respiration-induced weathering patterns of two endolithically growing lichens: respiration-induced...
TRANSCRIPT

Respiration-induced weathering patterns of two endolithicallygrowing lichensBETTINA WEBER,1 CLAUDIA SCHERR,1 FRITZ BICKER,1 THOMAS FRIEDL2 AND BURKHARD
BUDEL1
1Department of Biology, Plant Ecology and Systematics, University of Kaiserslautern, Kaiserslautern, Germany2Department of Experimental Phycology and SAG, University of Gottingen, Gottingen, Germany
ABSTRACT
The two endolithic lichen species Hymenelia prevostii and Hymenelia coerulea were investigated with regard to
their thallus morphology and their effects on the surrounding substrate. The physiological processes responsible
for the observed alterations of the rock were identified. Whereas the thallus surface of H. coerulea was level,
H. prevostii formed small depressions that were deepest in the thallus center. In a cross-section, both species
revealed an algal zone consisting of algal cavities parallel to the substrate surface and a fungal zone below. How-
ever, H. prevostii revealed significantly larger cavities with more than twice the cell number and a denser pattern
of cavities than H. coerulea, resulting in a biomass per surface area being more than twice as large. Below
H. prevostii the layer of macroscopically visibly altered rock material was about twice as deep and within this
layer, the depletion of calcium and manganese was considerably higher. In simultaneous measurements of the
oxygen uptake ⁄ oxygen release and pH shift, the isolated algal strains of both lichens revealed respiration-
induced acidification of the medium in the dark. At higher light intensities, H. coerulea and to a lesser extent also
H. prevostii alkalized the medium which may lessen the acidification effect somewhat under natural conditions.
In a long-term growth experiment, the isolated algal strains of both lichens revealed acidification of the medium
to a similar extent. Neither acidic lichen substances nor oxalic acid was identified. The significant differences
between the weathering patterns of both species are based on the same respiration-induced acidification mecha-
nism, with H. prevostii having a greater effect due to its higher biomass per area.
Received 2 March 2010; accepted 12 July 2010
Corresponding author: B. Weber. Tel.: +49 (0)631-205-2810; fax: +49 (0)631-205-2998; e-mail: weberb@
rhrk.uni-kl.de
INTRODUCTION
Lichens, representing symbiotic organisms of a fungus
together with at least one algal or cyanobacterial species, may
occupy different places within the lithobiontic habitat. Besides
the epilithic growth, where the substrate mainly serves as
holdfast, several endolithic growth forms can be distinguished
(Golubic et al., 1981). Chasmoendolithic organisms live
within existing cracks and fissures of the rock, whereas crypto-
endoliths inhabit existing cavities or pore spaces inside the
substrate matrix. In contrast, euendolithic organisms actively
create the cavities they live in by mechanical and ⁄ or chemical
action.
The occurrence of euendolithic lichens on calcareous sub-
strates has been described for a wide variety of geographic
regions, like the Negev Highlands in Israel (Danin & Garty,
1983), the Trieste karst in Northern Italy (Tretiach, 1995),
the Dachstein Mountains of the Alps (Pohl & Schneider,
2002), the Mendip Hills in England (Viles, 1987), the lime-
stone coasts of the northern Adriatic Sea (Istria, Croatia;
Schneider, 1976) and on desert pavement of the central Sono-
ran Desert (Bungartz et al., 2004). Also on calcareous build-
ing stones, like limestone walls in Jerusalem and marble
monuments in Rome (Danin & Caneva, 1990) endolithic
lichens are known to be a common feature. Within lithobion-
tic biofilms, endolithic lichens form communities together
with cyanobacteria, algae, epilithic algae and fungi in varying
proportions (Pohl & Schneider, 2002).
The mechanical or physical weathering patterns induced by
lichens can be separated in the processes of (1) penetration of
hyphae, sometimes leading to the incorporation of mineral
fragments into the thallus; (2) expansion and contraction of
the thallus, induced by water or ice; and (3) swelling of
organic or inorganic salts (Chen et al., 2000). Chemical
weathering mechanisms are induced by (1) oxalic acid;
(2) lichen compounds, mostly referred to as lichen acids; (3)
34 � 2010 Blackwell Publishing Ltd
Geobiology (2011), 9, 34–43 DOI: 10.1111/j.1472-4669.2010.00256.x

alkalinolysis, as observed in endolithic cyanobacteria (Budel
et al., 2004); and (4) respiratory CO2, resulting in the forma-
tion of carbonic acid in an aqueous environment (Brunsden,
1979; Golubic & Schneider, 1979; Berthelin, 1988; Chen
et al., 2000).
Observations of weathering patterns on limestone triggered
by lichens have been described for multiple cases, first in
1880, when Sollas observed minute hemispherical pits on
exposed limestone surfaces, that where produced by the
apothecia of the endolithic lichen Verrucaria rupestris
(=V. muralis Ach.). Funfstuck (1895) found hyphae of the
endolithically growing Verrucaria marmorea Scop. as deep as
19 mm within the calcareous substrate and in 1890, Bach-
mann proved by microscopical thin sections that Verrucaria
calciseda DC. grows actively into the rock refuting the
hypothesis of Zukal (1884), that the calcareous particles were
excreted by the organism. Also in later times, substrate-
dissolving calcicolous lichens and the resulting patterns were
described (e.g. Pia, 1937; Doppelbaur, 1959; Danin et al.,
1983; Gehrmann et al., 1992; Blazquez et al., 1995). With
regard to the underlying mechanisms, oxalic acid and lichen
substances have frequently been identified and claimed
responsible for the observed weathering patterns (e.g. Schatz,
1963; Ascaso et al., 1982). Respiratory CO2, however, has
been assumed to be of only minor importance in weathering
processes (Syers & Iskandar, 1973; Jones & Wilson, 1985). In
situ measurements of respiration-induced pH-shifts as possi-
ble weathering agents are to our knowledge restricted to one
study on the alkalization of sandstone (Budel et al., 2004).
In contrast to the discussed weathering effects, some
authors (Hoppert et al., 2004; Pohl & Schneider, 2005) pro-
pose an overall protective effect of endolithic lichens on the
rock, as they also found mechanisms that stabilize and pre-
serve the rock surface morphology. An overall protective effect
of the calcicolous epilithic species Aspicilia calcarea (L.) Korb.
and Diploschistes diacapsis (Ach.) Lumbsch was also described
by Mottershead & Lucas (2000), who observed conical
mounds colonized by these lichens on a flat gypsum surface.
However, a mechanism for this observed overall protective
effect could not be given by the authors.
The aim of this study was to investigate the effects of
H. prevostii and H. coerulea on mineral weathering and to
explore the underlying species-specific morphological and
physiological differences. As carbonates are easily soluble by
acids and weathering patterns on these rocks are widespread
and common, we wanted to explore if they are caused by spe-
cific substances (like lichen-acids or oxalic acid) or if respira-
tion-induced acidification of the substrate is sufficient to
provoke these processes.
