dexamethasone downregulates calcification-inhibitor molecules and accelerates osteogenic...
TRANSCRIPT

Dexamethasone Downregulates Calcification-InhibitorMolecules and Accelerates Osteogenic Differentiation
of Vascular PericytesImplications for Vascular Calcification
John Paul Kirton, Fiona L. Wilkinson, Ann E. Canfield, M. Yvonne Alexander
Abstract—Vascular calcification is present in many pathological conditions and is recognized as a strong predictor offuture cardiovascular events. Current evidence suggests that it is a regulated process involving inducing and inhibitorymolecules. Glucocorticoids have great clinical importance as antiinflammatory drugs and can act as potent inducers ofosteogenic differentiation in vitro. The effect of glucocorticoids on vascular cells in vivo remains obscure. Pericytes arepluripotent cells that can differentiate into osteoblasts, and recent evidence suggests that they could participate invascular calcification. We hypothesized that the synthetic glucocorticoid dexamethasone would enhance the rate ofpericyte differentiation and mineralization in vitro with a concomitant suppression of calcification-inhibitory molecules.Three weeks of dexamethasone treatment induced a 2-fold increase in (1) alkaline phosphatase activity, (2) calciumdeposition, and (3) the number of nodules formed in vitro; and a reduction in the expression of matrix Gla protein(MGP), osteopontin (OPN), and vascular calcification-associated factor (VCAF) mRNAs. The glucocorticoid receptorantagonist Org 34116 abolished dexamethasone-accelerated pericyte differentiation, nodule formation, and mineraliza-tion. Data obtained using Org 34116, the transcription inhibitor actinomycin D, and the protein synthesis inhibitorcyclohexamide suggest that MGP, OPN, and VCAF mRNA abundance are controlled at different and multiple levelsby dexamethasone. This is the first report showing that dexamethasone enhances the osteogenic differentiation ofpericytes and downregulates genes associated with inhibition of mineralization. Our study highlights the need for furtherinvestigation into the long-term consequences of prolonged glucocorticoid therapy on vascular calcification. (Circ Res.2006;98:1264-1272.)
Key Words: pericytes � differentiation � calcification � gene regulation � glucocorticoid
Calcification can lead to devastating clinical consequenceswhen present in blood vessels. It has been suggested that
cells maintain a balance between calcification inducing andinhibitory proteins in soft tissues in such a way that thedeposition of a mineralized matrix is normally avoided.Alterations in this balance may be brought about by injury ordisease, with the subsequent deposition of calcific deposits inthe vessel wall. It is now well established that vascularcalcification is associated with elevated expression of bone-related proteins1,2 and that pericyte-like cells exist in thevasculature, which, under appropriate conditions, can beinduced to display characteristics of the osteoblastic lin-eage.3,4 Our previous studies have demonstrated that culturedpostconfluent pericytes form large calcified multicellularnodules both in vitro and in vivo,5 the matrix of whichresembles that found in calcified atherosclerotic plaques andbone.5 Although factors such as transforming growth factor �(TGF-�),6 tumor necrosis factor � (TNF-�),7 and acetylated
LDL8 have been shown to be involved in the promotion ofcalcification in vitro, the underlying mechanisms of vascularcalcification have not been fully elucidated.
The main risk factors for atherosclerosis include not onlythe classic factors such as age, high cholesterol levels, andhypertension but also prolonged glucocorticoid therapy. Themechanisms underlying the adverse effects of glucocorticoidexcess on the vascular system are complex and have not yetbeen fully explained. It is well recognized that vascularossification and osteoporosis often coexist in patients,whereby bone mineral is formed within the vasculature, whileit is simultaneously lost from the skeleton.9 How this buildupand loss of mineral can occur simultaneously in patientsremains to be elucidated.
Glucocorticoids are a class of steroid hormones producedby the cortex within the mammalian adrenal gland and arenormally maintained at steady-state levels. A deregulation ofendogenous levels is induced by vascular injury in vivo10 and
Original received November 17, 2005; revision received March 31, 2006; accepted April 6, 2006.From the Wellcome Trust Centre for Cell-Matrix Research and Division of Cardiovascular and Endocrine Sciences, Faculty of Medical and Human
Sciences, University of Manchester, UK.Correspondence to Dr M. Yvonne Alexander, Division of Cardiovascular and Endocrine Sciences, University of Manchester, Core Technology Facility
(3rd Floor), 46 Grafton St, Manchester M13 9NT, United Kingdom. E-mail [email protected]© 2006 American Heart Association, Inc.
Circulation Research is available at http://circres.ahajournals.org DOI: 10.1161/01.RES.0000223056.68892.8b
1264 by guest on November 13, 2015http://circres.ahajournals.org/Downloaded from by guest on November 13, 2015http://circres.ahajournals.org/Downloaded from by guest on November 13, 2015http://circres.ahajournals.org/Downloaded from

could be a contributing factor in the development of vascularcalcification. The synthetic glucocorticoid dexamethasoneinduces an osteoblastic differentiation pathway in manydifferent cell types in vitro, including human bone marrow–derived stromal cells,11,12 fetal rat calvarial cells,13 bovinevascular smooth muscle cells (SMCs),14 and mouse embryo–derived NIH3T3 fibroblasts.15 This study tests the hypothesisthat dexamethasone enhances the in vitro osteoblastic differ-entiation of vascular pericytes and that this may be associatedwith a decrease in the expression of calcification-inhibitormolecules. We investigate several parameters within peri-cytes exposed to dexamethasone, in terms of their prolifera-tion, the time-course of nodule formation and mineralization,the expression of osteoblastic markers, and the mechanismsunderlying these effects.
Materials and Methods
Cell Culture and Nodule FormationPericytes were isolated from bovine retinas essentially as previouslydescribed,16 except that cells were isolated and subsequently main-tained either in the continued presence or absence of dexamethasone(10 nmol/L) in Eagle’s minimal essential medium (MEM) containing20% FCS, 2 mmol/L glutamine, 1 mmol/L sodium pyruvate, 50�g/mL ascorbate-2-phosphate, and nonessential amino acids. Con-trol cells were treated with vehicle alone. For experiments, pericyteswere plated at 1.5�104 cells/cm2, with a media change every 4 days.In some experiments, the glucocorticoid receptor (GR) antagonistOrg 34116 (1 �mol/L; a kind gift from Angela Fisher, Organon,Motherwell, Scotland, UK) was also included, and the cells weresubsequently harvested at weekly time points. The area occupied bynodules was determined using AnalySIS software. Mineralizednodules were visualized by washing in PBS, fixing in 4% formal-dehyde, and staining with 1% (wt/vol) Alizarin red S (pH 4.2).Actinomycin D and cyclohexamide were used at a final concentra-tion of 5 �g/mL and 35 mmol/L, respectively. Cells were grown for3 weeks to ensure detectable baseline levels of vascular calcification-associated factor (VCAF), osteopontin (OPN), and matrix Glaprotein (MGP) mRNA and protein; then actinomycin D or cyclo-hexamide was added to the media for 8 and 4 hours, respectively,before harvesting.
