Évolution de la structure diamétrale et de la composition des peuplements mixtes de sapin baumier...
TRANSCRIPT

Évolution de la structure diamétrale et de lacomposition des peuplements mixtes de sapinbaumier et d’épinette rouge de la forêt primitiveaprès une coupe à diamètre limite sur l’Aired’observation de la rivière Ouareau
Mathieu Fortin, Jean Bégin et Louis Bélanger
Résumé : La structure diamétrale et la composition des peuplements mixtes de sapin baumier (Abies balsamea (L.)Mill.) et d’épinette rouge (Picea rubens Sarg.) de la forêt primitive ont été caractérisées au moyen de 41 placettes.Nous avons regroupé la table de peuplement en cinq groupes de classes de diamètre et nous avons caractérisé la sur-face terrière de quatre espèces dans chaque groupe. L’analyse révèle que la structure diamétrale tend vers un J inversé.L’épinette rouge et le bouleau jaune (Betula alleghaniensis Britt.) sont plus abondants parmi les tiges de forts diamè-tres alors que le sapin baumier est plus abondant parmi les gaules. La comparaison avec 18 placettes permanentes issusd’une coupe à diamètre limite démontre que les placettes ont retrouvé, 47 ans après la coupe, une structure semblableà celle de la forêt primitive. Toutefois, les proportions d’épinette ont diminué alors que celles de sapin baumier et debouleau à papier (Betula papyrifera Marsh.) ont augmenté. La raréfaction de l’épinette rouge n’est pas due à une régé-nération déficiente, mais à un prélèvement trop important. Un jardinage par pied d’arbre serait un traitement adéquat sil’intervention était réalisée dans l’optique de protéger les tiges d’épinette rouge jusqu’à un certain diamètre.
Abstract: Using data from 41 plots, the diameter structure and composition of old-growth mixed stands composedmainly of balsam fir (Abies basalmea (L.) Mill.) and red spruce (Picea rubens Sarg.) were characterized. The stand ta-ble was organized in five groups of diameter classes, and we characterized the basal area of four tree species in eachgroup. Analysis showed that diameter structure tends toward a reverse J-shaped curve. Red spruce and yellow birch(Betula alleghaniensis Britt.) were more abundant among large diameter trees while balsam fir was more abundantamong saplings. A comparison with 18 permanent plots from a diameter-limit cutting showed that plots had come backto a structure similar to that of the old-growth forest 47 years after cutting. However, spruce ratios had decreased whilethose of balsam fir and paper birch (Betula papyrifera Marsh.) had increased. Red spruce decrease is not due to a lackof regeneration but to overcutting. Single-tree selection method would be a suitable treatment if aimed at protecting redspruce trees up to a given diameter.
[Journal translation] Fortin et al. 704
Introduction
L’épinette rouge (Picea rubens Sarg.) et le sapin baumier(Abies balsamea (L.) Mill.) forment des peuplements mixtessur l’ensemble de la zone tempérée du Québec méridional.On les retrouve abondamment dans les domaines bioclimati-ques de l’érablière à tilleul et de l’érablière à bouleau jaune(Grondin et al. 1996; Robitaille et Saucier 1998). Ces peu-plements sont également caractéristiques de la Région fores-tière acadienne (Rowe 1972). Ils sont donc présents dans lesprovinces maritimes du Canada et les états du Nord-Est
américain. Bien que ces peuplements couvrent toujoursd’importantes superficies, Gordon (1998) signale que le tauxd’occupation de l’épinette rouge ne représente plus que lecinquième de ce qu’il était dans la forêt précoloniale surl’ensemble de son aire de distribution. La coupe à blanc et laconversion de peuplements seraient principalement respon-sables de cette diminution généralisée des proportionsd’épinette rouge (Gordon 1998).
Offrant une qualité élevée pour le bois d’œuvre, ces peu-plements ont été exploités dès le milieu du 19e siècle (Heim-burger 1941; Ray 1941; Seymour 1992, 1995). L’essor del’industrie des pâtes et papiers au début du 20e siècle a eupour effet d’en intensifier la récolte. Les coupes d’intensitélégère, pratiquées jusque là, ont été graduellement rempla-cées par des coupes à diamètre limite et des coupes à blanc(Heimburger 1941; Ray 1941, 1956).
Les effets de la coupe à blanc sur ces peuplements sontaujourd’hui connus. Les conditions d’humidité et de tempé-rature qui sont observées après cette intervention créent unenvironnement défavorable à l’implantation et au développe-
Can. J. For. Res. 33: 691–704 (2003) doi: 10.1139/X02-205 © 2003 CNRC Canada
691
Reçu le 5 décembre 2001. Accepté le 29 novembre 2002.Publié sur le site Web des Presses scientifiques du CNRC, àhttp://rcrf.cnrc.ca, le 24 mars 2003.
M. Fortin1, J. Bégin et L. Bélanger. Département dessciences du bois et de la forêt, Université Laval, Sainte-Foy,QC G1K 7P4, Canada.
1. Auteur correspondant(courriel : [email protected]).
I:\cjfr\cjfr3304\X02-205.vpWednesday, March 19, 2003 4:48:20 PM
Color profile: DisabledComposite Default screen

ment des semis résineux (Frank et Bjorkbom 1973; Seymour1995). Lorsque la régénération préétablie est insuffisante, ils’ensuit un envahissement par la végétation compétitrice(Ray 1941; Seymour 1995). Lorsque la régénération rési-neuse préétablie est abondante, les semis de sapin surpassenten nombre ceux de l’épinette rouge (Ray 1941; Seymour1995; Grondin et al. 1996). Les peuplements de secondevenue contiennent, en conséquence, une plus faible propor-tion d’épinette rouge (Ray 1941; Seymour 1992). Par ail-leurs, la coupe à blanc a un effet marqué sur la structured’âge de ces peuplements en homogénéisant l’âge des tiges(Seymour 1992). Les peuplements issus de cette interventionont généralement une structure d’âge équienne (Oliver etLarson 1996; Smith et al. 1997).
Depuis les années 1950, les forestiers entrevoient les cou-pes partielles comme une alternative intéressante à la coupeà blanc pour l’aménagement de ces peuplements. Les coupespartielles permettent de maintenir une structure irrégulièreou étagée tout en préservant des conditions favorables àl’installation de la régénération des espèces résineuses(Westveld 1953; Ray 1956; Hatcher 1959a, 1960; Frank etBlum 1978; Seymour 1992; Brissette 1996). Une coupe par-tielle à diamètre limite réalisées dans les années 1950 à laForêt expérimentale du lac Édouard a même permis d’aug-menter les proportions d’épinette rouge dans les sapinières àbouleau jaune (Ménard 1999).
La compréhension de la structure et de la composition despeuplements mixtes de sapin baumier et d’épinette rougedans la forêt primitive demeure toutefois imprécise (Sey-mour 1992). Plusieurs auteurs suggèrent que la structure ir-régulière y était dominante (Oosting et Billings 1951; Leak1975; Seymour 1995). Outre cela, on ignore réellement laproportion de chaque espèce au sein des peuplements et lavariabilité qui lui est associée. Avec l’exploitation forestièrequi dure depuis plus d’un siècle, on considère que les peu-plements de sapin baumier et d’épinette rouge représentatifsde la forêt primitive sont aujourd’hui très rares (Oosting etBillings 1951; Foster et Reiners 1983; Seymour 1992). Aumilieu du 20e siècle, Hatcher (1954) et Ray (1956) préci-saient que l’Aire d’observation de la rivière Ouareau abritaitl’un des derniers tenants encore intacts de ce type de forêtdans la province de Québec. Les données recueillies dans cedispositif de recherche constituent une information privi-légiée sur les peuplements mixtes de sapin baumier et d’épi-nette rouge de la forêt primitive. Une coupe à diamètrelimite utilisant un diamètre de récolte unique pour toutes lesessences résineuses a été réalisée au moment de l’implanta-tion du dispositif. Le suivi de placettes permanentes permetd’observer les impacts de ce type de coupe partielle sur lacomposition et la structure diamétrale de la forêt primitive.
Nous posons comme hypothèse qu’une coupe à diamètrelimite comme celle réalisée sur l’Aire d’observation de la ri-vière Ouareau pourrait être un traitement sylvicole efficacedans une stratégie d’aménagement écosystémique, c’est-à-dire, une stratégie d’aménagement visant à imiter le régimede perturbations naturelles. Les objectifs de cette étude sont(i) de qualifier la structure diamétrale et la composition despeuplements mixtes de sapin baumier et d’épinette rouge dela forêt primitive et (ii) de comparer cette structure et cettecomposition à celles observées après une coupe à diamètrelimite sur une période de 47 ans. Nous utilisons la composi-
tion et la structure diamétrale de la forêt primitive comme cri-tères pour juger l’acceptabilité de cette coupe dans une stra-tégie d’aménagement écosystémique. Les recommandationsémises à partir des résultats obtenus viseront à améliorer lesperformances du traitement dans ce même contexte d’amé-nagement.
Aire d’étude
Cadre écologiqueL’Aire d’observation de la rivière Ouareau (46° 26′ N.,
74° 10′ O.) est située dans la région écologique des Hautescollines du bas Saint-Maurice (3c), à la limite septentrionaledu sous-domaine bioclimatique de l’érablière à bouleaujaune de l’est (Grondin et al. 1996; Gosselin et al. 1998;Robitaille et Saucier 1998). Le climat y est frais et humide(Hatcher 1954; Robitaille et Saucier 1998). L’altitudemoyenne est d’environ 500 m. Les températures et précipita-tions annuelles moyennes sont respectivement de 3,0 °C et1096 mm. Les dépôts de surface qui caractérisent l’Aired’observation sont représentatifs de la région écologique :des tills d’épaisseur variable et des dépôts fluvioglaciairescouvrent la majeure partie de sa superficie (Gagnon et Mar-cotte 1980; Gosselin et al. 1998; Robitaille et Saucier 1998).
