endogenous sleep-promoting substances and sleep regulation and permissive components: two-process...
TRANSCRIPT

PHYSIOLOGICAL REVIEWS Vol. 69, No. 2, April 1989
Printed in I%S.A.
Endogenous Sleep-Promoting Substances and Sleep Regulation
ALEXANDER A. BORBELY AND IRENE TOBLER
Institute of Pharmacology, University of Ziirich, Ziirich, Switzerland
I. Introduction .................................................. ....... 605 II. Processes Underlying Physiological Sleep Regulation ................... 606
A. Permissive components ............................................ 606 B. Circadian components ............................................. 607 C. Homeostaticcomponents .......................................... 608 D. Interaction of homeostatic, circadian, and permissive components:
two-process model of sleep regulation ........................... 612 III. Search and Identification of Sleep Factors .............................. 613
A. Approaches based on total sleep deprivation ........................ 613 B. Approaches based on rapid-eye-movement sleep deprivation .......... 620 C. Approaches based on electrical brain stimulation .................... 621 D. Approaches based on spontaneous sleep ............................. 629 E. Neuromodulators and neurotransmitters ............................ 631 F. Immunologically active polypeptides ................................ 642
IV. Discussion and Conclusions ........................................... 645 A. Criteria for an endogenous sleep factor or a sleep substance .......... 645 B. Sleep-promotingaction ............................................ 645 C. Changes associated with sleep and waking .......................... 651 D. Conclusions ....................................................... 652
I. INTRODUCTION
The field of endogenous sleep-promoting substances has become increas- ingly popular in recent years, and various reviews (2, 29, 39, 73, 77, 82, 85, 128a, 145,146,151,155,157,165,166,178,186,189,201,210,234,240,277,393, 394), two symposium volumes (148,156), and a monograph (147a) have been published on this topic. However, because many of the reviews have focused on a specific class of substances, they did not offer an extensive overview of endogenous sleep-promoting substances. In the present review we attempt to provide this information. Moreover, by presenting the various approaches within the framework of physiological sleep regulation, we made an effort to transcend the purely descriptive level. Ultimately, endogenous sleep-pro- moting substances must be understood as chemical mediators of physiologi- cal processes. It is therefore important to analyze their effects from this vantage point. To provide the necessary background, the relevant recent developments in the physiology of sleep regulation are summarized in the next section. Even though a considerable body of data cannot yet be fitted into physiological categories, we are convinced that physiology must remain
0031-9333/89 $1.50 Copyright 0 1989 the American Physiological Society 605

606 ALEXANDER A. BORBELY AND IRENE TOBLER Volume 69
a major guideline for future research. [Substances inducing hibernation are not covered; see Krueger and Shoham (206) for a recent review.]
II. PROCESSES UNDERLYING PHYSIOLOGICAL SLEEP REGULATION
Permissive, circadian, and homeostatic components play important roles in sleep regulation. This section provides an overview of recent developments in human and animal sleep research that are relevant to the topic of this review. The majority of the animal studies that are discussed were performed in the rat. This species has been increasingly used both in physiological investigations and in the search for endogenous sleep-promoting substances. Although the rat has a polyphasic sleep pattern, its distinct circadian sleep/ waking rhythm and the distribution of its sleep states resemble in various respects human sleep (28).
A. Permissive Cowaponents
Sleep may occur at virtually any time provided that the environmental conditions are propitious. In humans the capacity to fall asleep during the habitual waking hours is made use of in the multiple sleep latency test (MSLT) for the assessment of daytime sleep propensity (45). The subjects are asked to lie down in a darkened, sound-shielded room, to keep their eyes closed, and to try to fall asleep. Usually sleep sets in within the 20 min that are available in the test situation. The recumbent position as well as the shielding from visual and auditory stimuli are obviously factors that lower arousal and favor sleep. The sleep-conducive effect of stimulus reduction has been impressively demonstrated experimentally in the “enforced bed rest” paradigm (43). When subjects were confined to bed for 3 days and were not allowed to engage in any sort of activity, they not only showed multiple sleep episodes per 24 h but also showed a marked rise in total sleep time.
In animals, sleep is favored when the environmental conditions are not activating (for review see Ref. 370). In particular, light and ambient tempera- ture may exert a strong influence on sleep. Thus, in the rat, exposure to short light periods (5-60 min) (24) or a rise in ambient temperature (270,362,396) markedly enhanced sleep. Also, availability of food after a period of food deprivation (25) increased sleep. Electroencephalogram (EEG) synchroniza- tion has been reported after heating of the preoptic region (18), after electri- cal stimulation of vagal afferents (298), and postprandially (224). The former manipulations presumably mimic a rise in ambient temperature and satiety, respectively.
Exogenous factors not only affect sleep in general but may induce changes in a specific substate of sleep. For example, turning the light off tends to trigger a rapid-eye-movement (REM) sleep episode in the sleeping rat (24,219,307,372), whereas turning the light on has the same effect in the

April 1989 ENDOGENOUS SLEEP-PROMOTING SUBSTANCES 607
hamster (371). Also REM sleep can be triggered by brief peripheral thermal stimuli in the rat (363) or by auditory and somatosensory stimuli in the cat (74, 79).
B. Circadian Components
In humans, sleep latency, as measured by the MSLT, declined precipi- tously in the early part of the night (311). When sleep onset was delayed from the normal bedtime to the morning hours, sleep duration decreased (8). These results indicate that sleep onset and sleep maintenance are favored during the portion of the 24-h cycle corresponding to the habitual sleep period. The minima of the 24-h rhythms of body temperature and plasma cortisol levels typically coincide with the habitual sleep period (256,410). This relationship is present also when subjects live without time cues, and hence their circa- dian rhythms are no longer synchronized to the environmental 24-h cycle. Thus the onset of major sleep episodes coincided with the descending limb of the circadian temperature rhythm (429), and the highest amount of sleep occurred around the trough of the temperature cycle (59,409,428). Because body temperature and cortisol secretion are reliable indicators of the human circadian pacemaker (5, 256), the specific phase relation of sleep to these parameters indicates the involvement of circadian factors in sleep regula- tion. Sleeping at the “wrong circadian time” is considered as a main cause of sleep disturbances after time zone shifts or during shift work (6).
Among the substates of human sleep it is REM sleep that is most in- fluenced by circadian factors. The percentage of REM sleep typically in- creases in the course of a nocturnal sleep period and is particularly high when sleep is extended into the morning (15, 95). This rising trend of REM sleep is reversed to a declining trend if sleep onset occurs between 0700 in the morning and 1500 (8). Naps early during the day have a higher REM sleep content than naps late during the day (174). When sleep is phase shifted, REM sleep readjusts only after several days (411). Taken together, these observations constitute strong evidence for the powerful effect of circadian factors on REM sleep propensity.
In contrast to REM sleep, slow-wave sleep (stages 3 and 4) is little influenced by circadian factors (408). Regardless of the time of sleep onset, slow-wave sleep shows a declining trend throughout the sleep period (8, 9). When sleep is phase shifted, slow-wave sleep readjusts immediately (411). Although it has been claimed that the resurgence of slow-wave sleep in the course of extended bed rest periods reflects a semicircadian rhythm (107), the data are open to alternative interpretations (140).
A distinct circadian sleep/wake rhythm is present also in other mam- mals, such as the rat, that are awake during ~80% of the dark period and asleep during 80% of the light period (36, 399). The circadian sleep/wake rhythm persists in the absence of light (26,34,35,244). Other animals, such

608 ALEXANDER A. BORBELY AND IRENE TOBLER Vohww 69
as the cat (213,220) and rabbit (290), show much smaller circadian variations of sleep.
When circadian factors of sleep regulation are assessed, it is important to keep in mind that the circadian pacemaker (presumably located in the suprachiasmatic nucleus) imposes a rhythm on a variety of different pro- cesses. These include behavior (e.g., motor activity, feeding, drinking, sensory discrimination), thermoregulation, metabolism, hormone secretion (e.g., cor- ticosterone, vasopressin), and several neurotransmitter receptors (320,419). The circadian modulation of sleep propensity should be therefore regarded not in isolation but as part of the rhythmic changes in the state of the whole organism. Such a global view comes close to the old notion of a trophotropic and ergotropic state alternating during the course of 24 h (132).
C. Homeostatic Components
Sleep reduction gives rise to increased sleep propensity, whereas excess sleep has the opposite effect. Thus regulatory mechanisms exist that counter- act deviations from a physiological “reference level” of sleep. This aspect of sleep regulation is referred to as sleep homeostasis.
1. Sleep latency and sleep duration
In humans, sleep reduction at night gives rise to increased sleep propen- sity during daytime. Sleep latency during the day is a sensitive measure of the amount of prior sleep (44). In a recent experiment, mean daytime sleep latency was 13.6 min after a sleep duration of 7 h, 7.5 min after 4 h, and 1.6 min when no sleep had occurred in the previous night (31).
Sleep loss is compensated only to a small extent by a prolongation of sleep during the next sleep period. Thus when sleep was postponed by 24 h with respect to habitual bedtime, sleep duration increased from 7.9 to 8.7 h (8). Even after a vigil of 264 h the duration of recovery sleep in the first night was 14.4 h (126). In sleep-deprivation experiments in the rat, sleep duration was not related to prior waking (217, 375).
2. Electroencephalogram slow-wave activity: a measure of sleep intensity
The alternation of non-REM (NREM) and REM sleep is a typical feature of mammalian sleep. Non-REM sleep is not a homogenous substate of sleep, but it can be subdivided according to the predominance of EEG slow-wave activity. The percentage of slow waves (frequency O-2 Hz; minimum ampli- tude 75 pV) is the major criterion for scoring human NREM sleep into stages 2,3, or 4 (308). Stages 3 and 4 are commonly referred to as slow-wave sleep. However, the conventional sleep scoring method is inadequate for a quanti- tative analysis, because the sleep stages are based on gross and arbitrary

April 1989 ENDOGENOUS SLEEP-PROMOTING SUBSTANCES 609
criteria. Presently, EEG parameters can be assessed by computer-aided methods of signal analysis. The most important functional EEG parameter will be referred to as “slow-wave activity.” It is equivalent to a-activity and encompasses components of the EEG signal in the frequency range of -1-4 Hz. A quantitative measure of slow-wave activity has been obtained by spec- tral analysis (e.g., see Ref. 32), period amplitude analysis (e.g., see Refs. 52, 243), filtering and signal rectification (e.g., see Ref. 98), or similar methods. Although each method has its inherent limitations (see Ref. ZIZ), they all allow the quantitative determination of EEG components in the low-fre- quency range, which will be referred to as slow-wave activity. The amount of slow-wave sleep is a rough indication of the prevalence of slow-wave activity in NREM sleep.
It has been shown repeatedly that partial or total sleep deprivation gives rise to increased slow-wave sleep (19; see Ref. 27 for references). Webb and Agnew (408) presented compelling evidence that slow-wave sleep increases as a function of prior waking. It has been reported that the responsiveness to stimuli decreases as EEG slow waves become more predominant (e.g., see Refs. 20, 418). Under physiological conditions, this EEG parameter can be therefore regarded as an indicator of sleep depth or sleep intensity.
All-night spectral analysis of the sleep EEG revealed an exponential decline of average slow-wave activity over the first three NREM/REM sleep cycles of human sleep (32). After sleep deprivation, the initial level was increased but the exponential decline was preserved. The extension of sleep time in the previous night or napping during the day reduced slow-wave sleep or slow-wave activity in the following night (95, 96, 175). The rise of slow- wave propensity during the analysis of daytime naps th 2000 h revealed a mono tonic rise of slow -wave activity as a function of prior waki ng. Extrapol .ation of th e values to nap-onset time on the basis of the two-process model (see sect. IID) showed that the rise can be approximated by a saturating exponential function.
day has been recently documented (17, 71). The at took place at 2-h intervals between 1000 and
In contrast to human studies, in animals only NREM and REM sleep (i.e., paradoxical sleep) have usually been discriminated. Some authors further subdivided NREM sleep into light and deep substates on the basis of the prevalence of slow waves in the EEG. It is confusing that in animal studies the term “slow-wave sleep” often designates NREM sleep, whereas in humans it refers to the substate of NREM sleep with predominant EEG slow waves (i.e., stages 3 and 4). Here we use the terms NREM sleep, deep and light NREM sleep, and slow-wave activity to designate a quantitative measure of EEG slow waves within NREM sleep.
The application of quantitative EEG analyses revealed that in the rat EEG slow-wave activity as well as EEG amplitude progressively decline in the course of the light period, the animal’s circadian sleep period, and in- crease during the dark period (30,36,40,318,385). Similar results have been obtained in the hamster (378). An increase of slow-wave activity after sleep

610 ALEXANDER A. BORBELY AND IRENE TOBLER Volume 69
deprivation was first demonstrated in the rabbit (278) and then was exten- sively documented in the rat (36,105). Varying the duration of sleep depriva- tion showed that the increase in slow-wave activity is a function of the duration of prior waking (375). Sleep deprivation in other mammalian spe- cies (i.e., primates, dogs, cats, and dolphins) has elicited similar increases of slow-wave activity (for references see Ref. 369). In summary, in humans and other mammals, slow-wave activity may represent an intensity parameter of NREM sleep that is regulated as a function of prior waking.
3. Rapid-eye-movement sleep
In humans, 24-h sleep deprivation does not induce an increase in REM sleep during recovery (e.g., see Ref. 32). A more severe sleep deficit gives rise to a REM sleep rebound that appears sometimes only in the second night (for references see Ref. 27). Excess REM sleep during daytime naps does not lead to a reduction during the night. Furthermore, no EEG index of an intensity dimension has been described for REM sleep. Selective REM sleep depriva- tion augments REM sleep pressure, which is manifested by the increasing number of interventions required to prevent REM sleep episodes (for refer- ences see Ref. 27). However, the occurrence of a REM sleep rebound during recovery sleep is inconsistent (46). Thus, although human REM sleep re- sponds to a sleep deficit, it is not as finely regulated as slow-wave sleep.
In the rat, a 24-h total sleep deprivation is followed by an immediate REM sleep rebound (36,105). However, also in this species, slow-wave activ- ity responds more sensitively to sleep loss than REM sleep. Thus a nocturnal sleep deprivation of 6 or 12 h induced an increase in slow-wave activity but not in REM sleep (375). The increase in e-activity during REM sleep after 24-h sleep deprivation could reflect an enhancement of REM sleep intensity (40). In contrast, recovery from prolonged sleep deprivation elicited a large and immediate REM sleep rebound and no increase of deep NREM sleep (91). In the cat, total sleep deprivation for 72 h was also followed by an immediate REM sleep rebound (185).
Selective REM sleep deprivation is a procedure that has been often used in animal experiments. Whereas various methods have been applied [e.g., hypothermia (Bl)], the “flower pot” or “island” technique has been the most popular method. This procedure was originally devised for the cat (161) and has been adapted for the rat. The animal is placed on a small platform (an inverted flower pot) surrounded by water. Due to the muscle relaxation associated with REM sleep the animal falls off the platform whenever this sleep state is initiated. Electroencephalogram recordings have confirmed that little or no REM sleep occurs in this situation (e.g., see Refs. 3,125,235, 398, 402, 403). Another technique consists of keeping the rat’s cage in a pendulum-like motion so that postural imbalance prevents REM sleep (397). The effect of REM sleep deprivation by several methods has been carefully

April 1989 ENDOGENOUS SLEEP-PROMOTING SUBSTANCES 611
documented in the rat (398). After a 72-h deprivation, REM sleep was imme- diately augmented and returned to the control level within 2 days. When the duration of REM sleep deprivation was varied between 1 and 4 days, REM sleep rebound in the first 6 h of recovery sleep was a function of the duration of deprivation (3). A REM sleep rebound has been reported in many mam- malian species, including the monkey (67), cat (161), mouse (184), cow (319), and rabbit (291).
4 Sleep deprivation: problem of specificity
The investigation of homeostatic aspects in sleep regulation is heavily based on sleep-deprivation experiments. It is therefore important to assess the specificity of such manipulations. In human studies, sleep deprivation entails not only the lack of sleep but also an upright body posture, increased sensory stimulation, motor and cognitive activity, social interaction, as well as eating and drinking at night. In animal experiments, sleep deprivation is frequently associated with forced locomotion and possibly stress. The ques- tion arises therefore whether the changes in sleep observed after various deprivation procedures are a consequence of sleep loss or of some other parameter.
There is strong evidence that the level of EEG slow-wave activity is determined by the duration of prior waking and not by some unspecific factor. As shown in a human nap study (17,71), slow-wave propensity builds up gradually in the course of the habitual waking period. If sleep is pre- vented, this buildup proceeds further. These results show that the depriva- tion procedure does notscause a qualitatively different kind of response but only augments a physiological trend. The same argument applies to rat ex- periments where the physiological rise of slow-wave propensity during the dark period is augmented by enforced waking (40,375,384).
It is still unclear whether the increase of slow-wave activity in human and animal experiments is due to specific aspects of waking. Human studies yielded contradictory results concerning the relationship between physical exercise and slow-wave sleep (141, 344, 382). An i ncrease in slow-wave has been reported as a consequence of rai .sing body te mperature se
sleep veral
hours before sleep (142). Whether the alleged slow-wave-promoting action of physical exercise is due to hyperthermia remains to be established.
The influence of forced locomotion on sleep in the rat was investigated by varying the rotation rate of the deprivation apparatus (36, 105) and by allowing the animal to engage in voluntary running activity (127). No major effect on deep sleep was found. In the hamster, sleep deprivation by forced locomotion and by gentle handling affected sleep in very similar ways (378). Finally, the influence of a major stress effect seems to be unlikely in view of the finding that plasma corticosterone levels during forced locomotion did not differ significantly from those obtained in control animals (379). All

612 ALEXANDER A. BORBELY AND IRENE TOBLER hhme 69
these observations indicate that it is the duration of waking that is most directly related to the increase in slow-wave activity.
In REM sleep-deprivation studies, REM sleep is frequently deprived for several days. It is not unreasonable to assume that the risk of nonspecific effects is higher in such prolonged interventions. In addition, an effect of the deprivation procedure on other sleep stages must be seriously considered. In several REM sleep-deprivation studies NREM sleep was also decreased and during the following recovery period, deep NREM sleep was increased (125, 235, 323, 398). However, in most REM sleep-deprivation studies only the duration of vigilance states was determined, and a possible intensification of NREM sleep was not assessed.
Even though REM sleep is effectively deprived by the flower pot method, changes induced by this procedure are not necessarily a consequence of se- lective REM sleep deprivation. The influence of stress by restriction of motor activity and the repeated exposure to water cannot be excluded (e.g., see Refs. 55, 193, 398).
D. Interaction of Homeostatic, Circadian, and Permissive Components: Two-Process Model of Sleep Regulation
Two lines of evidence indicate that the homeostatic and circadian facets of sleep regulation are mediated by separate processes. First, a conflict can be created between the two tendencies in the rat by ending a 24-h sleep-de- privation period at the onset or in the middle of the dark period, the circadian period of predominant waking (36,244,384). In this experimental paradigm, slow-wave sleep showed a rebound in two stages: an immediate increase followed by waking and a second, delayed increase at light onset.
A second line of evidence stems from experiments in which the circadian facet of sleep regulation was disrupted by lesions of the suprachiasmatic nuclei (376). Arhythmic rats were subjected to 24-h sleep deprivation to test their ability to compensate for sleep loss. Both slow-wave activity in NREM sleep and the amount of REM sleep were increased. Similar results were obtained by another group (244). In addition, sleep deprivation affected nei- ther the phase nor the period of the free-running rest-activity rhythm *of the rat (17, 28). These results indicate that the homeostatic component of sleep regulation is morphologically and functionally distinct from the circadian component.
The interaction of the major processes underlying the regulation of human sleep has been conceptualized in the two-process model (27, 60). The model is based on a sleep/wake-dependent homeostatic process (process S) and a circadian, sleep/wake-independent process (process C). Process S shows a saturating exponential rise during the waking period and an expo- nential decline during the sleep period. The parameters determining the time course of process S have been estimated from the changes of slow-wave

April 1989 ENDOGENOUS SLEEP-PROMOTING SUBSTANCES 613
activity in the course of the habitual sleep period and during recovery sleep after extended waking. Process C represents the circadian modulation of two thresholds: one defining sleep termination and the other defining sleep onset. In the model, process S oscillates between the two thresholds. Experimental sleep-duration data obtained at various circadian phases served to character- ize process C as a skewed sinusoidal function (60). Permissive factors may shift the level of the two thresholds, thereby precipitating or delaying the onset or the end of sleep. Simulations with the two-process model were able to account for such diverse phenomena as recovery from sleep deprivation, circadian phase dependence of sleep duration, sleep during shift work, sleep fragmentation during continuous bed rest, and internal desynchronization in the absence of time cues (60). The regulation of REM sleep has been incorpo- rated so far only on a qualitative level (27). Basic features of the model apply also to animal sleep (30, 384, 386).
III. SEARCH AND IDENTIFICATION OF SLEEP FACTORS
A. Approaches Based on Total Sleep Deprivation
1. Early studies
Early in our century Pieron and co-workers (214,215,289) investigated the presence of sleep-inducing factors in the dog. After sleep-depriving ani- mals for 10 days they extracted cerebrospinal fluid (CSF) by cisternal punc- ture and injected it into the fourth ventricles of naive recipients. Sleep was induced 2-6 h after the infusion. The effect was attributed to a “hypnotoxin” that was nondialyzable and thermolabile. The interpretation of these early studies is hampered by the lack of quantitative measurements and proper control experiments, the increase in CSF pressure caused by the injections, as well as the pain and stress involved in the procedure. It is an interesting manifestation of the zeitgeist that Ishimori (158) conducted similar studies in Japan at just about the same time.
Pieron’s hypnotoxin theory was examined by Schnedorf and Ivy in 1939 (332). From dogs kept awake for 7-16 days, 8 ml of CSF was removed by aseptic cisternal puncture. Blood-tinged samples were discarded. From 20 naive recipient dogs, an equal amount of CSF was removed and replaced by the extracted fluid from the donor animal. On the basis of behavior and responsiveness to stimuli, induction of sleep was observed in 9 of 20 animals but only in 4 of 24 control animals that had received CSF from nondeprived donors. The responding animals showed a gradual reduction of vigilance and most were “soundly asleep” between 45 min and 3-4 h after the injection. It is important to note that any replacement of CSF induced a rise in body tem- perature and intraventricular pressure. Although the authors were careful to

614 ALEXANDER A. BORBELY AND IRENE TOBLER Vohm 69
point out that the induced sleep was not normal, they assumed that they had essentially confirmed Pieron’s findings.
2. Factor S
A major series of studies was initiated by Pappenheimer in 1965. He and his co-workers used the goat as donor animal that, due to its thick occipital bone and large ventricular cavity, allowed the continuous or repeated ex- traction of CSF from the cisterna magna. The procedure did not involve pain and major stress and could be therefore performed without anesthesia. The goats were sleep deprived for up to 72 h by using a conditioning procedure to prevent them from lying down (98). When CSF from sleep-deprived donors was infused into the ventricle of cats and rats, behavioral sleep was induced (279). This effect was confirmed in the rat on the basis of quantitative motor activity recordings (98, 279). After a slow infusion of CSF from sleep-de- prived donors before the dark period, motor activity was depressed during the first half but not during the second half of the night. Pyrogen-free saline served as the control infusate. The effect was enhanced as a function of the duration of sleep deprivation in the donor animal (0, 24, and 48 h) (98). However, a 72-h sleep deprivation caused no further depression of motor activity. The unknown depressive factor was designated as factor S. Early ultrafiltration experiments showed it to be present in the low-molecular- weight fraction (98). Interestingly, an excitatory factor (factor E), inducing a long-lasting hyperactivity, was also obtained.
After these early experiments the rabbit became the standard animal for bioassays because, unlike the rat, it has no prominent circadian sleep/ waking rhythm and the infusion procedure itself does not affect sleep notice- ably (see, however, Ref. 405). In addition, the rabbit has very little REM sleep, which allowed the analysis to be restricted to NREM sleep. The dura- tion of NREM sleep was assessed by visual scoring of the EEG records. In addition, slow-wave activity (0.5-4 Hz) was determined for successive 2-min epochs by integrating the filtered and rectified EEG signal (278). The mate- rial for purifying and concentrating factor S was derived not only from CSF but also from the brain tissue of sleep-deprived goats, sheep, rabbits, and later from the brain stem of slaughterhouse cattle (202). The fate of the latter animals before death was not specified. A 900min infusion of purified and concentrated material caused an increase in sleep duration and slow-wave / activity, which showed a maximum after 3-5 h (278). These changes were similar to those observed during recovery from 24-h sleep deprivation but the time course differed, since in sleep-deprived animals the maximum effect was observed already in the 1st h. Factor S had no effect on slow-wave activity in the waking EEG (202). Treatment of the fraction with pronase eliminated the biological activity. It should be noted that in the rat also the purified concentrate obtained from CSF of nondeprived goats showed a

April 1989 ENDOGENOUS SLEEP-PROMOTING SUBSTANCES 615
marked, long-lasting depression of nocturnal locomotor activity that was only slightly smaller than that caused by CSF from sleep-deprived donors (278).
Because insufficient CSF was available for the further purification of factor S, human urine was used for the chemical analysis. Its use was based on the observation that factor S was chemically stable and therefore would probably be absorbed from the CSF into the blood and concentrated in the kidney. The urinary factor was shown to affect sleep in the rabbit similar to the factor obtained from brain (195). Sleep increased in the 2nd h after administration, reached a maximum after 3-5 h, and remained above control up to 10 h. The animals showed periodic waking periods with eating, drink- ing, and grooming. Chemically the two factors were similar, but urinary factor S differed from the brain-derived factor in that it caused an inconsis- tent increase in EEG slow-wave activity. Intracerebroventricular infusion of the urinary factor to rats before dark onset enhanced nocturnal NREM sleep already in the first postinfusion hour. The effect lasted up to 12 h (195). The NREM sleep-enhancing effect of factor S from brain stem material from cattle was confirmed in the rabbit (330), whereas in the rat the result could not be replicated (240).
Urinary factor S infused intracerebroventricularly in cats increased NREM sleep (but not slow-wave activity) for 10 h with a maximum after 4 h (109). A transitory decrease was seen in REM sleep in the first 4 h, which was compensated subsequently. Normal sleeping postures were assumed by the animals.
In an effort to localize the site of action of urinary factor S, a highly purified fraction was administered by microinjection into various areas of the rabbit brain (110). At 8 of 52 injection sites NREM sleep was enhanced for 25 h. Most active sites were located in a region extending from the basal forebrain at the level of the optic chiasm to the mesodiencephalic junction.
3. Muramyl peptides
Factor S extracted and purified from human urine and from brain mate- rial was identified as a small glycopeptide containing glutamic acid, alanine, diaminopimelic acid, and muramic acid in molar ratios of 2:Z:l:l (195, 203). Muramyl peptide was found also in brain material (203). Intracerebroven- tricular infusion of 5 pmol/kg of final purified urinary factor S enhanced NREM sleep in rabbits by 50% and augmented EEG slow-wave activity. The time course was similar to that observed previously. Sleep postures appeared normal, and the animals were easily aroused by noise.
In a next step, the structure of urinary factor S was determined by fast atom bombardment-mass spectrometry (226). The major somnogenic constit- uent was N-acetylglucosaminyl-l,6-anhydro-2\r-acetylmuramylalanyl- glutamyldiaminopimelylalanine with a molecular weight of 921. The intra-

616 ALEXANDER A. BORBELY AND IRENE TOBLER Volume 69
cerebroventricular administration of 1 pmol of this anhydromuramyltetra- peptide (AMTP) was sufficient to induce excess NREM sleep in the rabbit (199). The effect was evident in the second hour and persisted during the remaining 5 h of the recording period. A second component of urinary factor S, an anhydromuramyltripeptide (mol wt 850) enhanced NREM sleep only at higher doses. A hyperthermic response paralleled the somnogenic action. At higher doses (20 pmol) abnormal responses, including seizures, were observed with both peptides. Amidation of several analogues to the urinary material blocked the biological activity. The authors proposed that amidation/deami- dation reactions as well as hydration/dehydration reactions (205) may serve to inactivate/activate the sleep substance.
Recently the two somnogenic components of urinary factor S were ob- tained from enzymatic digests of bacterial peptidoglycan (205). Of the 14 muramyl peptides tested, 6 compounds were both somnogenic and hyperther- mic, whereas 3 compounds were only hyperthermic. The high somnogenic potency of AMTP was confirmed (196). The lo-pmol dose caused a more consistent increase in NREM sleep duration and in slow-wave activity than the 1-pmol dose. Although the effect on REM sleep was variable, there were indications of a reduction of this sleep state after the higher dose. The time course of the hyperthermic response paralleled the increase in sleep. How- ever, the changes in brain temperature that normally accompany the transi- tion between vigilance states persisted after AMTP. The authors concluded that the two somnogenic components identified from urinary factor S (i.e., AMTP and the anhydromuramyltripeptide) are likely to be of bacterial ori- gin, because bacteria are the only known source of these substances (205). They cite evidence for the presence of muramic acid in mammalian tissues, for the absorption of peptidoglycans from the intestinal lumen, and for their release by macrophages. Thus muramyl peptides may have vitamin-like properties in that they may be required but cannot be synthetized by the organism.
The somnogenic action of muramyl peptides is not restricted to the substances isolated from urine or bacteria. The simple synthetic dipeptide N-acetylmuramyl-L-alanyl-D-isoglutamine (muramyl dipeptide or MDP), that has long been known as an immunostimulant, enhanced, on intracere- broventricular administration, NREM sleep in the rabbit and cat (66, 204, 348,350, 351). In the rabbit, an increase of NREM sleep and a depression of REM sleep was observed. The effect was present at different ambient tem- peratures (351). In the cat the somnogenic action was preceded by a suppres- sion of deep NREM sleep and REM sleep for 2-4 h. The somnogenic action in the rabbit was also seen after systemic administration of MDP. However, disturbances of autonomic functions were present. The febrile response oc- curred in both rabbit and cat. Pretreatment with acetaminophen attenuated the pyrogenic response to MDP (iv) in the rabbit without abolishing the somnogenic response. The inhibition of NREM sleep induced by amphet- amine was to some extent counteracted by pretreatment with MDP 125 pmol

