co-evolutionary patterns in congeneric monogeneans: a review of dactylogyrus species and their...
TRANSCRIPT

Co-evolutionary patterns in congeneric monogeneans:a review ofDactylogyrus species and their cyprinid hosts
A. SIMKOVA*† AND S. MORAND‡
*Department of Botany and Zoology, Faculty of Science, Masaryk University, Kotlarska2, 61137 Brno, Czech Republic and ‡Institut des Sciences de l’Evolution – CNRS,D�epartement G�en�etique Environnement, CC065, Universit�e Montpellier 2, 34095,
Montpellier cedex 05, France
Patterns associated with the evolution of parasite diversity, speciation and diversification were
analysed using Dactylogyrus species (gill monogeneans) and their cyprinid hosts as a model. The
aim of this study was to use this highly specific host–parasite systems to review: (1) the diversity
and distribution of Dactylogyrus species, (2) the patterns of organization and structure of
Dactylogyrus communities, (3) the evolution and determinants of host specificity and (4) the
mode of Dactylogyrus speciation and co-evolutionary patterns in this Dactylogyrus–cyprinid
systems. Dactylogyrus are a highly diverse group of parasites, with their biogeography and
distribution clearly linked to the evolutionary history of their cyprinid hosts. The coexistence of
several Dactylogyrus species on one host is facilitated by increasing niche distances and the
differing morphology of their reproductive organs. The positive interspecific and intraspecific
interactions seem to be the most important factors determining the structure of Dactylogyrus
communities. Host specificity is partially constrained by parasite phylogeny. Being a strict
specialist is an ancestral character for Dactylogyrus, being the intermediate specialists or
generalists are the derived characters. The evolution of attachment organ morphology is
associated with both parasite phylogeny and host specificity. Considering larger and long-lived
hosts or hosts with several ecological characters as the measures of resource predictability,
specialists with larger anchors occurred on larger or longer-living fish species. Intra-host
speciation, a mode of speciation not often recorded in parasites, was observed in Dactylogyrus
infecting sympatric cyprinids. Sister parasite species coexisting on the same host occupied niches
that differed in at least one niche variable. Intra-host speciation, however, was not observed in
Dactylogyrus species of congeneric hosts from geographically isolated areas, which suggested
association by descent and host-switching events. # 2008 The Authors
Journal compilation # 2008 The Fisheries Society of the British Isles
Key words: diversity; fish parasites; host specificity; interaction; molecular phylogeny; speciation.
INTRODUCTION
The patterns and processes associated with the evolution of parasite diversity,especially the role of diversification and speciation for increasing parasite diver-sity, have attracted the attention of evolutionary and ecological biologists for
†Author to whom correspondence should be addressed. Tel.: þ420 549497363; fax: þ420 541211214;
email: [email protected]
Journal of Fish Biology (2008) 73, 2210–2227
doi:10.1111/j.1095-8649.2008.02064.x, available online at http://www.blackwell-synergy.com
2210# 2008 The Authors
Journal compilation # 2008 The Fisheries Society of the British Isles

many years. Fish parasites have been increasingly studied in the ecological,evolutionary and molecular phylogenetic fields. Ectoparasites of freshwaterfishes are generally characterized by having direct life cycles. Moreover, mono-genean ectoparasites are highly host specific, so that a given parasite species in-fects a single host species, or a restricted number of host species, and havea limited range of distribution when compared with monogeneans of marinefishes. For this reason, freshwater monogeneans seem to be an appropriatemodel for testing the ecological and evolutionary hypotheses.This study reviews the knowledge on patterns of parasite diversity, the struc-
ture of parasite communities, the evolution of host specificity and parasite spe-ciation using the model of congeneric monogeneans. The first part concerns thespecies diversity of monogeneans. What are the determinants of parasite diver-sity in hosts? Why are several host species able to support high parasite speciesrichness, while others support only a limited number of species? Is monogeneandiversity explained by host life-history traits, phylogeny and abiotic factors?When investigating parasite diversity, information about the origin, historicaldispersal routes and the recent biogeography of hosts should be taken intoaccount (Gibson et al., 1996).The second part of this review concerns the structure of parasite communi-
