microarray analysis of immune challenged hemocytes
TRANSCRIPT

www.elsevier.com/locate/yexcr
Experimental Cell Researc
Microarray analysis of immune challenged Drosophila hemocytes
Karin C. Johansson, Christoph Metzendorf, Kenneth Sfderh7ll*
Department of Comparative Physiology, Evolutionary Biology Center, Uppsala University, Norbyvagen 18A, SE-752 36 Uppsala, Sweden
Received 27 August 2004, revised version received 17 December 2004
Available online 2 February 2005
Abstract
Insect hemocytes play multiple roles in immunity and carry out cellular responses like phagocytosis, encapsulation and melanization as
well as producing humoral effector proteins in the first line of defense after injury and invasion of microorganisms. In this work, we used the
Drosophila melanogaster hemocyte-like cell line mbn-2 and Affymetrix Drosophila GeneChips to investigate the transcriptome of a single
type of immune competent tissue exposed to Gram-negative cell wall components (crude LPS) or high dose infection with live Escherichia
coli. We found that gene expression profiles of both treatments overlap but show important differences in expression levels of several genes
involved in immunity. In addition, cell morphology during infection was monitored and revealed distinct alterations in cell shape and
adhesion. Presence of large numbers of bacteria also increased the number of cells taking on crystal cell fate. Taken together, our results
indicate that hemocytes sense and respond differently to purified bacterial surface molecules and infection with live and actively growing
bacteria both at the level of gene expression and in cell behavior.
D 2005 Elsevier Inc. All rights reserved.
Keywords: Drosophila; Microarray analysis; Hemocytes; Crude LPS; E. coli
Introduction
Microorganisms that breach protective epithelial barriers
and invade circulated tissues trigger in all animals an array
of host immune reactions that work in synergy to fight
infection. Insects lack the antibody-based adaptive immun-
ity of mammals and rely solely on the capacity of their
innate immunity for survival in environments that are often
thriving with microbes. In the fruit fly Drosophila mela-
nogaster, the innate immune system is thought to recognize
non-self through a set of germ line-encoded pattern
recognition receptors (PRRs), binding to and raising the
alarm against evolutionary conserved microbial epitopes
like peptidoglycans (PGN), mannans and h-1,3 glucans of
bacterial cell walls and fungi, respectively [1]. The defenses
are broadly divided into humoral and cellular responses. The
humoral branch involves the rapid synthesis of antimicrobial
peptides (AMPs), secreted into the hemolymph mainly from
the fat body and to a lesser extent from circulating blood
0014-4827/$ - see front matter D 2005 Elsevier Inc. All rights reserved.
doi:10.1016/j.yexcr.2004.12.018
* Corresponding author. Fax: +46 18 4716425.
E-mail address: [email protected] (K. Sfderh7ll).
cells (hemocytes) and surface epithelia [2,3]. In addition,
proteolytic enzymes are released and activated in the
hemolymph inducing localized melanization around wounds
and foreign objects [4]. Cellular defense mechanisms,
executed by circulating hemocytes of different types,
include phagocytosis by plasmatocytes, capsule formation
around larger foreign objects such as parasitoid wasp eggs
by large adherent lamellocytes and immobilization of large
numbers of bacteria by the binding of multiple hemocytes in
a process referred to as nodulation. Capsules and nodules
are subsequently melanized by the prophenoloxidase-acti-
vating system (proPO) [5–9]. When the animal is wounded,
a reaction is induced that involves aggregation of hemocytes
into clots sealing the wound to limit bleeding and the risk of
microbial entry into the hemocoel [10]. Recognition of
different form of PGN activates either of two NF-nB-likeintracellular signaling pathways, Toll and immune defi-
ciency (IMD). The IMD pathway responds mainly to Gram-
negative infection by recognition of diaminopimelic acid
(DAP)-type PGN while Toll responds to Gram-positive
lysine(Lys)-type PGN but also to fungal infection. Activa-
tion of IMD leads to expression of AMPs and PRRs like
h 305 (2005) 145–155

K.C. Johansson et al. / Experimental Cell Research 305 (2005) 145–155146
peptidoglycan recognition proteins (PGRPs). The Toll
pathway controls transcription of the antifungal peptide
drosomycin, PPRs and several serine proteases involved in
melanization. In addition, Toll signaling has been proposed
a role in the cellular defense [11–16]. Together, Toll and
IMD regulate the majority of Drosophila immune genes but
a recent genomic-scale expression analysis of LPS chal-
lenged Drosophila SL2 cells has also demonstrated con-
tributing roles for JNK- and JAK/STAT-signaling in
immunity [17]. To investigate the response to live bacterial
infection in a single type of immune responsive tissue, we
took advantage of a Drosophila hemocyte-like cell line,
malignant blood neoplasm-2 (mbn-2). Mbn-2 was originally
established from circulating larval hemocytes [18] and has
with retained ability to express AMPs and phagocytose
microorganisms proven a valuable in vitro model system for
studies of the immune response in Drosophila [19]. In an
earlier study using mbn-2 cells, we showed that co-infection
with crude LPS and high doses of live enteric bacteria
muted induced transcription of the antibacterial peptide
Diptericin [20]. This effect was attributed to a potentially
novel feature of bacterial pathogenesis, distinct from the
well described type III secretion-dependent injection of
effector proteins that can interrupt or hijack normal host cell
processes like signal transduction, actin polymerization, and
vesicular trafficking to benefit the pathogen [21]. In this
aspect, we decided to take a whole genome approach to find
out whether other genes were affected in the same way at
the level of host gene transcription.
Materials and methods
Cells and bacterial strains
TheDrosophila mbn-2 cell line was maintained at 258C in
Schneider’s Drosophila medium (PAN Biotech GmbH,
Aidenbach, Germany) supplemented with 10% FBS and 2
mM l-glutamine. Cells were routinely passaged every third
day to a density of ~1 � 106 cells/ml. Enteropathogenic
Escherichia coli 2348/69 was obtained from J. Kaper
(University of Maryland School of Medicine, Baltimore,
MD). Bacteria for infection was diluted 1:50 in fresh LB from
an over night culture and grown to mid log phase (OD600nm =
0.5) at 378C with shaking. After centrifugation the bacterial
pellet was resuspended in Schneider’sDrosophila medium to
1 � 1011 CFU/ml. Heat-inactivated bacteria was prepared in
the sameway after 15min incubation in a 658Cwater bath. 48
h prior to infection, mbn-2 cells were treated with 1 AMEcdysone (Sigma-Aldrich, St. Louis, MO) to promote
immune competence as described elsewhere [22].
Microarray analysis
Approximately 5 � 106 mbn-2 cells were challenged
with crude LPS (O127:B8) 10 Ag/ml (Sigma) or a
combination of crude LPS and 1 � 109 CFU of live E.
coli, corresponding to approximately 200 bacteria per
Drosophila cell. Cells were harvested after 6 h (258C) andtotal RNA was isolated using TRIzol (Life Technologies,
Bethesda, MD) followed by RNeasy clean-up (Qiagen,
Valencia, CA). When treated with crude LPS only, cultures
were spiked with 5.5 � 109 CFU heat-killed bacteria just
before harvest to compensate for the higher ratio of
Drosophila versus E. coli-specific RNA in these samples.