April 1989 ENDOGENOUS SLEEP-PROMOTING SUBSTANCES 617
icv, whereas the NREM sleep inhibition induced by physostigmine remained unaltered (350). A 10-h intracerebroventricular infusion of MDP (2 nmol) in the rat enhanced nocturnal NREM sleep without affecting REM sleep and increased brain temperature (152). On the other hand, in rats, MDP (50-500 pg/kg; 0.1-l pmol/kg ip) administered at the beginning of the 12-h light period was neither somnogenic nor pyrogenic (102). At the highest doses (250 and 500 pg/kg; 0.5 and 1 pmol/kg), REM sleep was reduced during the first 6 h after injection and showed a rebound increase in the second 6 h. Muramyl dipeptide (25-2,000 pg/kg; 51 nmol/kg to 4.1 pmol/kg iv) administered to rats at hour 3 of the light period did not affect NREM sleep (170). The REM sleep was enhanced by the lowest dose and reduced by the highest dose. However, the 25-pg/kg (51 nmol/kg) dose, when administered at dark onset, enhanced nocturnal NREM and REM sleep. In the squirrel monkey, MDP (50 nmol ip or iv) had a somnogenic and pyrogenic action when administered either during the circadian period of activity or rest (417). An initial suppression of REM sleep was noted.
Various MDP analogues have been tested for their somnogenic activity (177, 209, 351). Krueger and co-workers (196) conclude that to date every somnogenic muramyl peptide has also been found to be pyrogenic. Moreover, these substances can induce the synthesis and release of the immunologically active polypeptides interleukin 1 and interferon a-2, which in turn have a sleep-promoting action. Unlike factor S and muramyl peptides, the somono- genie effect of these substances was not discovered by sleep-deprivation ex- periments; therefore they are discussed later (sect. IIIF).
4 Sleep-Promoting Substance
In the early 197Os, Uchizono and co-workers (151,260) started a series of experiments to investigate the presence of endogenous sleep-promoting sub- stances in rat brain. Groups of rats were sleep deprived for 24 h by intermit- tent exposure to electric foot shocks (35 mA, 30-40 V, 50 Hz) through the grid floor of their cage. The shocks were applied every 3 min for 1 min and were signaled by a light so that the rats could avoid the shock by moving to another compartment of the cage. The stress involved in the deprivation procedure was reflected by the increased weight of the adrenal glands (151). After the sleep-deprivation period the animals were decapitated, and the brain stem, including mesencephalon and hypothalamus, was removed. The material was homogenized, dialyzed, and lyophilized. Injection of 6 units ip (the equivalent of the material obtained from 6 sleep-deprived rats) to naive recipient rats in the middle of the 12-h light period reduced their nocturnal locomotor activity and enhanced nocturnal EEG slow-wave activity (l-4 Hz) (260). The somnogenic component contained in the brain extract was desig- nated as Sleep-Promoting Substance (SPS). Nocturnal locomotor activity was also reduced when 5 units of SPS were infused intracerebroventricularly

618 ALEXANDER A. BORBELY AND IRENE TOBLER t%kwme 69
during the dark period (387). Mater,ial from nondeprived rats caused no change. Three units administered by the same route to mice had a similar effect and, in addition, enhanced EEG slow-wave activity (2-4 Hz). The im- mediate response during the infusion was followed by a delayed somnogenic response several hours later. A dose-dependent increase of NREM sleep (dose range 0.05-0.4 units) and REM sleep (0.05-0.2 units) was observed in mice after intraperitoneal injection 1 h before dark onset (261). Whereas the in- crease of NREM sleep occurred within 1.5 h, the rise in REM sleep was delayed by 9 h. Changes in both sleep states were still 18 and 20 after SPS administration. Mendelson and
present co-work
between hours .ers (240) were
unable to replicate these results in the rat. Further purification of SPS allowed them to identify three different
fractions with somnogenic activity (187,388). One of the fractions was iden- tified as uridine. The fraction designated as SPS-B was administered by continuous infusion into the third ventricle of unrestrained rats (136, 152, 153). Infusion was started 1 h before dark onset and proceeded throughout the first 9 h of the 12-h dark period. Infusion of saline for 24 h served as the control. Two brain stem equivalents of SPS-B caused a marked increase of nocturnal NREM and REM sleep. Whereas NREM sleep was at base-line level during the following light period, REM sleep was reduced below base line. Apart from uridine the somnogenic substances in SPS have not been identified.
5. Uridine and cytidine
Uridine, one of the somnogenic constituents of SPS, was shown to en- hance both NREM and REM sleep in the mouse at a dose of 1 pmol ip (187). In the rat the effect depended on the time of administration. Thus a lo-pm01 dose infused over 10 h during the dark period (start of infusion: 1 h before dark onset) enhanced both NREM and REM sleep, whereas no effect was observed when infusion was carried out 12 h later during the light period (137, 150, 152, 153). Similarly, intracerebroventricular injection of 1, 10, or 100 nmol in the light period was ineffective (306). The nocturnal infusion of high doses of uridine (loo-1,000 pmol) did not affect sleep (150). Uridine had no effect on brain temperature (152). Uracil (l-100 pmol), the base moiety of uridine, was ineffective (150). The combined administration of uridine with other sleep-promoting compounds [MDP and &sleep-inducing peptide (DSIP) alone or in combination] induced complex changes of sleep (147,183).
In the rat, uridine injected intraperitoneally (100 pmol, i.e., ~30 mg/kg) caused a l-h rise in NREM sleep but only when administered before dark onset and not at light onset (139). However, administration in the 3rd h of the light period induced a short-lasting increase in REM sleep. Higher or lower doses were also ineffective.
The action of uridine was studied also in the rhinoceros beetle, a night- active insect (154). Intra-abdominal injection of l-10 pmol reduced slightly nocturnal locomotor activity, whereas 0.1-10 nmol had the opposite effect.

April 1989 ENDOGENOUS SLEEP-PROMOTING SUBSTANCES 619
Cytidine, another pyrimidine ribonucleoside, was not somnogenic in the rat when administered at doses of 10 pmol icv during the night (138) or l-100 nmol icv during the day (306).
6. Specific changes in brain mRNA
The effect of sleep on RNA synthesis has been investigated in an early study in the rabbit (401). The relative proportion of different types of RNA synthetized in the cerebral cortex and separated by sedimentation on sucrose gradients depended on the amount of synchronized sleep occurring during the period of incorporation of orotate. Subsequently, it was shown that syn- chronized sleep is accompanied by a marked accumulation of newly synthe- tized heterogenous nuclear RNA in cortical neurons (114,115; for review see Ref. 113). The functional significance of the increased level of mRNA pre- cursors and/or regulatory sequences is still unknown.
Recently, the existence of sleep-related changes at the transcriptional level was investigated. A lo-fold-enriched recombinant library was gener- ated from rat cDNA transcribed from forebrain mRNA after subtraction of cerebellum sequences (310). A method was developed to detect clones that correspond to transcripts present at a very low abundance. Enriched, sub- tracted forebrain probes were prepared from control rats and from rats sleep deprived for 24 h by forced locomotion in a slowly rotating cylinder (36). Clones were identified that are specific to the central nervous system and the level of which is modulated by enforced waking (310a). The specificity of these changes remain to be ascertained.
7. Varia
The somnogenic effect of plasma dialysates from sleep-deprived rabbits was investigated (312). The rabbits were sleep deprived in a slowly revolving drum (1 rotation/3.7 min) and by a combined optic and acoustic alerting device activated for 20 s every 10 min. Plasma from sleep-deprived and con- trol rabbits was obtained by intracardiac puncture. The dialysates were ad- ministered intravenously to rabbits and rats, and sleep was recorded for 45 min after the injection. No difference was observed between animals receiv- ing material from sleep-deprived or control rabbits. Similarly, the motor activity of mice was not affected during the 30 min after the intravenous injection. It should be pointed out that due to the short recording period, delayed effects would not have been noted.
In a pilot study, the somnogenic effect of CSF obtained from sleep-de- prived rats was investigated (39). Donor rats were sleep deprived for 24 h by placing them into a slowly rotating (1 rotation/45 s) cylinder (36). Cerebro- spinal fluid from experimental and control rats was obtained by cisternal puncture. Slow infusion of the former material into the lateral ventricle of recipient rats at the beginning of the 12-h light period enhanced sleep in postinfusion hours z and 3 in comparison to the control infusion.

620 ALEXANDER A. BORBELY AND IRENE TOBLER Volume 69
B. Approaches Based on Rapid-Eye-Movement Sleep Deprivation
Evidence for the presence of a REM sleep-promoting factor was first obtained in the cat (322,323). Donor cats were REM sleep deprived for 17 h by the flower pot method. After the deprivation period, the cats were allowed to feed and then were recorded for 24 h. During the recovery period, REM sleep was increased by 9.2% (P < O.OOl), whereas deep NREM sleep (SWSZ) was enhanced by 3.6% (P < 0.1). Cerebrospinal fluid was withdrawn from the cisterna magna after the occurrence of the first REM sleep episode.
The recipient cats were pretreated with p-chlorophenylalanine (PCPA) (400 mg/kg ip on 2 consecutive days), which suppressed REM sleep and SWSZ. Cerebrospinal fluid from donor animals (250 ~1) was infused into the fourth ventricle l-2 days after the last PCPA injection. In 12 of 13 experi- ments REM sleep reappeared with a latency ranging from 14 to 178 min and was always preceded by light NREM sleep. The mean duration of the REM sleep-inducing effect was 324 min. Deep NREM sleep was observed in only two instances. Infusion of CSF from nondeprived donors induced REM sleep in 4 of 9 experiments with latencies and durations of action similar to those of the previous group. The concentration of serotonin (5HT) in the CSF was too low to account for this effect.
The results were essentially confirmed by another group (295). The CSF obtained from donor cats subjected to 24-h REM sleep deprivation was ad- ministered to recipient cats that had been pretreated with PCPA. During the 8-h recording period, REM sleep was partially restored. The CSF from non- deprived donors was ineffective. Likewise, heating the CSF before infusion or pretreatment of the donor animals with chloramphenicol resulted in a loss of the REM sleep-restoring action.
The suppression of REM sleep by PCPA could be antagonized by the systemic, intraventricular, or intracerebral administration of bovine neuro- intermediate lobe extracts (327). When this extract was administered sys- temically or intracerebroventricularly to chronic pontine cats in which all cerebral structures rostra1 to the pons, including the pituitary, had been ablated, REM sleep was enhanced and survival was prolonged (169). Lysine vasopressin, oxytocin, or adrenocorticotropic hormone (ACTH) were ineffec- tive in the lesioned cats.
There is evidence also in the rat that a REM sleep-promoting factor accumulates in the CSF in the course of REM sleep deprivation. Donor rats were REM sleep deprived for 1,2,3, or 4 days by the flower pot technique and by manual awakenings during 2-6 h per day (3, 4). This procedure reduced REM sleep by 93% and NREM sleep by ‘7%. Body weight was maintained throughout the deprivation period. After withdrawal of CSF via chronically implanted cannulas, the rats were recorded for a further 6 h to assess the effect of deprivation on sleep. A REM sleep rebound was observed that was a function of the duration of prior REM sleep deprivation.
The recipient rats were pretreated with DL-propranolol (10 mg/kg ip) 15-30 min before infusion of CSF into the fourth ventricle. This pretreatment

April 1989 ENDOGENOUS SLEEP-PROMOTING SUBSTANCES 621
caused a marked suppression of REM sleep. Infusion of CSF (20 ~1 icv) from REM sleep-deprived donor rats during 5 min in hour 4 of the 12-h light period increased REM sleep during the following 6 h. It took l-2 h until the first REM sleep episode appeared. The effect was most prominent in hours Z-6 postinfusion and was due to an increase in number, but not in duration, of REM sleep episodes. The extent of REM sleep restoration in the recipients was a function of the duration of REM sleep deprivation in the donors. Rapid-eye-movement sleep was restored almost to base-line levels after in- fusion of CSF from donors deprived for 4 days.
In a similar protocol the recipient rats were pretreated with DL-cw-meth- yldopa (50 mg/kg ip) 90 min before infusion with CSF (4). The deprivation and infusion procedure corresponded to that in the previous study. The CSF from REM sleep-deprived donors partially restored REM sleep in pretreated recipients without affecting NREM sleep. The main effect occurred during the second half of the 8-h recording period.
In an effort to localize the origin of the REM sleep-promoting factor, donor rats were killed after 4 days of REM sleep deprivation and their brains were removed (86). Tissue extracts were prepared from the cortex, hypothal- amus, and pituitary gland and were dissolved in artificial CSF and infused into the fourth ventricle of cu-methyldopa-pretreated rats. Recordings pro- ceeded for 20 h, and the records were analyzed in 4-h periods. Pituitary and hypothalamic extracts restored REM and deep NREM sleep to base-line levels 4-8 h after infusion, as well as during the subsequent periods. Neither the cortical extracts from deprived donors nor pituitary and hypothalamic extracts from nondeprived donors were effective.
A brain extract from donor rats that had been subjected to 100-h REM sleep deprivation was injected intraperitoneally in recipient rats in the light period (416). Compared with saline, the brain extract reduced REM sleep latency and enhanced REM sleep by 16% in the first 4 h. No effect was seen in the second 4-h period after injection. Brain extract from nondeprived ani- mals was ineffective.
C. Approaches Based on Electrical Brain Stimulation
1. Early studies
In his pioneering studies, Hess (for review see Ref. 131) reported that low-frequency electrical stimulation of the median thalamus induced sleep in the cat. Kornmiiller et al. (188) used this method to test the presence of sleep-promoting factors in the blood. In two pairs of cats with crossed arte- rial circulation, sleep was induced by electrical stimulation in one of the animals. Electroencephalogram recordings demonstrated that as early as 20-30 s after stimulation, sleep also occurred in the unstimulated cat. In view of the small number of animals and the lack of control experiments and

622 ALEXANDER A. BORBRLY AND IRENE TOBLER Volume 69
statistics, this early report cannot be regarded as strong evidence for a humoral mediation of sleep.
A cross-perfusion study in unanesthetized and paralyzed intact or enckphale isole cats revealed that stimulation of the bulbar reticular forma- tion produced cortical desynchronization not only in the stimulated animal but after a delay of 30-80 s also in the nonstimulated animal (300). The effect was attributed to a blood-borne activating factor.
Cross-circulation experiments were performed in unanesthetized rab- bits (254). Electrical stimulation of the intralaminar thalamic nuclei aug- mented EEG slow-wave activity (0.5-3.5 Hz) in the stimulated (donor) ani- mal as well as in the nonstimulated (recipient) animal. Four 30-s stimulation episodes administered at 2-min intervals were required to elicit a significant change in the recipient. Stimulation of the midbrain reticular formation reduced EEG slow-wave activity in both the donor and the recipient rabbit.
Z. Venous dialysate frm rabbit brain
Monnier and Hijsli (252, 253) induced sleep in donor rabbits by low-fre- quency electrical stimulation of the mediocentral thalamus. Drowsiness was observed within 20-30 s. The cerebral venous blood of these donors was subjected to extracorporeal dialysis during 80 min of repeated electrical stimulation. The dialysate was then injected intravenously to unrestrained recipient rabbits. The EEG recordings were obtained 20 min before and 40 min after the injection. The dialysate induced sleep that was readily revers- ible by sensory stimulation. The EEG slow-wave activity (2-3 Hz) was en- hanced lo-15 min after injection with a maximum effect between 25 and 40 min. No sleep-inducing action was observed after injection of dialysate from donors that had been subjected to sham stimulation. In contrast, dialysate from alert rabbits, activated by electrical stimulation in the mesencephalic reticular formation, induced in recipients alert behavior with EEG de- synchronization.
The active dialysate was shown to be effective not only on systemic administration but also after infusion into the third ventricle (251). Dialy- sate derived from cerebral blood was more effective than that from femoral blood (249). Fractions obtained from cerebral blood before thalamic stimula- tion caused only a minor rise in slow-wave activity of recipients. ’
Fractionation and purification procedures led to the identification of the nonapeptide Trp-Ala-Gly-Gly-Asp-Ala-Ser-Gly-Glu that was designated as &sleep-inducing peptide (DSIP) (341-343). Intracerebroventricular infusion of synthetic DSIP, but not of related peptides, caused an increase in slow- wave and spindle activity in the rabbit EEG (342). The DSIP (30 nmol/kg iv) induced a delayed and prolonged enhancement of slow-wave activity (250).
3. &Sleep-inducing peptide
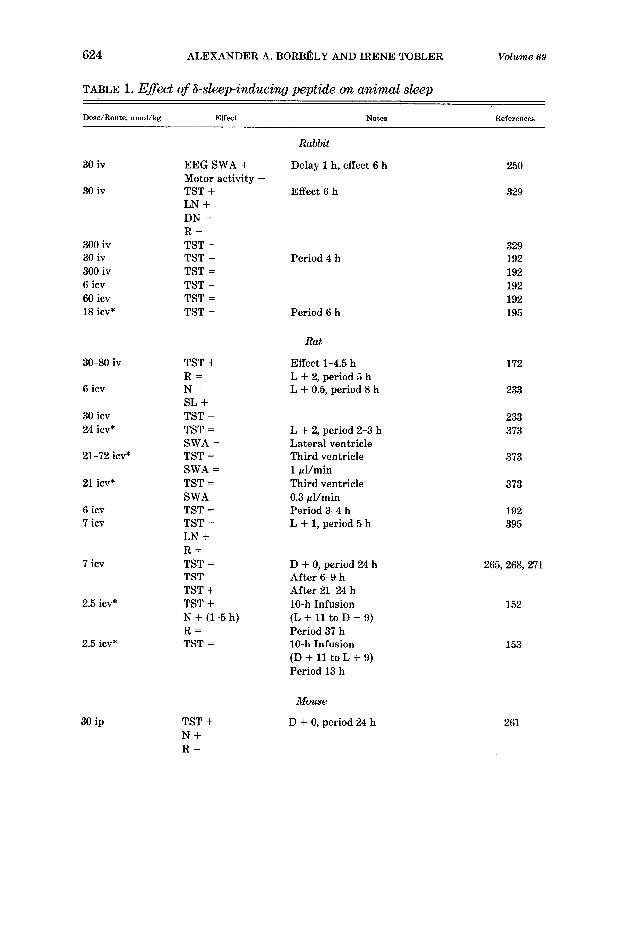
Of all putative sleep-promoting substances, DSIP is the compound that has been most extensively tested. Table 1 provides an overview of the animal

April 1989 ENDOGENOUS SLEEP-PROMOTING SUBSTANCES 623
studies. It is evident that a somnogenic action of DSIP could not be confirmed in all studies. Studies with negative results are present for every species that has been investigated by more than one group. Also a sleep-inhibiting action of DSIP has been reported in rats (233, 268, 271, 373) and in cats (352). The contradictory results cannot be accounted for by differences in the dose or in the route or time of administration.
In contrast to the early experiments with dialysates from cerebral blood, the somnogenic action of DSIP was generally prolonged and often delayed. Thus, whereas the intravenous administration of dialysate to rabbits en- hanced EEG slow-wave activity within IO-15 min and caused a maximum effect after 25-40 min (252, 253), the somnogenic effect of synthetic DSIP appeared only after 1 h and was still present after 6 h (250). Also in the rat, cat, and dog, the somnogenic actions of DSIP lasted several hours. Even effects occurring after 1 day or more have been reported (268, 373). On the other hand, during a 10-h continuous nocturnal intracerebroventricular in- fusion, the increase in sleep was limited to the first 5 h, and no late effects were observed during the next 24 h (152).
The effects of DSIP on sleep stages are inconsistent. In the rabbit, EEG slow-wave activity (corresponding presumably to deep NREM sleep) was enhanced (250), whereas an increase of light NREM sleep and a suppression of REM sleep have also been observed (329). In the eat, an increase in deep NREM sleep, a reduction or no change in light NREM sleep (359-361), or a prominent increase in REM sleep (292) have been ,meported.
The time of administration was an important variable in the rat. Intra- cerebroventricular infusion in the dark period had a somnogenic effect, whereas infusion in the light period did not enhance sleep (152, 153). A “bell-shaped” type of dose-response curve of DSIP was obtained in the cat and rabbit where a somnogenic action was observed after 30 nmol/kg but not after 300 nmol/kg (292, 329).
Microinjection of DSIP (7.5-22.5 nmol in 7 ~1) into the median thalamus, periaqueductal gray matter, or the nucleus tractus solitarii of rabbits en- hanced NREM sleep for up to 2 h (367). The response occurred within 10 min after injection and could be antagonized by local injection .of nalox- one (160 pg).
After intraperitoneal administration of DSIP or of various other unre- lated peptides, an increase in the EEG spectral power density in the low-fre- quency range was observed (241,242). However, since these results are based on 10-s samples of EEG without identification of the vigilance state or behav- ior, they are difficult to interpret.
In a recent paper (158a) it was claimed that in rats the intracerebroven- tricular injection of antiserum against DSIP (anti-DSIP) prevented the en- hancement of both NREM sleep and plasma growth hormone (GH) by 4-h sleep deprivation. Significant differences were reported for the values after anti-DSIP treatment in comparison with those after treatment with normal rabbit serum. However, the authors chose not to compare recovery sleep after anti-DSIP treatment with recovery sleep without any treatment. The data
624 ALEXANDER A. BORBfiLY AND IRENE TOBLER
TABLE 1. Eflect of &sleep-inducing peptide on animal sleep
voktcme 69
Dose/Route, nmol/kg Effect Notes References
30 iv
30 iv
300 iv 30 iv 300 iv 6 icv 60 icv 18 icv*
30-80 iv
6 icv
30 icv 24 icv*
21-72 icv*
21 icv*
6 icv 7 icv
7 icv
2.5 icv*
2.5 icv*
30 ip
EEG SWA + Motor activity TST + LN + DN= R- TST = TST = TST = TST = TST = TST =
TST + R= N- SL + TST - TST = SWA = TST = SWA = TST = SWA - TST = TST = LN+ R= TST = TST - TST + TST + N + (l-5 h) R= TST =
TST + N+ R=
-
Rabbit
Delay 1 h, effect 6 h 250
Effect 6 h 329
Period 4 h
Period 6 h
Rat
Effect l-4.5 h L + 2, period 5 h L + 0.5, period 8 h
L + 2, period 2-3 h Lateral ventricle Third ventricle 1 pl/min Third ventricle 0.3 jJ/min Period 3-4 h L + 1, period 5 h
D + 0, period 24 h After 6-9 h After 21-24 h 10-h Infusion (L + 11 to D + 9) Period 37 h 10-h Infusion (D + 11 to L + 9) Period 13 h
Mouse
D + 0, period 24 h 261
329 192 192 192 192 195
172
233
233 373
373
373
192 395
265,268,271
152
153

April1989 ENDOGENOUS SLEEP-PROMOTING SUBSTANCES 625
TABLE L-Continued
Dose/Route, nmol/kg Effect Notes References
30 ip
30 iv
300 iv 7 icv
7 icv
120 SC
Cat
TST - LN - R- RL + TST + N+ R+ TST = TST = N= R= LN - DN+ TST + N+ R= LN - DN+ SL - TST = N= R= LN = DN+ RL =
Effect/period 10 h Max 2-4 h
Effect/period 6 h
No. R episodes +
Period 8 h after 72-h R deprivation
Effect 7 h, period 10 h
Period 8 h
352
292
292 360
361
359
24 icv TST + Effect l-4 h N=l
365
+, Increase; =, no significant change; -, decrease (comparison to placebo). ip, Intraperito- neal; iv, intravenous; icv, intracerebroventricular; SC, subcutaneous. EEG, electroencephalo- gram; SWA, slow-wave activity; TST, total sleep time; R, rapid-eye-movement (REM) sleep; N, non-REM (NREM) sleep; LN, light NREM sleep (SWS-1); DN, deep NREM sleep (SWS-2); SL, sleep latency; RL, REM sleep latency; L + x, D + y, administration x h after light onset, y h after dark onset; period, recording period; max, maximum effect. *Dose in nanomoles per animal.
reported in the paper show that NREM sleep was at a similar level under the two conditions. It seems therefore that anti-DSIP treatment did not block the enhancement of NREM sleep but that the normal rabbit serum treatment itself increased the level of NREM sleep (to -40% of recording time in the first 2 h of recovery sleep). Note, however, that even this value is low in comparison to the rat’s typical base-line level in the light period (-60% of recording time) (36, 40, 384). The repeated withdrawal of blood samples via an implanted jugular catheter for determining GH levels may have disturbed

626 ALEXANDER A. BORBELY AND IRENE TOBLER Volume 69
sleep. The data presented also show that the plasma levels of GH after anti-DSIP treatment and without any treatment were similar. It is therefore premature to conclude that DSIP is a physiological stimulus for the induction of NREM sleep and for a sleep-related release of GH.
Motor activity, a reliable indicator of active waking, was not reduced when DSIP (40 or 80 nmol/kg ip) was administered to rats before the 12-h dark period (373). There was even evidence for a stimulating effect after the 80-nmol/kg dose. A delayed reduction of activity 1 and 2 days after adminis- tration was observed after 160 nmol/kg. In another study, 94 nmol/kg ip did not affect motor activity within the first 60 min after administration (242). An activity-reducing action of DSIP (120 nmol/kg ip) was reported for rats maintained at 4OC (425). Doses of 10 or 30 times higher were ineffective. In rats maintained at room temperature, none of the doses induced behavioral sleep (426). Finally, a reduction of nighttime activity and an increase in daytime activity was described whenDSIP (30 nmol/kg iv) was administered before dark onset (120). However, since no statistical analysis is reported, the results cannot be evaluated. The repeated daily injection (30 nmol/kg iv) in animals maintained in constant light caused a decrease of motor activity that was followed by an increase (119). The authors interpret the resulting 24-h rhythmicity as evidence for a “circadian programming” action of DSIP (see also Refs. 90,336). The unconventional way of assessing the changes of motor activity makes the evaluation of the results difficult.
A dose of 120 nmol/kg ip, but not larger doses (1.2 or 3.6 pmol/kg), induced hypothermia in rats maintained at 4OC (425). At room temperature (22”C), hyperthermia was observed after all three doses. These results are in contrast to those of another study in which no change of body temperature occurred after intracerebroventricular injection of 7 nmol/kg DSIP (271).
The effect of DSIP on model insomnia was investigated. The stress-in- duced sleep disturbance in rabbits that had been exposed to dogs was coun- teracted by pretreatment with DSIP (30 nmol/kg iv) (329,331). Pretreatment of cats with the same dose antagonized morphine-induced insomnia. Further interactions of DSIP with other pharmacological agents include nalorphine in morphine-dependent mice (329) and amphetamine (effect on motor activ- ity and body temperature) in the rat (425) and mouse (123).
The literature concerning effects of DSIP other than sleep has been reviewed (97, 121, 122, 124, 180). These studies deal with electrophysiological actions, behavioral effects, and drug interactions, as well as with the distri- bution and metabolism of DSIP-like immunoreactive material. In the context of the present review note that 24-h sleep deprivation by handling did not affect the level of DSIP-like material in rat brain (181,259). A 24-h rhythm in the plasma level of DSIP-like activity has been reported (100).
&Sleep-inducing peptide has been investigated also in humans. In a double-blind, placebo-controlled crossover schedule a dose of 25 nmol/kg was administered by a 200min intravenous infusion during the day (337). In the 130-min postinfusion recording period the median sleep time after DSIP was

April 1989 ENDOGENOUS SLEEP-PROMOTING SUBSTANCES 627
47.25 min and after placebo was 29.75 min (difference not significant). A tendency toward a reduced sleep latency and an increased sleep efficiency as well as a significant reduction of the percentage of stage 1 sleep was observed in the night after daytime DSIP administration. A significant increase of slow-wave and REM sleep was also reported, yet no data were presented. In another similar study, 10 subjects were administered 25 nmol/kg DSIP iv in the morning (21). Sleep duration after injection did not differ significantly from placebo, although a significant increase in sleep was noted between 120 and 160 min after administration. The same authors infused DSIP to eight subjects before bedtime. Total sleep time and the sleep stages did not differ from the preceding placebo night, but sleep was enhanced during the with- drawal night.
Several studies were devoted to the therapeutic effect of DSIP in insom- nia. When administered to six insomniac patients, DSIP (25 nmol/kg iv) decreased the number of arousals during sleep as well as the percentage of stage 1 sleep and increased total sleep time and sleep efficiency (337, 338). The somnogenic effect became manifest only in the 2nd h after injection, whereas a slight sleep-inhibiting effect was noted in the 1st h. Sleep latency was not shortened. In a further study, sleep in four insomniacs was prolonged during a 4-day treatment with DSIP (25 nmol/kg iv 1 h before bedtime) in comparison with the preceding placebo value (339). Beneficial effects of DSIP treatment on daytime activities were also reported. They include higher alertness and improved performance, stress tolerance, and coping ability. The authors suggest that beyond its somnogenic action, DSIP is involved in regulating behavioral patterns and their proper physiological functions. Fur- ther case reports have been published (333). The increase in total sleep time during a 1-wk treatment was reported to persist during the follow-up week (334). In an open study, beneficial effects of a lo-day treatment with DSIP were observed in six of seven insomniac patients (171). Recently, a dose of 30 nmol/kg iv administered on 7 successive days before bedtime was tested in 14 chronic insomniacs (335). On 2 days before drug administration and on the first withdrawal day, placebo was injected under double-blind conditions. Sleep recordings were obtained on three nights before DSIP (including the adaptation night), on the first and last night of drug treatment, ‘and on the withdrawal night. A significant increase in total sleep time and sleep effi- ciency and a decrease in sleep latency and waking after sleep onset was observed in the DSIP nights. Sleep was already improved in the first DSIP night, and the further improvement was not significant. The changes per- sisted in the withdrawal night. The treatment did not significantly affect the percentage of REM sleep and slow-wave sleep. It is unclear how the single base-line value that served as the reference for statistics was calculated from the three pre-DSIP nights.
Taken together, the various reports of this group suggest a beneficial effect of DSIP in the treatment of insomnia. However, since placebo was administered before and/or after the DSIP treatment period without a cross-

628 ALEXANDER A. BORBELY AND IRENE TOBLER Vohme 69
over design, nonpharmacological sleep-promoting effects due to the thera- peutic setting (e.g., repeated injections before bedtime, the high expectations of patients and physicians, change in daily routine in a clinical setting) cannot be ruled out. It is therefore premature to conclude that DSIP has a therapeutic action.
The sleep-promoting action of DSIP was not confirmed in a recent study in six insomniac patients (255). These authors administered on 4 consecutive nights either DSIP (25 nmol/kg iv) or placebo 1 h before bedtime in a dou- ble-blind crossover design. The treatment nights were preceded by two base- line nights and were followed by two withdrawal nights. A comparison be- tween the three periods (i.e., base line, DSIP/placebo, withdrawal) yielded no significant treatment effects on any polygraphically recorded sleep parame- ter. Non-REM sleep time and stage 2 sleep were significantly higher for DSIP than for placebo, but differences in the same directions were already present in the preceding base-line nights. The authors concluded that sleep improve- ment induced by DSIP was of little clinical significance.
4. b-Sleep-inducing peptide analogues
When the action of synthetic DSIP on rabbit EEG was first investigated, eight related peptides consisting of DSIP residues or analogues were found to be ineffective (343).
In an effort to stabilize the peptide against proteolytic degradation, D-amino acids were introduced in the DSIP molecule. Kovalzon and co- workers (190,191) studied the effect of such analogues on sleep in the rabbit and rat. In rabbits [D-Trpl]-DSIP, [D-Tyrl]-DSIP, and [D-Ala’]-DSIP (8 nmol/kg icv) were infused. A rise of NREM sleep occurred, while REM sleep was unaffected. Elevating the dose of the tryptophan analogue by a factor of 100 caused no further increase of NREM sleep but did cause a reduction of REM sleep. All three peptides were ineffective after intravenous injection. The authors proposed that the somnogenic action of DSIP analogues may be attributed to the preservation of the active structure due to their increased stability to enzymatic degradation.
In the rat, both the tryptophan and tyrosine analogues (7 nmol/kg) enhanced nocturnal sleep after intracerebroventricular administration be- fore dark onset (191,265,268). The analysis by 3-h periods revealed a signifi- cant increase of NREM (both analogues) and REM sleep (tyrosine analogue) (268). When administered in the light period, the tryptophan analogue was ineffective (190). The shortened analogue [D-Trpl]-DSIP-(l-6) was not som- nogenic in the rabbit and even reduced total sleep time in the rat during the first 3 nighttime h after injection (191, 268). A further analogue, Q-NH,-ca- prilyl-DSIP (7 nmol/kg iv), did not enhance sleep when injected in rats before dark onset (271). As has been observed for DSIP, the analogue reduced sleep in hours 6-9 after administration. It has been suggested that DSIP may contain a fragment that increases waking instead of sleeping (268).