Le cadre écologique des Basses-Laurentides est largementdétaillé par plusieurs auteurs (Gagnon et Marcotte 1980; Bé-langer 1985; Lalumière et Thibault 1988; Grondin et al.1996; Gosselin et al. 1998). Les peuplements mixtes de sa-pin baumier et d’épinette rouge sont généralement présentssur les tills mésiques au bas des pentes et en terrains plats(Heimburger 1941; Grondin et al. 1996; Gosselin et al.1998). On les retrouve également sur les tills hydriques, lessols minces et les dépôts fluvioglaciaires caractérisés par desdrainages rapides (Heimburger 1941; Bélanger 1985; Gron-din et al. 1996; Gosselin et al. 1998). Ce cadre écologiqueest similaire à celui de la Région forestière acadienne (Rowe1972). L’Aire d’observation de la rivière Ouareau en cons-titue une version nordique. Elle se démarque de ce cadre(i) par l’absence de la pruche (Tsuga canadensis (L.) Car-rière) dans les peuplements mixtes et résineux et (ii) par unetrès faible abondance du hêtre à grandes feuilles (Fagusgrandifolia Ehrh.) dans les peuplements mixtes et feuillus(Hatcher 1954).
Dynamique forestièreLe régime de perturbations des peuplements mixtes de sa-
pin baumier et d’épinette rouge est décrit comme une suc-cession de trouées de petites dimensions causées par la mortd’un individu ou d’un groupe restreint d’individus (Battles etFahey 1996, 2000; Hébert 2000). Les défoliations de la tor-deuse des bourgeons de l’épinette (Choristoneura fumife-rana Clem.), la carie et le chablis comptent parmi lesprincipales sources de mortalité (McLintock 1954; Hatcher1959b; Seymour 1992). On croit que des chablis de superfi-cies considérables pouvaient survenir occasionnellement surles stations où l’enracinement est superficiel (Seymour1992). Quant au cycle de feu, il demeure un sujet peu docu-menté pour le domaine bioclimatique de l’érablière à bou-leau jaune au Québec (Grondin et al. 1996). Dans le reste dela Forêt acadienne, on estime que le cycle de feu était supé-rieur à 760 ans alors que l’intervalle de récurrence des cha-blis de plus de 25 ha était de 1150 ans (Lorimer 1977; Fahey
© 2003 CNRC Canada
692 Can. J. For. Res. Vol. 33, 2003
I:\cjfr\cjfr3304\X02-205.vpWednesday, March 19, 2003 4:48:21 PM
Color profile: DisabledComposite Default screen

et Reiners 1981). Les trouées étaient le type de perturbationprédominant dans les peuplements de la forêt primitive(Lorimer 1977; Fahey et Reiners 1981; Foster et Reiners1983; Seymour 1992). Dans les vieux peuplements mixtesde sapin baumier et d’épinette rouge, la dimension destrouées est généralement inférieure à 200 m2 et le taux deremplacement du couvert (« forest turnover rate », p. ex.,Green 1996) est estimé à 303 ans (Foster et Reiners 1986;Worrall et Harrington 1988; Battles et Fahey 1996, 2000).
Comparativement à la régénération du sapin baumier, lessemis d’épinette rouge sont rarement abondants. Grondin etal. (1996) considèrent que cette espèce compte pour au plus15 % du nombre de tiges en régénération dans la sapinière àépinette rouge. Comme le sapin est de loin l’espèce la plusvulnérable à la tordeuse, au chablis et à la carie (McLintock1954; Hatcher 1959b; Seegrist et Arner 1982; Frank 1990;Seymour 1992; Grondin et al. 1996; MacLean et MacKinnon1997; Ruel 2000; Meunier et al. 2002), la dynamique natu-relle semble (i) limiter l’abondance du sapin baumier dans lecouvert dominant malgré sa capacité à produire une régéné-ration abondante sous couvert, (ii) favoriser le dévelop-pement d’une structure d’âge à plusieurs cohortes et(iii) favoriser le développement d’une structure diamétraleirrégulière telle que définie par Smith et al. (1997).
La composition des peuplements de la forêt primitive at-teindrait un équilibre entre la longévité des épinettes rougeset la régénération abondante du sapin baumier. Les différen-tes cohortes initialement composées d’une majorité de sapinverraient leur proportion d’épinette rouge augmenter avec letemps, les tiges de sapin étant éliminées progressivement parla tordeuse, la carie et le chablis. Au Québec, plusieurs au-teurs définissent la sapinière à épinette rouge comme la vé-gétation potentielle, c’est-à-dire, la végétation de fin desuccession, sur les stations forestières en terrain plat dansl’érablière à bouleau jaune de l’est (Grondin et al. 1996;Gosselin et al. 1998).
Historique des perturbationsDepuis 1938, les registres de la Société de la protection
des forêts contre les insectes et maladies du Québec recen-sent deux périodes épidémiques de tordeuse dans ce secteur.L’épisode de 1944–1954 est qualifié d’infestation légère tan-dis que celui de 1972–1986 est considéré comme une infes-tation sévère. Des pulvérisations aériennes d’insecticideschimiques ont été réalisées sur une base opérationnelle entre1972 et 1976. En 1999, des carottes ont été prélevées à hau-teur de poitrine à l’aide d’une sonde de Pressler dans 14 pla-cettes de l’Aire d’observation. Toutes les tiges d’un diamètreà hauteur de poitrine (DHP) de plus de 50 mm ont étééchantillonnées. L’analyse de ces carottes a révélé un ralen-tissement de l’accroissement en diamètre chez le sapin etl’épinette pour la dernière épidémie, ce ralentissement étantde loin plus important chez le sapin baumier.
En plus des épidémies de tordeuse, on signale égalementle dépérissement du bouleau à papier (Betula papyriferaMarsh.) sur l’Aire d’observation de la rivière Ouareau versle milieu du 20e siècle (Hatcher 1954). Comparativement aubouleau à papier, le bouleau jaune (Betula alleghaniensisBritt.) a été peu affecté (Hatcher 1954). Quant aux incendiesforestiers, aucune trace de feu n’a été répertoriée sur l’Aired’observation avant et depuis son implantation. Le dernierincendie remonte donc à une date indéterminée.
Matériel et méthode
Base de donnéesPrès des deux tiers de la superficie de ce qui allait devenir
l’Aire d’observation était toujours à l’état vierge en 1948(Hatcher 1954). La récolte forestière a été réalisée durant lessaisons hivernales entre 1948 et 1956. Le traitement alors envigueur était la récolte des résineux dont le diamètre à lasouche était équivalent ou supérieur à 18 cm (7 po). L’abat-tage a été effectué à l’aide de scies manuelles, l’utilisationde la scie mécanique restait encore marginale à cette époque.Quant au débardage, il a été réalisé à l’aide de chevaux. En1953, au moment de l’implantation, environ le tiers de la su-perficie du dispositif était toujours intact. À l’hiver de 1956,les opérations se terminaient dans le secteur. On considèrequ’environ 60 % du volume marchand (9,1 cm et plus) ontété récoltés lors de cette coupe à diamètre limite (Hatcher1954), la majorité de ce volume étant composée d’épinetterouge.
Le dispositif a été implanté en 1953 et mesuré à nouveauen 1963, entre 1974 et 1977 et, finalement, entre 1998 et1999. Le temps écoulé depuis la coupe à diamètre limiten’est pas identique pour chaque placette étant donné que lesopérations forestières se sont déroulées sur plusieurs saisons.En moyenne, les relevés de 1963 représentent l’état de la fo-rêt 12 ans après la coupe avec une étendue variant entre 8 et13 ans. Les relevés de 1974–1977 représentent l’état 24 ansaprès coupe (21–27 ans) et ceux de 1998–1999, l’état 47 ansaprès coupe (43–49 ans). Nous disposons donc de trois inter-valles pour documenter l’évolution de la structure diamétraleet de la composition après cette coupe à diamètre limite.
Les placettes du dispositif ont une superficie de 404 m2
(0,1 acre) chacune. À l’intérieur de chacune d’elles, toutesles tiges d’essences commerciales dont le DHP (1,37 m) estéquivalent ou supérieur à 1,5 cm (0,6 po) ont été dénom-brées par classe de 2,5 cm (1 po). Le dispositif a été créé aumoment où le système impérial était encore en vigueur. Tousles mesurages subséquents ont donc été faits en suivant cesystème.
En plus du dénombrement des tiges, chaque pointd’observation a été classifié selon les types forestiers deHeimburger (1941). Compte tenu des données disponibles,seules les placettes associées au type forestier Oxalis–Cornus ont été considérées dans l’analyse. Ce type forestiercorrespond essentiellement aux stations mésiques situées aubas de pente et en terrains plats. Les dépôts sont générale-ment des tills épais de faible pierrosité. Gosselin et al.(1998) définissent la végétation potentielle de ces stationscomme la bétulaie jaune à sapin (MJ2x).
La base de données comprend 41 placettes décrivant la fo-rêt primitive et 18 placettes décrivant l’état 12, 24 et 47 ansaprès la coupe à diamètre limite. Le tableau 1 présente unsommaire de ces placettes. Les 41 placettes établies dans lespeuplements vierges en 1953 ont été récoltées entre 1963 et1967 de sorte que leur suivi a été abandonné depuis. Nousprésumons dans cette comparaison que la composition et lastructure moyennes des deux groupes étaient similaires avantla coupe étant donné que (i) toutes les placettes apparte-naient à la forêt primitive avant 1948 (Hatcher 1954), (ii) lescaractéristiques stationnelles des placettes sont semblables etfinalement, (iii) le volume moyen avant la coupe était prati-quement identique pour ces deux groupes (Hatcher 1954).
© 2003 CNRC Canada
Fortin et al. 693
I:\cjfr\cjfr3304\X02-205.vpWednesday, March 19, 2003 4:48:22 PM
Color profile: DisabledComposite Default screen

© 2003 CNRC Canada
694 Can. J. For. Res. Vol. 33, 2003
La localisation des placettes à l’intérieur du dispositif est il-lustrée à la figure 1.
