detection of goldfish haematopoietic necrosis herpes virus (cyprinid herpesvirus-2) with multi-drug...
TRANSCRIPT

Dhio
PSNa
b
0c
d
a
ARRAA
KACCELM
1
attvfwW
t
h0
Acta Tropica 161 (2016) 8–17
Contents lists available at ScienceDirect
Acta Tropica
jo u r n al homep age: www.elsev ier .com/ locate /ac ta t ropica
etection of goldfish haematopoietic necrosis herpes virus (Cypriniderpesvirus-2) with multi-drug resistant Aeromonas hydrophila
nfection in goldfish: First evidence of any viral disease outbreak inrnamental freshwater aquaculture farms in India
.K. Sahoo a,∗, T. Raja Swaminathan b,1, Thangapalam Jawahar Abraham c, Raj Kumar b,
. Pattanayak a, A. Mohapatra a, S.S. Rath a, Avijit Patra c, Harresh Adikesavalu c,eeraj Sood d, P.K. Pradhan d, B.K. Das a, P. Jayasankar a, J.K. Jena d
ICAR-Central Institute of Freshwater Aquaculture, Kausalyaganga, Bhubaneswar 751 002, IndiaPeninsular and Marine Fish Genetic Resources Centre of ICAR-National Bureau of Fish Genetic Resources, CMFRI Campus, Ernakulum North PO, Kochi 68218, IndiaFaculty of Fishery Sciences, West Bengal University of Animal and Fishery Sciences, Chakgaria, Kolkata 700 094, IndiaICAR-National Bureau of Fish Genetic Resources, Canal Ring Road, Dilkusha, Lucknow 226 002, India
r t i c l e i n f o
rticle history:eceived 5 March 2016eceived in revised form 5 May 2016ccepted 6 May 2016vailable online 9 May 2016
eywords:eromonas hydrophilaarassius auratus
a b s t r a c t
This outbreak report details of a mortality event where Cyprinid herpes virus-2 (CyHV-2) was detectedin association with multidrug-resistant Aeromonas hydrophila infection in goldfish, Carassius auratus,from commercial farms. The goldfish exhibited large scale haemorrhages on the body, fins and gills,lepidorthosis, necrosed gills, protruded anus and shrunken eyes. White nodular necrotic foci in spleen andkidneys were noticed, along with necrosis and fusion of gill lamellae. Transmission electron microscopy ofaffected tissues revealed the presence of mature virus particles. Involvement of CyHV-2 was confirmed byPCR, sequencing and observed cytopathic effect in koi carp fin cell line along with experimental infectionstudy. A bacterium isolated from the internal organs of affected fish was found to be pathogenic Aeromonas
yprinid herpesvirus-2merging pathogenabeo rohitaultidrug-resistance
hydrophila having resistance to more than 10 classes of antibiotics. We postulate that CyHV-2 was theprimary etiological agent responsible for this outbreak with secondary infection by A. hydrophila. Theexperimental infection trials in Labeo rohita and koi carp by intraperitoneal challenge with CyHV-2 tissuehomogenates failed to reproduce the disease in those co-cultured fish species. This is the first report of aviral disease outbreak in organised earthen ornamental fish farms in India and bears further investigation.
© 2016 Elsevier B.V. All rights reserved.
. Introduction
Ornamental aquaculture has increased in popularity in India as result of its economic benefits due to a large internal market andhe potential for exports. The goldfish (Carassius auratus) is one ofhe most popular species both in terms of production and tradealue in India. Aquaculture of goldfish in ponds, ranging in size
rom 0.01 ha to more than 1.0 ha, either alone or in combinationith Indian Major Carps (IMC) is very common in some pockets ofest Bengal, India. While some of the broodstock are sourced from
∗ Corresponding author at: Fish Health Management Division, ICAR-Central Insti-ute of Freshwater Aquaculture, Kausalyaganga, Bhubaneswar 751 002, India.
E-mail address: [email protected] (P.K. Sahoo).1 Equally contributed as first author.
ttp://dx.doi.org/10.1016/j.actatropica.2016.05.004001-706X/© 2016 Elsevier B.V. All rights reserved.
home-grown stock, the vast majority are imported from neighbour-ing countries. Despite the large scale culture, no major viral diseaseoutbreak has been reported earlier in India, either from ornamentalfarms or the IMC farms that are the mainstay of Indian aquaculture.However, viral disease problems have been reported and charac-terized elsewhere in goldfish (Groff et al., 1998). Recently, Georgeet al. (2015) isolated and characterized a ranavirus from koi, Cypri-nus carpio L., experiencing mass mortalities in India and mortalitydue to viral nervous necrosis were reported in an experimental lotof goldfish and zebrafish in India (Binesh, 2013).
Herpes viral haematopoietic necrosis (HVHN) is a fatal diseaseof goldfish, leading to 100% mortality. It is caused by Cyprinid
herpesvirus-2 (CyHV-2) (Goodwin et al., 2009), a member of theCyprinid herpesvirus group that includes carp pox (CyHV-1) and koiherpesvirus (CyHV-3) (Goodwin et al., 2006b). CyHV-2 has mostly
a Trop
bae2iH1eeacalkheabce
ieasae2tf2hciwsogfo
2
2
tfibG(KibbvTwwctpc(T
P.K. Sahoo et al. / Act
een detected from goldfish (Goodwin et al., 2006b), although therere reports on mortalities in Prussian carp (Carassius gibelio) (Wut al., 2013) and crucian carp (Carassius carassius) (Bergmann et al.,010; Fichi et al., 2013).The disease has been reported world-wide
ncluding USA, UK, Australia, New Zealand, Taiwan, China, Italy,ungary, Czech Republic and most recently in France (Groff et al.,998; Stephens et al., 2004; Goodwin et al., 2006a,b, 2009; Jefferyt al., 2007; Fichi et al., 2013; Wu et al., 2013; Xu et al., 2013; Boitardt al., 2015) following its first report in juvenile goldfish (C. auratusuratus L.) in Japan in 1992 (Jung and Miyazaki, 1995). Commonharacteristics of the disease include high mortality in juvenilest temperature ranging from 15 to 25 ◦C, with the fish exhibitingethargy, anorexia, pale gills, and swollen and necrotic spleen andidney tissues (Goodwin et al., 2006a,b; Jeffery et al., 2007). Besidesistology and detection of the virus in tissues under ultrastructurexamination (Wu et al., 2013), isolation of virus using goldfish finnd koi fin cell lines (Xu et al., 2013; Ito and Maeno, 2014) and PCR-ased methods have been well developed for the identification andharacterization of CyHV-2 (Goodwin et al., 2006a,b, 2009; Waltzekt al., 2009; Fichi et al., 2013).
There is little evidence to suggest the presence of this virusn aquaculture systems in tropical countries like India. How-ver, bacterial disease outbreaks have been widely reported fromquaculture establishments, particularly due to motile aeromonadepticaemia involving Aeromonas hydrophila (Mohanty et al., 2008)nd other bacterial pathogens (Swann and White, 1991; Nielsent al., 2001; Rahman et al., 2002; Yesmin et al., 2004; Wahli et al.,005). Previously, Fichi et al. (2013) have reported the associa-ion of A. sorbia with CyHV-2 mortality in crucian carp C. carassiusrom Italy. This case report details a mortality event where CyHV-
was detected in association with multidrug-resistant (MDR) A.ydrophila infection in goldfish from commercial farms based onlinical signs, histology, electron microscopy, cell line infectiv-ty, experimental infection and PCR-based assay. It is not known
hether this virus infects other Cyprinids, or other ornamentalpecies being cultured with goldfish in the same ponds. The resultsbtained here, independently verified at two different labs, sug-est the complexity of the infection process and lay a foundationor further studies on the epidemiology, pathology and preventionf newly emerging freshwater viral diseases in India.