April 1989 ENDOGENOUS SLEEP-PROMOTING SUBSTANCES 629
The DSIP analogue with the terminal glutamine replaced by glycine was studied in the rat and cat (176). Suboccipital injection of 16 and 20 nmol/kg increased sleep (particularly deep NREM sleep), suppressed REM sleep, and reduced the responsiveness to sensory stimulation. No statistics were re- ported. The serine phosphorylated DSIP analogue was investigated in the rat (119, 120). A dose of 0.1 nmol/kg iv attenuated the circadian rest-activity rhythm in rats kept under light-dark conditions, changed the pattern of the rhythm, and increased activity in rats kept in continuous light. Statistics were not provided. Sleep-promoting effects have been reported after noctur- nal administration to rats [ZO or 200 pmol/kg icv (216a); 0.5 nmol icv (183a)]. Finally, [D-Ala4]-DSIP-NH2 (85 nmol/kg ip), an analogue that enters the brain more readily than DSIP, reduced the locomotor activity of rats and enhanced the low-frequency EEG activity (242). The latter effect was more prominent than for DSIP. However, the result is difficult to interpret, since data on sleep are lacking.
D. Approaches Based on Spontaneous Sleep
I. Crossed circulation studies
Pairs of rats were connected parabiotically, and their sleep was recorded polygraphically for Z-day periods Z-15 days after parabiotic union (228). The estimated blood exchange between the animals was 1% of total blood volume per minute. Control rats were united by their skins only. The rate of synchro- nous occurrence of NREM and REM sleep was higher in parabiotic rats than in controls. Although the synchronized occurrence of waking and sleeping may have been due to the mutual disturbance of the animals, this is not the case for the synchronization of REM sleep episodes. The authors therefore conclude that a humoral REM sleep-inducing factor may be present.
A dog was implanted with a second head so that the implanted head received its blood supply from the host animal (68). Electroencephalogram recordings were obtained simultaneously from both heads. During the 108-h survival period, the two recordings indicated independent sleep/waking cycles and little synchrony between the vigilance states.
Human cross-circulation studies of sleep were obtained from conjoined twins. In craniopagus twins with shunting of arterial blood from twin I to twin 2, complete independence of the sleep/waking cycle and of the occur- rence of active and quiet sleep was observed on days 4 and 11 after birth (216). The authors cite six other studies of craniopagus twins who manifested independent sleep and waking episodes despite extensive circulatory connec- tions. Conjoined twins with a common heart and circulatory system were observed during days 14 and 15 after birth for 11 h (407). Scoring of sleep, waking, active sleep, and quiet sleep for 1-min epochs based on observation revealed an independent occurrence of the vigilance states. These results

630 ALEXANDER A. BORBfiLY AND IRENE TOBLER vohme 69
contrast with a recent study in craniopagus twins (213a) whose sleep was recorded for 9.5 h at 120wk postgestation and for 24 h at 16-wk postgestation. With the use of scoring intervals of 5 min, the percentage of synchronous onset of sleep at 16 wk was 6’7%, of active sleep onset was 47%, and of quiet sleep onset was 65%. The author states that sleep appears to be synchronous beyond chance expectations and suggests that a maturational effect may account for the discrepancy between his results and those of the other au- thors mentioned above.
It should be mentioned in this context that in some species of aquatic mammals, EEG signs of NREM sleep have been reported to occur alternately in the two cerebral hemispheres, a phenomenon designated as “unihemi- spheric sleep” (257, 258).
2. Crossed-brain perfusion studies
By means of push-pull cannulas the mesencephalic reticular formation (MRF) of donor and recipient cats was cross-perfused (72,80). Perfusate from a spontaneously sleeping donor animal induced sleep when infused into an awake recipient animal. The effect consisted of a shortened sleep latency and an increased NREM sleep compared with sleep after perfusion with Ringer solution or without perfusion. Infusing the perfusate into the brain of sleep- ing cats augmented REM sleep (316). Perfusate from awake donors did not affect sleep latency in awake recipients (316) but prolonged REM sleep la- tency (72), whereas administration to sleeping recipients enhanced waking (316).
The same experimental procedure was used to analyze the protein con- tent of perfusates collected during waking and REM sleep (81,83,353). Cyclic variations in the protein level were observed in the course of the sleep/wake cycle. The protein concentration was higher during REM sleep than during waking. Two proteins were detected that were present in REM sleep perfus- ates but not in wake perfusates (353), a finding that was not confirmed in later experiments (85). Forebrain lesions resulted in reduced sleep and in fragmented REM sleep, changes that were associated with the disappearance of the cyclic release of proteins in the perfusate (76). When antibodies raised to MRF proteins were injected into the MRF of cats, REM sleep was reduced (84,85). However, as the authors themselves point out, this may represent a nonspecific effect.
3. Transfer of cerebrospinal jluid
In an effort to identify humoral correlates of the circadian rest-activity cycle, CSF was obtained from donor rats either during the dark (active) or, light (inactive) period of their 24-h cycle and infused intracerebroventricu- larly during 30 min in recipient rats (321). Infusion in the recipients was

April 1989 ENDOGENOUS SLEEP-PROMOTING SUBSTANCES 631
terminated either 2 h before light onset or 2 h after light onset. The CSF from light-exposed donors reduced the motor activity of recipients in the dark period for 2 h, whereas CSF from dark-exposed donors increased activity of recipients in the light for 1 h.
E. Neuromodulators and Neurotransmitters
In this section various classes of endogenous compounds are discussed that possess a somnogenic action or have been implicated in sleep regulation. Because the classic neurotransmitters do not fall under the heading of endog- enous SPSs, they are not extensively discussed. Nevertheless, some transmit- ters have been included in view of new developments regarding their possible role in sleep regulation or their putative involvement in the elaboration of endogenous sleep-promoting substances. Moreover, there are persistent at- tempts to use precursors of transmitters or drugs affecting receptors to promote physiological sleep for therapeutic or experimental purposes.
The majority of substances discussed in this section could be designated as neuromodulators, as they are not involved in synaptic neurotransmission in the strict sense. They are released not only by neurons but also by other cells (e.g., neurosecretory cells), and they may affect transmitter release or modulate receptor sensitivity. For most of these compounds the somnogenic action or the involvement in sleep regulation was discovered by serendipity or as a result of screening a large number of substances.
1. Prostaglandins
Prostaglandin Dz (PGD2) induced a dose-dependent increase in NREM sleep when microinjected into the preoptic area of the rat in the dose range of 0.3-1.25 nmol (391). The effect occurred within 1 min after injection and lasted 220-350 min. Electroencephalogram slow waves were prominent, and 50% of NREM sleep was classified as deep NREM sleep. No deep NREM sleep was present after injection of saline. Prostaglandin D2 also caused bradycar- dia and hypothermia. A dose of 2.5 nmol caused a slight hyperthermia with- out affecting sleep (229). Microinjections of PGD2 into other brain sites, such as the posterior hypothalamus, were not somnogenic (391). Intracerebroven- tricular administration of 2 and 20 pg caused reduced motor activity and induced sedation (103).
The somnogenic action of PGDz was investigated in detail by using the continuous infusion technique into the third ventricle (152, 390). Infusions started 1 h before dark onset and continued throughout the first 9 h of the 12-h dark period. Excess sleep was induced by a dose as small as 60 fmol/min. The effect occurred within the 1st h of infusion and subsided within 2 h after the end of administration. Whereas at 60 fmol/min only NREM sleep was increased, higher doses (600 and 6,000 fmol/min) also markedly augmented

632 ALEXANDER A. BORBELY AND IRENE TOBLER Vot?um 69
REM sleep. Diurnal infusion of PGDz (6 pmol/min) did not affect sleep (153). In the rabbit, PGDz (250 pmol icv) increased NREM sleep with enhanced slow-wave activity and raised rectal temperature (194).
A continuous intracerebroventricular infusion of PGDZ in five rhesus monkeys (Macaca muZatta) during 6 h of the light period enhanced drowsi- ness, NREM sleep, and REM sleep (l29,275a, 392). On discontinuation of the infusion, sleep and behavior reverted rapidly to their normal level, and sleep in the following night did not differ from base line. The PGD2 infusion induced in three animals a dose-dependent increase of sleep, whereas in two animals a bell-shaped dose-response curve was observed (275a). For infusions into the lateral ventricle, the effective dose range was 0.015-2.25 nmol/min. In a monkey with two implanted cannulas, infusion into the third ventricle of as little as 0.15 pmol/min PGDz elevated sleep, whereas for infusion into the lateral ventricle the minimal somnogenic dose was 1,000 times higher. Sleep induced by PGD2 occurred episodically, the animals could be easily aroused, and body temperature and heart rate. were little altered. Infusion of PGE2 and PGFzcy into the lateral ventricle reduced NREM sleep and increased body temperature and heart rate. A stereoisomer of PGDz, 9-P-PGDz, had little effect on sleep (392).
Both PGE2 and PGFZa had a much weaker somnogenic action in the rat than PGD2 (103,390,391). Prostaglandin Ez (0.25 pmol to 2.5 nmol) microin- jetted into the preoptic area of partially restrained rats caused a dose-de- pendent reduction of sleep during the 900min postinjection period (229). Hy- perthermia was present only at higher doses. Whereas 50 pg/kg (141 nmol) PGEl administered intracisternally to the rat even suppressed sleep, 5 pg/kg (14 nmol) was ineffective (227). A dose of 16-64 pg/kg (45-181 nmol/kg icv) did not affect sleep in the cat (88).
It has been proposed that PGD2 may be a regulator of physiological sleep, since it is synthetized and me tabolized i n the brai n, it induces an apparently physiological sleep, and it is effective at concentrations that are comparable to the endogenous level in the brain (128a, 390). Further data that are consistent with this proposition are the hypothermic action of PGDB and the diurnal rhythm in PGD2-synthetase activity in rat brain (389). How- ever, a sleep-related hypothermia has not been demonstrated.
2. Vasoactive intestinal polypeptide
The somnogenic action of vasoactive intestinal polypeptide (VIP) in the rat was recognized during the screening of numerous neuropeptides (315). Intracerebroventricular administration (100 ng; 30 pmol) in the first part of the light period enhanced diurnal REM sleep. A dose of 1 pg (0.3 nmol) increased REM sleep after a delay of 24 h. Administration of 100 ng (30 pmol) icv at dark onset enhanced both NREM and REM sleep during the dark period, an observation that was confirmed by another group (269). An analy-

April 1989 ENDOGENOUS SLEEP-PROMOTING SUBSTANCES 633
sis of the time course revealed a prompt action of VIP: both NREM and REM sleep were enhanced within the 1st h after injections, and the latency to the first NREM sleep episode was reduced. Rapid-eye-movement sleep remained elevated for 3 h and NREM sleep was elevated for 6 h. Brain temperature was not affected. A slow release of VIP into the lateral ventricle (estimated release rate 100 rig/day; 30 pmol/day) enhanced NREM and REM sleep in both the light and the dark period (211).
In the cat, VIP (100 ng; 30 pmol icv) enhanced REM sleep (75). In contrast to the prompt response in the rat, a significant difference of the hourly cumulative REM sleep values was present only after 5 h.
Vasoactive intestinal polypeptide was able to restore sleep in animals that had been rendered insomniac by pharmacological pretreatment. In rats pretreated with either PCPA or chloramphenicol (CAP), the suppression of sleep could be antagonized by VIP (100 ng; 30 pmol icv) (315). Also in the cat, PCPA-induced insomnia could be partially antagonized by VIP (ZOO ng; 60 pmol icv) (79,295). However, this effect was not confirmed for a dose of 1,250 ng (376 pmol icv) in another study (354; D. Stenberg, personal communica- tion). Vasoactive intestinal polypeptide (100 ng; 30 pmol icv) did not restore sleep when insomnia in the cat had been induced by CAP (75).
Finally, in accordance with a putative role of VIP in the regulation of REM sleep, VIP antiserum administered intracerebroventricularly to rats increased waking and depressed REM sleep without affecting NREM sleep (315).
3. Growth hormone, growth hormone-releasing factor, and somatostatin
The rise in the plasma level of GH after sleep onset is one of the best documented sleep-related neurochemical events (for reviews see Refs. 5,230, 236). In humans, the rise in GH is associated with slow-wave sleep (stages 3 and 4) (366). Sleep-associated variations in GH have been reported also for the baboon (280), dog (364), and rat (182,245).
Systemic administration of GH enhanced REM sleep in humans (human GH, 5 U) (239), cat (bovine GH, 50 pg to 1 mg ip) (356), and rat (rat or bovine GH, 0.1-l mg/kg ip) (83,355). A dose of 3 mg ip enhanced waking in the cat but did not enhance REM sleep (356,357).
Diurnal administration of growth hormone-releasing factor (GRF) (50 pg; 9.9 nmol iv) to humans did not affect sleep or sleep-related GH secre- tion (111).
Intracerebroventricular administration of GRF (2 nmol) to rats reduced motor activity, increased NREM sleep (the effect on REM sleep was not reported), and enhanced EEG slow-wave activity during the 30- to 320-min recording period (89). Doses of 0.5 and 5 nmol had no significant somnogenic action. Also, in the rat, NREM and REM sleep were enhanced in postinjection hour 1 after intracerebroventricular administration of GRF (0.01-l nmol/kg)

634 ALEXANDER A. BORBELY AND IRENE TOBLER vobwrne 69
at dark onset (265,267). Brain temperature was not affected. The intracere- broventricular administration of GRF (SO-100 ng; 11-22 pmol) caused a dose-dependent behavioral sedation and an increase in EEG slow-wave activ- ity (263). The effect started within 5-10 min and lasted for 2-4 h. Body temperature was not affected. Similar effects were obtained after microin- jection into the hippocampus (100 ng; 22 pmol), whereas injection into the head of the caudate (75 ng; 16.5 pmol) had an activating effect. Vigilance states were not scored in these experiments.
Intracerebroventricular infusion of GRF (0.01-l nmol/kg) in the rabbit caused a dose-dependent increase in NREM sleep, REM sleep, and EEG slow-wave activity during the first 2 h postinfusion (267). The effect on NREM sleep was most prominent in the 1st h.
In a recent study, a Z-day intracerebroventricular infusion of somato- statin (20 pg/day; 12 nmol/day) in rats enhanced REM sleep without affect- ing NREM sleep (62). After withdrawal, REM sleep reverted to its control level. Intracerebroventricular infusion of cysteamine (40 or 90 pg/day), a somatostatin-depleting drug, caused a dose-dependent reduction of REM sleep without affecting NREM sleep. The effect was not more prominent after a 180 pg/day dose. Infusion of ethanolamine (90 pg/day), a structural analogue of cysteamine, was ineffective. Local infusion of somatostatin anti- serum into the nucleus tractus solitarius in the rat blocked the REM sleep- enhancing action of local carbachol injection (64).
4 hsulin and alterations in glucose metabolism
Infusion of insulin (Actrapid, 200 pIU/day icv for 3 days) enhanced NREM sleep in the rat without affecting REM sleep, while anti-insulin serum (iv or icv) caused a decrease in NREM sleep (65). Induction of diabetes mel- litus by streptozotocin caused a long-lasting reduction in NREM sleep and a transient reduction in REM sleep (61). Both intravenous (2 IU/day) and intracerebroventricular infusion (50-200 pIU/day) of insulin restored NREM sleep. The latter manipulation showed a dose-dependent effect without low- ering the blood glucose level. The somnogenic action of insulin could be mediated by some as yet unspecified effect on brain metabolism or, alterna- tively, could be caused by a direct effect on neurons (see Ref. 63).
Administration of Z-deoxy-D-glucose (500 and ‘750 mg/kg ip), an inhibi- tor of glucose catabolism, to rats caused a dose-dependent/increase in NREM sleep and a suppression of REM sleep (276).
Sleep-related changes in brain glucose metabolism have been reported. The brain glycogen level in a small number of rats was rapidly increased during sleep (179). Animals having slept continuously for 5-11 min showed a 70% higher level than animals that had been awake. After sleep termination, glycogen returned to the waking level within 2-5 min. Glucose utilization in the choroid plexus of the cat was higher during NREM sleep than during

April 1989 ENDOGENOUS SLEEP-PROMOTING SUBSTANCES 635
waking (22). No significant changes were observed in other brain structures (288). The rate of glucose utilization was lower during sleep than during waking in various brain regions of the rhesus monkey (Macaca mulatta) (262). Taken together, the data are consistent with a close tween NREM sleep regulation and glucose metabolism.
relationship be-
In humans, measurements of regional cerebral glucose uptake by posi- tron emission tomography (PET) showed reduced values during slow-wave sleep in comparison to relaxed wakefulness or REM sleep (104). Metabolic rates measured in the left caudate during waking were reduced after a night of sleep deprivation (421).
5. Arginine vasotocin
Although the presence of a vasotocin-like peptide in the mammalian pineal gland has been advocated for a number of years, its structure still awaits conclusive identification (see Ref. 294). A striking seasonal variation of arginine vasotocin (AVT) immunoreactivity in rat pineal glands has been described (294).
The administration of minute quantities of AVT (10v21 mol; 10-l* g) into the third ventricle of cats induced NREM sleep within 5 min (285). The response lasted for 30 min and could be elicited also by intraperitoneal in- jection of 10B6 g (1 nmol). Rapid-eye-movement sleep was completely sup- pressed throughout the 5-h recording period. In a later study, the much higher intracerebroventricular dose of 1 pg (1 nmol) increased the percentage of NREM sleep by 50% and suppressed REM sleep completely in the 3-h recording period (118). Lesioning of the nucleus raphe dorsalis prevented the effect of AVT during the first 5 postoperative days, whereas lesioning the nucleus raphe pontis did not alter the action of AVT. An increase of REM sleep and the occurrence of REM sleep episodes at sleep onset was observed after the intracerebroventricular injection of an AVT antiserum (282).
The effect of AVT on sleep showed marked species differences. In the rat, a dose of 0.5 pg/kg (0.5 nmol/kg ip) administered after light onset was ineffective, whereas a dose of 50 ,ug/kg (50 nmol/kg) reduced REM sleep latency without affecting sleep latency, total sleep time, or the amount of REM sleep in the first 2 h after injection (232). A dose of 1 pg/kg (1 nmol/kg ip) injected in the first part of the light period was ineffective throughout the 8-h recording period (313). A dose-dependent reduction of total sleep time and REM sleep was observed after 10 and 50 pg/kg (10 and 50 nmol/kg). In another study, the injection of AVT (10-12-10-16 g; 10-15-10-1g mol) into the third ventricle in the light period did not alter total sleep time, NREM sleep, or REM sleep (373). However, a dose of lo-l4 g (lo-l7 mol) reduced slow-wave activity within NREM sleep.
In the rabbit, the large intracerebroventricular dose of 25 pg (25 nmol) induced an increase of NREM sleep within 5 min that lasted up to 1 h (195).

636 ALEXANDER A. BORBELY AND IRENE TOBLER Volume 69
Subcutaneous injection of AVT (2 lug; 2 nmol) to human subjects caused an increase in REM sleep but had no other effects on sleep (54). A dramatic enhancement of REM sleep was reported after intranasal administration of 100 rig/kg (100 pmol/kg) to prepubertal boys (283). The effect of AVT was antagonized by intranasal valproic acid (117). The same group observed a REM sleep-dependent rise of AVT in lumbar CSF of healthy volunteers (284) and narcoleptic patients (293). However, as has been mentioned, there are problems with respect to an unequivocal identification of AVT.
The effect of iontophoretically applied AVT on the activity of single neurons was recorded in the nucleus reticularis gigantocellularis of the brain stem in rats and rabbits (264). The responses were predominantly excitatory and showed a long time course and a rapid desensitization.
In conclusion, the somnogenic effect of AVT in animals and humans awaits confirmation. In view of the potent endocrinological actions of this substance, indirect effects on sleep should be considered. The status of AVT as an endogenous substance is controversial (see Ref. 294).
6. Pro-opiomelanocortin-derived peptides
Effects of pro-opiomelanocortin-derived peptides on sleep have been re- cently reviewed (51,210). Of the various peptides tested in the rat two com- pounds were somnogenic. When injected before dark onset, desacetyl-a-me- lanocyte-stimulating hormone (des-a-MSH) (1 ng ip; ~4 rig/kg; 2.4 pmol/kg) enhanced total sleep time and NREM sleep in the 12-h dark period (50). Doses of 0.1, 10, or 100 ng (0.06, 6.1, or 61 pmol, respectively) were ineffective. Corticotropin-like intermediate lobe peptide (CLIP; ACTH-(18-39) (10 ng; 4.1 pmol ip) did not affect total sleep time but enhanced REM sleep during two 12-h dark periods. Both the number and duration of REM sleep episodes were increased. Doses of 1, 20, and 100 ng (0.41, 8.1, and 40.6 pmol, respec- tively) were ineffective. The depression of sleep induced by apomorphine (500 pg/kg ip) was antagonized by CLIP (10 ng; 4.1 pmol ip) (51). No significant changes of sleep were observed after administration of ACTH (l-100 ng; 0.41-40.6 pmol ip) and a-endorphin (l-100 ng; 0.57-57.3 pmol ip) (50). A lack of a somnogenic action was reported also for P-endorphin (1 pg; 0.29 nmol icv) (314).
The effect of 72-h REM sleep deprivation on the level of endogenous opioid peptides was investigated in the rat (297). Groups of rats were de- prived by either the platform or the pendulum technique, and for each method a control group was used (i.e., exposure to a larger platform or to a pendulum not inducing postural imbalance). Rapid-eye-movement sleep de- privation reduced P-endorphin in the hypothalamus and increased its level in the plasma. However, since in both control groups the plasma level was higher than under home cage condition, a part of the change must be attrib- uted to a nonspecific treatment effect.

April 1989 ENDOGENOUS SLEEP-PROMOTING SUBSTANCES 637
7. Cholecgstokinin octapeptide
The effect of cholecystokinin octapeptide (CCK-8) on sleep was investi- gated in the rat (317). Doses of 10 and 20 pg/kg (8.7 and 17.4 nmol/kg, respectively) administered intraperitoneally in the light period reduced the latency to the first NREM sleep episode but did not affect sleep. Doses of 1 pg/kg (0.87 nmol/kg) (317) or 2 pg/kg (1.74 nmol/kg) intraperitoneal as well as the intracerebroventricular administration of 100 ng and 1 pg (87 pmol and 0.87 nmol, respectively) (314) were ineffective. In a recent study, CCK-8 (10 or 50 pg/kg; 8.7 or 43.7 nmol/kg ip) administered at dark onset caused a dose-dependent increase in NREM sleep and EEG slow-wave activity and a reduction in motor activity, food intake, and brain temperature (173). Rapid- eye-movement sleep was not affected. The changes induced by CCK-8 were limited to the 1st h postinjection, and no significant effects were observed during the remainder of the dark period and during the subsequent light period. A dose of 4 pg/kg (3.48 nmol/kg) had no somnogenic action. In an- other study, continuous intracerebroventricular infusion of CCK-8 (1.1 ng/ min; 1 pmol/min for 5 days) induced an increase in the frequency of REM sleep episodes in the two 3.5-h recording periods on days 4 and 5 (70). In cats rendered insomniac by pretreatment with PCPA, 100 ng (87 pmol) CCK-8 intracerebroventricular restored REM sleep (79,296).
8. Other peptides
No effect on total sleep time was reported for Met-enkephalin (1 lug; 1.7 nmol icv), Leu-enkephalin (1 ,ug; 1.8 nmol icv), neurotensin (300 ng and 2 pg; 179 pmol and 1.2 nmol icv), angiotensin II (l-100 ng; 0.96-95.6 pmol icv), renin (0.01 U icv), and substance P (0.1 and 1 /-cg; 74.2 pmol and 0.74 nmol icv) (314). Some of the peptides reduced REM sleep (angiotensin II: 100 ng, 95.6 pmol; renin: 0.01 U; substance P: 100 ng, 74.2 pmol). Neuropeptide Y (1.25 nmol icv) enhanced EEG synchronization in the rat (106,427).
9. Serotonin
The increasing recognition of the role of 5-HT and other monoamines in sleep regulation in the 1960s culminated in the “monoamine theory” of sleep (162, 163). However, recent data have proved to be incompatible with the theory. They include the observation that both the in vivo release of indole- amines and the firing rate of serotonergic neurons is higher during waking than during sleep (47, 49, 299). Moreover, the intraventricular or systemic administration of 5-hydroxytryptophan (5-HTP) in the PCPA-pretreated cat restores sleep only after a relatively long delay (287, 324). Accordingly, Jou- vet (164-166) proposed that 5-HT may promote in the course of the waking period the synthesis of one or several “sleep factors” that are stored until they trigger sleep. The role of 5-HT in sleep has been recently reviewed (99).

638 ALEXANDER A. BORBfiLY AND IRENE TOBLER votwne 69
In the rat, the forebrain 5-HT level was increased during recovery from 24-h sleep deprivation (37). The time course was similar to that of excess deep NREM and REM sleep. Interestingly, the raised 5-HT level was present only from the second hour of recovery sleep onward. The 5-HT content in the cortex and midbrain was higher when immobilized rats were killed by mi- crowave irradiation during sleep than during waking (272). A close associa- tion between 5-HT /level and sleep is suggested also by another study. When rats were allowed to sleep for 90 min after a 96-h REM sleep-deprivation period, the 5-HT level in the cortex and diencephalon was higher than in a group maintained on the deprivation schedule (130). However, a normal level of 5-HT is not a prerequisite for sleep regulation. Thus when 5-HT depletion by PCPA was combined with sleep deprivation, an increase of EEG slow- wave activity was still present during recovery sleep (374).
Not only the level of 5-HT but also the number of specific 5-HT binding sites in rat brain exhibit a prominent 24-h rhythm (413). The latter rhythm persisted unchanged after 24-h sleep deprivation (414,415). This observation extends the number of neurotransmitters, the rhythmic variations of which are unaffected by extended waking (420; see Ref. 419 for a review).
Radioligand binding studies have revealed four distinct 5-HT binding sites in rat brain that may function as 5-HT receptors (56). Studies with the specific 5-HTz antagonist ritanserin indicate that the 5-HT2 receptor may be implicated in the regulation of slow-wave sleep. An oral dose of 10 mg ad- ministered to human volunteers either in the morning or in the evening induced a marked increase in slow-wave sleep (stages 3 and 4) and a reduc- tion of REM sleep (after the evening dose) in the subsequent night (144). Similarly, after treatment with ritanserin (an oral dose of 5 mg in the morn- ing for 7 days), human subjects exhibited a marked increase in slow-wave sleep, whereas waking and REM sleep tended to decrease (69). A morning dose of ritanserin (10 mg) administered over 2 wk caused a persistent rise in slow-wave sleep (143). In the rat, ritanserin (0.63-2.5 mg/kg ip) enhanced deep NREM sleep and EEG slow-wave activity and suppressed REM sleep (40a, 87).
Tryptophan, the precursor of 5=HT, is being advocated as a natural sleep remedy. However, in view of the short-lasting acute effect, the high dose required for sedation (3 g and more), the insufficient documentation of a long-term hypnotic action, and the risk of toxicity, its clinical use is question- able (41). Other authors view the merits of this substance more positively (340). Irrespective of its therapeutic usefulness, there is little evidence that tryptophan promotes sleep by physiological mechanisms, since various me- tabolites other than 5-HT references see Ref. 41).
10. Adenosine
Administration of
are known to affect the central nervous system (for
adenosine (1, 10, or 100 nmol) into the lateral ventricle sleep during the 6-h postinjection in the light period of rats increased total

April 1989 ENDOGENOUS SLEEP-PROMOTING SUBSTANCES 639
(306). Deep NREM sleep was enhanced by the 1 and 100 nmol dose. Although the amount of REM sleep was not significantly changed, REM sleep latency was shortened. No dose-response relationship was present in the l- to lOO- nmol range. This result confirms earlier observations that intracerebroven- tricular administration of adenosine (190 nmol) causes sleep in dogs (128). Three metabolically stable adenosine analogues enhanced deep NREM and REM sleep after intraperitoneal administration to rats in the beginning of the light period (303, 305). A somnogenic effect was reported also for N6-l- methylcyclopentyladenosine, a specific adenosine Al-receptor agonist (301). Administration of deoxycoformycin, an adenosine deaminase inhibitor, en- hanced REM sleep and at a higher dose enhanced deep NREM sleep (304, 306). However, another adenosine deaminase inhibitor reduced sleep (237). The adenosine precursor S-adenosylhomocysteine enhanced at certain doses NREM, deep NREM, and/or REM sleep in the rat, cat, and rabbit (101,326). Taken together, the results indicate that increasing the brain level of adeno- sine or stimulating the adenosine receptor by an agonist, enhances deep NREM and REM sleep. It has been suggested that these effects are mediated by Al-receptors that are activated by nanomolar quantities of adenosine or its analogues, whereas micromolar quantities are required for AZ-receptor stimulation (305, 306). In accordance with this proposition, the somnogenic action was observed with nanomolar doses and diminished when the doses reached the micromolar range.
The effect of REM sleep deprivation by the platform method on adeno- sine receptor binding in the brain was determined by means of Al-specific ligands (424). Rats exposed to a large platform or maintained in the home cage served as controls. Deprivation for 48 or 96 h increased the number of specific binding sites in the cortex and striatum. The brain concentration of adenosine was not altered after 24 or 48 h REM sleep deprivation in contrast to an earlier study where an increase had been found (128).
Il. Acet ylcholine
There is a large literature concerning the role of the cholinergic system in sleep regulation (for reviews see Refs. 134,346). Recent studies confirmed that microinjection or microinfusion of cholinergic agonists into the pontine reticular formation of the cat (16, 347) and rat (116, 345) induce a dose-de- pendent and site-dependent increase of a REM sleeplike state. The effect could be blocked by muscarinic antagonists. At certain injection sites only some elements of REM sleep (i.e., muscle atonia, pontogeniculooccipital waves, rapid eye movements, and cortical desychronization) were elicited. In humans the intravenous infusion of a cholinergic agonist after sleep onset reduced the latency to the subsequent REM sleep episode, whereas the mus- carinic antagonist scopolamine had the opposite effect (for references see Ref. 346).
The relationship of cholinergic mechanisms to REM sleep-promoting factors in the CSF have not yet been investigated. Preliminary data in rat

640 ALEXANDER A. BORBELY .AND IRENE TOBLER Vohme 69
(86) and cat (168) indicate that these factors originate in the hypothalamus and pituitary rather than in the pons, one of the central structures of REM sleep regulation.
12. Catecholamines
The role of catecholamines in the control of waking and REM sleep has been extensively discussed in the framework of the “monoamine theory” (162163,166) and has been recently reviewed (108,406). Several studies have focused on changes in receptor sites after REM sleep deprivation. An initial report of a reduction in the number of P-receptors in rat cortex after 72 h but not after 24 or 48 h of REM sleep deprivation (246) has not been confirmed (1, 302). A similar inconsistent picture prevails for the number of dopamine- binding sites where an increase in the second part of a 24-h total sleep-de- privation period (420), no change after 96-h REM sleep deprivation (92), and a decrease after 48 and 96 h REM sleep deprivation (430) were reported. rH]- clonidine binding in rat cortex was not changed after 24 or 48 h REM sleep deprivation (247). In some studies, control rats maintained on a larger plat- form exhibited similar yet smaller changes than the experimental group on a small platform (246, 430). Thus, in addition to REM sleep deprivation, vari- ables related to the experimental procedure seem to play a role.
There is evidence that stimulation of dopamine De-receptors promotes sedation and EEG synchronization in rats (112). The relationship between catecholamines and putative endogenous sleep factors has not been investi- gated. It should be noted though that in the assay of REM sleep-promoting factors, the animals were pretreated with REM sleep-suppressing drugs that act via the catecholaminergic system (see sect. III@.
13. Melatonin
Because the effects of melatonin on sleep have been recently reviewed (29), only a short summary is provided here.
Melatonin has been reported to enhance sleep in various animal species (cat, rat, young chicken) (e.g., see Refs. 133,225) as well as in humans (e.g., see Refs. 10, 404). Whereas in the rat a dose-related somnogenic effect was ob- served (135), a sleep-reducing action was found with a low dose (231). In humans a wide dose range (1.7 mg to 1 g; 7.3 pmol to 4.3 mmol/day) was studied. The effects include a reduction of sleep latency after an intravenous bedtime dose (50 mg) (57,58) and the enhancement of sleepiness after three daytime oral doses of 80 mg (218). However, since even a small oral dose (2 mg) raises plasma melatonin to a level that exceeds the endogenous night- time level by a factor of 10 to 100 (13), it is doubtful whether a physiological effect was induced by these studies. A 1 or 5 mg oral dose administered before bedtime did not affect sleep except for a prolongation of REM sleep latency after the larger dose (159).

