an internal enhancer regulates heme and cadmium-mediated induction of human heme oxygenase-1
TRANSCRIPT

F-00137-2003
1
An internal enhancer regulates heme and cadmium-mediated induction ofhuman heme oxygenase-1
Nathalie Hill-Kapturczak, PhD*, Eric Sikorski, PhD*, Christy Voakes*, Jairo Garcia*,Harry S. Nick, PhD#, Anupam Agarwal, MD*
*Department of Medicine, Division of Nephrology, Hypertension and Transplantation#Department of Neuroscience, University of Florida, Gainesville, FL 32610
Running title:
Human heme oxygenase-1 gene regulation
Address for Correspondence:
Anupam Agarwal, MDDivision of Nephrology, Hypertension and TransplantationBox 100224 JHMHC1600 SW Archer RoadUniversity of FloridaGainesville FL 32610Tel: (352) 846 2367Fax: (352) 392 3581Email: [email protected]
Copyright (c) 2003 by the American Physiological Society.
Articles in PresS. Am J Physiol Renal Physiol (June 3, 2003). 10.1152/ajprenal.00137.2003

F-00137-2003
2
ABSTRACT
Heme oxygenase-1 (HO-1) catalyzes the rate-limiting step in heme degradation releasing
iron, carbon monoxide and biliverdin. Induction of HO-1 is an adaptive and beneficial response
in renal and non-renal settings of tissue injury. The purpose of this study was to characterize the
regulation of the human HO-1 gene in renal proximal tubule and aortic endothelial cells in
response to heme and cadmium. Evaluation of multiple human HO-1 promoter-reporter
constructs up to –9.1kb demonstrated only a partial response to heme and cadmium. In an effort
to mimic endogenous stimulus-dependent levels of HO-1 induction, we evaluated the entire 12.5
kb of the human HO-1 gene, including introns and exons, in conjunction with a –4.5 kb human
HO-1 promoter and observed significant heme and cadmium-mediated induction of the reporter
gene, suggesting the presence of an internal enhancer. Enhancer function was orientation
independent and required a region between –3.5 and –4.5kb of the human HO-1 promoter. Our
studies have identified a novel enhancer internal to the human HO-1 gene that, in conjunction
with the HO-1 promoter, recapitulates heme and cadmium-mediated induction of the endogenous
HO-1 gene. Elucidation of the molecular regulation of the human HO-1 gene will allow for the
development of therapeutic strategies to manipulate HO-1 gene expression in pathologic states.
Key words: gene transcription, renal proximal tubule cells, endothelial cells, heme proteins,
molecular regulation

F-00137-2003
3
INTRODUCTION
The cellular content of heme (ferriprotoporphyrin IX), derived from heme-containing
proteins such as hemoglobin, myoglobin, cytochromes, and enzymes like nitric oxide synthase,
catalase, peroxidases, respiratory burst oxidase and pyrrolases, requires a balance between heme
synthesis and degradation (31). In pathological states, destabilization of heme proteins leads to
liberation of free heme which has potential pro-oxidant effects (22). Increases in renal heme
content are observed in rhabdomyolysis, ischemia-reperfusion injury and nephrotoxin-induced
acute renal failure (2, 29, 33, 40). Heme damages multiple cellular targets including lipid
bilayers, mitochondria, cytoskeleton, nuclei and several intracellular enzymes (33). While
multiple enzymes are involved in heme synthesis, the major biochemical pathway for heme
detoxification is via the heme oxygenase enzyme system (28).
Heme oxygenase (HO) opens the heme ring producing equimolar quantities of biliverdin,
iron and carbon monoxide (CO). Biliverdin is subsequently converted to bilirubin by biliverdin
reductase. Recent studies have highlighted the important biological effects of the HO reaction
product(s) which possess antioxidant, anti-inflammatory and anti-apoptotic functions (reviewed
in 21). Two isoforms of HO have been characterized; an inducible enzyme, HO-1, and a
constitutive isoform, HO-2 (28). A putative isozyme, HO-3, isolated from rat brain and sharing
about 90% homology with HO-2, has also been described (30). HO-1 and HO-2 are products of
different genes and share ~40% amino acid homology (28). We and others have demonstrated
that induction of HO-1 by chemical inducers (2, 32), or selective overexpression (1, 41) is
cytoprotective both in vitro and in vivo; findings that have been further substantiated by studies
in HO-1 knock-out mice and a patient with HO-1 deficiency (34, 41, 48).

F-00137-2003
4
The mechanisms underlying HO-1 induction are complex and tightly regulated at the
transcriptional level. Adding to the complexity, regulation of the HO-1 gene is species and cell
specific. The promoter of the rat HO-1 gene, for instance, has a heat shock responsive element
that is not functional in the human HO-1 gene (50). A GT repeat region identified at position
–256/-196 in the human HO-1 promoter is absent in the mouse HO-1 gene (49). Interestingly,
length polymorphisms of this GT repeat correlate with disease development in atherosclerosis
(16), emphysema (49), and vascular restenosis (18) in humans.
The human HO-1 gene, located on chromosome 22q12 (26), has 5 exons and spans
approximately 14kb. A potential cadmium response element (TGCTAGATTT) has been
identified between –4.5 and –4.0kb of the 5’ flanking region (44) of the human HO-1 gene,
however the cadmium response element in the mouse HO-1 gene is located downstream of this
sequence (13). A 3-fold increase in luciferase activity was reported with a –4.5kb human HO-1
construct in response to cadmium, with only a minimal response to hemin, sodium arsenite or
cobalt protoporphyrin in HeLa cells (44). We have previously shown that the identical 4.5kb
human HO-1 gene promoter fragment responds, in part, to both heme and cadmium with a 2-3
fold increase in reporter gene activity compared to ~20-30 fold increase in endogenous HO-1
mRNA in human aortic endothelial and renal proximal tubular cells (4). Furthermore, this 4.5kb
promoter fragment does not respond to other stimuli such as 13-hydroxyperoxyoctadecadienoic
acid (13-HPODE), hydrogen peroxide (H2O2) and hyperoxia that directly increase de novo HO-1
gene transcription (4, 19). We report here the identification of an enhancer region internal to the
human HO-1 gene that, together with the 4.5kb promoter, recapitulates levels of heme and
cadmium induction observed using steady state northern analysis of the endogenous gene.

F-00137-2003
5
MATERIALS AND METHODS
Reagents: Hemin (ferriprotoporphyrin IX chloride), cadmium chloride (CdCl2), linoleic acid,
lipoxidase, H2O2, and diethylaminoethyl (DEAE)-dextran were purchased from Sigma Chemical
Co. (St. Louis, MO). Transforming growth factor β1 (TGF-β1) was obtained from R & D
systems (Minneapolis, MN). RNeasy mini and Maxi-prep kits were obtained from Qiagen
(Valencia, CA).
Cell Culture: Human renal proximal tubule cells (HPTC)(Clonetics, Walkersville, MD) were
grown in renal epithelial basal medium supplemented with 5% fetal bovine serum (FBS),
gentamicin (50µg/ml), amphotericin B (50µg/ml), insulin (5µg/ml), transferrin (10µg/ml),
triiodothyronine (6.5ng/ml), hydrocortisone (0.5µg/ml), epinephrine (0.5µg/ml), and human
epidermal growth factor (10ng/ml). These cells are positive for γ-glutamyltranspeptidase, an
enzyme marker for proximal tubule cells and the presence of microvilli, abundant mitochondria,
lysosomes and endocytotic vacuoles, morphological features of proximal tubule cells, were
confirmed by electron microscopy. Human aortic endothelial cells (HAEC) [derived from
segments of human aorta obtained from heart transplant donors and previously characterized by
positive staining for Factor VIII-related antigen and acetylated LDL (46)] were grown in
endothelial basal medium (Clonetics), supplemented with 10% FBS, gentamicin (50µg/ml),
amphotericin B (50µg/ml), hydrocortisone (1µg/ml), human epidermal growth factor (10ng/ml),
and bovine brain extract (6µg/ml) at 37°C in 95% air and 5% CO2. Studies were performed over
a range of no more than 6-8 passages. 16h prior to treatment with HO-1 inducers, growth media
was changed to a 0.5 and 1% FBS-containing media for HPTC and HAEC, respectively.
Preparation of HO-1 inducers: Hemin, CdCl2 and 13-HPODE were prepared fresh, daily. A
stock solution of hemin (1mM) was prepared using 10mM NaOH. CdCl2 (5mM stock solution)