. Materials and methods
.1. Case history and analysis
Late in the rainy season in August 2013, large scale mortali-ies of goldfish, C. auratus and koi carp C. carpio in ornamentalsh farms of Hooghly district, West Bengal, India were reportedy the Fishery Extension Officer of the Department of Fisheries,overnment of West Bengal to the Faculty of Fishery Sciences
FFSc), West Bengal University of Animal and Fishery Sciences,olkata, India. The cumulative mortalities of goldfish and koi carp
n the affected ponds were in the range of 70–90%. Preliminaryacteriology and microscopy revealed the etiological agents toe motile Aeromonas spp. along with Microcystis intoxication. Noiral involvement was looked into during that outbreak period.reatment for bacterial infection was attempted with the use ofater sanitizer, benzalkonium chloride and enrofloxacin in the diet,hich led to partial control of mortality (Abraham, T.J., personal
ommunication). In early winter (November 2014), the ornamen-al fish farmers reported mass mortality of goldfish in few of the
onds in the same locality. In the affected farms, goldfish, koiarp and barbs were reared along with other Indian major carpsIMCs). However, no mortality was observed in the IMCs and barbs.he mortality rate was recorded at 1–5% daily in different gold-ica 161 (2016) 8–17 9
fish ponds, with a cumulative mortality above 90% during thisperiod in goldfish. The affected fish showed clinical signs such assluggish movement, erratic swimming, anorexia, spinning, fin rot,focal cutaneous haemorrhages, reddish lateral line, protrusion andsloughing off of scales, enophthalmia, peeled skin, corneal opac-ity, blind eye, etc. Bacteriological analysis of goldfish samples atFFSc, Kolkata revealed Aeromonas veronii biovar sobria from kid-ney and Flavobacterium sp. from the skin ulcers (Abraham, T.J.,personal communication). Water disinfection along with antibiotictreatment had no apparent effect on disease control this time, withincreased and continuous mortality. The authors here were calledin following a subsequent disease outbreak in goldfish from otherfarms in the same locality in mid-December 2014.
2.2. Fish samples
The ornamental fish farms where the outbreak occurred arelocated on both sides of and fed by sewage and waste water chan-nels draining into the River Hooghly. The details of geographicallocation of three farms from which samples were collected areas follows: Piyarapur/Chakpathakoria, Serampore Block, HooghlyDistrict, West Bengal, India; Latitude-22046′27.1′′N, Longitude-88017′47.0′′E; Latitude-22046′40′′N, Longitude-88018′6′′E andLatitude-22047′49.5′′N, Longitude-88018′1.8′′E. The ponds weremostly stocked with goldfish (C. auratus) along with few koi carp(C. carpio), barbs (Barbus barbus) and few IMCs, acquired as seedsfrom Howrah, West Bengal, India, at a density of 20,000–40,000numbers per acre. Only goldfish, approximately 3 months old,were affected, and symptoms included protrusion of scales, dropsy,enophthalmia, necrotized gills, haemorrhagic patches on the bodysurface particularly towards the base of the caudal fin, sluggishmovement and gulping of air followed by large scale mortality.The diseased fish were transported to the laboratories both on ice(n = 15) and alive, in bags with oxygen (n = 10). According to the lab-oratory standard operating procedures, live fish were euthanisedwith clove oil and all samples underwent necropsy, parasitologi-cal, bacteriological, histological and virological examinations. Forexperimental infection, apparently healthy goldfish, koi carp androhu carp (Labeo rohita) juveniles were obtained from fish farmslocated at Bhubaneswar, Odisha, India and Kochi, Kerala, India. Thefish were kept in 500 L capacity FRP tanks with aerated water at atemperature of 25–27 ◦C and fed with commercial pellet feed forone week of acclimatization before the challenge experiment.
2.3. Post mortem findings
All the affected fish showed gill necrosis, splenomegaly withlarge white granular nodules, pale liver, swollen kidneys with smallwhite focal necrosis and empty intestine.
2.4. Bacteriology and parasitology
Inocula from the blood, gill and kidney of infected goldfish wasstreaked on to tryptone soy agar (TSA, HiMedia, India) and singlecolonies obtained were purified and used for taxonomic analysisaccording to Bergey’s Manual of Determinative Bacteriology (Holtet al., 1994), with biochemical reactions performed following stan-dard protocols. Sequences of the 16s rRNA gene following PCR wereused to screen for the pathogen, following Nayak et al. (2013). Basedon the results, further PCR for aerolysin and beta-haemolysin genesspecific to A. hydrophila were carried out for reconfirmation fol-
lowing Mohanty et al. (2008). OmpTs and AHCYTOEN gene basedprimers were also used to determine whether the strain involvedwas pathogenic or not (Cagatay and Sen, 2014).The mucus, gill andviscera samples were also screened for the presence of parasites.
1 a Trop
2
c1atwcizCwt(
2
1thm
2
i0owuifct
2
sp0w4cUmsoe
2
n1Itagieuw
0 P.K. Sahoo et al. / Act
.5. Antibiogram
The resistance or sensitivity of the isolated bacteria to 31 antimi-robial agents was determined by disc diffusion assay (Bauer et al.,966). The bacterial isolate was grown overnight in a nutrient broth,nd 0.1 mL of the broth was spread onto TSA plates to make bac-erial lawns. Different antibiotic discs (Octo Disc, HiMedia, India)ere placed onto the plates and incubated at 30 ◦C for 24 h. The
lear zones around each antibiotic disc were measured. Sensitiv-ty and resistance to the antibiotics were determined based onone diameter, following the manufacturer’s instructions as perLSI standard. The classification as Multi Drug Resistant (MDR)as based on acquired non-susceptibility to at least one agent in
hree or more antimicrobial categories, following Magiorakos et al.2012).
.6. Histopathological examination
Tissues, including gills, kidney and spleen, were fixed in0% neutral buffered formalin solution. Samples were rou-inely processed and 5 �m-thick sections were stained withaematoxylin-eosin for further examination under a compoundicroscope.
.7. Experimental infection study with bacteria
Healthy goldfish juveniles (20 numbers) were injectedntraperitoneally with the isolated bacterium (5 × 106 cfu.05 mL−1 PBS fish−1) and another group of the same numberf fish received only PBS intraperitoneally as a control. The fishere observed for clinical signs and rate of mortality was recorded
p to 10 days post-challenge. Kidney tissue samples from threendividuals each of moribund fish from the infected group and fishrom control group were collected for the isolation, biochemicalharacterization and further confirmation of bacterium fromissues directly by PCR of aerolysin gene as described above.
.8. Transmission electron microscopy
For transmission electron microscopy (TEM) the affected gillamples from diseased fish were removed, dissected into smallieces (1 mm3), and rapidly immersed in 2.5% glutaraldehyde in.1 M phosphate buffer (pH 7.4) for 6–8 h at 4 ◦C. These wereashed and post fixed in 2% osmium tetroxide (OsO4) for 1 h at◦C. The tissue was dehydrated through a graded series of ethanol
oncentrations to 100% ethanol and embedded in araldite CY212.ltrathin sections (60–70 nm) of the gill tissues were cut using aicrotome (Leica ultracut UCT), mounted onto copper grids and
tained with uranyl acetate and alkaline lead citrate. Sections werebserved and photographed under a Tecnai T12 Spirit transmissionlectron microscope at 60 kV.
.9. Virus isolation on cell culture and its confirmation
Tissues, viz., gills, kidney and spleen (0.5 g of each from 10umbers of diseased goldfish, averaging about 9.0 cm length and5.5 g in weight) were collected in Leibovitz 15 medium (L-15,
nvitrogen) with 5% FBS and 2% v/v of antibiotic–antimycotic solu-ion (penicillin G 100 U mL−1, streptomycin sulphate 10 mg mL−1,mphotericin B 25 �g mL−1 and kanamycin 10 mg mL−1) (Invitro-en) and transported to the laboratory on ice and used for virus
solation. A cell line, Cyprinus carpio koi fin (CCKF) (Swaminathant al., 2015), derived from the fin of koi carp, C. carpio koi wassed for virus isolation. Fin cells were cultured in L-15 mediumith 10% FBS at 28 ◦C. Pooled tissue extracts from gills, kidneyica 161 (2016) 8–17
and spleen of the infected fish were aseptically prepared in a tis-sue homogenizer at 1/10 dilution in L-15 medium containing 2%FBS and supplemented with antibiotics (200 IU mL−1 penicillin,200 �g mL−1 streptomycin, and 0.5 �g mL−1 amphotericin B). Thetissue homogenate was centrifuged at 3000g for 10 min at 4 ◦C, andclarified supernatant was filtered through 0.22 �m syringe mem-brane filter (Millipore). 500 �L of the filtrate was inoculated ontoa confluent monolayer of CCKF cells in a 25 cm2 flask (Corning,USA). The same volume of L-15 cell culture medium was used for amock infection in CCKF cells as control. After adsorption for 1 h at28 ◦C, 5 mL of the medium supplemented with 2% FBS was addedto the flasks and incubated at 28 ◦C in low temperature incubator.The infected cells were examined daily under an inverted phasecontrast microscope (TE2000, Nikon, Japan). Once the CPE was com-plete, the CCKF cell culture supernatant was collected, clarified andfiltered through 0.22 �m syringe filter, and DNA was isolated usingQiagen kit for confirmation of viral agent by PCR. The cell culturesupernatant with 80–90% CPE was centrifuged at 3000g for 15 minand stored in aliquots at −80 ◦C for further use.