April 1989 ENDOGENOUS SLEEP-PROMOTING SUBSTANCES 641
In humans as in other mammals, melatonin secretion is influenced by a, circadian pacemaker. The maximum occurs during the dark period regard- less of whether this constitutes the phase of sleep or waking. Human studies have shown that neither darkness nor sleep are prerequisites for a high level at night (7,160). Phase shifting the sleep/wake cycle or maintaining subjects under temporal isolation induced a dissociation between the circadian rhythms of melatonin and sleep (222,412).
Although it is unlikely that melatonin is directly involved in sleep regu- lation, it may affect sleep indirectly by altering the phase of the circadian pacemaker. Thus the daily administration of 2 mg melatonin in the late afternoon induced unusual tiredness in the early evening after 4-5 days, an observation that is compatible with a gradual phase advance of the circadian timing of sleep (13,14). A recent study indicated that melatonin may allevi- ate jet lag (11) and synchronize a disturbed sleep/waking cycle (12).
14. Adrenal steroid metabolites
The deoxycorticosterone metabolite 3&&etrahydrodeoxycorticoster- one, which is a brain constituent, showed a dose-dependent somnogenic ac- tion in the rat (238). Whereas a dose of 5 mg/kg ip administered in the light period had a marginal effect, 10 mg/kg reduced sleep latency and increased NREM sleep in the 2-h recording period. 3cu,ScY-Tetrahydrodeoxycorticoster- one appears to be a potent ligand of the benzodiazepine receptor complex.
15. Calcium
In early experiments, 0.1 mg CaC12 was injected in a volume of 20 ~1 into the diencephalon of rats that had been anesthetized by ether (53). After emergence from anesthesia, a sleeplike state was observed for 30-90 min. The state was characterized by closed eyelids, muscular hypotonia, and reduced respiratory rate. The animals could be briefly aroused by sensory stimuli. Based on experiments in 25 animals, the infundibular region was defined as the sensitive injection site. Higher doses induced anesthesia and occasionally respiratory arrest. The authors raised the possibility that calcium may be involved in the regulation of physiological sleep.
In the cat, a sleeplike state with cortical EEG slow-wave activity was induced by push-pull perfusion of distinct sites in the posterior hypothala- mus with excess calcium (400). Drowsiness became apparent within 5-15 min after the start of perfusion.
In the rat, sleep and behavior were recorded during a l-h infusion of artificial CSF into the lateral ventricle (383). When the infusate contained a physiological concentration of calcium, REM sleep tended to be reduced. Excess calcium in the perfusate (2-3 times the regular level) either antago- nized the REM sleep suppression or even enhanced REM sleep. Total sleep time was not changed. Still higher doses of calcium elicited feeding episodes

642 ALEXANDER A. BORB&LY AND IRENE TOBLER vot?ume 69
during which the cortical EEG exhibited a synchronized pattern. The authors suggested that the cortical synchronization may reflect a disinhibition of hypothalamic mechanisms from cortical control.
16. Endogenous organic bromine cornpound
An organic bromine compound was detected in human CSF (422) and identified as 1-methylheptyl-y-bromoacetate (y-Br) (423). Because a similar substance was found in cat CSF, the effect of r-Br on sleep was investigated in the encephale isole cat (381). Intravenous administration of 0.1 mg/kg enhanced REM sleep within 5-15 min. There is some evidence for a 24-h variation of the r-Br level in human and cat plasma and in cat CSF (274). In the cat, 72-h REM sleep deprivation elevated the r-Br level in plasma and CSF, whereas in humans there was no increase during total sleep depriva- tion. Butoctamide hydrogen succinate, a structurally related, bromine-free compound, was reported to enhance REM sleep in the cat (380) and in humans (275) and to slightly enhance human slow-wave activity (273).
F. Immunologically Active Polypeptides
1. Interleukin I
Interleukin 1 (IL-l) represents a family of polypeptides that are pro- duced by activated macrophages and other cell types and that mediate a wide range of biological activities. The genes coding for human IL-l have been recently cloned (223). There is evidence for the presence of IL-l immunore- activity (42) and IL-l mRNA (94) in cerebral neurons. The IL-l purified from human or rabbit mononuclear cells enhanced NREM sleep and EEG slow- wave activity when infused into the lateral ventricles of the rabbit (208,405). The response was dose dependent: after a low dose, excess NREM sleep was observed for 3 h, and after a higher dose it was observed throughout the 6-h recording period. After large doses the effect was observed in some cases for 18 h. In contrast to the delayed action of muramyl peptides, the increase of NREM sleep after IL-l occurred already in the 1st h. Administration of IL-l reduced REM sleep in comparison to noninfused controls but not in compari- son to rabbits administered heat-treated IL-l (405). These results document the REM sleep-inhibiting action of the intracerebroventricular infusion pro- cedure per se. Interleukin-1 exhibited its somnogenic action also after intra- venous administration (208, 349). Sleep induced by IL-l remained episodic, and the animals could be easily aroused. Human recombinant IL-lp (50 ng to 5 pg icv, 10 rig/kg iv) (349) had similar effects on sleep in the rabbit as native IL-l.
When astrocyte-derived IL-l was infused into the lateral ventricle of rats at the onset of the light period, EEG slow-wave activity was enhanced but total sleep time was not affected (377). The effect was most prominent in

April 1989 ENDOGENOUS SLEEP-PROMOTING SUBSTANCES 643
the second 2 h of the 4-h recording period (Tobler and Borbely, unpublished observations). Rapid-eye-movement sleep was significantly lower than in a group receiving control infusions (column fractions devoid of IL-l activity or pyrogen-free saline). However, also in this species, the infusion per se re- duced REM sleep.
In both rabbit and rat, IL-l elicited hyperthermia. This response could be prevented in the rabbit by the simultaneous intracerebroventricular infu- sion of anisomycin (‘700 pg) without abolishing the somnogenic effect of IL-l (ZOO). Although IL-l elevated brain temperature, it did not abolish the typi- cal temperature changes occurring at the transition from one vigilance state to another (405). The results indicate that the effect of IL-l on sleep is not a secondary effect of hyperthermia. This interpretation is supported by the observation in the rat that the increase of slow-wave activity was not corre- lated with the enhancement of body temperature (377).
Plasma IL-l activity was assayed during waking and sleep in six healthy young human subjects (248). Slow-wave sleep shortly after sleep onset was \ associated with a peak in the plasma IL-l level. Because the.assay system was sensitive to both IL-l and IL-Z activity, it would be important to confirm this interesting observation by a more specific assay. In a preliminary report, IL-l-like activity in CSF of three cats was higher during sleep than during waking (221).
2. Interferon
Human recombinant leukocyte interferon (IFN)-ar2 administered intra- venously or intracerebroventricularly to rabbits enhanced NREM sleep and EEG slow-wave activity (197). The effect became manifest in hour 2 after intracerebroventricular infusion and persisted throughout the 6-h recording period. The increase of NREM sleep was dose dependent. Intravenous injec- tion, but not intracerebroventricular infusion, reduced REM sleep during the first 3 h in comparison to controls treated with saline or artificial CSF. Sleep induced by IFN remained episodic, brain temperature showed the normal variations at state transitions, and the animals could be easily aroused. Interferon caused hyperthermia. In contrast to lipopolysaccharide (LPS) and lipid A, the hyperthermic and somnogenic effects of IFN could not be blocked by preincubation with polymyxin B.
Recombinant human IFN-aA was administered intravenously to pigtail monkeys (Macaca nemestrina) in the morning, and sleep was recorded during the following two nights (309). In both nights, REM sleep latency was shorter compared with a base-line night before IFN administration. The sleep dis- tribution has not been reported in detail.
3. Tumor necrosis factor
Human recombinant tumor necrosis factor (TNF) enhanced NREM sleep and EEG slow-wave activity in the rabbit (349). After intracerebroventricu-

644 ALEXANDER A. BORBfiLY AND IRENE TOBLER voi?ume 69
lar administration of a 5-pg dose, the effect was observed throughout the 6-h recording period, whereas a 0.5pg dose elicited changes only in the 1st h postinfusion. After intravenous administration, the increase of NREM sleep was significant only in the postinjection hours 2-6. Rapid-eye-movement sleep was inhibited after either route of administration. The animals still exhibited a normal sleep posture, episodic sleep, and the typical changes in brain temperature at transitions of vigilance states. Tumor necrosis factor elicited biphasic hyperthermia.
4. Lipopolysaccharide and lipid A
Lipopolysaccharide (endotoxin), a bacterial cell wall component, and its lipid A moiety enhanced NREM sleep and EEG slow-wave activity in the rabbit (ZOO). The time course of the response and the effect on REM sleep depended on the route of administration. Thus intravenous injection of LPS or lipid A augmented NREM sleep within 1 h, whereas after intracerebro- ventricular infusion of lipid A, excess NREM sleep appeared only after 3-4 h. Rapid-eye-movement sleep was inhibited after intravenous administration of lipid A but not after intracerebroventricular administration. Regardless of the route of administration, sleep remained episodic, brain temperature showed the normal variations at sleep-state transitions, and the rabbits could be easily aroused. Both LPS and lipid A caused hyperthermia. The hyperthermic as well as the somnogenic action of LPS could be blocked by preincubation with polymyxin B.
5. Possible relations between immunologically active substances
Krueger and co-workers (196-198,207,210,349) htave recently described a number of interactions between various endogenous sleep-promoting sub- stances and proposed that sleep may be involved in immunomodulatory pro- cesses. They emphasized the immune-related activity of many putative sleep-promoting substances (VIP, prostaglandins). Moreover, they suggested that sleep may play an active role in recuperative processes from damage induced by infectious disease. A cascade of events are envisaged, which are briefly discussed below. For the detailed arguments the reader is referred to the original papers.
Interleukin 1 is known to be produced in response to muramyl peptides, LPWlipid A, and TNF. The delayed increase of NREM sleep induced by muramyl peptides and the rapid effect of IL-1 indicate that muramyl pep- tides may influence sleep via IL-l. On the other hand, the observation that the time course of somnogenic action of either LPS or TNF differs from that of IL-l suggests partly independent mechanisms. Muramyl peptides and all the other immunologically active compounds discussed in this section can alter arachidonic acid metabolism and thereby possibly influence the synthe-

April 1989 ENDOGENOUS SLEEP-PROMOTING SUBSTANCES 645
sis of PGDz (see sect. IIIEI). A further link between various classes of sleep- promoting agents is suggested by reports that prostaglandins release VIP from the hypothalamus and that LPS raises the VIP plasma level. Finally, it has been shown that muramyl peptides may compete for 5-HT-binding sites on macrophages.
IV.
A .
DISCUSSION AND CONCLUSIONS
Criteria for an Endogenous Sleep Factor or a Sleep Substance
Different criteria have been proposed for defining an endogenous sleep factor or sleep substance (39,166,201,240). We propose the two basic criteria shown in Table 2. The first criterion is fulfilled by various compounds, whe reas the second criterion has not yet been rigorou any of the putative sleep factors or sleep subs tances.
sly shown to be met by
B. SleepPmmoting Action
I. Physiological sleep
An endogenous sleep-promoting substance should induce a physiological type of sleep. Physiological sleep can be defined by behavioral, electrogra- phic, and physiological criteria (368). The behavioral signs of sleep include a typical body posture, reduced responsivity to environmental stimuli, a rapid state reversibility, and a species-specific alternation between sleep and wak- ing episodes. The electrographic criteria encompass the typical EEG, electrooculogram, and electromyogram changes of NREM and REM sleep. Computer-aided methods of EEG analysis may serve to quantitatively define the sleep-related variations. Moreover, NREM and REM sleep should show cyclic alternations. The physiological criteria include typical variations of
TABLE 2. Basic criteria for an endogenous sleep factor or sleep substance
Promotion of physiological sleep Sleep-promoting action consists of (at least one of the following must apply)
l Sleep induction l Sleep maintenance l Increase of electroencephalogram slow-wave activity l Increase in amount ‘of rapid-eye-movement sleep
Sleep-associated change in the organism Variation of sleep factor/substance in organism in association with sleeping and waking
l Brain concentration l Turnover l Other functionally relevant parameters

646 ALEXANDER A. BORBfiLY AND IRENE TOBLER vohmae 69
body temperature, breathing, and heart rate, which are associated with the vigilance states (266).
In most reports it has not been explicitly stated that sleep induced by endogenous sleep-promoting substances fulfills the various criteria for phys- iological sleep. Notable exceptions are factor S (sect. IIIAZ), AMTP (sect. III/k?), and other immunologically active polypeptides (sect. IIIF) for which signs of physiological sleep, such as a normal sleep posture, rapid state reversibility, and the cyclic alternation of NREM and REM sleep, were re- ported. Moreover, despite the hyperthermic action of those compounds, the typical state-associated changes in brain temperature were still present. It would be important to apply refined methods of EEG analysis to demon- strate that the changes induced by the various compounds correspond to the physiological changes.
The induction of a nonphysiological type of sleep cannot be regarded as conclusive evidence that a compound is not involved in physiological sleep regulation. Sleep may be altered by an excessive dose or by a nonphysiologi- cal route of administration. For example, a somnogenic component of urinary factor S showed toxic effects in rabbits only when administered at a high dose (199). In addition, it is possible that endogenous compounds are not globally involved in sleep regulation but may mediate the manifestation of a specific substate of sleep. The enhancement of slow-wave activity in NREM sleep and the concomitant depression of REM sleep by AMTP and other immunologically active peptides may be interpreted in these terms (see sect. 1vA8).
2. Reliability
Ideally, the somnogenic action of a compound should be demonstrated in different laboratories for a specified dose range and in different species of animals. Because this has rarely been the case, it is often difficult to assess the reliability of the sleep-promoting action.
For factor S, the muramyl peptide AMTP, as well as the immunologi- cally active peptides, a somnogenic effect was reported in most studies. The substances were generally administered via the intracerebroventricular route, and the rabbit was usually the experimental animal. However, a som- nogenic effect (MDP) was present also in the rat, cat, and squirrel monkey. The SPS was somnogenic after systemic an’d intracerebroventricular admin- istration in the rat and mouse. However, this factor as well as its constitu- ents SPS-B and uridine were effective only during the nocturnal period. &Sleep-inducing peptide constitutes one of the most extensively studied pu- tative sleep substances for which a somnogenic effect was not consistently observed (Table 1). A similar inconsistent picture emerges for AVT. A sleep- promoting effect has been reported for melatonin in four species (humans, cats, rats, and young chickens), for PGD2 in three species
for VIP in two species (cats and rats). (rats, rabbits, and
monkeys), and

April 1989 ENDOGENOUS SLEEP-PROMOTING SUBSTANCES 647
3. Onset and duration of action
Various compounds induce sleep only after a delay. Thus after intrace- rebroventricular administration of factor S derived from CSF, brain, or urine (195) or of IFN-a2 to rabbits (197), the increase in sleep became manifest during the 2nd h postinfusion. A delayed onset of action was observed also for the REM sleep-promoting factor administered to the pretreated cat or rat (sect. IIBI) as well as for DSIP (Table 1). These observations may reflect a slow diffusion or transport from the ventricle to the active site in the brain or may indicate that the compound induces sleep indirectly via other biochemi- cal processes. The short sleep latency after microinjection of factor S; PGDz, or DSIP into active brain sites (sect. 1vB4) suggests the former possibility. On the other hand, there are other substances [e.g., GRF (263) or AVT (285)] that enhanced sleep within minutes even after intracerebroventricular ad- ministration.
Substances that were derived from approaches based on total or selec- tive sleep deprivation have almost invariably a long duration of action. Thus a somnogenic effect lasting for hours has been reported for factor S, the muramyl peptides, the immunologically active polypeptides, SPS, and the REM sleep-promoting factor. Changes lasting as long as 18-20 h were ob- served after intraperitoneal injection of SPS in the mouse (261) or after intracerebroventricular administration of IL-1 in the rabbit (208). A short sleep-inducing action was described in this class of compounds only for uri- dine [intraperitoneal administration in the rat (139)] and for the REM sleep- promoting factor [intraperitoneal administration to nonpretreated rats (416)]. A sleep-promoting action extending over several hours has been re- ported also for DSIP, PGD2, VIP, GRF, and adenosine. An effect lasting 11 h was described only for AVT (285) and CCK-8 (173).
Prolonged rebound effects of EEG slow-wave activity in NREM sleep and of REM sleep time during recovery from sleep deprivation are well documented (sect. IIC). The prolonged somnogenic effect of substances de- rived from sleep-deprived animals is therefore consistent with the assump- tion that these compounds mediate sleep homeostasis. Alternatively, the long-lasting effect may be due to excessive, nonphysiological doses or to mechanisms that are unrelated to physiological sleep regulation. These problems can be further elucidated once the sleep-associated substances in th .e brain have been specified (sect. WC).
changes of sleep
4 Efective smnogenic dose and dose-response relationship
An endogenous sleep-promoting substance should exert its effect at a concentration that prevails under physiological conditions at the active site in the brain. This requirement cannot be adequately assessed, since for most compoun action is
.ds neith .er the endogenous cerebral concentration nor the site of known. ‘Ne vertheless, it is interesting that for the following sub-

648 ALEXANDER A. BORBiilLY AND IRENE TOBLER Vokmze 69
stances the somnogenic intracerebroventricular dose lies in the lower pico- molar range: AMTP (muramyl peptide), recombinant IL-l, uridine, PGD2, VIP, and GRF. A dose in the low nanomolar range is required for a sleep- promoting action of DSIP and adenosine. The unusually high potency of AVT (10 -” mol) needs confirmation.
A dose-response relationship has been reported for the following puta- tive sleep factors or sleep substances: AMTP, SPS, PGDz, GRF, insulin, and CCK-8. On the other hand, the somnogenic action of some compounds seems to lack a clear dose dependence (e.g., adenosine) or exhibits a bell-shaped dose-response curve (DSIP, uridine, GH, des-a-MSH, CLIP). It has been pointed out that the latter is a typical property of peptides (180).
5. Site of actio?a
The site of action of somnogenic compounds has been rarely investi- gated. Microinjections of urinary factor S into the brain were somnogenic at 8 of 52 sites (110). These extended from the basal forebrain to the mesodien- cephalic junction. The injection of PGD2 into the preoptic area had a somno- genie effect, whereas injections into the posterior hypothalamus were inef- fective (391). In both these studies, the responsive sites were located in the diencephalon. &Sleep-inducing peptide was somnogenic when injected into the median thalamus, the periaquaeductal grey, and the nucleus tractus solitarii (367). However, the results were not presented in any detail, and ineffective sites were not described.
6. Species specificity
Are endogenous sleep-promoting substances largely species invariant or species specific? The former seems to be the case for factor S that was obtained from brain, CSF, and urine of several donor species (i.e., goats, rabbits, cattle, and humans) and had comparable somnogenic effects in var- ious recipient species (i.e., rabbits, rats, and cats). The species specificity of SPS is unknown, since it has been derived exclusively from rat brain and tested so far only in two rodent species. The REM sleep-promoting factor was obtained from cat and rat CSF and tested only within the same species. &Sleep-inducing peptide has been studied in six mammalian species, includ- ing humans. Whereas positive effects have been reported in all of them, negative results were also obtained in four species. The results within species are so inconsistent that species specificity cannot be assessed. Among the neuromodulators, PGD2 was effective in rats and monkeys; VIP in rats and cats; growth hormone in rats, cats, and humans; GRF in rats and rabbits; AVT in cats, rabbits, and humans; melatonin in cats, rats, and humans; and calcium in cats and rats. It should be noted in this context that PGDz is a major arachidonic acid metabolite in the rat but not in some other species.

April 1989 ENDOGENOUS SLEEP-PROMOTING SUBSTANCES 649
Prominent species differences were reported for AVT, which enhanced NREM sleep in animals and REM sleep in humans. However, these results require confirmation. Taken together, the data suggest a considerable degree of species invariance for endogenous sleep factors or sleep-promoting sub- stances.
By definition, any endogenous sleep factor or sleep substance promotes sleep. However, based on physiological considerations (sect. II), two classes can be discriminated. The first class favors sleep induction and sleep mainte- nance by exerting a permissive effect on sleep. Various exogenous factors (e.g., ambient temperature, lighting conditions) have such an effect. In, the framework of the two-process model (sect. II~), they facilitate sleep onset by lowering the upper threshold of process C and promote sleep maintenance bY lowering the lower threshold. The second cl .ass of agents are assumed to mediate sleep homeostasis. By increasing EEG slow-wave activity and/or the amount of REM sleep, they mimic the action of total or selective sleep depri- vation. Because the determination of sleep latency and sleep duration is insufficient to discriminate between the two classes, an intensity measure of NREM sleep should be obtained. However, even when using this measure, a strict discrimination is difficult to achieve. Thus a permissive factor may augment sleep and thereby favor the occurrence of EEG slow-wave activity and REM sleep. Nevertheless, as shown by the nap study in humans (?l), the amount of slow-wave activity is determined .by a homeostatic process even when it is elicited by permissive daytime conditions.
T. Permissive versus hmeostatic sleep-promoting action
Also Jouvet (167) discriminated sleep-facilitatory and sleep-promoting factors. The former induce drowsiness by facilitating permissive mecha- nisms but do not affect the postulated executive sleep mechanisms. The latter act directly on executive mechanisms and their level increases during sleep deprivation. Their inactivation should lead to total insomnia or to.a selective suppression of NREM and/or REM sleep. It should be possible to maintain these changes for a long time without subsequently inducing a compensatory rebound. In fact, the destruction of paramedial preoptic neurons in cats resulted in a reduction or disappearance of deep NREM and REM sleep, while light NREM sleep was largely maintained (325). The sleep changes were well tolerated by the animals. These results indicate that it is possible with homeostatic sleep-regulating systems. Endogenous sleep
to interfere substances
belonging to the homeostatic class may activate these systems or mediate their action.
A induce
sleep substance a type of sleep
isolated from sleep -deprived donor anim als should that corresponds to recovery sleep i ri the donor. An
increase of slow-wave activity was in fact observed after administration of brain-derived and urinary factor S as well as the muramyl peptide AMTP.

650 ALEXANDER A. BORBELY AND IRENE TOBLER vohrw 69
However, a REM sleep-enhancing action of these compounds was not ob- served. The REM sleep was either suppressed or unchanged. An increase of both NREM and REM sleep was reported for SPS, SPS-B, and uridine. How- ever, since these effects in rodents were obtained only in the dark period and uridine was ineffective in the light period, a permissive action on sleep ap- pears to be more likely than an effect on sleep homeostasis. This statement applies only to REM sleep, since EEG slow-wave activity was not recorded in these studies.
Selective REM sleep deprivation has yielded a REM sleep-promoting factor in the CSF and brain of donor animals (sect. 112). However, with one exception (416), the effect was tested in pharmacologically pretreated ani- mals in which REM sleep had been suppressed. The massive REM sleep rebound that is normally seen after REM sleep deprivation in intact animals has not yet been elicited in this type of experiment. Whether this is due to the small quantity of the active principle transferred or to some other cause remains to be established. Finally, it should be realized that the effects induced by total or selective sleep deprivation may be only in part due to a high level of putative sleep substances. Other changes (e.g., alteration in receptor sensitivity) might sleep in the recovery period
also contribute to the increased and intensified . They would not be present when putative sleep
substances are administered to normal recipients.
8. Are there specific norwapid-eye-movement and rapid-eye-movement sleep-promoting factors?
Factor S, the muramyl peptides, as well as the immunologically active polypeptides enhance slow-wave activity in NREM sleep and generally, but not invariably, depress REM sleep. Thus by specifically intensifying NREM sleep they may mediate NREM sleep homeostasis. Moreover, since an inhibi- tory interaction between REM sleep and slow-wave sleep has been postulated (27), the suppression of REM sleep may be due to a shift in balance between the two substates of sleep caused by the slow-wave-promoting compounds. Alternatively, since REM sleep is known to be a “fragile” state that is readily depressed by a variety of factors (e.g., stress, high or low ambient tempera- ture), the inhibition of REM sleep may represent a nonspecific effect (sect. 11C4).
Not all factors or substances that were derived from the sleep depriva- tion approach suppress REM sleep. Sleep-promoting substance, SPS-B, and uridine enhance both NREM and REM sleep. However, as has been men- tioned in the preceding section, these effects may be due to permissive rather than to homeostatic mechanisms.
The REM sleep-promoting factor contained in the CSF of REM sleep- deprived donors was able to counteract pharmacological REM sleep suppres- sion in recipients (sect. 1123). However, its specificity for REM sleep is ques-

April 1989 ENDOGENOUS SLEEP-PROMOTING SUBSTANCES 651
tionable in view of the observations that deep NREM sleep was restored as well in both the rat (86) and cat (322,323). It should be kept in mind that REM sleep-deprivation procedures may induce not only a REM sleep rebound but also an augmentation in deep NREM sleep (sect. 114). A close link in the regulation of NREM sleep intensity and REM sleep is suggested also by the observation that the destruction of the paramedian preoptic neurons abol- ished both deep NREM sleep and REM sleep, leaving only light NREM sleep (325).
In conclusion, whereas some compounds intensify specifically NREM sleep without a concomitant increase of REM sleep, the specificity of REM sleep-promoting factors has not yet been conclusively demonstrated.
9. Circadian diflerences
Circadian differences in the sleep-promoting action of various com- pounds have been reported for the rat. In contrast to the cat and rabbit, the rat has a distinct circadian sleep/waking rhythm. Substances showing a sleep-promoting action in the dark period (the rat’s circadian period of pre- dominant waking) were ineffective in the light period. Such a difference in activity has been reported for uridine, PGD2, and DSIP (153). The authors concluded that during the light period the endogenous sleep-promoting sys- tem is maximally operative, filling all the time available for sleep, and that, consequently, the administration of sleep-promoting compounds is not able to induce excess sleep. This interpretation is valid only for compounds that affect sleep via a permissive action. They favor sleep during the dark period when the intrinsic sleep tendency is low, but they are ineffective during the light period when the intrinsic sleep tendency is high. A similar circadian difference has been observed for the sleep-promoting action of short light periods (23). The argument of sleep saturation does not apply to compounds activating or mimicking homeostatic mechanisms. When the recovery period from sleep deprivation coincides with the light period, EEG slow-wave activ- ity is massively enhanced and the amount of REM sleep is increased (40,375). Thus even if sleep is “physiologically saturated,” NREM sleep can be inten- sified and REM sleep time expanded (167). The former effect could be induced in the rat by the administration of the 5-HT2-receptor antagonist ritanserin (40a, 87) and IL-l (377).
C Changes Associated With Sleep and Waking
The second, basic requirement that we postulate for an endogenous sleep factor or sleep substance is its variation in the organism in association with sleep and waking (Table 2). This criterion has not yet been shown to be fulfilled by any of the factors and substances covered in this review. With the development of sensitive techniques for determining the cerebral concentra-