Alkaline Phosphatase ActivityCells were seeded in 6-well plates and maintained up to 4 weeks.Proteins were extracted at weekly time points by freeze-thawing thecells in 0.05% Triton X-100 in PBS. Total cellular proteins weremeasured using Bradford’s assay (Bio-Rad). Alkaline phosphatase(ALP) activity was quantified in protein lysates (15 �g) made up to100 �L in 2-amino-2-methyl 1-propanol (AMP) buffer containing 8mmol/L �-nitrophenylphosphate (�-NPP) (Sigma-Aldrich) as sub-strate and incubated at 37°C for 2 hours before terminating thereaction with 0.5 mol/L NaOH (100 �L). The absorbance wasmeasured at 405 nm and ALP activity calculated as nmol/L�-nitrophenol converted per microgram of protein per minute.
Calcium Assay45Ca deposition was assayed as described.17 Cells were seeded in24-well plates and harvested weekly up to 4 weeks. 45CaCl2 (ICN)was added (1.0 �Ci/mL for 48 hours), and the cells were harvestedin PBS (0.3 mL), placed into vials containing perchloric acid (0.2mL) with 3% hydrogen peroxide (0.3 mL), and incubated at 80°C for1 hour. Samples were then dissolved in ethylene glycol monoethylether (0.6 mL), and the radioactivity was quantified by scintillationcounting.
RNA AnalysisCells were seeded in 25-cm2 flasks and the RNA harvested weeklyusing TRIzol reagent (Invitrogen). Northern blot analysis and probepreparation were performed as previously described.5,18 Densitome-try was used to express the mRNA abundance of MGP, OPN, andVCAF relative to 18S ribosomal RNA. To determine mRNAstability, total RNA was harvested at selected time points followingincubation with the transcriptional inhibitor actinomycin D. mRNAabundance at the time of addition of actinomycin D was set at 100%,and the rate of OPN, MGP, and VCAF mRNA decay was comparedin dexamethasone- versus vehicle-treated cells.
Western Blot and Immunocytochemical AnalysisWhole cell lysates were extracted as previously described,16 andcytosolic and nuclear fractions were separated using a BioVision kit(Mountain View, Calif) according to the protocol of the manufac-turer. Proteins (20 �g/lane) were separated by SDS-PAGE (10%gels) and transferred to nitrocellulose membranes (Amersham Bio-sciences). The membranes were incubated with a nuclear factor �B(NF-�B) antibody (1:500 dilution; Santa Cruz Biotechnology, SantaCruz, Calif) at 4°C for 16 hours and a horseradish peroxidase–conjugated swine anti-rabbit secondary antibody (1:1000 dilution)and developed as described.16 For immunocytochemistry, pericyteswere cultured in chamber slides for 48 hours, fixed in 100%methanol, and blocked in 5% goat serum/PBS for 1 hour at roomtemperature. The NF-�B polyclonal antibody (1:200 dilution) wasused with a goat anti-rabbit IgG, conjugated to AlexaFluor 488secondary antibody (Molecular Probes; 1:300 dilution). Cells werecounterstained with 4 �g/mL 4�,6-diamidino-2-phenylindole(DAPI).
Data AnalysisThe data for the ALP, nodule formation, and 45Ca assay are shown asthe mean�SD of 2 experiments performed in triplicate wells. Anunpaired, 2-tailed Student’s t test or ANOVA was used to determinestatistical significance between mean values, as appropriate. A valueof P�0.05 was considered statistically significant. For Northern andWestern blots, data are representative of at least 2 experiments.
ResultsDexamethasone Induces Differentiation andMineralization of Pericytes In VitroInitially, the effects of continued presence or absence ofdexamethasone on pericyte proliferation, alkaline phospha-tase activity, calcium incorporation, and mineralized noduleformation were investigated. Using a thymidine incorporationassay, pericyte growth was investigated during the first weekof culture, and dexamethasone was found to have no signif-icant effect on pericyte proliferation compared with controls(data not shown).
There was a 4.4-fold increase in ALP activity in pericytesafter dexamethasone treatment during 3 weeks of culturecompared with 2.1-fold increase in the vehicle-treated cells(Figure 1). At week 3, there was a 2.2-fold increase in ALPactivity in cells grown in the presence of dexamethasonecompared with vehicle-treated cells (Figure 1). No furtherincrease in ALP activity was detected at week 4.
Nodule formation was detected as early as 1 week afterplating in the presence of dexamethasone (Figure 2A),culminating in a 2.9-fold increase in the number of nodulesformed, compared with vehicle-treated cells at week 4 (Fig-ure 2B). In addition, nodules were larger (Table) and miner-alized earlier in the presence of dexamethasone comparedwith controls (Figure 2A). Furthermore, in the presence of the
Kirton et al Dexamethasone Accelerates Pericyte Differentiation 1265
by guest on November 13, 2015http://circres.ahajournals.org/Downloaded from

glucocorticoid antagonist Org 34116, the dexamethasone-in-duced increase in pericyte nodule number and size wasabolished (Table).
Culturing pericytes for 4 weeks in dexamethasone in-creased 45Ca deposition 2.4-fold compared with vehicle-
treated cells (Figure 3). The increased calcium incorporationin the dexamethasone-treated cells was also abolished in thepresence of Org 34116 (Figure 3).