Synthesis of cDNA, biotin-labeled cRNA and hybridization
to Affymetrix Drosophila genome GeneChipsk (Affyme-
trix, Santa Clara, CA) was performed by the Affymetrix
core facility (NOVUM, Karolinska Institutet, Huddinge,
Sweden) according to standard protocols. Each infection
experiment was made in two replicates, processed in parallel
and hybridized to two individual GeneChips. One single
GeneChip was hybridized with RNA from naive cells as a
control of basal level gene expression. GeneChips were
scanned with an Affymetrix GeneArrayR scanner. Data
were analyzed using Affymetrix Microarray Suite 5.0
software. To select for robust changes in transcript
abundance, transcripts with detection P values of P b
0.0025 for increased, and P N 0.9975 (Wilcoxon’s Signed
Rank test) for decreased, in both naive control and
challenged cells were included in data analyses. Cluster
analysis (SOM) was used to group genes with similar
expression over the experiment as a whole, focusing on the
bshapeQ of the expression pattern rather than absolute levels.
Replicate chips with RNA from cells treated with crude LPS
(control) and cells infected with live bacteria were pair-wise
compared (n = 4) and hierarchically clustered (GeneWeaver)
to identify unique and common expression changes.
Viability and gross cell morphology of cells was assessed
by phase contrast microscopy and Tryphan blue exclusion.
Northern blot
To confirm microarray predictions, mbn-2 cells were
challenged as described above. RNA gel electrophoresis and
transfer to Hybond-N membranes (Amersham Pharmacia
Biotech, Little Chalfont, UK) was performed according to
standard procedures. Hybridizations with radiolabeled
probes that were either inserts from cDNA clones [23], or
PCR products verified by DNA sequence were performed
under high stringency conditions (6� SSC, 658C). Filterexposure time to IP-screens varied between 16 and 48 h for
the individual probes depending on the signal strength of the
target gene.
Scanning electron microscopy (SEM)
Mbn-2 cells were prepared for electron microscopy
essentially by following the infection protocol above, except
that sterile round glass cover slips were placed in each cell
culture well. Challenge with live or heat-inactivated bacteria
at a dose of approximately 20 bacteria per Drosophila cell

Fig. 1. Genome wide response of Drosophila mbn-2 cells to live bacterial
infection and treatment with crude LPS. Distribution of up- or down-
regulated transcripts in response to either or both treatments.
K.C. Johansson et al. / Experimental Cell Research 305 (2005) 145–155 147
was carried out for 1, 2, 3, 4, 5, and 6 h (258C). Cells werefixed in 2.5% glutaraldehyde (48C) and then washed 3 � 15
min in PBS (pH 7.0), post-fixed in 1% osmium tetroxide for
2 h and washed again 3� with PBS. Dehydration was
carried out in an ascending ethanol series to 95% overnight.
Two further dehydration steps in 99% ethanol for 20 min
were carried out before drying in a critical point dryer and
gold coating. Images were captured with a Phillips XL30
electron microscope.
Immunohistochemistry
Mbn-2 cells were seeded onto SuperfrostR Plus micro-
scope slides and challenged with live E. coli and/or crude
LPS as described in the microarray section above. Cell
monolayers were fixed in 3.7% paraformaldehyde/PBS,
washed three times with PBS, blocked with 5% normal
rabbit serum (Dako Cytomation, Glostrup, Denmark)/PBS
and incubated with IgG mouse mAb HC12F6 against
Hyalophora cecropia hemocytes over night at 48C. mAb
HC12F6 recognizes Drosophila crystal cells with very high
specificity both in hemocyte monolayers and in the
hematopoietic organ and co-stains the same cells as a
polyclonal anti-PO Ab raised against Bombyx mori PO.
MabHC12F6 recognizes a band with the right molecular
weight of PO on western blots of Drosophila hemolymph
and can be competed with a B. mori anti-PO antibody that
also binds to Drosophila PO (T. Trenczek, personal
communication). Slides were washed 3� with PBS and
incubated with a FITC-conjugated rabbit anti-mouse Ab
(Dako) (1:1000) for 1 h at 258C and washed again several
times before mounting in Vectashield (Vector Laboratories,
Burlingame, CA). Confocal images were captured with a
Leica TCS SP.
Results and discussion
Microarray gene expression analysis
Affymetrix Drosophila GeneChips were probed with
RNA from mbn-2 cells challenged with live E. coli and
crude LPS or crude LPS only (control). We then compared
the transcriptional responses evoked by bacterial cell wall
products (crude LPS preparations are always contaminated
by PGN) to those matching a live actively proliferating
Gram-negative bacterial infection. Of 13197 unique genes
and ESTs represented on the chip, 4970 (35.6%) were
detected as present in naive mbn-2 cells. Control crude LPS
treatment increased this number to 5188 (37.2%), whereas
infection with bacteria decreased the number of present calls
to 4308 (30.8%). Of the 926 transcripts that were diffe-
rentially expressed in response to either type of challenge,
387 were increased and 539 decreased (Fig. 1). To bring
primary structure to the data and uncover fundamental gene
expression patterns, we used a SOM method for sorting
genes with common responses across the experiment in
clusters. This enabled us to identify several clusters of clear
relevance for immunity, which we grouped in three main
classes: (1) genes that were induced by crude LPS and
further up-regulated by live bacterial challenge; (2) genes
that were only up-regulated by live bacterial challenge; and
(3) genes that were up-regulated by crude LPS but down-
regulated by challenge with live E. coli (Fig. 2).
Class 1—Inducible genes with enhanced expression by live
E. coli challenge
In class 1 (Fig. 2A), we found genes with defined roles
both in humoral and cellular immunity like Attacin-A and
actin binding/cytoskeletal proteins (spire, inscuteable). Up-
regulation of Pvf2, one of three putative ligands to the
Drosophila homologue of the mammalian PDGF/VEGF
receptor suggested active cell proliferation. Overexpression
of Pvf2 in larvae has been shown to induce dramatic
proliferation of circulating hemocytes [24], proposing that
encounter with a massive bacterial infection may trigger
maturation or differentiation of hemocytes from precursor
cells in the mbn-2 system as a way of boosting cell mediated
defenses. Up-regulation of an extracellular sulphatase
(Sulphated) required for developmental patterning of the
embryo as a regulator of Wingless signaling [25] was
unexpected but suggests that it may be playing a yet
undefined role in immunity.
Class 2—Genes induced only by challenge with E. coli
For induction of some genes actual presence of whole
bacteria seemed critical (Fig. 2B). Among them were
several proteins involved in tissue remodeling and stress
signaling including the MAP kinase phosphatase Puckered,
a transcriptional target and negative regulator of JNK
signaling [26]. Up-regulation of several genes in this class
is consistent with results from a microarray study of the

Fig. 2. Gene expression patterns in immune challenged mbn-2 cells. SOM analysis clustered genes with similar expression profiles for different types of
challenge; naRve (N), crude LPS (cL) and bacterial challenge with E. coli (E.c). Clusters of clear relevance for immunity were grouped together: (A) Class 1—
Genes induced by crude LPS and further up-regulated by live bacterial infection. (B) Class 2—Genes induced only by bacterial infection. (C) Class 3—Genes
induced by crude LPS and down-regulated by bacterial infection. Genes within each cluster are listed in the tables. Graphs show the average expression level
for genes in the cluster (–o–), in-cluster variation is visualized by the surrounding lines.
K.C. Johansson et al. / Experimental Cell Research 305 (2005) 145–155148
LPS response in SL2 cells by Boutros et al. [17], such as
Ets21C, a member of the ETS (erythroblast transformation
specific)—family of transcription factors and the matrix
metalloproteinase 1 (Mmp 1). However, in contrast to Ref.
