immunosuppressive properties of purified immune t-interferon
TRANSCRIPT

CELLULAR IMMUNOLOGY 54, 58-69 (1980)
Immunosuppressive Properties of Purified Immune T-Interferon
MIGUEL A. LUCERO, JUANA WIETZERBIN, SIMON STEFANOS, CLAUDE BILLARDON, ERNESTO FALCOFF, AND WOLF H. FRIDMAN*
lnsiitut Curie, Section de Biologic, FRA. No. 22, 26, rue d’Ulm. 75231 Puris. Ckdex 05, France. and *Lnhoratorir d’lmmunologie Cellulaire, IRSC, Villyjuif, France
Received Mav IS, 1979
Purified T-interferon (specific activity l-2 x 10 units/m@, induced by PHA, inhibits antibody synthesis in vitro to sheep red blood cells (SRBC). Upon fractionation on different chromatographic sorbents (Con A-Sepharose, Blue Sepharose CL-68), the inhibitory activity could not be dissociated from antiviral activity. The kinetics of action of T-interferon on antibody synthesis show that it interferes with the early events of lymphocyte activation. Inhibition was maximum when T-interferon was added 6 hr before or together with antigen but significant effect was still obtained when T interferon was added up to 24-48 hr after antigens. These kinetics were similar to those obtained with viral interferon. Pretreatment of T-interferon for 24 hr at pH 2 destroyed both the antiviral and immunosuppressive activities. A large increase in the number of anti-SRBC plaque-forming cells was observed when crude T-interferon preparation was added 24-48 hr after SRBC. However, purified T-interferon did not display any enhancing activity at any time of addition and always inhibited the plaque-forming cell response to SRBC. When titrated on the basis of antiviral activity no quantitative difference in the inhibition of antibody synthesis could be found between purified T-interferon and viral interferon.
INTRODUCTION
Classically, interferons are considered to be antiviral substances, synthesized by somatic cells in response to viral infection. In addition to its role as a defense against viruses a large body of evidence has been obtained during the last few years, showing that viral-induced, also called type I interferon, has a multiplicity of effects on the immune system both in vivo and in vitro (1). For instance, viral interferon has been shown to inhibit DNA synthesis in lymphocytes stimulated by T- or B-cell mitogens (2,3) or allogenic cells (2) or the in viva and in vitro antibody responses to a variety of antigens (4-6). Recently, interferon has also been shown to be produced during stimulation of natural killer (NK) cells by tumor cells (7,8) and to be a potent inducer of NK cells when added to lymphocytes cultures (8).
Another type of interferon called immune, or type II interferon, is synthesized during the course of immune reactions such as a mixed leukocyte culture (MLC) (9) or antigen (lo- 12) and mitogen stimulation of T and B cells (12- 14). Among immune interferons, B- and T-dependent mitogens stimulate the production of dif- ferent types of interterons (10, 14) in the mouse. Interferons induced by B-cell mitogens are antigenically related to viral interferon (type I) while interferon induced by T-cell mitogens-such as PHA-are unstable at pH 2 and antigenically
58
0008.8749/80/110058-12$02.00/O Copyright 0 1980 by Academic Press, Inc. All rights of reproduction in any form reserved

IMMUNOSUPPRESSIVE PROPERTIES OF T-INTERFERON 59
different (10, 12-15). PHA-induced (T-type) interferon is probably similar to interferon produced in MLC (9) and to interferon induced by antigenic stimulation, e.g., interferon induced by injecting tuberculin in BCG-sensitized mice (IO).
During the last few years, a great amount of data have accumulated on physicochemical properties, purification, mechanism of action, and immunologi- cal properties of virus-induced interferons (1, 16- 18) while no studies have been conducted on the properties of immune interferons. The interest in these interferons has been recently accelerated by the findings of Johnson and Baron (19) using T-interferon inducers, and of Sonnenfeld of ~1. (20) using BCG-tuberculin- induced interferon (type II), which indicate that T-type interferon or type II interferon seems to have exceptionally high immunosuppressive activity in comparison to virus-induced interferon. It has to be stated, however, that these studies have been done with nearly crude preparations of type II interferon.