F-00137-2003
6
and H202 (10mM stock) were prepared in water and sterile filtered. 13-HPODE was prepared as
previously described (4). Lyophilized TGF-β1 was dissolved in 4mM HCl containing 0.1%
bovine serum albumin to obtain a 2ng/µl stock solution, aliquoted and stored at –80˚C.
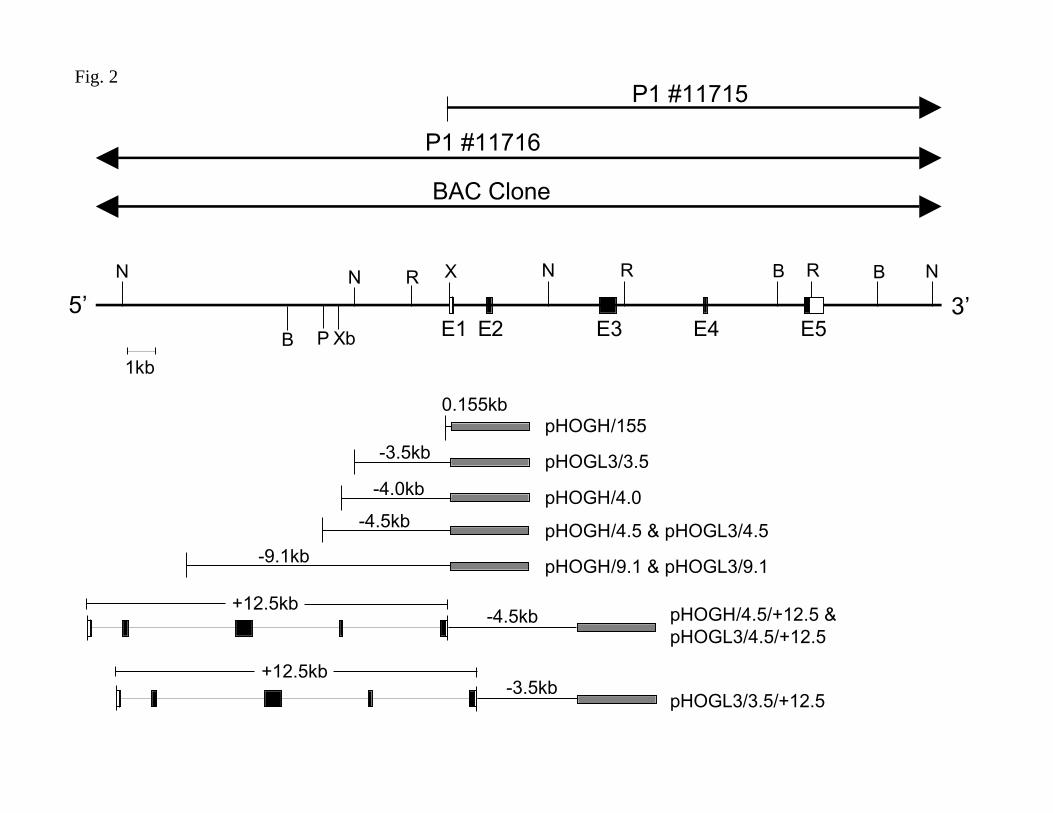
Cloning and characterization of a human HO-1 genomic clone: A 4.5kb fragment of the 5'
flanking region of the human HO-1 gene, including the transcription initiation site, was
generated by long range PCR (Perkin-Elmer, Foster City, CA) using human genomic DNA as a
template, as described previously (4). In order to obtain additional genomic sequence for the
human HO-1 gene, two single copy probes were designed, one from the –4.5/-4.0kb region and
one from an internal exon, and a commercially available human P1 bacteriophage library
(Genome systems, Inc. St. Louis, MO) was screened. Two P1 clones (P1 11715 and 11716) were
identified by both probes. These clones were characterized by restriction map and sequence
analysis and the presence of the entire human HO-1 gene, as well as >15kb of the 5’ flanking
region was verified in P1 11716. More recently, the efforts of the human genome project resulted
in the sequencing of a bacterial artificial chromosome (BAC) clone, containing portions of
chromosome 22 (Accession no. Z82244). Using this BAC clone, sequences of the human HO-1
gene promoter fragments were confirmed and larger fragments, further upstream, were generated
by long-range PCR and restriction digestion (Fig. 2).
Plasmid Construction: 5’ flanking promoter fragments of the human HO-1 gene, extending from
–4.0kb, –4.5kb and –9.1kb to +80bp position were generated using oligonucleotide primers
(Table 1) specific to the HO-1 gene by long range PCR (Perkin-Elmer, Foster City, CA). The 4.0
and 4.5kb PCR products were ligated into TA cloning vectors (Invitrogen Corp., Carlsbad, CA)
and the 9.1kb PCR product was cloned into pCR-XL-TOPO (Invitrogen). BamHI sites were
incorporated in the primers for the 4.0 and 4.5kb fragments and SalI sites were incorporated in

F-00137-2003
7
the primers for the 9.1kb fragment to enable subcloning into a promoterless human growth
hormone (hGH) reporter gene, generating pHOGH/4.0, pHOGH/4.5, and pHOGH/9.1. A 155bp
HO-1 promoter hGH plasmid was generated by deletion of a 4.345 kb HO-1 promoter fragment
using NdeI followed by religation resulting in the formation of pHOGH/155. The 4.5kb promoter
fragment was also cloned into the BglII site in a luciferase reporter (pGL3) generating
pHOGL3/4.5. The 9.1kb promoter fragment (with SalI sites on each end) was cloned into the
XhoI site of pGL3, generating pHOGL3/9.1. A 3.5kb HO-1 promoter reporter vector was
generated by cutting the pHOGL3/9.1 with NheI, which cuts HO-1 at ~3.5kb 5’ from the start
site, and cuts pGL3 once 5’ from the XhoI site, and religating to generate pHOGL3/3.5.
The entire +1 to +12.5kb region of the human HO-1 gene (including all exons and
introns) was generated using long range PCR with oligonucleotide primers specific to the HO-1
gene (Table 1). The 12.5kb PCR product was cloned into pCR-XL-TOPO and then subcloned
into pHOGH/4.5, pHOGL3/4.5 or pHOGL3/3.5 generating pHOGH/4.5/+12.5,
pHOGL3/4.5/+12.5 and pHOGL3/3.5/+12.5. SalI sites were included at each end of the 12.5kb
fragment to enable cloning into the SalI site of the hGH and luciferase vectors. We obtained both
5’-3’ and 3’-5’ orientations of the +12.5kb fragment in the hGH vector containing the –4.5kb
HO-1 promoter region [pHOGH/4.5/+12.5 and pHOGH/4.5/+12.5(3’-5’), respectively]. The
+12.5kb fragment was also cloned into the HindIII site of an hGH vector containing a
heterologous herpes-virus thymidine kinase (TK) minimal promoter to generate
pHOTKGH/+12.5. All constructs were verified for orientation by sequencing and restriction
analysis.
Transfection of the Reporter Gene: HPTC and HAEC were transiently transfected with the
reporter gene vectors by the DEAE-dextran method using a batch transfection protocol (4). The

F-00137-2003
8
DEAE-dextran method was optimized to achieve reproducible transfection efficiency (~20-25%)
in HPTC and HAEC as assessed by co-transfection with pcDNA3.1/Lac-z (Invitrogen). The
transfection protocol for HPTC was as follows: 60-70% confluent cells in 150 mm tissue culture
plates were washed once with HBSS and once with Tris-buffered saline solution (TBS). A 1.0ml
solution containing equimolar amounts of the plasmid(s) (normalized to 8.1µg of pHOGH/4.5)
and DEAE-dextran (10 mg/ml) was added per plate and rocked for 1h at room temperature. The
cells were shocked for 1 minute at room temperature with 10% DMSO in TBS, and then washed
once with HBSS. The cells were incubated in complete media (18ml/plate) containing
chloroquine diphosphate (100µM), to inhibit lysosomal degradation of the DNA. After 4h
incubation at 37˚C in 5% CO2/room air, the cells were washed twice with HBSS and incubated in
normal growth media overnight.
The transfection protocol for HAEC was as follows: 60-70% confluent cells in 150 mm
tissue culture plates were washed once with PBS. Complete media containing 10% NuSerum
(BD Biosciences, San Jose, CA) was added to the plates. The transfection solution, 1.5ml
DEAE-dextran solution (10mg/ml) containing equimolar amounts of the plasmid(s) (normalized
to 8.1µg of pHOGH/4.5), was added to each dish dropwise and then gently swirled to ensure that
the DNA/DEAE-dextran was uniformly applied to the cells. Chloroquine diphosphate (100µM)
was then added to the medium. Cells were incubated for 4h at 37˚C in 5% CO2/room air and then
shocked for 1 minute at room temperature with 10% DMSO in PBS, washed twice with PBS and
incubated in growth media containing 10% FBS overnight.
24h post-transfection, the cells (both cell types) were passaged from 150mm dishes into
several 100mm dishes (for hGH assays) or 12-well trays (for luciferase assays) in order to insure
equal transfection efficiency between experimental treatments. Co-transfection with

F-00137-2003
9
pcDNA3.1/Lac-z demonstrated equal amounts of DNA in each of the wells. After the cells were
allowed to recover for 24h, they were treated with stimulus or vehicle (control). 16-72h
following stimulus exposure, cells were evaluated for hGH mRNA content by northern analysis,
levels of secreted hGH protein in the culture media, as described previously (4), and luciferase
activity was monitored using the luciferase reporter assay system (Promega) according to the
manufacturer’s instructions. Luminescence was measured with a Sirius Luminometer (Berthold
Detection Systems, Pforzheim, Germany).
RNA Isolation, Molecular Probes, and Northern Analysis: Total cellular RNA was isolated by
the method of Chomczynski and Sacchi (17). Samples were electrophoresed on a 1% agarose
formaldehyde gel and electrotransferred to a charged nylon hybridization membrane. The cDNA
probes for hGH, human HO-1 and human GAPDH were radiolabelled with [α-32P]dATP using a
random primer labeling kit, according to the instructions of the manufacturer (Invitrogen) and
purified over a G-50 column (Amersham Pharmacia Biotech, Piscataway, NJ). Membranes were
hybridized overnight at 60˚C, washed in a high stringency buffer (0.04M sodium phosphate,
1mM EDTA, 1% SDS) at 65˚C and subjected to autoradiography. To quantitate expression
levels, autoradiographs were scanned on a Hewlett-Packard ScanJet 4C using DeskScan II
software and densitometry was performed using NIH Image 1.63 software. Experiments were
internally controlled by hybridization with GAPDH, then normalized and expressed as a
percentage of maximal expression. All experiments were repeated with at least 2-3 independent
RNA preparations to show reproducibility.
Statistical Analysis: Data are expressed as mean ± SEM. Statistical analyses were performed
using ANOVA and the Student-Newman-Keuls test. All results are considered significant at p <
0.05.