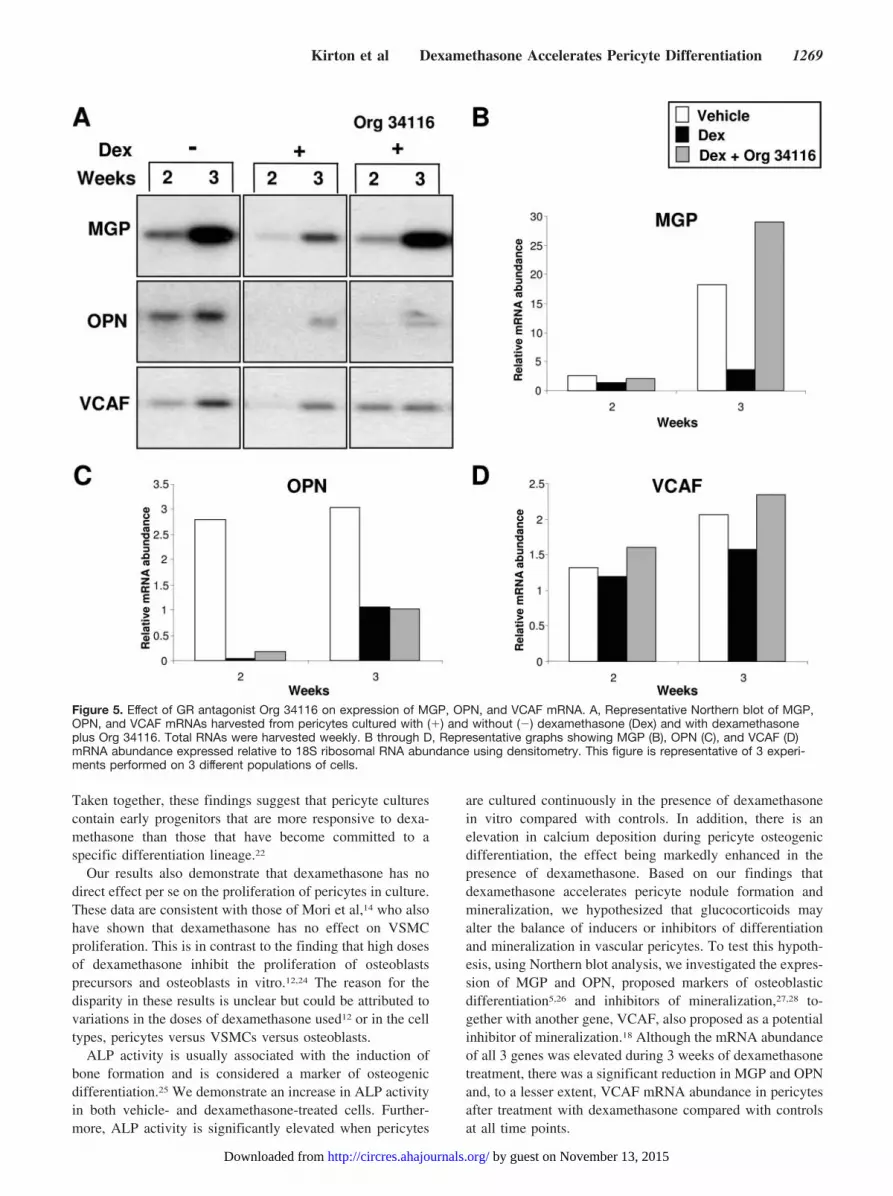
Dexamethasone Reduces OPN, MGP, and VCAFmRNA Abundance in PericytesMGP, OPN, and VCAF mRNAs were examined in cellsmaintained in the presence and absence of dexamethasone.As reported previously, we detected an increase in expressionof OPN,5 MGP, and VCAF mRNA18 in vehicle-treatedcontrol cells with increasing time in culture (Figure 4). All 3genes were also upregulated in pericytes cultured in thepresence of dexamethasone for 2 to 3 weeks (Figure 4);however, dexamethasone caused a decrease in the mRNAabundance of all 3 transcripts compared with vehicle-treatedcells (Figure 4), with the decrease in MGP and OPN mRNAbeing particularly striking (Figure 4B and 4C). These differ-ences in gene expression were maintained when pericyteswere cultured with and without dexamethasone for 4 weeks(results not shown).
Control of MGP, OPN, and VCAF GeneExpression by DexamethasoneThe decline in abundance of MGP, OPN, and VCAF mRNAscould be caused directly by either a suppression of transcrip-
Figure 1. Effect of dexamethasone (Dex) on alkaline phospha-tase activity, measured using the �-NPP assay and expressedas picomole of �-nitrophenol formed per microgram of proteinper minute. Results confirm significant differences betweendexamethasone-treated and untreated cells. *P�0.05,**P�0.001.
Figure 2. Effect of dexamethasone (Dex) on the formation of mineralized nodules by pericytes. A, Pericytes were seeded with growthmedium in the absence (a through d) or presence (e through h) of dexamethasone (10 nmol/L) and cultured for 1 to 4 weeks. Alizarinred staining was performed to confirm the deposition of a mineralized matrix. The figure is representative of 3 experiments. Bar repre-sents 100 �m. B, Time course of nodule formation in the presence of vehicle (open bars) or dexamethasone (closed bars). Nodulenumbers were counted at weekly time points and analyzed using AnalySIS software. Values are presented as mean�SD of 2 experi-ments performed in triplicate wells in 6-well plates. Statistical significance between dexamethasone-treated and control cells orbetween 1 and 4 weeks is represented as *P�0.001.
1266 Circulation Research May 26, 2006
by guest on November 13, 2015http://circres.ahajournals.org/Downloaded from

tion or accelerated degradation of RNA or indirectly by theinvolvement of a second transcription factor. mRNA stabilitywas investigated using the transcription inhibitor actinomycinD. MGP, OPN, and VCAF mRNAs were found to have longhalf-lives of more than 8 hours, which were not altered byculturing cells in the presence of dexamethasone (data notshown), suggesting that dexamethasone had not induceddegradation of these mRNAs.
To investigate transcriptional control mediated by dexa-methasone binding to its receptor, the cells were treated withOrg 34116. Pericytes were divided into 3 groups: (1) withoutdexamethasone (�), (2) with dexamethasone (�), and (3) inthe presence of dexamethasone plus Org 34116. The presenceof Org 34116 blocked the dexamethasone-induced downregu-lation of MGP and VCAF mRNAs (Figure 5A, 5B, and 5D),adding support for a transcriptional mechanism of regulationof these genes by dexamethasone. However, the GR antago-nist had no effect on OPN expression after dexamethasonetreatment (Figure 5A and 5C), indicating that dexamethasoneregulates OPN gene expression by a different mechanism tothat shown for MGP and VCAF.
Because OPN expression was unaffected by the GR antag-onist, it is likely that another protein is involved in theregulation of OPN by dexamethasone. To investigate thispossibility and to assess the role of de novo protein synthesisin the downregulation of MGP, VCAF, and OPN mRNAduring continuous exposure to dexamethasone, pericyteswere treated with the protein synthesis inhibitor cyclohexam-ide for 4 hours (Figure 6). As previously shown, dexameth-asone downregulated expression of MGP and OPN mRNAcompared with control cells. However, the mRNA abundanceof both MGP and OPN was found to be unaffected bycyclohexamide treatment (Figure 6). In contrast, cyclohex-amide abolished the modest downregulation of VCAF mRNAexpression after exposure of the cells to dexamethasone,suggesting involvement of another protein in VCAF regula-tion by dexamethasone (Figure 6).