ties. Is their composition random or predictable? In the case of predictability,what are the factors driving the formation of parasite communities? Generally,two fundamental forces are considered to be important in the structuring ofparasite communities: interspecific interactions (competition) mainly due tothe limited space in available niches, and species coexistence in the case ofabsence of niche saturation (Rohde 1979, 1991; Bush & Holmes, 1986; Holmes,1990). In the case of species coexistence, intraspecific interactions that facilitatemating may determine niche size (Rohde, 1977). Competition is considered tobe the most frequent selective factor responsible for site segregation in endo-parasites (Holmes, 1973). For ectoparasitic monogeneans, niche restriction byspecialization, reduction of competition in evolutionary time and reinforcementof reproductive barriers to prevent hybridization were suggested as mechanismspromoting species coexistence (Rohde, 1977, 1979; Rohde & Hobbs, 1986).The third part of this review concerns the evolution of host specificity and
the processes that may contribute to host specialization in monogeneans, animportant topic in the field of evolutionary ecology. Generally, a parasite in-fecting one host species is described as a specialist, while a parasite infectingmore then one host species is a generalist (Humphery-Smith, 1989). Host spec-ificity plays an important role for the structure of parasite communities whenspecies-rich parasite communities are formed by both specialists and general-ists, whereas species-poor communities are formed mainly by generalist para-sites (Poulin, 1997; Simkova et al., 2001a). The hypothesis of specializationon predictable resources was introduced by Ward (1992) predicting that organ-isms tend to specialize on stable resources, which minimizes the risk of theirextinction. This hypothesis was proposed for host-specific monogeneans para-sitizing fish gills (Simkova et al., 2001a; Desdevises et al., 2002). In thesehost–parasite systems, larger hosts harbour more specialists than smaller hosts,which could be related to resource predictability. Fish longevity, abundance,position at the top of a food chain and all other factors that might decrease
EVOLUTIONARY PATTERNS IN FISH MONOGENEANS 2211
# 2008 The Authors
Journal compilation # 2008 The Fisheries Society of the British Isles, Journal of Fish Biology 2008, 73, 2210–2227

the risk of parasite extinction could be considered as potential measures ofresource predictability for parasites. Rohde (1979, 1989) hypothesized thatthe evolution of host specificity is associated with the evolution of morpholog-ical adaptation, that is the haptor in highly host-specific monogeneans. Finally,host specificity could possibly be constrained by parasite or host phylogeny asproposed by Desdevises et al. (2002). The important aspect connected with theevolution of host specificity is the determination of an ancestral state: is a gen-eralist evolutionarily older than a specialist as hypothesized by Futuyma &Moreno (1988)?The fourth part of this review is devoted to the illustration of patterns of
parasite speciation. In the case of allopatry, parasite speciation follows geo-graphical isolation (and sometimes ecological isolation) of their hosts (Hafner& Nadler, 1988) and therefore speciation in host lineages corresponds to thespeciation in parasite lineages. The sympatric speciation of parasites may occurwhen closely related hosts live in the same or similar ecological conditions,which facilitate host switching (Gu�egan & Agnese, 1991). Finally, parasite spe-ciation without a corresponding host speciation event leads to two or more par-asite species being present on a single host species (Paterson & Gray, 1997). Suchan example of speciation is not often observed in host–parasite systems (Pater-son & Gray, 1997; Paterson & Poulin, 1999; Paterson & Banks, 2001).The aim of this study was to use the highly specific host–parasite systems,
Dactylogyrus spp. (Monogenea) on members of the family Cyprinidae, toreview and discuss current knowledge about: (1) the diversity and distributionof Dactylogyrus species, (2) patterns of organization and structure in Dactylogyruscommunities, (3) the evolution and determinants of host specificity and (4) themode of speciation and co-evolutionary patterns in this system.
