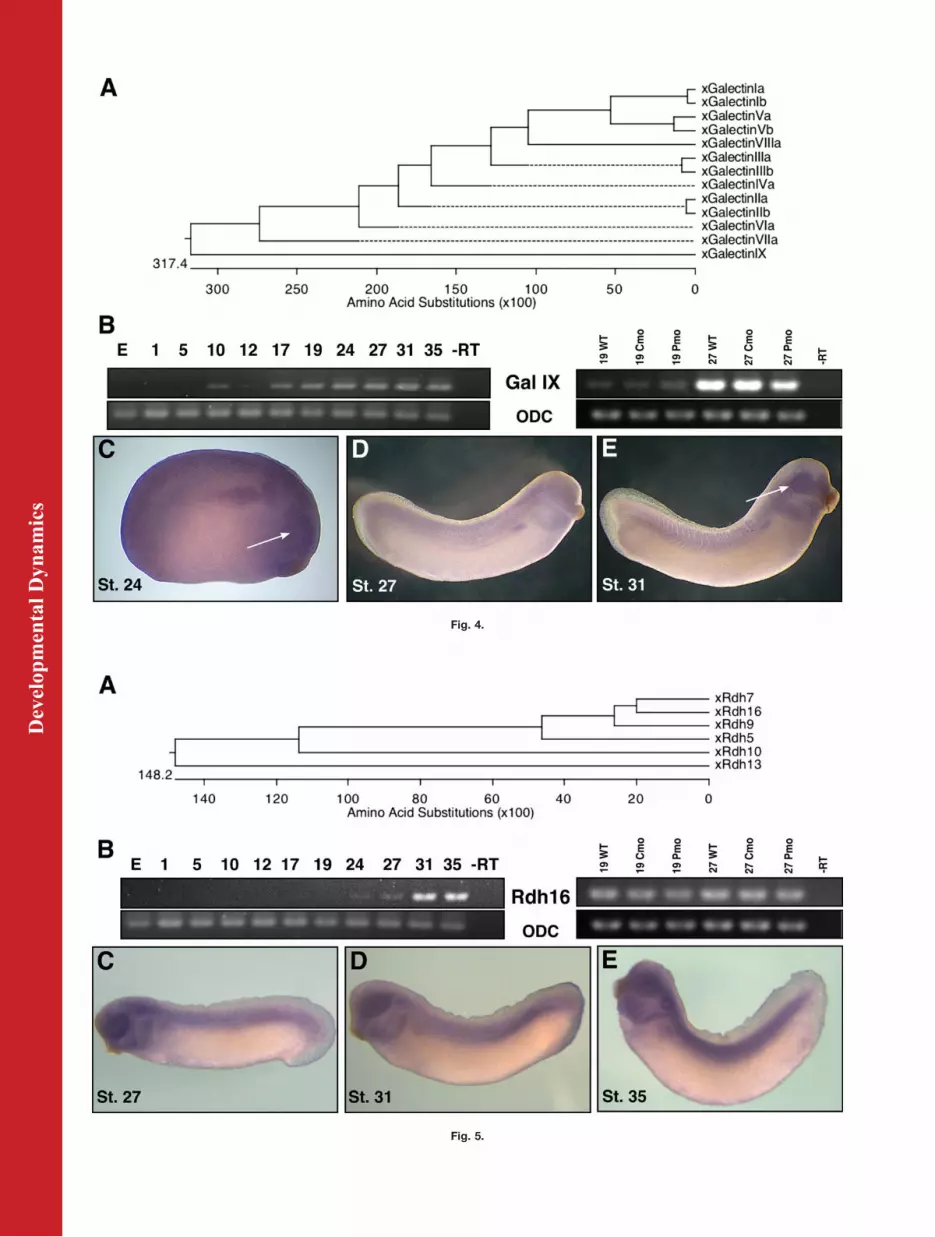
Fig. 5. Characterization of a novel transcript, Rdh16, in X. laevis. A: Protein alignment showingamino acid similarities between Xenopus retinol dehydrogenase (rdh) family members. B: Tem-poral expression of Rdh16 throughout embryonic stages of development shows faint expressionbeginning at stage 24 and 27, then increasing at stages 31 and 35. We were unable to confirmthe microarray predictions by means of reverse transcriptase-polymerase chain reaction (RT-PCR), as no change in expression was detected between wild-type (WT) control-morpholino(Cmo), or Pitx3-morpholino (Pmo) embryos. C–E: In situ hybridization with antisense riboprobeagainst Rdh16 transcript, shows expression at stages 27 (C), 31 (D), and 35 (E) concentrated inthe eyecup, branchial arches, and otic vesicle, as well as along the lateral plate mesoderm, witha focus on the posterior half (D), and on in the developing myotomes. GenBank accession num-bers used to generate phylogenetic tree (A) are as follows: xRdh16 NP_001083356, xRdh7NP_001079189, xRdh13 NP_001085680, xRdh5 NP_001086194, xRdh9 NP_001090337, xRdh10ACN32204.
MICROARRAY OF PITX3 TARGETS 1495

Dev
elop
men
tal D
ynam
ics
Fig. 6.
Fig. 7.
Fig. 8.

the other RBPs, including Rbp4l’sclosest human homologue RBP4, aresynthesized in the liver, bind to reti-nol in the blood (serum RBPs), andthey transport retinol throughout thebody to target cells (Goodman, 1981).Human PITX3 maps to 10q25, andthis is close to human RBP4 and sev-eral retinoid synthetic CYP loci at10q24 (Gray et al., 1997). According tothe Ancora resource, the region nearPitx3 is replete with highly conservednon-coding elements, so it is temptingto speculate that the genes are embed-ded within a conserved genome regu-latory block (Kikuta et al., 2007; Eng-strom et al., 2008).
Expression of Rbp4l is first detectedby RT-PCR around stage 17 andincreases past stage 35 (Fig. 3B). Insitu hybridization shows that expres-sion of this transcript concentrates inthe lens area and as a pronouncedspot along the midline on the top ofthe brain. It expresses at lower levelsin the craniofacial region and somites(Fig. 3C–E). These expression pat-terns are distinct from those reportedfor RBP4 and purpurin. RT-PCR anal-ysis was performed and confirmedmicroarray trends: morphants demon-
strated an increase in expression atstage 19 (1.84-fold) and 27 (2.88-fold;Fig. 3B). Consistent with the microar-ray and RT-PCR data, the gene under-goes up-regulation as a consequenceof Pitx3 knockdown (Fig. 3F). BecauseRbp4l expression in Pitx3 morphantsis broadly up-regulated in the cranio-facial region, our supposition is thatPitx3 exerts its effects upon this geneearlier than the lens stage, and whenPitx3 expression is more expansive.The murine homolog, Rbp4, is alsoaffected by Pitx3 depletion in Aphakiamutants (Munster, 2005). Taken to-gether, the results for this novel reti-nol binding protein show the possibil-ity of acting downstream of Pitx3 inlens developmental pathways, whereboth genes are expressed.