MéthodeL’Aire d’observation est un dispositif de recherche systé-
matique qui a été établi dans l’optique de documenter l’évo-lution d’une forêt suite à une coupe à diamètre limite. Lesplacettes et traitements n’ont pas été distribués au hasard etcette absence de randomisation empêche l’utilisation des sta-tistiques inférencielles. Cette étude traite un cas unique biendocumenté et les comparaisons ont été effectuées à partir destatistiques descriptives.
Une approche en fonction de la dimension des tiges a étéutilisée. Celle-ci consiste à diviser la structure diamétrale ensous-groupes représentant l’un des cinq groupes diamétrauxprésentés au tableau 2. Les divisions retenues sont des inter-valles de trois classes de pouce. Ces intervalles ont été déter-minés arbitrairement et font référence à des dimensionsgénéralement utilisées en sylviculture, notamment pour lesgaules et les petites tiges marchandes. Nous avons définil’abondance des espèces par leur surface terrière (m2·ha–1).Pour chacun des groupes diamétraux, nous avons analysécette abondance (i) pour toutes les essences confondues et(ii) pour chacune des essences principales : le sapin baumier,l’épinette rouge, le bouleau jaune et le bouleau à papier.
En premier lieu, nous avons observé et défini la structurediamétrale de la forêt primitive. Sa composition a ensuite étédécrite pour chacun des groupes diamétraux. Par la suite,nous avons comparé la structure diamétrale de la forêt primi-tive et celle de la forêt issue de la coupe à diamètre limite.Pour cette comparaison, nous avons utilisé la distribution dela surface terrière par classe de pouce pour ne pas diminuerl’importance des grosses tiges qui sont généralement peu re-présentées. En dernier lieu, dans chacun des groupes diamé-traux, nous avons observé l’évolution de la surface terrièreaprès la coupe pour toutes les essences confondues et pourchacune des espèces principales. Nous avons comparé cetteévolution à la surface terrière observée dans la forêt primi-tive. Nous nous sommes basés sur les moyennes arithméti-ques et l’erreur type de ces moyennes.
Résultats et analyses
Structure diamétrale et composition de la forêtprimitive
La distribution diamétrale moyenne de la forêt primitive
adopte la forme d’un J inversé (fig. 2). Cependant, peu deplacettes correspondent exactement à cette distribution.Parmi l’ensemble des structures diamétrales observées, 34des 41 relevés affichent une distribution qui s’apparentegrossièrement à un J inversé. Les placettes 138 et 139 ensont des exemples (fig. 2c et 2d). Les sept autres relevésprésentent des distributions très irrégulières comme celle dela placette 135 (fig. 2b).
La distribution de la surface terrière par essence et pargroupe diamétral est présentée au tableau 3. En moyenne,l’épinette rouge représente 47 % de la surface terrière, le sa-pin baumier, 30 %, le bouleau jaune, 13 %, le bouleau à pa-pier, 3 % et les autres essences, principalement l’érable àsucre (Acer saccharum Marsh.), l’érable rouge (Acer ru-brum L.) et le thuya (Thuja occidentalis L.), 7 %. Les grou-
Variable n Minimum Moyenne MaximumÉcarttype
Surface terrière (m2·ha–1)Forêt primitive 41 10,3 29,8 58,1 9,212 ans après coupe 18 3,7 16,4 37,7 8,324 ans après coupe 18 13,2 26,2 38,8 7,647 ans après coupe 18 23,4 33,8 46,1 6,1Diamètre moyen quadratique (cm)Forêt primitive 41 6,5 14,4 20,5 3,512 ans après coupe 18 5,1 10,3 17,7 3,124 ans après coupe 18 5,9 9,1 13,4 2,147 ans après coupe 18 8,1 13,0 19,2 3,4
Tableau 1. Caractéristiques dendrométriques des placettes pour la forêt primitive etchacun des intervalles de temps après coupe.
Fig. 1. Localisation des placettes dans le dispositif de recherche.
Groupe diamétral DHP
Très grosses tiges ≥32,0 cm (classes ≥13 po)Grosses tiges 24,4–31,9 cm (classes 10 à 12 po)Moyennes tiges 16,8–24,3 cm (classes 7 à 9 po)Petites tiges 9,1–16,7 cm (classes 4 à 6 po)Gaules 1,5–9,0 cm (classes 1 à 3 po)
Tableau 2. Divisions de la structure diamétrale pourl’analyse.
I:\cjfr\cjfr3304\X02-205.vpWednesday, March 19, 2003 4:48:23 PM
Color profile: DisabledComposite Default screen
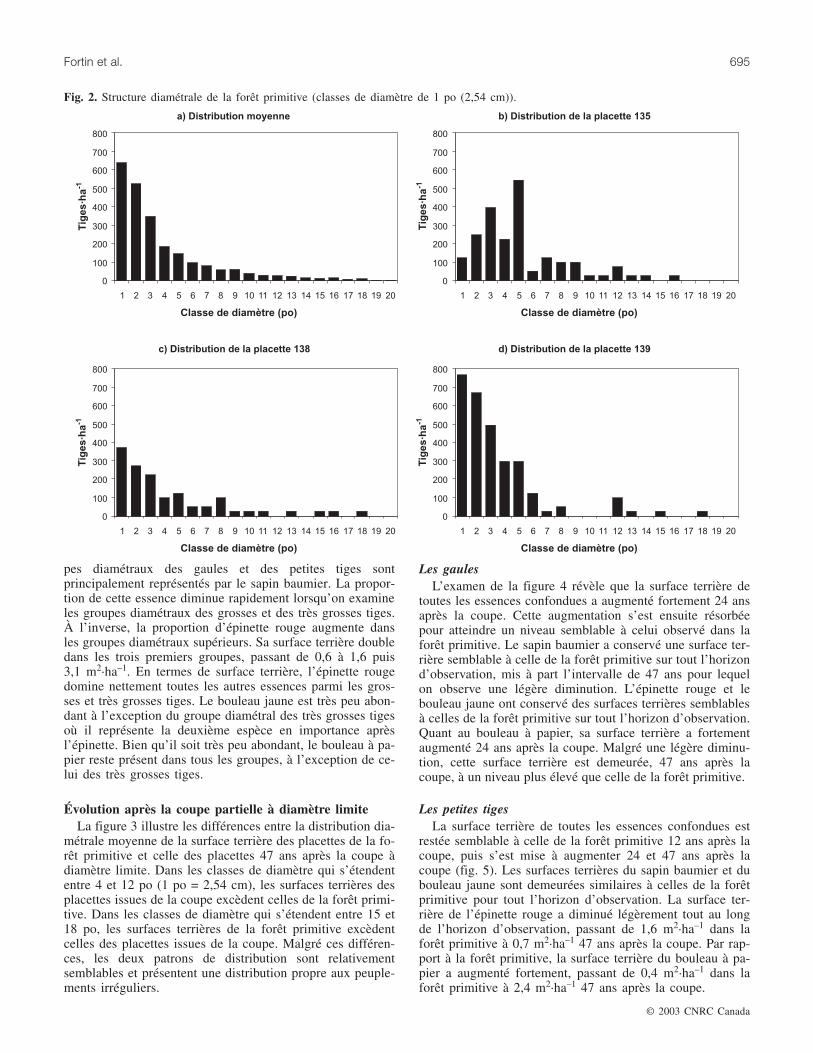
pes diamétraux des gaules et des petites tiges sontprincipalement représentés par le sapin baumier. La propor-tion de cette essence diminue rapidement lorsqu’on examineles groupes diamétraux des grosses et des très grosses tiges.À l’inverse, la proportion d’épinette rouge augmente dansles groupes diamétraux supérieurs. Sa surface terrière doubledans les trois premiers groupes, passant de 0,6 à 1,6 puis3,1 m2·ha–1. En termes de surface terrière, l’épinette rougedomine nettement toutes les autres essences parmi les gros-ses et très grosses tiges. Le bouleau jaune est très peu abon-dant à l’exception du groupe diamétral des très grosses tigesoù il représente la deuxième espèce en importance aprèsl’épinette. Bien qu’il soit très peu abondant, le bouleau à pa-pier reste présent dans tous les groupes, à l’exception de ce-lui des très grosses tiges.
Évolution après la coupe partielle à diamètre limiteLa figure 3 illustre les différences entre la distribution dia-
métrale moyenne de la surface terrière des placettes de la fo-rêt primitive et celle des placettes 47 ans après la coupe àdiamètre limite. Dans les classes de diamètre qui s’étendententre 4 et 12 po (1 po = 2,54 cm), les surfaces terrières desplacettes issues de la coupe excèdent celles de la forêt primi-tive. Dans les classes de diamètre qui s’étendent entre 15 et18 po, les surfaces terrières de la forêt primitive excèdentcelles des placettes issues de la coupe. Malgré ces différen-ces, les deux patrons de distribution sont relativementsemblables et présentent une distribution propre aux peuple-ments irréguliers.