2.10. Polymerase chain reaction assay and sequence analysis
Different tissues (liver, gill, kidney, spleen, muscle, and heart)of infected goldfish from the field samples were processed for DNAisolation. Tissue samples (approximately 100 mg) were treatedwith proteinase K in lysis buffer (50 mM Tris/HCl, 100 mM NaCl,100 mM EDTA, 1% [w/v] SDS, pH 8.0) and subjected to extractionwith phenol/chloroform/isoamyl alcohol, followed by ethanol pre-cipitation. The DNA was diluted in TE (50 mM Tris/HCl, 1 mM EDTA,pH 7.5). Concentration and purity of the extracted DNA was deter-mined by measuring OD at 260 and 280 nm using a NanoDropND1000 spectrophotometer (Nano Drop Technologies Inc., USA).The samples were stored at −20 ◦C for further analysis. PCR wasperformed using three sets of published oligonucleotide primersfor confirmation. In the first set, CyHVpol-FOR and CyHVpol-REVprimers (Jeffery et al., 2007) were used in a final volume of 25 �Lcontaining 1 �L of total DNA, 1.5 �L (10 pmol) of each primer,0.25 �L of Taq DNA polymerase (5 U �L−1), 2.5 �L of 10X Taq bufferA, 0.5 �L of dNTPs (2 mM) and ddH2O to make final volume to25 �L. Amplification included the following steps: denaturation at95 ◦C for 2 min, followed by 40 cycles of denaturation at 95 ◦C for1 min, annealing at 55 ◦C for 1 min, extension at 72 ◦C for 1 minand final extension at 72 ◦C for 10 min. In the second set, oligonu-cleotide primers, CyHVGW-FOR and CyHVGW-REV (Goodwin et al.,2006b) were used. The reaction mixture for PCR consisted of 5 �Lof template, 0.5 �L of each primer (10 pmol), 0.5 �L 2 mM dNTPs,2.5 �L 10 X buffer A, 0.25 �L Taq polymerase mix (5 U �L−1) and15.75 �L PCR water. The reaction conditions were 95 ◦C for 2 minfor one cycle followed by 95 ◦C for 30 s, 58 ◦C for 30 s, and 72 ◦C for45 s and final extension 72 ◦C for 2 min for 35 cycles. In the thirdset, CyHV helicase primers, CyHV-2HelF and CyHV-2HelR (Waltzeket al., 2009) were used. The 25 �L PCR reaction mixture consisted of2.5 �L 10X Taq buffer A, 0.5 �L (10 �mol L−1) of each of the primers,1 �L of template, 0.25 �L Taq polymerase, 0.5 �L of dNTPs (2 mM)and 19.75 �L ddH2O. The PCR was conducted as follows: denatur-ing at 95 ◦C for 2 min; followed by 35 cycles at 95 ◦C for 30 s, 60 ◦Cfor 45 s, and 72 ◦C for 30 s; and final extension at 72 ◦C for 2 min.In addition, samples were further screened for CyHV-3 using OIErecommended Gray Sph primers. The primer details are given inTable 1. The PCR products were checked in 1.5% agarose gel. Theamplicons obtained (three from each product) were purified usinga gel extraction kit (Genei, India). The obtained fragments were
ligated to T-vector and cloned into Escherichia coli DH5� bacterialstrain using InsTAclone PCR Cloning Kit (Thermo Scientific, USA)according to manufacturer’s instructions. Three clones were con-firmed by sequencing (Xcelris, India) for each fragment. Sequence
P.K. Sahoo et al. / Acta Tropica 161 (2016) 8–17 11
Table 1Details of the primers used in this study.
Sl No Primer Nucleotide base sequence (5′–3′) Amplicon size(bp)
1a CyHVpol-FOR CCCAGCAACATGTGCGACGG 3621b CyHVpol-REV CCGTARTGAGAGTTGGCGCA
2a CyHV-2 GW-FOR TCGGTTGGACTCGGTTTGTG 932b CyHV-2 GW-REV CTCGGTCTTGATGCGTTTCTTG
3a CyHV-2HelF GGACTTGCGAAGAGTTTGATTTCTAC 3663b CyHV-2HelR CCATAGTCACCATCGTCTCATC
4a CyHV-3Gray Sph- FOR GACACCACATCTGCAAGGAG 2924b CyHV-3Gray Sph- REV GACACATGTTACAATGGTCGC
5a CCBA-2f TTGGCAATGAGAGGTTCAGGT 1395b CCBA-2r TTGGCATACAGGTCCTTACGG
6a 16S-f AGAGTTTGATCATGGCTCAG 15006b 16S-r GGTTACCTTGTTACGACTT
7a AhBhaeF GCTATGAAAAAACTAAAAATAACTG 15887b AhBhaeR CAGTATAAGTGGGGAAATGGAAAG
8a AeroF F-TGTCGGSGATGACATGGAYGTG 7208b AeroR R-CCAGTTCCAGTCCCACCACTTCA
9a AHCYOENf GAGAAGGTGACCACCAAGAACAA 2329b AHCYTOENr AACTGACATCGGCCTTGAACTC
GCATTA
asss
2
ifid(aEeTfam1ecc
2c
gm5fi−solwww
10a OmpTS-f
10b OmpTS-r
lignment was done in ClustalW as implemented in BioEdit ver-ion 7.0.0. Phylogenetic analysis based on partial polymerase geneequence obtained here was carried out using other Cyprinivirusequences.
.11. Real-time PCR
As a viral disease outbreak was suspected, for the first timen the country, multiple diagnostic tests were performed to con-rm our diagnosis. Real-time PCR was carried out using primersetailed in Goodwin et al. (2006b) in a Light Cycler 96 SW 1.1Roche, Germany). Briefly, 1 �L of DNA was used as template in
total reaction mixture of 10 �L containing 5 �L of 2X Fast Startssential DNA Green Master (Roche, Germany), 0.5 �L (5 pmole) ofach primer (Table 1) and 3 �L of PCR grade H2O provided in the kit.he real-time PCR programme included a pre-denaturation at 95 ◦Cor 2 min and 35 cycles of amplification at 45 s at 58 ◦C, 45 s at 72 ◦C,nd 30 s at 95 ◦C and a final extension of 2 min at 72 ◦C followed byelt curve analysis at of 95 ◦C for 10 s, 65 ◦C for 1 min and 95 ◦C for
min. Housekeeping gene �-actin was run as a calibrator (Sahoot al., 2013). Each sample was taken in duplicate and no templateontrols were run each time. Tm analysis was also performed toheck primer specificity.
.12. Experimental infection with suspected virus to goldfish, koiarp and rohu carp
Gill, spleen, liver and kidney tissues obtained from diseasedoldfish were homogenized in phosphate buffer saline (PBS) toake a 10% (w/v) suspension. The homogenate was centrifuged at
000 rpm for 5 min at 4 ◦C. The supernatant was filtered (0.22 �mlter) and the suspension (inoculum) stored in a sterile tube at20 ◦C until required. Healthy goldfish of weight 12–14 g (pre-
creened, PCR negative to CyHV-2) were divided into three groupsf 9 fish each (Trial I). Group 1 was injected with 0.1 mL of inocu-
um intraperitoneally and group 2 was challenged by immersionith viral inoculum (0.1 mL L−1 of water) for one hour. Group 3as injected with 0.1 mL sterile PBS and served as control. The fishere observed for any mortality for ten days and tissues were col-
GTGGTTTATGACAAAGAC 1008GAAGTTGTATTGCAGGGC
lected from euthanized or dead fish for PCR screening as describedbelow.
In another laboratory (Trial II), the tissues transported in tissueculture medium with antibiotics on ice were thawed and homog-enized in a sterile homogenizer and a 10% (w/v) suspension wasmade with L-15 medium without antibiotics. The homogenate wascentrifuged at 4000g for 20 min at 4 ◦C, and the supernatant was re-centrifuged at 13,000 g for 20 min at 4 ◦C. The final supernatant wasfiltered through a 0.22 �m membrane filter. The filtrate was thenstored at −80 ◦C for infectivity studies. Healthy juvenile goldfish,of average weight 9.0 g were maintained in laboratory conditionsfor 7 days in well-aerated glass aquaria of 100-L capacity and fedwith commercial pellet feed twice daily. Following acclimatiza-tion, the fish were split into two groups of 10 fish each in aerated40 L aquaria. Group A received intraperitoneal injections of 50 �Lof tissue homogenate. The fish of group B were intraperitoneallyinjected with 50 �L of L-15 medium and served as control. Thefish were maintained under observation for 15 days and fed ad libi-tum with commercial pellet feed. Detection and isolation of thevirus from dead and infected fish was attempted by inoculatingtissue homogenates of pooled tissue samples onto CCKF cells. Chal-lenge experiments on healthy koi carp (weighing 12 g) were alsocarried out. The virus suspension in L-15 medium (100 �L) wasinjected intraperitoneally into koi carp (n = 10). The control fish(n = 10) received 100 �L of L-15 medium. The infectivity patternwas observed for 10 days.