MATERIAL
The two crustose endolithic lichen species Hymenelia prevostii
and Hymenelia coerulea (DC.) Massal. were collected in the
eastern Alpine mountains, at the Untersberg (47�43¢N,
13�03¢E) south of the Austrian city Salzburg. The Untersberg
consists of Dachsteinkalk (limestone) and has two summits,
the Berchtesgadener Hochthron (1972 m) and the Salzburg-
er Hochthron (1853 m). The samples were collected at a
steep westward facing outcrop, only a few meters below the
Salzburger Hochthron (Fig. 1A).
Hymenelia prevostii has an oval, crustose, whitish-gray thal-
lus. Brownish-pink apothecia (=fruiting bodies) are sunk into
the thallus, separated from it by a narrow crack (Fig. 1B).
Hymenelia coerulea is characterized by its bluish to bluish-gray
thallus that in some areas remains almost completely below
the surface of the substrate. The mostly rimless apothecia are
small (0.2–0.4 mm) and are often situated on small elevations
within the limestone (Fig. 1C).
METHODS
Structural and molecular characterization of the endolithic
habitat
On eight representative cross-sections of each endolithic
lichen species, the distance of the upper and lower termina-
tion of the algal zone from the rock surface, the number of
cavities lodging algae per 5 mm distance, the number of algal
cells per cavity and the distance between the rock surface and
the macroscopically non-modified rock were examined under
an inverted transmitted-light microscope (Axiovert 200M;
Carl Zeiss, Jena, Germany). Differences between the two
species were tested for statistical significance by the Mann–
Whitney U-test utilizing SPSSTM 16.0 (SPSS Inc., Chicago,
IL, USA).
Isolation of algae
The algal partners of both lichen species were isolated for
later molecular and physiological analyses: from a clean
thallus edge, small lichen pieces were removed with a
razor blade and carefully squeezed between a microscope
slide and cover slip to obtain a green suspension of algal
cells and fungal hyphae. Under the microscope, a group
of algal cells was then absorbed with a pipette and trans-
ferred to a petri dish with Bold’s Basal Medium (MBB;
Friedl, 1989) at pH 5.5 solidified with 1.5% agar. After
3–4 weeks, algal colonies became visible with the bare eye
and were streaked on new plates for purification and to
obtain single-cell based colonies.
The algae were grown in liquid MBB medium (Friedl,
1989) with 1 mM HEPES buffer to promote the growth of
algae and to impede that of fungi. All cultures were main-
tained at about 20 lE m)2 s)1 at a light–dark regime of
14:10 hours and 17 �C. The cultures in Erlenmeyer flasks
were shaken several times per day to allow gas exchange with
the atmosphere. Every 6–8 weeks the cultures were trans-
ferred to fresh medium.
Respiration-induced weathering of lichens 35
� 2010 Blackwell Publishing Ltd
Molecular determination of photobionts
DNA was extracted from algal cells of both Hymenelia photo-
biont strains as described by Muller et al. (2005) and the ITS-
1, 5.8S and ITS-2 regions of the nuclear-encoded rDNA were
amplified as described in Friedl (1996). Cycle sequencing was
performed with the ABI PRISM� Big DyeTM Terminator
Cycle Sequencing Kit v2.0 (Applied Biosystems, Foster City,
CA, USA) with primers as in Mikhailyuk et al. (2008). The
sequencing reactions were analyzed on an ABI 3100 DNA
Sequencer (Applied Biosystems) and the individual sequences
were assembled into contigs using SeqAssem (Hepperle,
2004). For comparison, also the ITS rDNA sequence of strain
Asterochloris phycobiontica Tschermak-Woess SAG 26.81
(obtained from the Culture Collection of Algae at Gottingen
University, Germany) was determined using the same
methods. Sequence alignments were prepared by eye using
BIOEDIT v.6.0.7 (Hall, 1999). To find the next closest rela-
tives of the Hymenelia photobionts, the newly determined
sequences were compared to the NCBI database (http://
www.ncbi.nlm.nih.gov/) using BLAST (Altschul et al., 1990).
The sequences of the next closest relatives were incorporated
into the alignment and corrected (Kimura two-parameter
model of nucleotide substitution); pairwise genetic distances
were calculated in MEGA 3.1 (Kumar et al., 2004).
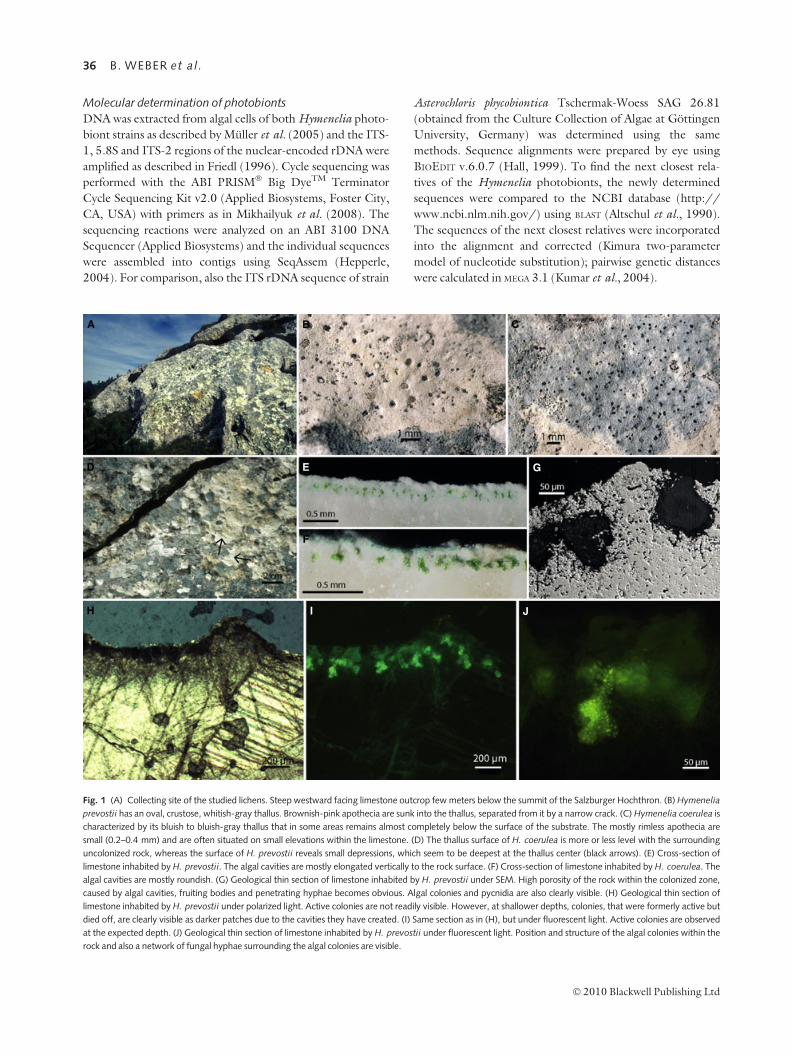
A B C
D E
I JH
F
G
Fig. 1 (A) Collecting site of the studied lichens. Steep westward facing limestone outcrop few meters below the summit of the Salzburger Hochthron. (B) Hymenelia
prevostii has an oval, crustose, whitish-gray thallus. Brownish-pink apothecia are sunk into the thallus, separated from it by a narrow crack. (C) Hymenelia coerulea is
characterized by its bluish to bluish-gray thallus that in some areas remains almost completely below the surface of the substrate. The mostly rimless apothecia are
small (0.2–0.4 mm) and are often situated on small elevations within the limestone. (D) The thallus surface of H. coerulea is more or less level with the surrounding
uncolonized rock, whereas the surface of H. prevostii reveals small depressions, which seem to be deepest at the thallus center (black arrows). (E) Cross-section of
limestone inhabited by H. prevostii. The algal cavities are mostly elongated vertically to the rock surface. (F) Cross-section of limestone inhabited by H. coerulea. The
algal cavities are mostly roundish. (G) Geological thin section of limestone inhabited by H. prevostii under SEM. High porosity of the rock within the colonized zone,
caused by algal cavities, fruiting bodies and penetrating hyphae becomes obvious. Algal colonies and pycnidia are also clearly visible. (H) Geological thin section of
limestone inhabited by H. prevostii under polarized light. Active colonies are not readily visible. However, at shallower depths, colonies, that were formerly active but
died off, are clearly visible as darker patches due to the cavities they have created. (I) Same section as in (H), but under fluorescent light. Active colonies are observed
at the expected depth. (J) Geological thin section of limestone inhabited by H. prevostii under fluorescent light. Position and structure of the algal colonies within the
rock and also a network of fungal hyphae surrounding the algal colonies are visible.