THE DIVERSITY AND DISTRIBUTION OF DACTYLOGYRUS
Dactylogyrus spp. represent a group of monogeneans living predominantly onthe gills of cyprinids. Gibson et al. (1996) reviewed more than 900 nominalDactylogyrus species restricted mainly to the Cyprinidae. They included infor-mation on host genera and geographical locations for each nominal species,but information could not be extracted on host specificity as no indicationof host range was mentioned. The overall biogeographical distribution ofDactylogyrus coincides with the area of natural distribution of cyprinids, whichincludes Africa, Asia, North America and Europe. Simkova et al. (2004)hypothesized that the biogeography of Dactylogyrus is linked to the evolution-ary history of cyprinids and supported this hypothesis by molecular phylogenyof Dactylogyrus species on Central European cyprinid subfamilies. The totalphylogenetic reconstruction of Dactylogyrus evolution including a wide inter-continental range of cyprinids, however, is presently lacking. Reports ofDactylogyrus on non-cyprinids are rare but include e.g. ruffe Gymnocephaluscernuus (L.) and Japanese seaperch Lateolabrax japonicus (Cuvier) (Gusev,1985; Cone et al., 1994; Simkova et al., 2006a).Several Dactylogyrus species can coexist on the gills of a host species.
Records of Dactylogyrus species from Central European cyprinids indicatea high diversity on common cyprinids: 14 species on chub Leuciscus cephalus
2212 A. S IMKOVA AND S. MORAND
# 2008 The Authors
Journal compilation # 2008 The Fisheries Society of the British Isles, Journal of Fish Biology 2008, 73, 2210–2227

(L.), 13 species on roach Rutilus rutilus (L.), 10 species on common bream Abra-mis brama (L.) and 10 species on white bream Abramis bjoerkna (L.) (Simkova,2001). Many naturally occurring cyprinids in Central Europe, however, onlyharbour from one to three Dactylogyrus species. In Barbus species from theIberian Peninsula, the endemism of Dactylogyrus species corresponds to theendemism of their cyprinid hosts and congeneric host species share severalDactylogyrus species (El Gharbi et al., 1992).In Africa, cyprinids are dominated by the genera Labeo and Barbus and
Dactylogyrus diversity has been investigated in several species of these genera,particularly from North and West Africa. Gu�egan & Lambert (1990) recordedeight Dactylogyrus species on seven West African barbels. Gu�egan & Agnese(1991) found 14 Dactylogyrus species on West African Labeo species with a max-imum of seven species per host species. These species are believed to berestricted to the host on which they are found. For North America and Asia,there is only limited information about Dactylogyrus (Gibson et al., 1996). East-ern Asia is considered to be the centre of origin for cyprinids (at least for Cyp-rininae) or an important centre of cyprinids interchange (Durand et al., 2002).Recently, the origin of Dactylogyrus species was investigated using molecularphylogeny and the suggestion that Dactylogyrus species originated on the AsianCyprininae was proposed (Simkova et al., 2004).
