intrinsic pump-conduit behavior of lymphangions
TRANSCRIPT

doi:10.1152/ajpregu.00258.2006 292:1510-1518, 2007. First published Nov 22, 2006;Am J Physiol Regulatory Integrative Comp Physiol
and Randolph H. Stewart Christopher M. Quick, Arun M. Venugopal, Anatoliy A. Gashev, David C. ZawiejaIntrinsic pump-conduit behavior of lymphangions
You might find this additional information useful...
48 articles, 28 of which you can access free at: This article cites http://ajpregu.physiology.org/cgi/content/full/292/4/R1510#BIBL
including high-resolution figures, can be found at: Updated information and services http://ajpregu.physiology.org/cgi/content/full/292/4/R1510
can be found at: and Comparative PhysiologyAmerican Journal of Physiology - Regulatory, Integrativeabout Additional material and information
http://www.the-aps.org/publications/ajpregu
This information is current as of April 5, 2007 .
http://www.the-aps.org/.ISSN: 0363-6119, ESSN: 1522-1490. Visit our website at Physiological Society, 9650 Rockville Pike, Bethesda MD 20814-3991. Copyright © 2005 by the American Physiological Society. ranging from molecules to humans, including clinical investigations. It is published 12 times a year (monthly) by the Americanilluminate normal or abnormal regulation and integration of physiological mechanisms at all levels of biological organization,
publishes original investigations thatThe American Journal of Physiology - Regulatory, Integrative and Comparative Physiology
on April 5, 2007
ajpregu.physiology.orgD
ownloaded from

Intrinsic pump-conduit behavior of lymphangions
Christopher M. Quick,1,2,3,4 Arun M. Venugopal,1,3 Anatoliy A. Gashev,2,5
David C. Zawieja,2,5 and Randolph H. Stewart1,2,3
1Michael E. DeBakey Institute, Texas A&M University, College Station; 2Cardiovascular Research Institute, Texas
A&M University System, Health Science Center, College Station; 3Department of Physiology and Pharmacology
and 4Department of Biomedical Engineering, Texas A&M University, College Station, Texas; 5Department of
Medical Physiology, Texas A&M University System, Health Science Center, College Station, Texas
Submitted 14 April 2006; accepted in final form 20 November 2006
Quick CM, Venugopal AM, Gashev AA, Zawieja DC, StewartRH. Intrinsic pump-conduit behavior of lymphangions. Am J Physiol
Regnl Integr Comp Physiol 292: R1510–R1518, 2007. First publishedNovember 22, 2006; doi:10.1152/ajpregu.00258.2006.—Lymphan-gions, segments of lymphatic vessels bounded by valves, have char-acteristics of both ventricles and arteries. They can act primarily likepumps when actively transporting lymph against a pressure gradient.They also can act as conduit vessels when passively transportinglymph down a pressure gradient. This duality has implications forclinical treatment of several types of edema, since the strategy tooptimize lymph flow may depend on whether it is most beneficial forlymphangions to act as pumps or conduits. To address this duality, weemployed a simple computational model of a contracting lym-phangion, predicted the flows at both positive and negative axialpressure gradients, and validated the results with in vitro experimentson bovine mesenteric vessels. This model illustrates that contractionincreases flow for normal axial pressure gradients. With edema, limbelevation, or external compression, however, the pressure gradientmight reverse, and lymph may flow passively down a pressuregradient. In such cases, the valves may be forced open during theentire contraction cycle. The vessel thus acts as a conduit, andcontraction has the effect of increasing resistance to passive flow, thusinhibiting flow rather than promoting it. This analysis may explain apossible physiological benefit of the observed flow-mediated inhibi-tion of the lymphatic pump at high flow rates.
lymphangion; time-varying elastance; shear stress-induced dilation
THE RELATIVELY SIMPLE STRUCTURE of collecting lymphatic ves-sels yields complex behavior. Like veins, lymphatic vessels areremarkably thin-walled and contain unidirectional valves.Whereas the initial lymphatics are typically passive structures,the conducting lymphatics contain smooth muscle that cycli-cally contracts at a rate of 1–15 cycles per minute (2, 13, 27,42, 53). The section of vessel between valves forms a func-tional unit, called a lymphangion (32). Lymphangions arearranged to form a converging structure that returns interstitialfluid, called lymph once it enters the lymphatic system, to thegeneral circulation through the great veins of the neck. Cyclicalcontractions of lymphangions usually actively pump lymphagainst a local pressure gradient. The concept that lymph isactively transported by lymphatic vessels took years to beaccepted (17) and has become the focus of investigators inter-ested in the role of lymphatic function in the genesis andresolution of edema (2, 4, 6, 25, 27, 42, 53).
This pumping behavior has lead investigators to describelymphangion function in terms typically applied to ventricular
function. For instance, lymphangions have discernable systolicand diastolic periods, and ejection fraction, stroke volume, andstroke work have been used to describe active lymphangionoutput per cycle (3, 25). Borrowing the terms describingventricular function is not unwarranted. Transmural pressure-volume loops, for instance, reveal behavior similar to that ofventricles, including an identifiable end-systolic pressure-vol-ume relationship (25). Furthermore, lymphangion output in-creases with preload, like the Frank-Starling effect (28). Lym-phangion and lymphatic vessel output also is sensitive toafterload (6, 23). Characterization of lymphatic function usingventricular analogies is complicated, in part, because the af-terload of one lymphangion forms the preload of the next oneor more (16), yielding complex and variable interactionsamong lymphangions within a lymphatic vessel (53).
The use of ventricular analogies to describe lymphangions,however, is insufficient, because lymphangions exhibit somebehaviors best described by analogy to muscular arteries.Although lymphangions cyclically contract, they can exhibit abasal tone (26) that is sensitive to vasoactive substances suchas NO donors (43), prostaglandins, and thromboxane (22).Furthermore, high-flow conditions cause lymphatic vessels torelax (15), much like the shear stress-induced vasodilationfound in muscular arteries. Wall tension-length relationshipsare similar to that of muscular arteries, with an optimal radiusthat yields an optimal developed tension (10, 42). Interest inthis constellation of artery-like behaviors has not been recon-ciled with descriptions of lymphangions as pumps.
Interpreting lymphatic vessels only as a series of pumpsbecomes problematic when they are exposed to conditionscommon to arteries. Both high transmural pressures, leading topump failure (4, 25, 28, 36), and high flow rates, leading toshear stress-induced relaxation (15), can inhibit active lym-phatic pumping. Since these behaviors occur with edema, theymay be perceived as maladaptive. However, if the inlet pres-sure of a lymphatic vessel rises significantly higher than itsoutlet pressure, a fundamentally new situation arises. In thiscase, the lymphatic inlet pressure is high enough that lym-phangions are no longer pumping against a local pressuregradient. Lymphatic function, assessed by lymph flow, has notbeen sufficiently investigated for the special case in which inletpressure is greater than outlet pressure. Since lymph flowdepends on preload, afterload, contractility, temperature, timedelay between lymphangion contractions, tissue pressure, andlymph viscosity, it is understandable that many investigators
Address for reprint requests and other correspondence: C. M. Quick, Collegeof Veterinary Medicine, 4466 Texas A&M Univ., College Station, TX 77843-4466 (e-mail: [email protected]).