To investigate the susceptibility of co-cultured IMCs to this viralpathogen, rohu (L. rohita) weighing average 50 g each were dividedinto three groups of 10 fish each. The first group was challenged asdescribed earlier with an intraperitoneal injection of 0.2 mL viralsuspension in PBS. The second group was co-habited with threeacutely infected (CyHV-2 tested positive from gill samples) gold-fish. The control group received only 0.2 mL PBS intraperitoneally.Tissues viz., gill, liver, spleen and kidney were collected from twoeuthanised fish from each group at 2, 5, 10 and 15 days post-
infection (dpi). The tissues were processed for PCR analysis to detectthe presence of virus.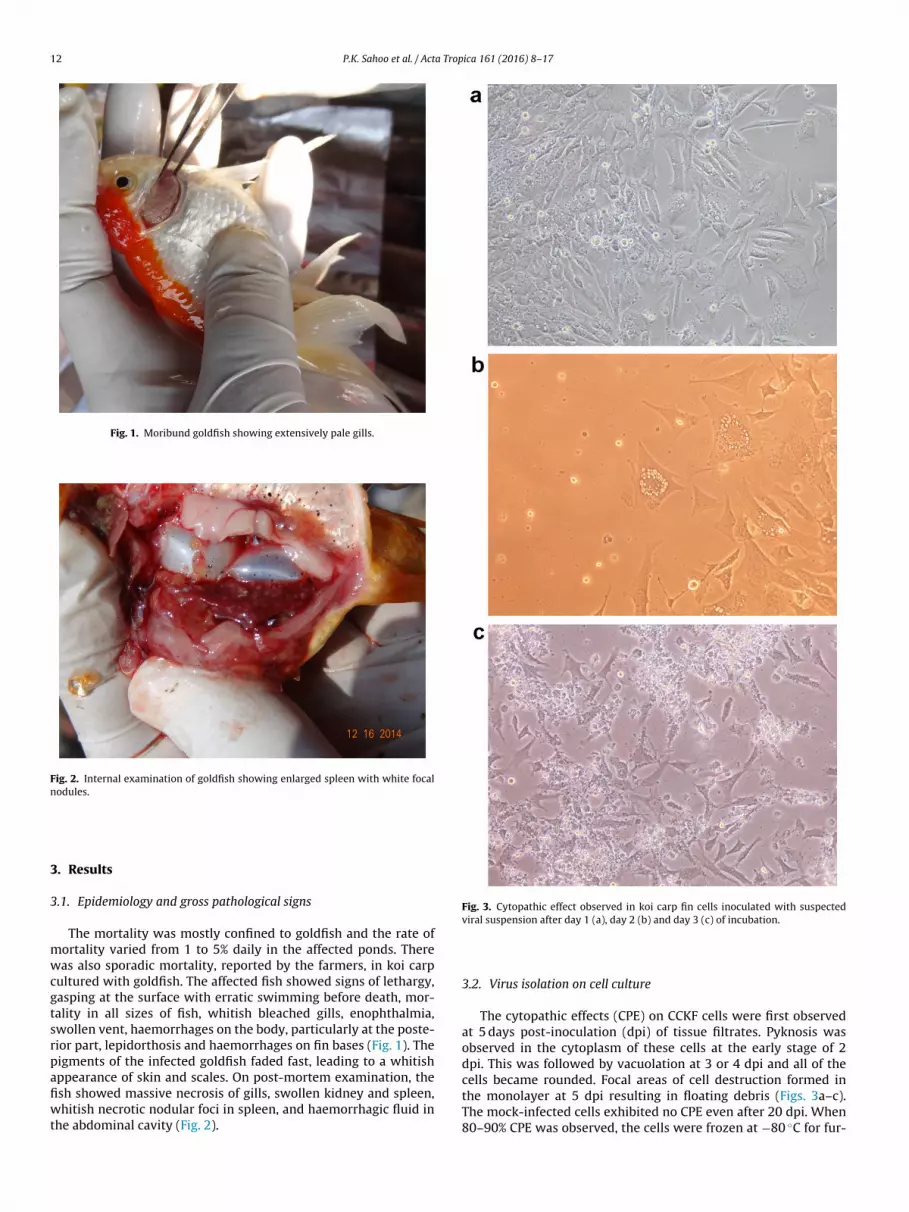
12 P.K. Sahoo et al. / Acta Tropica 161 (2016) 8–17
Fig. 1. Moribund goldfish showing extensively pale gills.
Fig. 2. Internal examination of goldfish showing enlarged spleen with white focalnodules.
3
3
mwcgtsrpafiwt
Fig. 3. Cytopathic effect observed in koi carp fin cells inoculated with suspected
The mock-infected cells exhibited no CPE even after 20 dpi. When80–90% CPE was observed, the cells were frozen at −80 ◦C for fur-
. Results
.1. Epidemiology and gross pathological signs
The mortality was mostly confined to goldfish and the rate ofortality varied from 1 to 5% daily in the affected ponds. Thereas also sporadic mortality, reported by the farmers, in koi carp
ultured with goldfish. The affected fish showed signs of lethargy,asping at the surface with erratic swimming before death, mor-ality in all sizes of fish, whitish bleached gills, enophthalmia,wollen vent, haemorrhages on the body, particularly at the poste-ior part, lepidorthosis and haemorrhages on fin bases (Fig. 1). Theigments of the infected goldfish faded fast, leading to a whitishppearance of skin and scales. On post-mortem examination, thesh showed massive necrosis of gills, swollen kidney and spleen,hitish necrotic nodular foci in spleen, and haemorrhagic fluid in
he abdominal cavity (Fig. 2).
viral suspension after day 1 (a), day 2 (b) and day 3 (c) of incubation.
3.2. Virus isolation on cell culture
The cytopathic effects (CPE) on CCKF cells were first observedat 5 days post-inoculation (dpi) of tissue filtrates. Pyknosis wasobserved in the cytoplasm of these cells at the early stage of 2dpi. This was followed by vacuolation at 3 or 4 dpi and all of thecells became rounded. Focal areas of cell destruction formed inthe monolayer at 5 dpi resulting in floating debris (Figs. 3a–c).

P.K. Sahoo et al. / Acta Tropica 161 (2016) 8–17 13
Table 2Physical and biochemical characteristics of the isolated bacterium.
Characteristics Reaction Characteristics Reaction
Gram stain − Acid production fromMotility + Glucose +Cytochrome oxidase + Fructose +Triple sugar iron agar A/A Galactose +Gelatin liquefaction + Raffinose +Methyl red − Mannose +Catalase + Trehalose −Growth on TSA + Cellobiose −Growth on RSA + (yellow colony) Dulcitol −Citrate + Xylose −Indole + Inulin −Voges Proskauer reaction + Salicin −
tb
3
otvicmw
3
ttpwfsosh
FgeS
Table 3Antibiogram of A. hydrophila strain isolated from diseased C. auratus.
Name of the Antibiotics Disc Concentration Resistant(R)/IntermediateSensitive (IS)/Sensitive(S)
Amoxyclav (Ac) 30 mcg RAmpicillin (As) 10 mcg RAzetreonam (Ao) 30 mcg RCeftazidime (Ca) 30 mcg RCefuroxime (Cu) 30 mcg RCephalothin (Ch) 30 mcg RCephotaxime (Ce) 30 mcg RCloxacillin (Cx) 1 mcg RImipenam (I) 10 mcg RLincomycin (L) 2 mcg RMethicillin (M) 5 mcg ROxacillin (Ox) 1 mcg RPenicillin G (P) 10 unit RSulphatriad (S3) 300 mcg RVancomycin (Va) 30 mcg RCefaperazone (Cs) 75 mcg ISCo-trimoxazole (Co) 25 mcg ISErythromycin (E) 15 mcg ISAmikacin (Ak) 30 mcg SChloramphenical (C) 25 mcg SCiprofloxacin (Cf) 5 mcg SColistin (Cl) 10 mcg SDoxycycline (Do) 30 mcg SGentamicin (G) 10 mcg SLevofloxacin (Le) 5 mcg SNorfloxacin (Nx) 10 mcg SOxytetracycline (O) 30 mcg SPiperacillin (Pc) 100 mcg S
Congo red binding + Inositol −Arabinose −
her use. The virus caused CPE in CCKF cells at the early passages,ut not beyond the fourth passage.
