ecology of endolithic lichens colonizing granite in continental antarctica
TRANSCRIPT

The Lichenologist 37(?): 000 (2005) � 2005 The British Lichen Societydoi:10.1017/S0024282905014969 Printed in the United Kingdom
Ecology of endolithic lichens colonizing granite in continentalAntarctica
Asunción DE LOS RIuOS, Jacek WIERZCHOS, Leopoldo G. SANCHO,Allan GREEN and Carmen ASCASO
Abstract: In this study, the symbiont cells of several endolithic lichens colonizing granite incontinental Antarctica and the relationships they have with the abiotic environment were analyzed insitu, in order to characterize the microecosystems integrating these lichens, from a microecologicalperspective. Mycobiont and photobiont cells, the majority classified as living by fluoresecent vitalitytesting, were observed distributed through the fissures of the granite. The fact that extracellularpolymeric substances were commonly observed close to these cells and the features of thesecompounds, suggest a certain protective role for these substances against the harsh conditions of theenvironment. Different chemical, physical and biological relationships take place within the endo-lithic biofilms where the lichens are found, possibly affecting also the survival and distribution of theseorganisms. The alteration of bedrock minerals and synthesis of biominerals in the proximity of theselichens gives rise to different chemical microenvironments and suggests their participation in mineralnutrient cycling.
Key words: Antarctica, endoliths, EPS, lichens, microecology, mineral-lichen interactions
Introduction
The Antarctic continent is the coldest, dri-est, windiest and most isolated landmass onEarth, yet despite these harsh conditions,there is still substantial terrestrial vegetation(Robinson et al. 2003). As part of this veg-etation, many lichen species are componentsof lithobiontic communities (Seppelt et al.1995; Castello & Nimis 1997; Øvstedal &Lewis Smith 2001). Some of these lichenscan withstand extreme environmental con-ditions and live on the surface of rocks.However, others appear to avoid these con-ditions by colonizing the inside of lithicsubstrata (Wierzchos & Ascaso 2001; Ascaso
& Wierzchos 2003; De los Ríos et al. 2005).Thus, we can find microbial life in areaswhere the environmental conditions pre-clude the colonization of rock surfaces(Friedmann 1982; Green et al. 1999).
The study of lithobiontic microorganismsin general, and endolithic lichens in particu-lar, has met with numerous difficulties. Thesymbionts of the latter are not only hidden inthe hard lithic substratum but their mostimportant interactions are those establishedwith their immediate physical environment,i.e. the lithic substratum. Hence endolithiclichens require analyses on a microecologicalscale, for which the approaches normallyused for ecological studies on epilithiclichens are inappropriate. The physicalorganization of lichen symbiont cells withinlocal lithic microenvironments and inter-relations between these and the surroundingenvironment must be analysed from a micro-ecological perspective. Microscopy is anideal technique for such studies, especiallywhen applied in situ, that is, when the bio-logical components are not separated from
A. de los Ríos and C. Ascaso: Centro de CienciasMedioambientales (CSIC), Serrano 115 bis, 28006Madrid, Spain.J. Wierzchos: Servei de Microscopia Electronica, UdL,Rovira Roure 44, Lleida, Spain.L. G. Sancho: Biologia Vegetal II, Universidad Com-plutense de Madrid, 28040 Madrid, Spain.A. Green: Biological Sciences, Waikato University,Hamilton, New Zealand.

the lithic material (Ascaso & Wierzchos2003; De los Ríos et al. 2005). Using thistechnique, the zone of the lithic substratumcolonized by lichens is seen as complexorganic-mineral interfaces composed of arange of microorganisms, the organic com-pounds generated by them, together withinorganic materials. At these interfaces, li-chenized algae and fungi, sometimes withfree-living microorganisms, are organized asbiofilms (De los Ríos et al. 2002). The suiteof microorganisms comprising these biofilmsand their biotopes where different inter-relationships are established, constitute atiny ecosystem or microecosystem (De losRíos et al. 2003). In the present study, wehave used in situ microscopy techniques tocharacterize both the biological and abioticcomponents of microecosystems integratingendolithic lichens from two sites in conti-nental Antarctica, and examined the inter-actions occurring in the system on amicroecological scale. This work forms partof our ongoing project designed to charac-terize different ecological aspects of endo-lithic lichens in an effort to improve ourunderstanding of how these lithic micro-ecosystems function.
Materials and MethodsMaterials
Fragments of granite rock were collected from severalsites near Canada Glacier Taylor Valley (Dry Valleys,77(37#S 162(59#E) and Cape Geology, GraniteHarbour (Ross Sea coast 77(00#S 162(00#E). Rocksamples, 0·5–1 cm thick, with visible signs of endolithicmicrobial growth were placed in sterile polyethylenebags, transported by air to our laboratory and stored dryat �20(C until processing.
Transmission electron microscopy examination
Fragments of endolithic biofilm were removed fromthe rock under the stereomicroscope using a sterileneedle and embedded in 2% (w/v) agar. Small pieces ofagar containing the microbial cells were then immedi-ately fixed in glutaraldehyde and osmium tetroxidesolutions, dehydrated in a graded ethanol series, andembedded in Spurr’s resin following the protocol de-scribed in De los Ríos & Ascaso (2001). Ultrathinsections were post-stained with lead citrate (Reynolds1963) and observed in a Zeiss EM910 transmissionelectron microscope (TEM).