The costs of publication of this article were defrayed in part by the paymentof page charges. The article must therefore be hereby marked “advertisement”in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
Am J Physiol Regul Integr Comp Physiol 292: R1510–R1518, 2007.First published November 22, 2006; doi:10.1152/ajpregu.00258.2006.
0363-6119/07 $8.00 Copyright © 2007 the American Physiological Society http://www.ajpregu.orgR1510
on April 5, 2007
ajpregu.physiology.orgD
ownloaded from

have restricted themselves to studying flow through lymphaticvessels with a zero axial (end-to-end) pressure gradient (8, 19,28, 29).
The complexity arising from the interaction of lymphangionsis contrasted with the comparative simplicity of lymphaticvessel structure and fluid flow. The mechanics of fluid flowthrough vessels is fairly well understood, and computationalmodels characterizing this flow have been well established (35,52). Furthermore, ventricular contraction also has been wellcharacterized, and computational models have been used suc-cessfully to describe their function in a variety of loadingconditions (45, 49, 50). Computational modeling, combiningmodels of arteries and ventricles, provides the means to char-acterize lymphatic function without neglecting many dynamiccomplexities while focusing in particular on the response tochanges in axial pressure gradients.
The purpose of this study was to determine the effect ofreversing the axial pressure gradient on lymph flow, using acomputational model of a single lymphangion, and to verifypredicted results with focused in vitro experiments. We pro-pose that under normal conditions, lymphangions act primarilyas active pumps, but with elevated inlet pressure, they canbegin to act as passive conduits.
METHODS
Theory
Characterizing lymphangion contraction. The need to characterizethe inherent contractility of a lymphatic vessel independent of itsloading conditions is similar to the historic need to characterizeventricular contractility independent of both preload and afterload.Suga and colleagues (47–49), in a series of seminal papers, developeda relatively simple description of the heart that related ventricularchamber pressure, Pt, to chamber volume, V. This relationship, namedthe time-varying elastance, E(t), is a function of time and can becalculated from measured Pt and V
E�t� �Pt�t�
V�t� � Vo
(1)
where Vo is the dead volume (the theoretical volume at zero pressure).The maximum value, Emax, is the slope of the end-systolic pressure-volume relationship, and the minimum value, Emin, is the slope of theend-diastolic pressure-volume relationship. The value of Vo can becalculated from the intercept of the end-systolic pressure-volumerelationship. Pt is a transmural pressure, the difference in luminal andexternal pressures, and thus can be modified by external compression.Although it has been subject to numerous criticisms and improve-ments since its inception (37, 44), this simple description formed thebasis for successful models of heart-arterial system interaction (3, 11)and has been used in the present work to describe lymphangioncontraction.
Characterizing lymph dynamics. The need to characterize pulsatilelymph flow as a function of pressure is similar to the historic need tocharacterize the pulsatile pressure-flow relationship in arteries. Reddyand colleagues (39, 40) recognized the need for a fundamental basisfrom which to predict lymph flow and derived equations from sim-plifications of the Navier-Stokes equation. The results are equivalentto equations described by Noordergraaf (35) for blood flow in arteries.In essence, the resulting relationship depends on tube radius, r, length,L, fluid density, �, and viscosity, �. Assuming a cylindrical shape, thepressure drop across a segment of vessel, �P, depends on a resistiveterm and a term related to fluid inertia.
�P �8�L
�r4 Q ��L
�r2
dQ
dt(2)
Unlike Eq. 1, the pressure in this case is a difference in upstream anddownstream luminal pressures. We have added the inertial term to themodel to ensure that the model accounts for fluid acceleration duringejection, even though it is expected to have a smaller effect than theresistive term. Continuity is assured if the rate at which volume isstored is equal to the difference of the flow into the vessel, Qin, and theflow out of the segment, Qout.
dV
dt� Qin � Qout (3)
In addition, a small resistance is added before and after the inlet andoutlet of the vessel, to simulate the resistance of the tubing used for invitro experiments, and is analogous to upstream and downstreamresistances in vivo. These equations have been successfully used as abasis to describe the interaction of multiple vessels in a vascularnetwork (21, 51) and are used in the present work to describe lymphflow. The fundamental difference, however, is that radius in Eq. 2 isconsidered a variable, rather than a constant.
Description of valves. The valves in lymphangions are very thinand prevent, or at least minimize, retrograde flow (12). In concert withcomputational descriptions of ventricular valves, the lymphatic valvesare assumed to open when the pressure immediately proximal tothe valve is greater than the pressure immediately distal to the valve.The valve is assumed to close when the pressure immediately distalto the valve is greater than the pressure immediately proximal to thevalve. Because there is a rigid annulus surrounding the valve struc-ture, it is explicitly assumed to depend only on the axial pressuregradient, and not transmural pressure.
External compression. Notably, the axial pressure gradient in Eq. 2is expressed in terms of luminal pressures only, but the pressure inEq. 1 is expressed in terms of transmural pressure (luminal pressureminus external pressure). Characterizing lymphangion contraction interms of transmural pressure allows this system of equations to predictthe effects of both intrinsic and extrinsic propulsion of lymph. Theunderlying assumption, however, is that external compression isuniform. This may be a questionable assumption for compression byskeletal muscle in vivo but is a fair approximation for compressiondue to submersion of a lymphangion in a vessel bath in vitro.
Boundary conditions and solution technique. With lymphangionsdescribed by Eqs. 1–3, the flow resulting from a pressure gradient canbe predicted. To simplify, the inlet and outlet pressures were set tomimic those in vitro. All inlet and outlet pressures were maintained�8 mmHg to ensure that transmural pressure remained below a levelthat would be expected to induce pump failure (13). All equationswere solved using a multistep solver (implementing Gear’s method) inMatLab, which is a particularly stable solver for these nonlinearequations (Eqs. 1–3). Flow was solved as a function of time, and thesolution was obtained after the system achieved steady state. Toestimate the sensitivity of the solution to the particular parameters, wecalculated the ranges of parameters necessary to cause a 10% changein predicted mean flow.
