genotypic heterogeneity within giardia lamblia isolates demonstrated by m13 dna fingerprinting
TRANSCRIPT

INFECTION AND IMMUNITY, May 1994, p. 1875-1880 Vol. 62, No. 5001 9-9567/94/$04.0()+0Copyright (© 1994. American Society for Microbiology
Genotypic Heterogeneity within Giardia lamblia IsolatesDemonstrated by M13 DNA Fingerprinting
SIMON CARNABY,? PETER H. KATELARIS, ARIFA NAEEM, AND MICHAEL J. G. FARTHING
Department of Gastroenterology, St. Bartholomew's Hospital, London, ECIA 7BE, United Kingdom
Received 18 October 1993/Returned for modification 7 December 1993/Accepted 7 Februarv 1994
There has been considerable speculation regarding the possible relationship between the phenotypic andgenotypic heterogeneity seen among human isolates of Giardia lamblia and the wide clinical spectrum of humangiardiasis. Several workers have suggested that human giardiasis may be a mixed infection consisting ofvariant strains or subgroups which are present in the same infection and which are selectable, but it is not clearwhether these apparent variant strains represent a truly heterogeneous infection or whether the genotypicheterogeneity observed is due to the susceptibility of the Giardia genome to a high rate of structural geneticrearrangement. We have therefore studied variation in Giardia intestinalis genotypes in 19 isolates in vitro andin vivo by using the technique of M13 DNA fingerprinting. Genotypes of isolates changed with time whencultured under standard conditions and when pressured with bile. Sequential isolates and their clones takenfrom a patient with chronic giardiasis both before and after several treatments with metronidazole haddifferent genotypes. Finally, clones of isolate WB had different initial genotypes, which changed after 4 monthsin culture. These findings suggest that the apparent genotypic heterogeneity at least in these G. intestinalisisolates is more likely to be due to the plasticity of the Giardia genome than to the presence of a truly mixedpopulation of strains within the same infection.
Giardia lamblia is an amitochondrial binucleate protozoalparasite of the small intestine and represents the earliestdiverging lineage of the eukaryotic line of descent on the basisof rRNA sequence analysis (23). The clinical spectrum ofhuman giardiasis varies from an asymptomatic carrier state tosevere diarrhea that is often accompanied by malabsorption(12). The reasons for this clinical heterogeneity have not beendefined, and disease expression in the host has not beenrelated to any putative markers of virulence or biologicalcharacteristics of the organism.
Several reports have suggested that human giardiasis may bea mixed infection by phenotypically and genotypically variantstrains or subgroups (3, 9, 19) which exhibit different sensitiv-ities to drugs and environmental conditions and which areselectable (4). It is not certain, however, whether the individualgenotypes within these mixed populations are representative ofa heterogeneous population of organisms in the environment,which, when present in the same infection, are affected byselection pressures or whether the different genotypes occur asa result of selection pressures magnifying some form ofmutational event such as the extensive genomic rearrange-ments seen by pulsed-field gel analysis of the Giardia chromo-somes (1, 18).There is some evidence that the classification Giardia intes-
tinalis may contain subgroups on the basis of ribosomalsequence information (30) and isoenzyme electrophoresis (20),representatives of which may be present in the same infectionand may therefore be selectable by drug or environmentalpressures (4). This would lead to the conclusion that all thelimited allelic variations seen between and within isolates aredue to selection of specific genotypes from a putative mixedpopulation. However, these and other extensive genomic poly-morphisms (18) could be brought about by the highly plasticnature of the Giardia genome, which is susceptible to a highrate of genetic rearrangement and therefore heterogeneity.
Corresponding author.
This would imply that the selection pressures are acting on agenetic population with a high degree of spontaneous hetero-geneity as in Trypanosoina (28) and Plasmodium (10, 17)species rather than on a truly mixed population. The individualGiardia chromosomes vary in size by up to 200 kb in differentisolates, with changes taking place at a rate of up to 3% perdivision in in vitro cultures without any overt pressures beingapplied (18). Changes in the rDNA (DNA encoding rRNA)repeat number can account for only some 30% of the chromo-some size changes seen (18). The nonribosomal component ofthe rearranged segments of the variable chromosomes (-70%)could be made up predominantly of some form of simplesequence or repetitive DNA as it is mostly devoid of restrictionenzyme sites (18). It has been estimated by Cot analysis thatsome 14% of the Giardia genome could be made up ofrepetitive DNA, which would allow for such large subtelomericrepeated elements (18). These long stretches of putativerepetitive DNA are likely to be important and may even bedirectly involved in genetic rearrangement. It is therefore likelythat such genotypic rearrangement is important in the devel-opment of biologically significant genotypic changes underenvironmentally selective conditions such as the developmentof drug resistance or immune evasion by some form ofrearrangement of the surface antigen gene family (2).
