interferon-mediated enhancement of metastasis. are mhc antigens involved?
TRANSCRIPT

CLIN. EXPL. METASTASIS, 1987, VOL. 5, NO. 4, 2 7 7 - 2 8 7
Interferon-mediated e n h a n c e m e n t of metastasis . Are MHC ant igens involved?
P I E R - L U I G I L O L L I N I , C A R L A D E G I O V A N N I , B R U N E L L A D E L RE, G I O R D A N O N I C O L E T T I , G I O R G I O P R O D I and P A T R I Z I A N A N N I
Istituto di Cancerologia, Universitfi di Bologna, Viale Filopanti 22, 1-40126 Bologna, Italy
(Received 19 June 1986; accepted 8 December 1986)
The relationship between major histocompatibility complex (MHC) antigens and metastasis was investigated on B16 melanoma variants. B16 cell lines express low amounts of murine M H C (H-2) antigens. A high expression can be induced in line B16-A by in vitro treatment with immune interferon ( IFN-gamma) or by in vivo transplant in allogeneic mice. The increase of H-2 antigens correlated with an enhancement of lung colonization in young syngeneic mice. The higher metastatic capacity of B16-A cells with induced high levels of H-2 antigens was observed also in adult mice and in young mice pretreated with cyclophos- phamide. These results were confirmed investigating the behaviour of a mutant B16 clone (B78H1) which was selectively resistant to the H-2-inducing action of IFN-gamma: lung colonization ability was not increased by I F N pretreatment. The study of variants derived from individual B16-A lung colonies revealed a wide range of H-2 levels. Variants with a low expression had a low colonization ability; one out of two variants with a high H-2 expression also was poorly colonizing. IFN-gamma-media ted H-2 expression appeared to act as an enhan- cer, rather than a determinant of B16 metastatic capacity.
I n t r o d u c t i o n T h e role p l ayed by m a j o r h i s t o c o m p a t i b i l i t y c o m p l e x ( M H C ) an t igens in the
pa thogenes i s of cancer me tas t a s i s has no t ye t been p rec i se ly clarif ied. Severa l types of me ta s t a t i c h u m a n t u m o r s show a l te ra t ions (usua l ly a decrease) of M H C class I an t igens expres s ion [ 1 , 2 , 3 , 8 , 13, 16], b u t no genera l co r re l a t ion has ye t been p r o p o s e d wi th me ta s t a t i c sp read or p rognos i s [22]. T h e da ta o b t a i n e d so far wi th rou t ine t u m o r s [4, 6, 24] c o n c o r d a n t l y show tha t the t r e a t m e n t of me tas t a t i c cells wi th in t e r f e rons induces a c o n c o m i t a n t increase of H - 2 expres s ion and of me ta s t a t i c po ten t i a l .
T w o di f ferent exp l ana t ions of th is p h e n o m e n o n have been p r o p o s e d . T h e da ta o b t a i n e d by F e l d m a n and c o - w o r k e r s wi th Lewis lung c a r c i n o m a [4] and T 1 0 f i b ro sa r coma [11] ind ica te tha t d i f ferent class I reg ions cou ld have an tagon i s t i c effects on the con t ro l of me tas t a s i s by T l y m p h o c y t e s . In cont ras t , T a n i g u c h i et al. [24], s t u d y i n g B16 m e l a n o m a , f o u n d an inverse r e l a t ionsh ip be tw e e n g loba l H - 2 expres s ion and me tas t a s i s on the one side and N K sens i t iv i ty of t u m o u r cells on the o ther .
W e have r ecen t ly cha rac t e r i zed the expres s ion of H - 2 an t igens in va r ious B16 cell l ines [14] and we have shown tha t the r e d u c t i o n of H - 2 expres s ion o b s e r v e d in vitro can be reve r sed b y i n t e r f e r o n - g a m m a ( I F N - g a m m a ) t r e a t m e n t or by in v ivo passage in a l logeneic mice [12]. W e p r e sen t here da ta on the inf luence of H - 2 an t igens on the

278 P.-L. Lollini et al.
metastatic ability of several B16 variants differing f rom each other in H-2 expression or interferon sensitivity. The i r metastatic potential has been compared using both normal and NK-depres sed mice.
M a t e r i a l s a n d m e t h o d s
Mice C57BL/6NCr lBR (C57BL/6) and B A L B / c A n N C r l B R (BALB/c) male mice
were purchased f rom Charles River, Calco, Italy. Mice used for lung colonization assay were either 8-12 weeks old (hereafter referred to as young mice) or 20-25 weeks old (referred to as adult mice) or young mice treated with a single intraperitoneal administrat ion of cyclophosphamide (240mg/kg, Asta Werke, Bielefeld, F .R.G.) four days before the injection of B16 cells (referred to as young CY-treated mice).