We have recently developed a method for producing relatively large amounts of phytohemagglutinin-induced interferon in mouse spleen cell cultures (21, 22). Furthermore, in order to evaluate some biological properties of this type of interferon. and in particular its immunosuppressive activity, it appeared necessary to develop simple and efficient methods for its partial purification. For this purpose we have used affinity chromatography techniques to purify murine interferon on several sorbents with different chromatographic properties (21. 22). The present report shows that purified T-interferon inhibits antibody synthesis in t’itro to sheep red blood cells (SRBC). Upon fractionation on different matrices the inhibitory activity cannot be dissociated from antiviral activity. The study of the kinetics of the effect of T-interferon on antibody synthesis shows that it interferes with the early events of lymphocyte activation. These kinetics are similar to those obtained where using viral interferon (5). When titrated on the basis of antiviral activity. no difference in the inhibition of antibody synthesis could be found between T-interferon and viral interferon. The present report indicates therefore that the T-interferon molecule itself expresses immunosuppressive activity which is quantitatively similar to immunosuppression mediated by viral interferon.
MATERIALS AND METHODS
Anitntrls.
Nude heterozygous mice (nu/+) and BalbiC mice were supplied by Centre de Selection et d’Elevage d’animaux de laboratoire (CSEAL, Orleans, France) and B,,D,F, mice purchased from Bomholtgard, Ry (Denmark).
Con A-Sepharose and Blue Sepharose CL-6B were purchased from Pharmacia, Uppsala, Sweden, Methyl cr-D-mannopyranoside (a-MM) was obtained from Roth Light Laboratory. Coin Brook, Bucks, England. Fluorescamine was from Roche, Basel, Schwitzerland. Purified phytohemagglutinin (PHA) was obtained from Wellcome, Beckenham, England.
VitwI inrc~~:f~~t~~n. Mouse L-cell interferon induced by Newcastle disease virus (NDV) was prepared as described elsewere (23).

60 LUCERO ET AL.
T-interferon. PHA-interferon preparations were prepared using spleen cells from nude heterozygous mice (nu/+) or Balb/C mice, as described previously (21,22). In short, spleen cells were suspended at the concentration of 1 x 10’ cells/ml in RPM1 1640 medium (Flow Laboratories, United Kingdom) supplemented with 5% fetal calf serum (Flow), 2 mM glutamine and 4 mg% gentamicin, and incubated with 3 &ml or PHA in petri dishes (90 mm, Nunc, Roskilde, Denmark) for 24 hr at 37°C in a humidified incubator (5% CO,). Cells were then spun off and the supernatant frozen at -70°C until used. Antiviral titers were in the range of 200 units/ml (BalbK) to 500 units/ml (nude heterozygous mice).
Interferon preparations were precipitated with ammonium sulfate to a final saturation of 42% at 4”C, the precipitate removed, and the supernatant concentrated (5 to IO-fold) under vacuum and dialyzed against phosphate-buffered physiological saline (crude interferon). This crude interferon was purified by chromatography on Blue Sepharose CL-6B as described under Results and in the legend to Fig. 2. The interferon eluted from this column will be referred to as purified T-interferon and has a specific activity of 1-2 x lo” unitsimg protein.
PHA-interferon from BalbiC or nude heterozygous mice showed similar chromatographic behavior and their properties were indistinguishable.
Mock Intecferon
Mock T-interferon was prepared and processed in the same way as PHA-interferon but without addition of the mitogen. No significant antiviral activity (less than 10 units/ml) was detected.
InteTferon assay
Titration of interferon was performed by a cytopathogenic inhibition test (24), in L-cell monolayers using vesicular stomatitis virus, a challenge virus. All interferon titers are expressed in international reference units.
Protein Determination
Protein concentration was measured by absorbance at 280 nm or by a fluorometric assay (25) with bovine serum albumin as standard.
Chromatographic Procedure
All interferon preparations were dialyzed against appropriate buffers (as indicated under figures) at 4°C for 16 hr. The columns were equilibrated initially with buffers used for dialysis of interferon preparations. The equilibration and development of the column were performed at 4°C. The flow rate from the columns, 15 mlihr for absortion and 3 ml/hr for elution, was maintained by means of a peristaltic pump. Fractions were collected into plastic test tubes and dialyzed for 16 hr at 4°C against PBS; 100 ~1 was used for interferon titration and 100 ~1 for determination of their effect on the number of anti-SRBC plaque-forming cells.
