developmental expression of heme oxygenase-1 (hsp32) in rat brain: an immunocytochemical study
TRANSCRIPT

Ž .Developmental Brain Research 105 1998 181–194
Research report
ž /Developmental expression of heme oxygenase-1 HSP32 in rat brain:an immunocytochemical study
Marcelle Bergeron a,c,), Donna M. Ferriero b,c, Frank R. Sharp a,c
a Department of Neurology, Veterans Affairs Medical Center, San Francisco, CA 94121, USAb Department of Pediatrics, UniÕersity of California, San Francisco, CA 94121, USAc Department of Neurology, UniÕersity of California, San Francisco, CA 94121, USA
Accepted 23 September 1997
Abstract
Ž .Heme oxygenase HO is a microsomal enzyme that oxidatively cleaves heme molecules to produce bile pigments, iron and carbonmonoxide. In normal adult rat brain, HO-2 is the most abundant isozyme whereas HO-1 is present at very low levels except in select cellpopulations. Because its promoter region has NF-kB and AP-1 sites, heat-shock and heme-responsive elements, the HO-1 isozyme can beinduced by a variety of stimuli. Since the expression and activity of several transcription factors such as NF-kB, FosrJun, and CREBshow specific changes during development, we postulated that HO-1 expression may show similar developmental regulation. Usingimmunocytochem istry and Western blotting, this study demonstrates the developmental changes of HO-1 protein expression in normal
Ž .brain from rats at postnatal day 7 P7 , P14, P21 and adult. Brain HO-1 immunoreactivity was highest at P7 in most brain regionsincluding the white matter in areas of myelinogenesis, cerebral cortex, hippocampus, thalamus and hypothalamus and, in the blood vesselendothelial cells throughout the brain. In most regions, the adult pattern was reached by P21 with HO-1 protein localized almostexclusively to the dentate regions of hippocampus, some thalamic and hypothalamic nuclei, with little or no staining of endothelium,white matter and cortex. In a few select areas such as the substantia nigra, globus pallidus, ventromedial hypothalamic nucleus and thelateral preoptic nuclei area, little or no cellular HO-1 staining was observed at P7 whereas increased staining was found with maturationand adulthood. These results show that HO-1 protein expression is regulated in different cell types of specific regions of the rat brainduring development. q 1998 Elsevier Science B.V.
Keywords: Brain; Development; Heat-shock protein; Heme oxygenase; HSP32; Neonatal
1. Introduction
Ž .Heme oxygenase HO is the rate-limiting enzyme thatmetabolizes heme molecules to biliverdin, ferrous ironŽ 2q. Ž .Fe and carbon monoxide CO . Biliverdin reductase
w xconverts biliverdin to bilirubin 20 . Two different geneŽ .products of HO microsomal isozymes HO-1 and HO-2
w xhave been characterized 38,53 . In normal mature ratbrain, the HO-2 isozyme is responsible for most of HOconstitutive activity since the HO-1 isozyme is present at
Abbreviations: BSA, bovine serum albumin; HO, heme oxygenase;HSP, heat-shock protein; mRNA, messenger RNA; NF-kB, nuclear factorkappa B; P7, postnatal day 7; PB, sodium phosphate buffer; SDS, sodiumdodecyl sulfate; SDS-PAGE, SDS–polyacrylamide gel electrophoresis
) Ž .Corresponding author. Department of Neurology V127 , UCSF andVA Medical Center, 4150 Clement Street, San Francisco, CA 94121,USA. Tel.: q1-415-750-2011; Fax: q1-415-750-2273; E-mail:[email protected]
w xvery low levels 17,35,58 . In contrast to HO-2 expressionwhich is not inducible by most types of stress, brain HO-1
w xexpression is markedly induced by hyperthermia 17 , glu-w x w xtathione depletion 19 , subarachnoid hemorrhage 61 ,
w x w xtrauma 22 and ischemia 8,43 .During the course of development, brain function and
plasticity depend on the coordinated expression of multiplegenes that are controlled by specific transcription factors.
ŽTranscription factors such as nuclear factor kappa B NF-.kB , FosrJun and CREB are developmentally regulated in
rat brain, the highest activity often occurring just afterw xbirth 2,5,13,48,49 . HO-1 is a potential target gene for
these transcription factors. Indeed, the promoter of HO-1gene contains several regulatory elements including: aNF-kB site that may mediate a response to oxidative stressw x33 ; an AP-1-binding site that may mediate a response to
w xFosrJun family members 1,41 ; a heat-shock element thatmakes it responsive to heat shock andror denatured pro-
0165-3806r98r$19.00 q 1998 Elsevier Science B.V. All rights reserved.Ž .PII S0165-3806 97 00169-7

( )M. Bergeron et al.rDeÕelopmental Brain Research 105 1998 181–194182
w xteins 47 ; and a heme-response element that renders thew xgene responsive to heme 41 .
Studies in rodents have demonstrated that the brainbecomes vulnerable to excitotoxic neuronal damage duringpostnatal development, in parallel with glutamate receptor
w xsensitivity 29,39 . Stimulation of glutamate receptors canw xactivate the transcription factor NF-kB 30 and Fos-related
w x Ž .factors 55,59 in neurons. CO like nitric oxide has beenproposed to act as a putative neurotransmitter and regula-
w xtor of guanylyl cyclase 36,62 . Since activation ofmetabotropic glutamate receptors modulates neuronal HO
w xactivity 24,42 and, in turn, CO may modulate the releasew xof glutamate at the synapse 54 , changes in HO-1 activity
could modulate brain development and maturation. Ac-cordingly, changes in HO-1 expression may explain, inpart, the known difference in vulnerability of newborn rat
Ž .brain to hypoxicrischemic excitotoxic cell injury com-w xpared to adult 29 . In order to define the role of HO-1 in
brain development, the present study describes the devel-opmental profile of brain HO-1 protein expression in P7-,P14-, P21-day-old and adult rats by immunocytochemistryand Western blot analyses. Regions of the forebrain anddiencephalon were primarily studied because of their par-
Ž .ticular vulnerability to hypoxicrischemic excitotoxicw xbrain damage 8,21,43 .
2. Materials and methods
2.1. Animals
All experiments were performed in strict accordancewith the National Institutes of Health Guide for Care andUse of Laboratory Animals, and all protocols were ap-proved by the University of California at San FranciscoCommittee on Animal Research. Male and female
Ž .Sprague–Dawley rats Bantam Kingman, Fremont, CA at7, 14, 21 days after birth and adult rats were used for the
Ž .study ns6rage . To minimize the potential variations inHO-1 expression that may arise from changes in circadianrhythms, excessive handling and stress, animals from P7,P14, P21 and adult groups were all received on the sameday and processed upon arrival. Hence, all animals wouldlikely to have experienced handling stress though thisshould have been similar for each group. Preliminaryresults suggest that the stress of handling does not signifi-cantly affect HO-1 staining in the brain.