PATTERNS OF ORGANIZATION AND STRUCTURE INDACTYLOGYRUS COMMUNITIES
The distribution of Dactylogyrus communities follows non-random patterns(Simkova et al., 2000, 2001b). Generally, the species are aggregated both inter-specifically and intraspecifically. Positive interspecific aggregation indicatesthat interspecific competition is not an important factor in the structure ofDactylogyrus assemblages (Simkova et al., 2000). Following the prediction ofthe aggregation model of coexistence (Shorrocks, 1996), species coexistence isfacilitated by reducing the overall intensity of competition via aggregated utili-zation of fragmented resources; hosts in the case of host–parasite systems(Shorrocks & Rosewell, 1986; Jaenike & James, 1991). If species are distributedin such a way that interspecific aggregation is reduced relative to intraspecificaggregation, then species coexistence is facilitated (Shorrocks et al., 1979; Ives &May, 1985; Shorrocks & Rosewell, 1986; Ives, 1988, 1991). This was tested andconfirmed in the case of nine Dactylogyrus species infecting R. rutilus (Simkovaet al., 2000). High levels of intraspecific aggregation as well as a positive inter-specific aggregation have been found when considering either individual hostsor gill arches as independent samples (Simkova et al., 2000, 2001c). Moreover,Dactylogyrus species with similar haptor morphology tend to be more positivelyaggregated at both levels studied: among their hosts and among gill microha-bitats (Simkova et al., 2000, 2001c).Simkova et al. (2000, 2004) observed that each Dactylogyrus species occupies
a specific microhabitat on a host species. Niche breadth, calculated as the sumof the proportion of individuals of a species found in all microhabitats (Fig. 1),was positively correlated with parasite abundance, indicating that interspecificinteractions are not the most important factors for determining the structure of
EVOLUTIONARY PATTERNS IN FISH MONOGENEANS 2213
# 2008 The Authors
Journal compilation # 2008 The Fisheries Society of the British Isles, Journal of Fish Biology 2008, 73, 2210–2227

ectoparasite communities, as previously suggested by Rohde (1979, 1989). Thecoexistence of Dactylogyrus species within one host is facilitated by niche dis-tances and morphological differences among reproductive organs (Simkovaet al., 2002) (see Fig. 2 for morphology of attachment and reproductive or-gans). Thus, species with dissimilar haptor morphology show a smaller nicheoverlap within the host than species with similar haptor morphology [Fig. 3(a)].In addition, the species with similar haptor morphology possess greater differ-ences in size and the form of their reproductive organs [Fig. 3(b), (c)]. Parasitespecialists infecting a single host species occupied the niches situated near eachother within a gill arch. Generalists infecting more than one host species occu-pied the distant niches within individual gill arches (Fig. 4).
FIG. 1. The delimitation of spatial distribution of monogeneans on fish gills. Each gill arch was divided
into three gill segments (D, dorsal; M, medial; V, ventral), three gill areas (p, proximal; c, central; d,
distal), two gill surfaces (in, inner; out, outer) and two gills hemibranchs (A, anterior; P, posterior).
The microhabitats occupied by parasites correspond to the sectors of fish gills.
2214 A. S IMKOVA AND S. MORAND
# 2008 The Authors
Journal compilation # 2008 The Fisheries Society of the British Isles, Journal of Fish Biology 2008, 73, 2210–2227

EVOLUTION AND DETERMINANTS OF HOST SPECIFICITY
Dactylogyrus species are highly host specific, commonly infecting only onehost species or several congeneric hosts. As mentioned, parasites infectingone host species are classified as specialists, while parasites infecting at leasttwo species are classified as generalists (Humphery-Smith, 1989). Such a classi-cal formulation, however, does not distinguish between those parasites infectingnon-related host species and those parasites infecting congeneric host species,both being classified as generalists. Recently, several studies have proposedan index of host specificity, which includes information on phylogenetic
FIG. 2. Morphometric measurements of haptor and copulatory organ of Dactylogyrus species (modified
from Gusev, 1985). The following sclerotized parts of the haptor were measured (measured
characters are included for each sclerotized part): (a) anchor [total length of anchor (1), length of
base (2), length of inner root (3), length of outer root (4) and length of point (5)], (b) marginal hook
[total length (6)], (c) dorsal connective bar [total length (7) and width (8)], (d) ventral connective bar
[total length (9) and width (10)] and (e) copulatory organ [total length (11)].