Dexamethasone Prevents Nuclear Accumulation ofNF-�B in PericytesGrowing evidence suggests that the antiinflammatory effectsof glucocorticoids are mediated by a downregulation ofNF-�B. Therefore, to establish a candidate transcriptionfactor that may be involved in VCAF gene regulation, weinvestigated the effect of dexamethasone on the NF-�Bprotein in pericytes. Immunocytochemical analysis confirmedsignificant NF-�B staining in both the cytoplasmic compart-ment and to a lesser extent, in the nuclear compartment incontrol pericytes (Figure 7A, a and c). However, afterdexamethasone treatment, there was an apparent reduction innuclear staining of NF-�B (Figure 7A, b and d). Western blotanalysis of total protein lysates obtained from pericytescultured with and without dexamethasone revealed a decreasein total levels of NF-�B protein in the dexamethasone-treatedversus control cells (Figure 7B). NF-�B was restored tocontrol levels when cells were incubated in the presence ofdexamethasone plus Org 34116, indicating a transcriptionallevel of control of this gene via the GR (Figure 7B).Fractionation of the cells, followed by Western blot analysis,confirmed a change in the subcellular distribution of NF-�B,whereby a pronounced decrease in NF-�B in the nuclearcompartment of pericytes treated with dexamethasone com-pared with controls was demonstrated (Figure 7C).
DiscussionThis is the first report demonstrating that dexamethasoneenhances the osteogenic differentiation of bovine retinal
Figure 3. Dexamethasone (Dex)-induced calcium depositionduring pericyte differentiation. [45Ca] accumulation within thematrix of pericyte nodules in response to dexamethasone isrepresented as average cpm per well. Results are shown asmean�SD of 2 experiments, performed in triplicate wells, andconfirm significant differences between dexamethasone-treatedand vehicle-treated cells and between dexamethasone-treatedand Org 34116–treated cells. *P�0.05, #P�0.01.
Effects of Dexamethasone and Dexamethasone Plus Org 34116on Nodule Formation
Size of Nodule (�m2)
Weeks
1 2 3 4
Vehicle
0–4999 3 34 66.3 83.3
5000–9999 1.3 29 68 100.7
10 000–19 999 11.7 48.3
�20 000 0.3 4
Dexamethasone
0–4999 6.3 50 69.7 90
5000–9999 7 34.3 63.3 132
10 000–19 999 12.7 44.3 73
�20 000 2.7 20
Dexamethasone�Org34116
0–4999 4.3 36 71.3 100.7
5000–9999 24.3 62.3 48.3
10 000–19 999 1 15.3 46.7
�20 000 0.3 3
Nodule numbers were counted and size measured using AnalySIS imagingsoftware. Values presented as a mean�SD of 2 experiments performed intriplicate wells.
Kirton et al Dexamethasone Accelerates Pericyte Differentiation 1267
by guest on November 13, 2015http://circres.ahajournals.org/Downloaded from

pericytes in vitro and investigating the mechanism by whichthis glucocorticoid exerts its effects. We demonstrate thatdexamethasone treatment results in enhanced osteogenicdifferentiation of pericytes, an effect abolished in the pres-ence of the GR antagonist Org 34116. Dexamethasoneinduced an upregulation of the osteoblast-specific markerALP, increased nodule formation, and calcium deposition, aswell as downregulated the postulated calcification-inhibitorgenes MGP, OPN, and VCAF. Proliferation was unaffectedby dexamethasone treatment of pericytes.
Previous studies show that vascular pericytes are capableof differentiating into osteoblast-like cells in vitro and invivo.5 Bone formation has been shown to occur in thevasculature,19 and Bostrom et al propose a developmentalretention of pluripotent cells coupled with a loss of molecularregulatory control that unmasks an embryonic osteogenicprogram when given an appropriate stimulus.20 We speculatethat pericyte-like cells represent a potential source of osteo-progenitor cells in the adult and, under certain stress situa-tions, have their normal signaling pathways deregulated. Forexample, under the microenvironmental conditions associatedwith atherosclerosis, eg, inflammation and elevated oxidative
stress, these cells could adopt an osteogenic phenotype anddeposit a calcified matrix in the vasculature.
Many reports have been made on the paradox of osteopo-rosis and vascular calcification occurring together in manypatients,17,21 adding strength to the idea that vascular calcifi-cation is a cellular and genetic process, rather than nonspe-cific calcium precipitation. Although glucocorticoids areknown to induce osteoporosis, it appears that in vitro,glucocorticoids promote recruitment, differentiation, andmaturation of osteoblasts.12,13,22 In the present study, dexa-methasone was used at a physiological concentration of 10nmol/L, which routinely induces the differentiation of humanbone marrow stromal cells in vitro.11,12 We demonstrate asignificant increase in the number and size of nodules formedby pericytes, continuously cultured in the presence of dexa-methasone, compared with control cells. It is important tonote that cells isolated in the presence and absence ofdexamethasone appeared identical in terms of their expres-sion of phenotypic markers of pericytes.5,16 However, inaccordance with previous data,22,23 we cannot exclude thepossibility that dexamethasone may have increased the pro-portion of osteoprogenitor cells in the original cell isolates.
Figure 4. Effect of dexamethasone on VCAF, OPN, and MGP mRNA expression in pericytes. A, Cells were cultured in the presence (�)or absence (�) of dexamethasone (Dex), and total RNAs were harvested at weeks 2 and 3. A representative Northern blot is shownwith autoradiograph exposure times varying from 24 hours for VCAF, MGP, and OPN to 6 hours for 18S. B through D, Representativegraphs showing MGP (B), OPN (C), and VCAF (D) mRNA abundance expressed relative to 18S ribosomal RNA abundance using densi-tometry. This figure is representative of 4 experiments performed on 4 different populations of cells.
1268 Circulation Research May 26, 2006
by guest on November 13, 2015http://circres.ahajournals.org/Downloaded from
Taken together, these findings suggest that pericyte culturescontain early progenitors that are more responsive to dexa-methasone than those that have become committed to aspecific differentiation lineage.22
Our results also demonstrate that dexamethasone has nodirect effect per se on the proliferation of pericytes in culture.These data are consistent with those of Mori et al,14 who alsohave shown that dexamethasone has no effect on VSMCproliferation. This is in contrast to the finding that high dosesof dexamethasone inhibit the proliferation of osteoblastsprecursors and osteoblasts in vitro.12,24 The reason for thedisparity in these results is unclear but could be attributed tovariations in the doses of dexamethasone used12 or in the celltypes, pericytes versus VSMCs versus osteoblasts.