This situation is similar to that found in other parasiticprotozoa such as Plasmodium falciparum and Trypanosomabrucei, in which gene rearrangement is known to be involved indifferential gene expression. In T brucei, programmed generearrangement of variant surface antigens, aided by repeatsequences, is involved in rearrangement of abundant minichro-mosomes to bring antigen genes into telomere-associatedexpression sites (8, 28). Extensive chromosomal rearrange-ment in Plasmodium species has also been speculated to beinvolved in regulation of surface antigen gene expression (10,17) and has been shown to play a role in development ofresistance to antimalarial drugs. For example, chloroquineresistance has been shown to occur in vivo by amplification ofthe multidrug resistance gene pfmdrl (26) and resistance to
1875

1876 CARNABY ET AL.
TABLE 1. Isolates of G. lamblia used in this study
Isolate Geographic origin Method of isolation Year Source oraxenized reference"
Portland I Oregon Jejunal aspirate 1976 21P01 United States Jejunal aspirate 1983 6BI United Kingdom Jejunal aspirate 1987 SBHB2 United Kingdom Jejunal aspirate 1988 SBHB2 clone United Kingdom Jejunal aspirate 1988 SBHB3 United Kingdom Jejunal aspirate 1988 SBHB3 clone United Kingdom Jejunal aspirate 1988 SBHB4 United Kingdom Jejunal aspirate 1988 SBHVNBI Asia Excystation 1981 7VNB2 Asia Excystation 1981 7VNB3 Asia Excystation 1981 7VNB4 Asia Excystation 1981 7VNB5 Asia Excystation 1981 7RW6 United Kingdom Jejunal aspirate 1989 SBHRW6 clone United Kingdom Jejunal aspirate 1989 SBHGP Oregon Jejunal aspirate (animal) 1976 15Ness United Kingdom Jejunal aspirate 1982 HVSBH1 United Kingdom Jejunal aspirate 1984 SBHWB Afganistan Jejunal aspirate 1982 22
a SBH, axenized by the authors at St. Bartholomew's Hospital, London, United Kingdom; HVS, axenized and supplied by H. V. Smith, Department of Bacteriology,Stobhill General Hospital, Glasgow, United Kingdom.
the dihydrofolate reductase antagonist has been shown todevelop in vitro as a result of duplication and rearrangement ofthe dihydrofolate reductase gene-containing chromosome(25).The genome of G. intestinalis is therefore susceptible to a
high degree of heterogeneity, and multiple genotypes can befound within a single isolate. It is not known, however, whethersuch genotypically mixed cultures are brought about by thegenomic plasticity of the organism, whether they reflect infec-tion by multiple genotypic variants, or whether they are simplyan artifact of axenic culture without in vivo biological rele-vance. It is therefore important to examine these possibilitiesin more detail before genetic markers are assigned to Giardiastrains or subgroups in an attempt to classify their virulencecharacteristics. The question therefore arises of whether thesechanges (i) are representative of the natural slow genetic driftof Giardia intestinalis, (ii) are brought about by environmentalpressures, or (iii) represent a change in the predominantgenotype from a genetically mixed population within theisolate.
In the present study we have attempted to answer thisquestion by using M13 DNA fingerprinting to characterize thegenotypes of isolates and their clones subjected to variousenvironmental stresses including bile, drugs, and prolonged invitro culture.
MATERIALS AND METHODS
Giardia isolates. Trophozoites used to establish axenic cul-tures were obtained from human duodenal intestinal contentsor feces with the exception of GP, which is a guinea pig isolate(Table 1). Three sequential isolates were taken from a patientwith chronic giardiasis and panhypogammaglobulinemia. Thefirst isolate, B2, was taken upon initial presentation, and thesecond isolate, B3, was taken 3 months later after three coursesof the antigiardial drug metronidazole, which did not clear theinfection. A third isolate, RW6, was obtained after a further 6months following two additional courses of metronidazole,which again did not eradicate the infection. The infection wasfinally cleared with mepacrine. A clone of each isolate was
made by the limiting-dilution method immediately after axeni-zation.