652 ALEXANDER A. BORBeLY AND IRENE TOBLER Vohme 69
tion of endogenous substances it may become possible to experimentally address this problem. For example, the in vivo application of electrochemical detection methods (48) allows the concomitant recording of neurochemical and electrophysiological parameters from specific brain sites. However, there is a large number of biologically.active substances (e.g., IL-l, IFN) that currently cannot be directly determined. Of course, the failure to demon- strate sleep/wake-associated variation in the brain or CSF does not necessar- ily imply that a substance is not involved in sleep regulation. The changes may be restricted to a specific site in the brain or they may not be reflected by a variation of the concentration of the substance. On the other hand, the demonstration of changes linked to sleep and waking would constitute evi- dence for its involvement in sleep regulation. There is some preliminary evidence for sleep-related variations in the level of substances that in turn may modify sleep. This includes 1) the rise in the plasma level of GH during sleep that is well documented in humans and that may occur in other mam- mals as well (sect. IIIES), 2) the increase in the cerebral level of glycogen after sleep onset (sect. IIIB), and 3) the rise in IL-l-like activity in human plasma and cat CSF in association with sleep (sect. IIIF~). Because these neurochemi- cal events seem to be linked to sleep onset, they may be involved in the control of state transitions rather than in the long-term regulation of sleep propen- sity. Whether the diurnal rhythm in cerebral PGD2-synthetase activity is related to diurnal variation in sleep propensity remains to be established.
Even though a direct link between neurochemical parameters and sleep homeostasis has not yet been demonstrated, two observations provide strong evidence for such an association. First, the suppression of motor activity of rats receiving goat CSF depended on the duration of enforced waking in the donor animals (98). Second, the REM sleep-restoring potency of CSF admin- istered to pharmacologically pretreated recipients depended on the duration of REM sleep deprivation of the donor animals (3, 4). The results of both studies are consistent with the assumption that deprivation of total sleep or REM sleep leads to a progressively increasing brain level of a sleep factor that is responsible for the compensatory changes in sleep during the recovery period. There is recent evidence that the hypothalamopituitary system may be involved in the elaboration of the REM sleep-promoting factor (86,327).
D. Cmclusim
1. Relations to sleep physiology
Sleep deprivation constitutes a most direct experimental approach to the problem of the chemical control of sleep. It is based on the assumption that the increase in “sleep pressure” activates endogenous sleep-promoting neu- rochemical processes. After Pi&on’s pioneering experiments it was Pappen- heimer and co-workers who obtained convincing evidence for the existence of

April 1989 ENDOGENOUS SLEEP-PROMOTING SUBSTANCES 653
a transferable sleep factor. They showed that its administration induces the type of deep NREM sleep that is typically seen during recovery from sleep deprivation. It was to be expected that the identification of the active constit- uent would provide basic insights into the neurochemistry of physiological sleep. These expectations have not yet been fulfilled. The main problem is that the physiological role of muramyl peptides and of the other immunologi- cally active peptides is largely obscure. This is the case also for SPS for which only one constituent has been chemically identified and for the REM sleep- promoting factors, the identity of which remains obscure.
E led to
llectrical stimulation of the recognition of a som
the median thal .amus was the technique that nogenic factor in venou s blood that was eventu-
ally identified as the nonapeptide DSIP. The mechanisms involved in electri- cally induced sleep are largely unknown, and it is unclear whether the stimu- lation does activate systems that are involved in physiological sleep regula- tion brai
Moreo .h that
ver, thalamic stimulation may affect a variety of pathways are unrelated to sleep. In the light of these. problems it
in the is not
surprising that more than 10 years after its isolation, the involvement of DSIP in sleep regulation is still open to debate.
Approaches based on spontaneous sleep h ave not led to the them ical identification of a sleep-promoting substance. Neverth eless, they have re- vealed a somnogenic factor as well as a REM sleep-related increase in level in perfusates of the mesencephal ic reticular formati .on.
The recognition of the somnogenic action of neuromodu lators and neuro- transmitters has not been the result of approach .es based on sleep deprivation or spontaneous sleep. Consequently ‘, their role in physiological sleep regula- tion is even more difficult to specify than for the aforementioned corn #pounds. On the other hand, the large body of neurochemical and physiological data provides a background for further investigating the sleep-promoting mechanisms.
protein
2. Speci$city for sleep
Is it reasonable to postulate than an endogenous sleep-promoting sub- stance be involved exclusively in sleep regulation? All substances that have been chemically identified are involved in functions other than sleep. Thus the immunologically active polypeptides mediate a wide variety of effects in addition to their actions on the immune system (e.g., see Ref. 93). In view of the many different effects of DSIP one may ask whether the designation of this compound as a sleep-inducing peptide was not a misnomer (121). It is evident that the neuromodulators and neurotransmitters are not specific to sleep, since they are involved in other functional systems.
Because sleep substances may mediate functions other than sleep, the sleep promotion may represent a secondary effect. This concern has been particularly serious in view of the hyperthermic action of muramyl peptides

654 ALEXANDER A. BORBELY AND IRENE TOBLER Vohme 69
and immunologically active polypeptides. Because the somnogenic action was still present after attenuation of the hyperthermic action and because the physiological sleep-related shifts in body temperature persisted, separate mechanisms may underlie the effects on sleep and body temperature (sect. 110’). It is possible that the somnogenic action of these compounds originates at specific sites in the brain and that the large doses administered intraven- tricularly affect also other systems in the brain. Further progress will be made when sleep-associated changes of sleep substances in specific cerebral structures or systems are documented (sect. IvC).
3. Activation and deactivation of sleep-promoting substances
Increasing “sleep pressure” by total or selective sleep deprivation or by electrical brain stimulation has been the method for inducing transferable sleep-promoting substances in the brain or CSF of donor animals. In some experiments material from donor animals that had not been subjected to sleep manipulations were used as controls. Brain stem extracts of donor rats that had not been sleep deprived failed to augment sleep in recipients (387). Similarly, CSF (295), brain extracts (416), and hypothalamic or pituitary extracts (86) from control animals lacked the REM sleep-promoting activity that was present in material derived from REM sleep-deprived donors. Fi- nally, venous dialysate obtained from sham-stimulated donor animals did not increase sleep in the recipients (252,253). These results may suggest that sleep-promoting substances are not present in the brain unless sleep pressure is incr*eased. However, it appears more likely that in the experiments men- tioned above, the concentration in the material obtained from control ani- mals was not sufficient to induce a significant change in the recipients. This interpretation is supported by the fact that factor S was obtained from the brains of animals that had not been experimentally sleep deprived and from human urine where the donors are not known to have suffered a sleep deficit. Furthermore, specific brain mRNA, the level of which was modulated by sleep deprivation, was present also in the brain of nondeprived control ani- mals (3lOa). Finally, CSF obtained from control goats also depressed loco- motor activity in recipient rats (278), and CSF obtained from donor cats that had not been REM sleep deprived restored in some cases REM sleep in pharmacologically pretreated recipients (322, 323). These results indicate that sleep-promoting substances are continuously present in the brain and that it is their concentration or their activity that is modulated as a function of sleep and waking.
A methodological aspect merits attention when sleep is induced by in- traventricular infusion. Infusing saline or artificial CSF into the cerebral ventricles had a REM sleep-suppressing effect in the rat (38,39,195,377,383), cat (109,323), and rabbit (405) and a sleep-suppressing effect in the monkey& (275a). Even continuously infusing saline at a low infusion rate reduced the

&rill989 ENDOGENOUS SLEEP-PROMOTING SUBSTANCES 655
circadian amplitude of the sleep/waking rhythm in the rat (149). The REM sleep both
I-suppressing the rate and
and motor activity-reducing effects in the rat depended on the volume of the infusate (38). These changes may repre-
sent nonspecific effects due to the nonphysiological stimulation of the “inner surface” of the brain (219a). However, it is not excluded that the stimulation of the ependymal cells lining the ventricular wall triggers the release of substances into the CSF. Regardless of the mechanisms, one should be aware that the intraventricular infusion technique may interfere with sleep regulation.
Although the. mechanisms by which sleep substances are activated are still completely obscure, some hypotheses have been advanced. Thus, since the muramyl peptide AMTP loses its somnogenic activity by amidation or hydration, it was proposed that amidation/deamidation or hydration/dehy- dration reactions may be involved in the physiological inactivation/activa- tion of these compounds (196). However, experimental evidence in support of this assumption is not yet available. The sleep-promoting peptides des-a- MSH and CLIP have pro-opiomelanocortin as the common precursor mole- cule (sect. IIIEG). It is conceivable that the specific enzymatic cleavage of the precursor regulates the level of the sleep-promoting peptide (51). The mecha- nisms considered so far are based on the assumption that specific enzymatic processes play a key role in regulating the level of sleep-promoting sub- stances. The question then arises how the activity of the enzymes are regu- lated and whether crucial regulatory processes occur at the transcriptional level (see sect. 11~46). Finally, the possibility should be considered that total or selective sleep deprivation induces changes in receptors of neurotransmit- ters or neuromodulators (see sect. IVB7). As has been mentioned, the data regarding catecholamine receptor changes after REM sleep deprivation are controversial (sect. IILZW). However, it should be realized that alterations of receptors bound to neuronal membranes cannot account for the fact that sleep-promoting substances can be transferred in the CSF from donors to recipients.
4. Therapeutic implications
The identification of endogenous sleep-promoting substances may even- tually lead to their use in the therapy of sleep disorders. It is reasonable to expect that, unlike the currently used benzodiazepine hypnotics (33), endoge- nous substances will induce a more physiological type of sleep. It is even conceivable that they may be administered to substitute for a deficiency in the brain. Unfortunately, there is presently little evidence in support of such speculations. The effectiveness of the two compounds that have been used in the therapy of insomnia are questionable. The beneficial effect of DSIP has yet to be confirmed in rigorous crossover schedules (sect. IIIB3), and the usefulness of tryptophan is controversial (sect. IIG!B). Neither DSIP nor

656 ALEXANDER A. BORBELY AND IRENE TOBLER Volume 69
tryptophan and its metabolites have been shown to be deficient in insomniac patients, and for neither compound has an activation of physiological sleep- inducing mechanisms been documented. The somnogenic action of AVT in humans (sect. IIIES) requires confirmation. Preliminary results indicate that melatonin may alleviate jet lag (11), but it is unlikely that sleep regulation is specifically influenced. In conclusion, therapeutic applications in humans must await further clarification of basic mechanisms.
5. Concepts and strategies for future research
The research strategies in the search for endogenous sleep-promoting substances depend on the conceptual framework as well as on the techniques available. It has been the leitmotiv of the present review that sleep physiol- ogy should serve as a guideline in exploring the neurochemistry of sleep regulation. In particular, the distinction between homeostatic and permis- sive factors was proposed for conceptualizing the mode of action of endoge- nous compounds.
Although the nature of the neurochemical processes mediating sleep are still obscure, two rather explicit hypotheses have been recently advanced (164,166,201). According to Jouvet (164,166), a sequence of events underlies the regulation of sleep, which can be analyzed by experimental manipulation. Thus the observation that CSF obtained from REM sleep-deprived donors was capable of restoring REM sleep in recipients pretreated with PCPA (322, 323) indicated that an intact serotonergic system is not a prerequisite for the occurrence of REM sleep. In the chain of events, the REM sleep-promoting factor is therefore “downstream” with respect to 5-HT. The more rapid restoration of REM sleep by the REM sleep-promoting factor than by 5-HTP (324) is consistent with this interpretation. It has been suggested that 5-HT might be involved in the elaboration of a sleep-promoting factor during waking (164-166).
Krueger et al. (201) have proposed a cascade of biochemical events lead- ing to the induction of slow-wave sleep. Their scheme includes 12 sleep-pro- moting substances that interact at cellular and extracellular sites. A role of glial cells in sleep regulation is suggested by the binding of muramyl peptides and 5-HT to glial membranes as well as by the glial production of IL-l and INF. Although this highly speculative hypothesis is still based on observa- tions made in vitro, it shows how a variety of different sleep-promoting substances might interact and how they may therefore serve as a guide for further experiments.
The chemical identity of a sleep-promoting factor derived from CSF or brain was initially unknown. Nevertheless, simple tests have yielded impor- tant preliminary information. Thus the peptidic nature of factor S has been early recognized by the loss of its somnogenic effect after treatment with pronase (98). Similarly, the observations that the REM sleep-promoting fac-

April 1989 ENDOGENOUS SLEEP-PROMOTING SUBSTANCES 657
tor obtained from CSF loses its effectiveness after either heat exposure or after pretreatment of the donor animals with the protein synthesis inhibitor chloramphenicol (295) suggest the involvement of a polypeptide. This as- sumption is supported by reports that REM sleep in cats is suppressed by chloramphenicol (78,286) but is’markedly augmented by bacitracin, a pepti- dase inhibitor (328). Cycloheximide induced a prolonged elevation of REM sleep during the period when protein synthesis reverted to its normal state (358). Taken together, the involvement of protein synthesis in the elaboration of a sleep-promoting substance raises the possibility of using a molecular genetic approach for its chemical identification provided that the relevant changes occur at the transcriptional level (see sect. IIIA~).
Karnovsky and Reich (178) concluded their review on the biochemistry of sleep with the following statement.
Despite conceptual and technical advances in neurophysiology, psychology, and bio- chemistry, investigators in this field may be said to be toiling more on the virgin plains than in orderly vineyards. Perhaps the matter of most importance is the realization that so many studies have been done, and many must be currently under way. This, coupled with increasing rigor of approach and imaginative design, leads one to the optimistic feeling that there is indeed a legitimate field to be explored.
Since these lines were written a decade ago the field has considerably ( ex- panded, partly in unforeseen dire&ions. Although it is still outside the or- derly vineyards of neuroscience, it attracts an increasing number of qualified scientists who are ready to explore this uncharted territory. There is room for many more.
We gratefully acknowledge the extremely helpful comments by Dr. J. M. Krueger on an earlier version of the manuscript. We thank the following colleagues for kindly making available their recent, unpublished data: Drs. R. Cespuglio, C. Dugovic, 0. Hayaishi, S. Inoue, V. Kovalzon, J. M. Krueger, W. B. Mendelson, F. Obal, N. Okudaira, and R. Scherschlicht. We gratefully acknowledge the help of Karin Jaggi in preparing this review,
This work was supported by the Swiss National Science Foundation Grant 3.234-0.85.
REFERENCES
1. ABEL, M. S., F. VILLEGAS, J. ABREU, F. GIMINO, S. STEINER, B. BEER, AND L. R. MEYERSON. The effect of rapid eye movement sleep deprivation on cortical /3- adrenergic receptors. Brain Res. BulL 11: 729-734,1983.
2. ADRIEN, J. Endogenous sleep factors-a critical ap- proach. In: Sleep ‘86, edited by W. P. Koella, F. Obirl, H. Schulz, and P. Visser. Stuttgart, FRG: Gustav Fischer- Verlag, 1988, p. 161-181.
3. ADRIEN, J., AND C. DUGOVIC. Presence of a paradoxi- cal sleep (PS) factor in the cerebrospinal fluid of PS-de- prived rats. Eur. J. PharmacoL 100: 223-226,1984.
4. ADRIEN, J., AND C. DUGOVIC. PS-inducing factors and the noradrenergic system. In: Endogenous Sleep Sub stunces and Sleep Reguh%m, edited by S. Inoue and A. A. Bortiiy. Tokyo: Jpn. Sci. Sot. Press, 1985, p. 227-235.
5. AKERSTEDT, T. Hormones and sleep. In: Esqwrimtal
Brain Research, Sleep Mechanisms, edited by A. A. Bortily and J. L. Valatx. Berlin: Springer-Verlag, 1984, suppl. 8, p. 193-203.
6. AKERSTEDT, T. Sleepiness as a consequence of shift work. Sleep 11: 17-34,1988.
‘7. AKERSTEDT, T., J. E. FRijBERG, Y. FRIBERG, AND L. WETTERBERG. Melatonin excretion body temperature and subjective arousal during 64 hours of sleep depriva- tion. Pq&wneur~~ 4: 219~225,1979.
8. AKERSTEDT, T., AND M. GILLBERG. The circadian variation of experimentally displaced sleep, Sleep 4: 159-169,1981.
9. AKERSTEDT, T., AND M. GILLBERG. Sleep duration and the power spectral density of the EEG. EZectroence- phabgr. Clin. Neuroph&oL 64: 119-122,1986.
10. ANTON-TAY, F., J. L. DIAZ, AND A. FERNANDEZ-

658 ALEXANDER A. BORBELY AND IRENE TOBLER vohne 69
GUARDIOLA. On the effect of melatonin upon human brain. Its possible therapeutic implications. L$e Sci. 10: 841-850,197l.
11. ARENDT, J., M. ALDHOUS, AND V. MARKS. Allevia- tion of jet lag by melatonin: preliminary results of con- trolled double blind trial. Br. Med J. 292: 1170, 1986.
12. ARENDT, J., M. ALDHOUS, AND J. WRIGHT. Synchronisation of a disturbed sleep-wake cycle in a blind man by melatonin treatment. Lancet 1: 772-773, 1988.
13. ARENDT, J., C. BOJKOWSKI, S. FOLKARD, C. FRANEY, V. MARKS, D. S. MINORS, J. M. WATER- HOUSE, R. A. WEVER, C. WILDGRUBER, AND J. WRIGHT. Some effects of melatonin and the control of its secretion in man. In: Phdoperiodism, the Pined Gland and MeZutonin, edited by D. Evered and S. Clark. 1985, p. 266-279. (Ciba Symp.)
14. ARENDT, J., A. A. BORBELY, C. FRANEY, AND J. WRIGHT. The effects of chronic, small doses of melato- nin given in the late afternoon on fatigue in man: a pre- liminary study. Neurosti Lett. 45: 317-321,1984.
15. ASERINSKY, E. The maximal capacity for sleep: rapid eye movement density as an index of sleep satiety. Biol, Ps@chiatqj 1: 147-159,1969.
16. BAGHDOYAN, H. A., M. L. RODRIGO-ANGULO, R. W. McCARLEY, AND A. HOBSON. A neuroanatomical gra- dient in the pontine tegmentum for the cholinoceptive induction of desynchronized sleep signs. Bruin Res. 414: 245-261,1987.
17. BEERSMA, D. G. M., S. DAAN, AND D. J. DIJK. Sleep intensity and timing-a model for their circadian con- trol. Lect. Math L$e Sci 19: 39-62,1987.
18. BENEDEK, G., F. OBAL, JR., L. SZEKERES, AND F. OBAL. Cortical synchronization induced by thermal stimulation of the preoptic area in immobilized rats. Actu Ph@ol Acad Sci Hung. 48: 65-72,1976.
19. BERGER, R. J., AND I. OSWALD. Effects of sleep depri- vation on behavior, subsequent sleep and dreaming. J. Merit. Sci. 108: 457-465,1962.
20. BLAKE, H., AND R. W. GERARD. Brain potentials dur- ing sleep. Am J PhysioL 119: 692-703, 1937.
21. BLOIS, R., M. MONNIER, R. TISSOT, AND J.-M. GAIL- LARD. Effect of DSIP on diurnal and nocturnal sleep in man. In: Sleep ‘80, edited by W. P. Koella. Basel: Karger, 1981, p. 301-303.
22. BOBILLIER, P., S. SEGUIN, F. PETITJEAN, C. BUDA, D. SALVERT, M. JANIN, G. CHOUVET, M. JOUVET, AND M. H. DES ROSIERS. Glucose utilization increases in choroid plexus during slow wave sleep. A [‘“Cl deoxy- glucose study in the cat. Bruin Res. 240: 359-363,1982.
23. BORBELY, A. A. Circadian rhythm of vigilance in rats: modulation by short light-dark cycle. Neurosci. Lett. 1: 67-71,1975.
24. BORBELY, A. A. Sleep and motor activity of the rat during ultra-short light-dark cycles. Bruin Res. 114: 305-317,1976.
25. BORBELY, A. A. Sleep in the rat during food deprivation and subsequent restitution of food. Bruin Res. 124: 457-471,1977.
26. BORBELY, A. A. Effects of light and circadian rhythm on the occurrence of REM sleep in the rat. Sleep 2: 289-298,198O.
27. BORBELY, A. A. A two process model of sleep regula- tion. Hum NeurobioL 1: 195-204,1982.
28. BORBELY, A. A. Sleep regulation: circadian rhythm and homeostasis. In: Sleep. Clinical and Experimental Aspects, edited by D. Ganten and D. Pfaff. New York: Springer-Verlag, 1982, vol. 1, p. 83-103. (Curr. Top. Neuroendocrinol.)
29. BORBELY, A. A. Endogenous sleep-substances and sleep regulation. J. Neural Trunsm. SuppL 21: 243-254,1986.
30. BORBELY, A. A., P. ACHERMANN, L. TRACHSEL, AND I. TOBLER. Sleep homeostasis in humans and rats. In: Clinical Ph2/siologp of Skep, edited by R. Lydic and J. F. Biebuyck. Bethesda, MD: Am. Physiol. Sot., 1988, chapt. 13, p. 191-198.
31. BORBELY, A. A., G. BALDERER, L. TRACHSEL, AND I. TOBLER. Effect of midazolam and sleep deprivation on day-time sleep propensity. Arzneim. Forsch. 35: 1696-1699,1985.
32. BORBELY, A. A., F. BAUMANN, D. BRANDEIS, I. STRAUCH, AND D. LEHMANN. Sleep-deprivation: ef- fect on sleep stages and EEG power density in man. Electroencephulogr. Clin Neuroph~sioL 51: 483-493,198l.
33. BORBELY, A. A., P. MATTMANN, M. LOEPFE, I. STRAUCH, AND D. LEHMANN. Effect of benzodiaze- pine hypnotics on all-night sleep EEG spectra. Hum. NeurobioL 4: 189-194,1985.
34. BORBELY, A. A., AND H.-U. NEUHAUS. Daily pattern of sleep, motor activity and feeding in the rat: effects of regular and gradually extended photoperiods. J. Comp. PhysioL 124: l-14,1978.
35. BORBELY, A. A., AND H.-U. NEUHAUS. Circadian rhythm of sleep and motor activity in the rat during skeleton photoperiod, continuous darkness and continu- ous light. J. Comp. Ph@oL 128: 37-46,1978.
36. BORBELY, A. A., AND H.-U. NEUHAUS. Sleep-depriva- tion: effects on sleep and EEG in the rat. J. Camp. Phg- SiOL 133: 71-87, 1979.
37. BORBELY, A. A., P. STEIGRAD, AND I. TOBLER. Effect of sleep deprivation on brain serotonin in the rat. Behuv. Bruin Res. 1: 205-210,198O.
38. BORBELY, A. A., AND I. TOBLER. Cerebroventricular infusion in the rat: depression of motor activity and par- adoxical sleep. Neurosci L&t. 12: 75-80,1979.
39. BORBELY, A. A., AND I. TOBLER. The search for an endogenous “sleep-substance.” Trends PhwmucoL Sci 1: 356-358,198O.
40. BORBELY, A. A., I. TOBLER, AND M. HANAGA- SIOGLU. Effect of sleep deprivation on sleep and EEG power spectra in the rat. Behuv. Bruin Res. 14: 171-182, 1984.
4Oa.BORBELY, A. A., L. TRACHSEL, AND I. TOBLER. Ef- fect of ritanserin on sleep stages and sleep EEG in the rat. Eur. J PhurrruzcoL 156: 275-278,1988.
41. BORBELY, A. A., AND G. YOUMBI-BALDERER. Effects of tryptophan on human sleep. Interdiscip. Top. GerontoL 22: ill-127,1987.
42. BREDER, C. D., C. A. DINARELLO, AND C. B. SAPER. Interleukin-1 immunoreactive innervation of the human hypothalamus. science Wash. DC 240: 321-324,1988.
43. CAMPBELL, S. S. Duration and placement of sleep in a “disentrained” environment. Psychophysiology 21: 106-113,1984.
44. CARSKADON, M. A., AND W. C. DEMENT. Cumulative effects of sleep restriction on daytime sleepiness. Psg- choph&Zogjj 18: 107-113,198l.
45. CARSKADON, M. A., W. C. DEMENT, M. M. MITLER, T. ROTH, P. R. WESTBROOK, AND S. KEENAN. Guide- lines for the multiple sleep latency test (MSLT): a stan- dard measure of sleepiness. Sleep 9: 519-524,1986.
46. CARTWRIGHT, R. D., L. J. MONROE, AND C. PALMER. Individual differences in response to REM deprivation. Arch. Gen Psgchiutr/ 16: 297-303,1967.
47. CESPUGLIO, R., H. FARADJI, M. E. GOMEZ, AND M. JOUVET. Single unit recordings in the nuclei raphe dor- salis and magnus during the sleep waking cycle of semi- chronic prepared cats. Neurosci. L&t. 24: X33-138,1981.

April 1989 ENDOGENOUS SLEEP-PROMOTING SUBSTANCES 659
48.
49.
50.
51.
52
53
54
55
56.
57.
58.
59.
60.
61.
62.
63.
CESPUGLIO, R., H. FARADJI, G. GUIDON, AND M. JOUVET. Voltametric detection of brain 5-hydroxyindo- lamines: a new technology applied to sleep research. In: Experimentul Brain Research, Sleep Mechanisms, edited by A. A. Borbely and J. L. Valatx. Berlin: Springer-Ver- lag, 1984, suppl. 8, p. 95-105. CESPUGLIO, R., H. FARADJI, F. RIOU, M. BUDA, F. GONON, J. F. PUJOL, AND M. JOUVET. Differential pulse voltammetry in brain tissue. II. Detection of 5-hy- droxyindolacetic acid in the rat striatum. Brain Res. 223: 299-311,198l. CHASTRETTE, N., AND R. CESPUGLIO. Influence of proopiomelanocortin-derived peptides on the sleep-wak- ing cycle of the rat. Neurosci Lett. 62: 365-370,1985. CHASTRETTE, N., H. W. CLEMENT, H. PREVAUTEL, AND R. CESPUGLIO. Proopiomelanocortin components: differential sleep-waking regulation? In: Sleep Peptides: Basic and Clinical Approaches, edited by S. Inoue and D. Schneider-Helmert. Tokyo: Jpn. Sci. Sot. Press, 1988, p. 27-52. CHURCH, M. W., J. D. MARCH, S. HIBI, C. CAVNESS, AND I. FEINBERG. Changes in frequency and amplitude of delta activity during sleep. Electroencephulogr. Clin. Neuroph@oL 39: l-7,1975. CLOETTA, M., AND H. FISCHER. Ueber die Wirkung der Kationen Ca, Mg, Sr, Ba, K und Na bei intrazere- braler Injektion. Arch. Exp. PathoL Pharmakol. 158: 254-280,193o. COCULESCU, M., A. SERBANESCU, AND E. TEMELI. Influence of arginine vasotocin administration on noc- turnal sleep of human subjects. Waking Sleeping 3: 273-277,1979. COENEN, A. M. L., AND E. L. J. M. VAN LUIJTELAAR. Stress induced by three procedures of deprivation of paradoxical sleep. Ph@oL Behuv. 35: 501~504,1985. CONN, P. J., AND E. SANDERS-BUSH. Central seroto- nin receptors: effector systems, physiological roles and regulation. Ps~chophu~ 92: 267-277,1987. CRAMER, H., W. BijHME, K. KENDEL, AND M. DON- NADIEU. Freisetzung von Wachstumshormon und von Melanozyten stimulierendem Hormon im durch Melato- nin gebahnten Schlaf beim Menschen. Arzneim Forsch 26: 1076-1078,1976. CRAMER, H., J. RUDOLPH, U. CONSBRUCH, AND K. KENDEL. On the effects of melatonin on sleep and be- havior in man. Adv. Biochem. PsychopharmacoL 11: 187-191,1974. CZEISLER, C. A., J. C. ZIMMERMAN, J. M. RONDA, M. C. MOORE-EDE, AND E. D. WEITZMAN. Timing of REM sleep is coupled to the circadian rhythm of body temperature in man. Sleep 2: 329-346,198O. DAAN, S., D. G. M. BEERSMA, AND A. A. BORBELY. Timing of human sleep: recovery process gated by a cir- cadian pacemaker. Am J. Ph@oL 246 (ReguUny Inte- grative Comp. PhgsioL 15): R161-R178,1984. DANGUIR, J. Sleep deficits in diabetic rats: restoration following chronic intravenous or intracerebroventricular infusions of insulin. Brain Res. B&L 12: 641~645,1984. DANGUIR, J. Intracerebroventricular infusion of soma- tostatin selectively increases paradoxical sleep in rats. Brain Res. 367: 26-30,1986. DANGUIR, J. Internal milieu and sleep homeostasis. In: Sleep Peptides: Basic and Clinical Approaches, edited by S. Inoue and D. Schneider-Helmert. Tokyo: Jpn. Sci. Sot. Press, 1988, p. 53-72.
64. DANGUIR, J., AND S. DE SAINT-HILAIRE-KAFI. So- matostatin antiserum blocks carbachol-induced increase of paradoxical sleep in the rat. Brain‘Res. Bull. 20: 9-12, 1988.
65.
66.
67.
68.
69.
70.
71.
72.
73.
74.
75.
76.
77.
78
DANGUIR, J., AND S. NICOLAIDIS. Chronic intracere- broventricular infusion of insulin causes selective in- crease of slow wave sleep in rats. Brain Res. 306: 97-103, 1984. DAVENNE, D., AND J. M. KRUEGER. Enhancement of quiet sleep in rabbit neonates by muramyl dipeptide. Am J. PhysioL 253 (Regulutoy/ Integrative Camp. Phy- sioL 22): R646-R654,1987. DAVID, J., R. S.. GREWAL, AND G. P. WAGLE. Re- stricted sleep regime in rhesus monkeys: differential ef- fect of one night’s sleep loss and selective REM depriva- tion. Life Sti 16: 1375-1386,1975. DE ANDRES, I., E. GUTIERREZ-RIVAS, E. NAVA, AND F. REINOSO-SUAREZ. Independence of sleep- wakefulness cycle in an implanted head “encephale isole.” Neurosci. L&t. 2: 13-18, 1976. DECLERCK, A. C., A. WAUQUIER, P. H. M. VAN DER HAM-VELTMAN, AND Y. GELDERS. Increase in slow- wave sleep in humans with the serotonin-Ss antagonist ritanserin. Cum: Ther. Res. Clin, Exp. 41: 427-432,1987. DEMESQUITA, S., AND W. HERSHEL HANEY. Effect of chronic intracerebroventricular infusion of cholecys- tokinin on respiration and sleep. Brain Res. 378: 127-132, 1986. DIJK, D. J., D. G. M. BEERSMA, AND S. DAAN. EEG power density during nap sleep: reflection, of an hour- glass measuring the duration of prior wakefulness. J. BioL [email protected] 2: 207-219,1987. DRUCKER-COLfN, R. R. Crossed perfusion of a sleep inducing brain tissue substance in conscious cats. Brain Res. 56: 123-134,1973. DRUCKER-COLfN, R. R., R. AGUILAR-ROBLERO, AND G. ARANKOWSKY-SANDOVAL. Sleep factors released from brain of unrestrained cats: a critical appraisal. Ann. NYAcud Sci 473: 449-460,1986. DRUCKER-COLIN, R. R., J. G. BERNAL-PEDRAZA, F. FERNANDEZ-CANCINO, AND A. R. MORRISON. In- creasing PGO spike density by auditory stimulation in- creases the duration and decreases the latency of rapid eye movement (REM) sleep. Brain Res. 278: 308-312, 1983. DRUCKER-COLiN, R. R., J. G. BERNAL-PEDRAZA, F. FERNANDEZ-CANCINO, AND A. OKSENBERG. Is va- soactive intestinal polypeptide (VIP) a sleep factor? Peptides Fagetteville 5: 837~840,1984. DRUCKER-COLiN, R. R., AND M. DEL CARMEN GUTIERREZ. Effects of forebrain lesions on release of proteins from the midbrain reticular formation during the sleep-wake cycle. Exp. NewoL 52: 339-344,1976. DRUCKER-COLIN, R. R., M. DEL CARMEN
GUTIERREZ, AND J. G. BERNAL-PEDRAZA. Polypep- tide regulation of REM sleep. Front. Horm. Res. 6: 138-155,198O. DRUCKER-COLiN, R. R., AND M. ESPEJEL. Chronic administration of chloramphenicol, a protein synthesis inhibitor, selectively decreases REM sleep. Behav. Neural Bid 29: 410-413,198O. DRUCKER-COLfN, R. R., 0. PROSPERO-GARCIA, G. ARANKOWSKY-SANDOVAL, AND R. PEREZ-MONT- FORT. Gastropancreatic peptides and sensory stimuli as REM sleep factors. In: Sleep Peptides. Basic and Clinical Approaches, edited by S. Ino& and D. Schneider-Hel- mert. Tokyo: Jpn. Sci. Sot. Press, 1988, p. 73-94. DRUCKER-COLfN, R. R., J. A. ROJAS-RAMIREZ, J. VERA-TRUEBA, G. MONROY-AYALA, AND R. HER- NANDEZ-PEON. Effect of crossed-perfusion of the midbrain reticular formation upon sleep. Brain Res. 23: 269-273,197O.
81. ---- -~ DRUCKER-COLfN. R. R.. AND C. W. SPANIS. Neurohu- I I