Experimental Procedures
Postnodal mesenteric lymphatic vessels were isolated from eutha-nized cattle and placed in a tubular organ bath (Harvard Apparatus,Holliston, MA). Vessel segments containing no valves (allowingmeasurement of transmural pressure) were used for experiments toestablish elastance parameters. Vessel segments containing two valveswere used for experiments to demonstrate the effect of axial pressuregradient on flow. The vessels were perfused and bathed with abalanced buffered polyionic solution with 1% albumin, gassed withroom air, and set to 37°C and pH 7.4 as shown in Fig. 1. Because the
R1511PUMP-CONDUIT BEHAVIOR
AJP-Regul Integr Comp Physiol • VOL 292 • APRIL 2007 • www.ajpregu.org
on April 5, 2007
ajpregu.physiology.orgD
ownloaded from
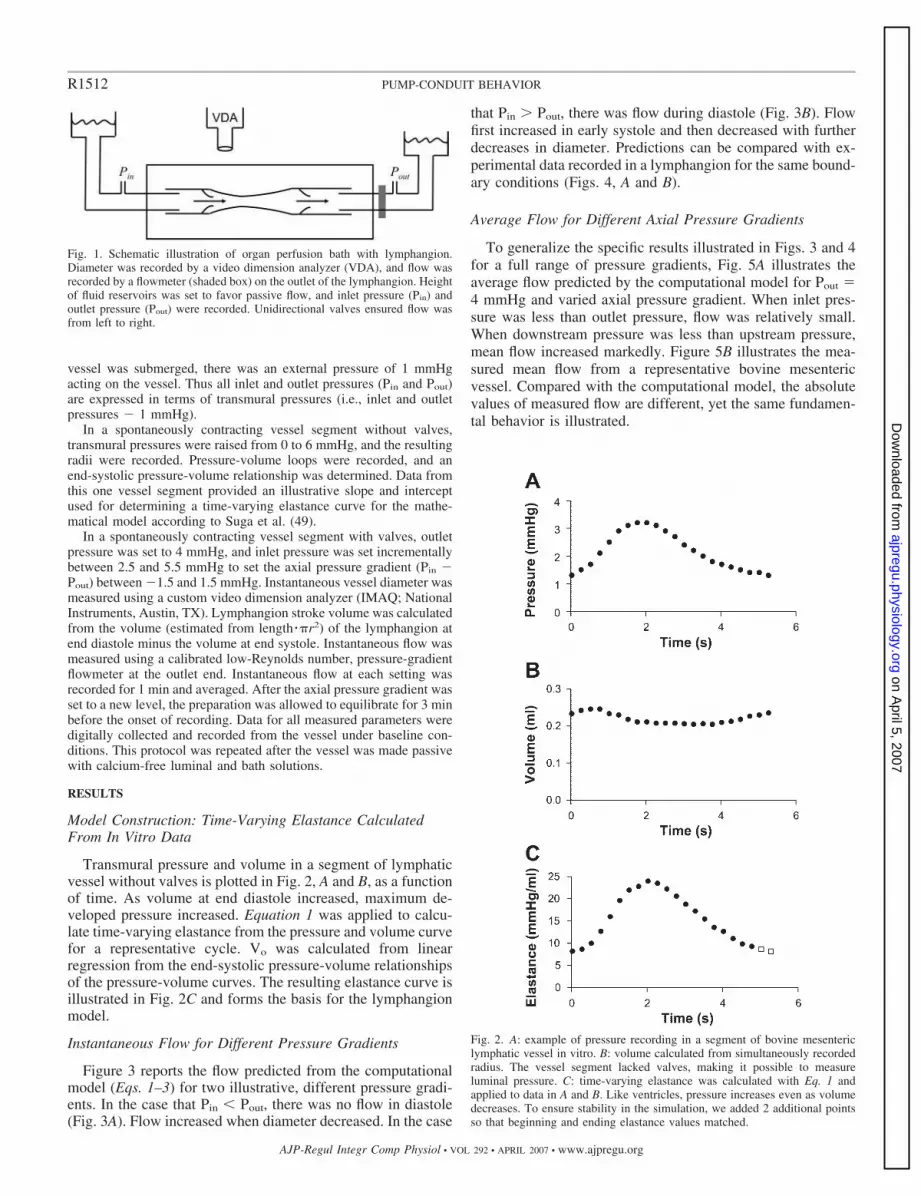
vessel was submerged, there was an external pressure of 1 mmHgacting on the vessel. Thus all inlet and outlet pressures (Pin and Pout)are expressed in terms of transmural pressures (i.e., inlet and outletpressures � 1 mmHg).
In a spontaneously contracting vessel segment without valves,transmural pressures were raised from 0 to 6 mmHg, and the resultingradii were recorded. Pressure-volume loops were recorded, and anend-systolic pressure-volume relationship was determined. Data fromthis one vessel segment provided an illustrative slope and interceptused for determining a time-varying elastance curve for the mathe-matical model according to Suga et al. (49).
In a spontaneously contracting vessel segment with valves, outletpressure was set to 4 mmHg, and inlet pressure was set incrementallybetween 2.5 and 5.5 mmHg to set the axial pressure gradient (Pin �Pout) between �1.5 and 1.5 mmHg. Instantaneous vessel diameter wasmeasured using a custom video dimension analyzer (IMAQ; NationalInstruments, Austin, TX). Lymphangion stroke volume was calculatedfrom the volume (estimated from length ��r2) of the lymphangion atend diastole minus the volume at end systole. Instantaneous flow wasmeasured using a calibrated low-Reynolds number, pressure-gradientflowmeter at the outlet end. Instantaneous flow at each setting wasrecorded for 1 min and averaged. After the axial pressure gradient wasset to a new level, the preparation was allowed to equilibrate for 3 minbefore the onset of recording. Data for all measured parameters weredigitally collected and recorded from the vessel under baseline con-ditions. This protocol was repeated after the vessel was made passivewith calcium-free luminal and bath solutions.