Galectin IX is expressed in eyefield and retina. One of the ESTsequences from the microarray dataidentified mostly with the Galectinfamily, and represents a new familymember (Fig. 4). We identify thissequence as a Galectin IX (GenBankaccession no. JN975639). It is relatedto the tectonin family that encodebeta-propeller repeats: the microar-
ray reports a change in transcript lev-els at stage 19 (diminished to a frac-tional level of 0.15) and stage 27(diminished to 0.25 of its formerlevel). The function of a galectin canbe extremely varied: it has intracellu-lar and extracellular functions in celladhesion, migration, proliferation,and apoptosis and that are stage- andtissue-specific (Cooper and Barondes,1999).Galectin IX, a gene uncharacterized
with regard to expression patternsuntil this study, expresses in eye fieldand later in both lens and retina (Fig.4). Little is known of its promoterstructure, so it is early to speculatewhether or not the gene is a directtarget of Pitx3. In Xenopus alone, 12different galectin proteins have beenidentified, numbered in order of dis-covery, and can be identified by meansof galactose-binding ability and pro-tein motifs, specifically carbohydraterecognition domains (Shoji et al.,2003). Other Galectin family mem-bers are expressed throughout theembryo in specific spatiotemporal pat-terns, suggesting varied developmen-tal roles for each protein (Shoji et al.,2003). Additional galectins were iden-tified in the microarray data: GalectinIIb (St.19 2.37-Fold), Galectin I (St.192.26-Fold, St.27 0.44-Fold), GalectinIIIb (St.19 0.30-Fold), Galectin IIa(St.19-0.29), Galectin IIIa (St.27-0.41). As a candidate Galectin, furtherfunctional assessment for galactose-binding affinity will be necessary tofirmly classify this novel proteinwithin the galectin family (Cooper andBarondes, 1999). Using an NIBB clone(XL103j23) we performed in situhybridization to visualize the expres-sion pattern of this novel transcript,which appears to be concentrated inthe presumptive pronephros and eyeregions (Fig. 4C–E). Expression beginsat gastrulation, fades and thenincreases gradually beginning at neu-rulation (Fig. 4B). Curiously, RT-PCRfor microarray confirmation (Fig. 3B)shows a fractional increase in expres-sion at stage 19 (5.28), but the expectedslight decrease at stage 27 (0.83) inmorphants. This interaction is likelyindirect because, even though expres-sion patterns of Pitx3 and Galectin IXoverlap, in situ hybridizations do notdemonstrate obvious changes of Galec-tin IX expression in morphants.
Dev
elop
men
tal D
ynam
ics
Fig. 6. In situ hybridization analysis for putative Pitx3 target genes Obscnl and Baz2b. Visualcomparisons of gene expression patterns between control-morpholino (Cmo) and Pitx3-morpho-lino (Pmo) right side-injected embryos. A: Obscnl shows a loss of expression in the branchialarches (black arrow), otic vesicle, and retina when treated at stage 27 with Pmo (A0) versus Cmo(A). B: Baz2b is substantially reduced in response to Pmo (B0) in the retinal layer of the opticprotuberance (white arrow), as well as in the pronephros and in the anterior region of the dorsalaxis, when compared with Cmo (B).
Fig. 8. In situ hybridization analysis for putative brain targets of Pitx3. Comparisons of geneexpression patterns between right-side injected control-morpholino (Cmo) or Pitx3-morpholino(Pmo) embryos and their untreated contralateral control. A–B0: Hes7.1 at stage 19 showsdecreased expression in the midbrain hindbrain boundary or isthmus (black arrow) in responseto Pmo (A0) versus Cmo (A) and again at stage 27 Pmo (B0) (black arrow) versus Cmo (B). C–D0:Spr1 stained embryos show increased expression (black arrow) at stage 19 when treated withPmo (C0), where no change in expression is observed with Cmo (C). At stage 27, Spr1 expres-sion in the isthmus is abolished on the Pmo side (D0) (white arrow). Dotted line represents themidline of the embryo, separating injected right-side from contralateral left-side control.
Fig. 7. Characterization of a novel transcript, Baz2b, in X. laevis. A: Protein alignment showingamino acid similarities between Baz2B homologs across organisms. B: Temporal expression ofBaz2b throughout embryonic stages of development show expression as a maternal transcriptin the egg ‘‘E’’ and throughout development to tailbud stage, with slight reductions in transcriptlevel at stages 10 and 19. B Confirmation of microarray predictions by means of reverse tran-scriptase-polymerase chain reaction (RT-PCR) show abolished expression at stage 27 inresponse to Pitx3-morpholino (Pmo) when compared with control-morpholino (Cmo) and wild-type (WT) embryos. C–E: Baz2b expression at stages 21 (C), 31 (D), and 35 (E) is concentratedin the developing eye, as well as the branchial arches and otic vesicle. Dark expression is seenin the pronephros, persisting in the tubules (E). F: A schematic diagram of Baz2b protein depict-ing various domains characteristic of Baz2B: methyl-CpG binding domain (MBD), DNA bindingdomain (DDT), zinc finger domain (Z), adjacent to the bromodomain (BR). GenBank accessionnumbers used to generate phylogenetic tree (A) are as follows: xBaz2b BQ400337 (X. laevis),mBaz2b BC150814 (mouse), rBaz2b NM_001108260 (rat), hBAZ2B NM_013450 (human),cBaz2b NM_204677 (chick), xtBaz2b BC166361 (X. tropicalis).
MICROARRAY OF PITX3 TARGETS 1497

Dev
elop
men
tal D
ynam
ics
Fig. 9. In situ hybridization analysis for putative segmentation targets of Pitx3. Visual comparisons of gene expression patterns between right-side injected control-morpholino (Cmo) or Pitx3-morpholino (Pmo) embryos and contralateral control. A–B0: Ripply2 expression, showing as twostripes in the presomitic mesoderm, shows an anterior shift (black arrow) in expression at stage 19 when treated with Pmo (A0) instead of Cmo (A).At stage 27, Ripply2 expression pattern loses its distinct shape and becomes unrestricted in response to Pmo (B0), whereas with Cmo treatment,precise patterning of this gene expression remains intact (B). C–D0: Hes4 expression becomes blurred in Pmo-treated embryos at stage 19 (C0)and at stage 27 (D0) Hes4 expression is absent in the presomitic mesoderm (black arrow) and pronephros areas, compared with Cmo-treatedembryos (D). E–F0: Hes7 no longer expresses in the most anterior stripe (black arrow), and the remaining two stripes are shifted anteriorly in com-parison to the contralateral control (E0). At stage 27, on the Pmo side of the embryo (F0), Hes7 shows increased expression in the presomitic mes-oderm (white arrow) and again an anterior shift of the striped pattern (black arrow). Dotted line represents the midline of the embryo, separatinginjected right-side from contralateral left-side control.
Fig. 10.
1498 HOOKER ET AL.

Novel Xenopus RetinolDehydrogenase (Rdh16). An ESTsequence found in the microarraydata can be identified as retinol dehy-drogenase 16 (Rdh16) (Fig. 5).Because retinoic acid is pertinent tomany developmental processes, andPitx3 has already been shown to regu-late an aldehyde dehydrogenase,AHD2 (Jacobs et al., 2007), thissequence is interesting as a putativedownstream target of Pitx3. Retinoldehydrogenases are enzymes that cat-alyze the conversion of retinol (vita-min A) to retinal, an intermediate inthe biosynthesis pathway of retinoicacid (Pares et al., 2008). Theseenzymes belong to the short-chain de-hydrogenase/reductase (SDR) family.Their substrate is retinol bound toCRBP (cellular retinol binding pro-tein) (Napoli et al., 1991) and theyappear to be differentially expressedin different tissues (Chai et al., 1996).Their differential expression suggeststissue-specific roles for different fam-ily members. Xenopus Rdh16 shows51% similarity to human 11-cis RDH.The 11-cis RDH is synthesized in theretinal pigmented epithelium, is nec-essary for the generation of 11-cis ret-inaldehyde from retinol, and bindsvisual pigments in the eye (Wald,1968; Simon et al., 1995, 1996). Micro-array predicts a fold change of 6.288at stage 19 and 2.758 at stage 27 forthis transcript. We were unable toconfirm this by RT-PCR (Fig. 5B) or insitu hybridization. We rule this geneout as a Pitx3 target.