36 B. WEBER et al.
� 2010 Blackwell Publishing Ltd

Preparation and analysis of geological thin sections
Rock material with inhabiting organisms (n = 3) was fixated,
dehydrated and embedded in acrylic resin (LR-white;
London Resin Company, London, UK) according to
Bungartz et al. (2004). All steps were conducted in a vacuum
chamber. The embedded samples were polished on one side,
fixed to glass slides and ground down to a thickness of
approximately 50 lm and screened with a covering glass.
Sections were examined with a light microscope (Axiophot;
Zeiss, Oberkochen, Germany) under white, polarized and
fluorescent light and with a scanning electron microscope
(SEM; Hitachi S 2300, Krefeld, Germany) equipped with
energy dispersive X-ray spectroscopy (EDX; Rontec GmbH,
Berlin, Germany) and WinDISS-ColorMAP analytical
software (Point Electronic GmbH, Halle ⁄ Saale, Germany).
Elemental analysis
To analyze possible modifications of the substrate by the
inhabiting organisms, carbonized thin sections were prepared
in the same manner as geological thin sections (see above). In
the last step, samples were ground down and polished to a
thickness of 35 lm with carborundum.
Mineral components within rock samples were analyzed
with an X-ray microprobe system (SX 50; Cameca, Paris,
France) at an acceleration voltage of 15 kV and a ray current
of 10 nA. Measurements were performed in a Faraday cage,
shielding the sample off the ray between measurements. The
following standards were used: dolomite ([MgCa]CO3) for
Mg, calcite (CaCO3) for Ca, MnTiO3 for Mn, ferric oxide
(Fe2O3) for Fe and a synthetic andradite (Ca3Fe2(SiO4)3) for
Si. Analyzed values were converted into the corresponding
compounds using conversion factors according to Kuster &
Thiel (1985).
In order to see possible variations in element concentration
caused by the organisms, perpendicular transects from the
rock surface into the unmodified rock were established at
5 lm steps (H. coerulea: n = 5; H. prevostii: n = 3). In each
transect, microscopical observation allowed to define a depth
at which the alternating effect of the organisms ceased. For
each element, the values above and below this height were
pooled within each species and tested on significant differ-
ences utilizing a paired t-test in SPSSTM 16.0.
Unfortunately, there were no uncolonized rock surfaces at
our study site, allowing a comparison to surfaces without bio-
logical weathering effects.
Analysis of different growth and weathering mechanisms
Short-term O2 and pH development
In order to analyze photosynthesis- and respiration-induced
short-term pH shifts, about 2 mL of algal suspension in liquid
MBB medium was transferred to an oxygen electrode cham-
ber (DW1; Hansatech Instruments, England, UK) mounted
on its control unit (Oxyview Electrode Control Unit; Hansa-
tech Instruments). A hole in the lid of the electrode chamber
was enlarged to a diameter of about 2 mm for a pH minitrode
(Hamilton Bonaduz AG, Bonaduz, Switzerland) to fit snugly
through. The electrode was controlled and calibrated with a
pH meter (PHM 210; Radiometer Analytical, Lyon, France).
The samples were measured at variable light intensities
(given with a halogen lamp) and in the dark. Simultaneous
measurements of O2 content and pH were taken every 2 min
and after 8–10 min, the light intensity was changed. Mean
values of the O2 content were calculated from three sensor
readings in equilibrium, pH changes were summarized over
6 min and then upscaled to hourly changes. O2 contents and
pH changes were related to 1 mL of solution with the respec-
tive chlorophyll contents.
Of each algal strain, three sample replicates were measured.
For chlorophyll determination, the samples were filtered to
remove the aqueous faction and then extracted two times with
dimethyl sulfoxide (DMSO) at 65 �C for 45 min. The chloro-
phylla+b content was determined and calculated according to
Ronen & Galun (1984).
Long-term pH measurements
To analyze long-term pH changes in the medium, algal colo-
nies of both strains were derived from liquid culture by centri-
fugation. The colonies were washed two times in MBB
medium, then 2 mL suspension was added to 50 mL sterile
solution of pure MBB medium, MBB medium with 3 mM
HEPES buffer, and the buffered solution with 10 mM NaH-
CO3 added, respectively. The pH-value was set to 6.5 and the
cultures were kept at 17 �C with light of approximately
60 lE m)2 s)1 photon flux density and a light–dark regime of
14:10 hours. In regular intervals, under sterile conditions
2 mL solution was taken from each culture and the pH was
determined as the mean of two single measurements (TS-1;
Suntex Instruments Co., Taipei, Taiwan).
Thin layer chromatography
Thin layer chromatography for the identification of secondary
lichen compounds was conducted according to Culberson &
Kristinsson (1970), Culberson (1972) and Culberson &
Johnson (1982) applying solvents A, B and C.
Identification of oxalate
In order to analyze the growth zone of the organisms with
regard to the presence of calcium oxalate, 2 mg of rock mate-
rial from the growth zone was finely ground and homoge-
nized with 200 mg of potassium bromide. The material was
compacted and analyzed by IR-Spectroscopy (FT-IR Spec-
trum BX; Perkin Elmer, Waltham, MA, USA). For compari-
son purposes, a second sample, consisting of 2 mg potassium
oxalate with 200 mg of potassium bromide was analyzed.