ALP activity is usually associated with the induction ofbone formation and is considered a marker of osteogenicdifferentiation.25 We demonstrate an increase in ALP activityin both vehicle- and dexamethasone-treated cells. Further-more, ALP activity is significantly elevated when pericytes
are cultured continuously in the presence of dexamethasonein vitro compared with controls. In addition, there is anelevation in calcium deposition during pericyte osteogenicdifferentiation, the effect being markedly enhanced in thepresence of dexamethasone. Based on our findings thatdexamethasone accelerates pericyte nodule formation andmineralization, we hypothesized that glucocorticoids mayalter the balance of inducers or inhibitors of differentiationand mineralization in vascular pericytes. To test this hypoth-esis, using Northern blot analysis, we investigated the expres-sion of MGP and OPN, proposed markers of osteoblasticdifferentiation5,26 and inhibitors of mineralization,27,28 to-gether with another gene, VCAF, also proposed as a potentialinhibitor of mineralization.18 Although the mRNA abundanceof all 3 genes was elevated during 3 weeks of dexamethasonetreatment, there was a significant reduction in MGP and OPNand, to a lesser extent, VCAF mRNA abundance in pericytesafter treatment with dexamethasone compared with controlsat all time points.
Figure 5. Effect of GR antagonist Org 34116 on expression of MGP, OPN, and VCAF mRNA. A, Representative Northern blot of MGP,OPN, and VCAF mRNAs harvested from pericytes cultured with (�) and without (�) dexamethasone (Dex) and with dexamethasoneplus Org 34116. Total RNAs were harvested weekly. B through D, Representative graphs showing MGP (B), OPN (C), and VCAF (D)mRNA abundance expressed relative to 18S ribosomal RNA abundance using densitometry. This figure is representative of 3 experi-ments performed on 3 different populations of cells.
Kirton et al Dexamethasone Accelerates Pericyte Differentiation 1269
by guest on November 13, 2015http://circres.ahajournals.org/Downloaded from

The dexamethasone-induced downregulation of OPNand VCAF in pericytes, together with the increased min-eral deposition by these cells, lends support to the hypoth-esis that these proteins may act as inhibitors of mineral-ization.18,27 The effects of the downregulation of MGPexpression by dexamethasone on the osteogenic differen-
tiation and mineralization of pericytes is more complexbecause MGP has been shown to both (1) regulate theosteogenic differentiation of vascular cells by controllingthe availability of bone morphogenetic protein 2 (BMP-2)26,29,30 and (2) inhibit mineralization in vivo.28 Therefore,in future studies, it would be interesting to determine
Figure 6. Effect of cyclohexamide (CHX) on MGP, OPN, and VCAF mRNA expression in pericytes. Cells were grown for 3 weeks in thepresence or absence of dexamethasone (Dex) and then treated with or without CHX for a 4-hour period before harvesting. Northernblots were probed with 32P-labeled OPN, MGP, VCAF, or 18S cDNA probes, visualized by autoradiography, and quantified by densi-tometry expressing OPN, MGP, VCAF mRNAs relative to 18S RNA. This figure is representative of 3 experiments performed on 3 differ-ent populations of cells.
Figure 7. Effect of dexamethasone (Dex) onNF-�B protein in pericytes. A, Immunofluores-cent staining of NF-�B in pericytes grown for 3weeks in the presence or absence of dexa-methasone. a and b, Representative micro-graphs of cells from four fields of view and 2experiments. Nuclei are counterstained withDAPI. Bar�100 �m. c and d, Magnified imagesof the boxed cells in a and b, respectively. B,Representative Western blot analysis of proteinextracts from pericytes cultured for 2 to 3weeks in the presence of dexamethasone withor without the addition of a GR antagonist, Org34116, showing a downregulation of NF-�B bydexamethasone and restoration to control lev-els after GR antagonist treatment. The blotswere reprobed with �-tubulin to act as a load-ing control. C, Western blot analysis of nuclearand cytoplasmic proteins probed with anNF-�B antibody, demonstrating downregulationof NF-�B in the nucleus after treatment of peri-cytes with dexamethasone.
1270 Circulation Research May 26, 2006
by guest on November 13, 2015http://circres.ahajournals.org/Downloaded from

whether BMP-2 expression is also modulated by dexa-methasone and whether this protein might also be involvedin the accelerated mineralization observed in dexametha-sone-treated pericytes.
There are reports indicating that dexamethasone-mediatedsuppression of mRNA occurs at the posttranscriptional levelas a result of mRNA destabilization.31 We investigatedwhether dexamethasone could downregulate MGP, OPN, andVCAF mRNA by increasing their mRNA degradation. How-ever, when transcription was blocked by actinomycin D, therewas no evidence for posttranscriptional control of gene expres-sion, as the half-lives of all 3 mRNAs were more than 8 hoursand unaffected by the presence of dexamethasone. These resultseliminate destabilization of MGP, OPN, and VCAF mRNAs asa mechanism by which dexamethasone downregulates theirexpression during pericyte differentiation.
Endogenous systemic glucocorticoids modulate the expres-sion of genes by activating the GR, which is a ligand-acti-vated transcription factor of the nuclear-receptor family.32
The GR resides in the cytoplasm as a multiprotein complex,which, on ligand binding, dissociates and translocates into thenucleus to up- or downregulate mRNA synthesis.32 We usedthe GR antagonist Org 34116 and demonstrate that it (1)blocks the dexamethasone-induced increase in nodule forma-tion and size, (2) blocks the dexamethasone-induced increasein calcium deposition, and (3) blocks the inhibitory effect ofdexamethasone on MGP mRNA expression. These datastrongly suggest that the dexamethasone–GR complex is alikely mediator not only of accelerated osteogenic differenti-ation and mineralization of pericytes but also of MGP generepression, possibly by direct action of the GR on a negativeglucocorticoid response element (GRE).32 In support of thissuggestion, 2 potential negative GREs33 are present in theMGP promoter sequence.34 In contrast, cyclohexamide treat-ment had no effect on the dexamethasone-induced decreaseof MGP, ruling out the involvement of another protein in thedownregulation of MGP mRNA by dexamethasone andadding strength to a direct effect on a negative GRE.
OPN mRNA remained downregulated after dexametha-sone plus Org 34116 treatment, eliminating transcriptionalregulation as a mechanism of control by dexamethasone. Thiswas a surprising result, because a GRE has been identifiedwithin the human osteopontin promoter.35 However, Wang etal have also suggested that under different conditions andwithin different cell types, the context of the GRE and othertranscription factors could influence the binding of the GR toa promoter.35 Cyclohexamide also had no effect on dexameth-asone-induced OPN downregulation, suggesting posttransla-tional control by the GR, perhaps involving a phosphorylationevent via another protein. In this regard, it is recognized thatphosphorylation of proteins is an important mechanism thatcontrols cellular events in the nucleus.36 These data indicatethat some form of alternative signaling and/or crosstalkbetween pathways is in play, adding support to the suggestionthat the GR is present in a range of multiprotein complexes(receptosomes) with different functions and means of generegulation.37 Clearly, a future study is warranted to under-stand the effects of dexamethasone on OPN gene regulation,because GR-interacting protein complexes may play a role in
the variability of glucocorticoid responsiveness observed insome patients.