Cells B16-A cell line has been previously characterized [12, 14]. Clone B78H1 [7] was
obtained through the courtesy of Dr T. Boon, Ludwig Institute, Brussels, Belgium.
Inter f erons
Natural mouse I F N - g a m m a was a kind gift of S. Landolfo, Ist i tuto di Microbiologia, Tur in , Italy. Recombinant I F N - g a m m a was obtained through the courtesy of G. R. Adolf, Ernst-B6hringer Insti tut , Wien, Austria. The antiprolifer- ative activity of I F N was measured adding increasing concentrations of recombinant I F N - g a m m a to B16 cells seeded 24h before into Falcon (Oxnard, U.S.A.) T25 flasks; each day one control and one IFN- t r ea t ed flask were harvested and the cell yield determined.
In vitro and in vivo induction of H-2 antigens In vitro, cells were treated with 50 U/ml of I F N - g a m m a for 24 h (except when
otherwise stated), then processed for quantitative absorption or for lung colonization assay. T h e protocol used to induce H-2 expression in B16 cells by means of in vivo t ransplant has been described [12]; briefly, allogeneic BALB/c mice were inoculated subcutaneously with 10 6 cells; one month later the ensuing tumour was dissociated mechanically and enzymatically, and the resulting cells were cultured in vitro for 2-4 passages to get rid of normal cells.
Absorption assay Quantitat ive in vitro absorption of complement -dependent cytotoxicity was
carried out as described [12, 14]. The following ant i -H-2 reagents were used: H-142-23, monoclonal a n t i - H - 2 K b, working dilution 1 : 55-1 : 60, purchased f rom Biotest, Milan, Italy, and E-2, monospecific an t i -H-2D b, working dilution 1 : 10, obtained from the N I H , Bethesda, M D , U.S.A. Cytotoxic activity against C57BL/6 lymph-node cells was assessed in a t rypan blue dye exclusion assay.
Lung colonies C57BL/6 mice were inoculated with 2"5 x 10"-105 cells in the tail vein (i.v.). Mice
were killed 21 days later and their lungs were removed and fixed. T h e lungs of mice

IFN-mediated enhancement of metastasis 279
inoculated with the amelanotic clone B78H1 were treated as previously described [15] to detect white nodules. Colonies were scored with the aid of a dissecting microscope. In some experiments individual lung colonies were punched out with the tip of a Pasteur pipette and briefly cultured in vitro (2-4 passages); cell cultures were then tested for H-2 expression and colonization ability.
Statistical analysis The non-parametric Wilcoxon's rank sum test was used.
R e s u l t s The B16-A cell line used in this work was an unselected line directly established
from a B16 tumor [14], and had a low metastatic efficiency in syngeneic im- munocompetent hosts. To obtain a better evaluation of the relationship between H-2 antigens and lung colonization, we used (in addition to normal young (C57BL/6 mice) young mice pretreated with cyclophosphamide according to a protocol which is reported to reduce the natural killer activity of hosts and to enhance B16 colonization [10, 18, 27], and adult mice, in which we have previously found a number of lung colonies higher than in young mice. This enhancement of lung colonization in adult mice could be mediated by an impairment of the natural response as well: the levels of natural cytotoxity detected in the spleens of adult C57BL/6 mice against sensitive adherent target cells were significantly lower than those of young mice (data not shown).
Induction of H-2 antigens and of high colonization ability in wild-type B16 cells We have previously shown that B16 cells cultured in vitro for more than 10
passages have a very low expression of H - 2 K b and a reduced amount of O b antigens [14], and that a higher expression can be reinduced by in vivo passage in allogeneic hosts [12]. Table 1 shows that, after transplant in allogeneic mice, B16 cells possess a lung-colonizing capacity higher than that of cells passed in syngeneic hosts. Concordant results were obtained using young or adult mice.
A high H-2 expression can be induced also by in vitro treatment with I F N - gamma [12], thus we studied the colonizing capacity of B16-A cells treated with natural (table 2 (a)) or recombinant (table 2 (b)) IFN-gamma. Again IFN-t rea ted cells, with high H-2 expression, gave more lung colonies, regardless of the age or the CY treatment of the host.