2.2. Western blot analysis
Rats were deeply anesthetized with an intraperitonealŽ . Ž .injection of ketamine 80 mgrkg and xylazine 8 mgrkg
and decapitated. The brains were quickly removed anddissected on ice. The hippocampus and cerebral cortex
Žwere dropped in Laemmli solubilizing buffer 2.5% sodiumŽ .dodecyl sulfate SDS , 10% glycerol, 62.5 mM Tris-HCl,
.pH 6.8, 5% 2-mercaptoethanol and boiled for 10 min.Whole tissue extracts were stored at y708C. Westernimmunoblot analysis was performed as described previ-
w xously 8 . Protein concentration was determined using theŽ .bicinchoninic acid method Pierce, Rockford, IL . Equal
Ž .protein samples 55 mg were separated on 12% SDSŽpolyacrylamide gels with 4.5% SDS stacking gels SDS-
.PAGE . After electrotransfer onto a nitrocellulose mem-Ž .brane 0.2 mm; Schleicher and Schuell, Keene, NH , im-
mobilized proteins were stained with Ponceau solution toverify equal protein loading and homogeneous transfer.Membranes were incubated overnight at 48C in 0.1 M
Ž .sodium phosphate buffer PB , pH 7.4 containing 5%Ž .non-fat dry milk, 1% bovine serum albumin BSA and
0.1% Tween-20. Membranes were then washed with 0.1 MPB containing 1% BSA and 0.1% Tween-20 and incubatedfor 1.5 h with a 1:3500 dilution of a rabbit polyclonal
Ž .anti-rat HO-1 antibody StressGen, Victoria, BC, Canada .This polyclonal antibody was raised against rat liver puri-fied HO-1 protein and characterized by Maines and collab-
w xorators 38 . Since this antibody was not affinity purified,we verified its specificity by analyzing samples of rat liver
Ž .HO-1 purified protein StressGen, Victoria, BC, Canadain parallel with the brain tissue samples. In the purifiedliver HO-1 protein samples, the antibody recognized onemajor band of 32 kDa and two minor bands of approxi-
Ž .mately 30 and 34 kDa, respectively not shown . In crudebrain extracts, the same antibody only recognized a single32 kDa band and not any other bands. Following washes,membranes were incubated with a 1:2500 dilution of anti-
Žrabbit Ig-horseradish peroxidase antibody Amersham, Ar-.lington Heights, IL for 1.0 h and bound antibody was
visualized with the ECL chemiluminescence systemŽ . ŽAmersham . A computer-based imaging system MCID,
.Imaging Research, St. Catherines, Ont., Canada was usedto measure the densities and areas of HO-1 protein expres-sion on Western immunoblots. The relative density of
Ž .HO-1 protein 32 kDa bands was analyzed followingsubtraction of the film background.
2.3. Immunocytochemistry
Rats were deeply anesthetized as described above andŽ .perfused through the left ventricle via the ascending aorta
with cold 0.9% NaCl solution followed by cold 4% para-Ž .formaldehyde PFA made up in 0.1 M PB, pH 7.4. Brains
were removed, post-fixed in 4% PFA for 4 h and stored in30% sucrose overnight at 48C. Fifty-micrometer-thickcoronal sections were cut on a vibratome and washedtwice with 0.05 M PB, pH 7.4. Following inhibition ofendogenous peroxidase in 0.05 M PB, pH 7.4 containing0.65% sodium azide and 0.2% hydrogen peroxide, sectionswere incubated for 1–2 h in 0.1 M PB containing 5%non-fat dry milk, 1% BSA, 2% goat serum, 0.1% TritonX-100 and 0.1% rat serum. Sections were then incubatedovernight at 48C with the same anti-HO-1 antibody used

( )M. Bergeron et al.rDeÕelopmental Brain Research 105 1998 181–194 183
Fig. 1. Western immunoblot analysis of developmental changes in ratŽbrain HO-1 protein level. Whole tissue extracts of cerebral cortex lanes
. Ž .1, 3, 5, 7 and hippocampus lanes 2, 4, 6, 8 were prepared as describedŽ .in Section 2 and equal protein samples 55 mg were analyzed by
SDS-PAGE and immunoblotting using rabbit anti-rat HO-1 antibody.Lanes 1,2: rat at P7; lanes 3,4: rat at P14; lanes 5,6: rat at P21; lanes 7,8:adult rat. Molecular weight markers in kilodalton are indicated on theright. This experiment was repeated 3 times with similar results.
Žfor Western blotting rabbit polyclonal anti-rat HO-1;.StressGen, Victoria, BC, Canada diluted 1:4000 in 0.1 M
PB containing 1% BSA, 2% goat serum and 0.1% Tritonw xX-100. As previously described in our laboratory 8,22,61 ,
negative controls included alternate sections from eachbrain incubated either without the primary antibody orwith some antiserum which had been pre-absorbed with
Žexcess rat HO-1 purified protein StressGen, Victoria, BC,.Canada . After 3 washes in 0.05 M PB, sections were
incubated at room temperature for 2 h with a 1:200Ždilution of biotinylated goat anti-rabbit IgG antibody Vec-
.tor Laboratories, Burlingame, CA . Sections were thenincubated in an avidin-horseradish peroxidase solutionŽ .Elite Vectastain, Vector Laboratories for 2 h, followedby washes with 0.05 M PB. Visualization of HO-1 stainingwas performed by incubating the sections with 0.015%3,3-diaminobenzidine tetrachloride as the chromogenŽ .Sigma, St. Louis, MO in the presence of 0.01% H O2 2
made up in 0.05 M PB. After washing, sections weremounted on gelatin-coated slides, dried and coverslipped.
3. Results
Western immunoblots showed an age-dependent reduc-Ž .tion in brain HO-1 protein expression Fig. 1 . HO-1
protein, visualized as a single ;32 kDa band on im-munoblots, was highest at P7 in all regions studied includ-
Ž .ing the hippocampus, the cerebral cortex Fig. 1 as well asŽ .the thalamic and hypothalamic areas not shown . HO-1
protein decreased 2–3-fold by P21 and was at lowest levelŽ .by adulthood Fig. 1 .
As summarized in Table 1, immunocytochemistryshowed similar results with cellular HO-1 immunoreactiv-
Žity being generally higher at P7 Figs. 2–6A–D,E–HFig.. Ž7A–DFig. 8C,D with some regional exceptions Fig.
.7E–HFig. 8A,B,E,FFig. 8G,H . The region-specific HO-1Ž .expression observed in adult rat brain see Table 1 was
w xsimilar to that reported previously 17,18,63 . Interestingly,the background staining was also higher at P7 compared
Ž .with more mature brains Fig. 4 . Previous studies withdifferent antibodies have also reported similar observations
w x20,21 . This phenomenon could be due to differences inthe degree of brain myelination andror protein composi-
w xtion. As shown previously in our laboratory 8,22,61 ,immunostaining was not noted in brain sections when theprimary antibody was pre-absorbed with its peptide oromitted from the incubation. Consistent with the microso-
w xmal localization of HO-1 protein 35,38 , cellular staininggenerally appeared to be mostly perinuclear in nature.HO-1 immunostaining was prominent in several distinctbrain regions at P7 and was particularly intense in areas ofmyelinogenesis of the white matter such as the cingulumŽ .Fig. 2A . The morphology of the HO-1-positive cellsfound in the cingulum was characterized by small round
Ž .and dark cell bodies with few processes Fig. 2A . HO-1immunoreactivity in P7 rat brain was also prominent in the
Žcorpus callosum, the external and internal capsule Fig.