The expression of this retinol dehy-drogenase appears only in tailbudstages and is concentrated in the reti-nal layer of the developing retina, pe-ripheral lens, otic vesicle, branchialarches, and along the anteroposterioraxis in a gradient intensified at theposterior half (Fig. 5C–E). If this geneis a homolog of human 11-cis RDH,the expression in the eye would sup-port a conserved functional role.
Novel Xenopus Genes Obscurin-Like and Chromatin-RemodelingProtein Baz2b. Other genes maybe indirect targets of Pitx3 such asobscurin-like (Obscnl) in the eye fieldand branchial arches (Fig. 6A), and achromatin remodeling gene Baz2b(Figs. 6B, 7). Obscnl, is an ESTweakly similar to obscurin, cytos-keletal calmodulin, and titin-interact-ing RhoGEF. Because neither gene’sexpression pattern is altered in allPitx3-expressing domains, it seemslikely that they are affected by themorphological changes induced byPitx3 knockdown, and thus should beconsidered indirectly affected.
The EST with homology to the BAZfamily of bromodomain-containingproteins (bromodomain adjacent tozinc finger) is tentatively assigned thedesignation Xenopus Baz2b (Gen-Bank accession no. JN975638). Theclone represents the 50 half of asequence encoding the N-terminus(921aa). This protein family containsa conserved bromodomain at the C-terminus, adjacent to a PHD zinc fin-ger motif (Fig. 7F). Bromodomains,capable of binding acetyl-lysine resi-dues, are often found in proteins withhistone acetyltransferase (HAT) activ-ity, and they are thought to play arole in chromatin-dependent generegulation by unwinding histone-DNA complexes (Zeng and Zhou,2002). Baz2b may have the ability tobind methylated CpG regions througha methyl-CpG binding domain (MBD;Fig. 4F). There is some evidence ofBAZ proteins having the ability tointeract with human homologs ofISWI, which in Drosophila, binds theBAZ1 protein homolog Acf1 to formthe ACF chromatin remodeling com-plex (Ito et al., 1999; Jones et al.,2000a,b).
The microarray predicts that atstage 19 this transcript decreases inmorphants to a fraction of 0.4 and atstage 27 to a fraction of 0.27. Unfortu-
nately, by RT-PCR stage 19 transcriptis just at the limit of detectability. RT-PCR shows expression throughoutembryogenesis, beginning as a mater-nal transcript in the oocyte and per-sisting through tailbud stages, andconfirms the microarray data by show-ing a drastic decrease in expression atstage 27 (to a fraction of 0.086), withundetected expression at stage 19(Fig. 7B). Its spatial expression pat-tern, initially quite diffuse (notshown), condenses around the devel-oping eye and pronephric structuresduring tailbud stages (Fig. 7C–E).Since Pitx3 has been shown to play
major roles in both the lens and retinadevelopment, these genes correlatewith a role for this transcription fac-tor in specifying lens placode, initiat-ing lens differentiation, and in induc-ing retina (Khosrowshahian et al.,2005).
Brain expression.
One candidate sequence was highlysimilar to Hes-related 1, and is tenta-tively re-assigned the name Hes7.1based upon homology to the X. tropica-lis and human genes. This gene likelyspecifies the frog midbrain/hindbrainboundary, or isthmus (Shinga et al.,2001; Takada et al., 2005). The isth-mus is an important organizer of brainregionalization and consequent pat-terning (Nakamura and Watanabe,2005). When murineHes1 is disrupted,brain patterning mediated through theisthmus is damaged, and the mesence-phalic dopaminergic (mDA) neuronsfail to thrive. The same authors reportthat expression of both Pitx3 and tyro-sine hydroxylase is abnormal (Kamedaet al., 2011). Because the related Xeno-pus homolog possesses 11 putative Pitxbinding motifs, future studies shouldbe sensitive to the possibility thatHes1/Hes7.1 and Pitx3 are engaged ina reciprocally regulatory relationship.Spr1, a Xenopus laevis transcriptionfactor that is related to the human Sp1and mouse Sp5 zinc finger genes, isexpressed in the forebrain as well asthe isthmus, where eFGF also plays arole (Isaacs et al., 1992; Ossipova et al.,2002). Both Spr1 and Hes7.1 showdecreased expression in the isthmusin response to Pitx3-morpholino asassessed by in situ hybridization(Fig. 8).
Dev
elop
men
tal D
ynam
ics
Fig. 10. In situ hybridization analysis for putative tailbud targets of Pitx3. A–B0: HoxA11 showsdecreased posterior expression in the tailbud region (white arrows) of Pmo embryos at stages19 (A0) and 27 (B0); C–D0: Spr2 displays a broader and larger domain of expression (blackarrows) when treated with Pmo, both at stage 19 (C0) and 27 (D0), compared with Cmo-treatedembryos (C, D). E–F0: Lim1 expression disappears from paraxial mesoderm (red arrow) and isup-regulated in lateral mesoderm (black arrow) at stage 19 when treated with Pmo (E0). At stage27 (F0), Pmo reduces Lim1 expression in the developing pronephros (black arrow) and in thehead mesenchyme and along the dorsal axis.
MICROARRAY OF PITX3 TARGETS 1499

Unfortunately, tyrosine hydroxy-lase, a gene critical to differentiationof dopaminergic neurons (mDA) of thesubstantia nigra, is not representedon the microarray. However, Wnt1, anearly stage marker for murine isth-mus (Wurst et al., 1994), is both rep-resented on the microarray anddown-regulated (Table 2). Only anunworkably small fragment of thegene has been cloned in frog (Woldaand Moon, 1992). Because Pitx3 isespecially pertinent for the differen-tiation and maintenance of mDA neu-rons and because the isthmus is criti-cal to development of the substantianigra (Marchand and Poirier, 1983), itis tempting to speculate that thisPitx3 effect is mediated through con-trol of isthmus patterning at early de-velopmental stages.
The expression patterns of Lim1will be discussed a greater lengthlater, however, it is worth noting inthe context of isthmus and substantianigra (structures that are inducedand patterned early by Lim1 [Shawlotand Behringer, 1995]), that althoughthe RT-PCR assays did not confirmthe microarray data, nevertheless, insitu hybridization did. Moreover,Lim1 possesses 5 evolutionarily con-served Pitx3 binding motifs. Basedupon our preliminary slate of putativesignaling partners, our suspicion isthat Pitx3 plays a heretofore unchar-acterized role during gastrulation topattern anterior-most structures—previous work has indicated that itexpresses in fish hypoblast (Duttaet al., 2005), and somewhere in Xeno-pus pregastrula (RT-PCR, uncharac-terized and low-expression location;Khosrowshahian et al., 2005).
Segmentation and tailbud
signaling.