Culture and cloning procedures. Giardia trophozoites werecultured axenically in vitro in borosilicate glass screw-cap tubesat 37°C in TYI-S-33 medium (16) and were subcultured every3 days at stationary phase. The Giardia isolates were cloned bythe limiting-dilution method (5) such that there was only a 1%chance of any one clonal culture being derived from more thanone viable cell. The cell count of the log-phase culture wasassessed by hemacytometry, and a dilution was then made inTYI-S-33 medium to give a final cell count of 1 cell per ml.This was aliquoted into sterile 96-well microtiter plates at arate of 100 pl per well. The plates were cultured for 72 h at37°C in sealed bags containing an Anaerocult P sachet (Merck)to maintain an anaerobic atmosphere. Samples in wells con-taining trophozoites as assessed by microscopy were thensubcultured into 16-ml borosilicate glass tubes and culturedconventionally.
Preparation of genomic DNA. Genomic DNA was isolatedfrom trophozoites by proteinase K-sodium dodecyl sulfate(SDS) digestion and organic extraction as previously described(11). DNA was purified by treatment with preboiled RNase Aand reextraction with phenol-chloroform-isoamyl alcohol andprecipitated with 0.1 M NaCl in 70% ethanol. Pellets werewashed twice in 70% ethanol to remove any excess salt, dried,and redissolved in TE buffer (10 mM Tris-HCl, 1 mM EDTA[pH 8]). DNA concentrations were assessed and comparedamong isolates in a conventional 1% agarose gel with 1 K.g ofEcoRI-HindIII-cut lambda DNA for comparison.M13 DNA fingerprinting. The restriction enzymes PvuII,
Sau3A, RsaI, and CfoI were used as specified by the manufac-turer (Boehringer Mannheim). Genomic DNA (1 ,ug perisolate) was digested to completion with a 10-fold excess ofrestriction enzyme for 16 h at 37°C. The cleaved DNA wasseparated at 4 V/cm for 6 h in a 1% agarose gel made up in 1 xTBE buffer (45 mM Tris-borate, 1 mM EDTA [pH 8.3]).Southern transfer onto a nylon membrane (Hybond N; Amer-sham) was performed by standard techniques (24), and theDNA was immobilized on the membrane by exposure to astrong UV source (312 nm) for 2 min.
INFECT. IMMUN.

GENOTYPIC HETEROGENEITY OF GIARDIA LAMBLIA 1877
kb 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19
20--
5--
2-
0.5--
FIG. 1. M13 DNA fingerprint analysis of Giardia lamblia. Shown isan autoradiographic fingerprint of Rsal-digested total genomic DNAfrom 19 G. lamblia isolates. Lanes: 1, Portland 1; 2, P01; 3, Bi; 4, B2;5, B2 clone; 6, B3; 7, B3 clone; 8, B4; 9, VNB1; 10, VNB2; 11, VNB3;12, VNB4; 13, VNB5; 14, RW6; 15, RW6 clone; 16, GP; 17, Ness; 18,BH1; 19, WB.
I 2 3 4 5
kb N 0 N 0 N 0 N 0 N 0
20
0.5 --
FIG. 2. M13 DNA fingerprint analysis of Giardia lamblia. Shown isa fingerprint comparison of PvuII-digested DNA preparations, made 2years apart (N, new DNA preparation; 0, 2-year-old DNA prepara-tion), from five G. lamblia isolates. Pairs: 1, B4; 2, VNB4; 3, VNB5; 4,Ness; 5, WB.
A 1,446-bp AluI restriction fragment from the M13 replica-tive-form DNA (Stratagene) gene III region containing bothrepeat elements (29) was labeled with [32P]dCTP by therandom-priming method (14) to a specific activity greater than109 cpm/,ug of template DNA. Prehybridization was carriedout for 4 h at 60°C in 7% SDS-0.263 M Na2HPO4 (pH 7.2)-imM EDTA-0.1cc bovine serum albumin. Hybridization wascarried out for 24 h in the same buffer including 10 ng oflabeled M13 probe per ml, again at 60°C.
Blots were washed twice in 2 x SSC (1 x SSC is 150 mMsodium chloride and 15 mM trisodium citrate)-0.1% SDS atroom temperature for 15 min and twice in the same solution at42°C for 15 min. Moist filters were wrapped in Saran Wrap andautoradiographed with B-max film (Amersham) and two in-tensifying screens at - 85°C for 48 h.