Table 1. Lung c o l o n i z a t i o n capac i ty in s y n g e n e i c C57BL/6 m i c e of B16-A c e l l s after in v i v o transplant in s y n g e n e i c or a l l o g e n e i c host.
Lung coloniest Intermediate Significance
Mice host Incidence Median Range of treatment
Young Syngeneic 7/8 3 0-5 P<0-05 Allogeneic 8/8 12 2-20
Adult Syngeneic 10/11 4 0 11 P<0"01 Allogeneic 7/7 42 10-113
"~C57BL/6 mice were inoculated with l0 s cells i.v.

280 P.-L. Lollini et al.
Table 2. L u n g c o l o n i z a t i o n capac i ty in s y n g e n e i c C57BL/6 m i c e o f B16-A cells treated in vitro w i t h 5 0 U / m l or natural or r e c o m b i n a n t m o u s e I F N - g a m m a .
Lung colonies Cell Cell Significance
Mice dose treatment Incidence Median Range of treatment
(a) Natural IFN-gamma Young 105
Adult 105
None 2/9 0 0-2 P<0-01 IFN-gamma 9/10 14 0-29
None 6/6 12 10 20 P<0"01 1FN-gamma 7/7 79 40 221
(b) Recombinant IFN-gamma Young 2"5 × 104 None 0/10 0 - - P<0"01 IFN-gamma 8/9 6 0-10
5 × 104 None 2/10 0 0-2 P<0-01 IFN-gamma 8/10 16 0~54
Young CY 2'5 × 104 None 7/7 28 3 101 treated IFN-gamma 10/10 61 6 157 n.s.
5 x 104 None 9/9 57 21-120 P<0"01 IFN-gamma 10/10 232 90-376
W h e n graded doses of r ecombinan t I F N - g a m m a were used, a directly p ropor - t ional effect on H-2 expression (figure 1 (a)) and on colonizing capaci ty (table 3) was observed. I t should be noted that, in the absorpt ion exper iment depicted in figure 1 (a), the an t ibody di lut ion was kept at 1 : 40 (instead of the usual 1 : 55-1 : 60) in order to resolve bet ter the effects of h igher I F N doses.
Lack of H-2 induction and of high colonization capacity in clone B78H1 W e have recently obta ined (from T. Boon, L u d w i g Inst i tute, Brussels, Belgium)
a B16 clone (B78H1, originally isolated by Silagi and co-workers [7]) which shows some interest ing features. B78H1 cells do not display any detectable amoun t of H-2 antigens on their surface, and H-2 expression could not be induced with I F N - g a m m a (figure 1 (b) and T. Boon, personal communica t ion) . T o rule out the possibil i ty that the absence of response could be due to an insufficient t rea tment with I F N - g a m m a , a h igher dose (500 U / m l instead of 50 U/ml ) for a longer per iod (96 hr instead of 24 hr) was tested, bu t again no induct ion of H-2 antigens was observed (figure 1 (b)). Similar results were obta ined with I F N - a l p h a + beta (data not shown) or in vivo t ransplant in allogeneic mice (figure 2).
T h e lack of I F N - m e d i a t e d H-2 induct ion in these cells seems to be a rather specific defect, since B78H1 cells are able to respond to the antiproliferative activity of I F N - g a m m a (figure 3). In addit ion, B78H1 cells are sensitive to the antiviral act ion of I F N - g a m m a and possess I F N - g a m m a receptors (S. Landolfo , personal communica t ion) .

100- _ _ _ _
U,
P-
5O
R
IFN-mediated enhancement of metastasis
10(3- . . . . . . . . . . . . ~ -
,'s 1 ~ ~ ABSORBING CELLS x 106 (a)
o. -H-2K D
~_ [] ~_
i i i
2.5 5 10
ABSORBING CELLS x 10 6
100-
50
0
281
,~. -H-2D b
. . . . ~_ ~ ~ ' - - -
J f i
2.5 5 10
ABSORBING CELLS x 10 6 (b)
Figure 1. (a) Residual C'-dependent cytotoxicity of ant i-H-2K b antibody after absorption with B16-A cells either untreated (©) or treated with 0-5 ( • ) , 5 ( • ) , or 50 ( • ) U/ml of recombinant murine IFN-gamma. (b) Residual C'-dependent cytotoxicity of anti-H-2 reagents after absorption with normal C57BL/6 lymph-node cells (triangles), B16-A (circles) or B78H1 (squares). Open symbols: untreated cells; half-closed symbols: cells treated with 50 U/ml of recombinant IFN-gamma for 24 h; closed symbols: cells treated with 500 U/ml of IFN-gamma for 96h. Dashed line represents unabsorbed reagent.