RESULTS
Model Construction: Time-Varying Elastance CalculatedFrom In Vitro Data
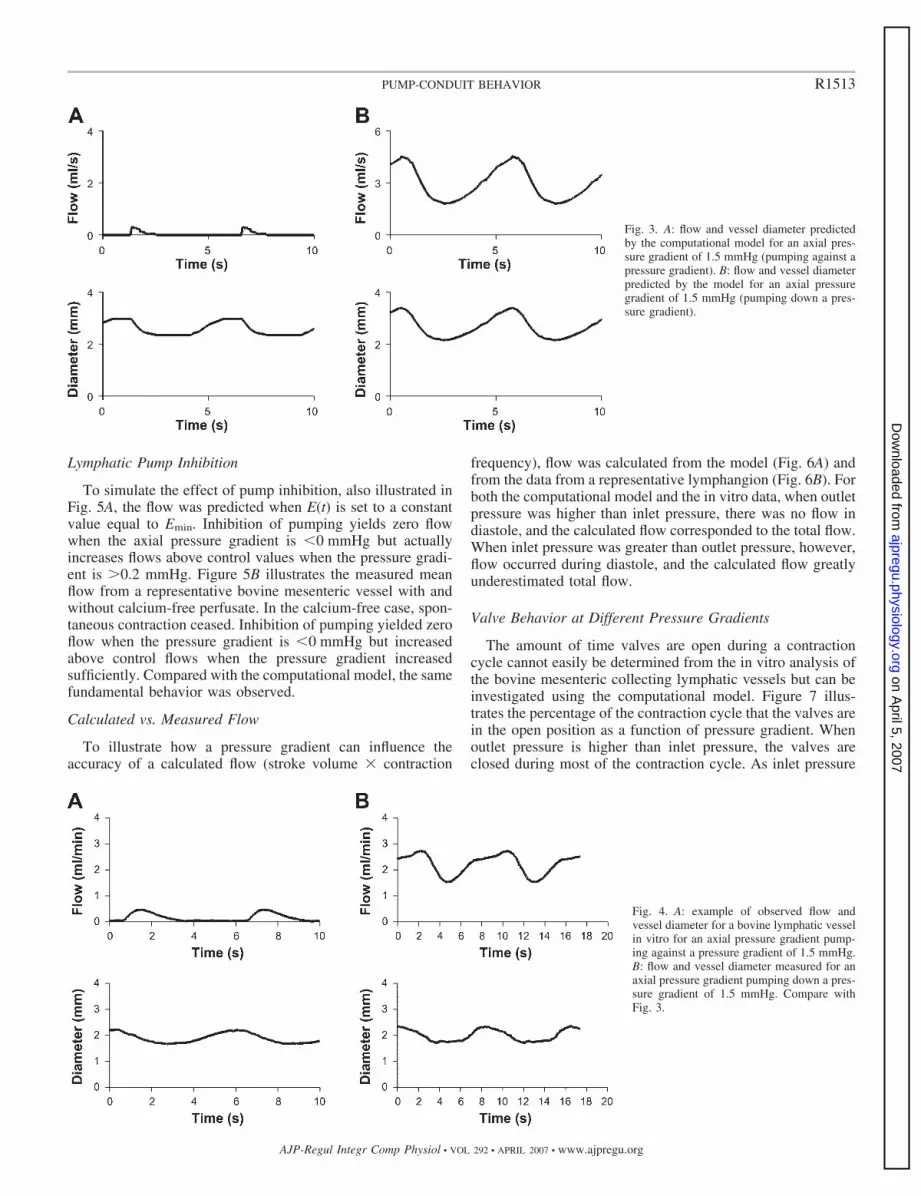
Transmural pressure and volume in a segment of lymphaticvessel without valves is plotted in Fig. 2, A and B, as a functionof time. As volume at end diastole increased, maximum de-veloped pressure increased. Equation 1 was applied to calcu-late time-varying elastance from the pressure and volume curvefor a representative cycle. Vo was calculated from linearregression from the end-systolic pressure-volume relationshipsof the pressure-volume curves. The resulting elastance curve isillustrated in Fig. 2C and forms the basis for the lymphangionmodel.
Instantaneous Flow for Different Pressure Gradients
Figure 3 reports the flow predicted from the computationalmodel (Eqs. 1–3) for two illustrative, different pressure gradi-ents. In the case that Pin � Pout, there was no flow in diastole(Fig. 3A). Flow increased when diameter decreased. In the case
that Pin Pout, there was flow during diastole (Fig. 3B). Flowfirst increased in early systole and then decreased with furtherdecreases in diameter. Predictions can be compared with ex-perimental data recorded in a lymphangion for the same bound-ary conditions (Figs. 4, A and B).
Average Flow for Different Axial Pressure Gradients
To generalize the specific results illustrated in Figs. 3 and 4for a full range of pressure gradients, Fig. 5A illustrates theaverage flow predicted by the computational model for Pout 4 mmHg and varied axial pressure gradient. When inlet pres-sure was less than outlet pressure, flow was relatively small.When downstream pressure was less than upstream pressure,mean flow increased markedly. Figure 5B illustrates the mea-sured mean flow from a representative bovine mesentericvessel. Compared with the computational model, the absolutevalues of measured flow are different, yet the same fundamen-tal behavior is illustrated.
Fig. 2. A: example of pressure recording in a segment of bovine mesentericlymphatic vessel in vitro. B: volume calculated from simultaneously recordedradius. The vessel segment lacked valves, making it possible to measureluminal pressure. C: time-varying elastance was calculated with Eq. 1 andapplied to data in A and B. Like ventricles, pressure increases even as volumedecreases. To ensure stability in the simulation, we added 2 additional pointsso that beginning and ending elastance values matched.
Fig. 1. Schematic illustration of organ perfusion bath with lymphangion.Diameter was recorded by a video dimension analyzer (VDA), and flow wasrecorded by a flowmeter (shaded box) on the outlet of the lymphangion. Heightof fluid reservoirs was set to favor passive flow, and inlet pressure (Pin) andoutlet pressure (Pout) were recorded. Unidirectional valves ensured flow wasfrom left to right.
R1512 PUMP-CONDUIT BEHAVIOR
AJP-Regul Integr Comp Physiol • VOL 292 • APRIL 2007 • www.ajpregu.org
on April 5, 2007
ajpregu.physiology.orgD
ownloaded from
Lymphatic Pump Inhibition
To simulate the effect of pump inhibition, also illustrated inFig. 5A, the flow was predicted when E(t) is set to a constantvalue equal to Emin. Inhibition of pumping yields zero flowwhen the axial pressure gradient is �0 mmHg but actuallyincreases flows above control values when the pressure gradi-ent is 0.2 mmHg. Figure 5B illustrates the measured meanflow from a representative bovine mesenteric vessel with andwithout calcium-free perfusate. In the calcium-free case, spon-taneous contraction ceased. Inhibition of pumping yielded zeroflow when the pressure gradient is �0 mmHg but increasedabove control flows when the pressure gradient increasedsufficiently. Compared with the computational model, the samefundamental behavior was observed.
Calculated vs. Measured Flow
To illustrate how a pressure gradient can influence theaccuracy of a calculated flow (stroke volume � contraction
frequency), flow was calculated from the model (Fig. 6A) andfrom the data from a representative lymphangion (Fig. 6B). Forboth the computational model and the in vitro data, when outletpressure was higher than inlet pressure, there was no flow indiastole, and the calculated flow corresponded to the total flow.When inlet pressure was greater than outlet pressure, however,flow occurred during diastole, and the calculated flow greatlyunderestimated total flow.
Valve Behavior at Different Pressure Gradients
The amount of time valves are open during a contractioncycle cannot easily be determined from the in vitro analysis ofthe bovine mesenteric collecting lymphatic vessels but can beinvestigated using the computational model. Figure 7 illus-trates the percentage of the contraction cycle that the valves arein the open position as a function of pressure gradient. Whenoutlet pressure is higher than inlet pressure, the valves areclosed during most of the contraction cycle. As inlet pressure
Fig. 3. A: flow and vessel diameter predictedby the computational model for an axial pres-sure gradient of 1.5 mmHg (pumping against apressure gradient). B: flow and vessel diameterpredicted by the model for an axial pressuregradient of 1.5 mmHg (pumping down a pres-sure gradient).
Fig. 4. A: example of observed flow andvessel diameter for a bovine lymphatic vesselin vitro for an axial pressure gradient pump-ing against a pressure gradient of 1.5 mmHg.B: flow and vessel diameter measured for anaxial pressure gradient pumping down a pres-sure gradient of 1.5 mmHg. Compare withFig. 3.