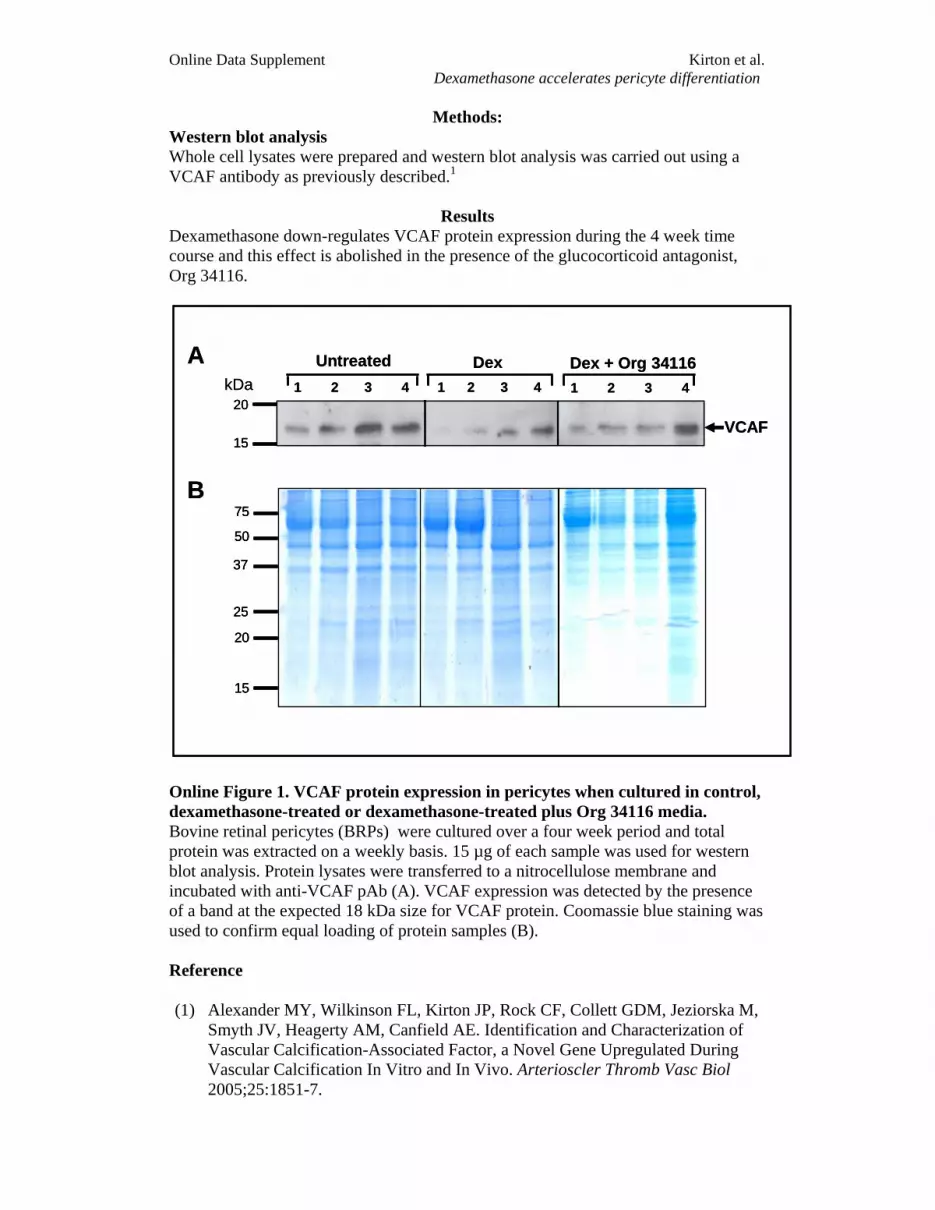
Org 34116 blocked the inhibitory effect of dexamethasoneon VCAF mRNA and protein expression (see the online datasupplement available at http://circres.ahajournals.org), which,like MGP, suggests involvement of the dexamethasone–GRcomplex possibly via a negative GRE. We also investigatedthe involvement of de novo protein synthesis in VCAFregulation by dexamethasone and found that, in contrast toMGP and OPN, cyclohexamide blocked dexamethasone-in-duced downregulation of VCAF mRNA. These results sug-gest a requirement for de novo protein synthesis as well,perhaps indicating protein–protein interaction with othersequence-specific transcription factors.
The most widespread advantage of glucocorticoids is theirantiinflammatory effect, thought to be attributable to a directinhibitory interaction between activated GRs and transcrip-tion factors, such as NF-�B.38 Inactive NF-�B is located inthe cytoplasmic compartment of the cell in association withthe I�-B inhibitory protein. On stimulation by inflammatoryagents, I�-B dissociates from NF-�B, allowing translocationof NF-�B to the nucleus, where it acts as a transcriptionfactor.39 We now demonstrate that NF-�B is negativelycontrolled at the level of transcription by dexamethasone,resulting in less NF-�B being translated into protein. Inaddition, we show, by immunocytochemistry, that dexameth-asone treatment results in an apparent decrease in NF-�Bprotein in the nucleus. Although immunofluorescence studiesmay be limited by the fact that cell shape changes caninfluence the amount of cytoplasm over the nucleus andhence the apparent amount of labeled NF-�B within thenucleus, we confirm these results using Western blot analysisand demonstrate a reduced level of NF-�B protein in thenuclear compartment after dexamethasone treatment. Theseresults suggest that this transcription factor may be involvedin the regulation of pericyte differentiation and mineralizationby dexamethasone. Furthermore, we suggest that the reduc-tion of NF-�B in the nucleus could be responsible for reducedtranscription of VCAF in pericytes after treatment withdexamethasone. However, further studies are required toverify the interaction and regulation of the GR multiproteincomplex, its effects on calcification-inhibiting proteins suchas VCAF, and the signaling pathways involved in theirregulation.
In conclusion, our results support the proposal that thevasculature could have a population of cells that, in responseto dexamethasone, differentiate toward an osteoblastic lin-eage by upregulating osteogenic markers and downregulatingspecific inhibitor molecules in both a direct and indirectmanner. Our data identify downstream targets of glucocorti-coid activation and suggest that elevated differentiation andmineralization of vascular pericytes by dexamethasone isassociated with the loss of expression of mineralizationinhibitory genes, which are controlled by dexamethasone atmultiple levels. This study unravels a novel mechanism fortranscriptional regulation by the GR and may be an importantcomponent in the cellular differentiation associated withvascular calcification. We speculate that glucocorticoids perse promote vascular calcification mostly by inhibiting
Kirton et al Dexamethasone Accelerates Pericyte Differentiation 1271
by guest on November 13, 2015http://circres.ahajournals.org/Downloaded from

calcification-inhibitor molecules as the main target, partly bydecreasing NF-�B transcription and activation. These dataprovide novel concepts regarding glucocorticoid-signalingpathways and vascular calcification. Identification of tissue-specific gene regulatory networks could yield insights intothe molecular basis of vascular calcification and may lead toimproved treatment regimes for diseases where glucocorti-coid therapy is in use.