[17], Mmp 1 was only induced in the presence of bacteria
and not by treatment with crude LPS alone. Matrix
metalloproteinases have demonstrated roles in tissue
remodeling in Drosophila [27] and have in mammals
been implicated in wound healing responses [28]. Detailed
functional characterization of Ets21C in Drosophila is
currently not available but its human counterpart ERG has
been implicated in development of the central nervous
system [29]. Many genes associated with cytoskeletal
function showed unique up-regulation in the presence
of whole bacteria. One, (CG13503) contains a WH2-
domain—an actin interacting domain found in WASP/
SCAR proteins. D-SCAR and D-WASP are required for
efficient phagocytosis in Drosophila hemocytes [30].
Several proteins with demonstrated or suggested functions
in the development of the nervous system also show this
profile, like the plasma membrane axon guidance receptor
Leak, the RAB GTPase Chrowded and E(spl)m, a member
of the Brd-family proteins that when overexpressed
resemble a Notch loss-of-function phenotype in the eye

K.C. Johansson et al. / Experimental Cell Research 305 (2005) 145–155 149
[31]. Indication of active Toll signaling was given by up-
regulation of the NF-nB-like transcription factor Relish.
Relish is involved in transcription of most of the AMPs
and several of the PGRPs, targets of the Imd pathway,
while the transcriptional control of relish is partly depend-
ent on the Toll pathway during infection [15]. Since active
Toll signaling also controls hemocyte division and differ-
entiation in larvae [16], this is in support of the hypothesis
that large doses of bacteria can elicit cell differentiation in
mbn-2 cells.
Class 3—Genes down-regulated by challenge with E. coli
Somewhat contradictory in a situation with a galloping
bacterial infection and up-regulation of Relish, but in
accordance with our previous results [20] many of the
AMPs; Attacin-A and -C, Diptericin, Drosomycin, and
Drosocin showed induction by crude LPS to varying
degrees but were down-regulated by bacterial challenge
(Fig. 2C). Other immune genes, like the short soluble
PGRPs, -SA, -SB1, and -SD [32], displayed the same
profile. The two serine-type endopeptidases (CG30086,
CG4259) have to our knowledge not previously been
identified as immune responsive. Decrease of cytochrome
P450 at the level of gene expression seems to be a
conserved signature of immune stimulation, since it is
observed in insects [15] as well as in association with
cytokine action in vertebrates [33]. A transcription factor,
Bigmax, involved in regulating autophagic cell death during
normal metazoan development was also found in this class
[34].
Active E. coli infection versus crude LPS treatment evoke
different responses in mbn-2 cells
To account for transcripts missing or of very low
abundance in naive cells, we let the transcriptional profile
mbn-2 cells challenged with crude LPS represent a baseline
to which we compared the profile of bacteria infected cells.
A total of 517 differentially regulated gene transcripts were
detected, as defined by the criteria in the Affymetrix
software (difference call), in four pair-wise comparisons
between replica chips of crude LPS and E. coli challenge.
Of these, 210 were up-regulated and 274 down-regulated.
The relative mRNA expression in E. coli challenged cells
versus crude LPS control varied from 1.3- to 11-fold for up-
regulated genes with the majority of genes (71%) showing
relatively little change with ratios in the range between 1.3-
and 2-fold. For down-regulated genes, 84% had ratios
between �1.2- and �2-fold, 45 genes (18%) had decreased
expression level between 2- and 14-fold. Genes that
changed 2-fold or more are listed in Table 1. A complete
list of genes induced by control crude LPS treatment and
differentially regulated genes by infection with live E. coli
can be found in Tables 2 and 3 in the Supplementary
material.
Up-regulated genes
Strongest up-regulation by live bacterial infection (z4-
fold) was recorded for two transcripts encoding actin
binding proteins (CG32030, CG13505), a matrix metal-
loproteinase 1 (Mmp 1), an extensin-like protein, a lipase
(CG4267), the JNK-signaling protein Puckered and two
transcription factors (Ets21C, CG11798). Many up-regu-
lated genes could be assigned to functional groups
reflecting different aspects of cellular immunity. Adhe-
sion between cells, cells and microbes and cells and the
extracellular matrix, is an essential part of invertebrate
immunity which in the array data is recognized by
several transcripts for extracellular matrix proteins (Glu-
tactin, Trol) and integrins (Scab, aPS4) in addition to
many actin and Ca2+-binding proteins. Arthropods have
open circulatory systems, which makes blood coagulation
one of the most important defense reactions that prevent
the animal from bleeding out when injured and aid in-
limiting the spread of microbes in the hemocoel.
Utilization of transglutaminases in cross-linking of clot-
table proteins and hemocytes seem to be phylogenetically
conserved in all clotting systems studied to date [10].
Expression of the only transglutaminase gene found in
the Drosophila genome (CG7356) was unchanged by
either type of immune challenge (data not shown).
However, an extracellular lipid transport protein
(CG15828) with similarity to Vitellogenin was found to
be up-regulated. Vitellogenin-like proteins have docu-
mented roles in clotting and melanization in other
arthropod species [35,36]. In addition, a number of other
genes with motifs frequently shared between clotting
proteins in different species like the immunoglobulin-
fold, (EGF)-like, sushi-, C-type lectin-, clip-, and
domains similar to von Willebrand Factor (vWF) [37]
were up-regulated (Table 1 and Supplementary material).
We also found up-regulation of Annexin IX, a Ca2+-