Acid Stability
Interferon samples were dialyzed for 24 hr at 4°C against saline, pH 2, followed by dialysis against 0.02 M sodium phosphate-O. 145 M sodium chloride, pH 7.4 (PBS).

IMMUNOSUPPRESSIVE PROPERTIES OF T-INTERFERON 61
Controls were dialyzed only against PBS for the same time. Interferon titers of treated and control samples were measured simultaneously.
The method used was described by Gisler and Fridman (26) with minor modifications. Spleen cells (8 x 10s) from 8- to 12-weeks-old B,,D,F, mice were cultured in 12 x 75-mm Falcon plastic tubes (Falcon, Oxnard, Calif.) in I ml of RPM1 1640 medium (Eurobio, France) supplemented with 5% fetal calf serum (Gibco Diagnostic, Grand Island, N.Y.), I% horse serum (Gibco Diagnostic), 3% sodium bicarbonate, and antibiotics (penicillin and streptomycin, 5000 units each). Cultures were stimulated with 3 x 10” SRBC and incubated for 5 days at 37°C in a CO, incubator (5% in air).
Direct plaque-forming cells (PFC) were assayed by the local hemolysis technique in liquid medium (27).
RESULTS
I-Inhibition of‘ in Vitro Antibody Synthesis to SRBC by T-IntcrfiJrm
Mitogen stimulation of spleen cells induces cell proliferation and the release of a variety of mediators (28) including interferon (12- 15). Some of these mediators interfere with antibody synthesis either enhancing (29) or suppressing (26, 30) the generation of antibody-producing cells. To test, among these different activities, the specific effect of interferon, it is therefore obligatory to purify interferon activity. In fact, when crude supernatants were added together with antigen to spleen cells stimulated by SRBC, and the PFC response measured 5 days later, we observed either no effect, enhancement, or inhibition of the response (data not shown). We, therefore, applied PHA-induced interferon-containing supernatant to a Con A-Sepharose column and chromatography was performed in conditions previously described (22). One hundred microliters of each fraction recovered from the column was tested both for the presence of antiviral activity and for its effect on antibody synthesis in vitro by adding it, together with antigen, to cultures of spleen cells stimulated by SRBC. Figure I shows a representative experiment in which the antiviral activity (demonstrative of the presence of interferon) is distributed among two fractions: (i) the breakthrough fraction containing about 30-40s of the total applied antiviral activity and (ii) a fraction eluted with Q-MM containing 60-70s of the interferon activity. This fraction is recovered with about lOO-fold purification and a specific activity of IO” unitsimg). Strikingly, inhibitory activity forilr vitro antibody synthesis to SRBC, as measured by inhibition of PFC at Day 5, was found in the very same fractions as the antiviral activity (Fig. 1). The true chromatographic character of both fractions was established by their individual rechromatography on freshly prepared lectin columns. They were recovered in their original positions (data not shown).