F-00137-2003
10
RESULTS
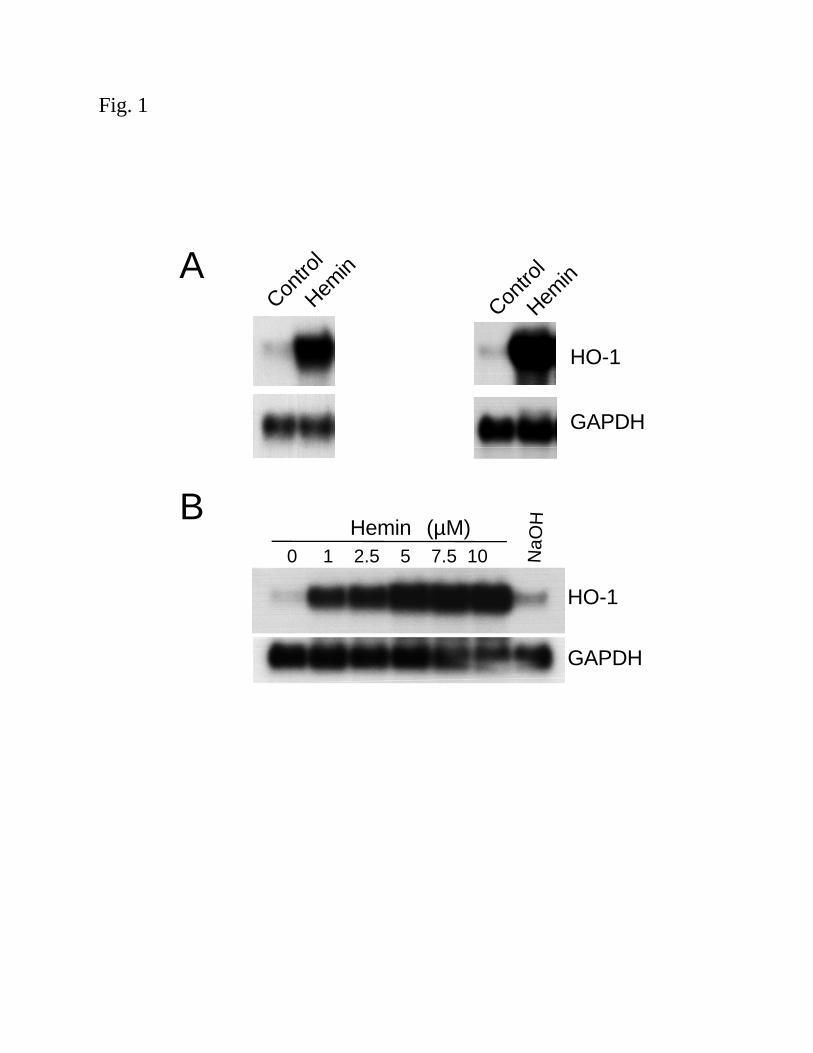
Induction of HO-1 mRNA by heme in human renal proximal tubular cells and aortic endothelial
cells: Heme serves not only as a substrate for the heme oxygenase enzyme, but is also a potent
inducer of the HO-1 gene both in vivo and in vitro (11, 32, 50). In order to understand the
molecular mechanism involved in heme-mediated induction of HO-1, HPTC and HAEC were
treated with heme (in the form of hemin, 5µM) for 4h. Northern analysis demonstrated a ~20-30
fold induction of HO-1 mRNA in both cell types (Fig. 1A). Fig. 1B and C demonstrate that the
induction of HO-1 mRNA by hemin was both time- and dose-dependent in HPTC. Similar
results were obtained in HAEC (data not shown). Sodium hydroxide (40 µM) alone, the vehicle
used in the preparation of hemin, did not induce HO-1 mRNA (Fig. 1B).
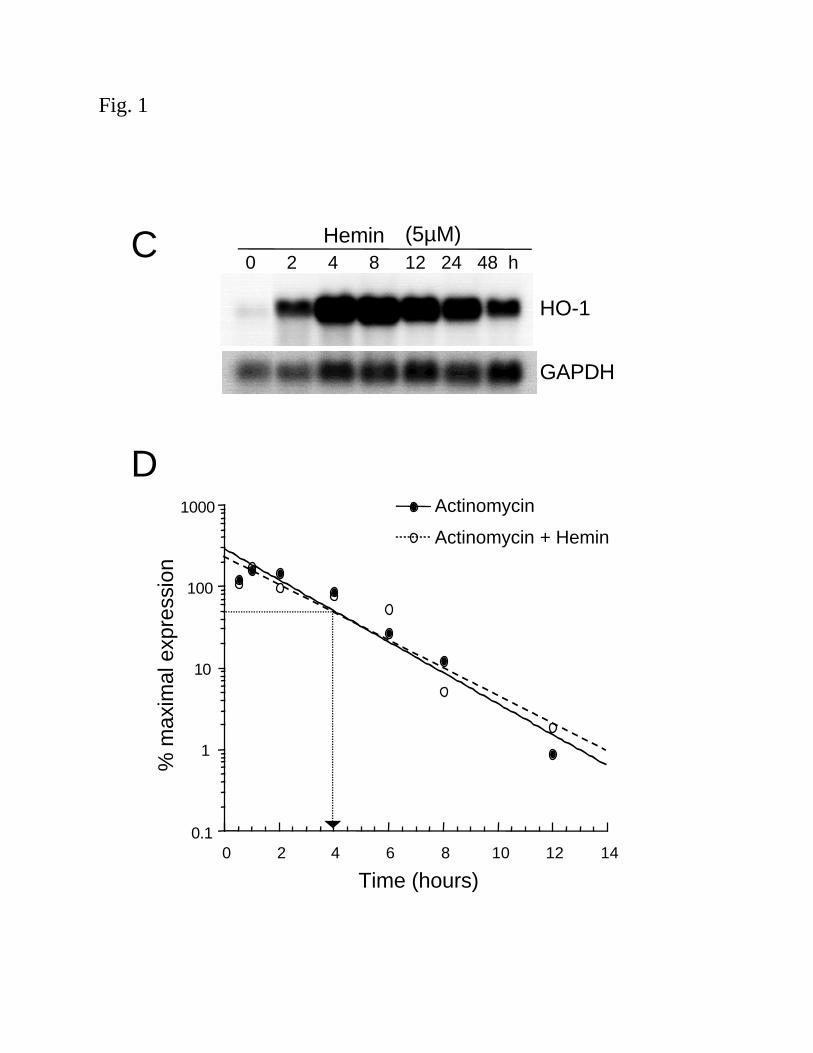
We have previously demonstrated the importance of de novo transcription by nuclear run-
on analysis in HAEC and HPTC treated with hemin (5µM) and observed increased HO-1 gene
transcription (4). To evaluate the role of message stability in the induction of HO-1 mRNA
following stimulation with hemin, the half-life of HO-1 mRNA was measured. Confluent HPTC
were exposed to hemin (5µM) for 4 h, washed with HBSS followed by the addition of fresh
media containing actinomycin D (4µM) with or without additional hemin. As shown in Fig. 1D,
the half-life of HO-1 mRNA was similar (~4h) with and without additional hemin treatment,
suggesting that message stability is not involved. These findings are consistent with previous
studies in other cell types (10) and enable us to use these human primary cultured cells for
further studies to evaluate the transcriptional activation of the human HO-1 gene by hemin.
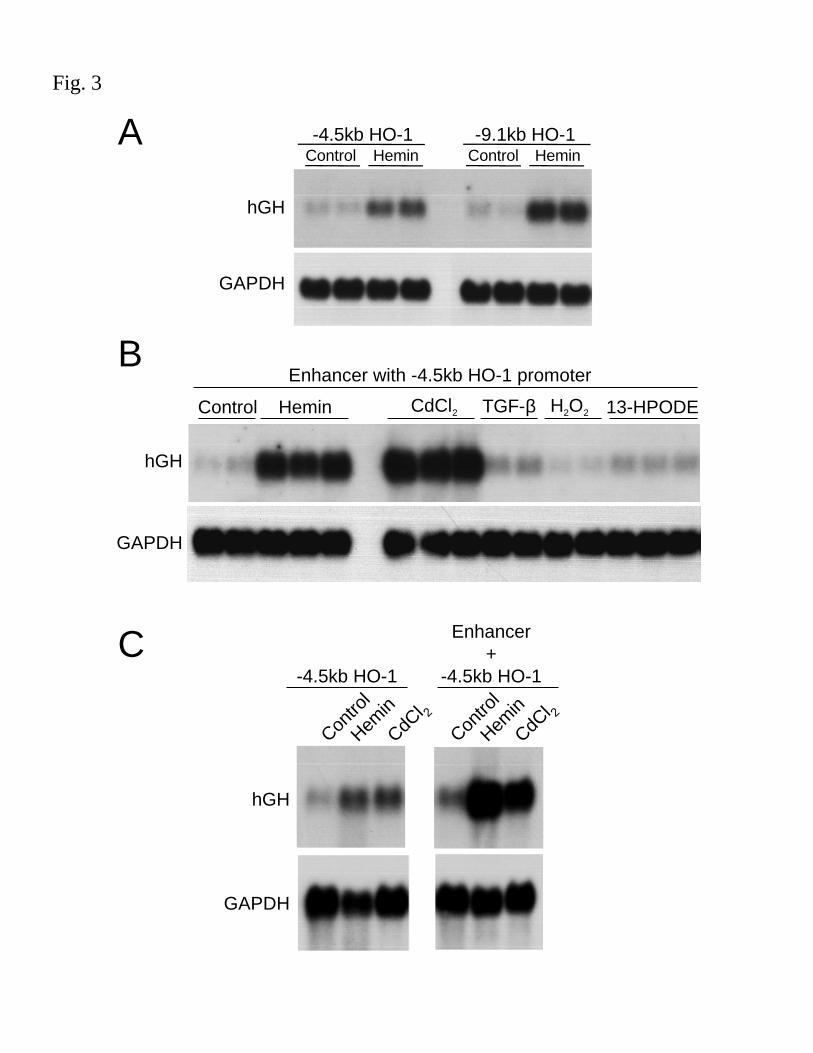
Identification of the enhancer region internal to the human HO-1 gene: To identify regulatory
elements in the human HO-1 gene that mediate its transcription following stimulation with
hemin, multiple promoter fragments (155bp, 4.0kb, 4.5kb and 9.1kb) were generated and