Scanning Electron Microscopy
Low TemperatureFragments of rock colonized by lichens were
examined using Low Temperature Scanning ElectronMicroscopy (LTSEM). Small fragments were mountedusing O.C.T. compound (Gurr) and mechanically fixedonto the specimen holder of the cryotransfer system(Oxford CT1500). These samples were plunge-frozenin subcooled liquid nitrogen and then transferred to thepreparation unit. The frozen specimens were cryo-fractured and etched for 2 min at �90(C. After icesublimation, the etched surfaces were gold sputtercoated and the specimens then placed on the cold stageof the SEM chamber. Fractured surfaces were observedunder a DSM960 Zeiss SEM microscope at �135(Cunder conditions of a 15 kV acceleration potential,10 mm working distance and 5–10 nA probe current.
Back-Scattered Electron Mode and Energy DispersiveX-ray Spectroscopy
Fragments of granite colonized by endolithic lichenswere processed for scanning electron microscopy withbackscattered electron imaging (SEM-BSE) and/orenergy dispersive X-ray spectroscopy (EDS) analysisaccording to a method described by Wierzchos andAscaso (1994). The fragments, after fixing and de-hydration in an ethanol series, were embedded inLR-White resin. Transverse sections of polishedsurfaces of the rock were examined using a DSM 960 AZeiss microscope.
Confocal Scanning Laser Microscopy
For Confocal Scanning Laser Microscopy (CSLM),the rock samples were prepared according to a pro-cedure based on the glutaraldehyde-enhanced fluor-escence method used for plant tissues (Wierzchos &Ascaso 2001). In brief, the samples were first fixed inglutaraldehyde, dehydrated in a graded series of ethanolsolutions, and embedded in LR-White resin. Blocks ofresin-embedded rock samples were cut transversally,finely polished and observed using an LSM 310 Zeissconfocal microscope.
Fluorescence labelling
Fluorescence assays were performed on fresh piecesof rock containing endolithic lichens. The LIVE/DEADBacLight Kit L-13152 (Molecular Probes) was used todistinguish living and dead symbiont cells following themethod described in Wierzchos et al. (2004). This kitcontains two proprietary nucleic acid stains that differ intheir ability to penetrate bacterial cell membranes. Thegreen fluorescence nucleic acid stain, SYTO 9, labels allcells and the red fluorescence nucleic acid stain, pro-pidium iodide, only penetrates cells with damagedmembranes and quenches the green SYTO 9 stain.After fluorescence staining, the samples were processedfor CSLM examination. They were fixed in paraformal-dehyde, dehydrated in a graded ethanol series, andembedded in LR-White resin. The fluorescenceintensity of living and dead microorganisms was then
2 THE LICHENOLOGIST Vol. 37

determined by CSLM double-channel scanning usingan excitation wavelength of 488 nm. SYTO 9 (greenfluorescent dye) fluorescence intensity was measured atemission wavelengths between 515–545 nm for thedetection of living microorganisms, and fluorescenceintensity generated by propidium iodide was measuredat emission wavelengths longer than 570 nm to detectdead microorganisms. Each channel was displayed in asingle colour (green and red respectively) and theimages were superimposed using the Adobe Photoshopprogram. In the final image, cells with intact cellmembranes were stained only green whereas cells withdamaged membranes had red nuclei.
Results
Endolithic biota colonizing Antarcticgranite
The endolithic microecosystems of GraniteHarbour and the Dry Valleys materials ex-amined contained many different micro-organisms. In some samples, the symbiontsof endolithic lichens constituted the pre-dominant biota. These lichen dominated en-dolithic zones coincided with the presence ofepilithic lichens or areas where the fruitingbodies of endolithic lichens occurred (Fig.1A). Figure 1B provides a diagram of howthe algal and fungal cells are distributedthrough the fissures of the granite. The as-sociation observed between fungal and algalcells (Fig. 2A), as well as the connection inseveral cases with identified external fruitingbodies allows the identification of lichensymbiont cells. Cyanobacteria were also
observed but these occupied different zoneswithin the rocks and were not observed closeto the lichens. Heterotrophic bacteria werefrequently observed in cyanobacterium-richcommunities but were less common aroundendolithic lichen symbionts.
Most lichenized algal and fungal cells inthese endolithic habitats appeared embed-ded in a polymeric matrix. Figure 2B showsan endolithic mycobiont cell with numerousvacuoles within a thick extracellular layer(arrows). Extracellular polymeric substances(EPS) were detected around the differentmicroorganisms forming the biofilm. Theimage in Figure 2C, obtained by LTSEM,shows the polymeric nature of the extra-cellular compounds (arrows) associated withmycobiont cells at the interface between anepilithic lichen thallus and the lithic substra-tum. These EPS produced by epilithiclichens extended as far as the endolithicbiofilms observed in nearby fissures (arrowin Fig. 2D).