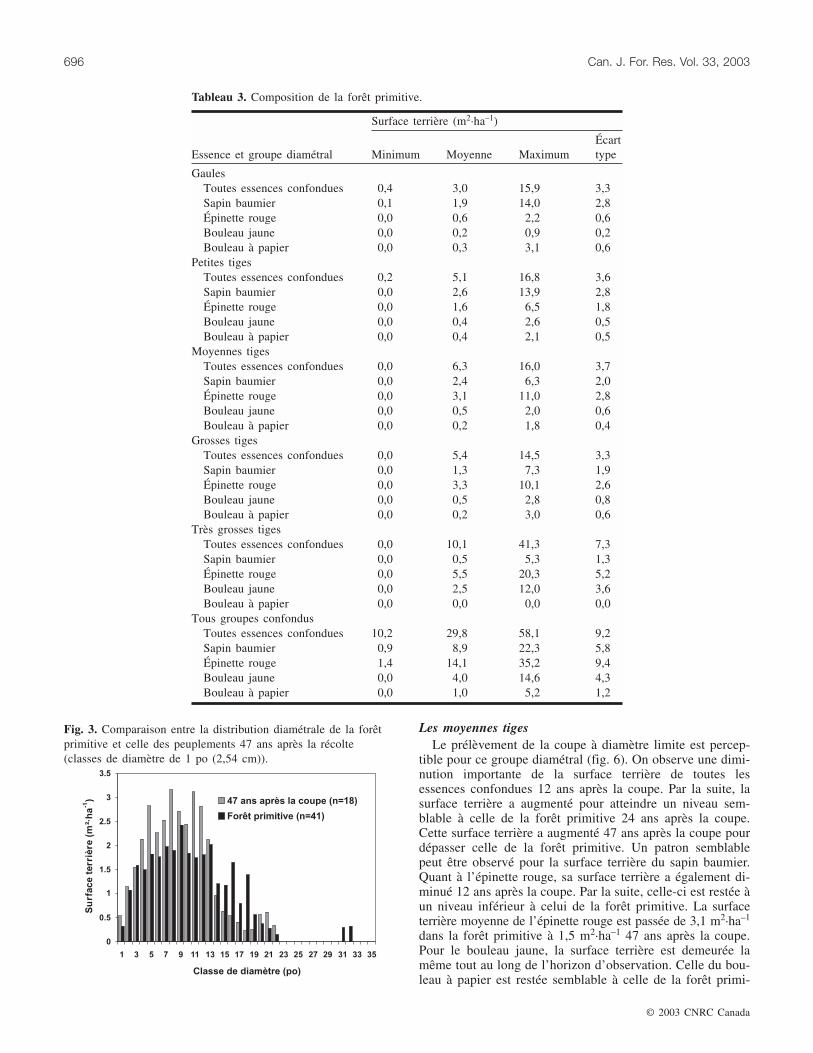
Les gaulesL’examen de la figure 4 révèle que la surface terrière de
toutes les essences confondues a augmenté fortement 24 ansaprès la coupe. Cette augmentation s’est ensuite résorbéepour atteindre un niveau semblable à celui observé dans laforêt primitive. Le sapin baumier a conservé une surface ter-rière semblable à celle de la forêt primitive sur tout l’horizond’observation, mis à part l’intervalle de 47 ans pour lequelon observe une légère diminution. L’épinette rouge et lebouleau jaune ont conservé des surfaces terrières semblablesà celles de la forêt primitive sur tout l’horizon d’observation.Quant au bouleau à papier, sa surface terrière a fortementaugmenté 24 ans après la coupe. Malgré une légère diminu-tion, cette surface terrière est demeurée, 47 ans après lacoupe, à un niveau plus élevé que celle de la forêt primitive.
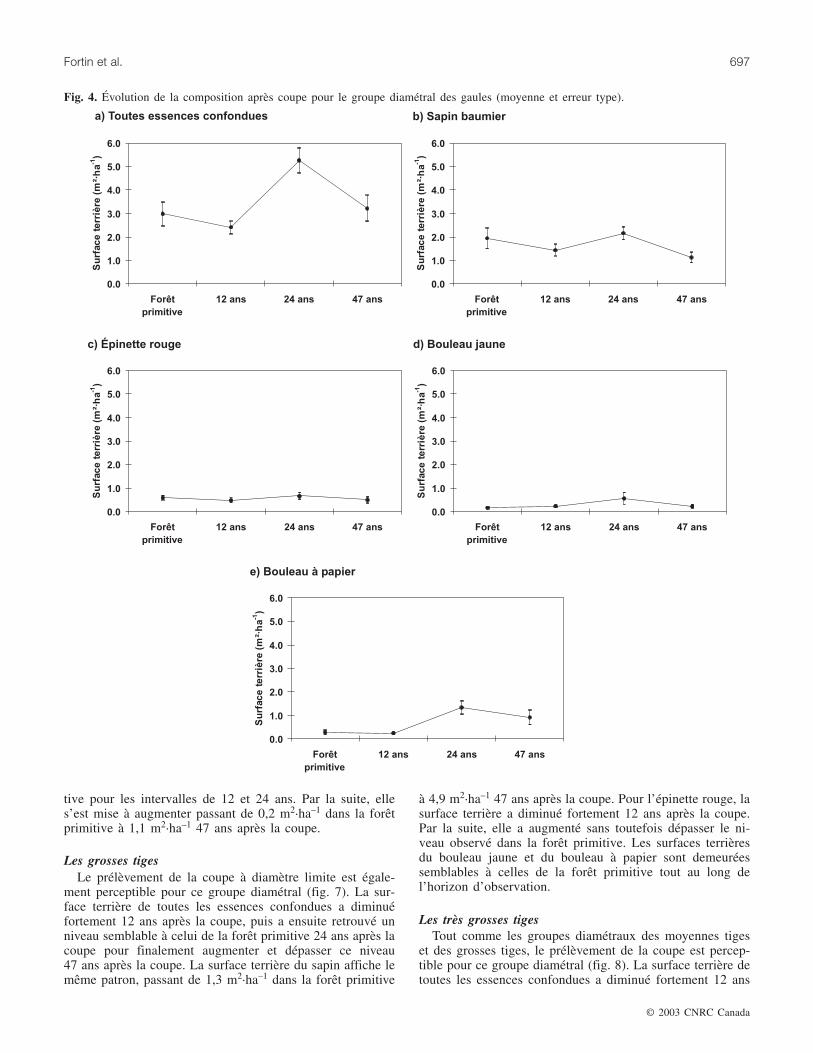
Les petites tigesLa surface terrière de toutes les essences confondues est
restée semblable à celle de la forêt primitive 12 ans après lacoupe, puis s’est mise à augmenter 24 et 47 ans après lacoupe (fig. 5). Les surfaces terrières du sapin baumier et dubouleau jaune sont demeurées similaires à celles de la forêtprimitive pour tout l’horizon d’observation. La surface ter-rière de l’épinette rouge a diminué légèrement tout au longde l’horizon d’observation, passant de 1,6 m2·ha–1 dans laforêt primitive à 0,7 m2·ha–1 47 ans après la coupe. Par rap-port à la forêt primitive, la surface terrière du bouleau à pa-pier a augmenté fortement, passant de 0,4 m2·ha–1 dans laforêt primitive à 2,4 m2·ha–1 47 ans après la coupe.
© 2003 CNRC Canada
Fortin et al. 695
Fig. 2. Structure diamétrale de la forêt primitive (classes de diamètre de 1 po (2,54 cm)).
I:\cjfr\cjfr3304\X02-205.vpWednesday, March 19, 2003 4:48:24 PM
Color profile: DisabledComposite Default screen
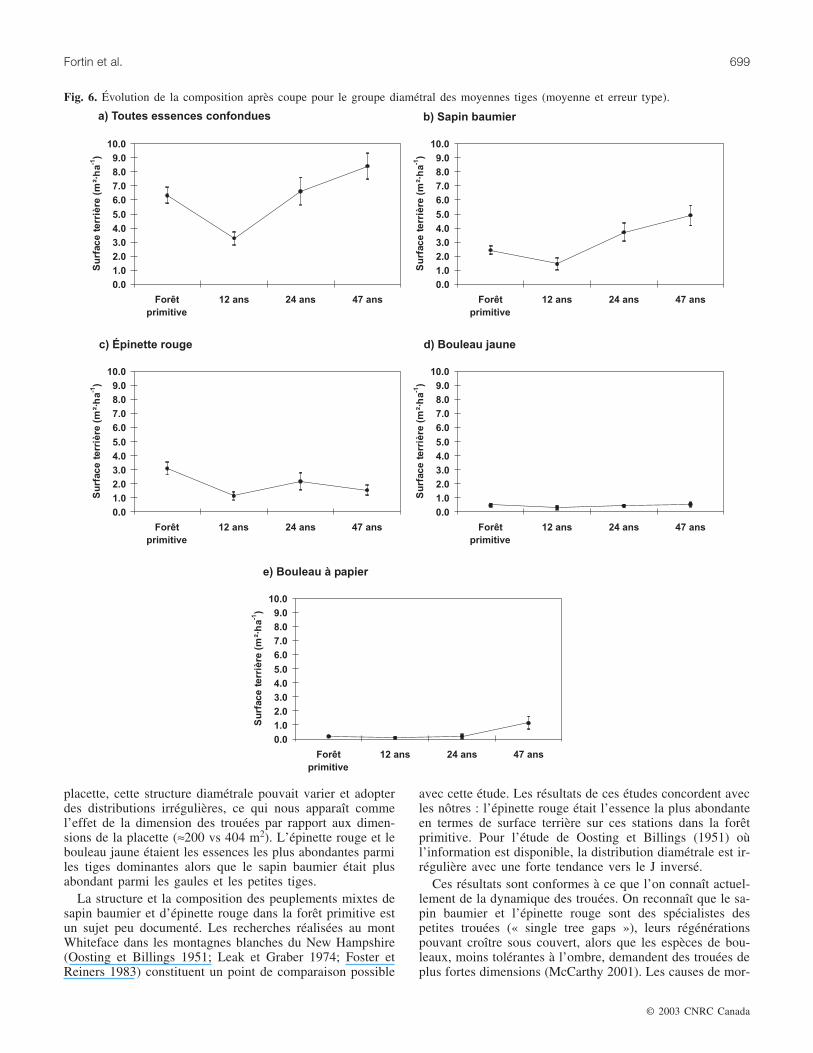
Les moyennes tigesLe prélèvement de la coupe à diamètre limite est percep-
tible pour ce groupe diamétral (fig. 6). On observe une dimi-nution importante de la surface terrière de toutes lesessences confondues 12 ans après la coupe. Par la suite, lasurface terrière a augmenté pour atteindre un niveau sem-blable à celle de la forêt primitive 24 ans après la coupe.Cette surface terrière a augmenté 47 ans après la coupe pourdépasser celle de la forêt primitive. Un patron semblablepeut être observé pour la surface terrière du sapin baumier.Quant à l’épinette rouge, sa surface terrière a également di-minué 12 ans après la coupe. Par la suite, celle-ci est restée àun niveau inférieur à celui de la forêt primitive. La surfaceterrière moyenne de l’épinette rouge est passée de 3,1 m2·ha–1
dans la forêt primitive à 1,5 m2·ha–1 47 ans après la coupe.Pour le bouleau jaune, la surface terrière est demeurée lamême tout au long de l’horizon d’observation. Celle du bou-leau à papier est restée semblable à celle de la forêt primi-
© 2003 CNRC Canada
696 Can. J. For. Res. Vol. 33, 2003
Surface terrière (m2·ha–1)
Essence et groupe diamétral Minimum Moyenne MaximumÉcarttype
GaulesToutes essences confondues 0,4 3,0 15,9 3,3Sapin baumier 0,1 1,9 14,0 2,8Épinette rouge 0,0 0,6 2,2 0,6Bouleau jaune 0,0 0,2 0,9 0,2Bouleau à papier 0,0 0,3 3,1 0,6
Petites tigesToutes essences confondues 0,2 5,1 16,8 3,6Sapin baumier 0,0 2,6 13,9 2,8Épinette rouge 0,0 1,6 6,5 1,8Bouleau jaune 0,0 0,4 2,6 0,5Bouleau à papier 0,0 0,4 2,1 0,5
Moyennes tigesToutes essences confondues 0,0 6,3 16,0 3,7Sapin baumier 0,0 2,4 6,3 2,0Épinette rouge 0,0 3,1 11,0 2,8Bouleau jaune 0,0 0,5 2,0 0,6Bouleau à papier 0,0 0,2 1,8 0,4
Grosses tigesToutes essences confondues 0,0 5,4 14,5 3,3Sapin baumier 0,0 1,3 7,3 1,9Épinette rouge 0,0 3,3 10,1 2,6Bouleau jaune 0,0 0,5 2,8 0,8Bouleau à papier 0,0 0,2 3,0 0,6
Très grosses tigesToutes essences confondues 0,0 10,1 41,3 7,3Sapin baumier 0,0 0,5 5,3 1,3Épinette rouge 0,0 5,5 20,3 5,2Bouleau jaune 0,0 2,5 12,0 3,6Bouleau à papier 0,0 0,0 0,0 0,0
Tous groupes confondusToutes essences confondues 10,2 29,8 58,1 9,2Sapin baumier 0,9 8,9 22,3 5,8Épinette rouge 1,4 14,1 35,2 9,4Bouleau jaune 0,0 4,0 14,6 4,3Bouleau à papier 0,0 1,0 5,2 1,2
Tableau 3. Composition de la forêt primitive.