regulated protein that interacts with phospholipids of the
cell membrane and the actin cytoskeleton. Reports of
Annexin IX up-regulation in response to immune challenge
has come from two other microarray analyses of bacteria
challenged adult flies [38] and LPS challenged Drosophila
SL2 cells [17]. Furthermore, it came up in a recent
proteomic screening of LPS induced proteins in mbn-2
cells [39]. Together, these results confirm Annexin IX as an
immune responsive gene whose function should be further
studied. Several genes with defined or putative function in
transcriptional regulation like, Psc, Ets21C, Jra, CG11798,
CG6051, were also up-regulated. The increase in expression
of Psc (Posterior sex combs), a negative regulator of the
polycomb group of homeotic genes (PcG), is of special
interest. PcG genes regulate Hox gene expression in
vertebrates and invertebrates during development and the
Psc homologues Bmi-1 and Mel 18 also participate in
hematopoiesis control in the mouse [40]. The first report
that homeobox genes may play a role in macrophage
responses to LPS came from a recent study of a murine cell
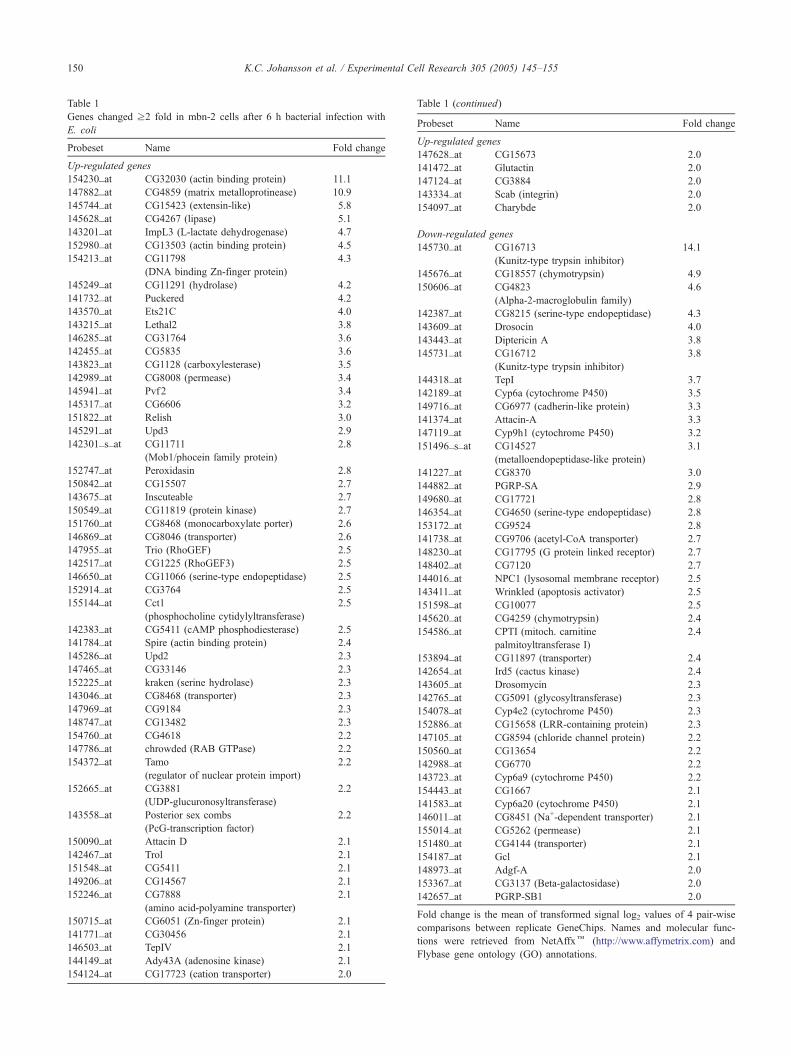
Table 1
Genes changed z2 fold in mbn-2 cells after 6 h bacterial infection with
E. coli
Probeset Name Fold change
Up-regulated genes
154230_at CG32030 (actin binding protein) 11.1
147882_at CG4859 (matrix metalloprotinease) 10.9
145744_at CG15423 (extensin-like) 5.8
145628_at CG4267 (lipase) 5.1
143201_at ImpL3 (L-lactate dehydrogenase) 4.7
152980_at CG13503 (actin binding protein) 4.5
154213_at CG11798
(DNA binding Zn-finger protein)
4.3
145249_at CG11291 (hydrolase) 4.2
141732_at Puckered 4.2
143570_at Ets21C 4.0
143215_at Lethal2 3.8
146285_at CG31764 3.6
142455_at CG5835 3.6
143823_at CG1128 (carboxylesterase) 3.5
142989_at CG8008 (permease) 3.4
145941_at Pvf2 3.4
145317_at CG6606 3.2
151822_at Relish 3.0
145291_at Upd3 2.9
142301_s_at CG11711
(Mob1/phocein family protein)
2.8
152747_at Peroxidasin 2.8
150842_at CG15507 2.7
143675_at Inscuteable 2.7
150549_at CG11819 (protein kinase) 2.7
151760_at CG8468 (monocarboxylate porter) 2.6
146869_at CG8046 (transporter) 2.6
147955_at Trio (RhoGEF) 2.5
142517_at CG1225 (RhoGEF3) 2.5
146650_at CG11066 (serine-type endopeptidase) 2.5
152914_at CG3764 2.5
155144_at Cct1
(phosphocholine cytidylyltransferase)
2.5
142383_at CG5411 (cAMP phosphodiesterase) 2.5
141784_at Spire (actin binding protein) 2.4
145286_at Upd2 2.3
147465_at CG33146 2.3
152225_at kraken (serine hydrolase) 2.3
143046_at CG8468 (transporter) 2.3
147969_at CG9184 2.3
148747_at CG13482 2.3
154760_at CG4618 2.2
147786_at chrowded (RAB GTPase) 2.2
154372_at Tamo
(regulator of nuclear protein import)
2.2
152665_at CG3881
(UDP-glucuronosyltransferase)
2.2
143558_at Posterior sex combs
(PcG-transcription factor)
2.2
150090_at Attacin D 2.1
142467_at Trol 2.1
151548_at CG5411 2.1
149206_at CG14567 2.1
152246_at CG7888
(amino acid-polyamine transporter)
2.1
150715_at CG6051 (Zn-finger protein) 2.1
141771_at CG30456 2.1
146503_at TepIV 2.1
144149_at Ady43A (adenosine kinase) 2.1
154124_at CG17723 (cation transporter) 2.0
Table 1 (continued)
Probeset Name Fold change
Up-regulated genes
147628_at CG15673 2.0
141472_at Glutactin 2.0
147124_at CG3884 2.0
143334_at Scab (integrin) 2.0
154097_at Charybde 2.0
Down-regulated genes
145730_at CG16713
(Kunitz-type trypsin inhibitor)
14.1
145676_at CG18557 (chymotrypsin) 4.9
150606_at CG4823
(Alpha-2-macroglobulin family)
4.6
142387_at CG8215 (serine-type endopeptidase) 4.3
143609_at Drosocin 4.0
143443_at Diptericin A 3.8
145731_at CG16712
(Kunitz-type trypsin inhibitor)
3.8
144318_at TepI 3.7
142189_at Cyp6a (cytochrome P450) 3.5
149716_at CG6977 (cadherin-like protein) 3.3
141374_at Attacin-A 3.3
147119_at Cyp9h1 (cytochrome P450) 3.2
151496_s_at CG14527
(metalloendopeptidase-like protein)
3.1
141227_at CG8370 3.0
144882_at PGRP-SA 2.9
149680_at CG17721 2.8
146354_at CG4650 (serine-type endopeptidase) 2.8
153172_at CG9524 2.8
141738_at CG9706 (acetyl-CoA transporter) 2.7
148230_at CG17795 (G protein linked receptor) 2.7
148402_at CG7120 2.7
144016_at NPC1 (lysosomal membrane receptor) 2.5
143411_at Wrinkled (apoptosis activator) 2.5
151598_at CG10077 2.5
145620_at CG4259 (chymotrypsin) 2.4
154586_at CPTI (mitoch. carnitine
palmitoyltransferase I)
2.4
153894_at CG11897 (transporter) 2.4
142654_at Ird5 (cactus kinase) 2.4
143605_at Drosomycin 2.3
142765_at CG5091 (glycosyltransferase) 2.3
154078_at Cyp4e2 (cytochrome P450) 2.3
152886_at CG15658 (LRR-containing protein) 2.3
147105_at CG8594 (chloride channel protein) 2.2
150560_at CG13654 2.2
142988_at CG6770 2.2
143723_at Cyp6a9 (cytochrome P450) 2.2
154443_at CG1667 2.1
141583_at Cyp6a20 (cytochrome P450) 2.1
146011_at CG8451 (Na+-dependent transporter) 2.1
155014_at CG5262 (permease) 2.1
151480_at CG4144 (transporter) 2.1
154187_at Gcl 2.1
148973_at Adgf-A 2.0
153367_at CG3137 (Beta-galactosidase) 2.0
142657_at PGRP-SB1 2.0
Fold change is the mean of transformed signal log2 values of 4 pair-wise
comparisons between replicate GeneChips. Names and molecular func
tions were retrieved from NetAffxk (http://www.affymetrix.com) and
Flybase gene ontology (GO) annotations.
K.C. Johansson et al. / Experimental Cell Research 305 (2005) 145–155150
-

Fig. 3. Northern blot confirmation of differentially expressed genes. Total
RNA (15 Ag/lane) from mbn-2 cells, naive (N), challenged with crude LPS
(cLPS) or live E. coli for 6 h was separated by denaturing gel
electrophoresis, immobilized on nylon membranes and hybridized with32P-labeled DNA probes for selected genes. (A) Annexin IX probe revealed
two bands corresponding to CG5730-RB (1264 bp) and CG5730-RA
(1196 bp). (B) Matrix metalloproteinase (Mmp 1) was representative of
genes that increased by co-challenge. (C) Diptericin (Dipt) and Peptido-
glycan recognition protein-SA (PGRP-SA) confirmed the class (3) profile.