The fact that fractionation of interferon-containing supernatant on Con A-Sepharose resulted in recovery of T-interferon in two peaks, one in the breakthrough and the other upon elution with (U-MM, suggested an heterogeneity of
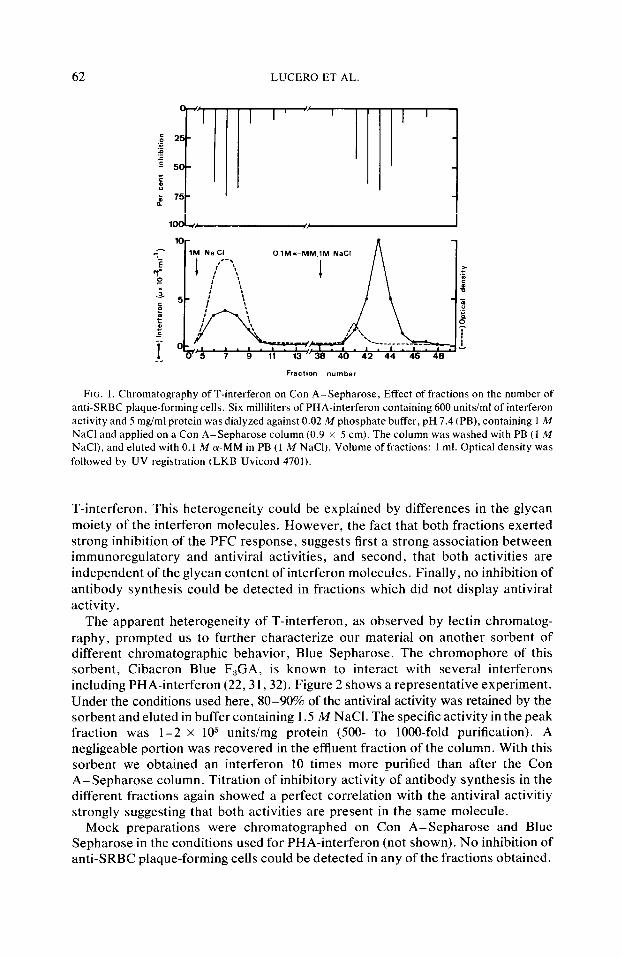
62 LUCERO ET AL.
= 2 .P f
r 5 5
‘; B
b 75
10 P-r ” f
7 9 11
FIG. 1. Chromatography of T-interferon on Con A-Sepharose, Effect of fractions on the number of anti-SRBC plaque-forming cells. Six milliliters of PHA-interferon containing 600 units/ml of interferon activity and 5 mgiml protein was dialyzed against 0.02 M phosphate buffer, pH 7.4 (PB), containing 1 M NaCl and applied on a Con A-Sepharose column (0.9 x 5 cm). The column was washed with PB (1 M NaCI), and eluted with 0.1 M (Y-MM in PB (1 M NaCl). Volume of fractions: 1 ml. Optical density was followed by UV registration (LKB Uvicord 4701).
T-interferon. This heterogeneity could be explained by differences in the glycan moiety of the interferon molecules. However, the fact that both fractions exerted strong inhibition of the PFC response, suggests first a strong association between immunoregulatory and antiviral activities, and second, that both activities are independent of the glycan content of interferon molecules. Finally, no inhibition of antibody synthesis could be detected in fractions which did not display antiviral activity.
The apparent heterogeneity of T-interferon, as observed by lectin chromatog- raphy, prompted us to further characterize our material on another sorbent of different chromatographic behavior, Blue Sepharose. The chromophore of this sorbent, Cibacron Blue F,GA, is known to interact with several interferons including PHA-interferon (22,3 1,32). Figure 2 shows a representative experiment. Under the conditions used here, 80-90% of the antiviral activity was retained by the sorbent and eluted in buffer containing 1.5 M NaCl. The specific activity in the peak fraction was l-2 x lo5 units/mg protein (500- to lOOO-fold purification). A negligeable portion was recovered in the effluent fraction of the column. With this sorbent we obtained an interferon 10 times more purified than after the Con A- Sepharose column. Titration of inhibitory activity of antibody synthesis in the different fractions again showed a perfect correlation with the antiviral activitiy strongly suggesting that both activities are present in the same molecule.
Mock preparations were chromatographed on Con A-Sepharose and Blue Sepharose in the conditions used for PHA-interferon (not shown). No inhibition of anti-SRBC plaque-forming cells could be detected in any of the fractions obtained.

IMMUNOSUPPRESSIVE PROPERTIES OF T-INTERFERON 63
50 “‘I ‘T I I
Fractmn “umber
FIG. 2. Chromatography of T-interferon on Blue Sepharose CL-6B. Effect offractions on the number of anti-SRBC plaque-forming cells. Ten milliliters of PHA-interferon containing 2500 units/ml of interferon activity and 12 mg/ml protein was dialyzed against PB and applied on a Blue Sepharose column (0.9 x 6 cm). The column was washed with 20 ml of PB, and then with 20 ml of 0.02 M sodium phosphate-O.145 M sodium chloride, pH 7.4, and eluted with PB containing 1.5 M NaCI. Volume of fractions: l- 18 = 3 ml; 19-29 = 1 ml. Optical density was followed by UV registration (LKB Uvicord 4701).