EVOLUTIONARY PATTERNS IN FISH MONOGENEANS 2215
# 2008 The Authors
Journal compilation # 2008 The Fisheries Society of the British Isles, Journal of Fish Biology 2008, 73, 2210–2227

relatedness of host species (Desdevises et al., 2002; Caira et al., 2003; Poulin &Mouillot, 2003). Host specificity and patterns associated with this feature inDactylogyrus species were analysed using a modification of the index of hostspecificity as defined by Desdevises et al. (2002) for Lamellodiscus species (gillmonogeneans infecting marine fishes of the family Sparidae). Desdeviseset al. (2002) applied the index of host specificity based only on four categoriesof specificity. This index was modified slightly for cyprinids because Dactylogyrusgeneralist species can infect cyprinids with different phylogenetic affinity (i.e.non-congeneric but phylogenetically related species or cyprinid species withoutclose phylogenetic affinity but belonging to the same subfamily or unrelatedcyprinid species belonging to different subfamilies). The different categoriesof host specificity are defined (Table I). Data on 51 Dactylogyrus species re-corded on freshwater fishes in the Czech Republic were used to analyse theevolution and determinants of host specificity (Simkova et al., 2006b). A differ-ence between local host specificity (the specificity for a given parasite specieswithin one locality) and regional host specificity (using the compiled data for
FIG. 3. (a) Dactylogyrus species with similar haptor morphology (i.e. low interspecies morphometrical
distances of attachment) occur in overlapping niches (i.e. high Renkonen niche overlap) following
the equation: y ¼ 612�175 � 1097�602x (n ¼ 36, r2 ¼ 0�353, P < 0�001). (b) Dactylogyrus species in
niches situated in close proximity to each other possess differences in morphology and size of
copulatory organs (i.e. high interspecies morphometrical distances of copulatory organ) following
the equation: y ¼ �14�3x þ 0�0 (n ¼ 36, r2 ¼ 0�203, P < 0�01). (c) Schematic position of two parasite
species with similar morphology of haptors and different morphology and size of copulatory organs
on gill arch is shown (see Fig. 2).
2216 A. S IMKOVA AND S. MORAND
# 2008 The Authors
Journal compilation # 2008 The Fisheries Society of the British Isles, Journal of Fish Biology 2008, 73, 2210–2227
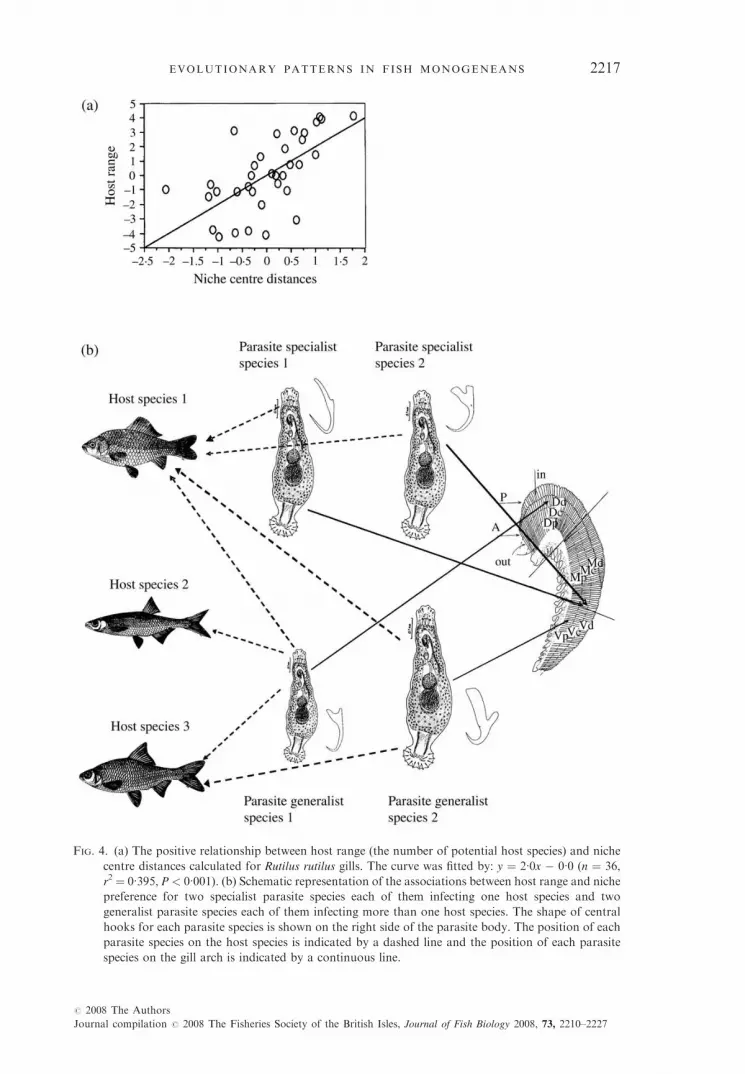
FIG. 4. (a) The positive relationship between host range (the number of potential host species) and niche
centre distances calculated for Rutilus rutilus gills. The curve was fitted by: y ¼ 2�0x � 0�0 (n ¼ 36,
r2 ¼ 0�395, P < 0�001). (b) Schematic representation of the associations between host range and niche
preference for two specialist parasite species each of them infecting one host species and two