(D) Unique down-regulation by bacterial challenge could be confirmed for
the GATA-factor Serpent and (CG4650), a serine-type endopeptidase. (E)
Ethidium bromide staining confirmed equal loading.
K.C. Johansson et al. / Experimental Cell Research 305 (2005) 145–155 151
line where several Hox genes were up-regulated by
Salmonella typhimurium infection [41]. Up-regulation of
Psc in Drosophila mbn-2 cells shows that immune
challenge provides a probable input into the developmental
programs regulating proliferation/differentiation of blood
cells that may be evolutionary conserved. Bacterial
challenge also induced the transcription of two Unpaired
(Upd)-related genes, upd3 (CG15062 and CG5963) and
upd2 (CG5988). Upd3 was recently demonstrated to
function as a cytokine produced by hemocytes in response
to septic injury in vivo and necessary for JAK/STAT and
Relish-dependent expression of the stress response Tur-
andot M (TotM) protein in the fat body of adult flies [42].
The function of Upd2 is at present uncertain but a role
similar to Upd3 seems likely. Of the other genes known to
function in the JAK/STAT pathway, only the cytokine-like
receptor Domeless (Dome) was up-regulated, but in
contrast to upd3 and upd2, crude LPS was enough for
up-regulation of (Dome) in mbn-2 cells. Moreover, crude
LPS induced the transcription of another Turandot peptide
TotF in mbn-2 cells (Table 2, Supplementary material).
The only inducible PGRPs in mbn-2 cells appear to be the
extracellular peptidoglycan receptor PGRP-SA which
recognizes (Lys)-type PGN leading to Toll activation
[13], a likely scavenger receptor PGRP-SB1 [43] and
PGRP-SD, which at present still awaits physiological
characterization.
Down-regulated genes
At 6 h post-E. coli challenge, the most negatively
affected genes, (z2-fold down-regulation), included pro-
teases, protease inhibitors, a significant number of AMP
genes in addition to the complement-like thiolester contain-
ing protein (TEP-family) Tep1 and the a-2-macroglobulin-
like CG4823, (Table 1 and Supplementary material). TEP-
family proteins have been proposed to function as opsonins
in the phagocytic defense of Anopheles mosquitoes [44] and
Tep1, -2, and -4 are induced after septic injury of
Drosophila adults [38]. In contrast to Tep1, Tep4 was up-
regulated in mbn-2 cells after bacterial challenge illustrating
differences between in vivo studies using whole animals and
in vitro studies with particular cell lines.
Unaffected immune genes
Neither of the proteins with demonstrated function in
phagocytosis, the scavenger receptor dSR-CI [45] and the
peptidoglycan recognition protein PGRP-LC [46] were
inducible by crude LPS or affected by bacterial infection
with the exception of the Croquemort [47], gene that was
slightly down-regulated by E. coli infection (Table 3 in
Supplementary material). Steady levels of mRNA from the
three phenoloxidase (proPO) genes; Bc, CG8193 and Dox-
A3 was also recorded (data not shown). Like in other
arthropods Drosophila phenoloxidase is produced as a
zymogen that is activated through cleavage by a serine
protease [9]. In contrast to the results of De Gregorio et al.
[38] where all of the putative proPO activating enzymes
(ProPO-AE); CG9733, CG3505, CG3066, CG16705,
CG1102 were up-regulated in flies challenged by septic
injury with a mixture of E. coli and Micrococcus luteus, we
could not detect any differential expression of these
transcripts. This may be because penetration of protective
barriers like cuticle and epithelium induces trauma and is
likely to involve additional tissues in the response.
Northern blot confirmation of the array expression data
To confirm gene expression profiles of the array
hybridization, we selected representative transcripts for
Northern analysis. Overall, there was a good correlation
between the two methods (Fig. 3) Annexin IX expression
followed the class 1 response, that is, induced expression
by crude LPS and further up-regulation by E. coli
infection. We could also confirm that Mmp1 is selec-
tively induced by bacterial infection in mbn-2 cells and
not like in SL2 cells induced by LPS alone [17]. Both
Diptericin and PGRP-SA expression correlated with the
profile of class 3 with strong up-regulation by exposure
to crude LPS and down-regulation by high-dose bacterial
infection.
K.C. Johansson et al. / Experimental Cell Research 305 (2005) 145–155152
High-dose infection with live bacteria induces
morphological changes in mbn-2 cells
The initial host response to most bacterial pathogens is
phagocytosis, however non-invasive enteropathogenic E.
coli avoids being engulfed and instead induces a spectacular
reorganization of the host cell actin cytoskeleton forming
characteristic pedestal-like structures, enriched in filamen-
tous actin, on the intestinal epithelial cells that the bacteria
resides on. The basis for these remodeling events is bacterial
adherence to the host cell surface followed by injection of the
receptor Tir (Translocated intimin receptor) and a number of
effector proteins via a type III secretion system. Interaction
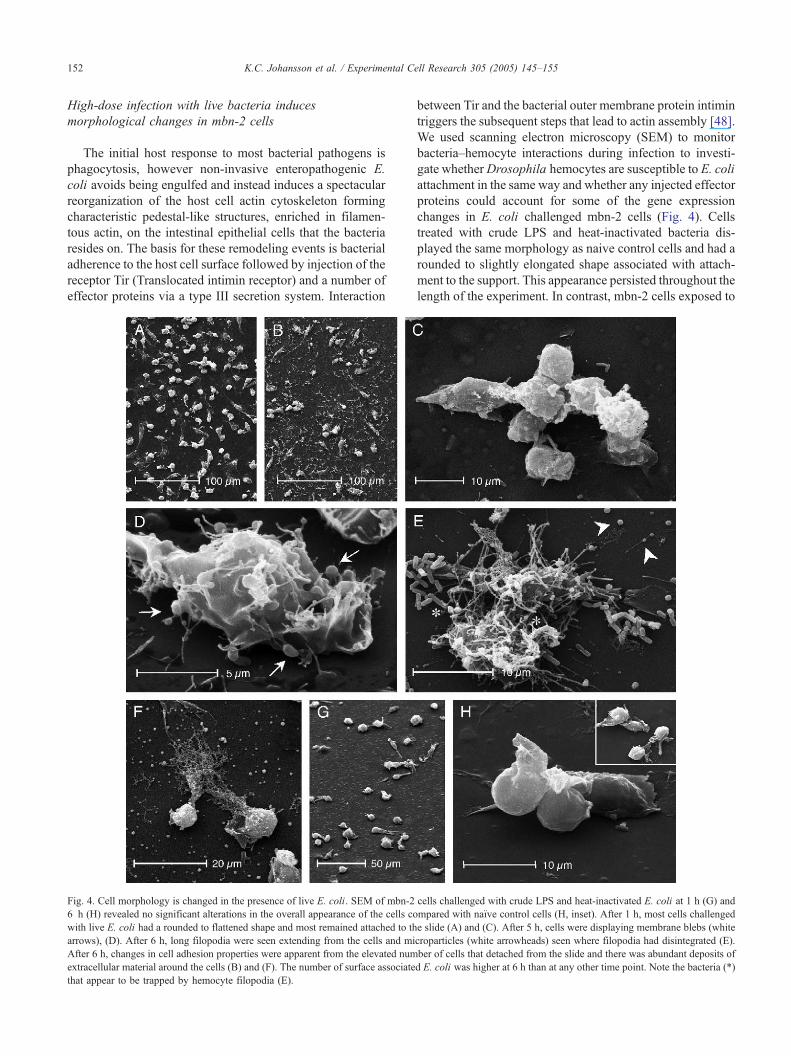
Fig. 4. Cell morphology is changed in the presence of live E. coli. SEM of mbn-2
6 h (H) revealed no significant alterations in the overall appearance of the cells co
with live E. coli had a rounded to flattened shape and most remained attached to th
arrows), (D). After 6 h, long filopodia were seen extending from the cells and mi
After 6 h, changes in cell adhesion properties were apparent from the elevated num
extracellular material around the cells (B) and (F). The number of surface associate
that appear to be trapped by hemocyte filopodia (E).
between Tir and the bacterial outer membrane protein intimin
triggers the subsequent steps that lead to actin assembly [48].