R1513PUMP-CONDUIT BEHAVIOR
AJP-Regul Integr Comp Physiol • VOL 292 • APRIL 2007 • www.ajpregu.org
on April 5, 2007
ajpregu.physiology.orgD
ownloaded from

approaches outlet pressure, the valves remain open a largerportion of the cycle. When inlet pressure rises above outletpressure, there is some flow in diastole, and the percentage oftime the valves are open increases markedly. At a criticalpressure gradient, the valves remain open for the entire con-traction cycle, indicating that the lymphangion is acting as aconduit. This prediction is difficult to confirm experimentallyin our study, given the difficulty of viewing valves through thelymphangion wall in the in vitro preparation.
Parameter Sensitivity
To determine the sensitivity of the model to assumed modelparameters, we calculated the range of parameters resulting ina 10% change in mean flow for the case of a pressure gradientof 1.5 mmHg. Experimental values for elastance used for themodel include both Emax and Emin. Emin can be varied up to 5%before altering calculated flow by 10%, and Emax can be variedup to 11% before altering calculated flow by 10%. The exper-imental errors in Emax and Emin measurement are estimated tobe �5%, given that pressure measurement error is �2% andradius measurement error is likewise �2%. Because input andoutput resistances were small, changes in their resistances hadnegligible effects on flow.
DISCUSSION
This work utilized multiple approaches to illustrate thatlymphangions can behave as either pumps or conduits. Underconditions in which inlet pressure is less than outlet pressure,flow through a particular lymphangion only occurs when ves-sels actively pump. Under conditions in which inlet pressureexceeds outlet pressure, however, there can be flow duringdiastole, and contraction may only partially augment flow.When the inlet pressure is high enough for the valves to beopen the entire contraction cycle, lymphangions become con-duits. When vessel segments are thus acting like conduits,cyclical contraction is unnecessary to propel lymph and, infact, cyclical contraction can impede flow. Because lym-phangions decrease pumping and decrease baseline tone with
Fig. 5. Effect of active contraction with mean flow out of a lymphangion whenoutlet pressure is set to 4 mmHg and with a variable axial pressure gradient. A:computational model results with cyclical contraction (F) and with cyclicalcontraction eliminated by setting Emax Emin (E). B: mean flow measuredfrom a representative bovine lymphatic vessel with (F) and without (E)contraction (i.e., contraction eliminated with calcium-free perfusate). Meanflow at a zero axial pressure gradient was 0.4 ml/min, consistent with otherreports (8, 19, 28, 29).
Fig. 6. Flow as a function of axial pressure gradient. True flow was measuredby flowmeter (F), and apparent flow was calculated from stroke volume �contraction frequency (E) in the computational model (A) and in representativelymphangion studied in vitro (B).
Fig. 7. Percentage of contraction cycle that the valves are open. Values arepredicted from the computational model under conditions similar to thosedescribed in Fig. 6. Valves open completely during the entire cycle when inletpressure is significantly higher than outlet pressure.
R1514 PUMP-CONDUIT BEHAVIOR
AJP-Regul Integr Comp Physiol • VOL 292 • APRIL 2007 • www.ajpregu.org
on April 5, 2007
ajpregu.physiology.orgD
ownloaded from

flow down a pressure gradient (14, 15), it is likely thatlymphangions act as self-regulating units that become betterconduits under high-flow conditions. Whereas early reportsdescribed lymphatic vessels as conduits, and newer functionalstudies describe them as pumps, the present work attempts toreconcile these two descriptions and delineate the conditions inwhich one description is better than the other.
Conditions That May Lead to Conduit Behavior In Vivo
The present work used experimental and theoretical ap-proaches to predict behavior of lymphangions when the normalaxial pressure gradient reverses. Although the scope of thepresent work did not include in vivo studies, four cases likelyresult in an altered lymphatic axial pressure gradient: intersti-tial edema, limb elevation, extrinsic compression, and contrac-tion of upstream lymphangions.
Interstitial edema. Many forms of interstitial edema will notresult in a reversal in the lymphatic axial pressure gradient,because interstitial fluid pressure may increase very little (1),especially with inflammatory edema (34, 38). However, sev-eral notable cases have suggested that interstitial pressure canrise significantly: 1) Mortillaro and Taylor (33) showed thatintestinal interstitial fluid pressure increased from �0.56 �0.193 to 7.3 � 0.63 mmHg as a result of venous hypertension;2) Laine et al. (24) showed that hepatic interstitial fluid pres-sure increased from 5.8 � 0.87 to 20 mmHg in response toinferior vena caval hypertension; 3) Stewart and Laine (46)showed that cysterna chili pressure increased from 1.0 � 0.2 to6.0 � 1.0 mmHg following partial occlusion of the inferiorvena cava; and 4) Granger et al. (18) estimated that intestinalinterstitial fluid pressure increased from 0.82 � 0.72 to 4.4 �0.76 mmHg during intestinal lipid absorption. In these cases, itmay be possible for average lymphangion inlet pressures to behigher than average outlet pressures, favoring the conduitbehavior described in the present work.
Limb elevation. Even if edema does not act to reverse thenormal pressure gradient in lymphatic vessels, a commontreatment may. Limb elevation has long been recognized as aneffective treatment of local edema (31). In this case, the inletlymphangion pressure can increase an amount equal to thepressure head generated from elevation. When a limb is ele-vated above the great veins of the neck, the axial pressuregradient could favor the passive flow of lymph (Fig. 7) andthus yield conduit behavior.
External compression. Cyclical lymphangion compressionresulting from locomotion, intestinal peristalsis, or respiration(20, 41) can augment or even obviate the need for intrinsiclymphatic pumping. External compression, however, also mayalter the inlet lymphangion pressure. McGeown et al. (30)illustrated that lymphatic outflow can increase fourfold withintermittent compression over metatarsal regions of sheep.
Contraction of upstream lymphangions. Lymphangions cangenerate pressures at least 20 mmHg, a value much higher thannormal central venous pressure. Although this may appear topreclude the possibility for the conduit behavior explored in thepresent work, it is the local axial pressure gradient that deter-mines whether a particular lymphangion acts as a conduit. If,for instance, an upstream lymphangion cycles between 3 and15 mmHg and a downstream lymphangion cycles in phasefrom 2 to 14 mmHg, the data in Fig. 7 suggest this axial
gradient would be enough (i.e., 1 mmHg) to ensure passivelymph flow. Valves separating intermediate lymphangions maynever close, and cyclical contraction of intermediate lym-phangions may inhibit this passive flow. Similarly, if contrac-tility of an upstream lymphangion is stimulated, or contractilityof a downstream lymphangion is inhibited, lymphangionscould generate a local pressure gradient that favors conduitbehavior. Since the pressure gradient between lymphangions istypically small (7), and regional variations in contractility canbe quite large (14), there may be numerous opportunities for a1-mmHg pressure gradient to arise, favoring local conduitbehavior.