Respiration-induced weathering of lichens 37
� 2010 Blackwell Publishing Ltd
RESULTS
Structural and molecular characterization of the endolithic
habitat
Macroscopic observations of the species in their natural habi-
tat revealed the thallus surface of H. coerulea to be more or
less level with the surrounding uncolonized rock, whereas the
surface of H. prevostii was characterized by small depressions,
which seemed to be deepest at the thallus center (Fig. 1D).
In cross-sections, it was observed that the algal partners of
both endolithic species live in cavities within the otherwise
dense limestone rock. The photobionts of both lichen
species were found to have identical ITS rDNA sequences
and belong to the genus Asterochloris Tschermak-Woess.
The highest sequence similarities (>99%) were found with
A. phycobiontica SAG 26.81 (sequences AM900490 and
GU017647), A. erici (Ahmadjian) Skaloud et Peksa UTEX
911 (AF345440) and an unidentified Asterochloris species,
strain LEP 28 (AM906010). Between the Hymenelia photo-
bionts and the latter three strains were no more than eight
sequence positions different, corresponding to 0.011 and
0.013 corrected pairwise distances. Assuming that ITS
rDNA provides molecular signatures clear enough for the
assignment to a certain species, this indicates that the Hyme-
nelia photobionts may represent a yet undescribed indepen-
dent species of Asterochloris.
The algal cavities of H. prevostii were mostly elongated
perpendicular to the rock surface, whereas those of H. coeru-
lea were more or less roundish (Fig. 1E,F). The algal zone
of H. prevostii started about twice as deep underneath the
rock surface and also reached deeper than that of H. coerulea
(Table 1). In H. prevostii the spatial density of cavities was
more than one-third higher than in H. coerulea. With a
mean value of 123 (n = 100) cells per cavity in H. prevostii,
compared to 47 ± 32.01 (n = 96) cells in H. coerulea,
the cell number within the cavities was also considerably
higher in H. prevostii. The thickness of the visibly altered
rock substrate was about twice as large in H. prevostii
(Table 1, Fig. 1E,F). All differences were highly significant
(P < 0.001, n = 40, Table 1). The substrate above the algal
zones looked whitish and fine-grained in contrast to the
underlying more translucent part (Fig. 1E,F).
In geological thin sections using fluorescent light, the
colonies active at the time of preparation were clearly visi-
ble in the expected depth (Fig. 1I). Using polarized light,
those colonies were difficult to image. However, at shal-
lower depths, colonies that had been active previously but
meanwhile had died off were clearly visible as darker areas
(Fig. 1H). At larger magnification, the position and struc-
ture of the algal colonies within the rock and also a net-
work of fungal hyphae surrounding the algal colonies was
observed (Fig. 1J).
Under SEM, the high porosity of the rock within the colo-
nized zone, caused by algal cavities, fruiting bodies and pene-
trating hyphae became obvious. The algal colonies and
pycnidia were also clearly visible (Fig. 1G).
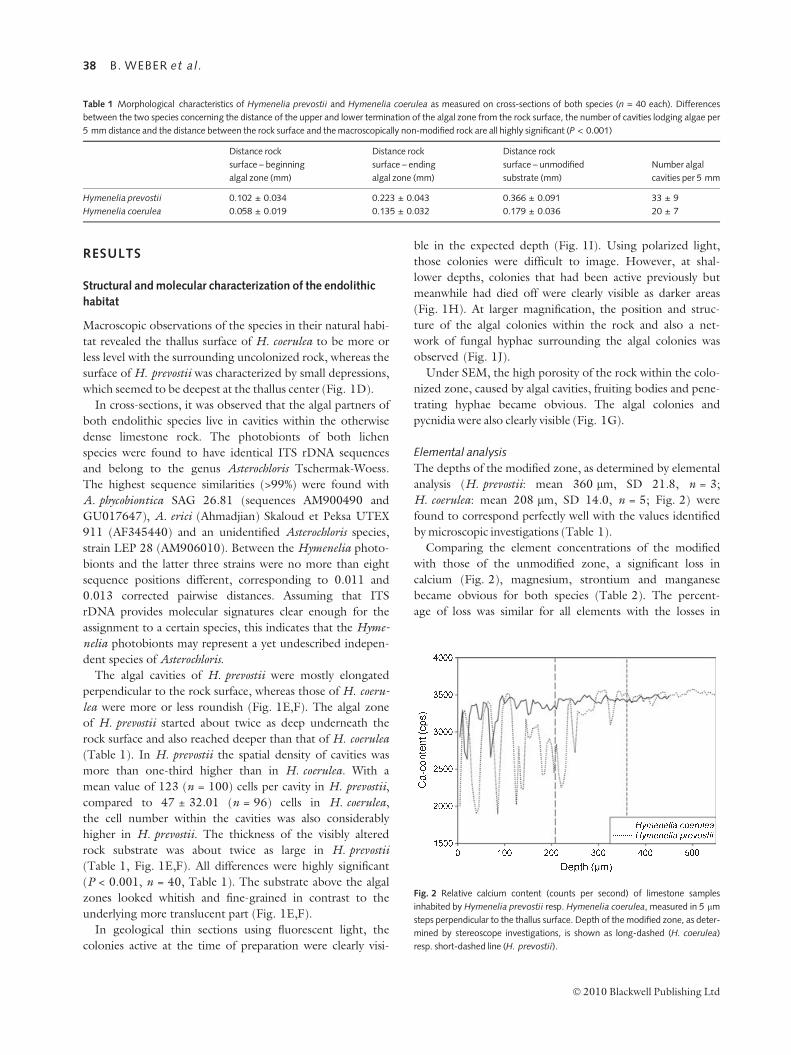
Elemental analysis
The depths of the modified zone, as determined by elemental
analysis (H. prevostii: mean 360 lm, SD 21.8, n = 3;
H. coerulea: mean 208 lm, SD 14.0, n = 5; Fig. 2) were
found to correspond perfectly well with the values identified
by microscopic investigations (Table 1).
Comparing the element concentrations of the modified
with those of the unmodified zone, a significant loss in
calcium (Fig. 2), magnesium, strontium and manganese
became obvious for both species (Table 2). The percent-
age of loss was similar for all elements with the losses in
Fig. 2 Relative calcium content (counts per second) of limestone samples
inhabited by Hymenelia prevostii resp. Hymenelia coerulea, measured in 5 lm
steps perpendicular to the thallus surface. Depth of the modified zone, as deter-
mined by stereoscope investigations, is shown as long-dashed (H. coerulea)
resp. short-dashed line (H. prevostii).
Table 1 Morphological characteristics of Hymenelia prevostii and Hymenelia coerulea as measured on cross-sections of both species (n = 40 each). Differences
between the two species concerning the distance of the upper and lower termination of the algal zone from the rock surface, the number of cavities lodging algae per
5 mm distance and the distance between the rock surface and the macroscopically non-modified rock are all highly significant (P < 0.001)
Distance rock
surface – beginning
algal zone (mm)
Distance rock
surface – ending
algal zone (mm)
Distance rock
surface – unmodified
substrate (mm)
Number algal
cavities per 5 mm
Hymenelia prevostii 0.102 ± 0.034 0.223 ± 0.043 0.366 ± 0.091 33 ± 9
Hymenelia coerulea 0.058 ± 0.019 0.135 ± 0.032 0.179 ± 0.036 20 ± 7
38 B. WEBER et al.
� 2010 Blackwell Publishing Ltd
the presence of H. prevostii being markedly higher than
those of H. coerulea. Strikingly, for iron, no difference was
observed within the modified zone of H. prevostii, whereas
a significant increase was registered for H. coerulea.