We used scanning electron microscopy (SEM) to monitor
bacteria–hemocyte interactions during infection to investi-
gate whetherDrosophila hemocytes are susceptible to E. coli
attachment in the same way and whether any injected effector
proteins could account for some of the gene expression
changes in E. coli challenged mbn-2 cells (Fig. 4). Cells
treated with crude LPS and heat-inactivated bacteria dis-
played the same morphology as naive control cells and had a
rounded to slightly elongated shape associated with attach-
ment to the support. This appearance persisted throughout the
length of the experiment. In contrast, mbn-2 cells exposed to
cells challenged with crude LPS and heat-inactivated E. coli at 1 h (G) and
mpared with naRve control cells (H, inset). After 1 h, most cells challenged
e slide (A) and (C). After 5 h, cells were displaying membrane blebs (white
croparticles (white arrowheads) seen where filopodia had disintegrated (E).
ber of cells that detached from the slide and there was abundant deposits of
d E. coli was higher at 6 h than at any other time point. Note the bacteria (*)
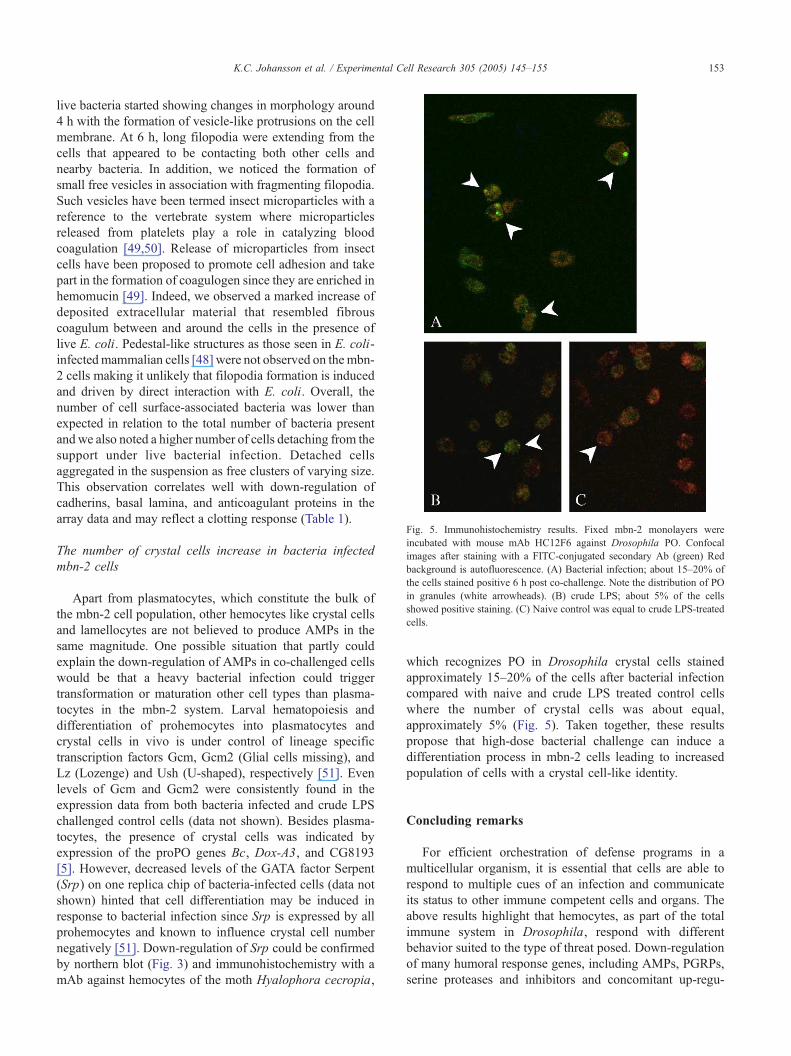
Fig. 5. Immunohistochemistry results. Fixed mbn-2 monolayers were
incubated with mouse mAb HC12F6 against Drosophila PO. Confocal
images after staining with a FITC-conjugated secondary Ab (green) Red
background is autofluorescence. (A) Bacterial infection; about 15–20% of
the cells stained positive 6 h post co-challenge. Note the distribution of PO
in granules (white arrowheads). (B) crude LPS; about 5% of the cells
showed positive staining. (C) Naive control was equal to crude LPS-treated
cells.
K.C. Johansson et al. / Experimental Cell Research 305 (2005) 145–155 153
live bacteria started showing changes in morphology around
4 h with the formation of vesicle-like protrusions on the cell
membrane. At 6 h, long filopodia were extending from the
cells that appeared to be contacting both other cells and
nearby bacteria. In addition, we noticed the formation of
small free vesicles in association with fragmenting filopodia.
Such vesicles have been termed insect microparticles with a
reference to the vertebrate system where microparticles
released from platelets play a role in catalyzing blood
coagulation [49,50]. Release of microparticles from insect
cells have been proposed to promote cell adhesion and take
part in the formation of coagulogen since they are enriched in
hemomucin [49]. Indeed, we observed a marked increase of
deposited extracellular material that resembled fibrous
coagulum between and around the cells in the presence of
live E. coli. Pedestal-like structures as those seen in E. coli-
infectedmammalian cells [48] were not observed on the mbn-
2 cells making it unlikely that filopodia formation is induced
and driven by direct interaction with E. coli. Overall, the
number of cell surface-associated bacteria was lower than
expected in relation to the total number of bacteria present
and we also noted a higher number of cells detaching from the
support under live bacterial infection. Detached cells
aggregated in the suspension as free clusters of varying size.
This observation correlates well with down-regulation of
cadherins, basal lamina, and anticoagulant proteins in the
array data and may reflect a clotting response (Table 1).
The number of crystal cells increase in bacteria infected
mbn-2 cells
Apart from plasmatocytes, which constitute the bulk of
the mbn-2 cell population, other hemocytes like crystal cells
and lamellocytes are not believed to produce AMPs in the
same magnitude. One possible situation that partly could
explain the down-regulation of AMPs in co-challenged cells
would be that a heavy bacterial infection could trigger
transformation or maturation other cell types than plasma-
tocytes in the mbn-2 system. Larval hematopoiesis and
differentiation of prohemocytes into plasmatocytes and
crystal cells in vivo is under control of lineage specific
transcription factors Gcm, Gcm2 (Glial cells missing), and
Lz (Lozenge) and Ush (U-shaped), respectively [51]. Even
levels of Gcm and Gcm2 were consistently found in the
expression data from both bacteria infected and crude LPS
challenged control cells (data not shown). Besides plasma-
tocytes, the presence of crystal cells was indicated by
expression of the proPO genes Bc, Dox-A3, and CG8193
[5]. However, decreased levels of the GATA factor Serpent
(Srp) on one replica chip of bacteria-infected cells (data not
shown) hinted that cell differentiation may be induced in
response to bacterial infection since Srp is expressed by all
prohemocytes and known to influence crystal cell number
negatively [51]. Down-regulation of Srp could be confirmed
by northern blot (Fig. 3) and immunohistochemistry with a
mAb against hemocytes of the moth Hyalophora cecropia,
which recognizes PO in Drosophila crystal cells stained
approximately 15–20% of the cells after bacterial infection
compared with naive and crude LPS treated control cells
where the number of crystal cells was about equal,
approximately 5% (Fig. 5). Taken together, these results
propose that high-dose bacterial challenge can induce a
differentiation process in mbn-2 cells leading to increased
population of cells with a crystal cell-like identity.