T a b l e 3 . L u n g c o l o n i z a t i o n c a p a c i t y o f B I 6 - A c e l l s t r e a t e d w i t h g r a d e d d o s e s o f
r e c o m b i n a n t I F N - g a m m a .
Lung colonies~
Cell treatment Incidence Median Range
None 11/12 1 0-14 IFN-gamma 0-5 U/ml 8/10 3 0-8 IFN-gamma 5 U/ml 11/11 11 2 25 IFN-gamma 50 U/ml 10/10 50 23-101
t After an injection of 105 cells i.v. into adult C57BL/6 mice.

282 P. -L . Lollini et al.
100 -
U,
50
P=
o~-H-2K b
_ _ _ . . . .
"0
100-
i i i
2,5 5 i0
50
0 O
o~-H-2D b
. . . . ~ .~ .Z . -Z~ - - - I -
2,5 5 i0
ABSORBING CELLS x 106 ABSORBING CELLS x 106
Figure 2. Residual C ' -dependent cytotoxicity of an t i -H-2 reagents after absorption with normal C57BL/6 lymph-node cells (triangles), B16-A (circles) or B78H1 (squares). Open symbols: untreated cells; closed symbols: cells grown for one month iri allogeneic B A L B / c mice then briefly cul tured in vitro. Dashed line represents unabsorbed reagent.
100 - 100-
50 50'
G , , 0 . . , 24 g8 72 5 50 500
HOURS AFTER TREATMENT WITH U/m] OF IFN- ~t
50 U/ml OF IFN- ~-
(o) (b)
Figure 3. Inhibi t ion of in vitro proliferation of B16-A (O) and B78H1 (m) by recombinant I F N - g a m m a . (a) T i m e - c o u r s e of inhibit ion; (b) dose-response of inhibit ion evaluated 72 h after I F N - g a m m a addition.
IFN-mediated enhancement of metastasis 283
We thought that B78H1 could contribute to elucidate whether the modulation of H-2 antigens induced by I F N - g a m m a was the true cause of the increase of the metastatic potential of B16 cells or if it was only a mere side effect of I F N treatment.
The number of lung colonies produced by B78H1 cells was not increased by in vivo transplant in allogeneic mice or by in vitro treatment with IFN-gamma. When 5 x 104 untreated B78H1 cells were injected i.v. into young C57BL/6 mice, the median number of resulting lung colonies was 1 (incidence 6/10, range 0-6); the same dose of cells pretreated with 50U/ml of recombinant I F N - g a m m a produced a median of 3 (incidence 7/10, range 0 5); l0 s untreated B78H1 cells produced a median of 3 colonies (incidence 9/10, range 0-5) and the same number of allogeneically passaged cells gave a median of 2 (incidence 8/10, range 0-4). Concordant findings were obtained using adult hosts treated i.v. with 105 cells: the median numbers of lung colonies obtained with untreated, I F N - g a m m a treated or allogeneically passaged cells were respectively 2 (incidence 7/10, range 0-7), 3 (incidence 7/10, range 0-11) and 3 (incidence 6/10, range 0-49). It should be noted that these results were not due to an intrinsically low metastatic potential of B78H1 cells, since, as with B 16-A, CY pretreatment of mice induced a significant (P < 0"01) increase of lung colonies, but, even in CY-pretreated mice, IFN-gamma-t rea ted B78H1 cells failed to show a colonization significantly higher than untreated cells: 5 x 104 untreated cells produced a median of 16 colonies (incidence 10/10, range 4-90), the same dose of IFN-gamma- t rea ted cells gave a median of 22 colonies (incidence 8/8, range 5-71).