Il-Comparutir!e Eflects c$ T-Interj2ron and Viral Interferon on Antibody Synthesis in Vitro
Acid treutment. As predictable by its previously known physicochemical characteristics (12, 13, 22)) pretreatment of different preparations of T-interferon (crude or purified on Blue Sepharose) for 24 hr at pH 2 destroyed both the antiviral and the immunosuppressive activities. A representative experiment, using purified T-interferon is given in Table 1. By contrast, as already shown by Gisler et al. (5), both activities were preserved when viral interferon preparations were submitted to the same acid treatment (Table 1).
(b) Kinetics ofaction of T-interft>ron us compurcd to virul interjeron. In order to study the kinetics of action of T-interferon on the in vitro generation of PFC to SRBC, a given dose of T-interferon was added at different times to SRBC- stimulated cultures, under the conditions used by Gisler et al. (5) for viral interferon. When blue Sepharose-purified T-interferon (400 units) was added to cultures of mouse spleen cells 6 hr before or simultaneously with SRBC, a strong inhibition (about 6.5%) of antibody synthesis was observed (Fig. 3). Addition of interferon 24 hr after antigen resulted in an inhibition of 48 and 30% if interferon was added at 48 hr.
By contrast, if T-interferon was added to spleen cell cultures 72 or 96 hr after SRBC (not shown), no inhibition was observed. In the experiment given in Fig. 4, we compared the effect of crude, Blue Sepharose-purified T-interferon and viral NDV-L-cell interferon. High increase in the number of PFC was observed when

64 LUCERO ET AL.
TABLE 1
Effect of pH2 Pretreatment of Viral and T-Interferon on the Antiviral Activity and on the Number of Anti-SRBC Plaque-Forming Cells
PHI Treatment SRBC
Interferon (units/ml)
PFCIlO” (viable cells)
Inhibition (%)
Control 20 k 3 Control + 722 t 41 T-interferon untreated” + 300 267 t 18 63 T-interferon treated + 30 747 5 33 0 Viral interferon untreated* + 300 310 t 19 57 Viral interferon treated + 300 252 i- 22 65
” PHA-interferon purified on Blue Sepharose. h Viral interferon used: NDV-L-cell interferon.
crude T-interferon-containing supernatant was added 24 or 48 hr after SRBC. Again, purified T-interferon did not display any enhancing activity and inhibits the PFC response when added between 0 and 24 hr. Therefore, the most likely explanation is that the enhancing activity exerted by crude interferon supernatants is due to other mediator(s) present in the crude preparation of T-interferon. Finally, the kinetics of action of murine T-interferon are similar to those of viral interferon. Both interferon expressed their immunosuppressive activity when added during the induction period of the response. When they were added more than 48-72 hr after antigenic stimulation, their inhibitory activity was no longer expressed (data not shown).
(c) Comparative dose-response effect between T- and viral interferon. Graded doses (between 12 and 1600 units) of purified T- and viral interferons were added, at the same time as antigen, to spleen cells stimulated by SRBC and the inhibitory effect on the generation of direct PFC was measured on Day 5. As shown in Figure 5, inhibition increased as a function of the dose of interferon added and no significant difference could be observed when the same dose of T- or viral interferon was added to the cultures.
II ’ I I I I -6 0 24 48 72
Time of Interferon additmn ( hours)
FIG. 3. Kinetics of the interferon effect on anti-SRBC plaque-forming cells. Control: 720 2 41 PFC/lO” viable cells; background: 20 2 3 PFC/lOG viable cells.

IMMUNOSUPPRESSIVE PROPERTIES OF T-INTERFERON 65
19500
19000
(0 = x
o e 18500 / :
i 1500
s
: Q
0
1000
%
K
i
500
FIG. 4. Effect of different interferon preparations on the number of anti-SRBC plaque-forming cells. Interferon added: 400 units/ml. Controls (0): background (W): purified T-interferon (0): crude T-interferon ( q ): NDV- L-cell interferon 0.
DISCUSSION
Many studies reported during the last few years indicate that viral interferon may interfere in the regulation of antibody synthesis (1,4-6). We were interested in the possible role of mitogen-induced interferon (T-type) in the regulation of the immune response. Very little work has been reported on this problem using immune interferons induced in immunocompetent cells by antigens or mitogens.