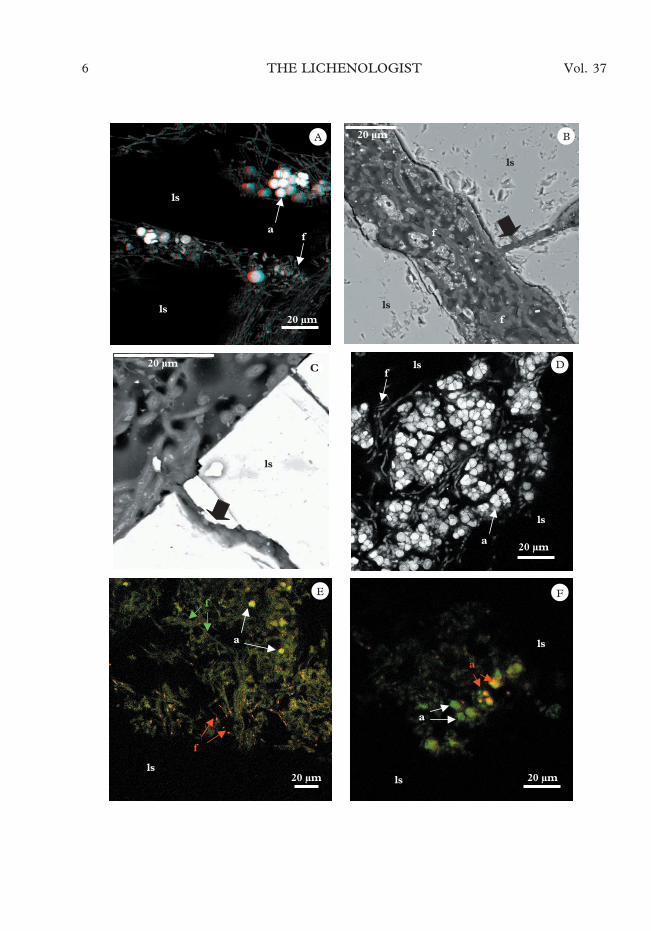
Several colonized fissures in granite fromboth geographical areas were seen to be fullyoccupied by lichenized algal and fungal cells(Figs 2A & 3A). Figure 3A is a 3-D imageobtained by CSLM showing the spatialarrangement of endoliths in different fissuresof a rock sample from Canada Glacier.Mycobiont cells were present in all fissuresoccupied by the lichen (Fig. 3B), even verynarrow fissures (Fig. 3C), but algal cells
F. 1. Distribution of endolithic lichen symbiont cells and their structures through the fissures. A, SEM-BSEimage of a rock colonized by Lecidea sp.: B, Model diagram corresponding to the area in Fig. 1A.
2005 Ecology of Antarctic endolithic lichens—de los Rı́os et al. 3
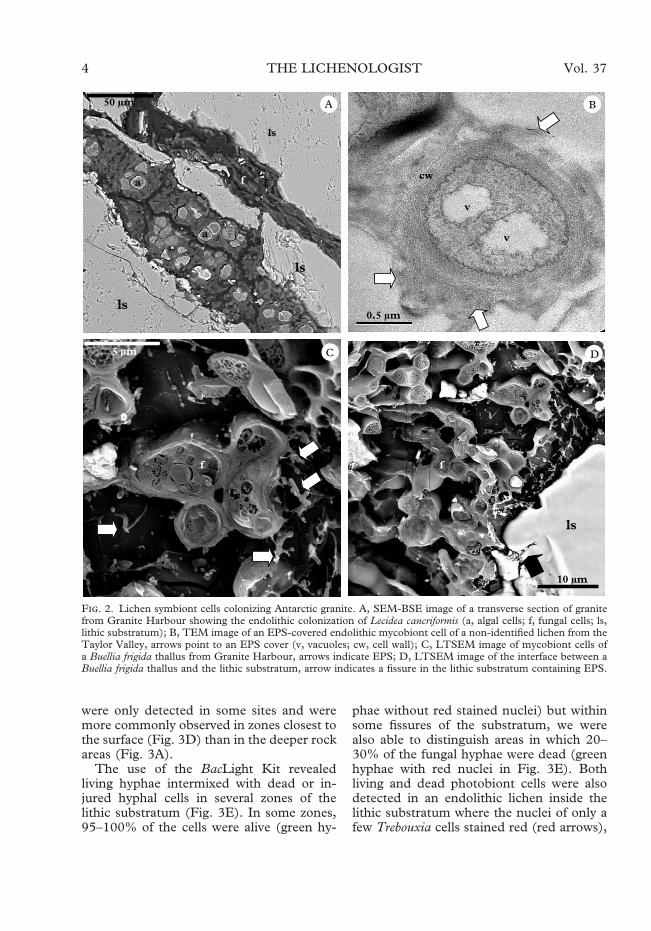
were only detected in some sites and weremore commonly observed in zones closest tothe surface (Fig. 3D) than in the deeper rockareas (Fig. 3A).
The use of the BacLight Kit revealedliving hyphae intermixed with dead or in-jured hyphal cells in several zones of thelithic substratum (Fig. 3E). In some zones,95–100% of the cells were alive (green hy-
phae without red stained nuclei) but withinsome fissures of the substratum, we werealso able to distinguish areas in which 20–30% of the fungal hyphae were dead (greenhyphae with red nuclei in Fig. 3E). Bothliving and dead photobiont cells were alsodetected in an endolithic lichen inside thelithic substratum where the nuclei of only afew Trebouxia cells stained red (red arrows),
F. 2. Lichen symbiont cells colonizing Antarctic granite. A, SEM-BSE image of a transverse section of granitefrom Granite Harbour showing the endolithic colonization of Lecidea cancriformis (a, algal cells; f, fungal cells; ls,lithic substratum); B, TEM image of an EPS-covered endolithic mycobiont cell of a non-identified lichen from theTaylor Valley, arrows point to an EPS cover (v, vacuoles; cw, cell wall); C, LTSEM image of mycobiont cells ofa Buellia frigida thallus from Granite Harbour, arrows indicate EPS; D, LTSEM image of the interface between aBuellia frigida thallus and the lithic substratum, arrow indicates a fissure in the lithic substratum containing EPS.
4 THE LICHENOLOGIST Vol. 37

indicating the damaged-membrane state ofthe cells (Fig. 3F).