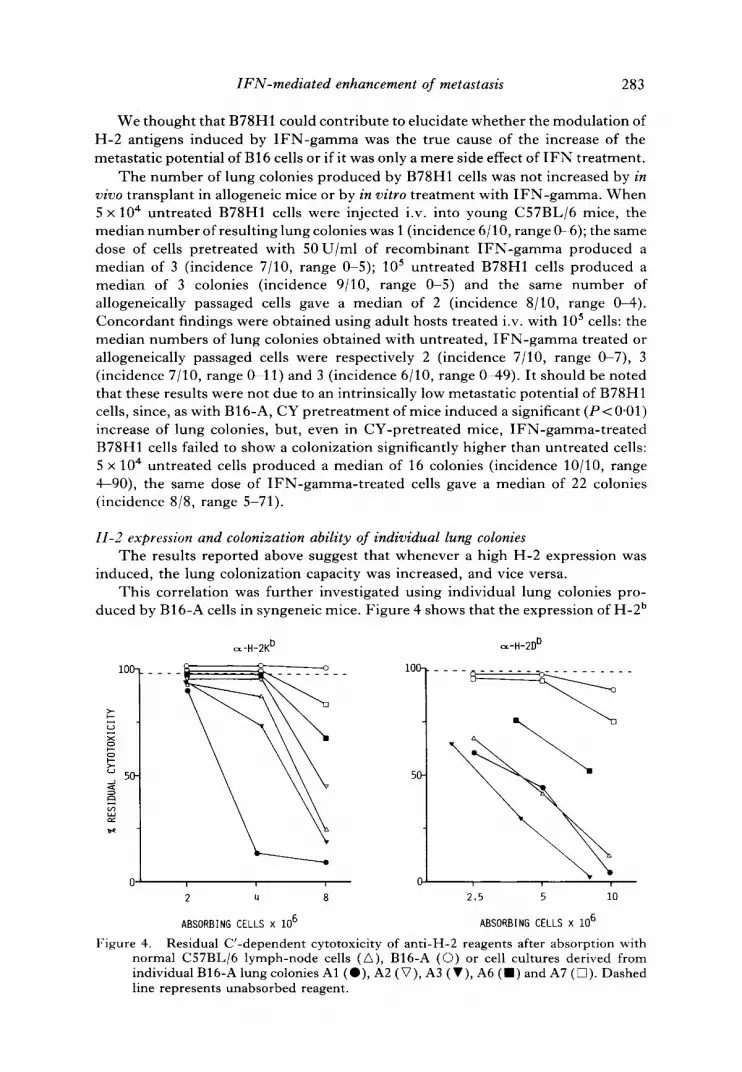
11-2 expression and colonization ability of individual lung colonies The results reported above suggest that whenever a high H-2 expression was
induced, the lung colonization capacity was increased, and vice versa. This correlation was further investigated using individual lung colonies pro-
duced by B16-A cells in syngeneic mice. Figure 4 shows that the expression of H-2 b
o.-H-2K b
c-)
x
50
R
i00- i00-
50
0 u u n 0
2 4 8
ABSORBING CELLS x 106
~-H-2D t)
. . . . ~ . . . . . . - ~ - -
r i r
2.5 5 i0
ABSORBING CELLS x 10 6
Figure 4. Residual C'-dependent cytotoxicity of anti-H-2 reagents after absorption with normal C57BL/6 lymph-node cells (A), B16-A (©) or cell cultures derived from individual B16-A lung colonies A1 (O), A2 (V), A3 (V), A6 ( ' ) and A7 ([~). Dashed line represents unabsorbed reagent.

284 P.-L . LoUini et al.
Table 4. Lung c o l o n i z a t i o n abi l i ty of cells der ived from lung co lon ies .
Lung coloniest
Variant Incidence Median Range
A1 7/10 3 0-37 A3 10/10 103 41-258 A6 8/9 5 0 12 A7 8/8 9 3 22
~'After an injection of 105 cells i.v. into adult C57BL/6 mice.
i n d i v i d u a l B16-A
antigens in cultures derived from individual lung colonies is widely dispersed, ranging from low levels, slightly above the parent cell line, to levels comparable to those obtained after allogeneic transplant of B16-A (see figure 2). It should be noted that the expression of the two class I regions appeared to be coordinated in these variants: the Kb/D b antigens ratio was similar to that of the parent cell line; only variant A1 showed a higher ratio.
We subsequently studied the lung-colonization ability of variants A6, A7 (low H-2 expression), A1 and A3 (high H-2 expression). The results (table 4) indicate that variants with a low H-2 expression do indeed have a low colonization ability, however, also variant A1, which has a high expression of H-2 antigens, produced a low number of lung colonies. Variant A3, like IFN- induced cells, expressed high levels of H-2 antigens and gave rise to a high number of lung nodules.
D i s c u s s i o n The first conclusion which can be drawn from the results presented here is that in
this B16 model there is a positive correlation between IFN-media ted M H C modulation and the metastatic potential. Whenever a high expression of H-2 antigens could be reinduced in B16-A cells, an increase of the number of lung colonies was observed. Conversely, B78H1 cells did not respond to H-2-inducing treatments and their metastatic capacity was unaffected. The results obtained with B16-A are in agreement with the findings recently reported by Taniguchi et al. [24], who used IFN-a lpha and beta to enhance H-2 expression in unselected B16 cells.