Analysis of different growth and weathering mechanisms
Short-term O2 and pH development
During measurements with the oxygen electrode, all sam-
ples containing algal suspensions isolated from H. coerulea
(Fig. 3A–C) and H. prevostii (Fig. 3D–F) revealed oxygen
consumption accompanied by acidification in the dark.
Above a certain light intensity (the so-called compensation
point) all samples produced oxygen and at higher light
intensities they also alkalized the solution. For all samples,
a clear correlation between oxygen uptake ⁄ release and a
pH shift is obvious and for some of them (Fig. 3A,B,D)
this correlation fits almost perfectly. All samples of H. co-
erulea alkalized the medium at relatively low light intensi-
ties (<50 lE m)2 s)1), whereas only one of the
H. prevostii samples alkalized it at 50 lE. Between the
physiological activity and the chlorophyll content of the
samples no clear correlations were observed.
Long-term pH measurements
In liquid cultures of both Asterochloris strains, the pH of the
surrounding medium decreased by 1.5–2 within 6 weeks,
with the unbuffered cultures reacting faster than the buffered
ones (Table 3). Despite the low pH, all cultures looked green
and healthy after 6 weeks’ time.
In the buffered cultures with bicarbonate, the pH increased
by 1.7 within 3 weeks. Already at this stage it was visible that
the cultures started to bleach and after 6 weeks they had died
off (Table 3).
Thin layer chromatography
In both Hymenelia species, no acidic lichen compounds were
detected. In H. prevostii, two unknown terpenoids were pres-
ent (1. A: 5, B: 6, C: 6, UV) ⁄ + ‘salmon pink’; 2. A: 6, B: 7, B:
7, UV) ⁄ + grayish white). In H. coerulea, no lichen com-
pounds were found.
Identification of oxalate
In none of the two Hymenelia species oxalate was identified,
as the samples did not show the characteristic spectral bands at
3000–3500, 1645, 1320 and 782 cm)1, respectively.
DISCUSSION
Although the two lichen species H. prevostii and H. coerulea
display a common thallus morphology with an algal zone
consisting of algal cavities parallel to the substrate surface, they
both revealed clear anatomical and morphological differences:
in H. prevostii the number of cavities per 5 mm substrate
was 1.65 times and the number of cells within each cavity
2.62 times higher. This corresponds very well to their chloro-
phyll values per surface area, which are more than twice as high
for H. prevostii (322,4 ± 39.4 mg Chla+b m)2, n = 3 vs.
H. coerulea: 141.7 ± 2.2 mg Chla+b m)2, n = 2) as analyzed
in a previous study (Weber et al., 2007).
The algal zone of H. prevostii was found to start twice as
deep and to be almost twice as large compared with H. coeru-
lea. If light attenuation within the thallus was comparable in
both species, the algae of H. prevostii would grow under lower
light intensities. The results of previous gas exchange mea-
surements, however, revealed that H. prevostii reaches the
light compensation point earlier (75 lE m)2 s)1 vs.
150 lE m)2 s)1) and is saturated (90% of maximum NP) at
higher light intensities than H. coerulea (1000 lE m)2 s)1 vs.
800 lE m)2 s)1) which are typical characteristics of high-
light organisms (Weber et al., 2007). A solution to this anta-
gonism is found if one re-inspects the thallus surface of both
species. Whereas in H. prevostii with its white thallus surface a
large percentage of light will be transmitted into deeper parts
of the wet thallus, the bluish surface of H. coerulea turns into
a dark-blue surface when wetted, absorbing a large percentage
of the incident light. Therefore, it can be expected, that at the
same depth light intensities are higher within H. prevostii
allowing for a larger algal zone that also reaches deeper into
the substrate.
For both Hymenelia species identical ITS rDNA sequences
were found, suggesting that they belong to the same species
of the genus Asterochloris (Skaloud & Peksa, 2010) or even
are genetically identical. Consequently, the observed morpho-
Table 2 Relative element concentrations (counts per 10 s) as determined by element analysis
Concentration
(counts per 10 s)
Hymenelia prevostii Hymenelia coerulea
Modified zone
(n = 167)
Unmodified
zone (n = 172) Percentage
Modified
zone (n = 187)
Unmodified
zone (n = 257) Percentage
Ca 31 724.8*** 35 366.7 89.7 32 205.1*** 34 093.9 94.5
Mg 112.9*** 127.7 88.4 175.6** 184.1 95.4
Fe 34.1 34.4 99.1 37.1* 35.5 104.5
Sr 342.7*** 367.0 93.4 347.3*** 357.8 97.1
Mn 270.5*** 286.0 94.6 302.0*** 316.4 95.4
Significant differences between original and modified zone of the rock are indicated by asterisks (level of significance 5%: *; 1%: **; 0.5%: ***).
Respiration-induced weathering of lichens 39
� 2010 Blackwell Publishing Ltd
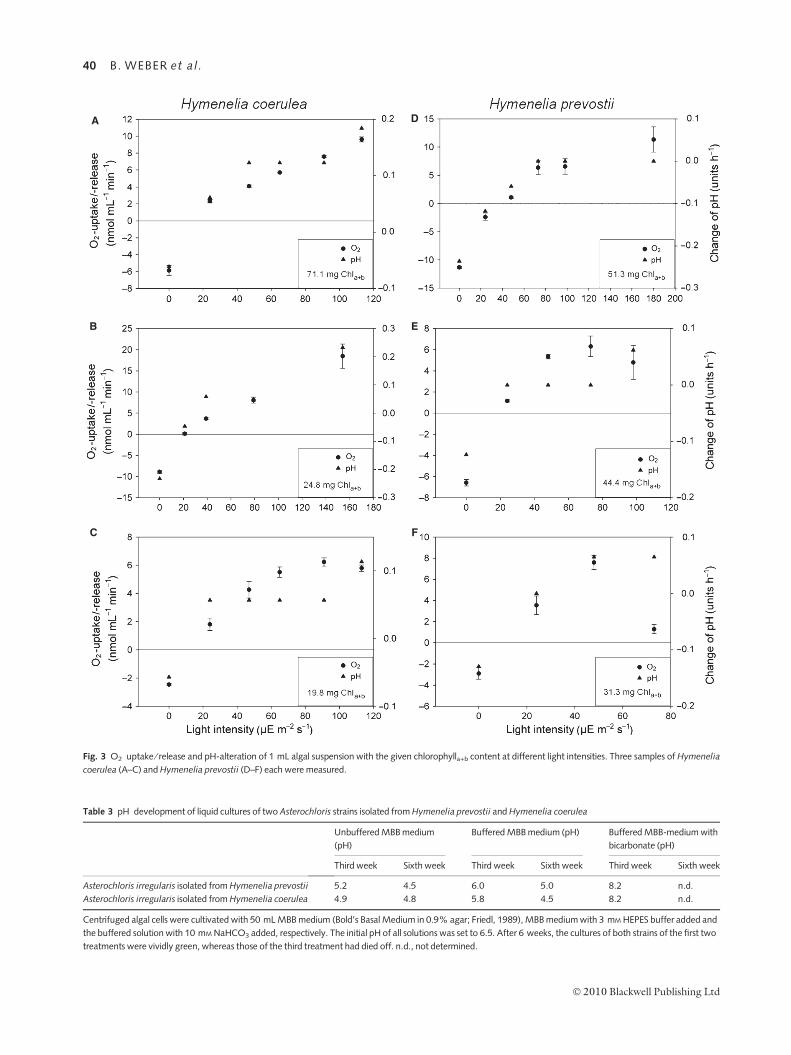
Table 3 pH development of liquid cultures of two Asterochloris strains isolated from Hymenelia prevostii and Hymenelia coerulea
Unbuffered MBB medium
(pH)
Buffered MBB medium (pH) Buffered MBB-medium with
bicarbonate (pH)
Third week Sixth week Third week Sixth week Third week Sixth week
Asterochloris irregularis isolated from Hymenelia prevostii 5.2 4.5 6.0 5.0 8.2 n.d.