Mineral components of biofilmscontaining endolithic lichens
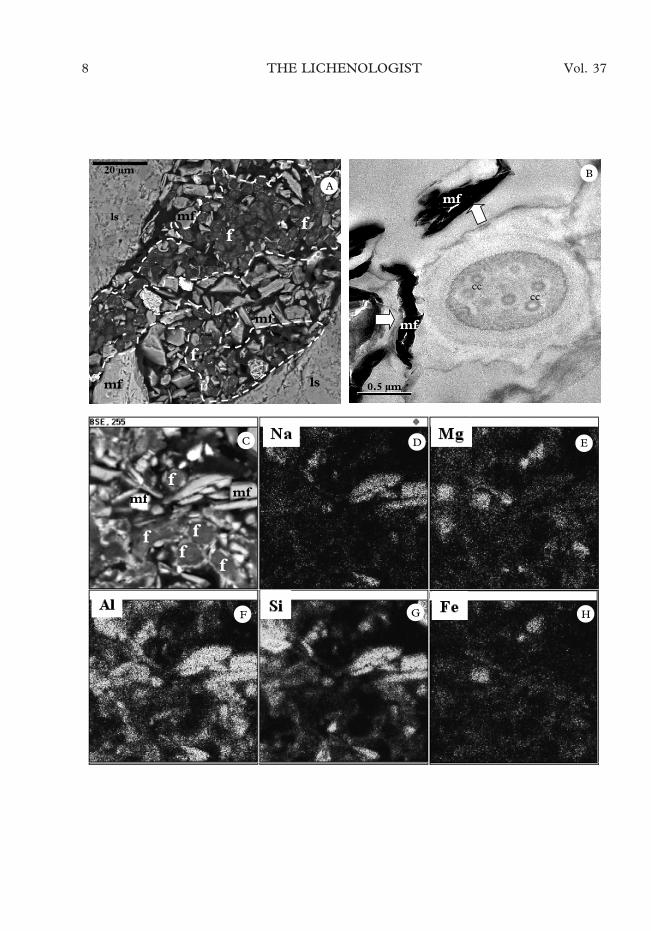
Mineral components mixed with the li-chen cells were observed in the fissures andcavities within the rock substratum. Some-times it was difficult to distinguish betweenthe part of a fissure occupied by mycobiontcells and that filled with mineral fragments(Fig. 4A). Aluminiosilicate minerals werefound to be especially closely associated withfungal cells (arrows in Fig. 4B). The elementdistribution map obtained by EDS analysisindicates that the minerals observed in theproximity of the cells were fairly hetero-geneous (Fig. 4C–H). Both closely associ-ated fine mineral fragments composed ofiron and magnesium and larger mineralfragments rich in aluminium and siliconwere observed. Aluminiosilicate fragmentscould be the result of physical weathering offissure walls but the source of the finelydisrupted minerals is unclear.
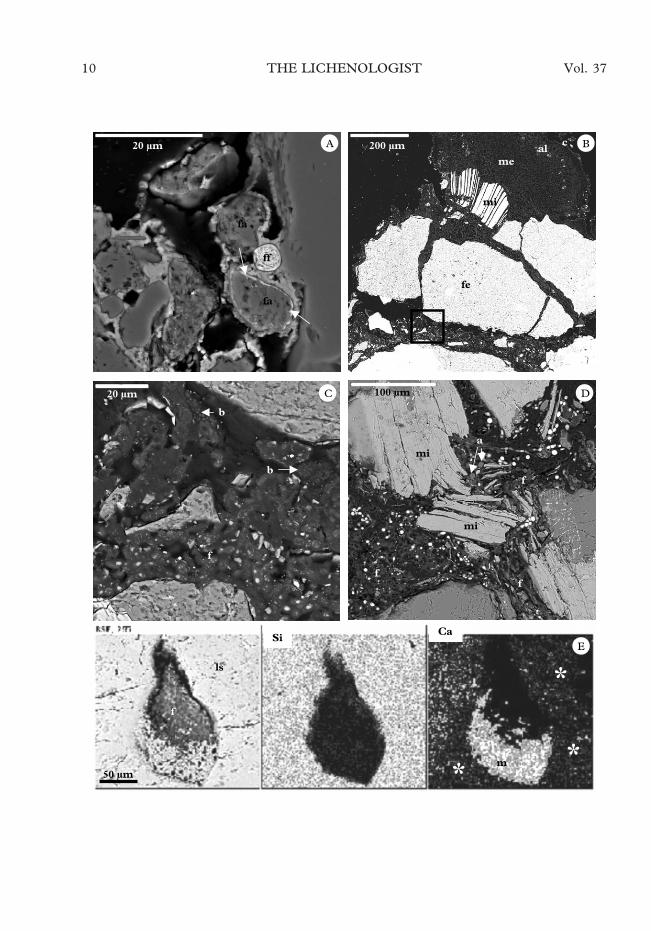
At the bottom of the Dry Valleys, livingendolithic lichens are scarce (Green et al.,1999) but their previous occurrence wasevidenced by the finding of mineralizedremains (fossils) of lichens (Fig. 5A),which comprise recognizable mineralizedstructures such as algal and fungal cells.