Johnson and Baron (19) have shown that T mitogens such as Con A or staphylococcal enterotoxin A were inhibitors of the PFC response against SRBC when added simultaneously with the antigen. They suggested that the immune interferon produced under these conditions was the mediator of the suppressive activity. The experimental results presented herein demonstrate directly that generation of PFC against SRBC in mouse spleen cell cultures can be inhibited by the addition of purified T-interferon during the inductive phase of the immune response. This interferon was produced in spleen cell suspensions stimulated with PHA. Using PHA-induced supernatants as starting material, we have determined the affinity chromatographic behavior of both antiviral and immunoregulatory activities on two sorbents of different chromatographic biases: Blue Sepharose and Con A-Sepharose, that we have already shown to interact with PHA-interferon (22). On Con A-Sepharose both activities were distributed into the same two major fractions, one in the breakthrough and the other which has a high affinity for the matrix and can be eluted after addition of cr-MM to the elution solvent which results in a considerable purification (about IOO-fold). Moreover these results indicate that the difference in the glycan moiety between the breakthrough and the retained fraction does not result in alteration of the biological activities since both fractions were antiviral and immunosuppressive.

66 LUCERO ET AL.
1500-
YI % "
P ~lOOO- B a:
i .;
'0
g soo-
,o 0
OL
FIG. 5. Dose effect of viral and T-interferon on the number of anti-SRBC plaque-forming cells. Interferons were added simultaneously with SRBC.
On the other sorbent, Blue Sepharose, again, only those fractions which contained immunosuppressive activity also contained antiviral activity. No immunosuppressive activity was found in fractions without antiviral activity. Under the conditions used in this work, 85% of the antiviral activity was recovered into a single peak which has a specific activity of 1-2 x 10” units/mg.
The immunosuppressive activity of Blue Sepharose-purified interferon was time and dose dependent. The effect of purified T-interferon was maximum when added 6 hr before or together with the antigen, but significant inhibition was still obtained when interferon was added up to 24-48 hr after antigen. No effect was observed when purified T-interferon was added after this time. These kinetics are similar to the kinetics of inhibition of antibody synthesis obtained with viral interferon (5), both interfering with the early phase of the immune response.
The most likely explanation for its immunosuppressive activity is that T-type interferon, as well as viral interferon (2, 3) inhibits proliferation of T or B cells and subsequently antibody production (33). Gisler et al. (5) utilizing “mosaic” cell cultures where T, B cells, or macrophages were treated separately by viral interferon have shown that it acts on B cells, probably by interfering with the proliferation of the precursors to antibody-producing cells. Our own experiments showing that T-type interferon inhibits antibody synthesis in spleen cell cultures from congenitally athymic nude (nu/nu) mice (data not shown) suggest that immune interferon also acts on B cells. However, inhibition of cell proliferation is not the only mechanism by which interferon may affect B-cell differentiation. It may interact with antigen binding by T or B cells; it has recently been proposed that interferon may enhance antigen binding by T cells (34). Interferon may also affect T-B cell cooperation, for example, by inducing T suppressor cells. However, the fact that T-type interferon inhibits antibody production in nude mice indicates that it does not act, on mature T cells at least. Finally it is quite possible that the observed inhibition of antibody synthesis results in additional effects of interferon

IMMUNOSUPPRESSIVE PROPERTIES OF T-INTERFERON 67
on different compartments of the immune response. It is clear that studies on cellular and molecular mechanisms by which interferons exert their im- munoregulatory activities are at their very beginning. The fact that when inhibition of antibody synthesis is expressed as a function of the antiviral activity, no significant difference can be observed between viral and T-type interferons (Fig. 5) brings additional evidence that the mechanisms of interactions of interferons with different cells are similar for the different interferons.