The Ripply family, Ripply1 (bowline),Ripply2 (ledgerline, stripy) and Rip-ply3 serve as transcriptional repress-ors that are necessary for properboundary formation during somito-genesis. The Ripply genes appear toact by balancing the FGF/RA signal-ing wave front and thereby regulatethe emergence of new somites: thisregulation is likely mediated by inter-action with T-box genes (Chan et al.,2006; Kawamura et al., 2008; Hitachiet al., 2009). It is interesting that
both Tbx4 and Tbx5 go down in ourdata set (0.237 and 0.436 for each ofthe two Tbx4 probands, and 0.432 forTbx5). Ripply2, Hes4, and Hes7 areperturbed in Pitx3 morphants (Fig. 9),and Ripply2 possesses 20 Pitx3 bind-ing sites in its 50-UTR. Hes7 expres-sion patterns confirmed the microar-ray data, however, triplicate RT-PCRreactions did not substantiate thisstatistically. We note that RT-PCRconsistency has historically been aproblem in microarray studies (Alt-mann et al., 2001; Buchtova et al.,2010), and given the presence of 10Pitx3 binding motifs within the 50-UTR of Hes7, we are inclined to pur-sue this gene’s candidacy further. Per-turbation of Hes4 is complex: itappears to up-regulate at earlystages, to remain unchanged throughneurulation, but to be inhibited attailbud stages (Smoczer et al., InPress). Hes4 and Hes7 are factors thatfunction downstream of the Notchpathway during somitogenesis andthat mediate segmental patterning ofthe presomitic mesoderm where theyserve as components of the segmenta-tion clock (Jen et al., 1999; Tsuji et al.,2003; Murato et al., 2007). Recently,presomitic expression has beenreported for Pitx3 and its perturbationresults in anomalous segmentationpresenting as a bent dorsal axis andaberrant somite morphogenesis(Smoczer et al., In Press). Ripply2morphants also produce bent dorsalaxes and shift Hes4 and Hes7 expres-sion patterns anteriorly (Chan et al.,2006). Further research is necessaryto deduce which of these are directdownstream targets of Pitx3, but agood starting point would be to test ifPitx3 modulates Ripply2 and therebyindirectly alters expression of the Hesgenes.
Both eFGF and RXRa are tran-scribed in the tailbud and thus maybe factors that are affected by Ripply2(Chan et al., 2006). eFGF extends tothe posterior of the body axis and intothe proliferating tailbud where noto-chord and somites continue toemerge. eFGF is also expressed laterin the myotome of the trunk (Isaacset al., 1992). Both eFGF and RXRaappear regulated by Pitx3 in themicroarray dataset, but neither con-firm by RT-PCR. The expression lev-els are too low to be reliably detected
by in situ hybridization at stage 19and 27; however, both possess consen-sus Pitx3 binding motifs in their re-spective 50-UTR. Given the effects ofPitx3 perturbation upon the somito-genesis- and tailbud-expressing genesHoxA11, Spr2, and Lim1 (Fig. 10), itmight be worth re-examining theirfailed candidacy as targets.Spr2 and HoxA11 are affected by
Pitx3 mis-regulation (Fig. 10).HoxA11 specifies positional identityalong the anteroposterior axis and islargely expressed in the posteriornotochord and tailbud mesoderm(Lombardo and Slack, 2001). OtherHox genes are affected to a lesser,though still significant fractionaldegree: HoxA13 (2.4), and HoxA10(0.37). The differential effect uponthese genes renders an indirect medi-ation by retinoid metabolism unlikely.Lim1 expression undergoes a complexmodulation of expression: lateral mes-oderm expression increases, while inparaxial mesoderm, expression isabolished. Spr2 and Vent2 areexpressed in the developing tailbud(Ladher et al., 1996; Ossipova et al.,2002), so effects in this domain wouldalso be reflected in the microarray.
Indirectly characterized early
perturbation effects.
Although the microarray data was an-alyzed for embryos at stages 19 and27, a significant number of candidatesare pertinent for early patterning ofthe embryo, and moreover, are knownto interact with each other in a man-ner consistent with Pitx3 impingingupon their respective regulatory net-works. Pitx3 has been detected atearly stages in the embryo (stage 8;Khosrowshahian et al., 2005) suggest-ing an unknown function for thistranscription factor at earlier stages.One of our candidate targets, Vent2,provides ventralizing information andperhaps signals for the differentiationof the epidermis (Ladher et al., 1996).This factor directly down-regulatesthe homeobox gene Goosecoid (Gsc),which is expressed in Spemann’s or-ganizer and then becomes undetect-able as the embryo undergoes neuru-lation (Cho et al., 1991; Trindadeet al., 1999). Gsc is responsible for thedevelopment of dorsal structures (Choet al., 1991). These two genes, Vent-2
Dev
elop
men
tal D
ynam
ics
1500 HOOKER ET AL.

and Gsc, play antagonistic roles in theestablishment of the dorsoventralaxis. Lim1 expression peaks at gas-trulation in Spemann’s organizer, andhas the ability to directly activate Gscand maintain its expression in theprechordal plate (Mochizuki et al.,2000). All three are represented asPitx3-sensitive in the microarray;however, Gsc expresses too early tohave been monitored in our riboprobein situ hybridization although itshould be noted that Gsc possesses 14Pitx3 motifs in its 50-UTR.
Bix4 is a Brachyury-induciblehomeobox-containing gene and isthought to induce both mesoderm andendoderm formation depending onthe concentration of its encoded pro-tein (Tada et al., 1998). It expressesearlier than we monitored by in situhybridization at stages 19 or 27. Simi-larly, eFGF and RXRa are alsoexpressed early in development, wellbefore the stages that we assessed.eFGF is most similar to FGF-6 andFGF-4 in mammals, yet may repre-sent a novel FGF secreted factor that
has both mesoderm-inducing proper-ties and roles in anteroposterior pat-terning (Isaacs et al., 1994). RXRaencodes a retinoid X receptor that ispart of the nuclear receptor familythat mediates the effects of retinoicacid upon embryos. Expression ofRXRa begins as a maternal transcriptin the oocyte, and then is temporarilyabolished before gastrulation, leadingto a role for this receptor in early pat-terning of the embryo (Blumberget al., 1992). RA provides positionalinformation and helps to pattern the
Dev
elop
men
tal D
ynam
ics
TABLE 3. Parameters and Primer Sequences Used in RT-PCR Experiments
Gene Primers Size
Anneal
temp GenBank no. Reference
ODC Sense: 50 - GTC AAT GAT GGA GTG TAT G - 30 385bp 57�C XenBaseAntisense: 50 - TCC ATT CCG CTC TCC TGA - 30