Concluding remarks
For efficient orchestration of defense programs in a
multicellular organism, it is essential that cells are able to
respond to multiple cues of an infection and communicate
its status to other immune competent cells and organs. The
above results highlight that hemocytes, as part of the total
immune system in Drosophila, respond with different
behavior suited to the type of threat posed. Down-regulation
of many humoral response genes, including AMPs, PGRPs,
serine proteases and inhibitors and concomitant up-regu-

K.C. Johansson et al. / Experimental Cell Research 305 (2005) 145–155154
lation of genes involved in cytoskeletal rearrangements, cell
proliferation and differentiation, suggests that cellular
response reactions, like increased phagocytosis, clotting,
and nodule formation, are favored when hemocytes are
presented with large numbers of whole bacteria. Combined
with synthesis of endocrine signaling molecules aimed at
the fat body—the main organ for production of humoral
effectors in the systemic defense in vivo, it would arguably
be a more effective way of dealing with a large bacterial
infection. Taken together, our results confirm the potency of
bacterial surface molecules (here in the form of crude LPS)
in activating humoral immunity but also shows that other
factors like bacterial titer should be taken into account as it
contributes to a balanced and adequate defense against
infection and hence, that the fly immune system may not
only discriminate between self and infectious non-self but
also respond to the way non-self is presented.
Acknowledgments
We thank Thomas Werner, Michael Williams, Carl-Johan
Zettervall and Dan Hultmark (UCMP, Ume3 University,
Sweden), Ylva Engstrfm (Department of Molecular Biol-
ogy and Functional Genomics, Stockholm University,
Sweden) and Lage Cerenius for helpful discussions, and
finally Tina Trenczek (Inst. of Zoology, Justus-Liebig
University of Giessen, Germany) for kindly providing us
with the hemocyte antibody.
This work was funded by the Wallenberg Consortium
North and the Swedish Research Council/NT and M.
Appendix A. Supplementary material
Supplementary data associated with this article can be
found, in the online version, at doi:10.1016/j.yexcr.2004.
12.018.
References
[1] R.M. Medzhitov, C.R. Janeway Jr., Innate immunity: the virtues of a
nonclonal system of recognition, Cell 91 (1997) 295–298.
[2] P. Tzou, S. Ohresser, D. Ferrandon, M. Capovilla, J.-M. Reichhart, B.
Lemaitre, J.A. Hoffmann, J.-L. Imler, Tissue-specific inducible
expression of antimicrobial peptide genes in Drosophila surface
epithelia, Immunity 13 (2000) 737–748.
[3] S. Naitza, P. Ligoxygakis, Antimicrobial defenses in Drosophila: the
story so far, Mol. Immunol. 40 (2004) 887–896.
[4] M. R7met, R. Lanot, D. Zachary, P. Manfruelli, JNK signaling
pathway is required for efficient wound healing in Drosophila,
Dev. Biol. 241 (2002) 145–156.
[5] M. Meister, M. Lagueux, Drosophila blood cells, Cell. Microbiol. 5
(2003) 573–580.
[6] Y. Carton, A.-J. Nappi, Immunogenic aspects of the cellular immune
response of Drosophila against parasitoids. Immunogenetics 52
(2001) 157–164.
[7] M.D. Lavine, M.R. Strand, Insect hemocytes and their role in
immunity, Insect Biochem. Mol. Biol. 32 (2002) 1295–1309.
[8] N.A. Ratcliffe, A.F. Rowley, Role of hemocytes in defense against
biological agents, in: A.P. Gupta (Ed.), Insect Hemocytes, Cambridge
Univ. Press, London, 1979, pp. 331–414.
[9] L. Cerenius, K. Sfderh7ll, The prophenoloxidase-activating system in
invertebrates, Immunol. Rev. 198 (2004) 116–126.
[10] U. Theopold, O. Schmidt, K. Sfderh7ll, M.S. Dushay, Coagulation in
arthropods: defense, wound closure and healing, Trends Immunol. 25
(6) (2004) 289–294.
[11] D. Hultmark, Drosophila immunity: paths and patterns, Curr. Opin.
Immunol. 15 (2003) 12–19.
[12] T. Kaneko, W.E. Goldman, P. Mellroth, H. Steiner, K. Fukase, S.
Kusumoto, W. Harley, A. Fox, D. Golenbock, N. Silverman,
Monomeric and polymeric Gram-negative peptidoglycan but not
LPS stimulate the Drosophila IMD pathway, Immunity 20 (2004)
637–649.
[13] F. Leulier, C. Parquet, S. Pili-Floury, J.-H. Ryu,M. Caroff,W.-J. Lee, D.
Mengin-Lecreulx, B. Lemaitre, TheDrosophila immune system detects
bacteria through specific peptidoglycan recognition, Nat. Immunol. 5
(2003) 478–484.
[14] T. Werner, K. Borge-Renberg, P. Mellroth, H. Steiner, D. Hultmark,
Functional diversity of the Drosophila PGRP-LC gene cluster in the
response to lipopolysaccharide and peptidoglycan, J. Biol. Chem. 278
(2003) 26319–26322.
[15] E. De Gregorio, P.T. Spellman, P. Tzou, G.M. Rubin, B. Lemaitre, The
Toll and Imd pathways are the major regulators of the immune
response in Drosophila, EMBO J. 21 (2002) 2568–2579.
[16] P. Qiu, P.C. Pan, S. Govind, A role for Drosophila Toll/Cactus
pathway in larval hematopoiesis, Development 125 (1998)
1909–1920.
[17] M. Boutros, H. Agaisse, N. Perrimon, Sequential activation of
signaling pathways during innate immune responses in Drosophila,
Dev. Cell 3 (2002) 711–722.
[18] E. Gateff, Malignant neoplasm of genetic origin in Drosophila
melanogaster, Science 200 (1978) 1448–1459.
[19] K.M. Choe, T. Werner, S. Stfven, D. Hultmark, K.V. Anderson,
Requirement for a peptidoglycan recognition protein (PGRP) in
Relish activation and antibacterial immune responses in Drosophila,
Science 296 (2002) 359–362.
[20] H. Lindmark, K.C. Johansson, S. Stfven, D. Hultmark, Y.
Engstrfm, K. Sfderh7ll, Enteric bacteria counteract lipopolysac-
charide induction of antimicrobial peptide genes, J. Immunol. 167
(2001) 6920–6923.
[21] S. Gruenheid, B. Finlay, Microbial pathogenesis and cytoskeletal
function, Nature 422 (2003) 775–781.
[22] J.-L. Dimarcq, J.-L. Imler, R. Lanot, R.A.B. Ezekowitz, J.A.
Hoffmann, C.A. Janeway, M. Lagueux, Treatment of l(2)mbn
Drosophila tumorous blood cells with the steroid hormone ecdysone
amplifies the inducibility of antimicrobial peptide gene expression,
Insect Biochem. Mol. Biol. 27 (1997) 877–886.