F-00137-2003
11
incorporated into promoterless reporter vectors as shown in Fig. 2 and described in the Methods.
Growth hormone mRNA levels, measured by northern analysis, were used as a direct measure of
reporter gene transcription rates. HAEC transfected with hGH vectors containing –155bp or
–4.0kb promoter constructs demonstrated no hGH mRNA induction in response to hemin (data
not shown) and the -4.5 and -9.1kb constructs resulted in a modest hemin-mediated induction in
hGH mRNA (~6.7 fold and ~9.9 fold, respectively), as shown in Fig. 3A.
Since levels of reporter gene activity did not correlate with steady state northern levels of
HO-1 induction with hemin, we hypothesized that additional regulatory sequences might exist
within the HO-1 gene. Therefore, construct containing the 12.5kb region internal to the HO-1
gene with the 4.5kb HO-1 promoter (pHOGH/4.5/+12.5) was generated. Unfortunately, we were
unable to subclone the 12.5kb fragment with the 9.1kb HO-1 promoter. The construct,
pHOGH/4.5/+12.5, demonstrated significant hemin- as well as CdCl2-inducible hGH mRNA in
HAEC (16.1 fold and 32.4 fold, respectively, Fig. 3B). Similar results were observed in HPTC,
pHOGH/4.5 yielded a 6.6 and 4.3 fold increase in response to hemin and CdCl2, respectively,
and pHOGH/4.5/+12.5 resulted in a 27 and 18 fold, increase in response to hemin and CdCl2,
respectively, over control (untreated cells) transfected with pHOGH/4.5 (Fig. 3C). These results
indicate the presence of sequences internal to the HO-1 gene that potentially function as an
enhancer. Interestingly, the putative enhancer region does not function for all known inducers of
HO-1 as pHOGH/4.5/+12.5 failed to respond to TGF-β1 (2ng/ml), H2O2 (200µM), or 13-
HPODE (25µM) (Fig. 3B).
Reporter activity, as measured by hGH protein or luciferase activity, yielded results
similar to hGH mRNA measurements. No promoter activity was detected in cells transfected
with hGH constructs containing –155bp or –4.0kb promoter fragments and treated with hemin