Table 1Summary of the age-dependent distribution of HO-1 immunoreactivity inthe brain of Sprague–Dawley rats
Brain region P7 P14 P21 Adult
Cerebral cortexFrontal qq qq q yRetrosplenial qqq qqq qq yPiriform-entorhinal qq qq q qParietal qq qq q y
White matterCorpus callosum qqq qq yrq yCingulum qqq qq yrq yInternal capsule qqq qq yrq yExternal capsule qqq qq yrq y
Vascular endothelium qqq qq yrq yHippocampus
CA1–CA3Stratum oriens qqq qqq qq qPyramidal layer qqq qqq qq qStratum lucidum qq q q y
Dentate gyrus qqq qqq qqq qqqBasal ganglia
Caudate-putamen yrq yrq yrq yrqGlobus pallidus yrq q q q
ThalamusMediodorsal n. qqq qq q yReuniens n. qq qq qq qqAnterodorsal n. qq qq qq qqVentromedial n. qq qq qq qq
HypothalamusVentromedial n. q qq qqq qqqParaventricular n. qq qq qq qqMagnocellular preoptic n. q qqq qqq qqq
Amygdala basal n. qqq qq q ySubstantia nigra
Pars compacta yrq q qq qqPars reticulata y yrq yrq q
Scores are based on the relative density and intensity of cells positivelyimmunostained for HO-1 within each respective brain region. Scoresrepresent a compilation of observations made by two different observers.y: not detectable; q: occasional HO-1-positive cells with low-to-mod-erate intensity staining; qq: moderate number of cells with moderate-to-high intensity staining; qqq: high cell density of moderate-to-highintensity staining.

Fig. 2. Developmental changes of HO-1 protein expression in rat cingulum. HO-1 immunoreactivity in 50-mm-thick comparable coronal sections from ratŽ . Ž . Ž . Ž . Ž . Ž .brain at P7 A , P14 B , P21 C , and in adult D . Note the intense staining of round cells at P7 A; arrow which gradually decreased by P14 B andŽ . Ž . Ž .with maturity C,D . HO-1 immunoreactive cells macrophage-like in the white matter tracts of the corpus callosum cc as well as HO-1-positive neurons
Ž . Ž . Ž .in the stratum oriens so , pyramidal cell layer py and stratum lucidum sl are also readily detectable at P7, with fewer cells staining for HO-1 by P21Ž . Ž .C and in adult D . Scale bar: 100 mm.

( )M. Bergeron et al.rDeÕelopmental Brain Research 105 1998 181–194 185
Ž . Ž .Fig. 3. Developmental changes of HO-1 expression in 50-mm-thick comparable coronal sections showing the cerebral cortex from rats at P7 A , P14 B ,Ž . Ž . Ž .P21 C , and in adult D . Note the characteristic endothelial and high background staining associated with P7 brain A , decreasing gradually with
Ž . Ž .maturity B–D . Of interest are some occasional clusters of intense HO-1 immunostained cells A; arrows which are characteristic of P7 animals.Scattered HO-1-positive neurons and some microglia-like cells were detected throughout cortex at all ages investigated. Scale bar: 100 mm.
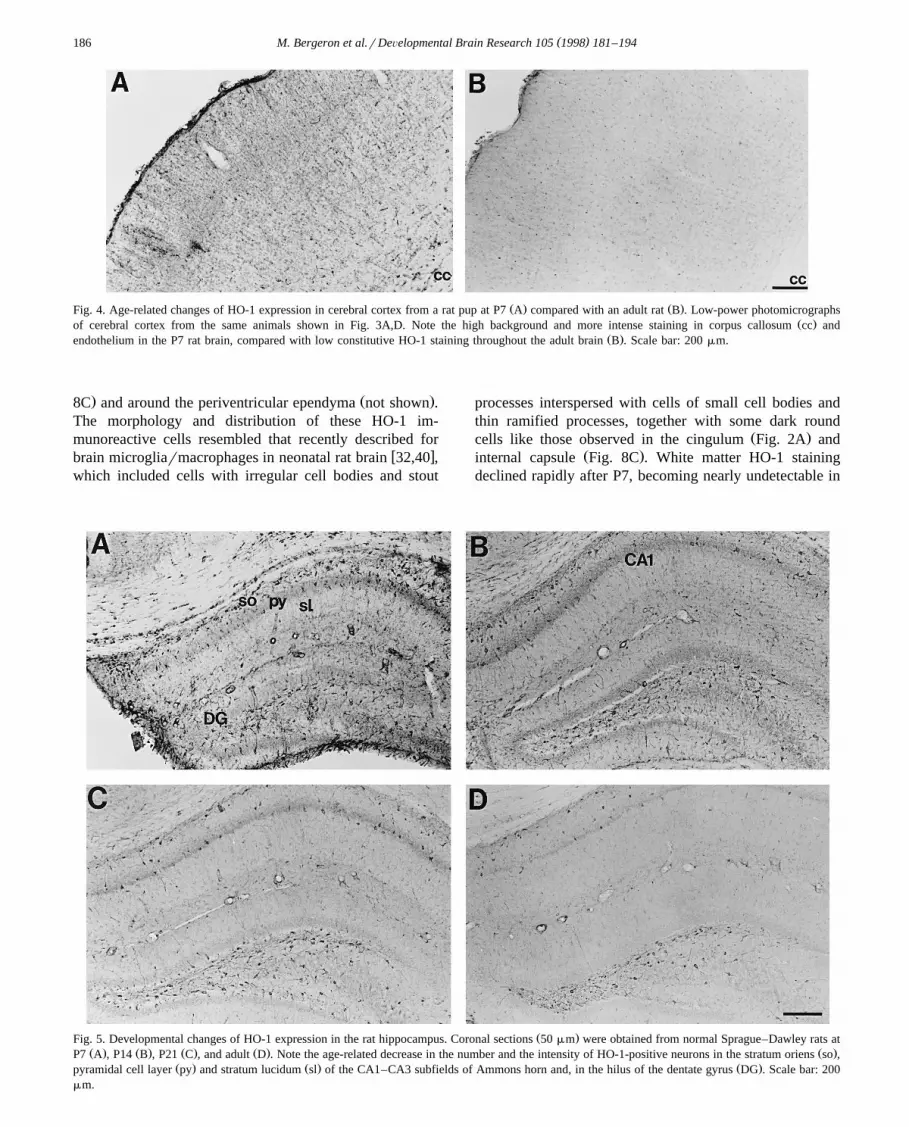
( )M. Bergeron et al.rDeÕelopmental Brain Research 105 1998 181–194186
Ž . Ž .Fig. 4. Age-related changes of HO-1 expression in cerebral cortex from a rat pup at P7 A compared with an adult rat B . Low-power photomicrographsŽ .of cerebral cortex from the same animals shown in Fig. 3A,D. Note the high background and more intense staining in corpus callosum cc and
Ž .endothelium in the P7 rat brain, compared with low constitutive HO-1 staining throughout the adult brain B . Scale bar: 200 mm.