660 ALEXANDER A. BORBfiLY AND IRENE TOBLER Vohrne 69
moral correlates of sleep: increase of proteins during rapid eye movement sleep. Experientia BaseZ51: 551-552, 1975.
82. DRUCKER-COLfN, R. R., AND C. W. SPANIS. Is there a sleep transmitter? Prog. Neurobiol O& 6: l-22,1976.
83. DRUCKER-COLfN, R. R., C. W. SPANIS, J. HUNYADI, J. F. SASSIN, AND J. L. McGAUGH. Growth hormone effects on sleep and wakefulness in the rat. Neuroerub crie 18: l-8,1975.
84. DRUCKER-COLgN, R. R., M. TUENA DE GOMEZ- PUYOU, M. DEL CARMEN GUTIERREZ, AND G. DREYFUS-CORTES. Immunological approach to the study of neurohumoral sleep factors: effects on REM sleep of antibodies to brain stem proteins. Exp. Neural 69: 563-575,198O.
85. DRUCKER-COLiN, R. R., AND C. VALVERDE. Endo- crine and peptide function in the sleep-wake cycle. In: Sleep: Clinical and Experimental Aspects, edited by D. Ganten and D. Pfaff. Berlin: Springer-Verlag, 1982, p. 37-81.
86. DUGOVIC, C., AND J. ADRIEN. Presence of PS-inducing substances in the hypothalamo-hypophyseal complex. In: Sleep ‘86, edited by W. P. Koella, F. Obil, H. Schulz, and P. Visser. Stuttgart, FRG: Fischer-Verlag, 1988, p. 297-299.
87. DUGOVIC, C., AND A. WAUQUIER. 5-HTs receptors could be primarily involved in the regulation of slow- wave sleep in the rat. Eur. J. PhuwuzoL 137: 145-146, 1987.
88. DZOLJIC, M. R. Prostaglandins and sleep. Awaking ef- fect of prostaglandins and sleep pattern of essential fatty acids deficient (EFAD) rats. Prostaglandins 15: 317-324,1978.
89. EHLERS, C. L., T. K. REED, AND S. J. HENRIKSEN. Effects of corticotropin-releasing factor and growth hormone-releasing factor on sleep and activity in rats. Neuroenduwinologzl42: 467-474,1986.
90. ERNST, A., AND G. A. SCHOENENBERGER. DSIP: basic findings in human beings. In: Sleep Peptides: Basic and Clinical Approaches, edited by S. Inoue and D. Schneider-Helmert. Tokyo: Jpn. Sci. Sot. Press, 1988, p. 131-173.
91. EVERSON, C. A., C. A. KUSHIDA, B. M. BERGMANN, M. A. GILLILAND, J. J. PILCHER, V. S. FANG, AND A. RECHTSCHAFFEN. Recovery from chronic sleep depri- vation in the rat (Abstract). Sleep &a. 16: 521,1987.
92. FARBER, J., J. D. MILLER, K. A. CRAWFORD, AND B. A. McMILLEN. Dopamine metabolism and receptor sensitivity in rat brain after REM sleep deprivation. PhurmacoL Biochem Behuv. 18: 509-513,1983.
93. FARRAR, W. L., J. M. HILL, A. HAREL-BELLAN, AND M. VINOCOUR. The immunological brain. ImmunoL Rev. 100: 361-378,1987.
94. FARRAR, W. L., P. L. KILIAN, M. R. RUFF, J. M. HILL, AND C. B. PERT. Visualization and characterization of interleukin 1 receptors in brain. J. Immunol. 139: 459-463,1987.
95. FEINBERG, I., G. FEIN, AND T. C. FLOYD. EEG pat- terns during and following extended sleep in young adults. Electroencephalogr. C&n. Neurophl/siol. 50: 467-476,198O.
96. FEINBERG, I., J. D. MARCH, T. C. FLOYD, R. JI- MISON, L. BOSSOM-DEMITRACK, AND P. H. KATZ. Homeostatic changes during post-nap sleep maintain baseline levels of delta EEG. Electroencephubgr. Clin, Neurophgsiol 61: 134-137,1985.
97. FELDMAN, S. C., AND A. J. KASTIN. Localization of neurons containing immunoreactive delta sleep-inducing
peptide in the rat brain: an immunocytochemical study. Neuroscience 2: 303-317,1984.
98. FENCL, V., G. KOSKI, AND J. R. PAPPENHEIMER. Factors in cerebrospinal fluid from goats that affect sleep and activity in rats. J. PhvsioL L+ond 216: 565-589, 1971.
99. FERNSTROM, J. D., AND R. H. PASTEL. The neurophar- macology of serotonin and sleep: an evaluation. In: Ani- rnal ModeZs of m&on, edited by G. F. Koob, C. L. Ehlers, and D. J. Kupfer. Boston, MA: Birkhiuser-Ver- lag. In press.
100. FISCHMAN, A. J., A. J. KASTIN, AND M. V. GRAF. Circadian variation of DSIP-like material in rat plasma. Life Sci 35: 2079-2084,1984.
101. FONLUPT, P., M. ROCHE, L. CRONENBERGER, AND H. PACHECO. La S-adenosyl-L-homocysteine. 1. Induc- trite de sommeil. Can. J. Physiol PhwmacoL 58: 160-166, 1980.
102. FORNAL, C., R. MARKUS, AND M. RADULOVACKI. Muramyl dipeptide does not induce slow-wave sleep or fever in rats. Peptides Fa~etteville 5: 91-95,1984.
103. FORSTERMANN, U., R. HELDT, AND G. HERTTING. Effects of intracerebroventricular administration of prostaglandin Dz on behaviour, blood pressure and body temperature as compared to prostaglandins Ez and F&. P&wp~~~ 80: 365-370,1983.
104. FRANCK, G., E. SALMON, R. POIRRIER, B. SADZOT, AND G. FRANCO. Etude du metabolisme glucidique cere- bral regional, chez l’homme, au tours de 1’6veil et du sommeil, par tomographie i emission de positrons. Reu. Electroencephalogr. NeurophysioL Clk 17: 71-77,1987.
105. FRIEDMAN, L., B. M. BERGMANN, AND A. RECHT- SCHAFFEN. Effects of sleep deprivation on sleepiness, sleep intensity, and subsequent sleep in the rat. SZeep 1: 369-391,1979.
106. FUXE, K., L. AGNATI, A. HARFSTRAND, I. ZINI, K. TATEMOTO, E. MERLO PICH, T. HijKFELT, V. MUTT, AND L. TERENIUS. Central administration of neuro- peptide Y induced hypotension bradypnea and EEG syn- chronization in the rat. Acta PhgsioL Scand 118: 189-192, 1983.
107. GAGNON, P., AND J. DE KONINCK. Reappearance of EEG slow waves in extended sleep. Electroencephalogr. C&L Neuroph&oL 58: 155-157,1984.
108. GAILLARD, J.-M. Involvement of noradrenaline in wakefulness and paradoxical sleep. In: Sleep, Neuro- transmitters and Neuromodulators, edited by A. Wau- quier, J.-M. Gaillard, J. M. Monti, and M. Radulovacki. New York: Raven, 1985, p. 57-67.
109. GARCIA-ARRARAS, J. E. Effects of sleep-promoting factor from human urine on sleep cycle of cats. Am. J. PhgsioL 241 (EndumimL Metab. 4): E269-E274,1981.
110. GARCIA-ARRARAS, J. E., AND J. R. PAPPEN- HEIMER. Site of action of sleep-inducing muramyl pep- tide isolated from human urine: microinjection studies in rabbit brains. J. Neuroph@oL 49: 528-533,1983.
111. GARRY, P., B. ROUSSEL, R. COHEN, S. BIOT-LA- PORTE, A. E. CHARFI, M. JOUVET, AND G. SAS- SOLAS. Diurnal administration of human growth hor- mone-releasing factor does not modify sleep and sleep- related growth hormone secretion in normal young men. Acta Endocrinol. 110: 158-163,1985.
112. GESSA, G. L., M. L. PORCEDDU, M. COLLU, G. MEREU, M. SERRA, E. ONGINI, AND G. BIGGIO. Se- dation and sleep induced by high doses of apomorphine after blockade of D-l receptors by SCH 23390. Eur. J. PharmucoL 109: 269-274,1985.
113. GIUDITTA, A., A. NEUGEBAUER-VITALE, G.

GRASSI-ZUCCONI, M. V. AMBROSINI, AND R. SCAR- 130. HERY, F., J.-F. PUJOL, M. LOPEZ, J. MACON, AND J. ONI. Synthesis of brain RNA and DNA during sleep. In: GLOWINSKI. Increased synthesis and utilization of se- Experiwwntal Brain Research, Sleep Mechanisms, edited rotonin in the central nervous system of the rat during by A. A. Borbely and J. L. Valatx. Berlin: Springer-Ver- paradoxical sleep deprivation. Bruin Res. 21: 391-403, lag, 1984, suppl. 8, p. 146-154. 1970.
114. GIUDITTA, A., B. RUTIGLIANO, AND A. VITALE- 131. HESS, W. R. Das Schlafsyndrom als Folge dienzephaler NEUGEBAUER. Influence of synchronized sleep on the Reizung. HeZv. PhgsioL PhurmacoL Actu 2: 305~344,1944. biosynthesis of RNA in two nuclear classes isolated from 132. HESS, W. R. Sleep as a phenomenon of the integral or- rabbit cerebral cortex. J. Neurochem 35: X%9-1266,1980. ganism. Prog. Brain Res. 18: 3-7,1965.
115. GIUDITTA, A., B. RUTIGLIANO, AND A. VITALE- 133. HISHIKAWA, Y., H. CRAMER, AND W. KUHLO. Natu- NEUGEBAUER. Influence of synchronized sleep on the ral and melatonin-induced sleep in young chickens-a biosynthesis of RNA in neuronal and mixed fractions behavioral and electrographic study. Exp. Bruin Res. 7: isolated from rabbit cerebral cortex. J. Neurocti 35: 84-94,1969. 126’7-1272,198O. 134. HOBSON, J. A., R. LYDIC, AND H. A. BAGHDOYAN.
116. GNADT, J. W., AND G. V. PEGRAM. Cholinergic brain- Evolving concepts of sleep cycle generation: from brain stem mechanisms of REM sleep in the rat. Bmin Res. centers to neuronal populations. Behav. Bruin Sci. 9: 384: 29-41,1986. 371-448,1986.
117. GOLDSTEIN, R., M. PETRESCU, AND M. POPA. Vaso- 135. HOLMES, S. W., AND D. SUGDEN. Effects of melatonin tocin and sleep in prepubertal boys: its suppression by on sleep and neurochemistry in the rat. Br. J. PhumnacoL valproic acid. E?zdocrinoZogg 25: 97-109, 1987. 76: 95-101,1982.
118. GOLDSTEIN, R., AND D. PSATTA. Sleep in the cat: 136. HONDA, K., Y. KOMODA, AND S. INOUE. Effects of raphe dorsalis and vasotocin. Sleep 7: 373-379,1984. sleep-promoting substances on the rat circadian sleep-
119. GRAF, M. V., H. CHRISTEN, AND G. A. SCHOENEN- waking cycles. In: Endogenous Sleep Substances and Sleep BERGER. DSIP/DSIP-P and circadian motor activity of Regulation, edited by S. Inou6 and A. A. Borbely. Tokyo: rats under continuous light. Peptides Fayetteville 3: Jpn. Sci. Sot. Press, 1985, p. 203-214. 623-626,1982. 137. HONDA, K., Y. KOMODA, S. NISHIDA, H. NAGA-
120. GRAF, M. V., H. CHRISTEN, H. J. TOBLER, P. F. SAKI, A. HIGASHI, K. UCHIZONO, AND S. INOUE. Ur- MAIER, AND G. A. SCHOENENBERGER. Effects of re- idine as an active component of sleep promoting sub- peated DSIP and DSIP-P administration on the circa- stance: its effects on nocturnal sleep in rats. Neurosci dian locomotor activity of rats. PharmacoL Biochem. Res. 1: 243~252,1984. Behav. 15: 717-721,198l. 138. HONDA, K., K. NAITO, Y. KOMODA, T. KIMURA, I.
121. GRAF, M. V., AND A. J. KASTIN. Delta-sleep-inducing YAMAMOTO, AND S. INOUE. Comparison of sleep-pro- peptide (DSIP): a review. Neurosci. Biobehav. Rev. 8: moting activity of uridine-related substances in unre- 83-93,1984. strained rats (Abstract). Sleep Res. 16: 62,1987.
122. GRAF, M. V., AND A. J. KASTIN. Delta-sleep-inducing 139. HONDA, K., Y. OKANO, Y. KOMODA, AND S. INOUE. peptide (DSIP): an update. Peptides Fuyetteville 7: Sleep-promoting effects of intraperitoneally adminis- 1165-1187,1986. tered uridine in unrestrained rats. Neurosci. Lett. 62:
123. GRAF, M. V., A. J. KASTIN, D. H. COY, AND J. E. ZA- 137-141,1985. DINA. DSIP reduces amphetamine-induced hyperther- 140. HORNE, J. A. Tissue restitution and sleep, with particu- mia in mice. PhysioL Behuv. 331291~295,1984. lar reference to human slow wave sleep. In: Endogenous
124. GRAF, M. V., A. J. KASTIN, AND A. J. FISCHMAN. The Sleep Substunces and Sleep Regulation, edited by S. Inoue natural occurrence and some extra-sleep effects of DSIP. and A. A. Borb&ly. Tokyo: Jpn. Sci. Sot. Press, 1985, vol. 8, In: Endogenous Sleep Substances and Sleep Regulation, p. 25-40. edited by S. Inoue and A. A. Borb&ly. Tokyo: Jpn. Sci. Sot. 141 HORNE, J. A., AND V. J. MOORE. Sleep EEG effects of Press, 1985, p. 155-165. exercise with and without additional body cooling. EZec-
125 . GRAHNSTEDT, S., AND R. URSIN. Platform sleep de- troencephuZogr. CZin Neurophy.@oL 60: 33-38,1985. privation affects deep slow wave sleep in addition to 142 HORNE, J. A., AND A. J. REID. Night-time sleep EEG REM sleep. Behuv. Bruin Res. 18: 233-239,1985. changes following body heating in a warm bath. EZec-
126 . GULEVICH, G., W. DEMENT, AND L. JOHNSON. Psy- troencephaZogr. CZin. Neuroph@oL 601 154-157,1985. / chiatric and EEG observations on a case of prolonged 143 IDZIKOWSKI, C., P. J. COWEN, D. NUTT, AND F. J. (264 h) wakefulness. Arch. Gen. Psgchiatrg 15: 29-35, MILLS. The effects of chronic ritanserin treatment on 1966. HANAGASIOGLU, M., AND A. A. BORBELY. Effect of
sleep and the neuroendocrine response to L-tryptophan. 127 .
voluntary locomotor activity on sleep in the rat. Behuv. Ps&wpharma&ogg 93: 416-420,1987.
144 Brain Res. 4: 359-368,1982.
IDZIKOWSKI, C., F. J. MILLS, AND R. GLENNARD.
128 . HAULCA, I., L. ARABEI, D. BRANISTEANU, AND F. 5-Hydroxytryptamine-2 antagonist increases human
/ TOPOLICEANU. Preliminary data on the possible hyp- slow wave sleep. Brain Res. 378: 164-168,1986.
nogenic role of adenosine. J. Neurocti 21: 1019-1020, 145 INOUE, S. Sleep substances: their roles and evolution.
1973. In: Endogenous Sleep Substances and SZeep Regulation,
128aHAYAISH1,O. Sleep-wake regulation by prostaglandins edited by S. Inoue and A. A. Borbely. Tokyo: Jpn. Sci. Sot.
Dt and I&. J. BioL Chem. 263: 14593-14596,1988. Press, 1985, vol. 8, p. 3-12.
129. HAYAISHI, O., R. UENO, H. ONOE, H. OSAMA, I. FU- 146. INOUE, S. Sleep and sleep substances. Brain Dev. 8:
JITA, H. NISHINO, AND Y. OOMURA. Prostaglandin Dz 469-473,1986.
induces sleep when infused into the cerebral ventricle of 147. INOUE, S. Multifactorial humoral regulation of sleep.
conscious monkeys. In: Advances in Prostaglandin, CZin NeurophurmucoL SuppL 9: 470-472,1986.
Thrornboxane, and Leukotriene Research, edited by B. 147aINOUE, S. Biologzl of SZeep Substances. Boca Raton, FL: Samuelson, R. Paoletti, and P. W. Ramwell. New York: CRC. In press. Raven, 1987, vol. 17, p. 946-948. 148. INOUE, S., AND A. A. BORBELY (Editors). Endogerwus
April 1989 ENDOGENOUS SLEEP-PROMOTING SUBSTANCES 661

662 ALEXANDER A. BORBfiLY AND IRENE TOBLER vohwq 69
Sleep Substances and Sleep Regulation Tokyo: Jpn. Sci. Sot. Press, 1985.
149. INOUfi, S., K. HONDA, AND Y. KOMODA. A possible mechanism by which the sleep-promoting substance in- duces slow wave sleep but suppresses paradoxical sleep in the rat. In: Sleep ‘82, edited by W. P. Koella. Basel: Karger, 1983, p. 112-124.
165.
166.
150. INOUE, S., K. HONDA, AND Y. KOMODA. Uridine as an active component of the sleep-promoting substance. In: Sleep ‘84, edited by W. P. Koella, E. Rtither, and H. Schulz. Stuttgart, FRG: Gustav Fischer-Verlag, 1985, p. 212-214.
167
151. INOUE, S., K. HONDA, AND Y. KOMODA. Sleep-pro- moting substances. In: Sleep: Neurotransmitters and Neuromodulators, edited by A. Wauquier, J.-M. Gaillard, J. M. Monti, and M. Radulovacki. New York: Raven, 1985, p. 305-318.
168
152. INOUE, S., K. HONDA, Y. KOMODA, K. UCHIZONO, AND R. UENO. Differential sleep-promoting effects of five sleep substances nocturnally infused in unrestrained rats. Proc. Nat1 Acud Sci. USA 81: 6240-6244,1984.
153. INOUE, S., K. HONDA, Y. KOMODA, K. UCHIZONO, R. UENO, AND 0. HAYAISHI. Little sleep-promoting effect of three sleep substances diurnally infused in unre- strained rats. ,Neurosci. Lett. 49: 207-211,1984.
- 154. INOUE, S., K. HONDA, Y. OKANO, AND Y. KOMODA. Behavior-modulating effects of uridine in the rhinoceros beetle. Zool. ti 3: 727-729,1986.
169
170.
171.
and paradoxical sleep rebound. In: SZeep ‘82, edited by W. P. Koella. Basel: Karger, 1983; p. 2-18. JOUVET, M. Neuromediateurs et facteurs hypnogGnes. Rev. Neural Paris 140: 389-400,1984. JOUVET, M. Indoleamines and sleep-inducing factors. In: Experimental Brain Research. Sleep Mechanisms, edited by A. A. Borbely and J. L. Valatx. Berlin: Springer-Verlag, 1984, suppl. 8, p. 81-94. JOUVET, M. Sleep factor(s)-reality or fiction. In: SZeep ‘86, edited by W. P. Koella, F. Obil, H. Schulz, and P.
Visser. Stuttgart, FRG: Gustav Fischer-Verlag, 1988, p. 161-164. JOUVET, M., C. BUDA, R. CESPUGLIO, N. CHAS- TRETTE, M. DENOYER, M. SALLANON, AND J. P. SASTRE. Hypnogenic effects of some hypothalamo-pitu- itary peptides. In: Clinical Neurophwmacology, edited by W. E. Bunney, Jr., E. Costa, and S. G. Potkin. New York: Raven, 1986, vol. 9, suppl. 4, p. 465-467. JOUVET, M., J.-P. SASTRE, C. BUDA, M. JANIN, K. KITAHAMA, AND M. SALLANON. La pars intermedia de I’hypophyse est-elle responsable du declenchement du sommeil paradoxal chez le chat pontique avec “hypo- physe isolee”? C! R. Acad Sci. Paris 299: 883-888, 1984. KADLECOVA, O., AND K. MASEK. Muramyl dipeptide and sleep in rat. Methods Find Exp. Clin. Phurmacol 8: ill-115,1986. KAESER, H. E. A clinical trial with DSIP. Eur. NeuroL 23: 386-388,1984.
155. INOUE, S., M. KIMURA, K. HONDA, AND Y. KOMODA. Sleep peptides: general and comparative aspects. In: Sleep Peptides Basic and Clinical Approaches, edited by S. Ino& and D. Schneider-Helmert. Tokyo: Jpn. Sci. Sot. Press, 1988, p. l-26.
156. INOUE, S., AND D. SCHNEIDER-HELMERT (Editors). Sleep Peptides: Basic and Clinical Approaches. Tokyo: Jpn. Sci. Sot. Press, 1988.
157. INOUE, S., K. UCHIZONO, AND H. NAGASAKI. Endog- enous sleep-promoting factors. Trends Neurosci. 5: 218-220,1982.
172. KAFI, S., M. MONNIER, AND J.-M. GAILLARD. The delta-sleep inducing peptide (DSIP) increases duration of sleep in rats. Neurosci Lett. 13: 169-172, 1979.
173. KAPAS, L., F. OBAL, JR., P. ALFOLDI, G. RUBICSEK, B. PENKE, AND F. OBAL. Effects of nocturnal intraperi- toneal administration of cholecystokinin in rats: simul- taneous increase in sleep, increase in EEG slow-wave activity, reduction of motor activity, suppression of eat- ing, and decrease in brain temperature. Brain Res. 438: 155-x4,1988.
158. ISHIMORI, K. True cause of sleep-a hypnogenic sub- stance as evidenced in the brain of sleep-deprived ani- mals. Tokyo Igaku Zasshi 23: 423-457,1909.
158a.IYER, K. S., G. A. MARKS, A. J. KASTIN, AND S. M. MCCANN. Evidence for a role of delta sleep-inducing peptide in slow-wave sleep and sleep-related growth hormone release in the rat. Proc NatL Acad Sci USA 85: 3653-3656,1988.
174. KARACAN, I., W. W. FINLEY, R. L. WILLIAMS, AND C. J. HURSCH. Changes in stage l-REM and stage 4 sleep during naps. BioL Psychiatv 2: 261-265,197O.
175. KARACAN, I., R. L. WILLIAMS, W. W. FINLEY, AND C. J. HURSCH. The effects of naps on nocturnal sleep: influence on the need for stage l-REM and stage 4 sleep. BioL Psychiatry 2: 391-399,197O.
159. JAMES, S. P., W. B. MENDELSON, D. A. SACK, N. E. ROSENTHAL, AND T. A. WEHR. The effect of melatonin on normal sleep. NeuropsychopharmacoZogy 1: 41-44, 1987.
160. JIMERSON, D. C., M. D. HARRY, H. D. LYNCH, R. M. POST, R. J. WURTMAN, AND W. E. BUNNEY. Urinary melatonin rhythms during sleep deprivation in de- pressed patients and normals. L$e Sti 20: 1501-1508, 1977.
176. KARMANOVA, I. G., V. F. MAKSIMUK, I. B. VOR- ONOV, M. M. BOGOSLOVSKII, N. N. DEMIN, N. L. RU- BINSKAYA, AND S. V. AL’BERTIN. Analysis of the ac- tion of the neuropeptide-inducing delta-sleep in cats and white rats. Neurosci. Behav. PhysioL 13: 476-481, 1983.
177. KARNOVSKY, M. L. Muramyl peptides in mammalian tissues and their effects at the cellular level. Federation Proc. 45: 2556-2560,1986.
161. JOUVET, D., P. VIMONT, F. DELORME, AND M. JOU- VET. Etude de la privation sblective de la phase para- doxale de sommeil chez le chat. C R. Sot. BioL Lyon 158: 756-759,1964.
178. KARNOVSKY, M. L., AND P. REICH. Biochemistry of sleep. In: Advance in Neurochemistry, edited by B. W. Agranoff and M. H. Aprison. New York: Plenum, 1977, vol. 2, p. 213-275.
162. JOUVET, M. The role of monoamines and acetylcholine containing neurons in the regulation of the sleep-waking cycle. Ergeb. PhysioL BioL Chem Exp. PharmakoL 64: 166-30’7,1972.
179. KARNOVSKY, M. L., P. REICH, J. M. ANCHORS, AND B. L. BURROWS. Changes in brain glycogen during slow-wave sleep in the rat. J. Neurochem 41: 1498-1501, 1983.
180. KASTIN, A. J., G. A. OLSON, A. V. SCHALLY, AND D. H. COY. DSIP-more than a sleep peptide? Trends Neurosci. 3: 163-165,198O.
163. JOUVET, M. Neuropharmacology of the sleep-waking 181. KATO, N., S. NAGAKI, Y. TAKAHASHI, I. NAMURA, cycle. Handb. PsycmharmucoL 8: 233-293,1977. AND Y. SAITO. DSIP-like material in rat brain, human
164. JOUVET, M. Hypnogenic indolamine-dependent factors cerebrospinal fluid, and plasma as determined by en-