F-00137-2003
12
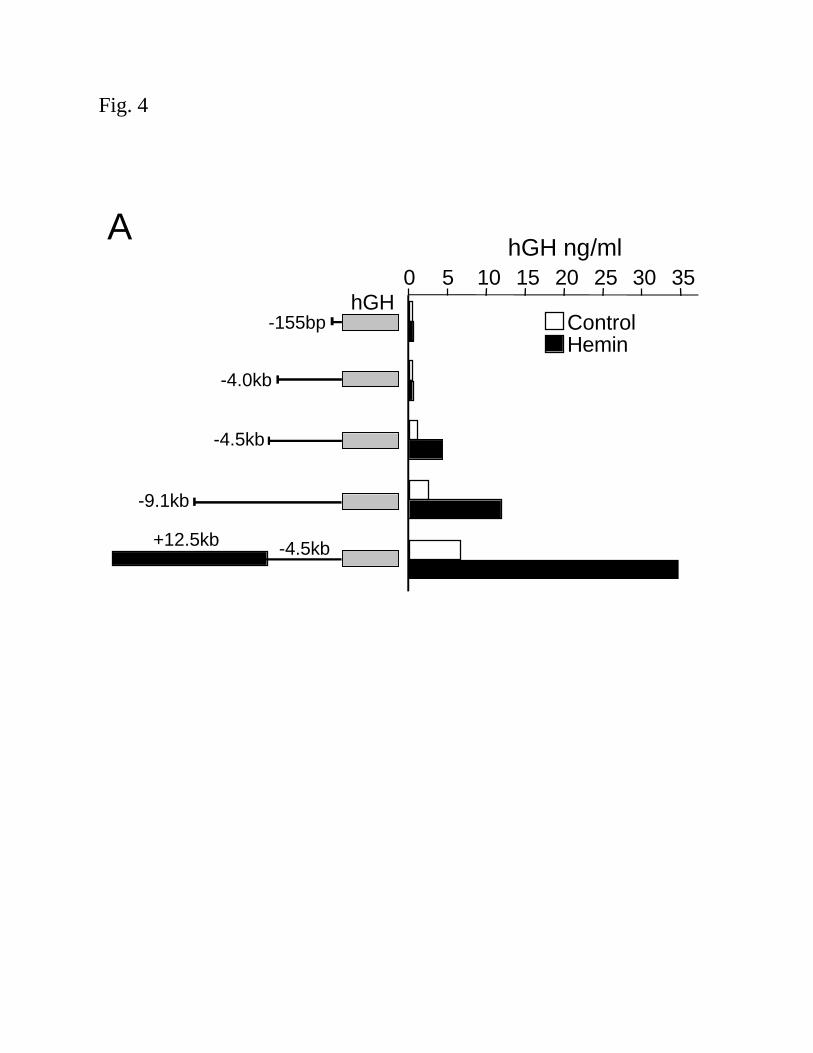
(5µM) (Fig. 4A). Cells transfected with vectors containing the -4.5kb or –9.1kb promoter
constructs and exposed to hemin (5µM) demonstrated a 3.6 and 4.5 fold, respectively, increase in
hGH protein, over untreated cells (Fig. 4A). The construct containing the 12.5kb region internal
to the HO-1 gene, in conjunction with the –4.5kb HO-1 promoter region, demonstrated elevated
basal levels as well as significant hemin-inducible hGH protein levels, ~5.1 fold over untreated
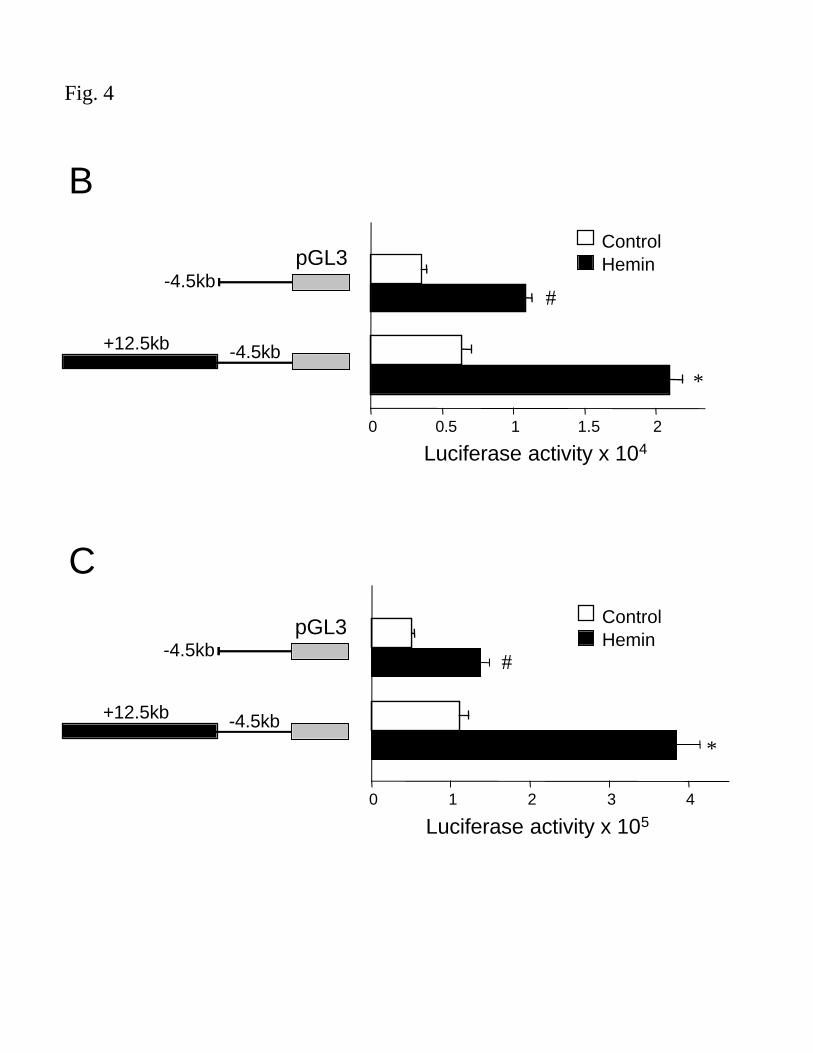
cells (Fig 4A). Luciferase reporter vectors containing HO-1 gene fragments were generated to
confirm the results with another reporter gene as well as ease of cloning and simplicity of the
assay. Luciferase/HO-1 vectors yielded results similar to hGH reporter vectors in both HAEC
and HPTC (Fig 4B and C, respectively).
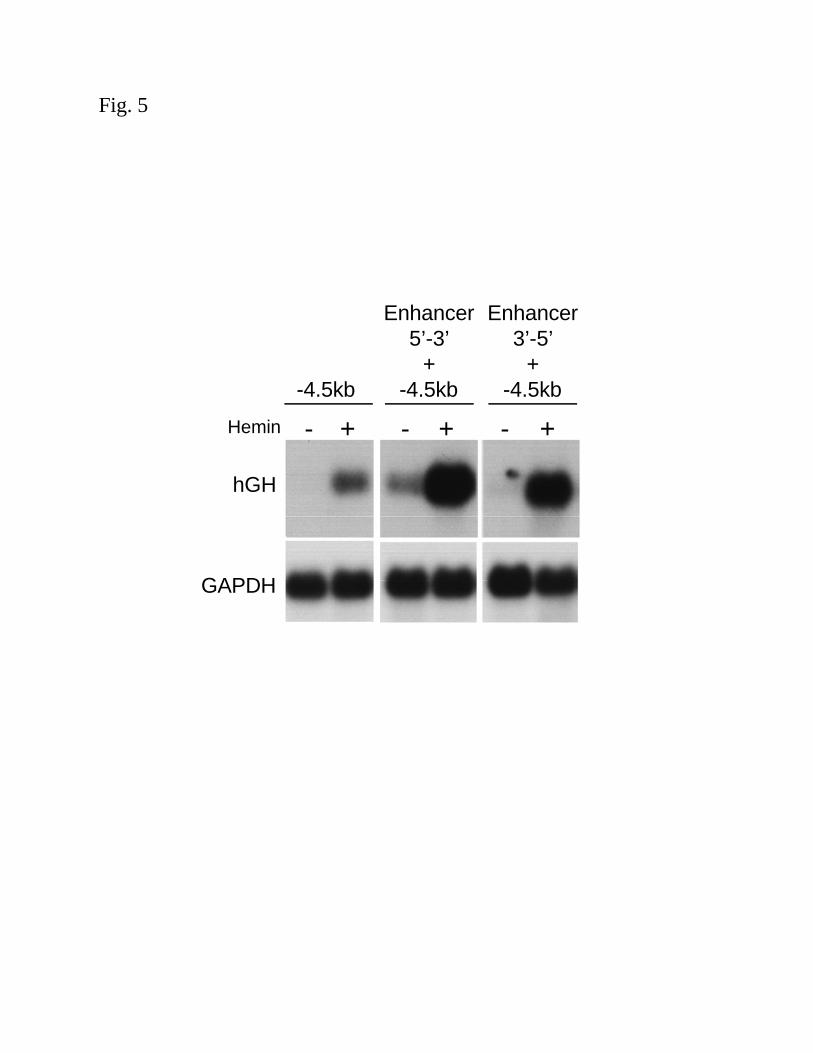
Orientation independent effects of the HO-1 enhancer region: In order to determine if the
+12.5kb fragment functions as a true enhancer, both orientations (5’-3’ and 3’-5’) were cloned
into pHOGH/4.5, to give pHOGH/4.5/+12.5 and pHOGH/4.5/+12.5(3’-5’). Similar levels of
heme-mediated induction, as measured by northern analysis of hGH mRNA were observed,
regardless of orientation (Fig. 5). In addition, to determine if the enhancer could function with a
heterologous promoter, the +12.5kb fragment was cloned into a hGH vector containing a TK
promoter (pHOTKGH/+12.5), which is a 200bp minimal, TATA-containing promoter. Transient
transfection of the 12.5kb fragment in conjunction with a TK promoter failed to respond to
hemin, as assessed by northern analysis of hGH mRNA (data not shown) suggesting that the
enhancer only functions with the HO-1 promoter.
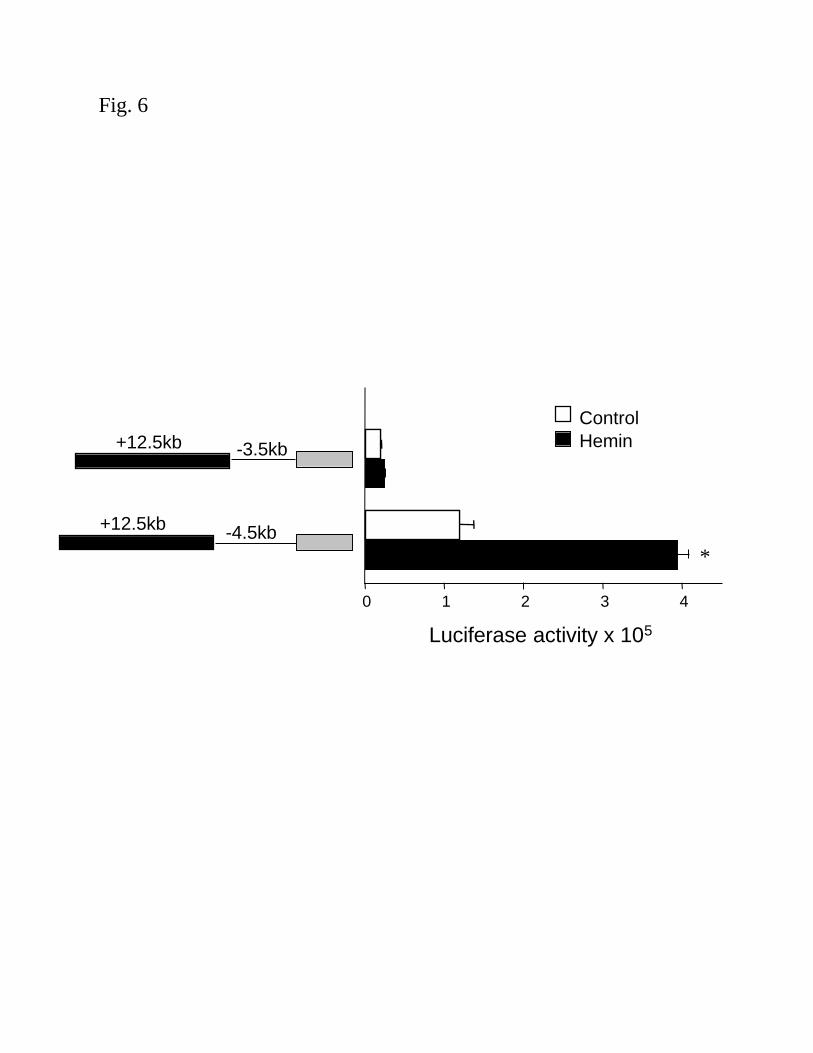
To further define the relevant sequences in the HO-1 promoter region that are required
for enhancer function, the +12.5 enhancer fragment was evaluated with a 3.5kb promoter of the
human HO-1 gene (pHOGH/3/5/+12.5). As shown in Fig. 6, the +12.5kb fragment requires the
4.5kb HO-1 promoter fragment to function, since the 3.5kb promoter with the +12.5kb enhancer

F-00137-2003
13
did not demonstrate any basal or hemin-inducible activity. The results demonstrate that enhancer
sequences internal to the human HO-1 gene (within 12.5kb), in conjunction with regions
between –3.5 and –4.5kb of the HO-1 promoter, mediate transcriptional activation of the human
HO-1 gene by hemin and the enhancer is likely composed of a complex set of interacting
elements.