. Ž .8C and around the periventricular ependyma not shown .The morphology and distribution of these HO-1 im-munoreactive cells resembled that recently described for
w xbrain microgliarmacrophages in neonatal rat brain 32,40 ,which included cells with irregular cell bodies and stout
processes interspersed with cells of small cell bodies andthin ramified processes, together with some dark round
Ž .cells like those observed in the cingulum Fig. 2A andŽ .internal capsule Fig. 8C . White matter HO-1 staining
declined rapidly after P7, becoming nearly undetectable in
Ž .Fig. 5. Developmental changes of HO-1 expression in the rat hippocampus. Coronal sections 50 mm were obtained from normal Sprague–Dawley rats atŽ . Ž . Ž . Ž . Ž .P7 A , P14 B , P21 C , and adult D . Note the age-related decrease in the number and the intensity of HO-1-positive neurons in the stratum oriens so ,
Ž . Ž . Ž .pyramidal cell layer py and stratum lucidum sl of the CA1–CA3 subfields of Ammons horn and, in the hilus of the dentate gyrus DG . Scale bar: 200mm.

Ž . Ž .Fig. 6. Developmental profile of HO-1 expression in the rat mediodorsal thalamic nucleus A–D and retrosplenial cortex E–H . Comparable coronalŽ . Ž . Ž . Ž . Ž .sections 50 mm were obtained from normal Sprague–Dawley rats at P7 A,E , P14 B,F , P21 C,G , and adult D,H . The mediodorsal thalamic nucleus
is located directly below the habenular nuclei and the third ventricle whereas the retrosplenial cortex is found in both hemispheres immediately adjacent tothe longitudinal fissure. Note the age-related decrease in the number of HO-1-positive neurons in both regions. Scale bar: 100 mm.
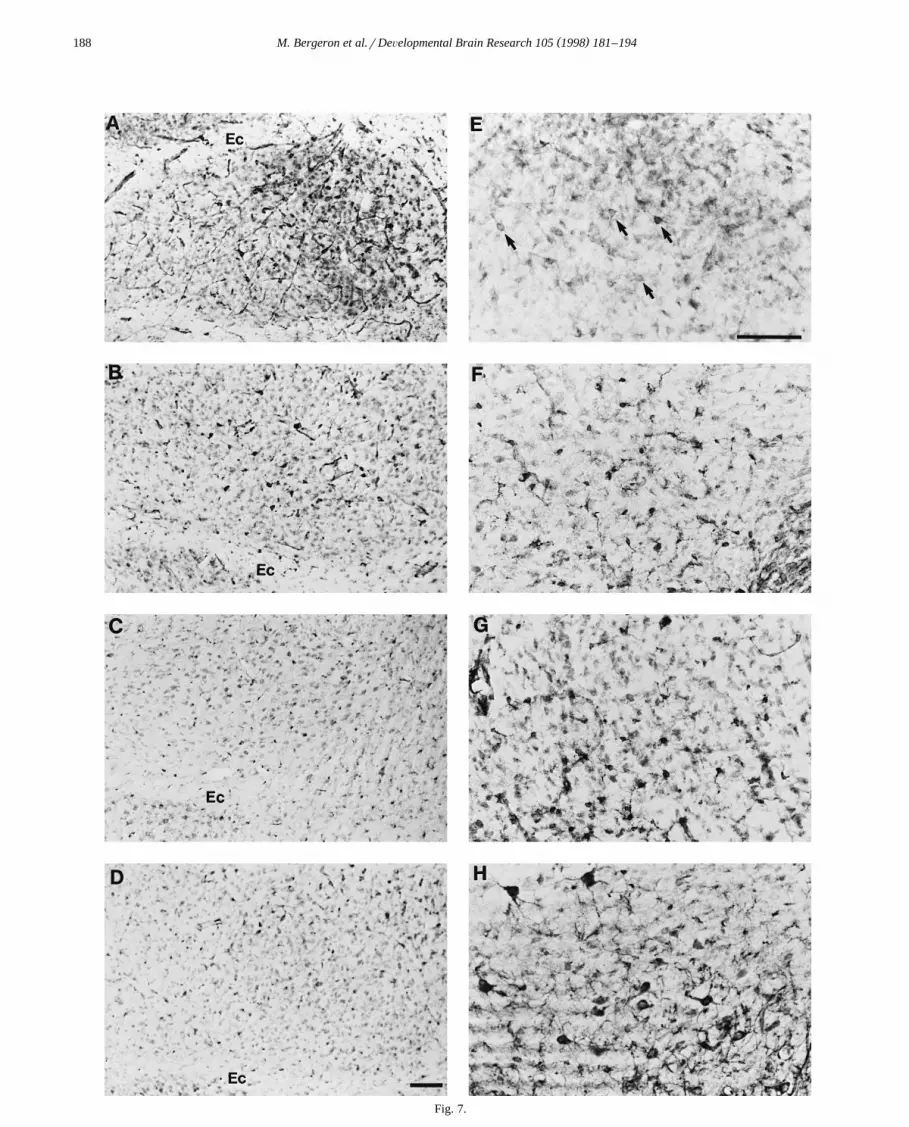
( )M. Bergeron et al.rDeÕelopmental Brain Research 105 1998 181–194188
Fig. 7.

( )M. Bergeron et al.rDeÕelopmental Brain Research 105 1998 181–194 189
Ž .adult brain Fig. 2B–DFig. 8D . HO-1 protein was alsoexpressed in the endothelium throughout the brain, with
Žmaximal staining at P7 Figs. 2A–5A, 6–7A,E, and.8A,C,E,G . Endothelial HO-1 expression declined progres-
sively with age, with no apparent staining in adult rat brainŽ .Figs. 2–8 .
In the cerebral cortex, HO-1 staining was highest in P7Ž .brains and declined with age Figs. 3 and 4 . HO-1 protein
was expressed in a variety of cortical cell types includingglia-like cells, endothelial cells and some scattered neuronsŽ . Ž .Fig. 3A–D . In the retrosplenial cortex Fig. 6E–H ,intense HO-1 expression was observed in several neuronsat P7–P14. The number and intensity of neuronal staining
Ž .in this area declined significantly by P21 Fig. 6G , be-Ž .coming undetectable in adults Fig. 6H . In the entorhinal-
Ž .piriform cortex not shown , moderately stained neuronswere found at P7, increasing slightly in apparent size andnumber by P14 and older. Notably, intensely stained cellclusters were found occasionally in different areas of
Ž .cortex, in P7 rat almost exclusively Fig. 3A, Fig. 4A . Onrare occasions, similar cell clusters were also observed inother brain areas such as the molecular and granular layersof the hippocampus, the striatum and the thalamus. Doublefluorescence labeling studies using antibodies recognizing
Žmarkers specific for neurons neuron-specific enolase;. Ž .NSE astrocytes glial fibrillary acidic protein; GFAP and
Ž .microgliarmacrophages OX-42 and ED-1 revealed thatsome of the cells in these clusters were either microglia or
Ž .astrocytes but not neurons data not shown . However, thetrue identity of some other cells in these clusters is stillunknown.