Sonnenfeld ct ul. (20) recently reported that serum containing antigen-induced interferon (type II) or in vitro preparations were 1.50 times more active than viral interferon in inhibiting the PFC anti-SRBC response. However, the increased immunosuppressive activity was observed only when cultures were treated for 24 hr with type II interferon prior to antigen stimulation. A major difference compared to Sonnenfeld’s work is the fact that we used mitogen-induced interferon. Type II interferon (antigen-induced) and T-interferon (mitogen-induced), shared similar properties (10, 12-15). They are unstable at pH 2 and are not neutralized by antiviral interferon serum. However, the intrinsic nature of mitogen- and antigen- induced interferon may differ and this would explain their apparently different immunosuppressive capacity.
Furthermore, although the purified T-interferon used in our study had a specific activity of l-2 x IO” unitsimg protein (the highest reported so far), it was not, of course completely purified. We therefore cannot exclude the possible presence of some residual contaminants which might affect the level of the anti-SRBC PFC inhibition. In addition contaminants in interferons prepared by antigen or mitogen stimulation are probably different and might possess different properties.
Certain crude preparations of PHA-interferon were found to cause a marked enhancement of the SRBC antibody response. Similar observations have been reported for viral (5) and type II interferon (35). However, in our case the kinetics of the enhancing activity (see Fig. 4) and the fact that this activity was absent from purified preparations of T-interferon suggest that the enhancing effect might be due to a factor(s) present in crude supernatants different from interferon.
Finally, the expression of the immunosuppressive activity and the antiviral activity were pH 2 labile. Taken together with the affinity chromatography patterns, our results suggest that the same molecule is responsible for both activities. To our knowledge this is the first report showing directly that a purified mitogen-induced interferon is able to modify the antibody immune response. More work needs to be done in order to determine precisely the role of various types of interferon in the regulation of the immune response.
Since synthesis of interferon can be induced by specific antigens (10, 11). it is present together with other regulatory factors (37) during the development of immune responses. With regard to antibody synthesis, T-type interferon takes its place among the nonspecific suppressor factors produced by T cells upon antigen or mitogen stimulation (28). Interferon is clearly different from suppressor immunoglobulin binding factor (IBF) (26) which is stable at pH 2 (37) and acts during the late phase of the generation of plaque-forming cells (26) without interfering with B-cell proliferation (38). While different in terms of molecular weight (39). interferon and concanavalin A-induced soluble immune response suppressor (SIRS) (29) have similar kinetics of action, the latter activating macrophages which would then inhibit B-cell proliferation (40). Immune interferon appears therefore to

68 LUCERO ET AL.
be another nonspecific early suppressor factor produced in each immune reaction, probably having the peculiar property of itself interacting with different compartments of the immune system, such as T cells, B cells, macrophages, or NK cells. To evaluate the importance and the role of these ubiquitous molecules, it is now an absolute requirement to characterize and purify T- and B-type interferons and to study the cellular and molecular bases of their interactions with lymphocytes.
ACKNOWLEDGMENTS
This work was supported by Direction GCnCrale de la Recherche Scientifique et Technique, Research Grant 77.7.1375 and the Institut National de la Sante et de la Recherche Medicale CRL 78.4.0821.
REFERENCES
1. Johnson, H. M., and Baron, S., ZRCS Med. Sci. 4, 50, 1976. 2. Lindahl-Magnusson, P., Leary, P., and Gresser, I., Nature Nrit, Bid/. 273, 120, 1972. 3. Pacheco, D., Falcoff, R., Catinot, L., Floc’h, F., Werner, G. H., and Fa]coff, E., Ann. [nst,
Pasteur, Puris 137C, 163, 1976. 4. Brodeur, R. B., and Merigan, T. C., J. Immunol. 4, 1323, 1975. 5. Gisler, R. H., Lindahl, P., and Gresser, I., Irnmunolv~y 115, 438, 1974. 6. Johnson, H. M., Smith, H. G., and Baron, S., J. Zmmunoi. 114, 403, 1975. 7. Trinchieri, G., Santoli, D., Dee, R. R., and Knowles, B. B., J. Exp. Med. 147, 1299, 1978. 8. Gidlund, M., Om, A., Wigzell, H., Senik, A., and Gresser, I., Nuture London 273, 759, 1978. 9. Virelizier, J. L., Allison, A. C., and De Maeyer, E., Infect. Immun. 17, 282, 1977.