AcknowledgmentsThis study was supported in part by grant PG/02/120/14492 from theBritish Heart Foundation (to M.Y.A. and A.E.C.) and by a Biologicaland Biochemical Sciences Research Council studentship (to J.P.K.).
References1. Bostrom K, Watson KE, Horn S, Wortham C, Herman I, Demer LL. Bone
morphogenetic protein expression in human atherosclerotic lesions. J ClinInvest. 1993;91:1800–1809.
2. Shanahan CM, Proudfoot D, Tyson KL, Cary NRB, Edmonds M,Weissberg PL. Expression of mineralisation-regulating proteins in asso-ciation with human vascular calcification. Z Kardiol. 2000;89:63–68.
3. Tintut Y, Alfonso Z, Saini T, Radcliff K, Watson K, Bostrom K, DemerLL. Multilineage potential of cells from the artery wall. Circulation.2003;108:2505–2510.
4. Collett GD, Canfield AE. Angiogenesis and pericytes in the initiation ofectopic calcification. Circ Res. 2005;96:930–938.
5. Doherty MJ, Ashton BA, Walsh S, Beresford JN, Grant ME, CanfieldAE. Vascular pericytes express osteogenic potential in vitro and in vivo.J Bone Min Res. 1998;13:828–838.
6. Watson KE, Bostrom K, Ravindranath R, Lam T, Nortin B, Demer LL.TGF-Beta-1 and 25-hydoxycholesterol stimulate osteoblast-like vascularcells to calcify. J Clin Invest. 1994;93:2106–2113.
7. Tintut Y, Patel J, Parhami F, Demer LL. Tumor necrosis factor-alphapromotes in vitro calcification of vascular cells via the cAMP pathway.Circulation. 2000;102:2636–2642.
8. Proudfoot D, Davies JD, Skepper JN, Weissberg PL, Shanahan CM.Acetylated low-density lipoprotein stimulates human vascular smoothmuscle cell calcification by promoting osteoblastic differentiation andinhibiting phagocytosis. Circulation. 2002;106:3044–3050.
9. Kiel DP, Kauppila LI, Cupples LA, Hannan MT, O’Donnell CJ, WilsonPWF. Bone loss and the progression of abdominal aortic calcificationover a 25 year period: The Framingham Heart Study. Calcif Tissue Int.2001;68:271–276.
10. Fishel RS, Eisenberg S, Shai SY, Redden RA, Bernstein KE, Berk BC.Glucocorticoids induce angiotensin-converting enzyme expression invascular smooth muscle. Hypertension. 1995;25:343–349.
11. Cheng S, Yang J, Rifas L, Zhang S, Avioli L. Differentiation of humanbone marrow osteogenic stromal cells in vitro: induction of the osteoblastphenotype by dexamethasone. Endocrinology. 1994;134:277–286.
12. Walsh S, Jordan GR, Jefferiss C, Stewart K, Beresford JN. High concen-trations of dexamethasone suppress the proliferation but not the differ-entiation or further maturation of human osteoblast precursors in vitro:relevance to glucocorticoid-induced osteoporosis. Rheumatology. 2001;40:74–83.
13. Bellows CG, Heersche JNM, Aubin JE. Determination of the capacity forproliferation and differentiation of osteoprogenitor cells in the presenceand absence of dexamethasone. Dev Biol. 1990;140:132–138.
14. Mori K, Shioi A, Jono S, Nishizawa Y, Morii H. Dexamethasoneenhances in vitro vascular calcification by promoting osteoblastic differ-entiation of vascular smooth muscle cells. Arterioscler Thromb Vasc Biol.1999;19:2112–2118.
15. Shui C, Scutt AM. Mouse embryo-derived NIH3T3 fibroblasts adopt anosteoblast-like phenotype when treated with 1a,25-dihydroxyvitamin D3and dexamethasone in vitro. J Cell Physiol. 2002;193:164–172.
16. Collett G, Wood A, Alexander MY, Varnum BC, Boot-Handford RP,Ohanian V, Ohanian J, Fridell YW, Canfield AE. Receptor tyrosinekinase Axl modulates the osteogenic differentiation of pericytes. CircRes. 2003;92:1123–1129.
17. Parhami F, Morrow AD, Balucan J, Leitinger N, Watson AD, Tintut Y,Berliner JA, Demer LL. Lipid oxidation products have opposite effects on
calcifying vascular cell and bone cell differentiation-A possible expla-nation for the paradox of arterial calcification in osteoporotic patients.Arterioscler Thromb Vasc Biol. 1997;17:680–687.
18. Alexander MY, Wilkinson FL, Kirton JP, Rock CF, Collett GDM,Jeziorska M, Smyth JV, Heagerty AM, Canfield AE. Identification andcharacterization of vascular calcification-associated factor, a novel geneupregulated during vascular calcification in vitro and in vivo. ArteriosclerThromb Vasc Biol. 2005;25:1851–1857.
19. Jeziorska M, McCollum C, Wooley DE. Observations on bone formationand remodelling in advanced atherosclerotic lesions of human carotidarteries. Virchows Arch. 1998;433:559–565.
20. Bostrom K, Watson KE, Stanford WP, Demer LL. Atheroscleroticcalcification-relation to developmental osteogenesis. Am J Cardiol. 1995;75:B88–B91.
21. Banks LM, Lees B, MacSweeney JE, Stevenson JC. Effect of degen-erative spinal and aortic calcification on bone density measurements inpost-menopausal women: links between osteoporosis and cardiovasculardisease? Eur J Clin Invest. 1994;24:813–817.
22. Purpura KA, Zandstra PW, Aubin JE. Fluorescence activated cell sortingreveals heterogeneous and cell non-autonomous osteoprogenitor differ-entiation in fetal rat calvaria cell populations. J Cell Biochem. 2003;90:109–120.
23. Purpura KA, Aubin JE, Zandstra PW. Sustained in vitro expansion ofbone progenitors is cell density dependent. Stem Cells. 2004;22:39–50.
24. Smith E, Coetzee GA, Frenkel B. Glucocorticoids inhibit cell cycleprogression in differentiating osteoblasts via glycogen synthase kinase-3beta. J Biol Chem. 2002;277:18191–18197.