In the hippocampus, HO-1 staining was observed at P7Ž .in neurons of the hilus of the dentate gyrus DG , the
Ž . Ž .stratum oriens so , the pyramidal cell layer py and theŽ .stratum lucidum sl of the CA1–CA3 subfields of Am-
Ž .mons horn Fig. 2AFig. 5A . This pattern was similar toŽ .that observed in later stages of development Fig. 5B–D
except that the apparent intensity and number of stainedcells diminished progressively with age in the molecular
Ž .and granular layers of CA1–CA3 fields Figs. 2 and 5 . ByP21 and adult stages, stained hippocampal neurons withsmall cell bodies and thin dendritic processes were foundprimarily in the polymorphic layer of the hilus of the DGŽ .Fig. 5C,D .
In some thalamic nuclei such as the mediodorsal nu-cleus which is located directly underneath the habenularnuclei and the third ventricle, the density and intensity of
neuronal staining was highest at P7 and declined rapidlyŽby P14 and P21, becoming undetectable by adulthood Fig.
.6A–D . In other thalamic regions such as the reuniens andŽ .anterodorsal nuclei not shown as well as the ventrome-
Ž .dial nucleus adjacent to the zona incerta Zi and mammil-Ž . Ž .lothalamic tract Mt Fig. 8A,B , HO-1 staining intensity
was low to moderate and the relative density did not seemto be affected by maturation.
In the ventral subcortical brain regions, an age-depen-dent reduction in the number and intensity of cell stainingwas observed in the nucleus basalis of the amygdala with
Žthe highest level of HO-1 expression found at P7 Fig..7A–D . In contrast, whereas very few cells expressed
HO-1 protein in the ventromedial nucleus of the hypothala-mus at P7, maturational increases in the number, size andstaining intensity of HO-1-positive neurons was observed
Ž .after P14 through adulthood Fig. 7E–H . In the magnocel-lular preoptic area, few faint HO-1-positive cells werefound at P7. In contrast, an increasing number of intenselystained neurons were found after P7, with a peak at
ŽP14–P21 which was maintained through adulthood Fig..8E,F . This particular profile may be related to the opening
of the eyes which generally occurs at around P12–P14 inw xthe rat 11 . Age-dependent changes in HO-1 expression
Ž .were also observed in the rat substantia nigra Fig. 8G,H .Whereas there was little neuronal HO-1 staining at P7,adult rat brain showed moderate HO-1 expression mainly
Ž .in neurons of the pars compacta Snc , and in someŽ .neurons scattered throughout the pars reticulata Snr . In
the caudate-putamen, a few faintly stained cells wereŽ .found at all ages except in the adult data not shown . In
the globus pallidus, little or no staining was observed at P7but low to moderate HO-1 staining was detectable in some
Ž .neurons by P14 through adulthood data not shown .
4. Discussion
The present study demonstrates developmental regula-tion of HO-1 protein expression in rat brain. HO-1 proteinis generally and maximally expressed in glia-like cells,neurons and endothelial cells 7 days after birth, decreasingthereafter to very low levels in adulthood. Interestingly,the transcription factors NF-kB, Fos and CREB also show
Ž .highest activity during the late prenatal E15–E19 rearlyŽ . w xpostnatal days P1–P7 in the rat 2,5,13,48,49 . This
raises the possibility that the HO-1 gene, which has pro-
Ž . Ž .Fig. 7. Developmental changes of HO-1 expression in the rat nucleus basalis of the amygdala A–D and the ventromedial hypothalamic nucleus E–H .Ž . Ž . Ž . Ž . Ž .Coronal sections 50 mm were obtained from normal Sprague–Dawley rats at P7 A,E , P14 B,F , P21 C,G , and adult D,H . Note the age-related
Ž .decrease in the number and intensity of cell staining in the nucleus basalis of the amygdala A–D located in the medial portion of the temporal lobeŽ .adjacent to the tip of the external capsule Ec . In contrast, an age-dependent increase in the number, size and staining intensity of HO-1-positive neurons is
Ž .observed in the ventromedial hypothalamic nucleus E–H located next to the ventral portion of the third ventricle, near the median eminance. Arrows inŽ . Ž .E indicate the faint and scarce neuronal staining at P7. The scale bars for the amygdala photomicrographs A–D and the ventromedial hypothalamic
Ž . Ž . Ž .nucleus photomicrographs E–H represent 100 mm and are shown in D and E , respectively.

( )M. Bergeron et al.rDeÕelopmental Brain Research 105 1998 181–194190
Fig. 8.

( )M. Bergeron et al.rDeÕelopmental Brain Research 105 1998 181–194 191
moter elements for each of these transcription factors, isdevelopmentally regulated by these transcription factors.Our findings, however, are in contrast with a previous
w xdevelopmental study of HO-1 by Sun and colleagues 58 ,which demonstrated the lack of developmental regulationof HO-1 mRNA and undetectable protein by Western blotanalysis in whole rat brain microsomes. These discrepan-cies could be due to a dilution of HO-1 protein during thepreparation of samples from whole rat brain in the study
w xby Sun and colleagues 58 , compared with our prepara-tions from microdissected brain regions which expressHO-1 protein in high abundance at P7–P14. Consistentwith this explanation, a recent preliminary study from thelaboratory of Sun and colleagues reported a high level ofHO-1 protein expression in the CA1–CA3 regions of theneonatal hippocampus which decreased with maturationw x37 .
Since a specific inhibitor of HO-1 activity does notcurrently exist, it is difficult to determine the real signifi-cance of increased HO-1 protein expression in terms of itsfunctional activity and contribution to brain development.However, it is known that heme oxygenase activity re-quires the presence of heme substrate and the concertedactivity of NADPH:cytochrome P450 reductase and
w xbiliverdin reductase for bilirubin formation 35,37 . Inter-estingly, brain NADPH:cytochrome P450 reductase enzy-matic activity demonstrates a developmental profile similarto that of HO-1 protein expression. In particular, theactivity is highest at P7 decreasing gradually by P14 and
w xreaching a 2-fold reduction by early adulthood 23 . More-over, the expression of NADPH:cytochrome P450 reduc-tase protein in adult rat brain is widespread and includesthe same regions found to express HO-1 protein at the
w xdifferent ages 44 . Although quite prominent at earlyŽ .stages of brain development P1–P7 , biliverdin reductase
w xactivity increases even more at P28 and adulthood 20 .Accordingly, biliverdin reductase protein is detectable at
w xall ages 20 in many regions including several cell popula-tions that display HO-1 protein immunoreactivity. Takentogether, these observations suggest that the biochemicalmachinery necessary for functional and regional activity ofHO-1 is present at the different stages of normal braindevelopment.
In contrast to the ubiquitous brain HO-2 mRNA andprotein levels which are relatively lower in early develop-
w xment compared with the adult 58 , the highest level ofconstitutive HO-1 protein expression observed in the pre-sent study coincides with the early critical developmental
period in rat brain. This period is characterized by en-hanced dendritic outgrowth and vascular capillary prolifer-ation, formation of mitochondria, increased formation ofsynapses, myelination of axons, glutamate receptor appear-ance, active reconstruction and naturally occurring cell
w xdeath 7,16,39,50 . Interestingly, this period of transientelevation of HO-1 protein expression in newborn brainalso corresponds to the period of highest iron and heme
Župtake and highest brain iron concentration total and.non-heme which occurs during the first two postnatal
w xweeks 15,52,60 . Heme is a potent catalyst of lipid per-w xoxidation 6 . HO-1 activity produces bilirubin which has
been suggested as a potent antioxidant and inhibitor ofw xlipid peroxidation 57 . Because the immature blood–brain
barrier is permeable to systemic heme, increased HO-1could play a neuroprotective role during this postnatalperiod of high heme turnover. This is consistent with theobserved elevated level of HO-1 protein in the vascularendothelium and the periventricular cells in newborn brainconsidering that these are gateways to the central nervoussystem.