Asterochloris irregularis isolated from Hymenelia coerulea 4.9 4.8 5.8 4.5 8.2 n.d.
Centrifuged algal cells were cultivated with 50 mL MBB medium (Bold’s Basal Medium in 0.9% agar; Friedl, 1989), MBB medium with 3 mM HEPES buffer added and
the buffered solution with 10 mM NaHCO3 added, respectively. The initial pH of all solutions was set to 6.5. After 6 weeks, the cultures of both strains of the first two
treatments were vividly green, whereas those of the third treatment had died off. n.d., not determined.
A
C
B
F
E
D
Fig. 3 O2 uptake ⁄ release and pH-alteration of 1 mL algal suspension with the given chlorophylla+b content at different light intensities. Three samples of Hymenelia
coerulea (A–C) and Hymenelia prevostii (D–F) each were measured.
40 B. WEBER et al.
� 2010 Blackwell Publishing Ltd

logical differences between the two species are most probably
not caused by the photobiont but by differing mycobionts.
As both lichens grow within dense limestone rock, they
must have a weathering mechanism allowing them to pene-
trate the substrate. Whereas the cavities of H. coerulea are
more or less roundish, those of H. prevostii are of an elon-
gated shape and their density is more than one-third higher.
This points to a higher weathering activity of H. prevostii,
which is consistent with the elemental analysis. Whereas in the
growth zone of H. coerulea, 5–6% of the calcium and manga-
nese, which are present within the unmodified substrate, are
removed, in H. prevostii even 11–12% of the original element
concentrations are missing. The element concentrations of
strontium and manganese are also depleted within the modi-
fied zone of both species, but to a somewhat smaller extent,
indicating a passive elution from the rock rather than an active
weathering mechanism. In contrast, the iron concentration in
the growth zone of H. prevostii does not reveal any changes,
whereas in H. coerulea, even an enrichment is observed, per-
haps caused by iron oxides being trapped within the lichen
thallus (Aghamiri & Schwartzman, 2002) or by the formation
of iron-containing pigments (Takeshi et al., 2003).
As possible weathering mechanisms, the activity of lichen
acids and oxalic acid is widely acknowledged (e.g. Schatz,
1962, 1963; Ascaso et al., 1982). In our case, however,
neither lichen acids nor oxalic acid could be identified as
weathering agents. In contrast, it was observed that the algal
strains of both Hymenelia species acidified the medium quite
effectively in the dark. Within a single hour, 1 mL of algal
solution was found to lower the pH by 0.12 (mean value of
H. coerulea samples) and 0.16 (mean value H. prevostii),
respectively, revealing that within a single night they may be
able to lower the pH by 1 unit. At somewhat higher light
intensities, however, the samples were also observed to alka-
lize the medium. Due to their endolithic growth, only about
0.1% of the incident light reaches the samples when the rock
is dry (Friedmann & Ocampo-Friedmann, 1984), whereas
in a wet stage, about 1% of the light arrives at the organ-
isms. As the exposed nearly vertical rock surfaces dry quite
rapidly after a rain shower, it can be expected that the sky is
still overcast when the rock is wet, causing maximum light
intensities of about 80 lE m)2 s)1 to reach the organisms
(at 800 lE m)2 s)1 ambient light). At these light intensities,
the alkalization rate of H. prevostii was by far lower than the
acidification rate in the dark. In contrast, H. coerulea
revealed alkalization at low light intensities, which may
correspond to its reduced weathering rate observed in the
morphological investigations. During these measurements,
however, only the algal partner has been measured, whereas
the fungal partner, which respires independently from light
conditions whenever enough water is available, has not been
taken into account. In the long-term observation of pH
development, both species did not reveal different alkaliza-
tion rates. Based on these studies, it can be concluded that
the carbonate dissolution is caused by respiratory CO2 as
described by Brunsden (1979) and Golubic & Schneider
(1979). An increase in CO2 content within the rock atmo-
sphere causes an increase in carbonic acid content within the
liquid phase and lowers the pH, which in turn enhances dis-
solution processes (Brunsden, 1979).
As described above, the biomass values of H. prevostii (mea-
sured as chlorophylla+b per surface area) are more than twice as
high compared with those of H. coerulea. Since in gas
exchange measurements, the respiration rates as related to the
chlorophylla+b content were found to be similar for both
species (Weber et al., 2007), the respiration and herewith
acidification rate of H. prevostii more than doubles that of
H. coerulea if related to the surface area. This species-specific
growth difference explains the much stronger weathering
effects of H. prevostii. Other fundamental differences between
the weathering mechanisms of both species were not apparent.
We observed that the mechanism of carbonate dissolution
caused by respiratory CO2 is crudely underestimated in the
literature. In their review, Syers & Iskandar (1973) estimated
the importance of biogenic CO2 in chemical weathering to be
of much less significance than lichen compounds. Wilson &
Jones (1983) did not discuss this mechanism, and later, Jones
& Wilson (1985) regarded this weathering process unlikely to
occur beneath lichen thalli. Conversely, Jones (1988) men-
tioned that endolithic lichens on limestone produce the deep-
est rock alteration not reflecting the underlying mechanism.
Ascaso et al. (1982) observed the dissolution of limestone
crystals beneath lichens without the formation of oxalates.
They did not even consider biogenic CO2 as a weathering
agent, but concluded that oxalic acid was responsible for the
solution processes not forming oxalates in the absence of
water molecules. This minor perception may partly be caused
by the fact that no new minerals are formed during the
CO2-induced weathering of limestone. We, in turn, hypothe-
size that carbonate dissolution by respiratory CO2 is one of
the main, if not the most important, biogenic weathering
mechanisms on carbonate substrates.