Effect of time in culture and bile on genotype. Parallelcultures of all isolates were maintained in culture medium withand without 0.6 mg of bovine bile per ml. DNA preparationswere made after 2 weeks and 4 months in continuous culture.Genotypic comparisons were made between these parallelisolates by M13 DNA fingerprinting.
RESULTSM13 DNA fingerprinting. M13 DNA fingerprinting revealed
genotypic differences among the 19 Giardia isolates studied(Fig. 1). Many bands appear to be common to all isolates, butmost isolates can be distinguished by a few variations inbanding pattern. These differences are reproducible when thesame DNA preparations are used (data not shown).To examine the stability of these differences with time, we
compared fingerprints in Fig. 1 with those of DNA prepara-tions made some 2 years earlier from five of the same isolates(Fig. 2). Several differences can be seen between temporallyseparated DNA preparations from the same isolate, indicatingthat the genotypes of these isolates changed without any overtculture pressure being applied, since the isolates were main-tained in continuous culture with standard media and cultureconditions throughout.
Effect of bile on genotype. M13 fingerprints of isolatescultured for 2 weeks with bile were similar to those of controlscultured without bile; however, after 4 months in continuousculture, two isolates (Ness and WB) did show quite markedgenotypic changes (Fig. 3), notably the appearance of astrongly positive band of approximately 1.5 kb in the WBisolate cultured with bile.
Effect of time alone on clonal cultures. To investigatewhether the apparent genotypic differences seen were the
2
kb
20 --
5 --
2 --
0.5 --
FIG. 3. Ml3 DNA fingerprint analysis of Giardia laniblia. Shown isa fingerprint comparison of RsaI-digested total genomic DNA fromtwo isolates of C. lamblia grown with (+) and without (-) 0.6 mg ofbovine bile per ml. Pairs: 1, Ness; 2, WB.