At P7, the cell morphology and distribution profilew xdescribed previously for iron-containing cells 15 is simi-
lar to that reported here for some populations of intenselystained HO-1-positive cells. In particular, most iron-con-taining cells are predominantly non-process bearing with
Žthe highest concentration found in the white matter mainly.in the areas of myelinogenesis like the cingulum , in layer
IV of somatosensory cortex and in the endothelium. Cur-rent knowledge suggests that perinatal iron flux may beassociated with macrophage invasion through the leaky
w ximmature blood–brain barrier 32,34 . Interestingly, ourdouble immunofluorescence studies have shown intenseHO-1 protein expression in ED-1-positive macrophages atP7 in the cingulum and throughout the corpus callosum,periventricular zones and adjacent areas of cortex and
w x Žhippocampus 8 . Although an essential nutrient, iron if.not sequestered properly can also become a critical factor
in the production of free hydroxyl radicals which in turncan initiate lipid peroxidation and other oxidative damagew x12 . Almost two-thirds of the iron in the brain is stored as
w xferritin 14 . However, in newborn brain the level ofw xferritin is relatively lower than in adult brain 52 . Since
HO-1 activity produces bilirubin which has potent antioxi-w xdant properties 57 , the higher levels of potentially lethal
free brain iron in the early stages of brain developmentcould also contribute, at least in part, to the higher level ofconstitutive HO-1 protein expression throughout the brain
Ž . Ž . Ž .Fig. 8. Effect of maturation on HO-1 expression in the rat ventromedial thalamic nucleus A,B , internal capsule C,D , magnocellular preoptic area E,FŽ . Ž . Ž . Ž .and substantia nigra G,H . Comparable coronal sections 50 mm were obtained from P7 newborn A,C,E,G and adult B,D,F,H Sprague–Dawley rats.
Ž .Taking into account the high background stain at P7 A , note that the cellular HO-1 staining intensity and relative density is only mildly influenced byŽ . Ž . Ž .maturation in the ventromedial thalamic nucleus A,B located adjacent to the zona incerta Zi and mammillothalamic tract Mt . In the internal capsule,
Ž . Ž .intense staining of microglia-like cells is observed at P7 C but not in adults D . In the preoptic area, note the increased number of intensely stainedŽ . Ž .neurons in the adult F compared to the faint and scarce neuronal staining at P7 arrows in E . In the substantia nigra, whereas P7 is associated with a high
Ž . Ž . Ž .background stain but little HO-1 cellular staining G , adult tissue H shows moderate HO-1 expression mostly in neurons of the pars compacta Snc ,Ž .with few scattered neurons in the pars reticulata Snr; arrows in H . Scale bar: 100 mm.

( )M. Bergeron et al.rDeÕelopmental Brain Research 105 1998 181–194192
in normal P7 animals compared with older animals. There-fore, the presence of high levels of free heme and free iron
Žduring early development when most antioxidant defense.mechanisms are not entirely developed could influence
the vulnerability of the newborn brain to oxidative stressand perhaps injury caused by bilirubin.
Hyperbilirubinemia is commonly observed during thew xfirst week of life in humans and rats 35 . Although
w xbilirubin may act as an antioxidant 57 , it may also bew xtoxic to the brain 25 . This dual effect of bilirubin depends
on the type of tissue, the local concentration and the lengthof exposure. Bilirubin can disrupt a number of metabolic
w xfunctions including glycolysis 31 , oxidative phosphoryla-w x w xtion 4 , cerebral glucose utilization 51 , synthesis of
w x w xneurotransmitters 46 , protein phosphorylation 26 , DNAand protein synthesis as well as NaqKq-ATPase activityw x4 . Interestingly, bilirubin toxicity decreases significantlywith cell differentiation and maturation as seen in primary
w x w xastrocyte cultures 3 and neural cell lines 45 . Theseobservations support the idea that the immature brain ismore sensitive to bilirubin toxicity than the older brain.Thus, the elevated levels of bilirubin produced by theincreases in HO-1 protein expression during the earlycritical growth period of the rat brain could contribute toincrease local bilirubin concentration and therefore predi-pose the immature brain to the toxic effects of hyperbiliru-binemia. Because the brain maturity of a rat at P7 isroughly comparable to that of a late-term gestation human
w xfetus or newborn infant 16 , this last suggestion may alsoapply to humans.
In addition to the production of bilirubin, HO-1 activityalso generates CO which has been proposed as a putativeneurotransmitter acting as a physiologic regulator ofguanylyl cyclase and cGMP-dependant protein kinasew x36,37,62 . The role of CO in brain development has notyet been elucidated. Recent studies have shown that neu-ronal HO activity could be modulated by the activation ofmetabotropic glutamate receptors and that CO could beinvolved in the signal transduction pathway coupling thesereceptors to the activity of the NaqKq-ATPase pumpw x24,42 . The critical growth period in newborn rat brain isassociated with increased glutamate receptor sensitivityw x29,39 . Additionally, the stimulation of ionotropic gluta-mate receptors in neuronal cultures induced the transcrip-
w x w xtion factors NF-kB 30 and c-fos 28,55,59 . Since COproduction may modulate neuronal excitability, synaptic
w xplasticity, long-term potentiation 27,42,56,64 and learn-w xing 9,10 , region-specific changes in CO production by
HO-1 could modulate local synaptic activity and glial andneuronal maturation during early brain development.
Acknowledgements
The present study was funded by National Institutes ofHealth Grants NS28167 and NS14543 to F.R.S. and
P20NS32553 to D.M.F. M.B. was the recipient of a post-doctoral fellowship from The Medical Research Council ofCanada.
References
w x1 J. Alam, D. Zhining, Distal AP-1 binding sites mediate basal levelenhancement and TPA induction of the mouse heme oxygenase-1
Ž .gene, J. Biol. Chem. 267 1992 21894–21900.w x2 A.A. Alcantara, W.T. Greenough, Developmental regulation of Fos
and Fos-related antigens in cerebral cortex, striatum, hippocampusŽ .and cerebellum of the rat, J. Comp. Neurol. 334 1993 75–85.
w x3 Y. Amit, T. Brenner, Age-dependent sensitivity of cultured rat glialŽ .cells to bilirubin toxicity, Exp. Neurol. 121 1993 248–255.
w x4 Y. Amit, G. Chan, S. Fedunec, M.J. Poznansky, D. Schiff, Bilirubintoxicity in a neuroblastoma cell line N-115. I. Effects on Naq Kq-
w3 x w35 xATPase, H thymidine uptake, L- S methionine incorporation, andŽ .mitochondrial function, Pediatr. Res. 25 1989 364–368.
w x5 G.Y. Bakalkin, T. Yakovleva, L. Terenius, NF-kB-like factors in themurine brain. Developmentally-regulated and tissue-specific expres-
Ž .sion, Mol. Brain Res. 20 1993 137–146.w x6 J. Balla, H.S. Jacob, G. Balla, Endothelial cell-heme uptake from
heme proteins: induction of sensitization and desensitization toŽ .oxidant damage, Proc. Natl. Acad. Sci. USA 90 1993 9285–9289.
w x7 S.A. Bayer, J. Altman, Neurogenesis and neuronal migration. in: G.Ž .Paxinos Ed. , The Rat Nervous System, 2nd edn., Academic Press,
Sydney, 1995, pp. 1041–1078.w x8 M. Bergeron, D.M. Ferriero, H.J. Vreman, D.K. Stevenson, F.R.