generalist parasite species each of them infecting more than one host species. The shape of central
hooks for each parasite species is shown on the right side of the parasite body. The position of each
parasite species on the host species is indicated by a dashed line and the position of each parasite
species on the gill arch is indicated by a continuous line.
EVOLUTIONARY PATTERNS IN FISH MONOGENEANS 2217
# 2008 The Authors
Journal compilation # 2008 The Fisheries Society of the British Isles, Journal of Fish Biology 2008, 73, 2210–2227

the host range of each Dactylogyrus species investigated) was observed, indicat-ing that several parasites recorded as specialists, at a local level, can be consid-ered as generalists at a regional level, that is an increase in area sampling isassociated with high host range. The problem of delimitation of host specificityat different spatial levels was highlighted by Fox & Morrow (1981). Neverthe-less, the most interesting observation was that fish species, recorded as a singlehost for a given Dactylogyrus species at a local level, were considered only asa preferred host species at a regional level. This observation was noted by Nor-ton & Carpenter (1998).The generalist towards specialist hypothesis was proposed by Futuyma &
Moreno (1988) and predicted that specialists evolve from generalists and manyspecialists evolve from other specialists. This hypothesis was tested using theDactylogyrus–Cyprinidae model. Mapping the index of host specificity ontoDactylogyrus phylogeny (Simkova et al., 2006b), however, showed that beinga specialist is the ancestral condition for this genus. Species with higher indexvalues of host specificity, that is with a higher host range, were situated in themore derived lineages of the phylogenetic tree (Fig. 5).The potential associations predicted between host specificity, evolution of
morphological adaptation, the haptor in this case, and molecular phylogenywere analysed using Dactylogyrus (Simkova et al., 2006b). Four different scler-otized structures (shape of central hooks, marginal hooks, dorsal connectivebar and ventral connective bar) on the haptor were mapped separately ontothe phylogeny. This mapping suggested that strict host specificity, or an inter-mediate state, or host preference (for a given host species considering generalistparasites) was associated with morphological adaptation. Similar morphologi-cal characters were observed mainly for parasites infecting congeneric host spe-cies or species belonging to the same host subfamily. Morphological adaptationis not only an advantage for specialists, however, because special morphologi-cal sclerotized structures on the haptor are also found in generalist parasitesinfecting a wide range of hosts.