.3. Transmission electron microscopy
Large numbers of enveloped, icosahedral virus particles werebserved in the transmission electron micrographs of the ultra-hin sections of gills. Characteristic cytoplasmic aggregations ofirus particles in special membranous envelopes were observedn the cell cytoplasm. Mature virus particles were observed in theytoplasm. The size of the nucleocapsid of the virus was approxi-ately 90–120 nm in diameter and envelope of the mature virionas 170–200 nm (Fig. 4).
.4. PCR
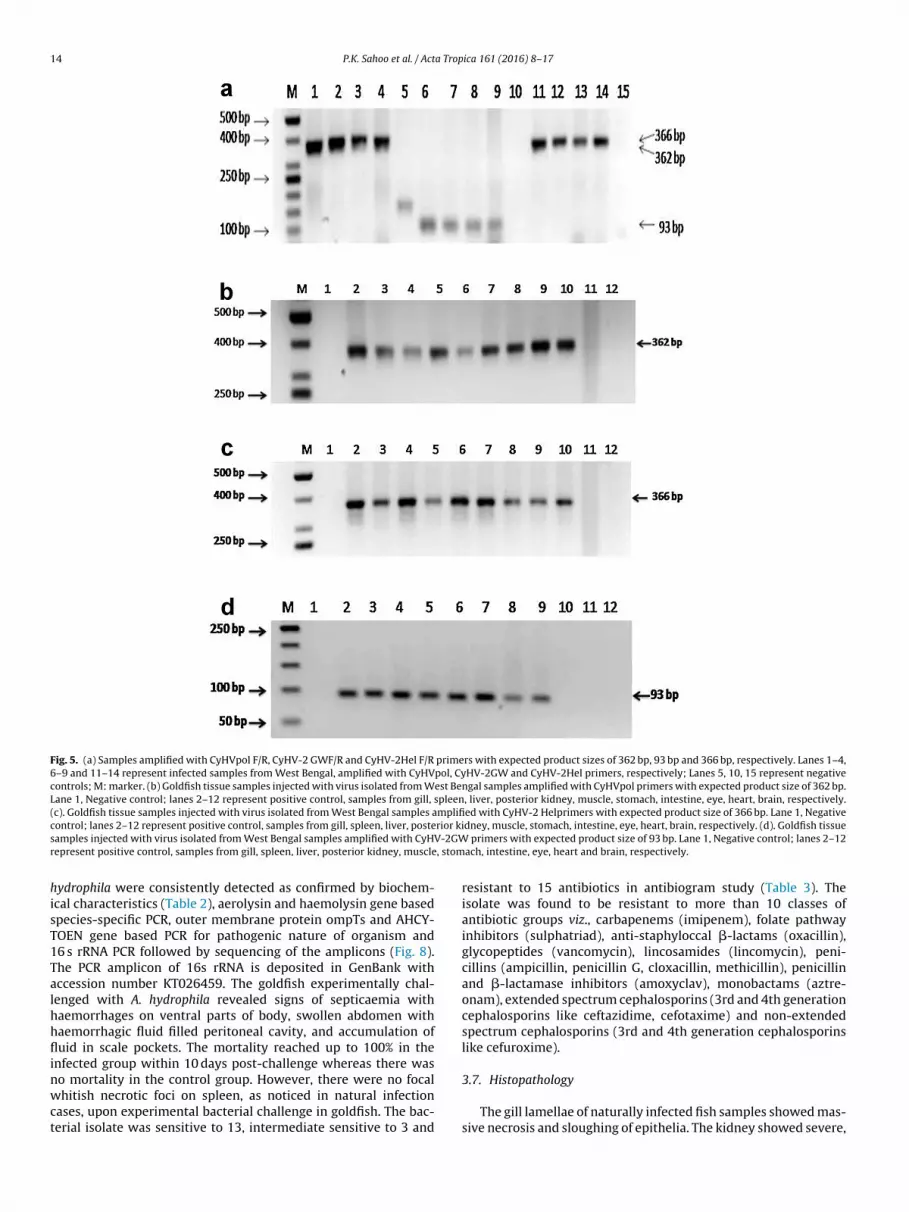
Tissue samples collected from natural infection, CCKF cell cul-ure suspension, and experimentally challenged fish were foundo be positive for CyHV-2 in PCR assay carried out using all threerimer sets with expected amplicon sizes of 362, 93 and 366 bpith polymerase, GW and helicase primers (Fig. 5a). Real-time PCR
urther confirmed the presence of amplicons in all the suspected
amples (Cq values ranging from 30.78 to 33.68). The productsbtained were cloned and sequenced and a BLAST search of theequence revealed 100% identity with known previously publishedelicase and polymerase gene sequences. The polymerase geneig. 4. Transmission electron microscopy (TEM) of tissues processed from diseasedoldfish, Carassius auratus infected with CyHV-2. Viral capsids in cytoplasm withlectron-dense cores and capsid surrounded by electron-dense material (arrows).cale bar = 200 nm.
Streptomycin (S) 10 mcg STetracycline (T) 25 mcg STobramycin (Tb) 10 mcg S
sequence has been submitted to GenBank (GenBank accessionnumber: KT900136). A phylogeny tree based on partial polymerasegene sequence was constructed (Fig. 6) and our isolate clusteredwith all CyHV-2 isolates forming a separate branch differing fromother Cyprinid herpesviruses groups. All the samples were foundto be negative for CyHV-3.
3.5. Experimental infection with virus suspension
The goldfish infected with virus suspension showed typical signsof gill necrosis within 48 h of post challenge in intraperitonealgroup and within 72 h in immersion group (Fig. 7). The mortal-ity reached up to 67% in intraperitoneal group within 10 days ofchallenge in one laboratory. Goldfish inoculated with 0.1 mL tis-sue homogenate filtrate began to die at 1 dpi, death rate increasedrapidly from 1 to 4 dpi and the death rate reached 100% at 10 dpi.The dead fish presented the same hemorrhagic signs as naturallyinfected fish. There was no mortality in mock-infection group. PCRdetections of dead fish infected with tissue homogenate filtratewere all positive, while the healthy fish of mock-infection grouptested negative at both laboratories. All the tissues (liver, mus-cle, spleen, kidney, eye, stomach and gill) except brain and heartrevealed positive amplicons in PCR with all three primer sets (Figs.5b–d). Rohu and koi carp challenged experimentally with viral sus-pension did not develop any infection or show any mortality ina 15-day trial. PCR analysis of samples collected at different timepoints from the injected animals failed to generate amplicons ofexpected size in all the tissues using helicase gene primers.
3.6. Bacteriology and parasitology
There was no evidence of parasitic infection in the exam-ined fish. However, from all the cases examined, pathogenic A.
14 P.K. Sahoo et al. / Acta Tropica 161 (2016) 8–17
Fig. 5. (a) Samples amplified with CyHVpol F/R, CyHV-2 GWF/R and CyHV-2Hel F/R primers with expected product sizes of 362 bp, 93 bp and 366 bp, respectively. Lanes 1–4,6–9 and 11–14 represent infected samples from West Bengal, amplified with CyHVpol, CyHV-2GW and CyHV-2Hel primers, respectively; Lanes 5, 10, 15 represent negativecontrols; M: marker. (b) Goldfish tissue samples injected with virus isolated from West Bengal samples amplified with CyHVpol primers with expected product size of 362 bp.Lane 1, Negative control; lanes 2–12 represent positive control, samples from gill, spleen, liver, posterior kidney, muscle, stomach, intestine, eye, heart, brain, respectively.(c). Goldfish tissue samples injected with virus isolated from West Bengal samples amplified with CyHV-2 Helprimers with expected product size of 366 bp. Lane 1, Negativec rior kis V-2GWr , stom
hisT1Talhhflinwct
ontrol; lanes 2–12 represent positive control, samples from gill, spleen, liver, posteamples injected with virus isolated from West Bengal samples amplified with CyHepresent positive control, samples from gill, spleen, liver, posterior kidney, muscle
ydrophila were consistently detected as confirmed by biochem-cal characteristics (Table 2), aerolysin and haemolysin gene basedpecies-specific PCR, outer membrane protein ompTs and AHCY-OEN gene based PCR for pathogenic nature of organism and6 s rRNA PCR followed by sequencing of the amplicons (Fig. 8).he PCR amplicon of 16s rRNA is deposited in GenBank withccession number KT026459. The goldfish experimentally chal-enged with A. hydrophila revealed signs of septicaemia withaemorrhages on ventral parts of body, swollen abdomen withaemorrhagic fluid filled peritoneal cavity, and accumulation ofuid in scale pockets. The mortality reached up to 100% in the
nfected group within 10 days post-challenge whereas there was
o mortality in the control group. However, there were no focalhitish necrotic foci on spleen, as noticed in natural infectionases, upon experimental bacterial challenge in goldfish. The bac-erial isolate was sensitive to 13, intermediate sensitive to 3 and
dney, muscle, stomach, intestine, eye, heart, brain, respectively. (d). Goldfish tissue primers with expected product size of 93 bp. Lane 1, Negative control; lanes 2–12
ach, intestine, eye, heart and brain, respectively.