However, the data obtained with the B16-A cells do not demonstrate that the action of I F N - g a m m a on lung colonization be mediated by H-2 antigens, but this hypothesis is enforced by the behaviour of B78H1 cells, which responded to some of the I F N actions tested but did not show any H-2 induction.
We cannot obviously exclude that the colonization-enhancing effect of I F N - gamma could be mediated by some unknown mechanism independent of H-2, but from the data presented here it could be hypothesized that this phenomenon is probably related to the antigen-inducing properties of IFN, rather than to its antiviral or antiproliferative effects.
A further possibility would be the expression of altered H-2 glycoproteins after the I F N - g a m m a treatment. We think that this was not the case, since the induced cells reacted with a panel of monoclonal and monospecific an t i -H-2K b or H-2D b reagents directed against different H-2 specificities (data not shown). The function-

IFN-media ted enhancement of metastasis 285
ality of the H-2 antigens expressed by allogeneically grown B16 cells was confirmed by the serological anti-H-2 b response of the hosts (De Giovanni et al., in preparation). The growth of allogeneic B16-A tumors was probably due to an imbalance between the kinetics of tumor growth and that of H-2 re-expression [12]: when the immune system recognizes the allogeneic antigens it is probably too late to reject an already large tumor. It should be noted however that the growth of B16 in allogeneic mice is significantly slower than in syngeneic mice [12, 14].
A point deserving further analysis is the role played by N K cells, and by I F N - mediated resistance to N K cells [9, 25], in determining the control of metastatic spread of B16 cells with different levels of H-2 expression [24]. In fact our findings confirm that B16 cells have a higher metastatic potential in mice with a depressed N K activity [10, 18, 27], but we show here that cells with a high H-2 expression produce significantly more lung colonies even in NK-depressed mice. We are currently studying the metastatic efficiency of our B16 variants in mice with altered T lymphocyte activities.
The involvement of a T cell-mediated response, which can be either stimulated or suppressed by different M H C antigens, has been demonstrated in other studies on metastasis: for example, H L A - D R positive human metastatic melanoma cells, which are able to suppress the IL-2-dependent generation of autologous cytotoxic lymphocytes, augment both DR expression and suppressive activity after I F N - gamma treatment [19].
A parallel stimulation of class I antigens and of metastatic potential by I F N - alpha + beta has been described [4] for 3LL cells. In that model the effects of I FNs were referred to a higher expression of H-2D b antigens (which could exert suppressive effects) than of K b. Unfortunately, the B16-A cell line does not seem to be suitable for studies of the relative influence of the two class I regions, since a coordinated increase of K and D antigens was observed after in vivo transplant in allogeneic hosts, I F N - g a m m a treatment [12] or after lung colonization. We observed an alteration of the K /D ratio in our model only among the colony-derived variants shown in figure 4:A1 cells display a K /D ratio (as calculated from zero-order probit regressions of the data in figure 4) higher than that of A3 cells. However the correlation between the K/D ratio and the colonization potential of these cells does not hold, since the low-colonizing A6 and A7 variants show a K/D ratio similar to or lower than that of the high-colonizing A3 variant. Clearly a more detailed analysis is required, but the study of H-2 expression and metastatic potential in clones, which allowed excellent studies in the 3LL model [4], has been discouraged by the fact that B16 clones have an unstable metastatic phenotype [17, 21]. We think that the most straightforward way of obtaining information on the relative importance of K and D regions would be to transform B16 cells with the relevant genes [26]; B78H1 cells appear particularly suitable for these kinds of studies [7].
In any case, it should be borne in mind that the metastatic spread is a multistep process [20, 23] and that M H C antigens can probably only modulate, rather than determine, the metastatic capacity of cells which already possess a complete and functional 'metastatic machinery' . We interpret in this way the results obtained with individual lung colonies: one of the colony-derived variants (A1) was indeed low colonizing despite its high H-2 expression. It is possible that it had an intrinsically low colonization capacity, determined for example by a defect in some essential step of the metastatic process. If this was the case, the relative advantage conferred by a high H-2 expression would probably have been meaningless for these cells.

286 P.-L. Lollini et al.
Finally, it remains to be determined why a high M H C expression favours metastatic spread. In the B16 model it has been demonstrated that cells selected in vitro for their resistance to cytotoxic lymphocytes have a decreased metastatic capacity [5]. Cells which interact poorly with the immune system, as is possibly the case with MHC-negat ive cells, could have a reduced probability of forming emboli with lymphocytes, and thus of organ arrest [5]. We are currently investigating this hypothesis by means of mice with immune system defects, either natural or induced.