VOL. 62, l199'4
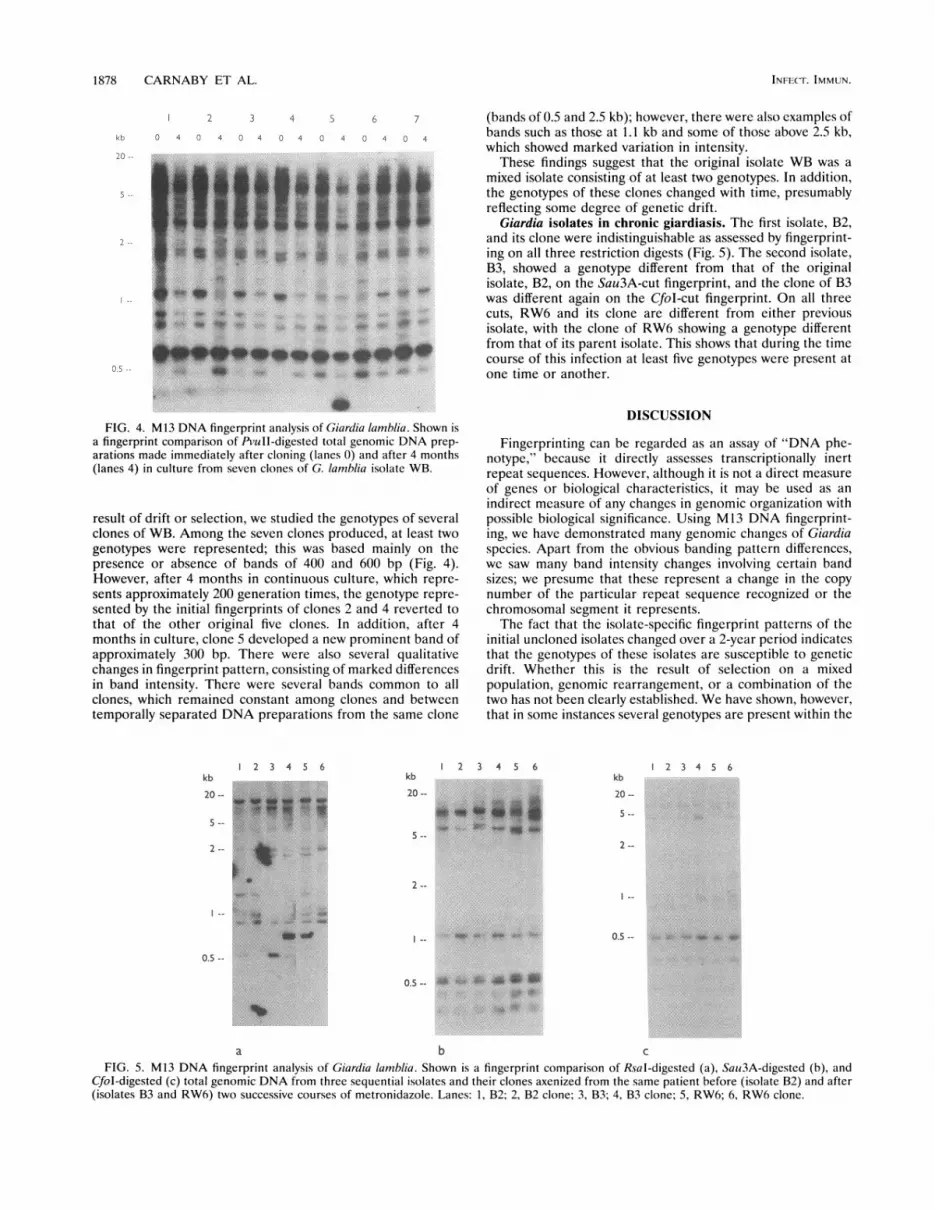
1878 CARNABY ET AL.
3 4 5
kb
20 --
5 --
2 --
I--
0.5 --
7
0 4 0 4 0 4 0 4 0 4 0 4 0 4
FIG. 4. M13 DNA fingerprint analysis of Giardia lamblia. Shown isa fingerprint comparison of PvuII-digested total genomic DNA prep-arations made immediately after cloning (lanes 0) and after 4 months(lanes 4) in culture from seven clones of G. lamblia isolate WB.
result of drift or selection, we studied the genotypes of severalclones of WB. Among the seven clones produced, at least twogenotypes were represented; this was based mainly on thepresence or absence of bands of 400 and 600 bp (Fig. 4).However, after 4 months in continuous culture, which repre-sents approximately 200 generation times, the genotype repre-sented by the initial fingerprints of clones 2 and 4 reverted tothat of the other original five clones. In addition, after 4months in culture, clone 5 developed a new prominent band ofapproximately 300 bp. There were also several qualitativechanges in fingerprint pattern, consisting of marked differencesin band intensity. There were several bands common to allclones, which remained constant among clones and betweentemporally separated DNA preparations from the same clone
kb
20 --
5 --
2 --
I--
0.5 --
2 3 4 5 6kb
20-
5 --
2--
I --
0.5 --
(bands of 0.5 and 2.5 kb); however, there were also examples ofbands such as those at 1.1 kb and some of those above 2.5 kb,which showed marked variation in intensity.
These findings suggest that the original isolate WB was amixed isolate consisting of at least two genotypes. In addition,the genotypes of these clones changed with time, presumablyreflecting some degree of genetic drift.
Giardia isolates in chronic giardiasis. The first isolate, B2,and its clone were indistinguishable as assessed by fingerprint-ing on all three restriction digests (Fig. 5). The second isolate,B3, showed a genotype different from that of the originalisolate, B2, on the Sau3A-cut fingerprint, and the clone of B3was different again on the CfoI-cut fingerprint. On all threecuts, RW6 and its clone are different from either previousisolate, with the clone of RW6 showing a genotype differentfrom that of its parent isolate. This shows that during the timecourse of this infection at least five genotypes were present atone time or another.
DISCUSSION
Fingerprinting can be regarded as an assay of "DNA phe-notype," because it directly assesses transcriptionally inertrepeat sequences. However, although it is not a direct measureof genes or biological characteristics, it may be used as anindirect measure of any changes in genomic organization withpossible biological significance. Using M13 DNA fingerprint-ing, we have demonstrated many genomic changes of Giardiaspecies. Apart from the obvious banding pattern differences,we saw many band intensity changes involving certain bandsizes; we presume that these represent a change in the copynumber of the particular repeat sequence recognized or thechromosomal segment it represents.The fact that the isolate-specific fingerprint patterns of the
initial uncloned isolates changed over a 2-year period indicatesthat the genotypes of these isolates are susceptible to geneticdrift. Whether this is the result of selection on a mixedpopulation, genomic rearrangement, or a combination of thetwo has not been clearly established. We have shown, however,that in some instances several genotypes are present within the
1 2 3 4 5 6kb
20 --
5 --
2 --
I--
0.5 --
1 2 3 4 5 6
a b c
FIG. 5. M13 DNA fingerprint analysis of Giardia lamlblia. Shown is a fingerprint comparison of RsaI-digested (a), Sau3A-digested (b), andCfol-digested (c) total genomic DNA from three sequential isolates and their clones axenized from the same patient before (isolate B2) and after(isolates B3 and RW6) two successive courses of metronidazole. Lanes: 1, B2; 2, B2 clone; 3, B3; 4, B3 clone; 5, RW6; 6, RW6 clone.