F-00137-2003
14
DISCUSSION
Induction of HO-1 is an adaptive and protective response in renal ischemia-reperfusion
injury (40), nephrotoxin-induced renal injury (1, 41), organ transplantation (23), acute
glomerulonephritis (47), rhabdomyolysis (32, 34) as well as non-renal settings of tissue injury
(3). The protective effects of HO-1 were first recognized due to its robust induction in
cells/tissues exposed to a wide variety of stimuli that are otherwise injurious (24, 32). The
mechanisms underlying HO-1 induction by multiple inducers including heme, CdCl2, nitric
oxide (NO), oxidized LDL and cytokines are complex and regulated predominantly at the
transcriptional level. Increased mRNA stability has been reported to contribute to induction of
HO-1 by NO donors in primary human embryonic fibroblasts with a strong correlation between
the rate of NO release and the half-life of HO-1 mRNA (14). Furthermore, HO-1 mRNA stability
was independent of RNA or protein synthesis. Our previous studies using nuclear run-on assays
(4) combined with the present half-life experiments with actinomycin D suggest that mRNA
stability is not involved in heme-mediated HO-1 induction in renal epithelial and endothelial
cells, results consistent with previously published observations in other cell types (10).
Thus far, studies on mammalian HO-1 gene regulation have focused mainly on the mouse
HO-1 gene (5-13), where multiple inducer-specific stress response (StRE) elements have been
localized within 10kb of the 5’ flanking region. Two distal promoter regions, E1 and E2, at –4.0
and –10kb, respectively, are required for induction of the mouse HO-1 gene in response to heme,
heavy metals, H2O2 and sodium arsenite (5-7, 13). It was proposed that activation of the mouse
HO-1 gene by most stimuli is mediated exclusively via the E1 and/or E2 regions. These regions
contain consensus binding sites for the NF-E2 related factor, Nrf2 and overexpression studies

F-00137-2003
15
coupled with mutations in these binding sites have implicated the involvement of Nrf2 in the
induction of the mouse HO-1 gene (12, 20).
To test whether promoter elements in the 5’ flanking region of the human HO-1 gene
can analogously control heme-mediated induction, we initially analyzed several human HO-1
promoter constructs up to –9.1kb, that contain sequences potentially similar to the E1 and E2
regions in the mouse gene. The 4.5 and the 9.1kb promoter fragments demonstrated a modest
induction with heme, however, unlike the E1 and E2 regions of the mouse gene these constructs
did not completely recapitulate steady state northern levels of HO-1 induction. In addition,
unlike the mouse promoter, we also found that the human HO-1 promoter constructs showed no
response to other known HO-1 stimuli, including 13-HPODE, hyperoxia, curcumin and TGF-β1
in human cultured cells (4, 19 and unpublished observations).
In our efforts to mimic endogenous, stimulus-dependent transcription levels, we
hypothesized that additional regulatory element(s) were necessary for heme and cadmium-
mediated induction of the human HO-1 gene. To identify other relevant regulatory sequences, we
tested the entire +12.5kb HO-1 gene, including introns and exons, in conjunction with the –4.5kb
human HO-1 promoter and observed significant heme and cadmium-dependent induction of
reporter activity. Furthermore, this enhancer region was specific for heme and cadmium and did
not function for other known HO-1 stimuli, such as TGF-β1, H2O2, or 13-HPODE. The large
+12.5kb fragment, in part, satisfied the characteristics of a true enhancer region by functioning in
an orientation independent manner. However, when the HO-1 promoter was replaced with a
heterologous TK promoter, no induction by heme was observed, thus demonstrating that this
region functions as a gene specific enhancer. This is substantiated by our findings that the
enhancer region requires a portion of the HO-1 promoter to mimic endogenous levels of
induction. Specifically, we have demonstrated that a region between –4.5 and –3.5kb is required

F-00137-2003
16
for enhancer function. Our data has also indicated the importance of an additional region
between –4.5 and –9.1 for heme-mediated HO-1 promoter activity. However, we were unable to
evaluate the +12.5kb enhancer in the context of the –9.1kb promoter construct due to our
inability to successfully subclone both of these large fragments into a single plasmid. Studies to
further delineate the regulatory sequences within the enhancer region are currently in progress.
As these regions become more well defined, constructs containing the relevant sequences within
the enhancer will be evaluated with the –9.1kb HO-1 promoter.
Regulatory sequences functioning as enhancers have been identified within introns of
several genes, including manganese superoxide dismutase, platelet-derived growth factor-A, and
alcohol dehydrogenase-1 (15, 37, 45). We have performed sequence comparisons with the 12.5
kb enhancer region and have not identified any sequences that are linked to known heme or
cadmium response elements. Alam and Den (8) had previously reported that the region
analogous to our enhancer region containing the entire protein coding region of the mouse HO-1
gene coupled to the mouse HO-1 promoter did not respond to heme or cadmium in rodent cells.
Two possibilities could account for the lack of response. On the one hand, the limited size
(~3.0kb) of the mouse HO-1 promoter used in these studies may account for the negative results
(8). On the other hand, perhaps it is a reflection of the differential regulation of the mouse versus
the human HO-1 genes.
Our observations indicate that the human HO-1 gene requires regulatory sequences that
differ from those previously implicated for the mouse gene. Other interspecies differences in
HO-1 gene regulation have also been noted. For example, the human HO-1 promoter contains a
GT repeat region that is absent in the mouse HO-1 gene (49). Deletion of this repeat region
results in a significant elevation of basal HO-1 promoter activity (unpublished observations),
similar to previously reported results using HO-1 promoter constructs with shorter GT repeats

F-00137-2003
17
(49), suggesting that this human specific region may function as a negative regulator of HO-1
gene expression. Studies have also demonstrated that HO-1 is induced by hypoxia in rat, bovine,
mouse and monkey cells, but is rather repressed in human cells (25, 27). Similarly, HO-1 has
been popularized as heat shock protein 32 (HSP32) based on its inducibility by heat shock in
rodent cells (38, 39). However, heat shock does not induce HO-1 in human cells (35). While
cytokines induce HO-1 in rodent cells, previous studies have reported that interferon-γ decreases
human HO-1 gene expression (43). Furthermore, a putative cadmium response element has been
identified in the human HO-1 promoter (44), however, the cadmium response element in the
mouse HO-1 gene is immediately 3’ to this region and is an Nrf2 consensus sequence (13).
Collectively, these observations suggest that the underlying molecular mechanisms that regulate
the human HO-1 gene are different from those described for mouse HO-1 gene expression.
Recent studies have demonstrated that the expression of the mouse HO-1 gene is
regulated through antagonism of transcription activators and the repressor, Bach1 (42). Under
normal physiological conditions, HO-1 expression is repressed by Bach1, and increased heme
levels displace Bach1 from the E1 and E2 regions, allowing activators to bind to the regulatory
sequences (42). The authors suggest that the transcriptional regulation of the mouse HO-1 gene
involves a direct sensing of heme levels by Bach1, generating a feedback loop by the substrate,
as in the lac operon. Studies to explore the involvement of Bach1 in human HO-1 gene
regulation will be of interest.
In summary, our data provide evidence that at least two regions in the human HO-1
promoter, one between –4.5 and –4.0 and another between –9.1 and –4.5, are required for
activation of the HO-1 gene by heme. Most importantly, we have identified a novel
transcriptional enhancer internal to the human HO-1 gene that, in conjunction with the HO-1
promoter, recapitulates steady state level expression of HO-1 by heme and cadmium. Studies on

F-00137-2003
18
regulation of the human HO-1 gene are biologically relevant because of the well documented
beneficial effects of HO-1 activity which include degradation of heme (a toxic pro-oxidant),
generation of bilirubin (an antioxidant), co-induction of ferritin (an intracellular repository for
iron), and the anti-apoptotic and anti-inflammatory effects of CO (36). We are continuing our
investigation of the molecular mechanisms involved in the regulation of the human HO-1 gene in
an effort to fine tune endogenous HO-1 gene expression to ultimately optimize its potential as a
therapeutic modality.

F-00137-2003
19
ACKNOWLEDGMENTS
This work was supported by National Institutes of Health Grants K08 DK02446 and R01
DK59600 (to AA) and a K01 DK02902 (to NHK).

F-00137-2003
20
REFERENCES
1. Abraham NG, Lavrovsky Y, Schwartzman ML, Stoltz RA, Levere RD, Gerritsen ME,
Shibahara S, Kappas A. Transfection of the human heme oxygenase gene into rabbit
coronary microvessel endothelial cells: protective effect against heme and hemoglobin
toxicity. Proc Natl Acad Sci U S A 92: 6798-6802, 1995.
2. Agarwal A, Balla J, Alam J, Croatt AJ, Nath KA. Induction of heme oxygenase in toxic renal
injury: A protective role in cisplatin nephrotoxicity in the rat. Kidney Int 48: 1298-1307,
1995.
3. Agarwal A, Nick HS. Renal response to tissue injury: lessons from heme oxygenase-1 gene
ablation and expression. J Am Soc Nephrol 11: 965-73, 2000.
4. Agarwal A, Shiraishi F, Visner GA, Nick HS. Linoleyl hydroperoxide transcriptionally
upregulates heme oxygenase-1 gene expression in human renal epithelial and aortic
endothelial cells. J Am Soc Nephrol 9: 1990-1997, 1998.
5. Alam J. Multiple elements within the 5' distal enhancer of the mouse heme oxygenase-1 gene
mediate induction by heavy metals. J Biol Chem 269: 25049-25056, 1994.
6. Alam J, Cai J, Smith A. Isolation and characterization of the mouse heme oxygenase-1 gene.
Distal 5’ sequences are required for induction by heme and heavy metals. J Biol Chem 269:
1001-1009, 1994.
7. Alam J, Camhi S, Choi AM. Identification of a second region upstream of the mouse heme
oxygenase-1 gene that functions as a basal level and inducer-dependent transcription
enhancer. J Biol Chem 270: 11977-11984, 1995.
8 . Alam J, Den Z. Distal AP-1 binding sites mediate basal level enhancement and TPA
induction of the mouse heme oxygenase-1 gene. J Biol Chem 267: 21894-218900, 1992.