The conclusions emerging from our, as well as other, studies on the possible adverse effects of IFN-gamma- induced M H C expression on the metastatic spread of tumors clearly cast a shadow on the therapeutic approaches involving the use of this molecule or the stimulation of immunologic reactions during which I F N release takes place.
Acknowledgments The authors thank Giorgio Parmiani for helpful discussions and Mr A.
Lorenzoni and Mrs M. Bertarini Soprani for their skilful technical assistance. This work was supported by a grant of the Italian National Research Council, Special Project 'Oncology' , contract 85.02268.44 and by Associazione Italiana per la Ricerca sul Cancro, Milan, Italy. P.-L. Lollini is in receipt of a PhD studentship (Dottorato di Ricerca in Oncologia) from Ministero della Pubblica Istruzione, Italy; B. Del Re is in receipt of a fellowship from Associazione Italiana per la Ricerca sul Cancro, Milan, Italy; G. Nicoletti is in receipt of a training grant from the Italian National Research Council.
References [1] ALBINO, A. P., LLOYD, K. O., HOUGHTON, A. N., OETTGEN, H. F., and OLD, L. J., 1981,
Heterogeneity in surface antigen and glycoprotein expression of cell lines derived from different melanoma metastases of the same patient. Journal of Experimental Medicine, 154, 1764-1778.
[2] BHAN, A. K., and DESMARAIS, C. L., 1983, Immunohistologic characterization of major histocompatibility antigens and inflammatory cellular infiltrate in human breast cancer. Journal of the National Cancer Institute, 71, 507-516.
[3] DOYLE, A., MARTIN, W. J., FUNA, K., GAZDAR, A., CARNEY, D., MARTIN, S. E., LINNOILA, I., CUTTITTA, F., MULSHINE, J., BUNN, P., and MINNA, J., 1985, Markedly decreased expression of class I histocompatibility antigens, protein, and mRNA in human small-cell lung cancer. Journal of Experimental Medicine, 161, 1135-1151.
[4] EISENBACH, L., SEGAL, S., and FELDMAN, M., 1983, MHC imbalance and metastatic spread in Lewis lung carcinoma. International Journal of Cancer, 32, 113-120.
[5] FIDLER, I. J., GERSTEN, D. M., and BUDMEN, M. B., 1976, Characterization in vivo and in vitro of tumor cells selected for resistance to syngeneic lymphocyte-mediated cytotoxicity. Cancer Research, 36, 3160-3165.
[6] GOODENOW, R. S., VOCEL, J. M., and LINSK, R. L., 1985, Histocompatibility antigens on murine tumors. Science, 230, 777-783.
[7] GaAF, L. H., KAPLAN, P., and SILAGI, S., 1984, Efficient DNA-mediated transfer of selectable genes and unselected sequences into differentiated and undifferentiated mouse melanoma clones. Somatic Cell and Molecular Genetics, 10, 139-151.
[8] GREINER, J. W., HORAN HAND, P., NOGUCHI, P., FISHER, P. B., PETSKA, S., and SCHLOM, J., 1984, Enhanced expression of surface tumor-associated antigens on human breast and colon tumor cells after recombinant human leukocyte alfa-interferon treatment. Cancer Research, 44, 3208-3214.
[9] GRONBERG, A., KIESSLING, R., MASUCCI, G., GUEVARA, L. A., ERIKSSON, E., and KLEIN, G., 1983, Gamma-interferon (IFN-gamma) produced during effector and target interactions renders target cells less susceptible to NK-cell-mediated lysis. International Journal of Cancer, 32, 609-616.

IFN-mediated enhancement of metastasis 287
[10] HANNA, N., and BURTON, R. C., 1981, Definitive evidence that natural killer (NK) cells inhibit experimental tumor metastasis in vivo. Journal of Immunology, 127, 1754-1758.
[11] KATZAV, S., DE BAETSELIER, P., TARTAKOVSKY, B., FELDMAN, M., and SEGAL, S., 1983, Alterations in major histocompatibili ty complex phenotypes of mouse cloned T10 sarcoma cells: association with shifts from nonmetastatic to metastatic cells. Journal of the National Cancer Institute, 71, 317-324.
[12] LOLLINI, P.-L., COLOMBO, M. P., DE GIOVANNI, C., NICOLETTI, G., PARMIANI, G., PRODI, G., and NANNI, P., 1985, In vivo reexpression of H-2 antigens in B16 melanoma cells. Experimental and Clinical Immunogenetics, 2, 14-23.