INFE(-F. IMMUN.

GENOTYPIC HETEROGENEITY OF GIARDIA LAMBLIA 1879
same infection. For example, the isolates taken from thepatient with chronic giardiasis, from which genotypically vari-ant clones were recovered, gave different fingerprint patternsafter drug pressure was applied in vivo. Selection from a mixedpopulation or genomic plasticity could account for theseresults. Bile, a normal component of the intestinal milieu anda known growth stimulant of Giardia trophozoites (13), wasexamined for its effects as an in vitro pressuring agent.Genotypic differences could be seen in some isolates whengrown with or without bile. This could be explained byselection pressures being applied to a mixed population orperhaps even by coincidence rather than by specifically in-duced genotypic changes. However, the clones of WB grown inbile-free medium showed genomic changes with time, whichimplies that some form of "mutational" event had taken placebecause these cultures were clonal in origin. The changes seenin the clones of WB during conventional culture must repre-sent the basal rate of genetic drift to which the organism issusceptible, because no specific pressures were applied to thecultures.The WB clones are very similar both to the parent isolate
and to each other. This would mean that for isolate WB, thepresence of several genotypes is unlikely to be due to themixed-population theory. It is more likely that this minorgenotypic heterogeneity between the clones has arisen as aresult of nonspecific genetic drift from the basic genotype ofWB. Similarly, the changes seen when pressuring isolates withbile in vitro or metronidazole in vivo seem to be only relativelyminor variations around a basic genotype for that isolate. Theycertainly do not show the same degree of genotypic differencesseen between different isolates, and it is thus unlikely that theyare due to selection from a truly mixed population.When studying changes in repeated loci which only repre-
sent a very small nontranscribed portion of the genome, onemust consider the relevance of these changes, in that they maysimply represent inert structural changes in what are biologi-cally inert nontranscribed genomic regions. The specificchanges seen in repeat sizes themselves may indeed be biolog-ically inert but may be markers of more relevant genomicchanges, such as the structural changes centered around thesubtelomeric repeat sequences seen in the P. falcipanim ge-nome, which have been associated with antigenic variation (10,17), and in T. brucei, in which transposition aided by repeatsequences from transcriptionally inactive regions to moreactive telomeric positions may be a way of controlling theexpression of surface antigen genes (28). Therefore, becausemultiply repeated elements are generally thought to be recom-binational hot spots, their involvement in biologically signifi-cant genomic rearrangement events in Giardia species is notunrealistic since it has been speculated that many of thesubtelomeric sequences involved in the genomic rearrange-ments in Giardia species may be composed of repeated ele-ments (18). This is supported by recent evidence of a geneassociated with cell division and drug resistance in Giardiaspecies, the copy number of which seems to be related togenomic rearrangement events (27). Therefore, although wehave not found and did not seek any relationship between thegenotypic rearrangements seen here and any changes in bio-logical characteristics of Giardia species, the genotypic changesseen in the present study do demonstrate the plastic nature ofthe Giardia genome necessary for the development of biolog-ically important genomic rearrangements. The doublet telo-mere sequence of Giardia species (TAGGGTAGGG) seemsto have some sequence homology to the consensus sequence ofthe M13 repeat (GAGGGTGGXGGXTCT) used as a probein this study. It has been speculated that under the low-
stringency hybridization conditions used in this study, theresults may be due to hybridization of the M13 repeat toGiardia telomeres. By hybridization of the M13 repeat and thetelomere repeat to identical PvuII-cut Southern blots, we haveshown this not to be the case even when the stringency waslowered significantly. The telomere repeat produced only fouror five faint smeared bands in the high-molecular-weightregion of the blot even when washed to a final stringency of 6 xSSC at 20'C, which in no way resembled the fingerprint patternproduced by the M13 repeat probe (data not shown).Many fewer clones were recovered than expected from the
cloning experiment. We expected to see a positive cloneculture in 1/10 of the wells, whereas we recovered a positiveculture from only approximately 1/50 of the wells. Assumingthat our original cell counts were reasonably accurate, it canonly be concluded that the cloning procedure itself had aserious effect on the viability of the cells because the startingculture was in log-phase growth and therefore should bcassumed to have contained mostly viable cells. Exposure tohigh oxygen tensions, which are known to be toxic to Giardiatrophozoites and which are an inevitable short-term conse-quence of the cloning procedure, and the physical stressesinvolved in pipetting out the culture are likely to be the majorcauses of this loss of viability. This means that the distributionof viable cells in the microtiter plates would actually be muchless than 0.1 per well, so that each clonal culture is even morelikely to be derived from only one viable cell and hence be trulyclonal. We cannot rule out the possibility that the differencesseen between clones are induced by the cloning process itself,because trophozoites are placed under great stress during theprocedure, which would represent a strong selection pressurenot normally encountered during natural infection or undernormal culture conditions (19). Similar changes are seen,however, in uncloned isolates, and so they are unlikely to bedue solely to the cloning procedure and must, at least in part,represent the natural genetic heterogeneity of Giardia species.