TABLE I. Index of host specificity applied for Dactylogyrus species according to Simkovaet al. (2006)
Index of host specificity Category of parasites Host species
1 Strict specialist ¼1 host species2 Intermediate specialist �2 congeneric host species3 Intermediate generalist �2 non-congeneric host
species forming onemonophyletic group
4 Generalist �2 different host speciesbelonging to onesubfamily
5 Wide range generalist �2 unrelated host speciesbelonging to differentsubfamilies
2218 A. S IMKOVA AND S. MORAND
# 2008 The Authors
Journal compilation # 2008 The Fisheries Society of the British Isles, Journal of Fish Biology 2008, 73, 2210–2227

In the study by Simkova et al. (2006b) of Dactylogyrus species, the determi-nants of host specificity were investigated. They included host body size, lon-gevity, density and two habitat characters: benthic, pelagic or withoutpreference and rheophilous, without preference, or limnophilous. For this anal-ysis, Dactylogyrus were evaluated in two groups using the classical definition ofspecialists and generalists. The association between morphometric measure-ments of the haptor (Fig. 2) after correcting for parasite body size and parasitephylogeny, and the potential determinants of host specificity were analysed.The hypothesis of specialization on a predictable resource proposed by Ward
FIG. 5. Mapping of host specificity onto the parasite (Dactylogyrus spp.) phylogenetic reconstruction.
Numbers along branches indicate bootstrap values resulting form the phylogenetic analyses
neighbour-joining, maximum parsimony and maximum likelihood (from Simkova et al., 2006b).
The values from 1 to 5 indicate the index of host specificity. , 1; , 2; , 3; , 4; , 5; , equivocal.
EVOLUTIONARY PATTERNS IN FISH MONOGENEANS 2219
# 2008 The Authors
Journal compilation # 2008 The Fisheries Society of the British Isles, Journal of Fish Biology 2008, 73, 2210–2227

(1992) was supported by the observation that Dactylogyrus specialists withlarger central hooks tended to live on hosts with larger body size and long-livedhosts (Simkova et al., 2006b). This could also be interpreted as a mechanismfor optimizing morphological adaptation. Furthermore, long-living hosts har-bour specialists with larger body sizes. It was also shown that host ecologyis an important determinant of host specificity. The hypothesis of specializationon predictable hosts was previously confirmed in marine gill parasites of theLamellodiscus genus, where host specificity increased with host body size. Inthat study, larger hosts are also assumed to be longer lived, thus being a morepredictable resource for parasites (Desdevises et al., 2002). This study of Lamel-lodiscus, however, revealed a relationship between host size and parasite bodysize in both specialists and generalists, suggesting that a preferred host of La-mellodiscus also represents a predictable resource for generalist parasites. In thecase of Dactylogyrus, there was no relationship between preferred host and gen-eralist parasites, that is preferred hosts for generalist parasites do not representpredictable hosts, supporting the hypothesis of Ward (1992).
MODE OF SPECIATION AND CO-EVOLUTIONARY PATTERNSOF DACTYLOGYRUS
MOLECULAR PHYLOGENY OF DACTYLOGYRUS SPECIES – ACASE OF INTRA-HOST SPECIATION
Dactylogyrus and the Cyprinidae represent a specific model system, for suchsystems it was hypothesized that host specificity promotes the parasite speciation(Brooks & McLennan, 1993). The molecular phylogenetic analyses includeda wide range of Dactylogyrus species including representatives of fauna livingin Central Europe, collected from sympatric cyprinids, and a single percid species(Simkova et al., 2004). In their study, it was hypothesized that highly specificcongeneric parasites coexisting on the same host species (Dactylogyrus speciesin this case) have speciated via intra-host speciation. Moreover, the different mi-crohabitats were hypothesized for phylogenetically related parasites.Three Dactylogyrus lineages were recognized using phylogenetic reconstruc-