resistant to 15 antibiotics in antibiogram study (Table 3). Theisolate was found to be resistant to more than 10 classes ofantibiotic groups viz., carbapenems (imipenem), folate pathwayinhibitors (sulphatriad), anti-staphyloccal �-lactams (oxacillin),glycopeptides (vancomycin), lincosamides (lincomycin), peni-cillins (ampicillin, penicillin G, cloxacillin, methicillin), penicillinand �-lactamase inhibitors (amoxyclav), monobactams (aztre-onam), extended spectrum cephalosporins (3rd and 4th generationcephalosporins like ceftazidime, cefotaxime) and non-extendedspectrum cephalosporins (3rd and 4th generation cephalosporinslike cefuroxime).
3.7. Histopathology
The gill lamellae of naturally infected fish samples showed mas-sive necrosis and sloughing of epithelia. The kidney showed severe,

P.K. Sahoo et al. / Acta Tropica 161 (2016) 8–17 15
Fig. 6. Phylogenetic tree based on the nucleotide sequence of herpesvirus homologues. The tree was constructed using neighbor-joining method of ClustalW2. 1. KC841411.1:Cyprinid herpesvirus 2 isolate BY-JS1 DNA polymerase catalytic subunit gene, partial cds; 2. JQ740766.1: Cyprinid herpesvirus 2 isolate CZ1736-3 DNA-dependent DNApolymerase gene, partial cds; 3. JQ740765.1: Cyprinid herpesvirus 2 isolate CZ1736-2 DNA-dependent DNA polymerase gene, partial cds; 4. JQ740764.1: Cyprinid herpesvirus2 isolate CZ1736-1 DNA-dependent DNA polymerase gene, partial cds; 5. HM014349.1: Cyprinid herpesvirus 2 DNA-dependent DNA polymerase gene, partial cds; 6.AY939863.1: Cyprinid herpesvirus 2 DNA polymerase gene, partial cds; 7. DQ085628.1: Cyprinid herpesvirus 2 DNA polymerase gene, partial cds; 8. DQ085627.1: Cyprinidherpesvirus 2 DNA polymerase gene, partial cds; 9. DQ085626.1: Cyprinid herpesvirus 2 DNA polymerase gene, partial cds; 10. KP976110.1: Cyprinid herpesvirus 2 isolateE erpesvirus DNA polymerase gene, complete cds; 12. AY939868.1: AY939868.1: Cyprinidh esvirus 1 DNA-dependent DNA polymerase gene, partial cds; 14. GU233800.1: Anguillidh
dshTsg
4
C1Jnhw(tna(hJia
ni(h(elaCed
S3060 DNA polymerase catalytic subunit gene, partial cds; 11. AY939862.1: Koi herpesvirus 1 DNA polymerase gene, complete cds; 13. HM014350.1: Cyprinid herperpesvirus 1 isolate C3P2 DNA polymerase gene, complete cds.
iffuse necrosis of the haematopoietic tissue. The lesions in thepleen were characterized by necrosis of the lymphoid tissue, andypertrophied nuclei with marginated chromatin material (Fig. 9).he bacteria infected fish in the experimental study revealed necro-is of spleen, kidney and liver tissues mostly with mild necrosis ofill epithelia.
. Discussion
The disease outbreak noticed here revealed many similarities toyHV-2 infection described earlier in goldfish (Jung and Miyazaki,995; Groff et al., 1998; Stephens et al., 2004; Goodwin et al., 2006b;
effery et al., 2007; Wu et al., 2013). The gross signs, typical whitishodular necrotic foci on enlarged spleen, enlarged kidney, massiveaemorrhages on body and sero-sanguinous peritoneal fluid alongith signs of mortality were also reported by earlier investigators
Jung and Miyazaki, 1995; Goodwin et al., 2006b, 2009). A charac-eristic leaching out of pigments before death observed here wasot noticed earlier and needs further investigation. The bleachedppearance of the gills as noticed here was also reported earlierJeffery et al., 2007), as was the margination of chromatin withypertrophic nuclei in spleen and kidney (Goodwin et al., 2006b;
effery et al., 2007). This study is the first detection of the CyHV-2n Indian aquaculture and also reports the first natural outbreak ofny virus infection in freshwater aquaculture system in India.
Several cases of mortality due to Herpesvirus haematopoieticecrosis of goldfish have been reported since its first record in Japan
n 1992 and its subsequent nomenclature as Cyprinid herpesvirus-2Goodwin et al., 2006b, 2009). In most cases, the diagnosis of HVHNas been based on histological findings and electron microscopyGoodwin et al., 2006a,b; Philbey, 2006) followed by PCR (Goodwint al., 2006b; Jeffery et al., 2007). In this case report, herpesvirus-ike particles were found in the tissues of fish by TEM observation
nd the DNA sequencing of three genes’ PCR product confirmedyHV-2 presence. CPE was noticed in CCKF cells inoculated withxtracts of infected fish tissue homogenates. From the screening ofifferent tissue samples from experimentally infected animals, itFig. 7. Immersion challenged goldfish showing whitish pale patches on gills on 2ndday post-challenge.
was noticed that heart and brain carried poor viral load and hencethe value of these organs as diagnostic materials must be appraised.On the contrary, Goodwin et al. (2006b) found brain and other tis-sues including spleen and trunk kidney suitable for viral isolationand screening.
OIE (World Organization for Animal Health) recommendsin vitro primary culture of cells infected by candidate pathogen asan accurate diagnostic method for viral pathogens. In this study,CCKF cells showed typical CPE within 5 days of infection. How-ever, CPE could not be detected after fourth passage. Earlier studieshave also indicated that CyHV-2 is difficult to grow in tissue cul-ture and is lost after several passages in vitro (Waltzek et al., 2005;Goodwin et al., 2006b). It has also been suggested that explorationof haematopoietic cells is the key to establish an effective cell line
for CyHV-2 isolation and propagation (Wu et al., 2013). The struc-tural characteristic of the viral particles were also consistent withthe characteristics of herpesviruses described in other reports (Xuet al., 2013; Lovy and Friend, 2014).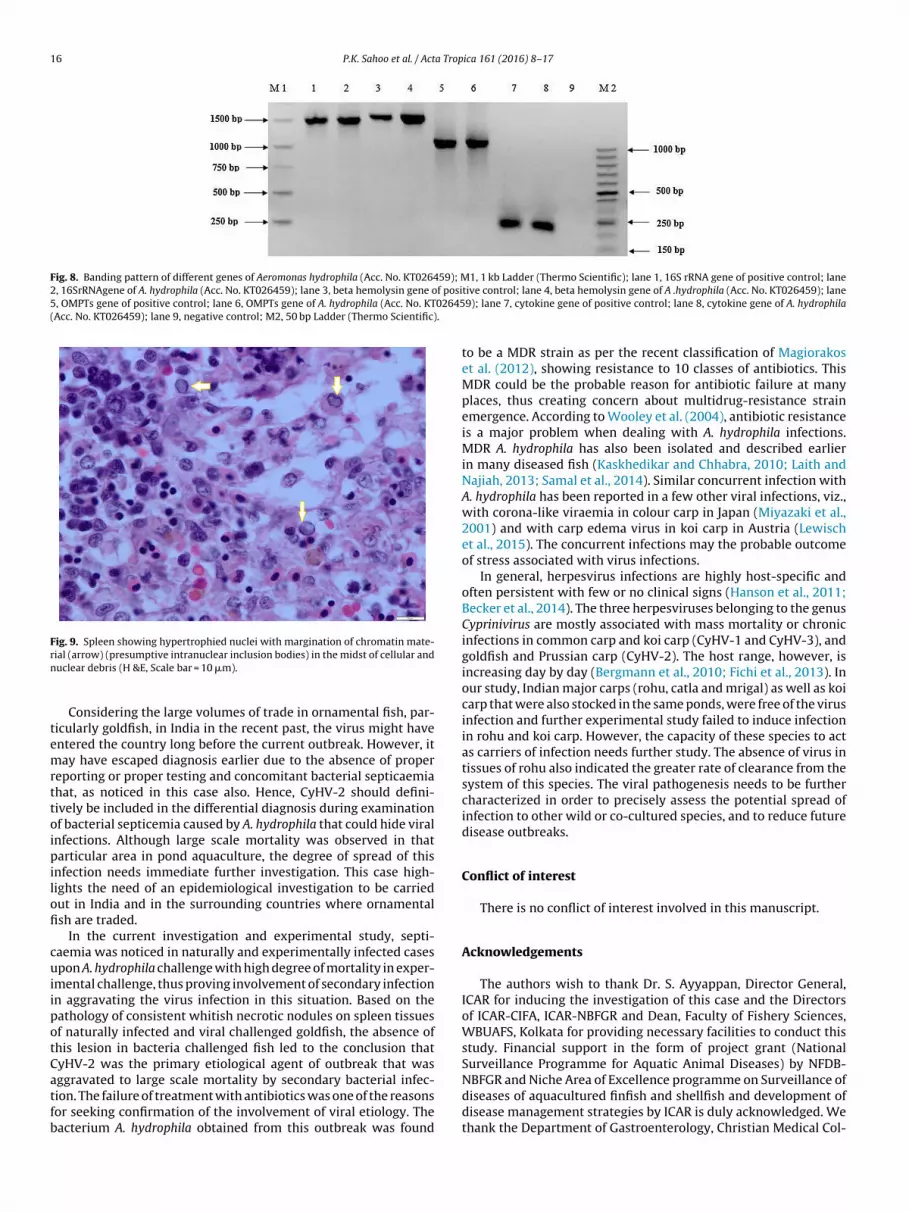
16 P.K. Sahoo et al. / Acta Tropica 161 (2016) 8–17
Fig. 8. Banding pattern of different genes of Aeromonas hydrophila (Acc. No. KT026459); M1, 1 kb Ladder (Thermo Scientific); lane 1, 16S rRNA gene of positive control; lane2, 16SrRNAgene of A. hydrophila (Acc. No. KT026459); lane 3, beta hemolysin gene of posi5, OMPTs gene of positive control; lane 6, OMPTs gene of A. hydrophila (Acc. No. KT02645(Acc. No. KT026459); lane 9, negative control; M2, 50 bp Ladder (Thermo Scientific).