Thus, the genotypic heterogeneity observed in the presentstudy is likely to be a natural phenomenon within the Giardiagenome that may be affected by selective pressures. From theevidence presented here, we consider that at least for theseisolates it is unlikely to be due to selection from a mixedpopulation and is more likely to be due to the genomicplasticity of the organism, resulting in a wide variety of rapidlychanging genotypes upon which selection pressures can act.We cannot, however, be sure that long-term in vitro culturedoes not irretrievably change an isolate by selecting for thoseorganisms within an isolate or indeed those isolates morecapable of growing in a very unnatural environment, with theconsequence that all work such as this on in vitro cultures maybe based on a biased sample of Giardia clones or genotypeswith a restricted range of biological capabilities. This at leastmay explain our failure to find substantive evidence for selec-tion from truly mixed populations. These findings have impli-cations for the development of typing systems for Giardiaspecies and in the assignment of virulence characteristics toisolates with apparently different genotypes.
ACKNOWLEDGMENTS
This work was supported by a grant from The Joint Research Boardof St. Bartholomew's Hospital (project 9G69). In addition, M.J.G.F.and P.H.K. gratefully acknowledge the financial support of the Wel-come Trust. London, United Kingdom.
REFERENCES1. Adam, R. D. 1992. Chromosome-size variation in Giarlia lamblia:
the role of rDNA repeats. Nucleic Acids Res. 20:3057-3061.
Vol- 62, 1994

1880 CARNABY ET AL.
2. Adam, R. D., A. Aggarwal, A. A. Lal, V. F. de la Cruz, T.McCutchan, and T. E. Nash. 1988. Antigenic variation of a
cysteine-rich protein in Giardia lamblia. J. Exp. Med. 167:109-118.3. Andrews, R. H., M. Adams, P. F. L. Boreham, G. Mayrhofer, and
B. P. Meloni. 1989. Giardia intestinalis: electrophoretic evidencefor a species complex. Int. J. Parasitol. 19:183-190.
4. Andrews, R. H., N. B. Chilton, and G. Mayrhofer. 1992. Selectionof specific genotypes of Giardia intestinalis by growth in vitro andin vivo. Parasitology 105:375-386.
5. Baum, K. F., R. L. Berens, R. H. Jones, and J. J. Marr. 1988. Anew method for cloning Giardia lamblia, with a discussion of thestatistical considerations of limiting dilution. J. Parasitol. 74:267-269.
6. Bertram, M. A., E. A. Meyer, J. D. Lile, and S. A. Morse. 1983. Acomparison of isozymes of five axenic Giardia isolates. J. Parasitol.69:793-801.
7. Bhatia, V. N., and D. C. Warhurst. 1981. Hatching and subsequentcultivation of cysts of Giardia intestinalis in Diamond's medium. J.Trop. Med. Hyg. 84:45.
8. Borst, P., and D. R. Greaves. 1987. Programmed gene rearrange-
ments altering gene expression. Science 235:658-667.9. Carnaby, S., T. D. McHugh, P. H. Katelaris, and M. J. G.
Farthing. 1991. Genomic evidence for multiple strain involvementin Giardia lamblia infection. Gut 32:A563.
10. Cowman, A. F., R. B. Saint, R. L. Coppel, G. B. Brown, R. F.Anders, and D. J. Kemp. 1985. Conserved sequences flank variabletandem repeats in two S-antigen genes of P. falciparum. Cell40:775-783.
11. De Jonckheere, J. F., A. C. Majewska, and W. Kasprzalk 1990.Giardia isolates from primates and rodents display the same
molecular polymorphism as human isolates. Mol. Biochem. Para-sitol. 39:23-28.