Fig. 3. Comparaison entre la distribution diamétrale de la forêtprimitive et celle des peuplements 47 ans après la récolte(classes de diamètre de 1 po (2,54 cm)).
I:\cjfr\cjfr3304\X02-205.vpWednesday, March 19, 2003 4:48:24 PM
Color profile: DisabledComposite Default screen
tive pour les intervalles de 12 et 24 ans. Par la suite, elles’est mise à augmenter passant de 0,2 m2·ha–1 dans la forêtprimitive à 1,1 m2·ha–1 47 ans après la coupe.
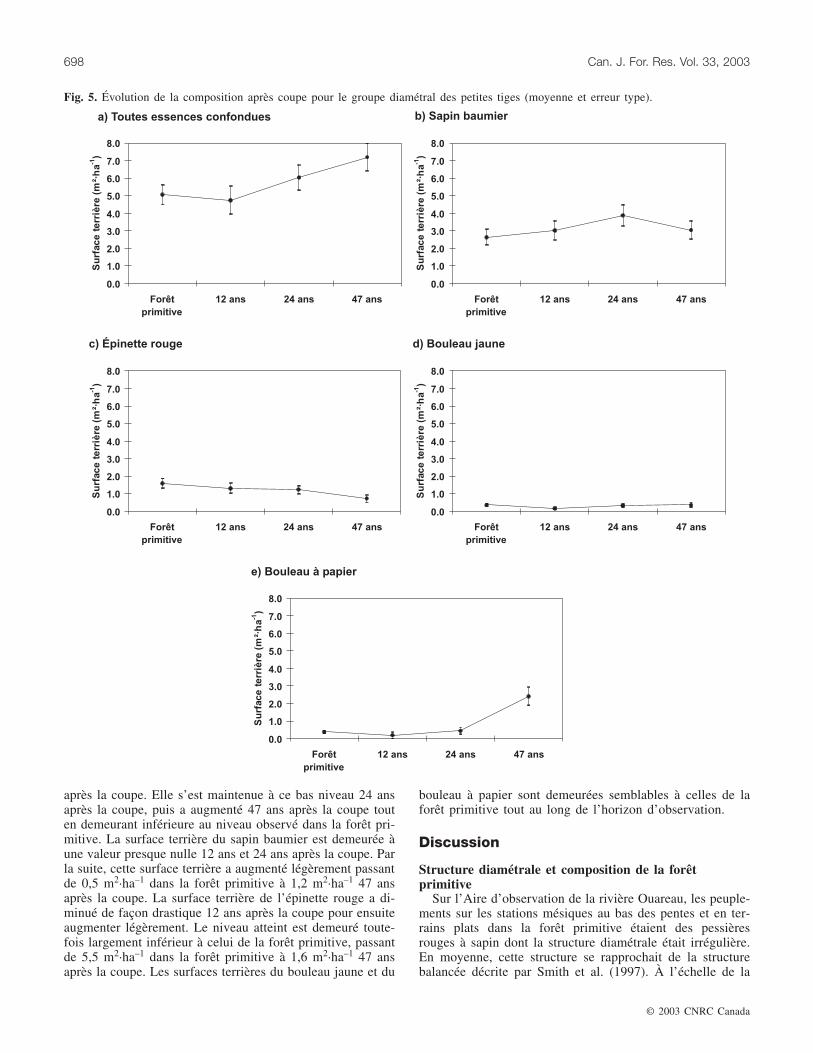
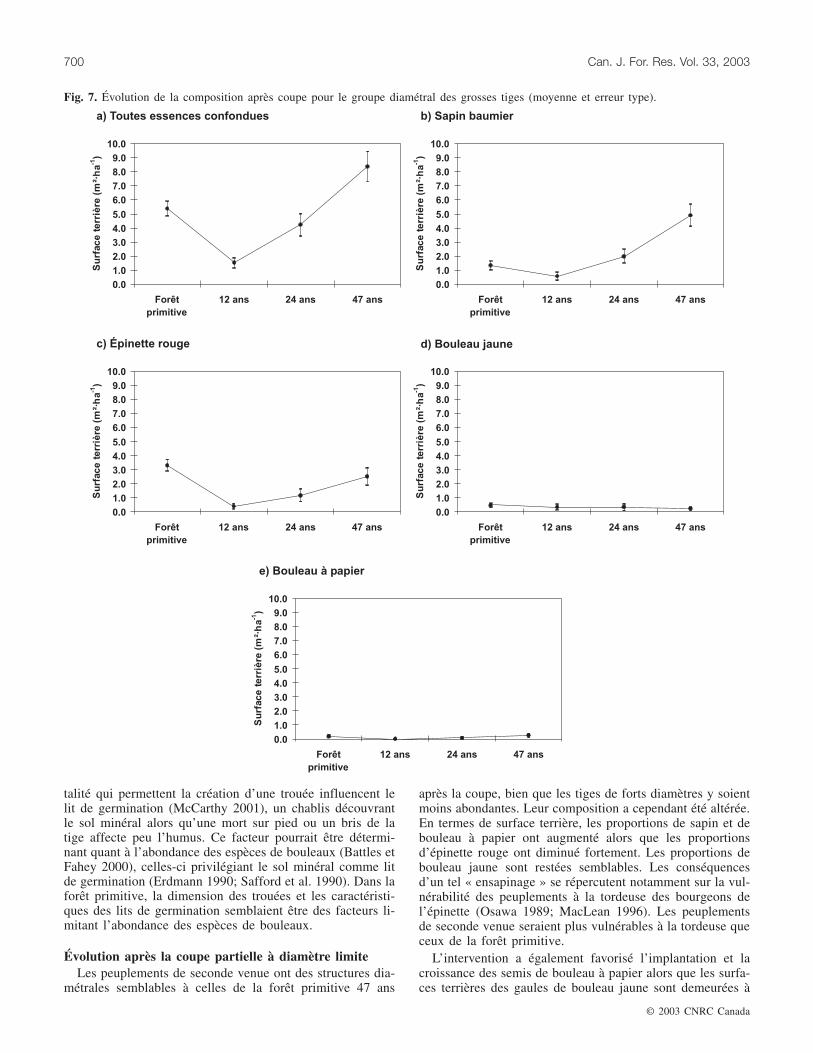
Les grosses tigesLe prélèvement de la coupe à diamètre limite est égale-
ment perceptible pour ce groupe diamétral (fig. 7). La sur-face terrière de toutes les essences confondues a diminuéfortement 12 ans après la coupe, puis a ensuite retrouvé unniveau semblable à celui de la forêt primitive 24 ans après lacoupe pour finalement augmenter et dépasser ce niveau47 ans après la coupe. La surface terrière du sapin affiche lemême patron, passant de 1,3 m2·ha–1 dans la forêt primitive
à 4,9 m2·ha–1 47 ans après la coupe. Pour l’épinette rouge, lasurface terrière a diminué fortement 12 ans après la coupe.Par la suite, elle a augmenté sans toutefois dépasser le ni-veau observé dans la forêt primitive. Les surfaces terrièresdu bouleau jaune et du bouleau à papier sont demeuréessemblables à celles de la forêt primitive tout au long del’horizon d’observation.