Lim1 Sense: 50 - CCG ACA CAT AAG GGA GCA GC - 30 573bp 60�C X63889 HomemadeAntisense: 50 - CTG GTG GGT GTG ACA AAT GG - 30
Spr1 Sense: 50 - CCA GGT ACA AGT CCT ACT GA - 30 752bp 54�C AY062264 Ossipovaet al., 2002Antisense: 50 - GAG TGC CAC CTC AAA TGA GC - 30
Spr2 Sense: 50 - CAA ACT GTT GCC TCT CAT GAG - 30 380bp 54�C AY062263 Ossipovaet al., 2002Antisense: 50 - CAC TTA CAC CTC CGG CAG CGC - 30
Vent2 Sense: 50 - GCT TTC TCC TCG GTT GAA TG - 30 461bp 57�C X98454 HomemadeAntisense: 50 - TCT CCT TCA GGG GCT GTA GA - 30
Hes4 Sense: 50 - GCA CGA ACG AAG TCA CAC GA - 30 297bp 65�C AF139914 HomemadeAntisense: 50 - GCT GGG TTG GGA ATG AGG AAA G - 30
Hes7.1 Sense: 50- TGT AAT GTG CTC AAA TGG CG - 30 336bp 54�C BJ088128 HomemadeAntisense: 50 - TCC GTC AGC CCT ACA AAG AC – 3’
Obscnl Sense: 5’ – ACA GTA TGG TTC ACA GCC – 3’ 283bp 57�C BJ085487 HomemadeAntisense: 5’ – CAG TTG GCA CAT CAA TCC AG – 3’
Gsc Sense: 50 - ACA ACT GGA AGC ACT GGA - 30 279bp 52�C M81481 XenBaseAntisense: 50 - TCT TAT TCC AGA GGA ACC - 30
RXRa Sense: 50 - AAG ATA CTT GAG GCG GAG CA - 30 531bp 54�C L11446 HomemadeAntisense: 50 - TTC GGG GTA TTT CTG TTT GC - 30
L-Maf Sense: 50 - CTT GCT CCT CCT CAA TCT CTG G - 30 331bp 54�C AF202059 Ishibashi andYasuda, 2001Antisense: 50 -CCG ACA AAG GCG AAA GCT GGT G - 30
eFGF Sense: 50 - TTA CCG GAC GGA AGG ATA - 30 222bp 56�C X62594 Kroll LabAntisense: 50 - CCT CGA TTC GTA AGC GTT - 30
Bix4 Sense: 50 - CAG AAC AGG AGA TCA AAA GC - 30 414bp 54�C AF079562 HomemadeAntisense: 50 - CGG GTA GGT ACT AGA TGC TG – 3’
Hes7 Sense: 5’ – TGT TGG CTT GAA AGG TTT GT - 30 394bp 60�C BJ058661 HomemadeAntisense: 5’ – CTC AAA ATG TGT CAT AAT CCA - 30
Ripply2 Sense: 5’ – ATG GAG CCG AAT CAA CAG C - 30 352bp 57�C AB073615 HomemadeAntisense: 5’ – TGT CTT CCT CTT CAG AGT CA - 30
Crybb1 Sense: 5’ – CGT GGT GAG ATG TTT ATC CTG GAG - 30 394bp 60�C CD303346 HomemadeAntisense: 5’ – CCT TCT GGT GCC ATT GAT TGT CTC - 30
Pax6 Sense: 5’ – GCA ACC TGG CGA GCG ATA AGC - 30 448bp 56�C U77532 Zuberet al., 2003Antisense: 5’ –CCT GCC GTC TCT GGT TCC GTA GTT - 30
HoxA11 Sense: 5’ – AAT CCC TCC AAT GTC TAC CAC C - 30 363bp 56�C AJ319668 Slacket al., 2001Antisense: 5’ – CTG GTA TTT GGT ATA CGG GCA C – 3’
Rbp4l Sense: 5’ – AGA TGC AAT GCT CAG TCC T – 3’ 432bp 54�C CD362061 HomemadeAntisense: 5’ – GCG GGA GAA TAT AAT AGA ATA – 3’
GalectinIX Sense: 5’ – CCC GTG CCT GGT ATT TCA – 3’ 448bp 55�C BJ056659 HomemadeAntisense: 5’ – ACC TGG CTG GAG TGA ACA – 3’
Baz2b Sense: 5’ – AAG ATG ATG ATG AGG ACG A – 3’ 837bp 55�C BQ400337 HomemadeAntisense: 5’ – CCA TTT TAG CCT GCT GTT TC – 3’
Rdh16 Sense: 5’ – CTG CGA CTC TGG GTT TGG A – 3’ 750bp 57�C BG514525 HomemadeAntisense: 5’ – TCA TAG CCG GCA GAG TAG – 3’
MICROARRAY OF PITX3 TARGETS 1501

anteroposterior body axis, mostly bymediating posterior transformation ofthe embryo (Durston et al., 1989).
Conclusion
Microarray analysis is a useful toolto monitor the influence of a geneupon the entire transcriptome of anorganism. However, the generateddata set is quite elaborate anddeducing pertinent trends can be achallenging process. The informationrepresented in this study provides aglobal view of general developmentalprocesses in which Pitx3 may beinvolved. New genetic players havebeen identified as putative Pitx3 tar-gets in the already established eyeand brain developmental processes.In addition, based on genes identi-fied by the microarray, novel rolesfor Pitx3 can be inferred for regula-tion of early patterning events andthe development of the anterior–pos-terior body axis.
EXPERIMENTAL
PROCEDURES
Embryo Collection and
Manipulation
Staging, de-jellying, and culturing ofXenopus laevis embryos were con-ducted as previously described(Nieuwkoop and Faber, 1967; Dry-sdale and Elinson, 1991). Animalswere reared and used in accordancewith University, Provincial, and Fed-
eral regulations. Fluorescently labeledmorpholinos for either control or ex-perimental Pitx3 treatments wereinjected as previously described (Khos-rowshahian et al., 2005; Smoczeret al., In Press). Essentially, 4.6-nlinjections were made into the animalpole of embryos at the 1-cell stages forRNA collection and one- or two-cellstages for in situ hybridization.Injected embryos were cultured in 0.3� MBS and 2% Ficoll-400 (Sigma) at17�C for at least 1 hr to allow healingbefore being removed and allowed todevelop at 12�C in 0.1�MBS.
RNA Preparation and
Microarray Analysis
At staged intervals, embryos wereremoved for RNA isolation, lysed, andprocessed in Trizol as per manufac-turer’s instructions (Invitrogen). Wethen used DNAseI to remove genomicDNA, and ran the product over Qia-gen RNeasy columns for purification.RNA quality was assessed using theAgilent 2100 Bioanalyzer (AgilentTechnologies Inc., Palo Alto, CA) andthe RNA 6000 Nano kit (Caliper LifeSciences, Mountain View, CA).
All GeneChips were processed fromtwo biological replicates at the Lon-don Regional Genomics Centre(Robarts Research Institute, London,Ontario, Canada; http://www.lrgc.ca).Biotinylated complimentary RNA(cRNA) was prepared from 10 mg oftotal RNA as per the Affymetrix Gen-eChip Technical Analysis Manual
(Affymetrix, Santa Clara, CA). Dou-ble-stranded cDNA was synthesizedusing SuperScriptII (Invitrogen,Carlsbad, CA) and oligo(dT)24 pri-mers. Biotin-labeled cRNA was pre-pared by cDNA in vitro transcriptionusing the BioArray High-Yield RNATranscript Labeling kit (Enzo Bio-chem, New York) incorporating biotin-ylated UTP and CTP. A total of 15 mgof labeled cRNA was hybridized toXenopus laevis GeneChips for 16 hrat 45�C as described in the AffymetrixTechnical Analysis Manual (Affyme-trix). GeneChips were stained withStreptavidin-Phycoerythrin, followedby an antibody solution and a secondStreptavidin-Phycoerythrin solution,with all liquid handling performed bya GeneChip Fluidics Station 400.GeneChips were scanned with theAffymetrix GeneChip Scanner 3000(Affymetrix).Signal intensities for genes were
generated using GCOS1.2 (Affyme-
trix) using default values for the Sta-
tistical Expression algorithm parame-ters and a Target Signal of 150 for all
probe sets and a Normalization Value
of 1. Normalization was performed in
GeneSpring 7.2 (Agilent TechnologiesInc., Palo Alto, CA). Data were first
transformed (measurements less than
0.01 set to 0.01) and then normalized
per chip to the 50th percentile, andper gene to control samples for each
stage. We performed two biological
replicates and filtered the data based
upon fold change with a cut off Pvalue set at 0.05.