F-00137-2003
21
9. Alam J, Killeen E, Gong P, Naquin R, Hu B, Stewart D, Ingelfinger JR, Nath KA. Heme
activates the heme oxygenase-1 gene in renal epithelial cells by stabilizing Nrf2. Am J
Physiol Renal Physiol 284: F743-F752, 2003.
10. Alam J, Shibahara S, Smith A. Transcriptional activation of the heme oxygenase gene by
heme and cadmium in mouse hepatoma cells. J Biol Chem 264: 6371-6375, 1989.
11. Alam J, Smith A. Receptor-mediated transport of heme by hemopexin regulates gene
expression in mammalian cells. J Biol Chem 264: 17637-17640, 1989.
12. Alam J, Stewart D, Touchard C, Boinapally S, Choi AM, Cook JL. Nrf2, a Cap'n'Collar
transcription factor, regulates induction of the heme oxygenase-1 gene. J Biol Chem 274:
26071-26078, 1999.
13. Alam J, Wicks C, Stewart D, Gong P, Touchard C, Otterbein S, Choi AM, Burow ME, Tou J.
Mechanism of heme oxygenase-1 gene activation by cadmium in MCF-7 mammary
epithelial cells. Role of p38 kinase and Nrf2 transcription factor. J Biol Chem 275: 27694-
27702, 2000.
14. Bouton C, Demple B. Nitric oxide-inducible expression of heme oxygenase-1 in human cells.
Translation-independent stabilization of the mRNA and evidence for direct action of nitric
oxide. J Biol Chem 275: 32688-32693, 2000.
15. Callis J, Fromm M, Walbot V. Introns increase gene expression in cultured maize cells.
Genes Dev 1: 1183-1200, 1987.
16. Chen YH, Lin SJ, Lin MW, Tsai HL, Kuo SS, Chen JW, Charng MJ, Wu TC, Chen LC,
Ding YA, Pan WH, Jou YS, Chau LY. Microsatellite polymorphism in promoter of heme
oxygenase-1 gene is associated with susceptibility to coronary artery disease in type 2
diabetic patients. Hum Genet 111: 1-8, 2002.

F-00137-2003
22
17. Chomczynski P, Sacchi N. Single-step method of RNA isolation by acid guanidinium
thiocyanate-phenol-chloroform extraction. Anal Biochem 162: 156-159, 1987.
18. Exner M, Schillinger M, Minar E, Mlekusch W, Schlerka G, Haumer M, Mannhalter C,
Wagner O. Heme oxygenase-1 gene promoter microsatellite polymorphism is associated with
restenosis after percutaneous transluminal angioplasty. J Endovasc Ther 8: 433-440, 2001.
19. Fogg S, Agarwal A, Nick HS, Visner GA. Iron regulates hyperoxia-dependent human heme
oxygenase 1 gene expression in pulmonary endothelial cells. Am J Respir Cell Mol Biol 20:
797-804, 1999.
20. Gong P, Stewart D, Hu B, Li N, Cook J, Nel A, Alam J. Activation of the mouse heme
oxygenase-1 gene by 15-deoxy-Delta(12,14)-prostaglandin J(2) is mediated by the stress
response elements and transcription factor Nrf2. Antioxid Redox Signal 4: 249-257, 2002.
21. Hill-Kapturczak N, Chang SH, Agarwal A. Heme oxygenase and the kidney. DNA Cell Biol
21: 307-321, 2002.
22. Jeney V, Balla J, Yachie A, Varga Z, Vercellotti GM, Eaton JW, Balla G. Pro-oxidant and
cytotoxic effects of circulating heme. Blood 100: 879-887, 2002.
23 . Katori M, Busuttil RW, Kupiec-Weglinski JW. Heme oxygenase-1 system in organ
transplantation. Transplantation 74: 905-912, 2002
24. Keyse SM, Tyrrell RM. Heme oxygenase is the major 32-kDa stress protein induced in
human skin fibroblasts by UVA radiation, hydrogen peroxide, and sodium arsenite. Proc Natl
Acad Sci U S A 86: 99-103, 1989.
25. Kitamuro T, Takahashi K, Ogawa K, Udono-Fujimori R, Takeda K, Furuyama K, Nakayama
M, Sun J, Fujita H, Hida W, Hattori T, Shirato K, Igarashi K, Shibahara S. Bach1 functions
as a hypoxia-inducible repressor for the heme oxygenase-1 gene in human cells. J Biol Chem
278: 9125-9133, 2003.

F-00137-2003
23
26. Kutty RK, Kutty G, Rodriguez IR, Chader GJ, Wiggert B. Chromosomal localization of the
human heme oxygenase genes: heme oxygenase-1 (HMOX1) maps to chromosome 22q12
and heme oxygenase-2 (HMOX2) maps to chromosome 16p13.3. Genomics 20: 513-516,
1994.
27. Lee PJ, Jiang BH, Chin BY, Iyer NV, Alam J, Semenza GL, Choi AM. Hypoxia-inducible
factor-1 mediates transcriptional activation of the heme oxygenase-1 gene in response to
hypoxia. J Biol Chem 272: 5375-5381, 1997.
28. Maines MD. The heme oxygenase system: A regulator of second messenger gases. Annu Rev
Pharmacol Toxicol 37: 517-554, 1997.
29. Maines MD, Mayer RD, Ewing JF, McCoubrey WK Jr. Induction of kidney heme
oxygenase-1 (HSP-32) mRNA and protein by ischemia/reperfusion: possible role of heme as
both promotor of tissue damage and regulator of HSP32. J Pharmacol Exp Ther 264: 457-
462, 1993.
30. McCoubrey WK Jr, Huang TJ, Maines MD. Isolation and characterization of a cDNA from
the rat brain that encodes hemoprotein heme oxygenase-3. Eur J Biochem 247: 725-732,
1997.
31. Nath KA, Agarwal A, Vogt BV. Heme oxygenase: Cytoprotective and cytotoxic effects.
Contemporary Issues in Nephrology 30: 97-118, 1996.
32. Nath KA, Balla G, Vercellotti GM, Balla J, Jacob HS, Levitt MD, Rosenberg ME. Induction
of heme oxygenase is a rapid, protective response in rhabdomyolysis in the rat. J Clin Invest
90: 267-270, 1992.
33. Nath KA, Grande JP, Croatt AJ, Likely S, Hebbel RP, Enright H. Intracellular targets in
heme protein-induced renal injury. Kidney Int 53: 100-111, 1998.

F-00137-2003
24
34. Nath KA, Haggard JJ, Croatt AJ, Grande JP, Poss KD, Alam J. The indispensability of heme
oxygenase-1 in protecting against acute heme protein-induced toxicity in vivo. Am J Pathol
156: 1527-1535, 2000.
35. Okinaga S, Takahashi K, Takeda K, Yoshizawa M, Fujita H, Sasaki H, Shibahara S.
Regulation of human heme oxygenase-1 gene expression under thermal stress. Blood 87:
5074-5084, 1996.
36. Platt JL, Nath KA. Heme oxygenase: protective gene or Trojan horse. Nat Med 4: 1364-
1365, 1998.
37. Rogers RJ, Chesrown SE, Kuo S, Monnier JM, Nick HS. Cytokine-inducible enhancer with
promoter activity in both the rat and human manganese-superoxide dismutase genes.
Biochem J 347: 233-242, 2000.
38. Shibahara S, Kitamuro T, Takahashi K. Heme degradation and human disease: diversity is
the soul of life. Antioxid Redox Signal 4: 593-602, 2002.
39. Shibahara S, Muller RM, Taguchi H. Transcriptional control of rat heme oxygenase by heat
shock. J Biol Chem 262: 12889-12892, 1987.
40. Shimizu H, Takahashi T, Suzuki T, Yamasaki A, Fujiwara T, Odaka Y, Hirakawa M, Fujita
H, Akagi R. Protective effect of heme oxygenase induction in ischemic acute renal failure.
Crit Care Med 28: 809-817, 2000.
41. Shiraishi F, Curtis LM, Truong L, Poss K, Visner GA, Madsen K, Nick HS, Agarwal A.
Heme oxygenase-1 gene ablation or expression modulates cisplatin-induced renal tubular
apoptosis. Am J Physiol Renal Physiol 278: F726-F736, 2000.
42. Sun J, Hoshino H, Takaku K, Nakajima O, Muto A, Suzuki H, Tashiro S, Takahashi S,
Shibahara S, Alam J, Taketo MM, Yamamoto M, Igarashi K. Hemoprotein Bach1 regulates
enhancer availability of heme oxygenase-1 gene. EMBO J 21: 5216-5224, 2002.

F-00137-2003
25
43. Takahashi K, Nakayama M, Takeda K, Fujia H, Shibahara S. Suppression of heme
oxygenase-1 mRNA expression by interferon-gamma in human glioblastoma cells. J
Neurochem. 72: 2356-2361, 1999
44. Takeda K, Ishizawa S, Sato M, Yoshida T, Shibahara S. Identification of a cis-acting element
that is responsible for cadmium-mediated induction of the human heme oxygenase gene. J
Biol Chem 269: 22858-22867, 1994.
45. Takimoto Y, Kuramoto A. Presence of a regulatory element within the first intron of the
human platelet-derived growth factor-A chain gene. Jpn J Cancer Res 84: 1268-1272, 1993.
46. Visner GA, Staples ED, Chesrown SE, Block ER, Zander DS, Nick HS. Isolation and
maintenance of human pulmonary artery endothelial cells in culture isolated from transplant
donors. Am J Physiol 267: L406-L413, 1994.
47. Vogt BA, Shanley TP, Croatt A, Alam J, Johnson KJ, Nath KA. Glomerular inflammation
induces resistance to tubular injury in the rat. A novel form of acquired, heme oxygenase-
dependent resistance to renal injury. J Clin Invest 98: 2139-2145, 1996.
48. Yachie A, Niida Y, Wada T, Igarashi N, Kaneda H, Toma T, Ohta K, Kasahara Y, Koizumi
S. Oxidative stress causes enhanced endothelial cell injury in human heme oxygenase-1
deficiency. J Clin Invest 103: 129-135, 1999.
49. Yamada N, Yamaya M, Okinaga S, Nakayama K, Sekizawa K, Shibahara S, Sasaki H.
Microsatellite polymorphism in the heme oxygenase-1 gene promoter is associated with
susceptibility to emphysema. Am J Hum Genet 66: 187-195, 2000.
50. Yoshida T, Biro P, Cohen T, Muller RM, Shibahara S. Human heme oxygenase cDNA and
induction of its mRNA by hemin. Eur J Biochem 171: 457-461, 1988.