Transition from a Pump to a Conduit
Most lymphatic studies in vitro or in situ are performed withthe axial pressure gradient set to zero. In our validation exper-iment with a bovine vessel, the flow at an axial pressuregradient of zero was 0.4 ml/min. This value corresponds toreported flows of 0.27–1.3 ml/min (29), 0.2–1.0 ml/min (8),and 0.2–1.0 ml/min (19). Also, McHale and Roddie (28)reported the flows from one vessel ranging from 0.38 to 0.8ml/min. The high sensitivity of lymphangions to axial pressuregradients, especially near a zero pressure gradient, allows anabrupt transition from a low-flow pump to a high-flow conduitand may explain, in part, the high degree of variability oflymphatic vessel behavior. Not only does the transition frompump to conduit result in very large changes in flow, there is arange of low pressure gradients where the behavior is notcompletely that of a pump or a conduit. Under these conditions,flow occurs during diastole, yet vessel contraction augmentstotal flow. Many investigators have characterized lymphaticvessel function in vitro or in situ with a zero axial pressuregradient. Because the pressure gradient across each lym-phangion is temporally variable due to the cyclic contraction ofupstream and downstream lymphangions, this type of experi-ment would ensure that some lymphangions would be actinglike pumps, some would be acting like conduits, and somewould be in transition from one to the other.
New Direction for Treatment of Edema
This work suggests a new direction in clinical researchfocusing on treatment of some kinds of edema, especially thosecaused by increased microvascular pressures or permeability.Typically, lymphatic vessels are viewed only as pumps, andthus only pump failure is understood to lead to edema (4, 5). Itfollows that an increase in the cyclical contraction of lym-phangions will increase lymph flow and thus is necessary toresolve edema. This could be true for edema resulting fromincreased downstream resistance or pressure for a given lym-phatic bed (e.g., for postinflammatory partial lymphatic ob-struction). However, if interstitial edema were to cause inletpressure in a given lymphatic bed to rise above outlet pressure,vessels may become conduits, thus vessel radius determinesthe resistance to passive flow. In fact, from the computationalmodel, validated by in vitro experimentation, flow can increasewhen cyclical contraction is inhibited. If the clinical goal inedema treatment for such cases is to increase drainage of fluidfrom the interstitium, then promoting vessel relaxation, notlymphatic pumping stimulation, may best achieve this goal.
R1515PUMP-CONDUIT BEHAVIOR
AJP-Regul Integr Comp Physiol • VOL 292 • APRIL 2007 • www.ajpregu.org
on April 5, 2007
ajpregu.physiology.orgD
ownloaded from

From Apparent Paradox to New Anti-Edema Mechanism:Reinterpreting Pump Failure
Gashev et al. (15) reported that small transport vesselsrelaxed when presented with progressively increasing axialpressure gradients and, presumably, high endothelial shearstresses. This behavior is consistent with shear stress-induceddilation found in arteries. This pump inhibition could beviewed as a maladaptive response to edema. This work, how-ever, provides evidence that pump inhibition is instead “areasonable physiological mechanism” (15) to save energy byreducing contractions and, thus, reducing resistance to lymphflow. Similarly, the observation that stroke volume of lym-phatic vessels diminishes when transmural pressure rises abovea critical value (28) may be described as “pump failure.” Sincelymphatic transmural pressure increases in edema, this behav-ior also has been viewed as a maladaptive response to edemaformation. Of course, the underlying assumption has been thatpumping is always necessary to resolve edema. The presentwork provides evidence that “pump failure” may be a benefi-cial adaptation that decreases resistance to passive lymph flow,a manifestation of a previously unappreciated anti-edemamechanism.
Increasing Pumping vs. Pump Inhibition in Responseto Edema
The question of whether “pump failure” is detrimental orbeneficial for resolving edema may not be answerable with theuse of in vitro experiments in which blood-borne or lymph-borne humoral factors, innervation, and tethering of the vesselto the surrounding extracellular matrix are removed, and everyeffort is made to keep inlet and outlet pressures constant. Inaddition, because the lymphangion interacts with its environ-ment, the critical pressures causing a transition from pump toconduit cannot be determined from in vitro experiments; lym-phangions can alter their own inlet and outlet pressures. Fur-thermore, the transition point from pump to conduit is notimmutable, since it depends on a lymphangion’s ability tocontract. For instance, endotoxin can inhibit lymphatic pump-ing at a particular transmural pressure (9), suggesting that theresponse to a significant edemagenic challenge is to become aconduit. However, endotoxin causes lymphangions to be betterpumps at higher pressures (9), possibly optimizing pumpingfor the higher interstitial pressures that result from endotoxin.Pump-conduit duality of lymphangions complicates the inter-pretation of both in vivo and in vitro data and puts into boldrelief the need to take into consideration the complex interac-tion of lymphatic vessels and their environment.
Apparent vs. True Lymph Flow
Because flow is difficult to measure in vivo, it is possiblethat the effect of pump inhibition in edemagenic states mayhave been misinterpreted. Lymph flow is difficult to measuredirectly with intravital microscopy in the microlymphaticsbecause lymph for the most part is transparent. To deal withthis limitation, an apparent flow has been calculated fromstroke volume multiplied by contraction frequency (2). Fromthis method, for instance, Benoit et al. (2) calculated that inresponse to volume loading, the calculated flow from smallcollecting lymphatics in the mesentery of a rat increased
threefold. The underlying assumption of this calculation is thatthere is no passive flow in diastole and that all flow out of alymphangion is equal to the volume change of the lym-phangion. However, the current study presents clear evidencethat it is possible for flow through a lymphangion to occur indiastole when inlet pressure is higher than outlet pressure. Infact, flow-induced pump inhibition can cause the calculatedlymph flow to actually decrease in edema. Failure to describelymphatic vessels as conduits in edema can yield misleadingresults.
Broadening the Scope
A pragmatic approach informed the scope of the currentwork. The model and the experiment were designed to becomplementary so that the transition from pump to conduitbehavior could be studied in the most elemental functional unitof the lymphatic system: the lymphangion. To interpret exper-imental results fully and place the model on a firm basis, themathematical model was designed to mimic the in vitro systemrather than a portion of the lymphatic vasculature in vivo. Thismodel also focused on intrinsic lymphatic pumping due tosmooth muscle activity, rather than the important effect ofextrinsic pumping arising from cyclical external compression(20, 30). Equation 1, however, was formulated in terms oftransmural rather than luminal pressure and could be used tofurther study the effects of cyclical compression. Nonetheless,the fundamental behavior of interest, the transition from apump to a conduit, can be expected in portions of a lymphaticvessel segment, a network of lymphatic vessels, and an entirelymphatic network. Each of these three hierarchical levels hasbeen modeled by Drake and Gabel (5), Stewart and Laine (46),and Reddy et al. (40), respectively. Only Drake and Gabel (5)tangentially described this transition. Potentially, pump-con-duit behavior of lymphangions could be complex, at eachhierarchical level. For instance, at the level of a single vessel,it is possible that an upstream lymphangion forces flow througha downstream lymphangion, causing it to transition from apump to a conduit. Similarly, with two convergent lymphaticvessels, the pumping in one vessel could raise the outletpressure of the other vessel, causing it to transition from aconduit to a pump. An entire lymphatic system could thereforehave multiple sections that are acting like both pumps andconduits. By limiting the scope of the present work to intrinsiccontraction of lymphangions, vessels, networks, and the entiresystem can now be modeled with more confidence and thephysiological and clinical implications of pump-conduit be-havior can be broadened.