Relationships established betweencomponents of endolithicmicroecosystems
The relationships within the analyzedliving lichen communities seemed to be ex-tremely diverse. For instance, in Figure 5B,the growth of an epilithic thallus of Buelliafrigida is seen to give rise to several differentendolithic microhabitats where other micro-organisms thrive. Figure 5C is an enlarg-ment of the area indicated with a square inFig. 5B where bacteria are observed inter-mixed with fungal cells. Many of the micro-organisms were intimately linked to theirrock substratum generating complexmineral-microbe interactions, such that thelithic substratum was highly altered in the
proximity of different Antarctic endolithiclichens (Fig. 2A, 5D). Figure 5D clearlyshows the detachment and separation ofmica minerals. Penetration by fungal hyphaewas also observed in fissures in feldspar(arrows in Fig. 3B and Fig. 3C). Physicalweathering by endolithic lichens was alsofrequently associated with a biogeochemicalaction, and evidence of the biomobilizationof different elements and biomineralizationprocesses could be observed around the or-ganisms. Figure 5E shows a cavity occupiedby mycobiont cells of Lecidea cancriformis,with a mineral deposit visible at the base ofthe cavity (m). Calcium and silicon distri-bution maps obtained by EDS revealed alarge amount of calcium in the mineraldeposits observed within the cavity in con-trast to the silicon composition of the sub-stratum (feldspar plagioclase). Thus, thesedeposits could be identified as calciumoxalate biominerals. In the lithic substratum,zones with small amounts of calcium can bedetected (indicated by asterisks in Fig. 5E)reflecting intense calcium depletion in thisfeldspar plagioclase in an area close to thalliof Lecidea cancriformis (low-calcium zone in-dicated by asterisks). This biomobilizationwas clearly related to a biomineralizationprocess, as indicated by the extensive cal-cium oxalate deposits in the cavity colonizedby L. cancriformis.
Discussion
We have shown that endolithic lichens arecomponents of endolithic biofilms, whereclose chemical, physical and biological rela-tionships are established among symbiontcells and their immediate environment. Itwas not usually possible to identify clearlydifferentiated thallus layers in these endo-lithic lichens, although a degree of organiza-tion of the algal and fungal cells wasobserved. Endolithic lichens colonizingBeacon sandstones occupy the airspaces ofthe rock and do not form an organizedthallus (Friedmann 1982). The distributionpatterns shown by the symbiont cells in thepresent study were similar in granite fromthe extremely dry Canada Glacier site on the
2005 Ecology of Antarctic endolithic lichens—de los Rı́os et al. 5
6 THE LICHENOLOGIST Vol. 37

floor of Taylor Valley and the more equablegranites at Granite Harbour on the Ross Seacoast. This suggests that the distribution ofsymbiont cells in the fissures and cracks ofthe granite is not determined solely by theexternal environmental conditions, but alsoby factors differing on a small spatial scalesuch as the physico-chemical features of therock (Matthes et al. 2001; De los Ríos et al.2005). Different ecological niches can beattributed to the different habitats occupiedby endolithic lichens: cells of chasmoendo-lithic lichens colonize the fissures of thesubstratum, cryptoendolithic forms colonizeits cavities and euendolithic ones activelypenetrating the rock (Golubic et al., 1981).To a large extent the type of rock determinesthe microbial ecological niche. In the graniteanalysed here, the predominant ecologicalniche was chasmoendolithic (Fig. 1),whereas the higher porosity of sandstonepromotes the presence of cryptoendolithicforms in other lichen dominated communi-ties of the Ross desert (Friedmann 1982).
The endolithic environment appeared tobe altered by the activity of the microbialcells. Several mineral-lichen interactionswere observed in the form of geophysicaland geochemical effects. Fungi were thebiological components that produced themost intense geophysical weathering ofthe rocks, and mica was the mineral mostsusceptible to this bioalteration; for examplemica minerals become more modifed thanfeldspar and quartz. Fungi not only partici-pate in the alteration processes because oftheir physiological properties, but they alsoexert mechanical forces because of theirability to penetrate and spread through hardsubstrata (Sterflinger 2000) and due to theirwater binding capacity causing volume
changes in wet and dry periods (De los Ríoset al., 1999). The ability of epilithic lichenthalli to disaggregate the rock surface hasbeen frequently described (Banfield et al.1999; Aghamiri & Schwatzman 2002;Bjelland & Thorseth 2002; De los Ríos et al.2002). However, the real extent of theweathering capacity of endolithic lichens hasyet to be fully evaluated. Although fungalaction is probably not the only cause ofthe fissure formation, the images obtainedduring this study suggest that fungal pen-etration of granite could contribute to fissureformation and certainly be involved in sub-sequent widening of the fissure. The coloni-zation of fissures by fungal cells, as observedhere, and the growth of apothecia at thesurface (De los Ríos et al. 2005), can havedramatic biogeophysical effects on graniterocks.
The present work has found compellingevidence that endolithic lichens are involvedin biogeochemical processes occurring ingranite. Biogeochemical processes prevail inthe early stages of lichenization (Cooks &Otto 1990) and these may be similar in theendolithic lichens examined here, in which itwas not possible to distinguish well-differentiated lichen thallus layers. Calciumbiomobilization related to Antarctic endo-lithic lichens was observed in the presentstudy, and potassium mobilization in bi-otites and feldspar has also been recentlyascribed to endolithic Antarctic lichens (Delos Ríos et al. 2005). In addition to calciumoxalate, the fine minerals composed of ironand magnesium observed here around thecells are also likely to be the consequence ofbiomineralization processes. The presenceof Fe-rich diagenetic minerals and biogenicclays has been previously reported around
F. 3. Spatial distribution and vitality of endolithic lichen cells. A, CSLM 3-D image of an endolithic lichen fromthe Canada Glacier area (a, algal cells; f, fungal hyphae; ls, lithic substratum); B, SEM-BSE image of a deep fissurein a rock zone colonized by Lecidea cancriformis from Granite Harbour, black arrow denotes a narrow fissureoccupied by fungal cells; C, SEM-BSE image showing Lecidea sp. mycobionts colonizing a narrow fissure in a rockfrom Granite Harbour. D, CSLM image of a cavity close to the surface colonized by an endolithic lichen fromCanada Glacier. E & F, CSLM images of a colony from the Canada Glacier area stained with the BacLight kit andthen processed for CSLM; E, fissures showing the presence of a zone composed by dead hyphae (red arrows) inthe proximity of other living algal cells (white arrows) and fungal cells (green arrows); F, fissure showing a mixture
of living (white arrows) and dead (red arrows) photobiont cells.