During long-term pH development, the presence of bicar-
bonate caused the algal strains not to lower the pH but to
alkalize the medium. This appears contradictory at first sight.
However, by utilization of mass spectroscopy it could be
shown that both species possess a carbon concentrating mech-
anism (CCM) that allows them to utilize small amounts of
bicarbonate (Scherr, 2000), releasing OH) ions back into the
substrate (D. Sultemeyer, pers. comm.). Nevertheless, the
organisms bleached under these conditions and after 6 weeks
completely died off. This may explain the fact that in their nat-
ural habitat, the two Hymenelia species are strictly limited to
steeply inclined limestone walls where CO2 is mostly available
as gas and only rarely has to be taken up as bicarbonate
dissolved in standing water.
Besides the effects of lichen respiration on the surrounding
substrate, the algal partners also take up CO2 for photosyn-
Respiration-induced weathering of lichens 41
� 2010 Blackwell Publishing Ltd

thesis. Due to the equilibrium, the concentration of carbonic
acid then decreases which may cause calcium carbonate to pre-
cipitate as fine-grained micrite. During macroscopic investiga-
tions it was observed that the substrate above the organisms
looked different from the unweathered rock being more fine-
grained and whitish. Elemental analyses revealed no major dif-
ferences compared to the underlying carbonate substrate.
Therefore, we think that this layer on top of the algae is
formed by biogenic precipitation processes. The observation
that endolithic lichens are responsible for both dissolution
and precipitation processes was also made by Bungartz et al.
(2004) who observed the endolithic lichen Verrucaria rubro-
cincta to be responsible for both biomineralization and
bioerosion processes of caliche plates in the southwestern
United States. Naylor et al. (2002) even identified the three
main processes bioerosion, bioconstruction and bioprotection
to be involved in biogeomorphic systems. As weathered
limestone surfaces were found to be less susceptible toward
weathering by carbonic acid (Carter & Viles, 2005; Thorn-
bush & Viles, 2007), bioprotection could also be involved in
our study, but was not investigated.
CONCLUSION
The two endolithically growing lichen species H. coerulea and
H. prevostii revealed the same general weathering patterns,
but at differing magnitude. Respiration-induced acidification
was proved to be the relevant weathering mechanism, whereas
lichen acids and oxalic acid were not involved in this process.
ACKNOWLEDGEMENTS
Prof. emeritus Dr. J. Schneider and Dr. W. Pohl are thanked
for the good cooperation, the support in the field and the
financial support of the field trips to Salzburg. Prof. J. Reitner
and Dr. G. Arp are acknowledged for the preparation of geo-
logical thin sections and for assistance in the evaluation of
those. Dr. U. Schussler is thanked for the elemental analysis.
We thank Dr. C. Printzen for DC and Hans Reichenberger
for chlorophyll analyses. The oxygen electrode was kindly
loaned by the Department of Plant Physiology (Prof. Neu-
haus) at the University of Kaiserslautern.
We herewith declare that the experiments comply with the
current laws of Germany.
REFERENCES
Aghamiri R, Schwartzman DW (2002) Weathering rates of bedrock
by lichens: a mini watershed study. Chemical Geology, 188, 249–
259.
Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ (1990) Basiclocal alignment search tool. Journal of Molecular Biology, 215,
403–410.
Ascaso C, Galvan J, Rodriguez-Pascual C (1982) The weathering of
calcareous rocks by lichens. Pedobiologia, 24, 219–229.
Bachmann E (1890) Die Beziehungen der Kalkflechten zu ihrem Sub-
strat. Berichte der Deutschen Botanischen Gesellschaft, 8, 141–145.
Berthelin J (1988) Microbial weathering processes in natural environ-ments. In Physical and Chemical Weathering in Geochemical Cycles(eds Lerman A, Meybeck M). Kluwer Academic Publishers,
Dordrecht, pp. 33–59.Blazquez F, Calvert F, Vendrell M (1995) Lichenic alteration and
mineralization in calcareous monuments of northeastern Spain.
Geomicrobiology, 13, 223–247.
Brunsden D (1979) Weathering. In Process in Geomorphology (edsEmbleton C, Thornes J). Edward Arnold Press, London,
pp. 73–129.
Budel B, Weber B, Kuhl M, Pfanz H, Sultemeyer D, Wessels D
(2004) Reshaping of sandstone surfaces by cryptoendolithic cyano-bacteria: bioalkalisation causes chemical weathering in arid land-
scapes. Geobiology, 2, 261–268.
Bungartz F, Garvie LAJ, Nash TH III (2004) Anatomy of the endo-
lithic Sonoran Desert lichen Verrucaria rubrocincta Breuss: impli-cations for biodeterioration and biomineralization. Lichenologist,36, 55–73.
Carter NEA, Viles HA (2005) Bioprotection explored: the story of alittle known earth surface process. Geomorphology, 67, 273–281.
Chen J, Blume H-P, Beyer L (2000) Weathering of rocks induced by
lichen colonization – a review. Catena, 39, 121–146.
Culberson CF (1972) Improved conditions and new data for the iden-tification of lichen products by a standardized thin-layer chromato-
graphic method. Journal of Chromatography, 72, 113–125.
Culberson CF, Johnson A (1982) Substitution of methyl tert.-butyl
ether for diethyl ether in the standardized thin-layer chromato-graphic method for lichen products. Journal of Chromatography,238, 483–487.
Culberson CF, Kristinsson H-D (1970) A standardized method forthe identification of lichen products. Journal of Chromatography,46, 85–93.
Danin A, Caneva G (1990) Deterioration of limestone walls in Jerusa-
lem and marble monuments in Rome caused by cyanobacteria andcyanophilous lichens. International Biodeterioration, 26, 397–
417.
Danin A, Garty J (1983) Distribution of cyanobacteria and lichens on
hillsides of the Negev Highlands and their impact on biogenicweathering. Zeitschrift fur Geomorphologie N F, 27, 423–444.
Danin A, Gerson R, Garty J (1983) Weathering patterns on hard lime-
stone and dolomite by endolithic lichens and cyanobacteria: sup-porting evidence for eolian contribution to Terra Rossa soil. SoilScience, 136, 213–217.
Doppelbaur HW (1959) Studien zur Anatomie und Ent-
wicklungsgeschichte einiger endolithischen pyrenocarpen Flechten.Planta, 53, 246–292.
Friedl T (1989) Comparative ultrastructure of pyrenoids in Trebouxia(Microthamniales, Chlorophyta). Plant Systematics and Evolution,
164, 145–159.Friedl T (1996) Evolution of the polyphyletic genus Pleurastrum
(Chlorophyta): inferences from nuclear-encoded ribosomal
DNA sequences and motile cell ultrastructure. Phycologia, 35,
456–469.Friedmann EI, Ocampo-Friedmann R (1984) Endolithic microorgan-
isms in extreme dry environments: analysis of a lithobiontic micro-
bial habitat. In Current Perspectives in Microbial Ecology (eds KlugMJ, Reddy CA). ATM Press, Washington, DC, pp. 177–185.