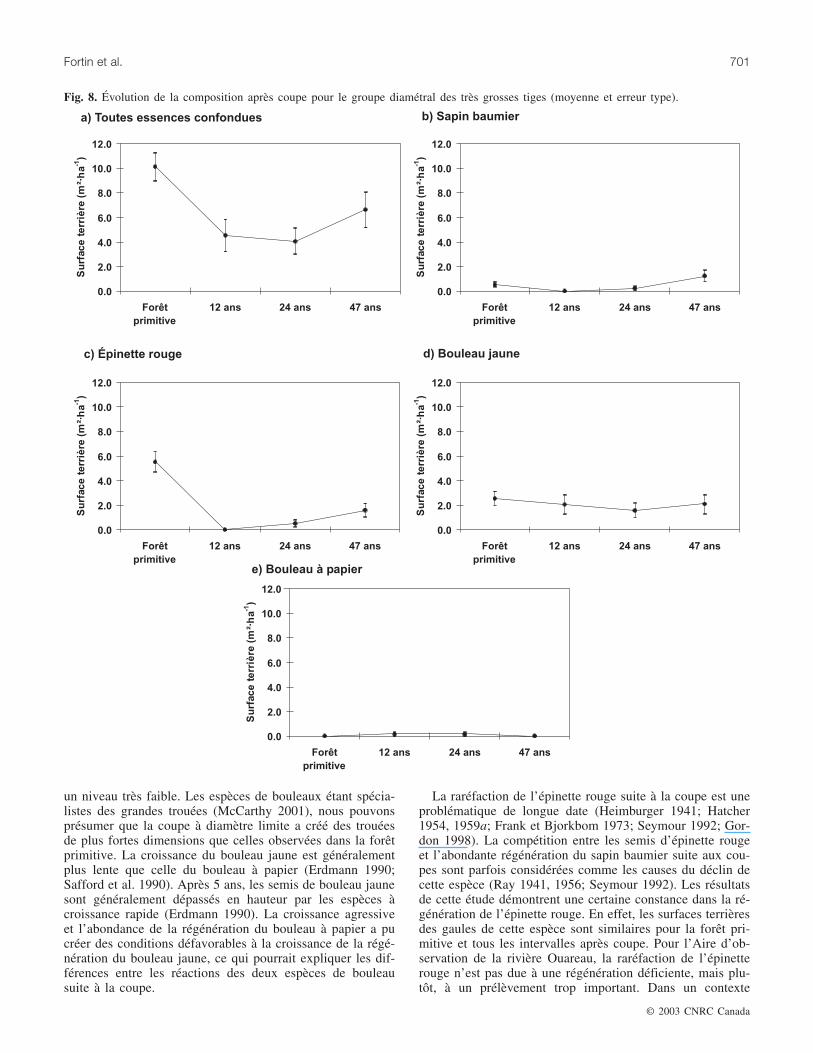
Les très grosses tigesTout comme les groupes diamétraux des moyennes tiges
et des grosses tiges, le prélèvement de la coupe est percep-tible pour ce groupe diamétral (fig. 8). La surface terrière detoutes les essences confondues a diminué fortement 12 ans
© 2003 CNRC Canada
Fortin et al. 697
Fig. 4. Évolution de la composition après coupe pour le groupe diamétral des gaules (moyenne et erreur type).
I:\cjfr\cjfr3304\X02-205.vpWednesday, March 19, 2003 4:48:25 PM
Color profile: DisabledComposite Default screen
après la coupe. Elle s’est maintenue à ce bas niveau 24 ansaprès la coupe, puis a augmenté 47 ans après la coupe touten demeurant inférieure au niveau observé dans la forêt pri-mitive. La surface terrière du sapin baumier est demeurée àune valeur presque nulle 12 ans et 24 ans après la coupe. Parla suite, cette surface terrière a augmenté légèrement passantde 0,5 m2·ha–1 dans la forêt primitive à 1,2 m2·ha–1 47 ansaprès la coupe. La surface terrière de l’épinette rouge a di-minué de façon drastique 12 ans après la coupe pour ensuiteaugmenter légèrement. Le niveau atteint est demeuré toute-fois largement inférieur à celui de la forêt primitive, passantde 5,5 m2·ha–1 dans la forêt primitive à 1,6 m2·ha–1 47 ansaprès la coupe. Les surfaces terrières du bouleau jaune et du
bouleau à papier sont demeurées semblables à celles de laforêt primitive tout au long de l’horizon d’observation.
Discussion
Structure diamétrale et composition de la forêtprimitive
Sur l’Aire d’observation de la rivière Ouareau, les peuple-ments sur les stations mésiques au bas des pentes et en ter-rains plats dans la forêt primitive étaient des pessièresrouges à sapin dont la structure diamétrale était irrégulière.En moyenne, cette structure se rapprochait de la structurebalancée décrite par Smith et al. (1997). À l’échelle de la
© 2003 CNRC Canada
698 Can. J. For. Res. Vol. 33, 2003
Fig. 5. Évolution de la composition après coupe pour le groupe diamétral des petites tiges (moyenne et erreur type).
I:\cjfr\cjfr3304\X02-205.vpWednesday, March 19, 2003 4:48:25 PM
Color profile: DisabledComposite Default screen
placette, cette structure diamétrale pouvait varier et adopterdes distributions irrégulières, ce qui nous apparaît commel’effet de la dimension des trouées par rapport aux dimen-sions de la placette (≈200 vs 404 m2). L’épinette rouge et lebouleau jaune étaient les essences les plus abondantes parmiles tiges dominantes alors que le sapin baumier était plusabondant parmi les gaules et les petites tiges.
La structure et la composition des peuplements mixtes desapin baumier et d’épinette rouge dans la forêt primitive estun sujet peu documenté. Les recherches réalisées au montWhiteface dans les montagnes blanches du New Hampshire(Oosting et Billings 1951; Leak et Graber 1974; Foster etReiners 1983) constituent un point de comparaison possible
avec cette étude. Les résultats de ces études concordent avecles nôtres : l’épinette rouge était l’essence la plus abondanteen termes de surface terrière sur ces stations dans la forêtprimitive. Pour l’étude de Oosting et Billings (1951) oùl’information est disponible, la distribution diamétrale est ir-régulière avec une forte tendance vers le J inversé.
Ces résultats sont conformes à ce que l’on connaît actuel-lement de la dynamique des trouées. On reconnaît que le sa-pin baumier et l’épinette rouge sont des spécialistes despetites trouées (« single tree gaps »), leurs régénérationspouvant croître sous couvert, alors que les espèces de bou-leaux, moins tolérantes à l’ombre, demandent des trouées deplus fortes dimensions (McCarthy 2001). Les causes de mor-
© 2003 CNRC Canada
Fortin et al. 699
Fig. 6. Évolution de la composition après coupe pour le groupe diamétral des moyennes tiges (moyenne et erreur type).
I:\cjfr\cjfr3304\X02-205.vpWednesday, March 19, 2003 4:48:26 PM
Color profile: DisabledComposite Default screen
talité qui permettent la création d’une trouée influencent lelit de germination (McCarthy 2001), un chablis découvrantle sol minéral alors qu’une mort sur pied ou un bris de latige affecte peu l’humus. Ce facteur pourrait être détermi-nant quant à l’abondance des espèces de bouleaux (Battles etFahey 2000), celles-ci privilégiant le sol minéral comme litde germination (Erdmann 1990; Safford et al. 1990). Dans laforêt primitive, la dimension des trouées et les caractéristi-ques des lits de germination semblaient être des facteurs li-mitant l’abondance des espèces de bouleaux.
Évolution après la coupe partielle à diamètre limiteLes peuplements de seconde venue ont des structures dia-
métrales semblables à celles de la forêt primitive 47 ans
après la coupe, bien que les tiges de forts diamètres y soientmoins abondantes. Leur composition a cependant été altérée.En termes de surface terrière, les proportions de sapin et debouleau à papier ont augmenté alors que les proportionsd’épinette rouge ont diminué fortement. Les proportions debouleau jaune sont restées semblables. Les conséquencesd’un tel « ensapinage » se répercutent notamment sur la vul-nérabilité des peuplements à la tordeuse des bourgeons del’épinette (Osawa 1989; MacLean 1996). Les peuplementsde seconde venue seraient plus vulnérables à la tordeuse queceux de la forêt primitive.
L’intervention a également favorisé l’implantation et lacroissance des semis de bouleau à papier alors que les surfa-ces terrières des gaules de bouleau jaune sont demeurées à
© 2003 CNRC Canada
700 Can. J. For. Res. Vol. 33, 2003
Fig. 7. Évolution de la composition après coupe pour le groupe diamétral des grosses tiges (moyenne et erreur type).
I:\cjfr\cjfr3304\X02-205.vpWednesday, March 19, 2003 4:48:26 PM
Color profile: DisabledComposite Default screen
un niveau très faible. Les espèces de bouleaux étant spécia-listes des grandes trouées (McCarthy 2001), nous pouvonsprésumer que la coupe à diamètre limite a créé des trouéesde plus fortes dimensions que celles observées dans la forêtprimitive. La croissance du bouleau jaune est généralementplus lente que celle du bouleau à papier (Erdmann 1990;Safford et al. 1990). Après 5 ans, les semis de bouleau jaunesont généralement dépassés en hauteur par les espèces àcroissance rapide (Erdmann 1990). La croissance agressiveet l’abondance de la régénération du bouleau à papier a pucréer des conditions défavorables à la croissance de la régé-nération du bouleau jaune, ce qui pourrait expliquer les dif-férences entre les réactions des deux espèces de bouleausuite à la coupe.
La raréfaction de l’épinette rouge suite à la coupe est uneproblématique de longue date (Heimburger 1941; Hatcher1954, 1959a; Frank et Bjorkbom 1973; Seymour 1992; Gor-don 1998). La compétition entre les semis d’épinette rougeet l’abondante régénération du sapin baumier suite aux cou-pes sont parfois considérées comme les causes du déclin decette espèce (Ray 1941, 1956; Seymour 1992). Les résultatsde cette étude démontrent une certaine constance dans la ré-génération de l’épinette rouge. En effet, les surfaces terrièresdes gaules de cette espèce sont similaires pour la forêt pri-mitive et tous les intervalles après coupe. Pour l’Aire d’ob-servation de la rivière Ouareau, la raréfaction de l’épinetterouge n’est pas due à une régénération déficiente, mais plu-tôt, à un prélèvement trop important. Dans un contexte
© 2003 CNRC Canada
Fortin et al. 701
Fig. 8. Évolution de la composition après coupe pour le groupe diamétral des très grosses tiges (moyenne et erreur type).