Sharp, Hypoxia-ischemia, but not hypoxia alone, induces the expres-Ž .sion of heme oxygenase-1 HSP32 in newborn rat brain, J. Cerebr.
Ž .Blood Flow Metab. 17 1997 647–658.w x9 R. Bernebu, F. Princ, M.L. de Stein, C. Fin, A.A. Juknat, A. Batile,
I. Izquierdo, J.H. Medina, Evidence for the involvement of hip-pocampal CO production in the acquisition and consolidation of
Ž .inhibitory avoidance learning, NeuroReport 6 1995 516–518.w x10 O. Bing, L. Grundemar, L. Ny, C. Moller, M. Heilig, Modulation of
carbon monoxide production and enhanced spatial learning by tinŽ .protoporphyrin, NeuroReport 6 1995 1369–1372.
w x11 K.E. Binns, T.E. Salt, Post eye-opening maturation of visual recep-tive field diameters in the superior colliculus of normal- and dark-re-
Ž .ared rats, Dev. Brain Res. 99 1997 263–266.w x12 J.M. Braughler, L.A. Duncan, R.L. Chase, The involvement of iron
in lipid peroxidation. Importance of ferric to ferrous ratios inŽ .initiation, J. Biol. Chem. 261 1986 10282–10289.
w x13 K. Cauley, I.M. Verma, kB enhancer-binding complexes that do notcontain NF-kB are developmentally regulated in mammalian brain,
Ž .Proc. Natl. Acad. Sci. USA 91 1994 390–394.w x14 J.R. Connor, K.L. Boeshore, S.A. Benkovic, S.L. Menzies, Isoforms
of ferritin have a specific cellular distribution in the brain, J.Ž .Neurosci. Res. 37 1994 461–465.
w x15 J.R. Connor, G. Palvick, D. Karli, S.L. Menzies, C. Palmer, Ahistochemical study of iron-positive cells in the developing rat brain,
Ž .J. Comp. Neurol. 355 1995 111–123.w x16 J. Dobbing, J. Sands, Comparative aspects of the brain growth spurt,
Ž .Early Hum. Dev. 3 1979 79–83.w x17 J.F. Ewing, S.N. Haber, M.D. Maines, Normal and heat-induced
Ž .patterns of expression of heme oxygenase-1 HSP32 in rat brain:hyperthermia causes rapid induction of mRNA and protein, J. Neu-
Ž .rochem. 58 1992 1140–1149.w x18 J.F. Ewing, M.D. Maines, In situ hybridization and immunohisto-
chemical localization of heme oxygenase-2 mRNA and protein innormal rat brain: differential distribution of isozyme 1 and 2, Mol.
Ž .Cell. Neurosci. 3 1992 559–570.

( )M. Bergeron et al.rDeÕelopmental Brain Research 105 1998 181–194 193
w x19 J.F. Ewing, M.D. Maines, Glutathione depletion induces heme oxy-Ž .genase-1 HSP32 mRNA and protein in rat brain, J. Neurochem. 60
Ž .1993 1512–1519.w x20 J.F. Ewing, M.D. Maines, Immunohistochemical localization of
biliverdin reductase in rat brain: age related expression of proteinŽ .and transcript, Brain Res. 672 1995 29–41.
w x21 D.M. Ferriero, H.Q. Soberano, R.P. Simon, F.R. Sharp, Hypoxia-Ž .ischemia induces heat shock protein-like HSP72 immunoreactivityŽ .in neonatal rat brain, Dev. Brain Res. 52 1990 145–150.
w x22 K. Fukuda, J.D. Richmon, M. Sato, F.R. Sharp, S.S. Panter, L.J.Ž .Noble, Induction of heme oxygenase-1 HO-1 in glia after traumatic
Ž .brain injury, Brain Res. 736 1996 68–75.w x23 J.F. Ghersi-Egea, A. Minn, J.L. Daval, Z. Jayyosi, V. Arnould, H.
Ž .Souhaili-El Amri, G. Siest, NADPH:cytochrome P-450 c reductase:biochemical and characterization in rat brain and cultured neuronsand evolution of activity during development, Neurochem. Res. 14Ž .1989 883–888.
w x24 S.R. Glaum, R.J. Miller, Zinc protoporphyrin-IX blocks the effectsof metabotropic glutamate receptor activation in the rat nucleus
Ž .tractus solitaris, Mol. Pharmacol. 43 1993 965–969.w x Ž .25 T.W.R. Hansen, Bilirubin in the brain, Clin. Pediatr. 33 1994
452–459.w x26 T.W.R. Hansen, D. Bratlid, S.I. Walaas, Bilirubin decreases phos-
phorylation of synapsin I, a synaptic vesicle-associated neuronalphosphoprotein, in intact synaptosomes from rat cerebral cortex,
Ž .Pediatr. Res. 23 1988 219–223.w x27 R.D. Hawkins, M. Zhuo, O. Arancio, Nitric oxide and carbon
monoxide as possible retrograde messengers in hippocampal long-Ž .term potentiation, J. Neurobiol. 25 1994 652–665.
w x28 K. Hisanaga, S.M. Sagar, K.J. Hicks, F.R. Sharp, NMDA antago-nists block Fos-like nuclear protein expression induced by multiplesignalling pathways in cultured cortical neurons, J. Neurochem. 58Ž .1992 1836–1844.
w x29 C. Ikonomidou, J.L. Mosinger, K. Shahid Salles, J. Labryere, J.W.Olney, Sensitivity of the developing rat brain to hypobaricrischemicdamage parallels sensitivity to N-methyl-aspartate toxicity, J. Neu-
Ž .rosci. 9 1989 2809–2818.w x30 C. Kaltschmidt, B. Kaltschmidt, P.A. Baeuerle, Stimulation of
ionotropic glutamate receptors activates transcription factor NF-kBŽ .in primary neurons, Proc. Natl. Acad. Sci. USA 92 1995 9618–
9622.w x31 R. Katoh-Semba, Studies on cellular toxicity of bilirubin: effect on
Ž .brain glycolysis in the young rat, Brain Res. 113 1976 339–348.w x32 C. Kaur, E.A. Ling, Transient expression of transferrin receptors and
localization of iron in amoeboid microglia in postnatal rats, J. Anat.Ž .186 1995 165–173.
w x33 Y. Lavrovsky, M.L. Schwartzman, R.D. Levere, A. Kappas, N.G.Abraham, Identification of binding sites transcription factors NF-kBand AP-2 in the promoter region of the human heme oxygenase 1
Ž .gene, Proc. Natl. Acad. Sci. USA 91 1994 5987–5991.w x34 E.A. Ling, W.C. Wong, The origin and nature of ramified and
amoeboid microglia: a historical review and current concepts, Glia 7Ž .1993 9–18.
w x35 M.D., Maines, Heme Oxygenase: Clinical Applications and Func-tions, CRC Press, Boca Raton, FL, 1992, 276 pp.
w x36 M.D. Maines, Carbon monoxide: an emerging regulator of cGMP inŽ .the brain, Mol. Cell. Neurosci. 4 1993 389–397.
w x37 M.D. Maines, The heme oxygenase system: a regulator of secondŽ .messenger gases, Ann. Rev. Pharmacol. Toxicol. 37 1997 517–554.
w x38 M.D. Maines, G.M. Trakshel, R.K. Kutty, Characterization of twoconstitutive forms of rat liver microsomal heme oxygenase, J. Biol.