2005 Ecology of Antarctic endolithic lichens—de los Rı́os et al. 7
8 THE LICHENOLOGIST Vol. 37

chasmoendolithic hyphae and bacterial cells(Wierzchos et al. 2003). Biomineralizationprocesses are related to the local chemicalcomposition of the substratum (Bjellandet al. 2002; De los Ríos et al. 2005) and cangive rise to chemical microenvironments thatcould affect the establishment of anothermicroorganism in the zone (De los Ríos et al.2002). These processes may have physio-logical consequences: biogenic mineralshave been found to completely envelopesymbiont cells or to occupy a large pro-portion of the colonized area, suggestingthese deposits could act as a barrier againstthe exchange of nutrients and affect nutrientcycling in the biofilm. The ‘fossilized’ cellsdescribed in endolithic systems could be theresult of these interactive processes ratherthan a change in the external habitat as hasbeen suggested (Friedmann et al. 1994). Inthe present work mycobiont cells were foundto be covered by EPS closely associated withmineral fragments, suggesting a certain levelof interaction of these compounds with rockcomponents and their possible involvementin the detected biogeochemical processes(Barker & Banfield 1996; Barker et al.1998). The formation of extracellular micro-bial polymeric layers may be controlled notonly by nutrient availability, but also by theavailability of metal ions to which the cell isexposed (Bevereidge 1989; Ferris 2000).
For the first time, we have been able toanalyse the physiological state of symbiontcells in situ, without removing the biofilmfrom the lithic substratum. This allows theprecise detection and localization of livingand dead cells within the lithic susbtratum.It was found that mycobiont and photobiontcells in different physiological states wereintermixed, and that the majority of cellswere viable. This confirms the results of aprevious study (De los Ríos et al. 2004) that
explored the physiological state of differentendolithic microorganisms in biofilms. Sucha high proportion of viable cells might indi-cate that these systems are very young andlittle dead material has yet accumulated, orthat they are very old and long-lived with avery low productivity and very long-livedcells. At present we cannot answer this ques-tion. Some endolithic systems have beenshown to be very old, even thousands ofyears (Johnston & Vestal 1991; Sun &Friedmann 1999) but dates of more samplesare needed in order to make any generaliza-tions. Few studies have assessed productivityon any time scale although it is suspectedthat productivity of endoliths is lower thanthat of epilithic lichens. The results ofFriedmann et al. (1993), derived from cryp-toendolithic communities, suggest that netecosystem productivity from long-termevents is only about 3 mg C m�2 y�1.
The biological success of these endolithiclichens requires the development of lifestrategies that will allow them to maintaintheir populations in this extreme habitat overa period of time (Block 1994). The survivalof endolithic lichens in continental Antarc-tica might be more a question of avoidingrather than tolerating the extreme conditionsoutside the microhabitat (Friedmann 1982;Friedmann et al. 1993; De los Ríos et al.2003; Cowan & Tow 2004). Production ofEPS is a survival mechanism and the matrixis thought to be one of the key biofilmcomponents involved in the protection ofthe component microorganisms from theenvironment (Davey & O’Toole 2000; Delos Ríos et al. 2004). Extracellular polymericsubstances are able to form microenviron-ments with conditions different from thoseof the bulk in terms of radiation received,capacity of water retention and pH, amongother factors, which could favour the
F. 4. Mineral components of biofilms containing endolithic lichens. A, SEM-BSE image of a fissure colonizedby mycobiont cells from Granite Harbour, dashed line outlines the part of the fissure occupied by mycobiont cells(a, algal cells; f, fungal hyphae; ls, lithic substratum, mf, mineral fragments); B, TEM image of an endolithicmycobiont cell of a non-identified lichen from Taylor Valley in the proximity of clay mineral fragments (arrows),note the fungal cells with several concentric bodies (cc); C-H, element distributions visualized by EDS in a fissureof a rock from Granite Harbour colonized by mycobiont cells of Lecidea sp.; C, SEM-BSE image; D, Na; E, Mg;
F, Al; G, Si; H, Fe.
2005 Ecology of Antarctic endolithic lichens—de los Rı́os et al. 9
10 THE LICHENOLOGIST Vol. 37

survival of lithobionts (De los Ríos et al.2003). In addition, EPS can be consideredlong-term accumulators of carbon and en-ergy (Wolfaardt et al. 1999) and are thus ofgreat interest in terms of adaptation to Ant-arctic conditions. The endolithic strategy isalso closely linked to the interactions of thelichen symbiont cells with the lithic substrata(Johnston & Vestal 1993). Interactions be-tween the components of the biofilms andtheir substrates determine the existence ofdifferent microhabitats and chemical micro-environments in these lithic microecosys-tems (Bungartz et al. 2004; De los Ríos et al.2002, 2003). The spatially close associationof endolithic symbiont cells with the lithicsubstratum, and the alteration of bedrockminerals and synthesis of biominerals ob-served in the antarctic granite analyzed, alsosupports this hypothesis.