[13] MACLEAN, G. D., SEEHAFER, J., SHAW, A. R. E., KIERAN, M. W., and LONGENECKER, B. M., 1982, Antigeneic heterogeneity of human colorectal cancer cell lines analyzed by a panel of monoclonal antibodies. I. Heterogeneous expression of Ia-like and HLA-l ike antigenic determinants. Journal of the National Cancer Institute, 69, 357 364.
[14] NANNI, P., COLOMBO, M. P., DE GIOVANNI, C., LOLLINI, P.-L., NICOLETTI, G., PARMIANI, G., and PBODI, G., 1983, Impaired H-2 expression in B16 melanoma variants. Journal of Immunogenetics, 10, 361-370.
[15] NANNI, P., DE GIOVANNI, C., LOLLINI, P.-L., NICOLETTI, G., and PRODI, G., 1983, TS/A: a new metastasizing cell line from a BALB/c spontaneous mammary adenocar- cinoma. Clinical and Experimental Metastasis, 1,373-380.
[16] NATALI, P. G., VIORA, M., NICOTRA, M. R., GIACOMINI, P., BIGOTTI, A., and FERRONE, S., 1983, Antigenic heterogeneity of skin tumors of nonmelanocyte origin: analysis with monoclonal antibodies to tumor-associated antigens and to histocompati- bility antigens. Journal of the National Cancer Institute, 71,439-447.
[17] NICOLSON, G. L., 1984, Tumor progression, oncogenes and the evolution of metastatic phenotypic diversity. Clinical and Experimental Metastasis, 2, 85-105.
[18] NISmMURA, J., MITSUI, K., TANAKA, Y., YAMAMOTO, R., SUHARA, Y., and ISHITSUKA, H., 1986, Restoration by recombinant interferon alpha A/D of host defense systems against tumor in immunosuppressed mice. Clinical and Experimental Meta- stasis, 4, 35~-4.
[19] PARMIANI, G., FOSSATI, G., TARAMELLI, D., ANICHINI, A., BALSAm, A., GAMBACORTI- PASSERINI, C., SCIORELLI, G., and CASCINELLI, N., 1985, Autologous cellular immune response to pr imary and metastatic human melanomas and its regulation by DR antigens expressed on tumor cells. Cancer and Metastasis Reviews, 4, 7-26.
[20] POSTE, G., and FIDLER, I. J., 1980, The pathogenesis of cancer metastasis. Nature, 283, 139 146.
[21] POSTE, G., DOLL, J., and FIDLER, I. J., 1981, Interactions among clonal subpopulations affect stability of the metastatic phenotype in polyclonal populations of B16 melanoma cells. Proceedings of the National Academy of Sciences, U.S.A., 78, 6226-6230.
[22] RUITER, D. J., VAN DUINEN, S. G., BR6CXER, E. B., SCnRIER, P. I., WELVAART, K., and FEBRONE, S., 1985, Low level of class I or high level of class II major histocompatibility complex antigens in metastatic melanoma is associated with a high grade of malignancy. Proceedings of the American Association for Cancer Research, 26, 315.
[23] SCHIRRMACHEB, V., 1985, Cancer metastasis: experimental approaches, theoretical concepts, and impact for treatment strategies. Advances in Cancer Research, 43, 1-73.
[24] TANICUCHI, K., KXRRE, K., and KLEIN, G., 1985, Lung colonization and metastasis by disseminated B16 melanoma cells: H-2 associated control at the level of the host and the tumor cell. International Journal of Cancer, 36, 503-510.
[25] TmNCHIERI, G., and SANTOLI, D., 1978, Anti-viral activity induced by culturing lymphocytes with tumor-derived or virus-transformed cells. Enhancement of human natural killer cell activity by interferon and antagonistic inhibition of susceptibility of target cells to lysis. Journal of Experimental Medicine, 147, 1314--1333.
[26] WALLtCH, R., BULBUC, N., HXMMERLING, G. J., KATZAV, S., SEGAL, S., and FELDMAN, M., 1985, Abrogation of metastatic properties of tumor cells by de novo expression of H-2K antigens following H-2 gene transfection. Nature, 315, 301 305.
[27] WILTROUT, R. H., HEBBERMAN, R. B., ZHANG, S.-R., CHIBIGOS, M. A., ORTALDO, J. R., GBEEN, K. M. JB., and TALMADGE, J. E., 1985, Role of organ-associated N K cells in decreased formation of experimental metastases in lung and liver. Journal of Immunology, 134, 42674275.