12. Farthing, M. J. G., and G. T. Keusch (ed.). 1989. Enteric infection:mechanisms, manifestations and management. Chapman & HallLtd., London.
13. Farthing, M. J. G., G. T. Keusch, and M. C. Carey. 1985. Effectsof bile and bile salts on growth and membrane lipid uptake byGiardia lamblia. J. Clin. Invest. 76:1727-1732.
14. Feinberg, A. P., and B. Vogelstein. 1983. A technique for radiola-beling DNA restriction enzyme fragments to high specific activity.Anal. Biochem. 132:6-13.
15. Fortess, E., and E. A. Meyer. 1976. Isolation and axenic cultivationof Giardia trophozoites from the guinea pig. J. Parasitol. 62:689.
16. Keister, D. B. 1983. Axenic culture of Giardia lamblia in TYI-S-33medium supplemented with bile. Trans. R. Soc. Trop. Med. Hyg.77:487-488.
17. Kemp, D. J., R. L. Coppel, and R. F. Anders. 1987. Repetitive
genes and proteins of malaria. Annu. Rev. Microbiol. 41:181-208.18. Le Blancq, S. M., S. H. Korman, and H. T. Van der Ploeg. 1991.
Frequent rearrangements of rRNA encoding chromosomes inGiardia lamblia. Nucleic Acids Res. 19:4405-4412.
19. Mayrhofer, G., R. H. Andrews, P. L. Ey, M. J. Albert, T. R.Grimmond, and D. J. Merry. 1992. The use of suckling mice toisolate and grow Giardia from mammalian faecal specimens forgenetic analysis. Parasitology 105:255-263.
20. Meloni, B. P., A. J. Lymbery, and R. C. A. Thompson. 1988.Isoenzyme electrophoresis of 30 isolates of Giardia from humansand felines. Am. J. Trop. Med. Hyg. 38:65-73.
21. Meyer, E. A. 1976. Giardia lamblia: isolation and axenic cultiva-tion. Exp. Parasitol. 39:101-105.
22. Smith, P. P., F. D. Gillin, W. M. Spira, and T. E. Nash. 1982.Chronic giardiasis: studies on drug sensitivity, toxin production,and host immune response. Gastroenterology 83:797-803.
23. Sogin, M. L., J. H. Gunderson, H. J. Elwood, R. A. Alonso, andD. A. Peattie. 1989. Phylogenetic meaning of the kingdom concept:an unusual ribosomal RNA from Giardia lamblia. Science 243:75-77.
24. Southern, E. M. 1975. Detection of specific sequences amongDNA fragments separated by gel electrophoresis. J. Mol. Biol.98:503-517.
25. Tanaka, M., H. M. Gu, D. J. Bzik, W. B. Li, and J. Inselburg. 1990.Mutant dihydrofolate reductase-thymidylate synthase genes inpyrimethamine-resistant Plasmodium falciparum with polymorphicchromosome duplications. Mol. Biochem. Parasitol. 42:83-92.
26. Triglia, T., S. J. Foot, D. J. Kemp, and A. F. Cowman. 1991.Amplification of the multidrug resistance gene pfmdrl in Plasmo-dium falciparum has arisen as multiple independent events. Mol.Cell. Biol. 11:5244-5250.
27. Upcroft, J. A., A. Healey, D. G. Murray, P. F. L. Boreham, and P.Upcroft. 1992. A gene associated with cell division and drugresistance in Giardia intestinalis. Parasitology 104:397-405.
28. Van der Ploeg, L. H. T., D. C. Schwartz, C. R. Cantor, and P.Borst. 1984. Antigenic variation in Trypanosoma brucei analysedby electrophoretic separation of chromosome sized DNA mole-cules. Cell 37:77-84.
29. Vassart, G., M. Georges, R. Monsieur, H. Brocas, A. S. Lequarre,and D. Christophe. 1987. A sequence of M13 phage detectshypervariable minisatellites in human and animal DNAs. Science235:683-684.
30. Weiss, J. B., H. Van Keulen, and T. E. Nash. 1992. Classification ofsubgroups of Giardia lamblia based upon ribosomal RNA genesequence using the polymerase chain reaction. Mol. Biochem.Parasitol. 54:73-86.
INFECT. IMMUN.
The minimal kinome of Giardia lamblia illuminates early kinase evolution and unique parasite biology
Prospective cohort study into post-disaster benzodiazepine use demonstrated only short-term increase