I:\cjfr\cjfr3304\X02-205.vpWednesday, March 19, 2003 4:48:27 PM
Color profile: DisabledComposite Default screen

d’aménagement écosystémique, le maintien de la proportionde cette essence au sein des peuplements tiendrait plus desvolumes laissés sur pied que d’une augmentation de l’abon-dance de sa régénération.
Les modalités de la coupe à diamètre limite réalisée surl’Aire d’observation de la rivière Ouareau ont fait en sortequ’un nombre important de tiges d’avenir d’épinette rougeont été récoltées. Ces tiges présentent un bon potentiel decroissance pour le peuplement futur (MRNQ 1997). D’aprèsles données de la forêt primitive, les surfaces terrières del’épinette rouge et du sapin baumier sont observées dans unrapport de 5 pour 3 (tableau 3). En considérant les relevés dela forêt vierge, le diamètre de récolte aurait dû être supérieurà 30 cm pour l’épinette rouge tout en restant sensiblement lemême, soit 18 cm, pour le sapin baumier si l’on avait voulumaintenir ce rapport immédiatement après la coupe. Unecoupe expérimentale menée en ce sens à la Forêt expérimen-tale du lac Édouard entre 1950 et 1956 a permis d’augmen-ter la proportion d’épinette rouge sur des stations semblables(Ménard 1999).
Dans le contexte forestier québécois, la coupe de jardi-nage par pied d’arbre (MRNQ 1997) pourrait s’avérer unesolution prometteuse. L’épinette rouge devrait être récoltéeen considérant que (i) cette essence est moins vulnérable auchablis, à la tordeuse et à la carie que le sapin baumier,(ii) qu’elle survit à de longues périodes de suppression,(iii) que sa régénération est peu abondante et (iv) qu’elleréagit aux éclaircies jusqu’à des âges avancés. Les tiges decette espèce sont peu abondantes et nous recommandons deles protéger systématiquement jusqu’à un certain diamètrelimite pour éviter de diminuer le potentiel de cette essence àreconstituer le volume prélevé. Le traitement de jardinagepar pied d’arbre pourrait être appliqué selon les normes ac-tuelles aux tiges d’épinette dont le diamètre excède la limiteainsi qu’à l’ensemble des tiges marchandes des autres espè-ces. Dans le cas présent, nous suggérons d’utiliser un dia-mètre limite de 30 cm à hauteur de poitrine et de ne récolterque les tiges d’épinette rouge moribondes sous cette limite.
Représentativité de la forêt primitiveLes placettes de la forêt primitive n’ont pu être mesurées
à nouveau après 1953. Il nous est donc impossible de distin-guer l’effet du temps de celui de la coupe dans les change-ments observés. La notion de « stabilité » des forêtsanciennes est discutable (McCarthy 2001). Malgré un tauxde remplacement (« forest turnover rate ») relativement long,le régime de perturbations des peuplements mixtes de sapinbaumier et d’épinette rouge pourrait varier dans le temps etêtre ponctué de périodes de forte mortalité (Battles et Fahey1996). Pour l’Aire d’observation de la rivière Ouareau, ledépérissement du bouleau à papier au milieu du 20e siècleest un exemple de l’irrégularité du régime de perturbationsnaturelles. La forêt primitive n’est pas un état stable et im-muable, mais nous apparaît plutôt comme une situationd’équilibre oscillant entre différentes périodes de fortes mor-talités. Le portrait de la forêt primitive que nous décrivonsdans cette étude est marqué par le dépérissement du bouleaublanc et peut-être par l’épidémie de tordeuse du début dusiècle. Nous ignorons où se situe ce portrait dans l’ « oscilla-tion » de la forêt primitive, mais il constitue la seule réfé-rence que nous ayons.
Les changements de composition observés après la coupe àdiamètre limite sont importants. Le phénomène « ensapinage –raréfaction de l’épinetterouge » ne peut être uniquement l’ef-fet du temps et de la dynamique naturelle si on considèrel’historique des perturbations tout au long de l’horizon d’ob-servation. Dans cette région, l’épinette rouge n’a pas été af-fectée par le dépérissement observé en Nouvelle-Angleterre(Siccama et al. 1982; Foster et Reiners 1983; Scott et al.1984), ni n’a connu une période évidente de forte mortalité.De plus, l’épidémie de tordeuse des années 1970 est consi-dérée comme sévère dans cette région, ce type de perturba-tion entraînant une hausse de mortalité chez le sapinbaumier (Blais 1983, 1985). L’historique des perturbationsnaturelles semble donc réunir les conditions favorables à uneaugmentation des proportions d’épinette rouge et non pasune diminution.
Conséquences pour l’aménagementLa diminution du taux d’occupation de l’épinette rouge si-
gnalée par Gordon (1998) est inquiétante et la coupe réaliséesur l’Aire d’observation de la rivière Ouareau ne constituepas la solution recherchée au maintien de l’intégrité de cetécosystème. Puisque la haute régénération de l’épinetterouge est demeurée aussi abondante qu’elle l’était dans laforêt primitive, ces peuplements retrouveront probablementla structure et la composition qu’ils avaient. Cependant, lesobjectifs de production ligneuse ne permettent pas actuelle-ment d’espacer les récoltes pour parvenir à un tel équilibre.Dans ce contexte, il importe de développer une sylviculturequi permette de maintenir la composante d’épinette rouge àun niveau acceptable.
Dans une stratégie d’aménagement écosystémique, l’utili-sation de coupes à blanc n’apparaît pas davantage commeune alternative valable. Les problèmes de régénération et devégétation compétitrice, l’ensapinage et la conversion d’unestructure d’âge inéquienne à une structure d’âge équiennesont des effets de cette intervention (Heimburger 1941; Ray1941, 1956; Westveld 1953; Hatcher 1959a; Seymour 1992,1995) qui ne cadrent pas avec les objectifs d’un aménage-ment écosystémique. Le maintien et l’augmentation des pro-portions d’épinette rouge dépendent nécessairement de laréduction du prélèvement pour cette espèce. Dans cette op-tique, des coupes partielles d’une intensité plus faible quecelle étudiée devraient être envisagées. Un jardinage parpied d’arbre pourrait représenter un traitement efficace sil’on modifiait quelques modalités d’intervention. Un dia-mètre limite devrait être fixé. Toutes les tiges d’épinetterouge (i) qui ne présentent pas de problème de vigueur et(ii) qui ont un diamètre inférieur à la limite devraient êtreprotégées systématiquement. Les tiges d’épinette rouge dontle diamètre est supérieur à cette limite ainsi que l’ensembledes tiges marchandes des autres espèces pourraient être ré-coltées selon les normes actuellement en vigueur pour cetraitement. Nous suggérons un diamètre limite de 30 cm àhauteur de poitrine.
Les effets des coupes partielles sur la structure d’âge etl’implantation des nouvelles cohortes méritent d’être appro-fondis. Seymour et Kenefic (1998) ont démontré que descoupes de jardinage ont réduit la diversité de la structured’âge des peuplements à la Forêt expérimentale de Penobs-cot. Par ailleurs, les coupes d’intensité trop légère comme la
© 2003 CNRC Canada
702 Can. J. For. Res. Vol. 33, 2003
I:\cjfr\cjfr3304\X02-205.vpWednesday, March 19, 2003 4:48:28 PM
Color profile: DisabledComposite Default screen

coupe progressive irrégulière pourraient inhiber l’apparitionde nouvelles cohortes : les peuplements de seconde venuetendraient alors vers des structures étagées (Seymour 1992).Même si la régénération de l’épinette rouge ne semble pasproblématique dans cette étude, il apparaît nécessaire de re-chercher les conditions optimales pour favoriser son installa-tion et sa croissance. L’intensité du prélèvement devrait êtreajustée en regard de ces éléments.
Remerciements
Cet article est basé sur les résultats d’un mémoire de maî-trise complété par le premier auteur à la Faculté de foresterieet de géomatique de l’Université Laval (Québec). Les au-teurs tiennent à remercier le ministère des Ressources natu-relles du Québec pour le financement de ce projet, le Servicecanadien des forêts pour le prêt des données de l’Aire d’ob-servation de la rivière Ouareau, le Parc du Mont-Tremblantet le personnel de terrain. Nous tenons également à remer-cier le Prof. Jean-Claude Ruel et le Prof. Hank Margolis, dela Faculté de foresterie et de géomatique, ainsi que deuxévaluateurs anonymes pour leurs précieux commentaires.
Bibliographie
Battles, J.J., et Fahey, T.J. 1996. Spruce decline as a disturbanceevent in the subalpine forests of the northeastern United States.Can. J. For. Res. 26 : 408–421.
Battles, J.J., et Fahey, T.J. 2000. Gap dynamics following forestdecline: a case study of red spruce forests. Ecol. Appl. 10 : 760–774.
Bélanger, L. 1985. Intégration de l’information écologique dans lagestion des terres de l’écoumène forestier par l’inventaire écolo-gique des terres. Thèse de doctorat, Université Laval, Sainte-Foy, Qc.
Blais, J.R. 1983. Trends in frequency, extent, and severity ofspruce budworm outbreaks in eastern Canada. Can. J. For. Res.13 : 539–547.
Blais, J.R. 1985. The ecology of the Eastern spruce budworm: a re-view and discussion. Dans Recent Advances in SpruceBudworms Research. Proceedings of the CANUSA SpruceBudworms Research Symposium, 16–20 sept. 1985, Bangor,Maine. Sous la direction de C.J. Sanders, R.W. Stark, E.J. Mul-lins et J. Murphy. Service canadien des forêts et United StatesDepartment of Agriculture Forest Service. p. 49–59.