Dev
elop
men
tal D
ynam
ics
TABLE 4. Gene-Specific Information Regarding Restriction Enzymes and RNA Polymerases Used to Generate
Riboprobes for In Situ Hybridization Experiments
Gene Plasmid Restriction enzyme RNA Polymerase Reference/source
Lim1 pBSKSþ/Xlim-1 (pXH32) XhoI T7 Dawid, I. (NICHD)Vent2 pBS-XOM EcoRI T7 Ladher et al., 1996Spr1 pBSTSp1 T3/597(2-3)-2/XSPR-1 NotI T3 Ossipova et al., 2002Spr2 pCS2þNLSmyc/XSPR-2 XhoI T7 Ossipova, O. (unpublished)Hes4 XL409i23ex EcoRI T7 NIBBRbp4l XL060f11 BamHI T7 NIBBGalectinIX XL103j23 EcoRI T7 NIBBBaz2b 6989392 SalI T7 I.M.A.G.E.Rdh16 9897030 SalI T7 I.M.A.G.EHes7.1 XL091p04 EcoRI T7 NIBBObscnl XL106a24 EcoRI T7 NIBBHes7 XL060b05 EcoRI T7 NIBBRipply2 pCS/ledgerline ClaI T7 Asashima M. labCrybb1 HindIII T7 Henry J. labPax6 EcoRI SP6 Lupo G. labHoxA11 pGMT/HoxA11 NotI T7 Slack et al., 2001
1502 HOOKER ET AL.

RT-PCR
cDNA was made using Omniscriptreverse transcriptase (Qiagen) andOligo(dT)18 primers (Sigma) from 1ugtotal RNA for microarray confirma-tion and from 10uL mRNA furtherisolated (GenElute Direct mRNAMiniprep Kit; Sigma) for stage analy-sis of novel EST sequences. RT-PCRwas performed at various annealingtemperatures and cycle numbers,resulting in five time-points that wereultimately graphed. A cycle at the lin-ear phase of amplification wasselected for each gene and standar-dized against ODC. Fold change formicroarray confirmation was deter-mined by comparing gene amplifica-tion of control-morpholino-treatedsamples with Pitx3-morpholino-treated samples. Primers and param-eters are outline in Table 3.
Whole-Mount In Situ
Hybridization
In situ hybridizations were performedaccording to established protocols(Harland, 1991) using digoxygenin-la-beled riboprobes. We probed genesthat were either two times up- ordown-regulated as a consequence ofPitx3-morpholino perturbation,deemed by the microarray analysis.The probes used were generated fromplasmids that were either the gener-ous gifts of colleagues, the NIBB/NIG/NBRP Xenopus laevis EST project, orwere purchased from ATCC (seeTables). When a probe revealed a tem-poral and spatial expression patternthat overlapped with the known activ-ity of Pitx3, further in situ hybridiza-tions were conducted on specimensthat had been unilaterally injectedwith morpholino (control- or Pitx3-morpholino) at the two-cell stage:expression on the perturbed sidecould be compared with the contralat-eral control, and the trend predictedby the microarray thereby confirmed.Probes were prepared from vectors asoutlined in Table 4.
Identification of Novel Genes
Some of the most differentiallyexpressed but previously uncharac-terized EST sequences were explored.Their spatial expression pattern was
visualized by means of in situ hybrid-ization and the temporal expressionpattern was then investigated usingRT-PCR throughout embryonic stagesof development. Varied stages wereused to determine specific develop-mental events: unfertilized egg (E)and stage 5 for maternal transcripts,stage 10 (early gastrula), stage 12(neural anlage), stage 17 (onset ofsomitogenesis), stage 19 (neuraltube), stage 24 (tailbud), stage 27(lens differentiation), stage 31 (car-diac looping), stage 35 (blood supply;Nieuwkoop and Faber, 1967). Phylo-genic profiles and functional attrib-utes were deduced using Blastpsearches within GenBank and homo-log alignments using the Megalignprogram of DNASTAR Lasergene 7.2.
ACKNOWLEDGMENTSThe authors thank the numerous col-leagues who provided us with probes,and particularly to Dr. Kitayama andDr. Ueno, at the NIBB, without whosepatient, generous, and numerous giftsthis work could not have proceeded.Thanks also to David Carter of theLondon Regional Genomics Center,Robarts Research Institute, London,Ontario. M.J.C. was funded by theNatural Sciences and EngineeringResearch Council (NSERC) of Canada.F.K.S. was supported by an NSERCPGS, and M.W. was partially sup-ported by an Ontario GraduateScholarship.
REFERENCES
Altmann CR, Bell E, Sczyrba A, Pun J,Bekiranov S, Gaasterland T, BrivanlouAH. 2001. Microarray-based analysis ofearly development in Xenopus laevis.Dev Biol 236:64–75.
Amendt BA, Sutherland LB, Semina EV,Russo AF. 1998. The molecular basis ofRieger syndrome. Analysis of Pitx2homeodomain protein activities. J BiolChem 273:20066–20072.
Berman P, Gray P, Chen E, Keyser K, Ehr-lich D, Karten H, LaCorbiere M, Esch F,Schubert D. 1987. Sequence analysis,cellular localization, and expression of aneuroretina adhesion and cell survivalmolecule. Cell 51:135–142.
Blumberg B, Mangelsdorf DJ, Dyck JA,Bittner DA, Evans RM, De RobertisEM. 1992. Multiple retinoid-responsivereceptors in a single cell: families of ret-inoid ‘‘X’’ receptors in the Xenopus egg.Proc Natl Acad Sci U S A 89:2321–2325.
Buchtova M, Kuo WP, Nimmagadda S,Benson SL, Geetha-Loganathan P,Logan C, Au-Yeung T, Chiang E, Fu K,Richman JM. 2010. Whole genomemicroarray analysis of chicken embryofacial prominences. Dev Dyn 239:574–591.
Cazorla P, Smidt MP, O’Malley KL, Bur-bach JP. 2000. A response element forthe homeodomain transcription factorPtx3 in the tyrosine hydroxylase genepromoter. J Neurochem 74:1829–1837.
Chai X, Zhai Y, Napoli JL. 1996. Cloningof a rat cDNA encoding retinol dehydro-genase isozyme type III. Gene 169:219–222.
Chan T, Satow R, Kitagawa H, Kato S,Asashima M. 2006. Ledgerline, a novelxenopus laevis gene, regulates differen-tiation of presomitic mesoderm duringsomitogenesis. Zoolog Sci 23:689–697.
Chepelinsky AB. 2009. Structural func-tion of MIP/aquaporin 0 in the eye lens;genetic defects lead to congenital inher-ited cataracts. Handb Exp Pharmacol:265–297.
Cho KW, Blumberg B, Steinbeisser H, DeRobertis EM. 1991. Molecular nature ofSpemann’s organizer: the role of theXenopus homeobox gene goosecoid. Cell67:1111–1120.
Cooper DN, Barondes SH. 1999. Godmust love galectins; he made so manyof them. Glycobiology 9:979–984.
Coulon V, L’Honore A, Ouimette JF,Dumontier E, van den Munckhof P,Drouin J. 2007. A muscle-specific pro-moter directs Pitx3 gene expression inskeletal muscle cells. J Biol Chem 282:33192–33200.