10. Youngner, J. S., and Salvin, S. B., J. Immunol. 111, 1914, 1973. 11. Valle, J. M., Bobrove, A. M., Straber, S., and Merigan, T. C., J. Immune/. 114, 435, 1975. 12. Epstein, L. B., Tex. Rep. Biol. Med. 35, 42, 1977. 13. Wietzerbin, J., Falcoff, R., Catinot, L., and Falcoff, E., Ann. Inst. Pnstrur, Puris 128C, 699,
1977. 14. Wallen, C. W., Dean, J. H., and Lucas, D. O., Cell. Immrrnol. 6, 110, 1973. 15. Wietzerbin, J., Stefanos, S., Falcoff, R., Lucero, M., Catinot, L., and Falcoff, E.,Infrct. Immunol.
21, 966, 1978. 16. Metz, D. H., Cc//. 6, 429, 1975. 17. Lewis, J., Falcoff, E., and Falcoff, R., Potho/. Biol. 25, 9, 1977. 18. De Maeyer-Guignard, J., Tovey, M. G., Gresser, I., and De Maeyer, E., Nrrture (London) 271,622,
1978. 19. Johnson, H. M., and Baron, S., Crli. Immunol. 25, 106, 1976. 20. Sonnenfeld, G., Mandel, A. D., and Merigdn, T. C., Cell. Immunol. 34, 193, 1977. 21. Wietzerbin, J., Stefanos, S., Lucero, M., Falcoff, E., Thang, D. C., and Thang, M. N., B&hem.
Biophys. Rrs. Commun. 85, 480, 1978. 22. Wietzerbin, J., Stefanos, S., Lucero, M., Falcoff, E., O’Malley, J., and Sulkowski, E., J. Gen.
Viral. 44, 773, 1979. 23. Falcoff, E., Falcoff, R., Lebleu, B., and Revel, M., J. Viral. 12, 421, 1973. 24. Have], E. A., and Vilcek, J., Antimicrob. Agents Chemother 2, 476, 1972. 25. Bohlen, P., Stein, S., Dairman, W., and Udenfriend, S., Arch. Biochem. Biophy. 155,213, 1973. 26. Gisler, R. H., and Fridman, W. H., J. Exp. Med. 142, 507, 1975. 27. Cunningham, A. J., Nature (London) 207, 1106, 1969. 28. Dutton, R. W., Falkoff, R., Hirst, J. A., Hoffmann, M., Kappler, J. W., Kettman, J. R., Lesley,
J. F., and Vann, D., Progr. Immunol. 1, 3, 1971. 29. Schimpl, A., and Wecker, E., Nature New Biol. 237, 15, 1972. 30. Rich, R. R., and Pierce, C. W., J. Erp. Med. 137, 649, 1973. 31. De Maeyer-Guignard, J., and De Maeyer, E., Compt. Rend. Acadh. Sci. Paris 283, 709, 1976. 32. Jankowski, W. J., Von Muench Hansen, W., Sulkowski, E., and Charter, W. A., Biochemisty 15,
5182, 1976.

IMMUNOSUPPRESSIVE PROPERTIES OF T-INTERFERON 69
33. Dutton. R. W., and Mishell, R. J., J. E.tp. Md. 126, 443, 1967. 34. Lonai, P.. and Steinman. J., Prw. Nor. Actrrl. SC;. USA 74, 5662. 1977. 35. Sonnenfeld, G., Mandel. A. D.. and Merigan, T. C.. Cell. Irwnrmol. 40, 285. 1978. 36. Bartfeld, H., and Vilceck, J., I,~,fi,c,r. I/nw~rrno/. 12, 1112, 1975. 37. Joskowicz. M.. Rabourdin, C.. Neauport-Sautes, C.. and Fridman. W. H., ./. Imr,~uno/. 121, 777.
1978. 3X. Yagello. M.. Rabourdin-Combe, C., and Fridman, W. H., 7rtrr1,cplrrnt. Pwc. 11, X95. 1979. 39. Tadakuma. T.. Kuhnar. A. L.. Rich, R. R.. David, J. R.. and Pierce. C. W., J. Irnmrrnol. 117, 323,
1976. 40. Tadakuma, T., and Pierce. C. W., .I. Iw~,nrrn~,/. 120, 481, 1978.