Computational Model Caveats
This work predicted the function of lymphangions based onknown properties of lymphangions and the principles govern-ing fluid motion. Although this computational model is basedon fundamental principles, several simplifications were made.First, the characterization of lymphangions with the time-varying elastance concept was convenient and provided asimple representation of the contraction of the lymphangionwall (54). The use of experimentally derived time-varyingelastance parameters Emax and Vo is not particularly sensitiveto measurement error. Rather, time-varying elastance, usedextensively to model the function of ventricles, has been
R1516 PUMP-CONDUIT BEHAVIOR
AJP-Regul Integr Comp Physiol • VOL 292 • APRIL 2007 • www.ajpregu.org
on April 5, 2007
ajpregu.physiology.orgD
ownloaded from

criticized for its simplicity. This approximation to describelymphangions in particular neglects the high degree of tempo-ral variability in the contractility and contraction frequencyfound in intact lymphatic vessels (25). This simple description,furthermore, does not fully account for “pump failure” athigher pressures, as described by McHale and Roddie (28), orthe flow-induced dilation described by Gashev et al. (15). Bothbehaviors are important aspects that affect lymphangion con-tractility but do not determine whether contraction itself in-creases or decreases total lymph flow. In fact, this simplicityhelps illustrate the effects of axial pressure gradient without theconfounding effects of these complications. Second, the com-putational model used simplified equations to describe the fluidmotion. Following the derivation of Noordergraaf (35), thesesimplifications notably neglect second-order terms and fluidmomentum in the radial direction. These descriptions werederived by Reddy and Kesavan (39) from a simplification ofthe Navier-Stokes equations, which is justified for vessels withlow Reynolds numbers. The results are consistent with the invitro measurements that were performed to validate the results.Figure 7 presents information from the computational modelthat is particularly difficult to obtain experimentally. We main-tain that when computational models based on fundamentalprinciples result in the same basic behavior observed in actualphysiological systems, the interpretation of the data is moresecure.
GRANTS
Portions of this work were supported by National Heart, Lung, and BloodInstitute Grants HL-070608 (to C. M. Quick) and HL-07688 (to D. C. Zawiejaand A. A. Gashev), the Texas Advanced Research Program (to R. H. Stewart),and American Heart Association Grant 0365127Y (to R. H. Stewart).
REFERENCES
1. Bates DO, Levick JR, Mortimer PS. Starling pressures in the human armand their alteration in postmastectomy oedema. J Physiol 477: 355–363,1994.
2. Benoit JN, Zawieja DC, Goodman AH, Granger HJ. Characterizationof intact mesenteric lymphatic pump and its responsiveness to acuteedemagenic stress. Am J Physiol Heart Circ Physiol 257: H2059–H2069,1989.
3. Berger DS, Li JK, Noordergraaf A. Differential effects of wave reflec-tions and peripheral resistance on aortic blood pressure: a model-basedstudy. Am J Physiol Heart Circ Physiol 266: H1626–H1642, 1994.
4. Drake RE, Anwar Z, Kee S, Gabel JC. Intestinal lymphatic pressureincreases during intravenous infusions in awake sheep. Am J Physiol
Regul Integr Comp Physiol 265: R703–R705, 1993.5. Drake RE, Gabel JC. Effect of outflow pressure on intestinal lymph flow
in unanesthetized sheep. Am J Physiol Regul Integr Comp Physiol 260:R668–R671, 1991.
6. Eisenhoffer J, Elias RM, Johnston MG. Effect of outflow pressure onlymphatic pumping in vitro. Am J Physiol Regul Integr Comp Physiol 265:R97–R102, 1993.
7. Eisenhoffer J, Kagal A, Klein T, Johnston MG. Importance of valvesand lymphangion contractions in determining pressure gradients in iso-lated lymphatics exposed to elevations in outflow pressure. Microvasc Res
49: 97–110, 1995.8. Elias RM, Johnston MG. Modulation of fluid pumping in isolated bovine
mesenteric lymphatics by a thromboxane/endoperoxide analogue. Prosta-
glandins 36: 97–106, 1988.9. Elias RM, Johnston MG, Hayashi A, Nelson W. Decreased lymphatic
pumping after intravenous endotoxin administration in sheep. Am J
Physiol Heart Circ Physiol 253: H1349–H1357, 1987.10. Ferguson MK, DeFilippi VJ, Reeder LB. Characterization of contractile
properties of porcine mesenteric and tracheobronchial lymphatic smoothmuscle. Lymphology 27: 71–81, 1994.
11. Fitchett DH. LV-arterial coupling: interactive model to predict effect ofwave reflections on LV energetics. Am J Physiol Heart Circ Physiol 261:H1026–H1033, 1991.
12. Gashev AA. The mechanism of the formation of a reverse fluid filling inthe lymphangions [in Russian]. Fiziol Zh SSSR Im I M Sechenova 77:63–69, 1991.
13. Gashev AA. The pump function of the lymphangion and the effect on itof different hydrostatic conditions [in Russian]. Fiziol Zh SSSR Im I M
Sechenova 75: 1737–1743, 1989.14. Gashev AA, Davis MJ, Delp MD, Zawieja DC. Regional variations of
contractile activity in isolated rat lymphatics. Microcirculation 11: 477–492, 2004.
15. Gashev AA, Davis MJ, Zawieja DC. Inhibition of the active lymph pumpby flow in rat mesenteric lymphatics and thoracic duct. J Physiol 540:1023–1037, 2002.
16. Gashev AA, Orlov RS, Borisov AV, Kliuchin’ski T, Andreevskaia
MV, Bubnova NA, Borisova RP, Andreev Iu A, Erofeev NP, Priklon-
skaia EG. The mechanisms of lymphangion interaction in the process oflymph movement [in Russian]. Fiziol Zh SSSR Im I M Sechenova 76:1489–1508, 1990.
17. Gashev AA, Zawieja DC. Physiology of human lymphatic contractility:a historical perspective. Lymphology 34: 124–134, 2001.
18. Granger DN, Korthuis RJ, Kvietys PR, Tso P. Intestinal microvascularexchange during lipid absorption. Am J Physiol Gastrointest Liver Physiol
255: G690–G695, 1988.19. Hanley CA, Elias RM, Movat HZ, Johnston MG. Suppression of fluid
pumping in isolated bovine mesenteric lymphatics by interleukin-1: inter-action with prostaglandin E2. Microvasc Res 37:218–229, 1989.