Cui W, Tomarev SI, Piatigorsky J, Chepe-linsky AB, Duncan MK. 2004. Mafs,Prox1, and Pax6 can regulate chickenbetaB1-crystallin gene expression. JBiol Chem 279:11088–11095.
Drysdale TA, Elinson RP. 1991. Develop-ment of the Xenopus laevis hatchinggland and its relationship to surfaceectoderm patterning. Development 111:469–478.
Durston AJ, Timmermans JP, Hage WJ,Hendriks HF, de Vries NJ, HeideveldM, Nieuwkoop PD. 1989. Retinoic acidcauses an anteroposterior transforma-tion in the developing central nervoussystem. Nature 340:140–144.
Dutta S, Dietrich JE, Aspock G, BurdineRD, Schier A, Westerfield M, Varga ZM.2005. pitx3 defines an equivalence do-main for lens and anterior pituitaryplacode. Development 132:1579–1590.
Eisen JS, Smith JC. 2008. Controllingmorpholino experiments: don’t stopmaking antisense. Development 135:1735–1743.
Engstrom PG, Fredman D, Lenhard B.2008. Ancora: a web resource for explor-ing highly conserved noncoding ele-ments and their association withdevelopmental regulatory genes. Ge-nome Biol 9:R34.
Flower DR. 1996. The lipocalin proteinfamily: structure and function. BiochemJ 318(pt 1):1–14.
Dev
elop
men
tal D
ynam
ics
MICROARRAY OF PITX3 TARGETS 1503

Goodman DS. 1981. Retinoid-binding pro-teins in plasma and in cells. Ann N YAcad Sci 359:69–78.
Gray IC, Fallowfield J, Ford S, Nobile C,Volpi EV, Spurr NK. 1997. An integratedphysical and genetic map spanning chro-mosome band 10q24. Genomics 43:85–88.
Halder G, Callaerts P, Gehring WJ. 1995.Induction of ectopic eyes by targetedexpression of the eyeless gene in Dro-sophila. [see comments]. Science 267:1788–1792.
Harland RM. 1991. In situ hybridization:an improved whole-mount method forXenopus embryos. Methods Cell Biol36:685–695.
Hitachi K, Danno H, Tazumi S, Aihara Y,Uchiyama H, Okabayashi K, KondowA, Asashima M. 2009. The XenopusBowline/Ripply family proteins nega-tively regulate the transcriptional activ-ity of T-box transcription factors. Int JDev Biol 53:631–639.
Ho HY, Chang KH, Nichols J, Li M. 2009.Homeodomain protein Pitx3 maintainsthe mitotic activity of lens epithelialcells. Mech Dev 126:18–29.
Huang B, He W. 2010. Molecular charac-teristics of inherited congenital cata-racts. Eur J Med Genet 53:347–357.
Hwang DY, Hong S, Jeong JW, Choi S,Kim H, Kim J, Kim KS. 2009. Vesicularmonoamine transporter 2 and dopaminetransporter are molecular targets ofPitx3 in the ventral midbrain dopamineneurons. J Neurochem 111:1202–1212.
Isaacs HV, Tannahill D, Slack JM. 1992.Expression of a novel FGF in the Xeno-pus embryo. A new candidate inducingfactor for mesoderm formation andanteroposterior specification. Develop-ment 114:711–720.
Isaacs HV, Pownall ME, Slack JMW.1994. eFGF regulates Xbra expressionduring Xenopus gastrulation. EMBO J13:4469–4481.
Ishibashi S, Yasuda K. 2001. Distinctroles of maf genes during Xenopus lensdevelopment. Mech Dev 101:155–166.
Ito T, Levenstein ME, Fyodorov DV,Kutach AK, Kobayashi R, Kadonaga JT.1999. ACF consists of two subunits,Acf1 and ISWI, that function coopera-tively in the ATP-dependent catalysis ofchromatin assembly. Genes Dev 13:1529–1539.
Jacobs FM, Smits SM, Noorlander CW,von Oerthel L, van der Linden AJ, Bur-bach JP, Smidt MP. 2007. Retinoic acidcounteracts developmental defects inthe substantia nigra caused by Pitx3deficiency. Development 134:2673–2684.
Jen WC, Gawantka V, Pollet N, Niehrs C,Kintner C. 1999. Periodic repression ofNotch pathway genes governs the seg-mentation of Xenopus embryos. GenesDev 13:1486–1499.
Jones MH, Hamana N, Nezu J, ShimaneM. 2000a. A novel family of bromodo-main genes. Genomics 63:40–45.
Jones MH, Hamana N, Shimane M. 2000b.Identification and characterization ofBPTF, a novel bromodomain transcrip-tion factor. Genomics 63:35–39.
Kameda Y, Saitoh T, Fujimura T. 2011.Hes1 regulates the number and ante-rior-posterior patterning of mesence-phalic dopaminergic neurons at themid/hindbrain boundary (isthmus). DevBiol 358:91–101.
Kawamura A, Koshida S, Takada S. 2008.Activator-to-repressor conversion of T-box transcription factors by the Ripplyfamily of Groucho/TLE-associated medi-ators. Mol Cell Biol 28:3236–3244.
Khosrowshahian F, Wolanski M, ChangWY, Fujiki K, Jacobs L, Crawford MJ.2005. Lens and retina formation requireexpression of Pitx3 in Xenopus pre-lensectoderm. Dev Dyn 234:577–589.
Kikuta H, Laplante M, Navratilova P,Komisarczuk AZ, Engstrom PG, Fred-man D, Akalin A, Caccamo M, Sealy I,Howe K, Ghislain J, Pezeron G, Mour-rain P, Ellingsen S, Oates AC, Thisse C,Thisse B, Foucher I, Adolf B, Geling A,Lenhard B, Becker TS. 2007. Genomicregulatory blocks encompass multipleneighboring genes and maintain con-served synteny in vertebrates. GenomeRes 17:545–555.
Ladher R, Mohun TJ, Smith JC, SnapeAM. 1996. Xom: a Xenopus homeoboxgene that mediates the early effects ofBMP-4. Development 122:2385–2394.
Lamonerie T, Trembley J, Lanctot C,Therrien M, Gautier Y, Drouin J. 1996.Ptx1, a bicoid-related homeo box tran-scription factor involved in the tran-scription of the pro-opiomelanocortingene. Genes Dev 10:1284–1295.
Landis SC, Siegel RE, Schwab M. 1988.Evidence for neurotransmitter plasticityin vivo. II. Immunocytochemical studiesof rat sweat gland innervation duringdevelopment. Dev Biol 126:129–140.
Lebel M, Gauthier Y, Moreau A, Drouin J.2001. Pitx3 activates mouse tyrosinehydroxylase promoter via a high-affinitybinding site. J Neurochem 77:558–567.
Lombardo A, Slack JM. 2001. AbdominalB-type Hox gene expression in Xenopuslaevis. Mech Dev 106:191–195.
Marchand R, Poirier LJ. 1983. Isthmic or-igin of neurons of the rat substantianigra. Neuroscience 9:373–381.
Messmer K, Remington MP, Skidmore F,Fishman PS. 2007. Induction of tyro-sine hydroxylase expression by thetranscription factor Pitx3. Int J DevNeurosci 25:29–37.