F-00137-2003
26
Figure Legends
Figure 1. Induction of HO-1 mRNA by hemin. (A) Confluent HAEC (left panel) or HPTC
(right panel) were incubated in 1% and 0.5% FBS containing media, respectively, with hemin
(5µM) for 4h as described in the Methods. (B) Confluent HPTC incubated in 0.5% FBS medium
containing hemin (0, 1, 2.5, 5, 7.5 or 10 µM) and vehicle, NaOH (40µM), for 4h. (C) Time
course of HO-1 mRNA induction in HPTC exposed to hemin (5µM) at the indicated times. (D)
A graphic representation showing the half-life of HO-1 mRNA following hemin stimulation.
HPTC were preincubated with hemin (5µM) for 4h, washed, and exposed to actinomycin D
(4µM) in the absence (closed circle, solid line) or presence (open circle, dashed line) of
additional hemin (5µM). The percent maximal expression of HO-1 mRNA corrected for the
internal control (GAPDH) versus time is plotted. RNA was isolated and subjected to northern
blot analysis with a 32P-labeled cDNA specific for HO-1 or GAPDH as described in the
Methods. Results are representative of at least 2-4 independent experiments.
Figure 2. Genomic structure of the human HO-1 gene and reporter gene constructs. A
schematic representation of the human HO-1 genomic clone isolated from the P1 and BAC
clones with a partial restriction map extending from –11.614kb to +16.9kb is shown in the upper
panel. The 5 exons are indicated (E). The restriction sites for NheI, N; BamHI, B; PstI, P; XbaI,
Xb; EcoRI, R are indicated. The plasmid constructs used to assess human HO-1 promoter and
enhancer activity are shown in the lower panel. Details for the generation of these constructs is
provided in the Methods.
Figure 3. Analysis of human growth hormone (hGH) mRNA in cells transfected with hGH-
HO-1 reporter constructs. HAEC were transiently transfected with equimolar amounts of

F-00137-2003
27
pHOGH/4.5 (-4.5kb HO-1 promoter in hGH vector) or pHOGH/9.1 (-9.1kb HO-1 promoter in
hGH vector) (A) or pHOGH/4.5/+12.5 (enhancer with –4.5kb HO-1 promoter in hGH vector)
(B) using DEAE/dextran and a batch transfection protocol as described in the Methods.
Transfected cells were exposed to hemin (5µM), cadmium chloride (CdCl2, 10µM), transforming
growth factor-β1 (TGF-β1, 2ng/ml), or 13-hydroxyperoxyoctadecadienoic acid (13-HPODE,
25µM) for 16h and analyzed for reporter gene expression by northern analysis. (C) HPTC were
transiently transfected with equimolar amounts of pHOGH/4.5 or pHOGH/4.5/+12.5 using
DEAE/dextran and a batch transfection protocol as described in the Methods. Cells were exposed
to hemin (5µM) or CdCl2 (10µΜ) for 16h prior to collection of RNA for reporter gene analysis.
Total RNA was isolated and northern analysis was performed using 32P-labeled hGH and
GAPDH cDNA probes. Results are representative of at least 3 independent experiments.
Figure 4. Analysis of reporter activity in cells transfected with HO-1-reporter constructs in
response to hemin. (A) HAEC were transiently transfected with equimolar amounts of
pHOGH/155 (-155bp HO-1 promoter), pHOGH/4 (-4.0kb HO-1 promoter), pHOGH/4.5 (-4.5kb
HO-1 promoter), pHOGH/9.1 (-9.1kb HO-1 promtoer) and pHOGH/4.5/+12.5 (enhancer with
–4.5kb HO-1 promoter) as described in the Methods. Cells were exposed to hemin (5µM) for
72h and secreted hGH protein in the media was measured using a radioimmunoassay kit as
described previously (4). Results are derived from 2 independent experiments. HAEC (B) and
HPTC (C) were transiently transfected with equimolar amounts of pHOGL3/4.5 (-4.5kb HO-1
promoter in luciferase vector) or pHOGL3/4.5/+12.5 (enhancer with –4.5kb HO-1 promoter in
luciferase vector), exposed to hemin (5µM) for 16h and luciferase activity was measured as
described in the Methods. Results are derived from 2 independent experiments with 3-12

F-00137-2003
28
replicates per group. Open bars represent untreated (control) cells and dark bars represent hemin
stimulated cells. #p<0.05 control vs. hemin; *p<0.001 vs. other groups (ANOVA).
Figure 5. Orientation independent effects of the HO-1 enhancer. HAEC were transiently
transfected with equimolar amounts of pHOGH/4.5 (-4.5kb HO-1 promoter in hGH vector),
pHOGH/4.5/+12.5 (5’-3’ orientation of the enhancer with –4.5kb promoter in hGH vector) and
pHOGH/4.5/+12.5(3’-5’) (3’-5’ orientation of the enhancer with –4.5kb promoter in hGH vector)
as described in the Methods. Cells were exposed to hemin (5µM) for 16h and total RNA was
isolated for northern analysis using 32P-labeled hGH and GAPDH cDNA probes.
Figure 6. Requirement of the –4.5kb HO-1 promoter for enhancer function. HPTC were
transiently transfected with equimolar amounts of pHOGL3/3.5/+12.5 (enhancer with –3.5kb
HO-1 promoter in a luciferase vector) or pHOGL3/4.5/+12.5 as described in the Methods. Cells
were exposed to hemin (5µM) for 16h and luciferase assays were performed. Results are derived
from two independent experiments with 6 replicates per group. Open bars represent untreated
(control) cells and dark bars represent hemin-stimulated cells. *p<0.001 vs. other groups
(ANOVA).

F-00137-2003
29
Table 1: Primers used for long-range PCR for human HO-1 constructs
Construct Forward Primer Reverse Primer
4.0 kb promoter GGATCCGGCAGAAGCCTCTTGTTTGTCT GGATCCGGCCGGTGCTGGGCTCGTTCGT
4.5 kb promoter GGATCCTGCAGAATCGAGCACATGTTC GGATCCGGCCGGTGCTGGGCTCGTTCGT
9.1 kb promoter GTCGACTGAGTAATCCTTTCCCGAG GTCGACGCTCCATCCGGCCGGTGCTGGGCTC
+12.5 kb
enhancer
GTCGACGGATGGAGCGTCCGCAACCCGACAG GTCGACTCTAGAAAGAAGGCCTTCCACCGGAC
B
A
HO-1
GAPDH
Contro
l
Hemin
Contro
l
Hemin
Fig. 1
Hemin (µM)0 1 2.5 5 7.5 10 N
aOH
HO-1
GAPDH
D
0.1
1
10
100
1000
0 2 4 6 8 10 12 14
Actinomycin
Actinomycin + Hemin
% m
axim
al e
xpre
ssio
n
Time (hours)
C
Fig. 1
HO-1
GAPDH
0 2 4 8 12 24 48 hHemin (5µM)
BAC Clone
P1 #11716
P1 #11715
E2E1 E5E4E3
R R RN NN NX
5’ 3’P Xb
B
B
B
-9.1kb
-4.5kb
-4.0kb
-3.5kb
-4.5kb
-3.5kb
0.155kb
1kb
pHOGH/155
pHOGL3/3.5
pHOGH/4.0
pHOGH/4.5 & pHOGL3/4.5
pHOGH/9.1 & pHOGL3/9.1
pHOGH/4.5/+12.5 & pHOGL3/4.5/+12.5
pHOGL3/3.5/+12.5
Fig. 2
+12.5kb
+12.5kb
GAPDH
hGH
-4.5kb HO-1 -9.1kb HO-1Control Hemin Control Hemin
Control Hemin CdCl2
Enhancer with -4.5kb HO-1 promoter
GAPDH
hGH
B
A
H2O2TGF-β 13-HPODE
C
Contro
l
Hemin
CdCl 2
hGH
GAPDH
Contro
l
Hemin
CdCl 2
-4.5kb HO-1
Enhancer+
-4.5kb HO-1
Fig. 3
0 5 10 15 20 25 30 35hGH ng/ml
ControlHemin
hGH-155bp
+12.5kb -4.5kb
-4.0kb
-4.5kb
-9.1kb
A
Fig. 4
B
+12.5kb -4.5kb
-4.5kb pGL3
0 1 2 3 4
HeminControl
Luciferase activity x 105
C
0 0.5 1 1.5 2
Luciferase activity x 104
+12.5kb -4.5kb
-4.5kb pGL3 Hemin
Control
#
*
#
*
Fig. 4
-4.5kb
Enhancer5’-3’
+-4.5kb
Hemin - + - + - +
GAPDH
hGH
Enhancer3’-5’
+-4.5kb
Fig. 5
0 1 2 3 4
+12.5kb -4.5kb
HeminControl
+12.5kb -3.5kb
Luciferase activity x 105
*
Fig. 6