Funfstuck M (1895) Die Fettabscheidungen der Kalkflechten.
Beitrage zur wissenschaftlichen Botanik, 1, 157–220.
Gehrmann CK, Krumbein WE, Petersen K (1992) Endolithic lichensand the corrosion of carbonate rocks – a study of biopitting.
International Journal of Mycology and Lichenology 5, 37–48.
42 B. WEBER et al.
� 2010 Blackwell Publishing Ltd

Golubic S, Schneider J (1979) Carbonate dissolution. In Biogeochemi-cal Cycling of Mineral-Forming Elements (eds Trudinger PA, Swa-
ine DJ). Elsevier Scientific Publishing Company, Amsterdam,Oxford, New York, pp. 107–129.
Golubic S, Friedmann I, Schneider J (1981) The lithobiontic ecologi-
cal niche, with special reference to microorganisms. Journal of Sedi-mentary Petrology, 51, 475–478.
Hall TA (1999) BioEdit: a user-friendly biological sequence align-
ment editor and analysis program for Windows 95 ⁄ 98 ⁄ NT. NucleicAcids Symposium Series, 41, 95–98.
Hepperle D (2004) SeqAssem�. A sequence analysis tool, contig
assembler and trace data visualization tool for molecular sequences.
Win32-Version. Distributed by the author at http://www.sequen-
tix.de/.Hoppert M, Flies C, Pohl W, Gunzl B, Schneider J (2004) Coloniza-
tion strategies of lithobiotic microorganisms on carbonate rocks.
Environmental Geology, 46, 421–428.
Jones D (1988) Lichens and pedogenesis. In CRC Handbook ofLichenology (ed. Galun M). CRC Press Inc., Boca Raton, Florida,
pp. 109–124.
Jones D, Wilson MJ (1985) Chemical activity of lichens on min-eral surfaces – a review. International Biodeterioration, 21, 99–
104.
Kumar S, Tamura K, Nei M (2004) MEGA3: integrated software for
Molecular Evolutionary Genetics Analysis and sequence alignment.Briefings in Bioinformatics, 5, 150–163.
Kuster FW, Thiel A (1985) Rechentafeln fur die chemische Analytik.
103. Ed. De Gruyter, Berlin.
Mikhailyuk TI, Sluiman HJ, Massalski A, Mudimu O, DemchenkoEM, Kondratyuk SY, Friedl T (2008) New streptophyte green algae
from terrestrial habitats and an assessment of the genus Interfilum(Klebsormidiophyceae, Streptophyta). Journal of Phycology, 44,1586–1603.
Mottershead D, Lucas G (2000) The role of lichens in inhibiting ero-
sion of a soluble rock. Lichenologist, 32, 601–609.
Muller J, Friedl T, Hepperle D, Lorenz M, Day JG (2005) Distinctionbetween mutliple isolates of Chlorella vulgaris (Chlorophyta,
Trebouxiophyceae) and testing for conspecificity using amplified
fragment length polymorphism and ITS rDNA sequences. Journalof Phycology, 41, 1236–1247.
Naylor LA, Viles HA, Carter NEA (2002) Biogeomorphology revis-
ited: looking towards the future. Geomorphology, 47, 3–14.
Pia J (1937) Die kalklosenden Thallophyten. Archiv fur Hydrobiolo-gie, 31, 264–328; 341–398.
Pohl W, Schneider J (2002) Impact of endolithic biofilms on
carbonate rock surfaces. In Natural Stone, Weathering Phenom-ena, Conservation Strategies and Case Studies (eds Siegesmund S,Weiss T, Vollbrecht A). Geological Society, London, pp. 177–
194.
Pohl W, Schneider J (2005) Geochemische Einflusse endolithischer
Mikroorganismen auf Gesteinsoberflachen. Zeitschrift der deutschenGesellschaft fur Geowissenschaften, 156, 81–92.
Ronen R, Galun M (1984) Pigment extraction from lichens
with dimethyl sulfoxide (DMSO) and estimation of chlorophyll
degradation. Environmental and Experimental Botany, 24,239–245.
Schatz A (1962) Pedogenic (soil forming) activity of lichen acids.
Naturwissenschaften, 49, 518–519.Schatz A (1963) Soil microorganisms and soil chelation. The pedo-
genic action of lichens and lichen acids. Journal of Agriculturaland Food Chemistry, 11, 112–118.
Scherr C (2000) Okophysiologische Untersuchungen alpinerGesteinsflechten und ihrer Photobionten. Diploma Thesis, Univer-
sity of Kaiserslautern, Kaiserslautern.
Schneider J (1976) Biological and inorganic factors in the destruction
of limestone coasts. Contributions to Sedimentology, 6, 1–112.Skaloud P, Peksa O (2010) Evolutionary inferences based on ITS
rDNA and actin sequences reveal extensive diversity of the common
lichen alga Asterochloris (Trebouxiophyceae, Chlorophyta). Molec-ular Phylogenetics and Evolution, 54, 36–46.
Sollas WJ (1880) On the activity of a lichen on limestone. Report ofthe British Association for the Advancement of Science, London, UK,
586.Syers JK, Iskandar IK (1973) Pedogenic significance of lichens. In The
Lichens (eds Ahmadjian V, Hale ME). Academic Press, New York,
London, pp. 225–248.
Takeshi K, Takeshi M, Toshihiko O (2003) Accumulation mecha-nisms of uranium, copper and iron by lichen Trapelia involuta. In
Biomineralization (BIOM2001): Formation, Evolution andApplication, Proceedings of the 8th International Symposium onBiomineralization (eds Kobayashi I, Ozawa H). Tokai UniversityPress, Kanagawa, pp. 298–301.
Thornbush MJ, Viles HA (2007) Simulation of the dissolution of
weathered versus unweathered limestone in carbonic acid solutionsof varying strength. Earth Surface Processes and Landforms, 32,
841–852.
Tretiach M (1995) Ecophysiology of calcicolous endolithic lichens:
progress and problems. Giornale Botanico Italiano, 129,159–184.
Viles H (1987) A quantitative scanning electron microscope study
of evidence for lichen weathering of limestone, Mendip Hills,
Sommerset. Earth Surface Processes and Landforms, 12,467–473.
Weber B, Scherr C, Reichenberger H, Budel B (2007) Fast reactiva-
tion by high air humidity and photosynthetic performance of alpinelichens growing endolithically in limestone. Arctic, Antarctic andAlpine Research, 39, 309–317.
Wilson MJ, Jones D (1983) Lichen weathering of minerals: implica-
tions for pedogenesis. In Residual Deposits: Surface-RelatedWeathering Processes and Materials (ed. Wilson RCL).
Blackwell Scientific, London, Oxford, pp. 5–12.
Zukal H (1884) Flechtenstudien. Denkschriften der mathematisch-naturwissenschaftlichen Klasse der kaiserlichen Akademie derWissenschaften zu Wien, 48, 1–44.
Respiration-induced weathering of lichens 43
� 2010 Blackwell Publishing Ltd