April 1989 ENDOGENOUS SLEEP-PROMOTING SUBSTANCES 663
zyme immunoassay. In: Endogenous Sleep Substances and Sleep Regulation, edited by S. Inot& and A. A. Bortily. Tokyo: Jpn. Sci. Sot. Press, 1985, p. 141-153.
182. KAWAKAMI, M., F. KIMURA, AND C.-W. TSAI. Corre- lation of growth hormone secretion to sleep in the im- mature rat. J. Ph&ol Land 339: 325-337,1983.
183. KIMURA, M., K. HONDA, Y. KOMODA, AND S. INOUfi. Interacting sleep-modulatory effects of simultaneously administered delta-sleep-inducing peptide, muramyl di- peptide and uridine in unrestrained rats. Neurosti Res. 5: 157-166,1987.
184.
183a.KIMURA, M., AND S. INOUE. The phosphorylated ana- logue of DSIP enhances slow wave sleep and paradoxical sleep in unrestrained rats. Psychopharmacology. In press. KITAHAMA, K., AND J. L. VALATX. Instrumental and pharmacological paradoxical sleep deprivation in mice: strain differences. Neurophurmaco+ 19: 529-535,198O. KIYONO, S., T. KAWAMOTO, H. SAKAKURA, AND K. IWAMA. Effects of sleep deprivation upon the paradoxi- cal phase of sleep in cats. Electroencephdogr. Clin Neu- roph~sd 19: 34-40,1965. KOELLA, W. P. Polypeptides and sleep. Trends Phur- mu.coL s& 4: 21Q-211,1983.
185.
186.
18’7
188.
189.
190.
191.
192.
193.
194.
195.
196.
KOMODA, Y., M. ISHIKAWA, H. NAGASAKI, M. IRIKI, K. HONDA, S. INOUE, A. HIGASHI, AND K. UCHIZONO. Uridine, a sleep-promoting substance from brainstems of sleep-deprived rats. Biomed Res. SuppL 4: 223-227,1983.
KORNMULLER, A. E., H. D. LUX, K. WINKEL, AND M. KLEE. Neurohumoral ausgeliiste Schlafzust%nde an Tieren mit gekreuztem Kreislauf unter der Kontrolle von EEG-Ableitungen. Naturwissenschafien 14: 503-505, 1961. KOVALZON, V. Humoral regulation of sleep. In: Issues in Science and Technob, Human and Animal Ph@ol- ogll Sleep Mechanisms, edited by 0. N. Barsova. Moscow: VINITI, 1986, vol. 31, p. 3-58. (In Russian.) KOVALZON, V., V. N. KALIKHEVICH, F. OBAL, JR., AND H. KLEINLOGEL. Hypnogenic effect of the peptide l-D-Trp-DSIP in rabbits and rats. In: Sleep %..& edited by W. P. Koella, E. Ruther, and H. Schulz. Stuttgart, FRG: Gustav Fischer-Verlag, 1985, p. 218-219. KOVALZON, V. M., F. OBAL, JR., AND V. N. KALIKHE- VICH. The peptidergic modulation of sleep: comparative investigation of analogs of the peptide DSIP. Zh, Ed Biokhim FizioL 22: 483-488,1986. KOVALZON, V. M., AND V. L. TSIBULSKII. Does syn- thetic delta sleep-inducing peptide possess hypnogenic properties? Neurosci Behav. Ph&oL 13: 243~244,1983. KOVALZON, V. M., AND V. L. TSIBULSKII. REM-sleep deprivation, stress and emotional behavior in rats. Behav. Brain Res. 14: 235-245,1984. KRUEGER, J. M. Muramyl peptides and interleukin-1 as promoters of slow wave sleep. In: Endog- Sleep Sub stances and Sleep Regulation, edited by S. Ino& and A. A. Borbbly. Tokyo: Jpn. Sci. Sot. Press, 1985, vol. 8, p. 181-192. KRUEGER, J. M., J. BACSIK, AND J. E. GARCIA-AR- RARAS. Sleep-promoting material from human urine and its relation to factor S from brain. Am. J. Ph&oL 238 (EndocrinoL Met&. 1): E116-E123,1980. KRUEGER, J. M., D. DAVENNE, J. WALTER, S. SHO- HAM, S. L. KUBILLUS, R. S. ROSENTHAL, S. A. MAR- TIN, AND K. BIEMANN. Bacterial peptidoglycans as modulators of sleep. II. Effects of muramyl peptides on
197.
198.
199.
200.
201.
202.
203.
204.
205.
206
207
208.
209.
210.
211.
the structure of rabbit sleep. Brain Res. 403: 258-266, 1987. KRUEGER, J. M., C. A. DINARELLO, S. SHOHAM, D. DAVENNE, J. WALTER, AND S. KUBILLUS. Interferon alpha-2 enhances slow-wave sleep in rabbits. Int. J. Im- munopharmad 9: 23-30,1987. KRUEGER, J. M., AND M. L. KARNOVSKY. Sleep and the immune response. Ann. NY Ad Sci. 496: 510-516, 1987. KRUEGER, J. M., M. L. KARNOVSKY, S. A. MARTIN, J. R. PAPPENHEIMER, J. WALTER, AND K. BIE- MANN. Peptidoglycans as promoters of slow-wave sleep. II. Somnogenic and pyrogenic activities of some natu- rally occurring muramyl peptides; correlations with mass spectrometric structure determination. J. BioL Chem 259: 12659-12662,1984. KRUEGER, J. M., S. KUBILLUS, S. SHOHAM, AND D. DAVENNE. Enhancement of slow wave sleep by endo- toxin and lipid. Am J PhysioL 251 (Regduby Integra- tive Comp. PhgsioL 20): R591-R597,1986. KRUEGER, J. M., F. OBAL, JR., L. JOHANNSEN, A. B. CADY, AND L. TOTH. Endogenous slow-wave sleep sub- stances: a review. In: Slow Wave Sleep: Ph&ological, Pathuph~siologicd and Function& Aspects, edited by A. Wauquier and C. Dugovic. New York: Raven. In press. KRUEGER, J. M., J. R. PAPPENHEIMER, AND M. L. KARNOVSKY. Sleep-promoting factor S: purification and properties. Proc NatL Ad Sci USA 75: 5235-5238, 1978. KRUEGER, J. M., J. R. PAPPENHEIMER, AND M. L. KARNOVSKY. The composition of sleep-promoting fac- tor isolated from human urine. J. BioL Chem. 257: 1664-1669,1982. KRUEGER, J. M., J. R. PAPPENHEIMER, AND M. L. KARNOVSKY. Sleep-promoting effects of muramyl peptides. Proc. NatL Ad S+i USA 79: 6102-6106,1982. KRUEGER, J. M., R. S. ROSENTHAL, S. A. MARTIN, J. WALTER, D. DAVENNE, S. SHOHAM, S. L. KU- BILLUS, AND K. BIEMANN. Bacterial peptidoglycans as modulators of sleep. I. Anhydro forms of muramyl pep- tides enhance somnogenic potency. Brain Res. 403: 249-257,1987. KRUEGER, J. M., AND S. SHOHAM. Homologies be- tween sleep factors and hibernation triggers. In: &v&g in the Cold, edited by H. C. Heller, X. T. Musacchia, and L. C. H. Wang. New York: Elsevier, 1986, p. 235-243. KRUEGER, J. M., L. A. TOTH, A. B. CADY, L. JO- HANNSEN, AND F. OBAL, JR. Immunomodulation and sleep. In: Sleep Peptides: Basic and Clinical Approaches, edited by S. Inou& and D. Schneider-Helmert. Tokyo: Jpn. Sci. Sot. Press, 1988, p. 95-129. KRUEGER, J. M., J. WALTER, C. A. DINARELLO, S. M. WOLFF, AND L. CHEDID. Sleep-promoting effects of endogenous pyrogen (interleukin-1). Am J. Ph&oL 246 (Regdatxxq Integrative Camp. PhysioL 15): R994-R999, 1984. KRUEGER, J. M., J. WALTER, M. L. KARNOVSKY, L. CHEDID, J. P. CHOAY, P. LEFRANCIER, AND E. LE- DERER. Muramyl peptides. Variation of somnogenic ac- tivity with structure. J. Exp. Med 159: 68-76,1984. KRUEGER, J. M., J. WALTER, AND C. LEVIN. Factor S and related somnogens: an immune theory for slow-wave sleep. In: Brain Mechanisms of Sleep, edited by D. J. McGinty, R. Drucker-Colin, A. Morrison, and P.-L. Par- meggiani. New York: Raven, 1985, p. 253-275. KRUISBRINK, J., M. MIRMIRAN, T. P. VAN DER WOUDE. AND G. J. BOER. Effects of enhanced cerebro-

664 ALEXANDER A. BORBELY AND IRENE TOBLER Vohme 69
spinal fluid levels of vasopressin, vasopressin antagonist or vasoactive intestinal polypeptide on circadian sleep- wake rhythm in the rat. Bruin Res. 419: 76-86,1987.
212. KTONAS, P. Y. Editorial comment: period-amplitude EEG analysis. SZeep 10: 505~507,1987.
213. KUWABARA, N., K. SEKI, AND K. AOKI. Circadian, sleep and brain temperature rhythms in cats under sus- tained daily light-dark cycles and constant darkness. Phl/sioL Behav. 38: 283-289,1986.
213a.LAHMEYER, H. W. Sleep in craniopagus twins. SZeep 11: 301-306,1988.
214. LEGENDRE, R., AND H. PIERON. Le problGme des fac- teurs du sommeil. Resultats d’injections vasculaires et intra-c&%brales de liquides insomniques. C. R. Hebd. Seance-s Mem, Sot. Bid 62: 1077-1079,191O.
215. LEGENDRE, R., AND H. PIERON. Recherches sur le be- soin de sommeil cons&utif B une veille prolong&e. 2. A&. PhysioL 14: 235-262,1913.
216. LENARD, H. G., AND F. J. SCHULTE. Polygraphic sleep study in craniopagus twins. (Where is the sleep trans- mitter?) J. NeuroL Neurosurg. Psychiatry 35: 756-762, 1972.
217. LEVITT, R. A. Sleep deprivation in the rat. Science Wash, DC 153: 85-87,1966.
218. LIEBERMAN, H. R., F. WALDHAUSER, G. GAR- FIELD, H. J. LYNCH, AND R. J. WURTMAN. Effects of melatonin on human mood and performance. Bruin Res. 323: 201-207,1’984.
219. LISK, R. D., AND C. H. SAWYER. Induction of paradoxi- cal sleep by lights-off stimulation. Proc Sot Exp. Biol Med 123: 664-667,1966.
219a.LOUIS-COINDET, J., AND C. DEBRU. Increase of para- doxical sleep episodes after electrical stimulation of the lateral and third ventricles in the rat. Neurosci Lett. 91: 165-171,1988.
220. LUCAS, E. A., AND M. B. STERMAN. The polycyclic sleep-wake cycle in the cat: effects produced by sensori- motor rhythm conditioning. Exp. Neural. 42: 347-368, 1974.
221. LUE, F. A., M. BAIL, R. GORCZYNSKI, AND H. MOL- DOFSKY. Sleep and interleukin-l-like activity in cat ce- rebrospinal fluid (Abstract). Sleep Res. 16: 51,1987.
222. LYNCH, H. J., D. C. JIMERSON, Y. OZAKI, R. M. POST, W. E. BUNNEY, AND R. J. WURTMAN. Entrainment of rhythmic melatonin secretion in man to a 12-hour phase shift in the light/dark cycle. Life Sci 23: 1557-1564,1978.
223. MARCH, C. J., B. MOSLEY, A. LARSEN, D. P. CER- RETTI, G. BRAEDT, V. PRICE, S. GILLIS, C. S. HEN- NEY, S. R. KRONHEIM, K. GRABSTEIN, P. J. CON- LON, T. P. HOPP, AND D. COSMAN. Cloning, sequence and expression of two distinct human interleukin-1 com- plementary DNAs. Nature Land 315: 641~647,1985.
224. MARCZYNSKI, T. J., AND L. L. BURNS. Reward contin- gent positive variation (RCPV) and postreinforcement EEG synchronization (PRS) in the cat: physiological aspects, the effect of morphine and LSD-25, and a new interpretation of cholinergic mechanisms. Gen. Phurmu- coL 7: 211~220,1976.
225. MARCZYNSKI, T. J., N. YAMAGUCHI, G. M. LING, AND L. GRODZINSKA. Sleep induced by the administra- tion of melatonin (5-methoxy-N-acetyltryptamine) to the hypothalamus in unrestrained cats. Experientia Base,? 20: 435-437,1964.
226. MARTIN, S. A., M. L. KARNOVSKY, J. M. KRUEGER, J. R. PAPPENHEIMER, AND K. BIEMANN. Peptidogly- cans as promoters of slow-wave sleep. I. Structure of the sleep-promoting factor isolated from human urine. J. Biol Chem. 259: 12652-12658,1984.
227. MASEK, K., 0. KADLECOVA, AND N. PijSCHLOVA. Effect of intracisternal administration of prostaglandin El on waking and sleep in the rat. Neurophurmucobg~ 15: 491-494,1976.
228. MATSUMOTO, J., K. SOGABE, AND Y. HORI-SAN- TIAGO. Sleep in parabiosis. Experientia Basel 28: 1043-1044,1972.
229. MATSUMURA, H., Y. GOH, R. UENO, T. SAKAI, AND 0. HAYAISHI. Awaking effect of PGEz microinjected into the preoptic area of rats. Brain Res. 444: 265-272,1988.
230. MENDELSON, W. B. Studies of human growth hormone secretion in sleep and waking. Int. Rev. Neurobiol. 23: 367-389,1982.
231. MENDELSON, W. B., J. C. GILLIN, S. D. DAWSON, A. J. LEWY, AND R. J. WYATT. Effects of melatonin and propranolol on sleep of the rat. Bruin Res. 201: 240244, 1980.
232. MENDELSON, W. B., J. C. GILLIN, G. PISNER, AND R. J. WYATT. Arginine vasotocin and sleep in the rat. Brain Res. 182: 246-249,198O.
233. MENDELSON, W. B., J. C. GILLIN, AND R. J. WYATT. Studies with the delta sleep-inducing peptide in the rat (Abstract). Sleep Res. 9: 55,198O.
234. MENDELSON, W. B., J. C. GILLIN, AND R. J. WYATT. The search for circulating sleep-promoting factors. In: Advances in Pharmacology & Therapeutics. II. CNS Phurmacolog~-Neuropeptides, edited by H. Yoshida, Y. Hagihara, and S. Ebashi. Oxford, UK: Pergamon, 1982, vol. 1, p. 227-240.
235. MENDELSON, W. B., R. D. GUTHRIE, C. FREDERICK, AND R. J. WYATT. The flower pot technique of rapid eye movement (REM) sleep deprivation. PharmacoL Bio- them Behuv. 2: 553-556,1974.
236. MENDELSON, W. B., L. S. JACOBS, J. C. GILLIN, AND R. J. WYATT. The regulation of insulin-induced and sleep-related human growth hormone secretion: a re- view. Ps@wneuroendwrinolog~ 4: 341~349,1979.
237. MENDELSON, W. B., A. KURUVILLA, T. WATLING- TON, K. GOEHL, S. M. PAUL, AND P. SKOLNICK. Seda- tive and electrographic actions of erythro-9-(2-hydroxy- 3-nonyl)-adenine (EHNA): relationship to inhibition of brain adenosine deaminase. Ps~chopharmucoZogy 79: 126-129,1983.
238. MENDELSON, W. B., J. V. MARTIN, M. PERLIS, R. WAGNER, M. D. MAJEWSKA, AND S. M. PAUL. Sleep induction by an adrenal steroid in the rat. Psychophar- mucolog~ 93: 226-229,1987.
239. MENDELSON, W. B., S. SLATER, P. GOLD, AND J. C. GILLIN. The effect of growth hormone administration on human sleep: a dose-response study. BioL Psychiatry 15: 613-618,198O.
240. MENDELSON, W. B., R. J. WYATT, AND J. C. GILLIN. Whither the sleep factors? In: Sleep Disorders: Basic and Clinical Research, edited by M. H. Chase and E. D. We&man. Jamaica, NY: Spectrum, 1983, p. 281-305.
241. MILLER, L. H., A. J. KASTIN, M. HAYES, A. STERSTE, J. GARCIA, AND D. H. COY. Inverse relationship be- tween onset and duration of EEG effects of six peripher- ally administered peptides. PhurmucoL B&hem, Behuv. 15: 845~848,198l.
242. MILLER, L. H., B. A. TURNBULL, A. J. KASTIN, AND D. H. COY. Sleep-wave activity of a delta sleep-inducing peptide analog correlates with its penetrance of the blood-brain barrier. Sleep 9: 80-84,1986.
243. MISTLBERGER, R., B. BERGMANN, AND A. RECHT- SCHAFFEN. Period-amplitude analysis of rat electroen- cephalogram: effects of sleep deprivation and exercise. Sleep 10: 508-522,1987.

April 1989 ENDOGENOUS SLEEP-PROMOTING SUBSTANCES 665
244.
245.
246.
247.
248.
249.
250
251
252
253
254
.
.
,
,
.
.
,
I
. MONNIER, M., T. KOLLER, AND S. GRABER. Humoral influences of induced sleep and arousal upon electrical brain activity of animals with crossed circulation. Exp. Neural. 8: 264-277,1963.
255. MONTI, J. M., J. DEBELLIS, P. ALTERWAIN, T. PEL- LEJERO, AND D. MONTI. Study of delta sleep-inducing peptide efficacy in improving sleep on short-term admin- istration to chronic insomniacs. Int. J Clin PharmacoL Res. 7: 105-110,1987.
MISTLBERGER, R. E., B. M. BERGMANN, W. WAL- DENAR, AND A. RECHTSCHAFFEN. Recovery sleep following sleep deprivation in intact and suprachiasma- tic nuclei lesioned rats. SZeep 6: 217~233,1983. MITSUGI, N., AND F. KIMURA. Simultaneous determi- nation of blood levels of corticosterone and growth hor- mone in the male rat: relation to sleep-wakefulness cycle. Neuroendocm~ 41: 125-130,1985. MOGILNICKA, E., S. ARBILLA, H. DEPOORTERE, AND S. Z. LANGER. Rapid-eye-movement sleep depriva- tion decreases the density of 8H-dihydroalprenolol and *H-imipramine binding sites in the rat cerebral cortex. Eur. J. PharmacoL 65: 289-292,198O. MOGILNICKA, E., AND A. PILC. Rapid-eye-movement sleep deprivation inhibits clonidine-induced sedation in rats. Eur. J. PharmacoL 71: 123-126,198l. MOLDOFSKY, H., F. A. LUE, J. EISEN, E. KEYSTONE, AND R. MI GORCZYNSKI. The relationship of interleu- kin-l and immune functions to sleep in humans. Psych mm. Med 48: 309-318,1986. MONNIER, M., L. DUDLER, R. GACHTER, AND G. A. SCHOENENBERGER. Humoral transmission of sleep. IX. Activity and concentration of the sleep peptide delta in cerebral and systemic blood fractions. Pfluegers Arch, 360:225-242,19'75. MONNIER, M., L. DUDLER, R. GACHTER, AND G. A. SCHOENENBERGER. Delta sleep-inducing peptide (DSIP): EEG and motor activity in rabbits following in- travenous administration. Neurosci L&t. 6: g-13,1977. MONNIER, M., AND A. M. HATT. Humoral transmission of sleep. V. New evidence from production of pure hemo- dyalisate. PcRuegers Arch. 329: 231~243,197l. MONNIER, M., AND L. HOSLI. Dialysis of sleep and waking factors in blood of the rabbit. science Wash. DC 146: 796-798,1964. MONNIER, M., AND L. HOSLI. Humoral transmission of sleep and wakefulness. II. Hemodialysis of a sleep induc- ing humor during stimulation of the thalamic somno- genie area. P#uegers Arch, 282: 60-75,1965.
256. MOORE-EDE, M. C., F. M. SULZMAN, AND C. A. FULLER. The Cbcks That Time Us, Ph@ob of the Circadian Timing System. Cambridge, MA: Harvard Univ. Press, 1982.
257. MUKHAMETOV, L. M. Unihemispheric slow wave sleep in the brain of dolphins and seals. In: Endogenous Sleep Substances and Sleep Regulation, edited by S. Inoue and A. A. Borbely. Tokyo: Jpn. Sci. Sot. Press, 1985, vol. 8, p. 67-75.
258. MUKHAMETOV, L. M. Unihemispheric slow-wave sleep in the amazonian dolphin, Inia geoflrensis. Neurosci. L&t. 79: 128-132,1987.
259. NAGAKI, S., AND N. KATO. Delta sleep-inducing pep- tide-like material in rat brain as determined by enzyme immunoassay: effect of sleep deprivation. New-ok. I&t. 51: 253-257,1984.
260. NAGASAKI, H., .M. IRIKI, S. INOUE, AND K. UCHI- ZONO. The presence of a sleep-promoting material in the brain of sleep-deprived rats. Proc. Jpn Acad 50: 241-246, 1974.
261. NAGASAKI, H., K. KITAHAMA, J.-L. VALATX, AND M. JOUVET. Sleep-promoting effect of the sleep-pro- moting substance (SPS) and delta sleep-inducing peptide (DSIP) in the mouse. Brain Res. 192: 276~280,198O.
26la.NAKAGAK1, K., S. EBIHARA, S. USUI, Y. HONDA,
262.
263.
264.
265.
266
267
268
AND Y. TAKAHASHI. Neurosti Lett. 91: 160-164,1988. NAKAMURA, R. K., C. KENNEDY, J. C. GILLIN, S. SUDA, M. ITO, F. I. STORCH, W. B. MENDELSON, L. SOKOLOFF, AND M. MISHKIN. Hypnogenic center theory of sleep: no support from metabolic mapping in monkeys. Brain Res. 268: 372-376,1983. NISTICO, G., G. B. DE SARRO, G. BAGETTA, AND E. E. MUELLER. Behavioural and electrocortical spectrum power effects of growth hormone releasing factor in rats. Neuropharmaco~ 26: 75-78,1987. NORMANTON, J. R., AND J. P. GENT. Comparison of the effects of two “sleep” peptides, delta sleep-inducing pep- tide and arginine-vasotocin, on single neurons in the rat and rabbit brain stem. Neuros~ 8: 107-114,1983. OBAL, F., JR. Effects of peptides (DSIP, DSIP analogues, VIP, GRF and CCK) on sleep in the rat. In: Clinical Neu- ropharmaco~, edited by W. E. Bunney, Jr., E. Costa, and S. G. Potkin. New York: Raven, 1986, vol. 9, suppl. 4, p. 459-461. OBAL, F., JR., P. ALFOLDI, G. BENEDEK, AND F. OBAL. Possible contribution of sleep to heat defence. In: Sleep '86, edited by W. P. Koella, F. Obbl, H. Schulz, and P. Visser. Stuttgart, FRG: Gustav Fischer-Verlag, 1988, p. 58-61. OBAL, F., JR., P. ALFOLDI, A. B. CADY, L. JOHANN- SEN, G. SARY, AND J. M. KRUEGER. Growth hor- mone-releasing factor enhances sleep in V.&s and rabbits. Am J PhgsioL 255 (ReguLatory Inte,rative Camp. Phy- siol. 24): R310-R316,1988. OBAL, F., JR., V. M. KOVALZO’,, V. N. KALIKHEVICH, A. TijRijK, P. ALFijLDI, G. &RY, M. HAJOS, AND B. PENKE. Structure-activity relationship in the effects of delta-sleep-inducing peptide (DSIP) on rat sleep. Phar- macoL Biochem Behav. 24: 889-894,1986.
269 . ( OBAL, F., JR., G. SARY, P. ALFOLDI, G. RUBICSEK, AND F. OBAL. Vasoactive intestinal polypeptide pro- motes sleep without effects on brain temperature in rats at night. Neurosci. L&t. 64: 236~240,1986.
270.
271.
272.
OBAL, F., JR., I. TOBLER, AND A. A. BORBELY. Effect of ambient temperature on the 24-hour sleep-wake cycle in normal and capsaicin-treated rats. PhysioL Behav. 30: 425-430,1983. OBAL, F., JR., A. TOROK, P. ALFOLDI, G. SARY, M. HAJOS, AND B. PENKE. Effects of intracerebroventric- ular injection of delta sleep-inducing peptide (DSIP) and an analogue on sleep and brain temperature in rats at night. PharmacoL Biochem Behav. 23: 953-957,1985. OGASAHARA, S., Y. TAGUCHI, AND H. WADA.
Changes in serotonin in rat brain during slow-wave sleep and paradoxical sleep: application of the microwave fix- ation method to sleep research. Brain Res 189: 570-575, 1980.
273. OKUDAIRA, N., H. KANEMOTO, H. KOIKE, K. SUEN- AGA, Y. IGUCHI, AND S. TORII. Butoctamide hydrogen succinate (BAHS) and human delta wave (Abstract). SleepRes 16:116,1987.
274. OKUDAIRA, N., H. KOIKE, S. INUBUSHI, A. ABE, M. YAMANE, I. YANAGISAWA, AND S. TORII. Organic bromine substance: an endogenous REM sleep modula- tor? In: Endogenous Sleep Substances and Sleep ReguLa- tion, edited by S. Inouk and A. A. Borb&ly. Tokyo: Jpn. Sci. Sot. Press, 1985, p. 237248.
275. OKUDAIRA, N., S. TORII, AND S. ENDO. The effect of

666 ALEXANDER A. BORBfiLY AND IRENE TOBLER vohme 69
butoctamide hydrogen succinate on nocturnal sleep: all- night polygraphical studies. Psychopharmacology 70: 117-121,198O.
276.
277.
278.
275a.ONOE, H., R. UENO, I. FUJITA, H. NISHINO, Y. OO- MURA, AND 0. HAYAISHI. Prostaglandin Dz, a cerebral sleep-inducing substance in monkeys. Proc. NutL Acad Sci USA 85: 4082-4086,1988. PANKSEPP, J., J. E. JALOWIEC, A. J. ZOLOVICK, W. C. STERN, AND P. J. MORGANE. Inhibition of glyco- lytic metabolism and sleep-waking states in cats. Phur- macoL Biochem Behav. 1: 117-119,1973. PAPPENHEIMER, J. R. Sleep factor in CSF, brain and urine. Front. Harm Res. 9: 173-178,1982. PAPPENHEIMER, J. R., G. KOSKI, V. FENCL, M. L. KARNOVSKY, AND J. KRUEGER. Extraction of sleep- promoting factor S from cerebrospinal fluid and from brains of sleep-deprived animals. J. NeurophysioL 38: 1299-1311,1975. PAPPENHEIMER, J. R., T. B. MILLER, AND C. A. GOODRICH. Sleep-promoting effects of cerebrospinal fluid from sleep-deprived goats. Proc. Nat1 Acud Sci. USA 58: 513-517,1967. PARKER, D. C., M. MORISHIMA, D. J. KOERKER, C. C. GALE, AND C. J. GOODNER. Pilot study of growth hor- mone release in sleep of the chair-adapted baboon: po- tential as model of human sleep release. EndocrinoZo~~ 91: 1462-1467,1972. PARMEGGIANI, P. L., AND C. RABINI. Sleep and envi- ronmental temperature. Arch. Ital. BioL 108: 369-387, 1970.
279.
280.
281
282
283
285.
PAVEL, S., AND R. GOLDSTEIN. Narcoleptic-like alter- ations of the sleep cycle in cats induced by a specific vasotocin antiserum. Bruin Res. BuZL 7: 453-454,198l. PAVEL, S., R. GOLDSTEIN, M. PETRESCU, AND M. POPA. REM sleep induction in prepubertal boys by va- sotocin: evidence for the involvement of serotonin con- taining neurons. Peptides Fuyetteville 2: 245~247,198l. PAVEL, S., R. GOLDSTEIN, L. POPOVICIU, 0. COR- FARIU, A. FGLDES, AND E. FARKAS. Pineal vasotocin: REM sleep dependent release into cerebrospinal fluid of man. Waking Sleeping 3: 347-352,1979. PAVEL, S., D. PSATTA, AND R. GOLDSTEIN. Slow- wave sleep induced in cats by extremely small amounts of synthetic and pineal vasotocin injected into the third ventricle of the brain. Bruin Res. Bull 2: 251~254,1977.
286. PETITJEAN, F., C. BUDA, M. JANIN, M. DAVID, AND M. JOUVET. Effets du chloramphenicol sur le sommeil du chat-comparaison avec le thiamphenicol, l’erythro- mycine et l’oxytetracycline. Ps~chopharmucolog~ 66: 147-153,1979.
287. PETITJEAN, F., C. BUDA, M. JANIN, M. SALLANON, AND M. JOUVET. L’insomnie provoquee par la P-chloro- phenylalanine chez le chat. Sa reversibilite par l’injec- tion intraventriculaire d’indolamines. C. R. Acad. Sci Paris 291: 1063-1066,198O.
288. PETITJEAN, F., S. SEGUIN, M. H. DES ROSIERS, D. SALVERT, C. BUDA, M. JANIN, G. DEBILLY, M. JOU- VET, AND P. BOBILLIER. Local cerebral glucose utiliza- tion during waking and slow wave sleep in the cat. A. r*CJdeoxyglucose study. Neurosci Lett. 32: 91-97,1982.
289. PIERON, H. Le probldme phgsiologique du sommeil. Paris: Masson, 1913.
299. PIVIK, R. T., F. W. BYLSMA, AND P. COOPER. Sleep- wakefulness rhythms in the rabbit. Behuv. NeuroL BioL 45: 275-286,1986.
291. PIVIK, R. T., F. W. BYLSMA, AND P. COOPER. Effects of paradoxical sleep deprivation in the rabbit. Ph@oL Behuv. 36: 671-676,1986.
298.
299.
300.
301.
302.
303.
304 .
305 .
306 .
,
307 .
/ 308 .
E. L. J. M. VAN LUIJTELAAR, AND A. M. L. COENEN. Deprivation of REM sleep in the rat and the opioid pep- tides /?-endorphin and dynorphin. Neurosci. Lett. 70: 138-142,1986. PUIZILLOUT, J. J., G. GAUDIN-CHAZAL, AND H. BRAS. Vagal mechanisms in sleep regulation. In: Experi- mental Brain Research. Sleep Mechanisms, edited by A. A. Borbely and J. L. Valatx. Berlin: Springer-Verlag, 1984, vol. 8, p. 19-38. PUIZILLOUT, J. J., G. GAUDIN-CHAZAL, A. DAS- ZUTA, N. SEYFRITZ, AND J. P. TERNAUX. Release of endogenous serotonin from “encephale isole” cats. II. Correlation with raphe neuronal activity and sleep and wakefulness. J PhgsioL Paris 75: 531-537, 1979. PURPURA, D. P. A neurohumoral mechanism of reti- culo-cortical activation. Am. J. Phgsiol. 186: 250-254, 1956. RADULOVACKI, M., P. HAJDUK, P. STEFANOVICH, A. RADETICH, AND N. PORTER. The effects of p-l- methylcyclopentyladenosine on sleep in rats. Res. Com- mun. Ps@wL Psgchiatm Behav. 12: l-8,1987. RADULOVACKI, M., AND N. MICOVIC. Effects of REM sleep deprivation and desipramine on @-adrenergic bind- ing sites in rat brain. Bruin Res. 235: 393-396,1982. RADULOVACKI, M., R. S. MILETICH, AND R. D.
GREEN. N6(L-phenylisopropyl)adenosine (L-PIA) in- creases slow-wave sleep (&) and decreases wakefulness in rats. Bruin Res. 246: 178-180,1982. RADULOVACKI, M., R. M. VIRUS, M. DJURICIC-NE- DELSON, AND R. D. GREEN. Hypnotic effects of deoxy- corformycin in rats. Bruin Res. 271: 392-395,1983. RADULOVACKI, M., R. M. VIRUS, M. DJURICIC-NE- DELSON, AND R. D. GREEN. Adenosine analogs and sleep in rats. J. PhurmucoL Exp. Ther. 228: 268-274,1984. RADULOVACKI, M., R. M. VIRUS, D. RAPOZA, AND R. A. CRANE. A comparison of the dose response effects of pyrimidine ribonucleosides and adenosine on sleep in rats. Ps&ophumulcdogzl87: 136-140,1985. RECHTSCHAFFEN, A., R. DATES, M. TOBIAS, AND W. E. WHITEHEAD. The effects of lights-off stimula- tion on the distribution of paradoxical sleep in the rat. Commun. Behuv. BioL 3: 93-99,1969. RECHTSCHAFFEN, A., AND A. KALES. A Manual of Standardized Termino~, Techniques and Scoring Sz/s- tem for Sleep Stages of Human Su&e&. Bethesda, MD: US Depart. Health, Education, and Welfare, 1968.
292. POLC, P., J. SCHNEEBERGER, AND W. HAEFELY. Ef- fect of the delta sleep-inducing peptide (DSIP) on the sleep-wakefulness cycle of cats. Neurosci. Lett. 9: 33-36, 1978.
293. POPOVICIU, L., 0. CORFARIU, A. FijLDES, E. FARKAS, R. GOLDSTEIN, AND S. PAVEL. REM sleep dependent release of vasotocin into cerebrospinal fluid of narcoleptics. Waking Sleeping 3: 341~346,1979.
294. PRECHEL, M. M., T. K. AUDHYA, AND D. H. SCHLE- SINGER. A seasonal variation in arginine vasotocin im- munoactivity in rat pineal glands. Endocrinology 112: 1474-1478,1983.
295. PROSPERO-GARCIA, O., M. MORALES, G. ARAN- KOWSKY-SANDOVAL, AND R. DRUCKER-COLIN. Va- soactive intestinal polypeptide (VIP) and cerebrospinal fluid (CSF) of sleep-deprived cats restores REM sleep in insomniac recipients. Bruin Res. 385: 169-173,1986.
296. PROSPERO-GARCIA, O., T. OTT, AND R. DRUCKER- COLIN. Cerebroventricular infusion of cholecystokinin (CCK-8) restores REM sleep in parachlorophenylalanine (PCPA)-pretreated cats. Neurosci. Lett. 78: 205-210,1987.
297. PRZEWLOCKA, B., E. MOGILNICKA, W. LASON,