[23] C. Wicker, J.-M. Reichhart, D. Hoffmann, D. Hultmark, C.
Samakovlis, J.A. Hoffmann, Insect immunity. Characterization
of a Drosophila cDNA encoding a novel member of the dip-
tericin family of immune peptides, J. Biol. Chem. 265 (1990)
22493–22498.
[24] A.-I. Munier, D. Doucet, E. Perrodou, D. Zachary, M. Meister, J.A.
Hoffmann, C.A. Janeway Jr., M. Lagueux, PVF2, a PDGF/VEGF-like
growth factor, induces hemocyte proliferation in Drosophila larvae,
EMBO Rep. 12 (2002) 1195–1200.
[25] D.M. Standiford, M. Lai, M.E. Gentile, W. Sun, X. Ai, Regulation of
Wingless-dependent patterning by an extracellular sulphatase in
Drosophila, A. Dros. Res. Conf. 44 (2003) 451A.
[26] E. Martin-Blanco, A. Gampel, J. Ring, K. Virdee, N. Kirov, A.M.
Tolkovsky, A. Martinez-Arias, Puckered encodes a phosphatase that
mediates a feedback loop regulating JNK activity during dorsal
closure in Drosophila, Genes Dev. 12 (1998) 557–570.

K.C. Johansson et al. / Experimental Cell Research 305 (2005) 145–155 155
[27] A. Page-McCaw, J. Serano, J.M. Sante, G.M. Rubin, Drosophila
matrix metalloproteinases are required for tissue remodeling, but not
embryonic development, Dev. Cell 4 (1) (2003) 95–106.
[28] L. Ravanti, V.M. Kahari, Matrix metalloproteinases in wound repair,
Int. J. Mol. Med. 6 (2000) 391–407.
[29] T. Chen, M. Bunting, F.D. Karim, C.S. Thummel, Isolation and
characterization of five Drosophila genes that encode ets-related DNA
binding domain, Dev. Biol. 151 (1992) 176–191.
[30] A.M. Pearson, K. Baksa, M. R7met, M. Protas, M. McKee, D. Brown,
R.A.B. Ezekowitz, Identification of cytoskeletal regulatory proteins
required for efficient phagocytosis in Drosophila, Microbes Infect. 5
(2003) 815–824.
[31] R. Bodner, E. Lai, J. Posakony, A new gene family encoding potential
antagonists of notch signaling in the eye, A. Dros. Res. Conf. 40
(1999) 509A.
[32] T. Werner, G. Liu, D. Kang, S. Ekengren, H. Steiner, D. Hultmark, A
family of peptidoglycan recognition proteins in the fruit fly
Drosophila melanogaster, Proc. Natl. Acad. Sci. U. S. A. 97 (2000)
13772–13777.
[33] K.W. Renton, Hepatic drug metabolism and immunostimulation,
Toxicology 142 (2000) 173–178.
[34] S.M. Gorski, S. Chittaranjan, E.D. Pleasance, J.D. Freeman,
C.L. Anderson, R.J. Varhol, S.M. Coughlin, S.D. Zuyderduyn,
S.J.M. Jones, M.A. Marra, A SAGE approach to discovery of
genes involved in autophagic cell death, Curr. Biol. 13 (2003)
358–363.
[35] M. Hall, R. Wang, R. van Antwerpen, L. Sottrup-Jensen, K. Sfderh7ll,The crayfish plasma clotting protein: a vitellogenin-related protein
responsible for clot formation in crustacean blood, Proc. Natl. Acad.
Sci. U. S. A. 96 (1999) 1965–1970.
[36] K.M. Lee, K.Y. Lee, H.W. Choi, M.Y. Cho, T.H. Kwon, S.
Kawabata, B.L. Lee, Activated prophenoloxidase from Tenebrio
molitor larvae enhances the synthesis of melanin by using a
vitellogenin-like protein in the presence of dopamine, Eur. J.
Biochem. 267 (2000) 3695–3703.
[37] U. Theopold, D. Li, M. Fabbri, C. Scherfer, O. Schmidt, The
coagulation of insect hemolymph, Cell. Mol. Life Sci. 59 (2002)
363–372.
[38] E. De Gregorio, P.T. Spellman, G.M. Rubin, B. Lemaitre, Genome-
wide analysis of the Drosophila immune response by using
oligonucleotide microarrays, Proc. Natl. Acad. Sci. U. S. A. 98
(2001) 12590–12595.
[39] O. Loseva, Y. Engstrfm, Analysis of signal-dependent changes in the
proteome of Drosophila blood cells during an immune response, Mol.
Cell. Proteomics 3 (2004) 796–808.
[40] N. Remillieux-Leschelle, P. Santamaria, N.B. Randsholt, Regulation
of larval hematopoiesis in Drosophila melanogaster: a role for the
multi sex combs gene, Genetics 162 (3) (2002) 1259–1274.
[41] C.M. Rosenberger, M.G. Scott, M.R. Gold, R.E.W. Hancock, B.B.
Finlay, Salmonella typhimurium infection and lipopolysaccharide
stimulation induce similar changes in macrophage gene expression,
J. Immunol. 164 (2000) 5894–5904.
[42] H. Agaisse, U.-M. Petersen, M. Boutros, B. Mathey-Prevot, N.
Perrimon, Signaling role of hemocytes in Drosophila JAK/STAT-
dependent response to septic injury, Dev. Cell 5 (2003) 441–450.
[43] P. Mellroth, J. Karlsson, H. Steiner, A scavenger function for a
Drosophila peptidoglycan recognition protein, J. Biol. Chem. 9
(2003) 7059–7064.
[44] E.A. Levashina, L.F. Moita, S. Blandin, G. Vriend, M. Lagueux, F.C.
Kafatos, Conserved role of a complement-like protein in phagocytosis
in revealed by dsRNA knockout in cultured cells of the mosquito
Anopheles gambiae, Cell 104 (2001) 709–718.
[45] M. R7met, A. Pearson, P. Manfruelli, X. Li, H. Koziel, V. Gfbel, E.Chung, M. Krieger, R.A.B. Ezekowitz, Drosophila scavenger receptor
CI is a pattern recognition receptor for bacteria, Immunity 15 (2001)
1027–1038.
[46] M. R7met, P. Manfruelli, A. Pearson, B. Mathey-Prevot, R.A.B.
Ezekowitz, Functional genomic analysis of phagocytosis and identi-
fication of a receptor for E. coli, Nature 416 (2002) 644–648.
[47] N.C. Franc, P. Heitzler, R.A. Ezekowitz, K. White, Requirement for
Croquemort in phagocytosis of apoptotic cells in Drosophila, Science
284 (1999) 1991–1994.
[48] K.G. Campellone, J.M. Leong, Tails of two Tirs: actin pedestal
formation by enteropathogenic E. coli and enterohemorrhagic E. coli
O157:H7, Curr. Opin. Microbiol. 6 (2003) 82–90.
[49] U. Theopold, O. Schmidt, Helix pomatia Lectin and Annexin V, two
molecular probes for insect microparticles; possible involvement in
hemolymph coagulation, J. Insect Physiol. 43 (1996) 667–674.
[50] J. Dachary-Prigent, J.M. Pasquet, J.M. Freyssinet, A.T. Nurden,
Calcium involvement in aminophospholipid exposure and micro-
particle formation during platelet activation—A study using Ca2+-
ATPase inhibitors, Biochemistry 34 (1995) 11625–11634.
[51] C.J. Evans, U. Banerjee, Transcriptional regulation of hematopoiesis
in Drosophila, Blood Cells, Mol. Dis. 30 (2003) 223–228.