Ž .Chem. 261 1986 411–419.w x39 J.W. McDonald, M.V. Johnston, Physiological and pathophysiologi-
cal roles of excitatory amino acids during central nervous develop-Ž .ment, Brain Res. Rev. 15 1990 41–70.
w x40 C.E. Milligan, T.J. Cunningham, P. Levitt, Differential immuno-chemical markers reveal the normal distribution of brain macrophages
and microglia in the developing rat brain, J. Comp. Neurol. 314Ž .1991 125–135.
w x41 R.M. Muller, H. Taguchi, S. Shibahara, Nucleotide sequence and¨organization of the rat heme oxygenase gene, J. Biol. Chem. 262Ž .1987 6795–6802.
w x42 J.A. Nathanson, C. Scavone, C. Scanlon, M. McKee, The cellularNaq pump as a site of action for carbon monoxide and glutamate: amechanism for long-term modulation of cellular activity, Neuron 14Ž .1995 781–794.
w x43 T. Nimura, P.R. Weinstein, S.M. Massa, S. Panter, F.R. Sharp,Ž .Heme oxygenase-1 HO-1 protein induction in rat brain following
Ž .focal ischemia, Mol. Brain Res. 37 1996 201–208.w x44 P.J. Norris, J.P. Hardwick, P.C. Emson, Localization of NADPH
cytochrome P450 oxidoreductase in rat brain by immunohistochem-istry and in situ hybridization and a comparison with the distribution
Ž .of neuronal NADPH-diaphorase staining, Neuroscience 61 1994331–350.
w x45 M.F.D. Notter, J.W. Kendig, Differential sensitivity of neural cellsŽ .to bilirubin toxicity, Exp. Neurol. 94 1986 670–682.
w x46 T. Ohno, Kernicterus: effect on choline acetyletransferase, glutamicacid decarboxylase and tyrosine hydroxylase activities in the brain
Ž .of Gunn rat, Brain Res. 196 1980 282–285.w x47 S. Okinaga, S. Shibahara, Identification of a nuclear protein that
constitutively recognizes the sequence containing a heat-shock ele-ment. Its binding properties and possible function modulating heat-shock induction of the rat heme oxygenase gene, Eur. J. Biochem.
Ž .212 1993 167–175.w x48 K.R. Pennypacker, D. Dreyer, J.S. Hong, M.K. McMillian, Elevated
basal AP-1 DNA binding activity in developing rat brain, Mol. BrainŽ .Res. 19 1993 349–352.
w x49 K.R. Pennypacker, P.M. Hudson, J.S. Hong, M.K. McMillian, DNAbinding activity of CREB transcription factors during ontogeny of
Ž .the central nervous system, Dev. Brain Res. 86 1995 242–249.w x50 P.L. Robertson, M. Du Bois, P.D. Bowman, G.W. Goldstein, Angio-
genesis in the developing rat brain: an in vivo and in vitro study,Ž .Dev. Brain Res. 23 1985 219–223.
w x51 C. Roger, V. Koziel, P. Vert, A. Nehlig, Regional cerebral metabolicconsequences of bilirubin in rat depend upon post-gestational age at
Ž .the time of hyperbilirubinemia, Dev. Brain Res. 87 1995 194–202.w x52 A.J.I. Roskams, J.R. Connor, Iron, transferrin, and ferritin in the rat
Ž .brain during development and aging, J. Neurochem. 63 1994709–716.
w x53 S. Shibahara, M. Yoshizawa, H. Suzuki, K. Takeda, K. Meguro, K.Endo, Functional analysis of cDNA for two types of human hemeoxygenase and evidence for their separate regulation, J. Biochem.
Ž .113 1993 214–218.w x54 T. Shinomura, S.-I. Nakao, K. Mori, Reduction of depolarization-in-
duced glutamate release by heme oxygenase inhibitor: possible roleof carbon monoxide in synaptic transmission, Neurosci. Lett. 166Ž .1994 131–134.
w x55 J.L. Sonnenberg, C. Mitchelmore, P.F. Macgregor-Leon, J. Hemp-stead, J.I. Morgan, T. Curran, Glutamate receptor agonists increasethe expression of Fos, Fra, and AP-1 DNA binding activity in the
Ž .mammalian brain, J. Neurosci. Res. 24 1989 72–80.w x56 C.F. Stevens, Y. Wang, Reversal of long-term potentia tion by
Ž .inhibitors of haem oxygenase, Nature 364 1993 147–149.w x57 R. Stocker, Y. Yamamoto, A.F. McDonagh, A.N. Glazer, B.N.
Ames, Bilirubin is an antioxidant of possible physiological impor-Ž .tance, Science 235 1987 1043–1046.
w x58 Y. Sun, M.O. Rotenberg, M.D. Maines, Developmental expressionof heme oxygenase isozymes in rat brain. Two HO-2 mRNAs are
Ž .detected, J. Biol. Chem. 265 1990 8212–8217.w x59 A.M. Szekely, M.L. Barbaccia, E. Costa, Activation of specific
glutamate receptor subtypes increases c-fos proto-oncogene expres-sion in primary cultures of neonatal rat cerebellar granule cells,
Ž .Neuropharmacology 26 1987 1779–1782.w x60 E.M. Taylor, E.H. Morgan, Developmental changes in transferrin

( )M. Bergeron et al.rDeÕelopmental Brain Research 105 1998 181–194194
Ž .and iron uptake by the brain in the rat, Dev. Brain Res. 55 199035–42.
w x61 C.P. Turner, M. Bergeron, P. Matz, L. Noble, S.S. Panter, F.R.Ž .Sharp, Heme Oxygenase-1 HO-1, HSP32 is induced in microglia
throughout brain by subarachnoid injection of hemoglobin, J. Cerebr.Ž .Blood Flow Metab., 1997 in press.
w x62 A. Verma, D.J. Hirsch, C.E. Glatt, G.V. Ronnett, S.H. Snyder,Ž .Carbon monoxide: putative neural messenger, Science 259 1993
381–384.
w x63 S.R. Vincent, S. Das, M.D. Maines, Brain heme oxygenase isoen-zymes and nitric oxide synthase are co-localized in select neurons,
Ž .Neuroscience 63 1994 223–231.w x64 M. Zhou, S.A. Small, E.R. Kandel, R.D. Hawkins, Nitric oxide and
carbon monoxide produce activity-dependent long-term synaptic en-Ž .hancement in hippocampus, Science 260 1993 1946–1950.