Fig. 9. Spleen showing hypertrophied nuclei with margination of chromatin mate-rn
temrttoipilofi
cuiipotCatfb
NBFGR and Niche Area of Excellence programme on Surveillance of
ial (arrow) (presumptive intranuclear inclusion bodies) in the midst of cellular anduclear debris (H &E, Scale bar = 10 �m).
Considering the large volumes of trade in ornamental fish, par-icularly goldfish, in India in the recent past, the virus might haventered the country long before the current outbreak. However, itay have escaped diagnosis earlier due to the absence of proper
eporting or proper testing and concomitant bacterial septicaemiahat, as noticed in this case also. Hence, CyHV-2 should defini-ively be included in the differential diagnosis during examinationf bacterial septicemia caused by A. hydrophila that could hide viral
nfections. Although large scale mortality was observed in thatarticular area in pond aquaculture, the degree of spread of this
nfection needs immediate further investigation. This case high-ights the need of an epidemiological investigation to be carriedut in India and in the surrounding countries where ornamentalsh are traded.
In the current investigation and experimental study, septi-aemia was noticed in naturally and experimentally infected casespon A. hydrophila challenge with high degree of mortality in exper-
mental challenge, thus proving involvement of secondary infectionn aggravating the virus infection in this situation. Based on theathology of consistent whitish necrotic nodules on spleen tissuesf naturally infected and viral challenged goldfish, the absence ofhis lesion in bacteria challenged fish led to the conclusion thatyHV-2 was the primary etiological agent of outbreak that wasggravated to large scale mortality by secondary bacterial infec-
ion. The failure of treatment with antibiotics was one of the reasonsor seeking confirmation of the involvement of viral etiology. Theacterium A. hydrophila obtained from this outbreak was foundtive control; lane 4, beta hemolysin gene of A .hydrophila (Acc. No. KT026459); lane9); lane 7, cytokine gene of positive control; lane 8, cytokine gene of A. hydrophila
to be a MDR strain as per the recent classification of Magiorakoset al. (2012), showing resistance to 10 classes of antibiotics. ThisMDR could be the probable reason for antibiotic failure at manyplaces, thus creating concern about multidrug-resistance strainemergence. According to Wooley et al. (2004), antibiotic resistanceis a major problem when dealing with A. hydrophila infections.MDR A. hydrophila has also been isolated and described earlierin many diseased fish (Kaskhedikar and Chhabra, 2010; Laith andNajiah, 2013; Samal et al., 2014). Similar concurrent infection withA. hydrophila has been reported in a few other viral infections, viz.,with corona-like viraemia in colour carp in Japan (Miyazaki et al.,2001) and with carp edema virus in koi carp in Austria (Lewischet al., 2015). The concurrent infections may the probable outcomeof stress associated with virus infections.
In general, herpesvirus infections are highly host-specific andoften persistent with few or no clinical signs (Hanson et al., 2011;Becker et al., 2014). The three herpesviruses belonging to the genusCyprinivirus are mostly associated with mass mortality or chronicinfections in common carp and koi carp (CyHV-1 and CyHV-3), andgoldfish and Prussian carp (CyHV-2). The host range, however, isincreasing day by day (Bergmann et al., 2010; Fichi et al., 2013). Inour study, Indian major carps (rohu, catla and mrigal) as well as koicarp that were also stocked in the same ponds, were free of the virusinfection and further experimental study failed to induce infectionin rohu and koi carp. However, the capacity of these species to actas carriers of infection needs further study. The absence of virus intissues of rohu also indicated the greater rate of clearance from thesystem of this species. The viral pathogenesis needs to be furthercharacterized in order to precisely assess the potential spread ofinfection to other wild or co-cultured species, and to reduce futuredisease outbreaks.
Conflict of interest
There is no conflict of interest involved in this manuscript.
Acknowledgements
The authors wish to thank Dr. S. Ayyappan, Director General,ICAR for inducing the investigation of this case and the Directorsof ICAR-CIFA, ICAR-NBFGR and Dean, Faculty of Fishery Sciences,WBUAFS, Kolkata for providing necessary facilities to conduct thisstudy. Financial support in the form of project grant (NationalSurveillance Programme for Aquatic Animal Diseases) by NFDB-
diseases of aquacultured finfish and shellfish and development ofdisease management strategies by ICAR is duly acknowledged. Wethank the Department of Gastroenterology, Christian Medical Col-

a Trop
lK
R
B
B
B
B
B
C
F
G
G
G
G
G
H
H
I
J
J
K
L
L
L
infection emerged in cultured gibel carp, Carassius auratus gibelio in China. Vet.Microbiol. 166, 138–144.
Yesmin, S., Rahmann, M.H., Hussain, M.A., Khan, A.R., Pervin, F., Hossain, M.A.,2004. Aeromonas hydrophila infection of swamps in Bangladesh. Pakistan J.
P.K. Sahoo et al. / Act
ege, Vellore, India for the TEM analysis of the samples and Mr R.G.umar, PMFGRC, Kochi for critically going through the manuscript.
eferences
auer, A.W., Kirby, W.M.M., Sherris, J.S., Turck, M., 1966. Antibiotic susceptibilitytesting by a standard single disc method. Am. J. Clin. Pathol. 45, 493–496.
ecker, J.A., Tweedie, A., Rimmer, A., Landos, M., Lintermans, M., Whittington, R.J.,2014. Incursions of Cyprinid herpesvirus 2 in goldfish populations in Australiadespite quarantine practices. Aquaculture 432, 53–59.
ergmann, S.M., Sadowski, J., Kielpiski, M., Barilomiejczyk, M., Fichtner, D., Riebe,R., et al., 2010. Susceptibility of koi x crucian carp and koi x goldfish hybrids tokoi herpesvirus (KHV) and the development of KHV disease (KHVD). J. Fish Dis.33, 267–272.
inesh, C.P., 2013. Mortality due to viral nervous necrosis in zebrafish Danio rerioand goldfish Carassius auratus. Dis. Aquat. Org. 104, 257–260.
oitard, P.M., Baud, M., Labrut, S., de Boisseson, C., Jamin, M., Bigarre, L., 2015. Firstdetection of Cyprinid herpesvirus 2 (CyHV-2) in goldfish (Carassius auratus) inFrance. J. Fish Dis., http://dx.doi.org/10.1111/jfd.12400.