Brissette, J.C. 1996. Effects of intensity and frequency of harvest-ing on abundance, stocking and composition of natural regenera-tion in the Acadian forest of Eastern North America. Silva Fenn.30 : 301–314.
Erdmann, G.G. 1990. Yellow birch. Dans Silvics of North Amer-ica. Vol. 1. Sous la direction de R.M. Burns et B.H. Honkala.U.S. Dep. Agric. Agric. Handb. 654. p. 133–147.
Fahey, T.J., et Reiners, W.A. 1981. Fire in the forests of Maine andNew Hampshire. Bull. Torrey Bot. Club, 108 : 362–373.
Foster, J.R., et Reiners, W.A. 1983. Vegetation patterns in a virginsubalpine forest at Crawford Notch, White Mountains, NewHampshire. Bull. Torrey Bot. Club, 110 : 141–153.
Foster, J.R., et Reiners, W.A. 1986. Size distribution and expansionof canopy gaps in a northern Appalachian spruce–fir forest. Veg-etation, 68 : 109–114.
Frank, R.M. 1990. Balsam fir. Dans Silvics of North America. Vol.1. Sous la direction de R.M. Burns et B.H. Honkala. U.S. Dep.Agic. Agric. Handb. 654. p. 26–35.
Frank, R.M., et Bjorkbom, J.C. 1973. A silvicultural guide forspruce–fir in the Northeast. USDA For. Serv. Gen. Tech. Rep.NE-6.
Frank, R.M., et Blum, B.M. 1978. The selection system of silvi-culture in spruce–fir stands — procedures, early results, andcomparisons with unmanaged stands. USDA For. Serv. Res. Pap.NE-425.
Gagnon, G., et Marcotte, G. 1980. Description des types écologi-ques et de leur productivité dans la section forestière lauren-tienne de Rowe (L-4a). Ministère des Ressources naturelles duQuébec, Québec. Mémoire de recherche forestière 61.
Gordon, A.G. 1998. The sweep of the boreal in time and space,from forest formations to genes, and implications for manage-ment. For. Chron. 72 : 19–30.
Gosselin, J., Grondin, P., et Saucier, J.-P. 1998. Rapport de classifi-cation écologique du sous-domaine bioclimatique de l’érablièreà bouleau jaune de l’est. Ministère des Ressources naturelles duQuébec, Direction de la gestion des stocks forestiers, Québec.
Green, P.T. 1996. Canopy gaps in rain forest on Christmas Island,Indian Ocean: size distribution and methods of measurement. J.Trop. Ecol. 12 : 427–434.
Grondin, P., Ansseau, C., Bélanger, L., Bergeron, J.-F., Bergeron, Y.,Bouchard, A., Brisson, J., De Grandpré, L., Gagnon, G., Lavoie,C., Lessard, G., Payette, S., Richard, P.J.H., Saucier, J.-P.,Sirois, L., et Vasseur, L. 1996. Écologie forestière. Dans Manuelde foresterie. Les Presses de l’Université Laval, Sainte-Foy, Qc.p. 133–279.
Hatcher, R.J. 1954. A report on the establishment of observationarea no. 12 on the limits of the Consolidated Paper CorporationOuareau River, P.Q. 1953. Can. For. Serv. Proj. Q-54.
Hatcher, R.J. 1959a. Partial cutting with diameter limit control inthe Lake Edward Experimental Forest, Quebec, 1950 to 1956.Can. For. Serv. Proj. Q-44.
Hatcher, R.J. 1959b. Mortality and regeneration following partialcutting of spruce – balsam fir – hardwood stands at lake Ed-ward, P.Q. Can. For. Serv. Proj. Q-44.
Hatcher, R.J. 1960. Management of a red spruce, balsam fir andyellow birch stand — 1935 to 1959. Pulp Pap. Mag. Can. 61 :168–171.
Hébert, R. 2000. Importance du processus naturel de régénérationde la forêt mature mixte pour l’alimentation du lièvre d’amé-rique, de l’orignal et de l’ours noir. Mémoire de maîtrise, Uni-versité Laval, Sainte-Foy, Qc.
Heimburger, C.C. 1941. Forest site classification and soil investiga-tion in the Lake Edward Forest Experimental Area. Can. For.Serv. Silv. Res. Note 66.
Lalumière, R., et Thibault, M. 1988. Les forêts du Parc national dela Mauricie au Québec. Étude écologique no 11. Les Presses del’Université Laval, Sainte-Foy, Qc.
Leak, W.B. 1975. Age distribution in virgin red spruce and north-ern hardwoods. Ecology, 56 : 1451–1454.
Leak, W.B., et Graber, R.E. 1974. Forest vegetation related to ele-vation in the White Mountains of New Hampshire. USDA For.Serv. Res. Pap. NE-299.
Lorimer, C.G. 1977. The presettlement forest and natural distur-bance cycle of northeastern Maine. Ecology, 58 : 139–148.
MacLean, D.A. 1996. The role of a stand dynamics model in thespruce budworm decision support system. Can. J. For. Res. 26 :1731–1741.
MacLean, D.A., et MacKinnon, W.E. 1997. Effects of stand andsite characteristics on susceptibility and vulnerability of balsamfir and spruce to spruce budworm in New Brunswick. Can. J.For. Res. 27 : 1859–1871.
© 2003 CNRC Canada
Fortin et al. 703
I:\cjfr\cjfr3304\X02-205.vpWednesday, March 19, 2003 4:48:29 PM
Color profile: DisabledComposite Default screen

© 2003 CNRC Canada
704 Can. J. For. Res. Vol. 33, 2003
McCarthy, J. 2001. Gap dynamics of forest trees: a review withparticular attention to boreal forests. Environ. Rev. 9 : 1–59.
McLintock, T.F. 1954. Factors affecting wind damage in selectivelycut stands of spruce and fir in Maine and northern New Hamp-shire. USDA For. Serv. Northeast. For. Exp. Stn. Stn. Pap. 70.
Ménard, B. 1999. Dynamique naturelle des forêts mixtes de la sta-tion expérimentale du lac Édouard, au Parc national de la Mau-ricie. Mémoire de maîtrise, Université Laval, Sainte-Foy, Qc.
Meunier, S., Ruel, J.-C., Laflamme, G., et Achim, A. 2002. Résis-tance comparée de l’épinette blanche et du sapin baumier aurenversement. Can. J. For. Res. 32 : 642–652.
Ministère des Ressources naturelles du Québec (MRNQ). 1997. Ma-nuel d’aménagement forestier. 3e éd. Ministère des Ressourcesnaturelles du Québec, Direction des relations publiques, Québec.
Oliver, C.D., et Larson, B.C. 1996. Forest stand dynamics. Up-dated edition. John Wiley and Sons, New York.
Oosting, H.J., et Billings, W.D. 1951. A comparison of virginspruce–fir forest in the northern and southern Appalachian sys-tem. Ecology, 32 : 84–103.
Osawa, A. 1989. Causality in mortality patterns of spruce trees dur-ing a spruce budworm outbreak. Can. J. For. Res. 19 : 632–638.
Ray, R.G. 1941. Site-types and rate of growth. Can. For. Serv.Silvic. Res. Note 65.
Ray, R.G. 1956. Site-types, growth and yield at Lake Edward Ex-perimental Area Quebec. Can. For. Serv. Tech. Note 27.
Robitaille, A., et Saucier, J.-P. 1998. Paysages régionaux du Qué-bec méridional. Ministère des Ressources naturelles du Québec,Québec.
Rowe, J.S. 1972. Les régions forestières du Canada. Ministère del’environnement du Canada, Service canadien des forêts,Ottawa, Ont. Publ. 1300F.
Ruel, J.-C. 2000. Factors influencing windthrow in balsam fir for-ests: from landscape studies to individual tree studies. For. Ecol.Manage. 135 : 169–178.
Safford, L.O., Bjorkbom, J.C., et Zasada, J.C. 1990. Paper birch.Dans Silvics of North America. Vol. 1. Sous la direction deR.M. Burns et B.H. Honkala. U.S. Dep. Agric. Agric. Handb.654. p. 158–171.
Scott, J.T., Siccama, T.G., Johnson, A.H., et Breisch, A.R. 1984.Decline of red spruce in the Adirondacks, New York. Bull.Torrey Bot. Club, 111 : 438–444.
Seegrist, D.W., et Arner, S.L. 1982. Mortality of spruce and fir inMaine in 1976–78 due to spruce budworm outbreak. USDA For.Serv. Res. Pap. NE-491.
Seymour, R.S. 1992. The red spruce – balsam fir forest of Maine:evolution of silvicultural practice in response to stand developmentpatterns and disturbances. Dans The ecology and silviculture ofmixed-species forests. Sous la direction de M.J. Kelty, B.C. Larsonet C.D. Olivier. Kluwer Publishers, Norwell, Mass. p. 217–244.
Seymour, R.S. 1995. The northeastern region. Dans Regional silvi-culture of the United States. 3e éd. Sous la direction deJ.W. Barrett. John Wiley and Sons, New York. p. 31–79.
Seymour, R.S., et Kenefic, L.S. 1998. Balance and sustainability inmultiaged stands — a northern conifer case study. J. For. 96 :12–17.
Siccama, T.G., Bliss, M., et Vogelmann, H.W. 1982. Decline of redspruce in the Green Mountains of Vermont. Bull. Torrey Bot.Club, 109 : 162–168.
Smith, D.M., Larson, B.C., Kelty, M.J., et Ashton, P.M.S. 1997.The practice of silviculture: applied forestry ecology. 9e éd. JohnWiley and Sons, New York.
Westveld, M. 1953. Ecology and silviculture of the spruce–fir for-ests of eastern North America. J. For. 51 : 422–430.
Worrall, J.J., et Harrington, T.C. 1988. Etiology of canopy gaps inspruce–fir forests at Crawford Notch, New Hampshire. Can. J.For. Res. 18 : 1463–1469.
I:\cjfr\cjfr3304\X02-205.vpWednesday, March 19, 2003 4:48:29 PM
Color profile: DisabledComposite Default screen