25. Stein GS, Lian JB. Molecular mechanisms mediating proliferation/differentiation interrelationships during progressive development of theosteoblast phenotype. Endocr Rev. 1993;14:424–442.
26. Canfield AE, Doherty MJ, Kelly V, Newman B, Farrington C, Grant ME,Boot-Handford RP. Matrix Gla protein is differentially expressed duringthe deposition of a calcified matrix by vascular pericytes. FEBS Lett.2000;487:267–271.
27. Jono S, Peinado C, Giachelli CM. Phosphorylation of osteopontin isrequired for inhibition of vascular smooth muscle cell calcification. J BiolChem. 2000;275:20197–20203.
28. Luo GB, Ducy P, McKee MD, Pinero GJ, Loyer E, Behringer RR,Karsenty G. Spontaneous calcification of arteries and cartilage in micelacking matrix GLA protein. Nature. 1997;386:78–81.
29. Zebboudj AF, Shin V, Bostrom K. Matrix GLA protein and BMP-2regulate osteoinduction in calcifying vascular cells. J Cell Biochem.2003;90:756–765.
30. Garfinkel A, Tintut Y, Petrasek D, Bostrom K, Demer LL. Patternformation by vascular mesenchymal cells. Proc Natl Acad Sci U S A.2004;101:9247–9250.
31. Ristimaki A, Narko K, Hla T. Down-regulation of cytokine-inducedcyclo-oxygenase-2 transcript isoforms by dexamethasone: evidence forpost-transcriptional regulation. Biochem J. 1996;318:325–331.
32. Schoneveld OJLM, Gaemers IC, Lamers WH. Mechanisms of glucocor-ticoid signalling. Biochim Biophys Acta. 2004;1680:114–128.
33. Moehren U, Eckey M, Baniahmad A. Gene repression by nuclearhormone receptors. In: McEwan I, ed. The Nuclear Receptor Superfamily.London: Portland Press; 2004:89–104.
34. Cancela L, Hsieh CL, Francke U, Price PA. Molecular-structure, chro-mosome assignment, and promoter organization of the human matrix Glaprotein gene. J Biol Chem. 1990;265:15040–15048.
35. Wang D, Yamamoto S, Hijiya N, Benveniste EN, Gladson CL. Tran-scriptional regulation of the human osteopontin promoter: functionalanalysis and DNA-protein interactions. Oncogene. 2000;19:5801–5809.
36. Carlson A, Yates KE, Slamon DJ, Gasson JC. Spatial and temporalchanges in the subcellular localization of the nuclear protein-tyrosinekinase, c-Fes. DNA Cell Biol. 2005;24:225–234.
37. Wikstrom A-C. Glucocorticoid action and novel mechanisms of steroidresistance: role of glucocorticoid receptor-interacting proteins for glu-cocorticoid responsiveness. J Endocrinol. 2003;178:331–337.
38. Nelson G, Wilde GJ, Spiller DG, Kennedy SM, Ray DW, Sullivan E,Unitt JF, White MR. NF-kappaB signalling is inhibited by glucocorticoidreceptor and STAT6 via distinct mechanisms. J Cell Sci. 2003;116:2495–2503.
39. Barnes PJ, Karin M. Nuclear factor-kappaB: a pivotal transcription factorin chronic inflammatory diseases. N Engl J Med. 1997;336:1066–1071.
1272 Circulation Research May 26, 2006
by guest on November 13, 2015http://circres.ahajournals.org/Downloaded from
Online Data Supplement Kirton et al. Dexamethasone accelerates pericyte differentiation
Methods: Western blot analysis Whole cell lysates were prepared and western blot analysis was carried out using a VCAF antibody as previously described.1
Results Dexamethasone down-regulates VCAF protein expression during the 4 week time course and this effect is abolished in the presence of the glucocorticoid antagonist, Org 34116.
Online Figure 1. VCAF protein expression in pericytes when cultured in control, dexamethasone-treated or dexamethasone-treated plus Org 34116 media. Bovine retinal pericytes (BRPs) were cultured over a four week period and total protein was extracted on a weekly basis. 15 µg of each sample was used for western blot analysis. Protein lysates were transferred to a nitrocellulose membrane and incubated with anti-VCAF pAb (A). VCAF expression was detected by the presence of a band at the expected 18 kDa size for VCAF protein. Coomassie blue staining was used to confirm equal loading of protein samples (B). Reference
(1) Alexander MY, Wilkinson FL, Kirton JP, Rock CF, Collett GDM, Jeziorska M,
Smyth JV, Heagerty AM, Canfield AE. Identification and Characterization of Vascular Calcification-Associated Factor, a Novel Gene Upregulated During Vascular Calcification In Vitro and In Vivo. Arterioscler Thromb Vasc Biol 2005;25:1851-7.
kDaUntreated
1 2 3 4 1 2 3 4
Dex 1 2 3 4
Dex + Org 34116 A
VCAF
B75
15
20
25
50
37
15
20kDa
Untreated1 2 3 4 1 2 3 4
Dex 1 2 3 4
Dex + Org 34116 A
VCAF
B75
15
20
25
50
37
75
15
20
25
50
37
15
20
15
20

John Paul Kirton, Fiona L. Wilkinson, Ann E. Canfield and M. Yvonne AlexanderOsteogenic Differentiation of Vascular Pericytes: Implications for Vascular Calcification
Dexamethasone Downregulates Calcification-Inhibitor Molecules and Accelerates
Print ISSN: 0009-7330. Online ISSN: 1524-4571 Copyright © 2006 American Heart Association, Inc. All rights reserved.is published by the American Heart Association, 7272 Greenville Avenue, Dallas, TX 75231Circulation Research
doi: 10.1161/01.RES.0000223056.68892.8b2006;98:1264-1272; originally published online April 20, 2006;Circ Res.
http://circres.ahajournals.org/content/98/10/1264World Wide Web at:
The online version of this article, along with updated information and services, is located on the
http://circres.ahajournals.org/content/suppl/2006/04/20/01.RES.0000223056.68892.8b.DC1.htmlData Supplement (unedited) at:
http://circres.ahajournals.org//subscriptions/
is online at: Circulation Research Information about subscribing to Subscriptions:
http://www.lww.com/reprints Information about reprints can be found online at: Reprints:
document. Permissions and Rights Question and Answer about this process is available in the
located, click Request Permissions in the middle column of the Web page under Services. Further informationEditorial Office. Once the online version of the published article for which permission is being requested is
can be obtained via RightsLink, a service of the Copyright Clearance Center, not theCirculation Researchin Requests for permissions to reproduce figures, tables, or portions of articles originally publishedPermissions:
by guest on November 13, 2015http://circres.ahajournals.org/Downloaded from