Mochizuki T, Karavanov AA, Curtiss PE,Ault KT, Sugimoto N, Watabe T, Shio-kawa K, Jamrich M, Cho KW, DawidIB, Taira M. 2000. Xlim-1 and LIM do-main binding protein 1 cooperate withvarious transcription factors in the reg-ulation of the goosecoid promoter. DevBiol 224:470–485.
Munster D. 2005. Pitx3 und seine Rolle inder Augen- und Gehirnentwicklung. In:GSF-Forschungszentrum fur Umwelt undGesundheit Institut fur Entwicklungsge-netik Neuherberg: Technischen Uni-versit€at Munchen. p 115.
Murato Y, Nagatomo K, Yamaguti M,Hashimoto C. 2007. Two alloalleles ofXenopus laevis hairy2 gene–evolution
of duplicated gene function from a de-velopmental perspective. Dev GenesEvol 217:665–673.
Nagy L, Saydak M, Shipley N, Lu S, Basi-lion JP, Yan ZH, Syka P, ChandraratnaRA, Stein JP, Heyman RA, Davies PJ.1996. Identification and characteriza-tion of a versatile retinoid response ele-ment (retinoic acid receptor responseelement-retinoid X receptor responseelement) in the mouse tissue transglu-taminase gene promoter. J Biol Chem271:4355–4365.
Nakamura H, Watanabe Y. 2005. Isthmusorganizer and regionalization of themesencephalon and metencephalon. IntJ Dev Biol 49:231–235.
Napoli JL, Posch KP, Fiorella PD, Boer-man MHEM. 1991. Physiological occur-rence, biosynthesis and metabolism ofretinoic acid: evidence for roles of cellu-lar retinol-binding protein (CRBP) andcellular retinoic acid-binding protein(CRABP) in the pathway of retinoicacid homeostasis. Biomed Pharmac-other 45:131–143.
Nieuwkoop PD, Faber J. 1967. NormalTable of Xenopus laevis (Daudin). Am-sterdam: North Holland Press.
Ossipova O, Stick R, Pieler T. 2002.XSPR-1 and XSPR-2, novel Sp1 relatedzinc finger containing genes, aredynamically expressed during Xenopusembryogenesis. Mech Dev 115:117–122.
Pares X, Farres J, Kedishvili N, Duester G.2008. Medium- and short-chain dehydro-genase/reductase gene and protein fami-lies: medium-chain and short-chaindehydrogenases/reductases in retinoid me-tabolism. Cell Mol Life Sci 65:3936–3949.
Pommereit D, Pieler T, Hollemann T.2001. Xpitx3: a member of the Rieg/Pitxgene family expressed during pituitaryand lens formation in Xenopus laevis.Mech Dev 102:255–257.
Reza HM, Ogino H, Yasuda K. 2002. L-Maf, a downstream target of Pax6, isessential for chick lens development.Mech Dev 116:61–73.
Rieger DK, Reichenberger E, McLean W,Sidow A, Olsen BR. 2001. A double-de-letion mutation in the pitx3 gene causesarrested lens development in aphakiamice. Genomics 72:61–72.
Sakazume S, Sorokina E, Iwamoto Y,Semina EV. 2007. Functional analysisof human mutations in homeodomaintranscription factor PITX3. BMC MolBiol 8:84.
Semina EV, Murray JC, Reiter R, HrstkaRF, Graw J. 2000. Deletion in the pro-moter region and altered expression ofPitx3 homeobox gene in aphakia mice.Hum Mol Genet 9:1575–1585.
Shawlot W, Behringer RR. 1995. Require-ment for Lim1 in head-organizer function.[see comments]. Nature 374:425–430.
Shi X, Bosenko DV, Zinkevich NS, FoleyS, Hyde DR, Semina EV, Vihtelic TS.2005. Zebrafish pitx3 is necessary fornormal lens and retinal development.Mech Dev 122:513–527.
Shinga J, Itoh M, Shiokawa K, Taira S,Taira M. 2001. Early patterning of the
Dev
elop
men
tal D
ynam
ics
1504 HOOKER ET AL.

prospectivemidbrain-hindbrain boundaryby the HES-related gene XHR1 in Xeno-pus embryos. Mech Dev 109:225–239.
Shoji H, Nishi N, Hirashima M, NakamuraT. 2003. Characterization of the Xenopusgalectin family. Three structurally differ-ent types as in mammals and regulatedexpression during embryogenesis. J BiolChem 278:12285–12293.
Simon A, Hellman U, Wernstedt C, Eriks-son U. 1995. The retinal pigment epi-thelial-specific 11-cis retinoldehydrogenase belongs to the family ofshort chain alcohol dehydrogenases. JBiol Chem 270:1107–1112.
Simon A, Lagercrantz J, Bajalica-Lager-crantz S, Eriksson U. 1996. Primarystructure of human 11-cis retinol dehy-drogenase and organization and chromo-somal localization of the correspondinggene. Genomics 36:424–430.
Smoczer C, Hooker L, Brode S, WolanskiM, KhosrowShahian F, Crawford M. (InPress). The Xenopus homeobox genePitx3 impinges upon somitogenesis andlaterality. Biochem Cell Biol.
Sorokina EA, Muheisen S, Mlodik N,Semina EV. 2011. MIP/Aquaporin 0 rep-
resents a direct transcriptional targetof PITX3 in the developing lens. PLoSOne 6:e21122.
Tada M, Casey ES, Fairclough L, SmithJC. 1998. Bix1, a direct target of Xeno-pus T-box genes, causes formation ofventral mesoderm and endoderm. De-velopment 125:3997–4006.
Takada H, Hattori D, Kitayama A, UenoN, Taira M. 2005. Identification of tar-get genes for the Xenopus Hes-relatedprotein XHR1, a prepattern factor spec-ifying the midbrain-hindbrain bound-ary. Dev Biol 283:253–267.
Tanaka M, Murayama D, Nagashima M,Higashi T, Mawatari K, MatsukawaT, Kato S. 2007. Purpurin expressionin the zebrafish retina during earlydevelopment and after optic nervelesion in adults. Brain Res 1153:34–42.
Trindade M, Tada M, Smith JC. 1999.DNA-binding specificity and embryolog-ical function of Xom (Xvent-2). Dev Biol216:442–456.
Tsuji S, Cho KW, Hashimoto C. 2003.Expression pattern of a basic helix-loop-helix transcription factor Xhairy2b dur-
ing Xenopus laevis development. DevGenes Evol 213:407–411.
van den Munckhof P, Luk KC, Ste-MarieL, Montgomery J, Blanchet PJ, Sadi-kot AF, Drouin J. 2003. Pitx3 isrequired for motor activity and forsurvival of a subset of midbrain dopa-minergic neurons. Development 130:2535–2542.
Varnum DS, Stevens LC. 1968. Aphakia,a new mutation in the mouse. J Hered59:147–150.
Wald G. 1968. The molecular basis of vis-ual excitation. Nature 219:800–807.
Wolda SL, Moon RT. 1992. Cloning anddevelopmental expression in Xenopuslaevis of seven additional membersof the Wnt family. Oncogene 7:1941–1947.
Wurst W, Auerbach AB, Joyner AL. 1994.Multiple developmental defects inEngrailed-1 mutant mice: an early mid-hindbrain deletion and patterningdefects in forlimbs and sternum. Devel-opement 120:2065–2075.
Zeng L, Zhou MM. 2002. Bromodomain:an acetyl-lysine binding domain. FEBSLett 513:124–128.
Dev
elop
men
tal D
ynam
ics
MICROARRAY OF PITX3 TARGETS 1505