Whilst lichen biogeochemical activity canlimit the bioavailability of some metals, itcan also release other nutrients to the micro-ecosystem; the present study is an exampleof how endolithic lichens can actively con-tribute to the mineral cycling (Wierzchos &Ascaso 1996, 1998). It is known that lichensoccupy extensive areas in the world, but thisarea is much greater if endolithic growthforms are also included in such global esti-mates. This means their global contributionto biogeochemical cycles both throughmineral cycling and metabolic activity mightneed to be re-assessed.
The authors thank Fernando Pinto, Charo Santos andSara Paniagua for technical assistance and Ana Burtonfor reviewing the English. This study was funded bygrants REN2003-07366-C01/C02 and BOS2003-02418 of the Plan Nacional I+D.
R
Aghamiri, R. & Schwartzman, D. W. (2002) Weather-ing rates of bedrock by lichens: a mini watershedstudy. Chemical Geology 188: 249–259.
Ascaso, C. & Wierzchos, J. (2003) The search forbiomarkers and microbial fossils in Antarctic rockmicrohabitats. Geomicrobiology Journal 20: 439–450.
Banfield, J. F., Barker, W. W., Welch, S. A. & Taunton,A. (1999) Biological impact on mineral dissolu-tion: application of the lichen model to understandmineral weathering in the rizosphere. Proceedings ofthe National Academy of Sciences 96: 3404–3411.
Barker, W. W. & Banfield, J. F. (1996) Biologicallyversus inorganically mediated weathering reac-tions: relationship between minerals and extra-cellular microbial polymers in lithobionticcommunities. Chemical Geology 132: 55–69.
Barker, W. W., Welch, S. A., Chu, S. & Banfield, J. F.(1998) Experimental observations of the effects ofbacteria on aluminiosilicate weathering. AmericanMineralogist 83: 1551–1563.
Beveridge, T. J. (1989) Role of cellular design inbacterial metal accumulation and mineralization.Annual Review of Microbiology 43: 147–171.
Bjelland, T., Saebo, L. & Thorseth, I. H. (2002) Theoccurrence of biomineralization products in fourlichen species growing on sandstone in westernNorway. Lichenologist 34: 429–440.
Bjelland, T. & Thorseth, I. H. (2002) Comparativestudies of the lichen-rock interface of four lichensin Vingen, western Norway. Chemical Geology 192:81–98.
Block, W. (1994) Terrestrial ecosystems: Antarctica.Polar Biology 14: 293–300.
Bungartz, F., Nash III, T. H. & Ryan, B. D. (2004)Morphology and anatomy of chasmolithic versusepilithic growth: a taxonomic revision of incon-spicuous saxicolous Buellia species from theSonoran Desert Region generally ascribed to the‘‘Buellia punctata’’ group. Canadian Journal ofBotany 82: 540–562.
Castello, M. & Nimis, P. L. (1997) Diversity of lichensin Antarctica. In Antarctic communities. Species,structure and survival (B. Battaglia, J. Valencia &D. W. H. Walton, eds.): 15–21. Cambridge:Cambridge University Press.
F. 5. Electron micrographs of Antarctic endolithic and epilithic lichens. A, SEM-BSE image of fossilized(mineralized) photobiont (fa) and mycobiont (ff) cells occupying a pore in sandstone rock from Mount Fleming,arrows point to a mineralized algal cell wall with large amounts of iron; B, SEM-BSE image of the interface madeby a Buellia frigida thallus and the surface of a Granite Harbour rock (c, upper cortex; al, algal layer; me, medulla;mi, mica; fe, feldspar); C, SEM-BSE image of the boxed area in Fig. 5B where bacterial (b) and fungal (f) cellsintermix under an epilithic lichen; D, endolithic fungal growth in a rock from Granite Harbour causing thedetachment and separation of mica layers (mi) (a, algal cells; f, fungal hyphae); E, SEM-BSE and EDS elementdistribution maps of a rock (Granite Harbour) zone colonized by mycobiont cells (f) of Lecidea cancriformis showingareas of plagioclase substratum with different amounts of calcium, suggesting the depletion of this element
(asterisks), and calcium rich mineral deposits (m) which could correspond to calcium oxalate accumulation.
2005 Ecology of Antarctic endolithic lichens—de los Rı́os et al. 11

Cooks, J. & Otto, E. (1990) The weathering effects ofthe lichen Lecidea aff. Sarcogynoides (Koerb.) onmagaliesberg quartzite. Earth Surface Processes andLandforms 15: 491–500.
Cowan, D. A. & Tow, L. A. (2004) EndangeredAntarctic environments. Annual Review of Micro-biology 58: 649–690.
Davey, M. E. & O’Toole, G. A. (2000) MicrobialBiofilms: from ecology to molecular genetics.Microbiology and Molecular Biology Reviews 64:847–867.
De los Ríos, A., Ascaso, C. and Wierzchos, J. (1999)Study of lichens with different state of hydrationby the combination of low temperature sacnningelectronand confocal laser scanning microscopies.International Microbiology 2: 251–257.
De los Ríos, A. & Ascaso, C. (2001) Preparativetechniques for transmission electron microscopyand confocal laser scanning microscopy of lichens.In Protocols in Lichenology (I. Kranner & R. P.Beckett & A. K. Varma, eds.): 87–151. Berlin:Springer.
De los Ríos, A., Wierzchos, J. & Ascaso, C. (2002)Microhabitats and chemical microenvironmentsunder saxicolous lichens growing on granite.Microbial Ecology 43: 181–188.
De los Ríos, A., Wierzchos, J., Sancho, L. G. & Ascaso,C. (2003) Acid microenvironments in microbialbiofilms of Antarctic endolithic microecosystems.Environmental microbiology 5: 231–237.