20. Ikomi F, Schmid-Schonbein GW. Lymph pump mechanics in the rabbithind leg. Am J Physiol Heart Circ Physiol 271: H173–H183, 1996.
21. Jager GN, Westerhof N, Noordergraaf A. Oscillatory flow impedance inelectrical analog of arterial system: representation of sleeve effect andnon-Newtonian properties of blood. Circ Res 16: 121–133, 1965.
22. Johnston MG, Gordon JL. Regulation of lymphatic contractility byarachidonate metabolites. Nature 293: 294–297, 1981.
23. Laine GA, Allen SJ, Katz J, Gabel JC, Drake RE. Outflow pressurereduces lymph flow rate from various tissues. Microvasc Res 33: 135–142,1987.
24. Laine GA, Hall JT, Laine SH, Granger J. Transsinusoidal fluid dynam-ics in canine liver during venous hypertension. Circ Res 45:317–323,1979.
25. Li B, Silver I, Szalai JP, Johnston MG. Pressure-volume relationships insheep mesenteric lymphatic vessels in situ: response to hypovolemia.Microvasc Res 56: 127–138, 1998.
26. Lobov GI, Orlov RS. Electrical and contractile activity of the lym-phangions of the mesenteric lymphatic vessels [in Russian]. Fiziol Zh
SSSR Im I M Sechenova 69: 1614–1620, 1983.27. McHale NG, Meharg MK. Co-ordination of pumping in isolated bovine
lymphatic vessels. J Physiol 450: 503–512, 1992.28. McHale NG, Roddie IC. The effect of transmural pressure on pumping
activity in isolated bovine lymphatic vessels. J Physiol 261: 255–269,1976.
29. McHale NG, Roddie IC. The effects of catecholamines on pumpingactivity in isolated bovine mesenteric lymphatics. J Physiol 338:527–536,1983.
30. McGeown JG, McHale NG, Thornbury KD. The role of externalcompression and movement in lymph propulsion in the sheep hind limb.J Physiol 387:83–93, 1987.
31. Moseley AL, Carati CJ, Piller NB. A systematic review of commonconservative therapies for arm lymphoedema secondary to breast cancertreatment. Ann Oncol. In press.
32. Mislin H. Active contractility of the lymphangion and coordination oflymphangion chains. Experientia 32: 820–822, 1976.
33. Mortillaro NA, Taylor AE. Interstitial fluid pressure of ileum measuredfrom chronically implanted polyethylene capsules. Am J Physiol Heart
Circ Physiol 257: H62–H69, 1989.34. Nedrebo T, Berg A, Reed RK. Effect of tumor necrosis factor- , IL-1�,
and IL-6 on interstitial fluid pressure in rat skin. Am J Physiol Heart Circ
Physiol 277: H1857–H1862, 1999.35. Noordergraaf A. Cardiovascular System Dynamics. New York: Aca-
demic, 1978.36. Ohhashi T, Azuma T, Sakaguchi M. Active and passive mechanical
characteristics of bovine mesenteric lymphatics. Am J Physiol Heart Circ
Physiol 239: H88–H95, 1980.
R1517PUMP-CONDUIT BEHAVIOR
AJP-Regul Integr Comp Physiol • VOL 292 • APRIL 2007 • www.ajpregu.org
on April 5, 2007
ajpregu.physiology.orgD
ownloaded from

37. Palladino J, Mulier J, Noordergraaf A. Defining ventricular elastance.20th Annual Conference IEEE/EMBS Hong Kong, 1998.
38. Reed RK, Rodt SA. Increased negativity of interstitial fluid pressureduring the onset stage of inflammatory edema in rat skin. Am J Physiol
Heart Circ Physiol 260: H1985–H1991, 1991.39. Reddy N, Kesavan S. Low Reynolds number liquid propulsion in con-
tracting tubular segments connected through valves. Math Comput Model
12: 839–884, 1989.40. Reddy NP, Krouskop TA, Newell PH Jr. A computer model of the
lymphatic system. Comput Biol Med 7: 181–197, 1977.41. Schmid-Schonbein GW. Microlymphatics and lymph flow. Physiol Rev
70: 987–1028, 1990.42. Shirasawa Y, Benoit JN. Stretch-induced calcium sensitization of rat
lymphatic smooth muscle. Am J Physiol Heart Circ Physiol 285: H2573–H2577, 2003.
43. Shirasawa Y, Ikomi F, Ohhashi T. Physiological roles of endogenousnitric oxide in lymphatic pump activity of rat mesentery in vivo. Am J
Physiol Gastrointest Liver Physiol 278: G551–G556, 2000.44. Shroff SG, Janicki JS, Weber KT. Evidence and quantitation of left
ventricular systolic resistance. Am J Physiol Heart Circ Physiol 249:H358–H370, 1985.
45. Shroff SG, Naegelen D, Clark WA. Relation between left ventricularsystolic resistance and contractile rate processes. Am J Physiol Heart Circ
Physiol 258: H381–H394, 1990.
46. Stewart RH, Laine GA. Flow in lymphatic networks: interaction betweenhepatic and intestinal lymph vessels. Microcirculation 8: 221–227, 2001.
47. Suga H, Sagawa K. Instantaneous pressure-volume relationships and theirratio in the excised, supported canine left ventricle. Circ Res 35: 117–126,1974.
48. Suga H, Sagawa K. Mathematical interrelationship between instanta-neous ventricular pressure-volume ratio and myocardial force-velocityrelation. Ann Biomed Eng 1: 160–181, 1972.
49. Suga H, Sagawa K, Shoukas AA. Load independence of the instanta-neous pressure-volume ratio of the canine left ventricle and effects ofepinephrine and heart rate on the ratio. Circ Res 32: 314–322, 1973.
50. Sunagawa K, Sagawa K. Models of ventricular contraction based ontime-varying elastance. Crit Rev Biomed Eng 7: 193–228, 1982.
51. Westerhof N, Noordergraaf A. Arterial viscoelasticity: a generalizedmodel. Effect on input impedance and wave travel in the systematic tree.J Biomech 3: 357–379, 1970.
52. Womersley J. An elastic tube theory of pulse transmission and oscillatoryflow in mammalian arteries. WADC Technical Report TR-56–614, 1957.
53. Zawieja DC, Davis KL, Schuster R, Hinds WM, Granger HJ. Distri-bution, propagation, and coordination of contractile activity in lymphatics.Am J Physiol Heart Circ Physiol 264: H1283–H1291, 1993.
54. Zawieja DC, Benoit J, Granger HJ. Coordination of lymphatic pumping.A theoretical model. Fed Proc 46: 1546, 1987.
R1518 PUMP-CONDUIT BEHAVIOR
AJP-Regul Integr Comp Physiol • VOL 292 • APRIL 2007 • www.ajpregu.org
on April 5, 2007
ajpregu.physiology.orgD
ownloaded from