April 1989 ENDOGENOUS SLEEP-PROMOTING SUBSTANCES 667
309. REITE, M., M. LAUDENSLAGER, J. JONES, L. CRNIC, AND K. KAEMINGK. Interferon decreases REM latency. BioL Psychiatry 22: 104-107,1987.
310. RHYNER, T. A., N. FAUCON BIGUET, S. BERRARD, A. A. BORBELY, AND J. MALLET. An efficient approach for the selective isolation of specific transcripts from complex brain mRNA populations. J. Neurosci. Res. 16: 167-181,1986.
3lOa.RHYNER, T. A., J. MALLET, AND A. A. BORBELY. A molecular genetic approach to endogenous sleep promot- ing substances. In: Sleep ‘88, edited by J. A. Horne and P. Lavie. Stuttgart, FRG: Gustav Fischer-Verlag. In press.
311. RICHARDSON, G. S., M. A. CARSKADON, E. J. ORAV, AND W. C. DEMENT. Circadian variation of sleep ten- dency in elderly and young adult subjects. Sleep 5, SuppL 2: S82-S94,1982.
312. RINGLE, D. A., AND B. L. HERNDON. Plasma dialysates from sleep-deprived rabbits and their effect on the elec- trocorticogram of rats. PJuegers Arch. 303: 344-349, 1968.
313. RIOU, F., R. CESPUGLIO, AND M. JOUVET. Endoge- nous peptides and sleep in the rat. I. Peptides decreasing paradoxical sleep. Neuropeptides 2: 243-254,1982.
314. RIOU, F., R. CESPUGLIO, AND M. JOUVET. Endoge- nous peptides and sleep in the rat. II. Peptides without significant effect on the sleep-waking cycle. Neuropep- tides 2: 255-264,1982.
315. RIOU, F., R. CESPUGLIO, AND M. JOUVET. Endoge- nous peptides and sleep in the rat. III. The hypnogenic properties of vasoactive intestinal polypeptide. New-+ peptides 2: 265-277,1982.
316. ROJAS-RAMIREZ, J. A. Waking induced by perfusates collected from alert cats. In: Sleep ‘7.4, edited by P. Levin and W. P. Koella. Basel: Karger, 1975, p. 306-310.
317. ROJAS-RAMIREZ, J. A., J. N. CRAWLEY, AND W. B. MENDELSON. Electroencephalographic analysis of the sleep-inducing actions of cholecystokinin. Neuropeptides 3: 129-138,1982.
318. ROSENBERG, R. S., B. M. BERGMANN, AND A. RECHTSCHAFFEN. Variations in slow wave activity during sleep in the rat. PhysioL Behuv. 17: 931-938,1976.
319. RUCKEBUSCH, Y., R. W. DOUGHERTY, AND H. M. COOK. Jaw movements and rumen motility as criteria for measurement of deep sleep in cattle. Am J Vet. Res. 35: 1309-1312,1974.
320. RUSAK, B., AND I. ZUCKER. Neural regulation of circa- dian rhythms. PhysioL Rev. 59: 449-526,1979.
321. SACHS, J., J. UNGAR, P. G. WASER, AND A. A. BORBELY. Factors in cerebrospinal fluid affecting motor activity in the rat. Neurosci I&t. 2: 83-86,1976.
322. SALLANON, M., C. BUDA, M. JANIN, AND M. JOUVET. L’insomnie provoquee par la p-chlorophenylalanine chez le chat. Sa reversibilit& par l’injection intraventriculaire de liquide cephalorachidien p&eve chez des chats priv& de sommeil paradoxal. C. R. Ad Sti Paris 292: 113-117, 1981.
323. SALLANON, M., C. BUDA, M. JANIN, AND M. JOUVET. Restoration of paradoxical sleep by cerebrospinal fluid transfer to PCPA pretreated insomniac cats. Bruin Res. 251: 137-147,1982.
324. SALLANON, M., C. BUDA, M. JANIN, AND M. JOUVET. 5-HT antagonists suppress sleep and delay its restora- tion after 5-HTP in p-chlorophenylalanine-pretreated cats. Eur. J. PhurmucoL 82: 29-35, 1982.
325. SALLANON, M., K. KITAHAMA, M. DENOYER, N. GAY, AND M. JOUVET. Insomnie de longue duree apr& lesions des pi%ykarions de l’aire preoptique paramediane chez le chat. C R. Ad Sci Paris 303: 403-409,1986.
326. SARDA, N., M. DUBOIS, A. GHARIB, J.-L. VALATX, AND M. JOUVET. Increase of paradoxical sleep induced by S-adenosyl-L-homocysteine. Neurosci L&t. 30: 69-72, 1982.
327. SASTRE, J.-P., C. BUDA, M. JANIN, S. SEGUIN, M. SALLANON, AND M. JOUVET. L’injection systemique ou intracerhbrale d’extraits de lobe neuro-intermhdiaire de boef peut induire le sommeil paradoxal au tours de l’insomnie provoquee par la P. chlorophenylalanine chez le chat. C. R. Acud Sci Paris 300: 13-17,1985.
328. SASTRE, J.-P., M. SALLANON, C. BUDA, AND M. JOU- VET. La bacitracine, un inhibiteur des peptidases, aug- ment le sommeil paradoxal chez le chat. C. R. Acad Sci. Paris 296: 965-968,1983.
329. SCHERSCHLICHT, R., L. AEPPLI, P. POLC, AND W. HAEFELY. Some pharmacological effects of delta- sleep-inducing peptide (DSIP). Eur. Neural 23: 346-352, 1984.
330. SCHERSCHLICHT, R., AND J. MARIAS. Effects of sleep promoting “factor S”, muramyl dipeptide (MDP) and L- cycloserine on the sleep of rabbits (Abstract). Experien- tiu BaaeZ39: 683, 1983.
331. SCHERSCHLICHT, R., J. MARIAS, J. SCHNEE- BERGER, AND M. STEINER. Model insomnia in ani- mals. In: Sleep ‘SO, edited by W. P. Koella. Basel: Karger, 1981, p. 147-155.
332. SCHNEDORF, J. G., AND A. C. IVY. An examination of the hypnotoxin theory of sleep. Am. J. Physiol. 125: 491-505,1939.
333. SCHNEIDER-HELMERT, D. DSIP in insomnia. Eur. NeuroL 23: 358-363,1984.
334. SCHNEIDER-HELMERT, D. Efficacy of DSIP to nor- malize sleep in middle-aged and elderly chronic insom- niacs. Eur. New-01 25: 448-453,1986.
335. SCHNEIDER-HELMERT, D. Effects of delta-sleep-in- ducing peptide on 24-hour sleep-wake behaviour in se- vere chronic insomnia. Eur. Neural 27: 120-129,1987.
336. SCHNEIDER-HELMERT, D. DSIP: clinical application of the programming effect. In: Sleep Peptides: Basic and Clinical Approaches, edited by S. Inoue and D. Schneider-Helmert. Tokyo: Jpn. Sci. Sot. Press, 1988,’ p. 175-198.
337. SCHNEIDER-HELMERT, D., M. GRAF, AND G. A. SCHOENENBERGER. Synthetic delta-sleep-inducing peptide improves sleep in insomniacs. Luncet 1: 1256, 1981.
338. SCHNEIDER-HELMERT, D., AND G. A. SCHOENEN- BERGER. The influence of synthetic DSIP (delta-sleep- inducing-peptide) on disturbed human sleep. Experientiu Base1 37: 913-1038,198l.
339. SCHNEIDER-HELMERT, D., AND G. A. SCHOENEN- BERGER. Effects of DSIP in man. Multifunctional psy- chophysiological properties besides induction of natural sleep. Neuropsych&ology 9: 197-206,1983.
340. SCHNEIDER-HELMERT, D., AND C. L. SPINWEBER. Evaluation of I.,-tryptophan for treatment of insomnia: a review. Psychopharmucolugy 89: l-7,1986.
341. SCHOENENBERGER, G. A., P. F. MAIER, H. J. TOBLER, AND M. MONNIER. A naturally occurring delta-EEG enhancing nonapeptide in rabbits. X. Final isolation, characterization and activity test. Pfluegers Arch 369: 99-109,1977.
342. SCHOENENBERGER, G. A., P. F. MAIER, H. J. TOBLER, K. WILSON, AND M. MONNIER. The delta EEG (sleep)-inducing peptide (DSIP). XI. Amino-acid analysis, sequence, synthesis and activity of the non- apeptide. &uegers Arch, 376: 119-129,197s.
343. SCHOENENBERGER, G. A., AND M. MONNIER. Char-

668 ALEXANDER A. BORB@LY AND IRENE TOBLER Votkme 69
344.
345
346
347
348
349.
350.
351.
352.
353.
354.
355.
356
357
358
359
acterization of a delta-electroencephalogram(-sleep)-in- ducing peptide. Proc. Nat1 Acad Sci USA 74: 1282-1286, 1977. SHAPIRO, C. M., R. BORTZ, D. MITCHELL, P. BAR- TEL, AND P. JOOSTE. Slow wave sleep: a recovery period after exercise. science Wash, DC 214: 1253-X%4,1981. SHIROMANI, P. J., AND W. FISHBEIN. Continuous pontine cholinergic microinfusion via mini-pump in- duces sustained alterations in rapid eye movement (REM) sleep. PhurmucoL B&hem Behuv. 25: 1253-1261, 1986. SHIROMANI, P. J., AND J. C. GILLIN. Acetylcholine and the regulation of REM sleep: basic mechanisms and clin- ical implications for affective illness and narcolepsy. Ann. Rev. PhwmacoL ToxicoL 27: 137-156,1987. SHIROMANI, P. J., AND D. J. McGINTY. Pontine neuro- nal response to local cholinergic infusion: relation to REM sleep. Brain Res. 386: 20-31,1986. SHOHAM, S., R. A. AHOKAS, C. M. BLATTEIS, AND J. M. KRUEGER. Effects of muramyl dipeptide on sleep, body temperature and plasma copper after intracerebral ventricular administration. Brain Res. 419: 223-228, 1987. SHOHAM, S., D. DAVENNE, A. B. CADY, C. A. DIN- ARELLO, AND J. M. KRUEGER. Recombinant tumor necrosis factor and interleukin 1 enhance slow-wave sleep. Am J. Phgsiol 253 (R&gulutoq Integrative Comp. PhgsioL 22): R142-R149, 1987. SHOHAM, S., D. DAVENNE, AND J. M. KRUEGER. Muramyl dipeptide, amphetamine, and physostigmine: effects on sleep of rabbits. Ph&oL Behuv. 41: 179-185, 1987. SHOHAM, S., AND J. M. KRUEGER. Muramyl dipep- tide-induced sleep and fever: effects of ambient tempera- ture and time of injections. Am J. PhysioL 255 (Rem- toq Integrative Comp. PhgsioL 24): R157-R165,1988. SOMMERFELT, L. Reduced sleep in cats after intraperi- toneal injection of b-sleep-inducing peptide (DSIP). Neurosci. L&t. 58: 73-77,1985. SPANIS, C. W., M. DEL CARMEN GUTIERREZ, AND R. DRUCKER-COLIN. Neurohumoral correlates of sleep: further biochemical and physiological characterization of sleep perfusates. Pharmacol. Biochem. Behav. 5: 165-173,1976. STENBERG, D., AND T. PORKKA-HEISKANEN. Sleep regulation and protein synthesis in cats. In: Sleep ‘86, edited by W. P. Koella, F. Obal, H. Schulz, and P. Visser. Stuttgart, FRG: Gustav Fischer-Verlag, 1988, p. 300-302. STERN, W. C., W. B. FORBES, J. E. JALOWIEC, AND P. J. MORGANE. Growth hormone administration and sleep in the rat (Abstract). Sleep Res. 4: 286,1975. STERN, W. C., J. E. JALOWIEC, H. SHABSHELOWITZ, AND P. J. MORGANE. Effects of growth hormone on sleep-waking patterns in cats. Harm Behuv. 6: 189-196, 1975. STERN, W. C., AND P. J. MORGANE. Sleep and memory: effects of growth hormone on sleep, brain neurochemis- try and behavior. In: Neurobiology of Sleep and Memory, edited by R. Drucker-Cohn and J. L. McGaugh. New York: Academic, 1977, p. 373-400. STERN, W. C., P. J. MORGANE, J. PANKSEPP, A. J. ZOLOVICK, AND J. E. JALOWIEC. Elevation of REM sleep following inhibition of protein synthesis. Brain Res. 47: 254-258,1972. SUSIC, V. The effect of subcutaneous administration of delta sleep-inducing peptide (DSIP) on some parameters of sleep in the cat. PhgsioL Behav. 40: 569-572,1987.
360. SUSIC, V., AND G. MASIREVIC. Effects of delta sleep inducing peptide on sleep cycle of cats deprived of para- doxical sleep. Arch. Int. Ph&oL B&him. 93: 271-277, 1985.
361. SUSIC, V., G. MASIREVIC, AND S. TOTIC. The effects of delta-sleep-inducing peptide (DSIP) on wakefulness and sleep patterns in the cat. Brain Res. 414: 262~270,1987.
362. SZYMUSIAK, R. S., AND E. SATINOFF. Maximal REM sleep time defines a narrower thermoneutral zone than does minimal metabolic rate. PhysioL Behav. 26: 687-690, 1981.
363. SZYMUSIAK, R. S., T. SCHALLERT, E. SATINOFF, AND I. Q. WHISHAW. Induction of REM sleep in rats by peripheral warming. Sot. Neurosci Al&r. 4: 544,1978.
364. TAKAHASHI, Y., S. EBIHARA, Y. NAKAMURA, AND K. TAKAHASHI. ‘A model of human sleep-related growth hormone secretion in dogs: effects of 3,6, and 12 hours of forced wakefulness on plasma growth hormone, cortisol, and sleep stages. Endocrinologll 109: 262-272, 1981.
365. TAKAHASHI, Y., N. KATO, Y. NAKAMURA, S. EBI- HARA, A. TSUJI, AND K. TAKAHASHI. Delta sleep in- ducing peptide (DSIP): a preliminary report of effects of intraventricular infusion on sleep and GH secretion, and of plasma DSIP concentrations in the dog and rat. In: Integrative Control Functions of the Brain, edited by M. Ito, N. Tsukahara, K. Kubota, and K. Yagi. Tokyo: Ko- dansha-Elsevier, 1980, vol. 2, p. 339-341.
366. TAKAHASHI, Y., D. M. KIPNIS, AND W. H. DAUGHA- DAY. Growth hormone secretion during sleep. J. Clin, Invest. 47: 2079-2090,1968.
367. TISSOT, R. Recepteurs de l’opium et sommeil. III. Effets de micro-injections de DSIP (delta sleep peptide) dans le thalamus median, la substance grise centrale p&i-aque- ducale et le noyau du tractus solitaire du lapin. Newo- psychobio~ 7: 321-325,198l.
368. TOBLER, I. Evolution of the sleep process: a phylogene- tic approach. In: Experimental Brain Research. Sleep Mechanisms, edited by A. A. Borbely and J. L. Valatx. Berlin: Springer-Verlag, 1984, suppl. 8, p. 207226.
369. TOBLER, I. Deprivation of sleep and rest in vertebrates and invertebrates. In: Endogenous Sleep Substances and Sleep Regulation, edited by S. Inoue and A. A. Bortily. Tokyo: Jpn. Sci. Sot. Press, 1985, vol. 8, p. 57-66.
370. TOBLER, I. Napping and polyphasic sleep in mammals. In: Napping: Biological, Ps2/chological, and Medical Aspects, edited by D. F. Dinges and R. J. Broughton. New York: Raven. In press.
371. TOBLER, I., AND A. A. BORBELY. Enhancement of par- adoxical sleep by short light periods in the golden ham- ster. Neurosci Lett. 6: 275-277,1977.
372. TOBLER, I., AND A. A. BORBELY. Short light-dark cycles and paradoxical sleep in the rat: effect of strain difference and hypophysectomy. Behav. BioL 23: 395-398, 1978.
373. TOBLER, I., AND A. A. BORBELY. Effect of delta sleep inducing peptide (DSIP) and arginine vasotocin (AVT) on sleep and motor activity in the rat. Waking Sleeping 4: 139-153,198o.
374. TOBLER, I., AND A. A. BORBELY. Sleep regulation after reduction of brain serotonin: effect of p-chlorophenyla- lanine combined with sleep deprivation in the rat. Sleep 5: 145-153,1982.
375. TOBLER, I., AND A. A. BORBELY. Sleep EEG in the rat as a function of prior waking. Electroencephabgr. Clin, Neuroph@oL 64: 74-76,1986.
376. TOBLER, I., A. A. BORBELY, AND G. GROOS. The effect

April 1989 ENDOGENOUS SLEEP-PROMOTING SUBSTANCES 669
of sleep deprivation on sleep in rats with suprachiasma- tic lesions. Neurosci Lett. 42: 49-54, 1983.
377. TOBLER, I., A. A. BORBELY, M. SCHWYZER, AND A. FONTANA. Interleukin-1 derived from astrocytes en- hances slow wave activity in sleep EEG of the rat. Eur. J. PhurmucoL 104: 191-192,1984.
378. TOBLER, I., AND K. JAGGI. Sleep and EEG spectra in the Syrian hamster (Mesocricetus aurutus) under base- line conditions and following sleep deprivation. J. Camp. PhysioL A Sens. Neural Behuv. Phgsiol. 161: 449-459, 1987.
379. TOBLER, I., R. MURISON, R. URSIN, H. URSIN, AND A. A. BORBELY. The effect of sleep deprivation and re- covery sleep on plasma corticosterone in the rat. Neuro- SC-i. Lett. 35: 297-300,1983.
380. TORII, S., S. INUBUSHI, AND A. SAKUMA. Effects of related compounds of naturally occurring bromosub- stance on the sleep-wakefulness cycle in the cat. In: Ad- vances in Sleep Research, edited by E. D. Weitzman. New York: Spectrum, 1976, vol. 2, p. 155-163.
381. TORII, S., K. MITSUMORI, S. INUBUSHI, AND I. YAN- AGISAWA. The REM sleep-inducing action of a natu- rally occuring organic bromine compound in the encephale isole cat. Ps@ophurmuco~ 29: 65-75,1973.
382. TORSVALL, L., T. AKERSTEDT, AND G. LINDBECK. Effects on sleep stages and EEG power density of differ- ent degrees of exercise in fit subjects. Ekctroencephu- bgr. Clin. Neuroph~sioL 57: 347-353,1984.
383. TOSZEGHI, P., I. TOBLER, AND A. A. BORBELY. Cere- bral ventricular infusion of excess calcium in the rat: effects on sleep states, behavior and cortical EEG. Ekc- troencephulogr. Clin Neuroph~siol 51: 407-416,1978.
384. TRACHSEL, L., I. TOBLER, AND A. A. BORBELY. Sleep regulation in rats: effects of sleep deprivation, light, and circadian phase. Am J. Physid 251 (Regulutorq Integra- tive Camp. PhysioL 20): R1037-R1044,1986.
385. TRACHSEL, L., I. TOBLER, AND A. A. BORBELY. Spectral EEG analysis of rat nonREM sleep: changes in the 24 hour cycle and within single sleep episodes (Ab- stract). Sleep Res. 16: 35,1987.
386. TRACHSEL, L., I. TOBLER, AND A. A. BORBELY. Elec- troencephalogram analysis of non-rapid eye movement sleep in rats. Am J. Ph&ol 255 (Regulutq Integrative Camp. Ph2/sioL 24): R27-R37, 1988.
387. UCHIZONO, K., A. HIGASHI, M. IRIKI, H. NAGA- SAKI, M. ISHIKAWA, Y. KOMODA, S. INOIJ& AND K. HONDA. Sleep-promoting fractions obtained from the brain-stem of sleep-deprived rats. In: Integrative Control Functions of the Brain, edited by M. Ito, K. Kubota, N. Tsukahara, and K. Yagi. Amsterdam: Elsevier/North- Holland, 1978, vol. 1, p. 392-396.
388. UCHIZONO, K., M. ISHIKAWA, M. IRIKI, S. INOUE, Y. KOMODA, H. NAGASAKI, A. HIGASHI, K. HONDA, AND A. McRAE-DEGUEURCE. Purification of sleep- promoting Substances (SPS). In: Advances in PhurmucoZ- og2/& Therapeutics II, edited by H. Yoshida, Y. Hagihara, and S. Ebashi. Oxford, UK: Pergamon, 1982, vol. 1, p. 217-225.
389. UENO, R., 0. HAYAISHI, H. OSAMA, K. HONDA, S. INOUE, Y. ISHIKAWA, AND T. NAKAYAMA. Prosta- glandin Ds regulates physiological sleep. In: Endogenous Sleep Substances and Sleep Regulation, edited by S. Inoue and A. A. Borbely. Tokyo: Jpn. Sci. Sot. Press, 1985, vol. 8, p. 193-201.
390. UENO, R., K. HONDA, S. INOUE, AND 0. HAYAISHI. Prostaglandin Ds, a cerebral sleep-inducing substance in rats. Proc. NatL Acad Sci USA 80: 1735-1737,1983.
391. UENO, R., Y. ISHIKAWA, T. NAKAYAMA, AND 0. HAYAISHI. Prostaglandin Ds induces sleep when mi- croinjected into the preoptic area of conscious rats. Bio- them, Biophys. Res. Commun, 109: 576-582,1982.
392. UENO, R., H. ONOE, H. MATSUMURA, 0. HAYAISHI, I. FUJITA, H. NISHINO, AND Y. OOMURA. Regulation of sleep by prostaglandins in conscious rhesus monkeys
393.
394.
395.
396.
(Abstract). Sleep Res. 16: 36,1987. URSIN, R. Endogenous sleep factors. In: Experimental Brain Research. Sleep Mechanisms, edited by A. A. Bortily and J. L. Valatx. Berlin: Springer-Verlag, 1984, suppl. 8, p. 118-132. URSIN, R., AND A. A. BORBELY. Endogenous sleep fac- tors. In: Sleep ‘8.2, edited by W. P. Koella. Basel: Karger, 1983, p. 106-125. URSIN, R., AND M. LARSEN. Increased sleep following intracerebroventricular injection of the delta sleep-in- ducing peptide in rats. Neurosci. Lett. 40: 145-149,1983. VALATX, J. L., B. ROUSSEL, AND M. CURE. Sommeil et temperature cerebrale du rat au tours de l’exposition chronique en ambiance chaude. Bruin Res. 55: 107-122, 1973.
397.
398 .
, (
399 .
400 .
VAN HULZEN, Z. J. M., AND A. M. L. COENEN. The pendulum technique for paradoxical sleep deprivation in rats. Ph@oL Behuv. 25: 807-811,198O. VAN LUIJTELAAR, E. L. J. M., AND A. M. L. COENEN. Electrophysiological evaluation of three paradoxical sleep deprivation techniques in rats. PhgsioL Behuv. 36: 603-609,1986.
401 I
402
403 .
/ 404 .
405 .
406 .
407 .
408 .
409 .
VAN TWYVER, H. Sleep patterns of five rodents. Phg- sioL Behuv. 4: 901-905,1969. VEALE, W. L., AND R. D. MYERS. Emotional behavior, arousal and sleep produced by sodium and calcium ions perfused within the hypothalamus of the cat. PhgsioL Behuv. 7: 601~607,197l. VITALE-NEUGEBAUER, A., A. GIUDITTA, B. VI- TALE, AND S. GIAQUINTO. Patterns of RNA synthesis in rabbit cortex during sleep. J. Neurochem. 17: 1263-1273,197O. VOGEL, G. W. A review of REM sleep deprivation. Arch, Gen Psychiatry 32: 749-761,1975. VOGEL, G. W., K. MINTER, AND B. WOOLWINE. Ef- fects of chronically administered antidepressant drugs on animal behavior. PhysioL Behuv. 36: 659-666, 1986. VOLLRATH, L., P. SEMM, AND G. GAMMEL. Sleep in- duction by intranasal application of melatonin. Bio- sciences 29: 327-329,198l. WALTER, J., D. DAVENNE, S. SHOHAM, C. A. DIN- ARELLO, AND J. M. KRUEGER. Brain temperature changes coupled to sleep states persist during interleu- kin l-enhanced sleep. Am. J. PhpsioL 250 (Regulatory Integrative Camp. Ph&oL 19): R96-R103,1986. WAUQUIER, A., G. H. C. CLINCKE, W. A. E. VAN DEN BROECK, AND E. DE PRINS. Active and permissive roles of dopamine in sleep-wakefulness regulation. In: Sleep: Neurotransmitters and Neuromodulutors, edited by A. Wauquier, J.-M. Gaillard, J. M. Monti, and M. Radulo- vacki. New York: Raven, 1985, p. 107-120. WEBB, W. B. The sleep of conjoined twins. Sleep 1: 205-211,1978. WEBB, W. B., AND H. W. AGNEW, JR. Stage 4 sleep: influence of time course variables. science Wash. DC 174: 1354-1356,197l. WEITZMAN, E. D., C. A. CZEISLER, J. C. ZIMMER- MAN, AND J. M. RONDA. Timing of REM and stages 3 + 4 sleep during temporal isolation in man. Sleep 2: 391-407,198O.

670 ALEXANDER A. BORBELY AND IRENE TOBLER Vohme 69
410. WEITZMAN, E. D., AND L. HELLMAN. Temporal orga- nization of the 24-hour pattern of the hypothalamic-pi- tuitary axis. In: Biorhythm and Human Reproduction, edited by M. Ferin, F. Halberg, R. M. Richart, and R. L. Vandewiele. New York: Wiley, 1974, p. 371-395.
411. WEITZMAN, E. D., D. F. KRIPKE, D. GOLDMACHER, P. MCGREGOR, AND C. NOGEIRE. Acute reversal of the sleep-waking cycle in man. Arch. NeuroL 22: 483-489, 1970.
412. WEITZMAN, E. D., U. WEINBERG, R. D’ELETTO, H. LYNCH, R. J. WURTMAN, C. CZEISLER, AND S. ER- LICH. Studies of the 24-hour rhythm of melatonin in man. J. Neural Transm. Suppl 13: 325337,1978.
413. WESEMANN, W., M. ROTSCH, E. SCHULZ, G. STURM, AND P. ZOFEL. Circadian rhythm of serotonin binding in rat brain. I. Effect of the light-dark cycle. ChronobioL Ink 3: 135139,1986.
414. WESEMANN, W., M. ROTSCH, E. SCHULZ, AND P. ZOFEL. Circadian rhythm of serotonin binding in rat brain. II. Influence of sleep deprivation and imipramine. ChronobioL Int. 3: 141-146,1986.
415. WESEMANN, W., N. WEINER, M. ROTSCH, AND E. SCHULZ. Serotonin binding in rat brain: circadian rhythm and effect of sleep deprivation. J. Neural. Transm. SuppL 1& 287-294,1983.
416. WETZEL, W., H. STARK, AND H. MATTHIES. Increase of paradoxical sleep (PS) by intraperitoneal injection of brain extract from PS-deprived rats. Pharmacol. Bb chmn Behav. 27: 537-539,1987.
417. WEXLER, D. B., AND M. C. MOORE-EDE. Effects of a muramyl dipeptide on the temperature and sleep-wake cycles of the squirrel monkey. Am J. Ph@oL 247 (Regu- luta7?y Integrative Camp. PhgsioL 16): R672-R680,1984.
418. WILLIAMS, H. L., J. T. HAMMACK, R. L. DALY, W. C. DEMENT, AND A. LUBIN. Responses to auditory stimu- lation, sleep loss and the EEG stages of sleep. EZectroen- cephulogr. C&n. Neuroph&L 16: 269-279,1964.
419. WIRZ-JUSTICE, A. Circadian rhythms in mammalian neurotransmitter receptors. Prog. NeurobioL 29: 219-259, 1987.
420. WIRZ-JUSTICE, A., I. TOBLER, M. S. KAFKA, D.
NABER, P. J. MARANGOS, A. A. BORBELY, AND ‘I’. A. WEHR. Sleep deprivation: effects on circadian rhythms of rat brain neurotransmitter receptors. Psgchiatr. Res. 5: 67-76,198l.
421. WU, J. C., M. S. BUCHSBAUM, AND J. C. GILLIN. Posi- tron emission tomography study of sleep deprivation (Abstract). Sleep Res. 16: 543,1987.
422. YANAGISAWA, I., AND H. YOSHIKAWA. Micro-deter- mination of bromine in cerebrospinal fluid. Clin. Chim Acta 21: 217~224,1968.
423. YANAGISAWA, I., AND H. YOSHIKAWA. A bromine compound isolated from human cerebrospinal fluid, and synthesis related compounds. B&him. Biophgs. Acta 329: 283-294,1973.
424. YANIK, G., AND M. RADULOVACKI. REM sleep depri- vation upregulates adenosine A1 receptors. Brain Res. 402: 362-364,1987.
425. YEHUDA, S., A. J. KASTIN, AND D. H. COY. Thermo- regulatory and locomotor effects of DSIP: paradoxical interaction with d-amphetamine. PharmacoL Biochem, Behav. 13: 895-900,198O.
426. YEHUDA, S., AND D. I. MOSTOFSKY. Possible seroton- ergic mediation of induction of sleep signs by DSIP and by d-amphetamine. Int. J. Neurosci 16: 221-226,1982.
427. ZINI, I., E. MERLO PICH, K. FUXE, P. L. LENZI, L. AGNATI, A. HARFSTRAND, V. MUTT, K. TATE- MOTO, AND M. MOSCARA. Actions of centrally admin- istered neuropeptide Y on EEG activity in different rat strains and in different phases of their circadian cycle. Acta PhysioL Scud 122: 71-77,1984.
428. ZULLEY, J., AND S. S. CAMPBELL. Napping behavior during “spontaneous internal desynchronization”: sleep remains in synchrony with body temperature. Hum. NeurobioL 4: 123-126,1985.
429. ZULLEY, J., R. WEVER, AND J. ASCHOFF. The depen- dence of onset and duration of sleep on the circadian rhythm of rectal temperature PjZuegers Arch. 391: 314-318,198l.
430. ZWICKER, A. P., AND H. M. CALIL. The effects of REM sleep deprivation on striatal dopamine receptor sites. PhawnucoL Biochem. Behuv. 24: 809-812,1986.