De los Ríos, A., Wierzchos, J., Sancho, L. G. & Ascaso,C. (2004) Exploring the physiological state ofcontinental Antarctic endolithic microorganismsby microscopy. FEMS Microbiology Ecology 50:143–152.
De los Ríos, A., Sancho, L. G., Grube, M., Wierzchos,J. & Ascaso, C. (2005) Endolithic growth of twoLecidea lichens in granite from continental Antarc-tica detected by molecular and microscopy tech-niques. New Phytologist 165: 181–190.
Ferris, F. G. (2000). Microbe-metal interactions insediments. In Microbial Sediments (R. E. Riding &S. M. Awramik, eds): 121–126. Berlin: Springer-Verlag.
Friedmann, E. I. (1982) Endolithic microorganisms inthe Antarctic cold desert. Science 215: 1045–1053.
Friedmann, E. I. & Ocampo-Friedmann, R. (1984).Endolithic microorganisms in extreme dry environ-ments: analysis of a lithobiontic microbial habitat.In Current Perspectives in Microbial Ecology Vol. XII(M. J. Klug & C. A. Reddy, eds.): 177–185.Washington, D.C.: American Society forMicrobiology.
Friedmann, E. I., Kappen, L., Meyer, M. A. & Nienow,J. A. (1993). Long-term productivity in the crypto-endolithic microbial community of the RossDesert, Antarctica. Microbial Ecology 25: 51–69.
Friedmann, E. I., Druk, A. Y. & Mckay, C. P. (1994)Limits of life and microbial extinction in theAntarctic desert. Antarctic Journal of USA 29:176–179.
Golubic, S., Friedmann, I. & Schneider, J. (1981) Thelithobiontic ecological niche, with special referenceto microorganisms. Journal of Sedimentary Petrology51: 475–478.
Green, T. G. A., Schroeter, B. & Sancho, L. G. (1999)Plant life in Antarctica. In Handbook of FunctionalPlant Ecology (F. I. Pugnaire & F. Valladares, eds.):495–543. New York, Basel: Marcel Dekker, Inc.
Johnston, C. G. & Vestal, R. (1991) Photosyntheticcarbon incorporation and turnover in Antarcticcryptoendolithic microbial communities: are theythe slowest growing communities on earth? Appliedand Environmental Microbiology 57: 2308–2311.
Johnston, C. G. & Vestal, R. (1993) Biogeochemistryof oxalate in the Antarctic cryptoendolithiclichen-dominated community. Microbial Ecology25: 305–319.
Matthes, U., Turner, S. J. & Larson, D. W. (2001)Light attenuation by limestone rock and its con-straint on the depth distribution of endolithic algaeand cyanobacteria. International Journal of PlantSciences 162: 263–270.
Øvstedal, D. O. & Lewis-Smith, R. I. (2001) Lichens ofAntarctica and South Georgia: A Guide to TheirIdentification and Ecology. Cambridge: CambridgeUniversity Press.
Reynolds, S. (1963) The use of lead citrate at high pHas an electron-opaque stain in electron microscopy.Journal of Cell Biology 17: 200–211.
Robinson, S. A., Wasley, J., & Tobin, A. K. (2003)Living on the edge – plants and global change incontinental and maritime Antarctica. GlobalChange Biology 9: 1681–1717.
Seppelt, R. D., Green, T. G. A. & Schroeter, B. (1995)Lichens and mosses from the Kar Plateau, South-ern Victoria Land, Antarctica. New Zealand Journalof Botany 33: 203–220.
Sterflinger, K. (2000) Fungi as geologic agents. Geo-microbiology Journal, 17: 97–124.
Sun, H. J. & Friedmann, E. I. (1999) Growth ongeological time scales in the Antarctic crytoendo-lithic microbial community. GeomicrobiologyJournal 16:193–202.
Wierzchos, J. & Ascaso, C. (1994) Application ofback-scattered electron imaging to the study of thelichen rock interface. Journal of Microscopy 175:54–59.
Wierzchos, J. & Ascaso, C. (1996) Morphological andchemical features of bioweathered granitic biotiteinduced by lichen activity. Clays and Clay Minerals44: 652–657.
Wierzchos, J. & Ascaso, C. (1998) Mineralogical trans-formation of bioweathered granitic biotite, studiedby HRTEM: evidence for a new pathway in lichenactivity. Clays and Clay Minerals 46: 446–452.
Wierzchos, J. & Ascaso, C. (2001) Life, decay andfossilisation of endolithic microorganisms from theRoss Desert, Antarctica: suggestions for in situfurther research. Polar Biology 24: 863–868.
Wierzchos, J., Ascaso, C., Sancho, L. G. & Green, A.(2003) Iron-rich diagenetic minerals are bio-
12 THE LICHENOLOGIST Vol. 37

markers of microbial activity in Antarctic rocks.Geomicrobiology Journal 20: 15–24.
Wierzchos, J., de los Rios, A., Sancho, L. G. & Ascaso,C. (2004) Viability of endolithic micro-organismsin rocks from the McMurdo Dry Valleys of Ant-arctica established by confocal and fluorescencemicroscopy. Journal of Microscopy 216: 57–61.
Wolfaardt, G. M., Lawrence, J. R. & Korber, D. R.(1999) Function of EPS. In Microbial ExtracellularPolymeric Substances (J. Wingender, T. R. Neu andH. C. Fleming, eds): 171–200. Berlin: Springer.
Accepted for publication 8 July 2005
2005 Ecology of Antarctic endolithic lichens—de los Rı́os et al. 13