agatay, I.T., Sen, T.B., 2014. Detection of pathogenic Aeromonas hydrophila fromrainbow trout (Oncorhynchus mykiss) farms in Turkey. Int. J. Agric. Biol. 16,435–438.
ichi, G., Cardeti, G., Cocumelli, C., Vendramin, N., Toffan, A., Eleni, C., et al., 2013.Detection of Cyprinid herpesvirus 2 in association with an Aeromonas sorbiainfection of Carassius carassius (L.) in Italy. J. Fish Dis. 36, 823–830.
eorge, M.R., John, K.R., Mansoor, M.M., Saravanakumar, R., Sundar, P., Pradeep, V.,2015. Isolation and characterization of a ranavirus from koi Cyprinus carpio L.,experiencing mass mortalities in India. J. Fish Dis. 38, 389–403.
oodwin, A.E., Merry, G.E., Sadler, J., 2006a. Detection of the herpesviralhematopoietic necrosis disease agent (Cyprinid herpesvirus 2) in moribundand healthy goldfish: validation of a quantitative PCR diagnostic method. Dis.Aquat. Org. 69, 137–143.
oodwin, A.E., Khoo, L., LaPatra, S.E., Bonar, C., Key, D.W., Garner, M., et al., 2006b.Goldfish haematopoietic necrosis herpesvirus (Cyprinid herpesvirus 2) in theUSA: molecular confirmation of isolates from diseased fish. J. Aquat. Anim.Health 18, 11–18.
oodwin, A.E., Sadler, J., Merry, G.E., Marecaux, E.N., 2009. Herpesviralhaematopoietic necrosis virus (CyHV-2) infection: case studies fromcommercial goldfish farms. J. Fish Dis. 32, 271–278.
roff, J.M., LaPatra, S.E., Munn, R.J., Jinkl, J.G., 1998. A viral epizootic in culturedpopulations of juvenile goldfish due to a putative herpesvirus etiology. J. Vet.Diagn. Invest. 10, 375–378.
anson, L., Dishon, A., Kotler, M., 2011. Herpesviruses that infect fish. Viruses 3,2160–2191.
olt, J.G., Krieg, N.R., Sneath, P.H.A., Staley, J.T., Williams, S.T., 1994. Bergey’sManual of Determinative Bacteriology, 9th edn. Lippincott Williams andWilkins, Baltimore.
to, T., Maeno, Y., 2014. Susceptibility of Japanese Cyprininae fish species toCyprinid herpesvirus 2 (CyHV-2). Vet. Microbiol. 169, 128–134.
effery, K.R., Bateman, K., Bayley, A., Feist, S.W., Hulland, J., Longshaw, C., et al.,2007. Isolation of a Cyprinid herpesvirus 2 from goldfish Carassius auratus (L.),in the UK. J. Fish Dis. 30, 649–656.
ung, S.J., Miyazaki, T., 1995. Herpesviral haematopoietic necrosis of goldfish,Carassius auratus (L.). J. Fish Dis. 18, 211–220.
askhedikar, M., Chhabra, D., 2010. Multiple drug resistance in Aeromonashydrophila isolates of fish. Vet. World 3, 76–77.
aith, A.R., Najiah, M., 2013. Aeromonas hydrophila: antimicrobial susceptibilityand histopathology of isolates from disease catfish, Clarias gariepinus(Burchell). Aquacult. Res. Dev. 5, 215.
ewisch, E., Gorgoglione, B., Way, K., El-Matbouli, M., 2015. Carp edema virus/koi
sleepy disease: an emerging disease in Central-East Europe. Transbound.Emerg. Dis. 62, 6–12.ovy, J., Friend, S.E., 2014. Cyprinid herpesvirus-2 causing mass mortality ingoldfish: applying electron microscopy to histological samples for diagnosticvirology. Dis. Aquat. Org. 108, 1–9.
ica 161 (2016) 8–17 17
Magiorakos, A.P., Srinivasan, A., Carey, R.B., Carmeli, Y., Falagas, M.E., Giske, C.G.,et al., 2012. Multidrug-resistant, extensively drug-resistant andpandrug-resistant bacteria: an international expert proposal for interimstandard definitions for acquired resistance. Bacteriology 18, 268–281.
Miyazaki, T., Kageyama, T., Miura, M., Yoshida, T., 2001. Histopathology ofviremia-associated ana-aki-byo in combination with Aeromonas hydrophila incolor carp Cyprinus carpio in Japan. Dis. Aquat. Org. 44, 109–120.
Mohanty, B.R., Mishra, J., Das, S., Jena, J.K., Sahoo, P.K., 2008. An outbreak ofaeromoniasis in an organized composite carp culture farm in India:experimental pathogenicity and antibiogram study. J. Aqua. 16, 27–37.
Nayak, S.P., Ray, P., Sahoo, P.K., 2013. Molecular characterization and identificationof an alkalophilic bacterial strain isolated from a local hot spring Atri KhurdaDistrict, Odisha, India. J. Pure Appl. Microbiol. 8, 743–747.
Nielsen, M.E., Hoi, L., Schmidt, A.S., Qian, D., Shimada, T., Shen, J.Y., et al., 2001. IsAeromonas hydrophila the dominant motile Aeromonas species that causesdisease outbreaks in aquaculture production in the Zhejiang Province ofChina? Dis. Aquat. Org. 46, 23–29.
Philbey, A.W., 2006. Herpesvirus haematopoietic necrosis in a goldfish (Carassiusauratus) in the UK. Vet. Rec. 158, 800–801.
Rahman, M., Colque-Navarro, P., Kuhn, I., Huys, G., Swings, J., Mollby, R., 2002.Identification and characterization of pathogenic Aeromonas veronii biovarsorbia associated with epizootic ulcerative syndrome in fish in Bangladesh.Appl. Environ. Microbiol. 68, 650–655.
Sahoo, P.K., Das, S., Mahapatra, K.D., Saha, J.N., Baranski, M., Ødegård, J., et al., 2013.Characterization of the ceruloplasmin gene and its potential role as an indirectmarker for selection to Aeromonas hydrophila resistance in rohu, Labeo rohita.Fish Shellfish Immunol. 34, 1325–1334.
Samal, S.K., Das, B.K., Pal, B.B., 2014. Isolation, biochemical characterization,antibiotic susceptibility study of Aeromonas hydrophila isolated fromfreshwater fish. Int. J. Curr. Microbiol. Appl. Sci. 3, 259–267.
Stephens, F.J., Raidal, S.R., Jones, B., 2004. Haematopoietic necrosis in a goldfish(Carassius auratus) associated with an agent morphologically similar toherpesvirus. Aust. Vet. J. 82, 167–169.
Swaminathan, T.R., Basheer, V.S., Rajkumar Kathirvelpandian, A., Sood, N., Jena, J.K.,2015. Establishment and characterization of fin derived cell line fromornamental carp, Cyprinus carpio var. Koi for virus isolation in India. In VitroCell Dev. Biol. Anim. 51, 705–713.
Swann, L., White, M.R., 1991. Diagnosis and treatment of Aeromonas hydrophilainfection of fish. Aquaculture Extension. Illionis Indiana Sea Grant Program.Fact Sheet As-461.
Wahli, T., Burr, S.E., Pugovkin, D., Mueller, O., Frey, J., 2005. Aeromonas sorbia, acausative agent of disease in farmed perch, Perca fluviatilis L. J. Fish Dis. 28,141–150.
Waltzek, T.B., Kelley, G.O., Stone, D.M., Way, K., Hanson, L., Fukuda, H., et al., 2005.Koi herpesvirus represents a third Cyprinid herpesvirus (CyHV-3) in the familyHerpesviridae. J. Gen. Virol. 86, 1659–1667.
Waltzek, T.B., Kurobe, T., Goodwin, A.E., Hedrick, R.P., 2009. Development of apolymerase chain reaction assay to detect Cyprinid herpesvirus 2 in goldfish. J.Aquat. Anim. Health 21, 60–67.
Wooley, R.E., Ritchie, B.W., Burnley, V.V., 2004. In vitro effect of a bufferedchelating agent and neomycin or oxytetracycline on bacteria associated withdiseases of fish, Dis. Aquat. Org. 59, 263–267.
Wu, T., Ding, Z., Ren, M., An, L., Xiao, Z., Liu, P., et al, 2013. The histo- andultra-pathological studies on a fatal disease of Prussian carp (Carassius gibelio)in mainland China associated with Cyprinid herpesvirus 2 (CyHV-2).Aquaculture 413, 8–13.
Xu, J., Zeng, L., Zhang, H., Zhou, Y., Ma, J., Fan, Y., 2013. Cyprinid herpesvirus 2
Biol. Sci. 7, 409–411.