tion on the combined data 18SrDNA and ITS1 (Fig. 6) suggesting a possibleassociation between Dactylogyrus and cyprinid evolution (Simkova et al.,2004). The phylogenetic analysis gave only weak support to the ancestral posi-tion of Dactylogyrus species of Cyprininae originating from Asia, i.e. commoncarp Cyprinus carpio L. and Prussian carp Carassius gibelio (Bloch). Thehypotheses of origin, historical migration routes and recent distribution of cyp-rinids suggested by several phylogenetic studies (Nelson, 1994; Zardoya &Doadrio, 1999; Zardoya et al., 1999; Durand et al., 2002) support the situationproposed for Dactylogyrus evolution (Simkova et al., 2004): Asian Cyprininaebeing a plesiomorphic group for Dactylogyrus and secondary colonization ofEuropean cyprinids of Gobioninae: gudgeon Gobio gobio (L.) and white-finnedgudgeon Romanogobio albipinnatus (Lukasch), Rasborinae: stone moroko Pseu-dorasbora parva (Temminck & Schlegel), an introduced species in Europe, Leu-ciscinae and one species of Percidae. The mapping of host subfamilies onto thephylogenetic reconstruction of Dactylogyrus species suggests that cyprinid
2220 A. S IMKOVA AND S. MORAND
# 2008 The Authors
Journal compilation # 2008 The Fisheries Society of the British Isles, Journal of Fish Biology 2008, 73, 2210–2227

species belonging to Alburninae, with their nested position within Leuciscinae(Gilles et al., 1998, 2001), were colonized by Dactylogyrus from Leuciscinae(Simkova et al., 2004). The colonization of barbel Barbus barbus (L.) (Cyprini-nae) by Dactylogyrus from Leuciscinae might be explained by the biogeographyof this species in Europe (Cunha et al., 2002; Durand et al., 2002).The comparison of host and parasite phylogenies (Fig. 6) demonstrated that
intra-host speciation is the process most often observed in Dactylogyrus species
FIG. 6. Tanglegram of Dactylogyrus species and their host species deduced from comparison of the
parasite tree topology and fish tree (from Simkova et al., 2004, with permission from John Wiley &
Sons). Intra-host duplications are depicted by shading on the trees. The co-evolution events are
indicated by a, b and c.
EVOLUTIONARY PATTERNS IN FISH MONOGENEANS 2221
# 2008 The Authors
Journal compilation # 2008 The Fisheries Society of the British Isles, Journal of Fish Biology 2008, 73, 2210–2227

of examined fish species. Thirteen intra-host speciation events were recorded in11 host species from a total of 20 host species examined. A small number of co-speciation events were observed using the comparison of host and parasite phy-logenies. The first represents a concordance of phylogenetic relationshipsbetween fish species of Gobioninae and Rasborinae and their parasites, the sec-ond includes G. gobio and R. albipinnatus with their two parasite species, andthe last, rudd Scardinius erythrophthalmus (L.) and bleak Alburnus alburnus(L.) with their respective parasites.The observed mode of speciation, i.e. intra-host speciation, was analysed
with respect to the pattern of species coexistence. The coexistence of nineDactylogyrus species was investigated in R. rutilus by Simkova et al. (2002)(Figs 3 and 4) who found that this coexistence may be facilitated by differencesin the morphology of the copulatory organ or increasing niche distances, whichprevent interspecific competition and increase mating contacts (Simkova et al.,2002). The mapping of preferred niches onto the molecular phylogenetic recon-struction showed that parasite species, speciating within one host species, tendto occupy niches different in at least one variable (gill arch, segment or area)(Simkova et al., 2006b).
MOLECULAR PHYLOGENY OF DACTYLOGYRUS SPECIESPARASITIZING CONGENERIC HOSTS: A CASE OFDACTYLOGYRUS ON BARBUS SPECIES
The molecular phylogenetic reconstruction was performed and aimed toinvestigate the mode of speciation in Dactylogyrus species parasitizing conge-neric hosts (Simkova et al., 2007). In this study, selected Barbus species col-lected from geographically isolated regions and their Dactylogyrus specieswere used (Table II). Geographically isolated Barbus species shared the samemonogenean species (D. carphathicus and D. dyki). The analyses of morphomet-ric variability (using the sclerotized parts of attachment and reproductive or-gans) and molecular variability (using partial 18S rDNA and ITS1),
TABLE II. Data on congeneric cyprinids (Barbus species) from geographically isolatedlocalities and their Dactylogyrus species from Simkova et al. (2007)
Barbus speciesCountry ofcollection Sea drainage Dactylogyrus species
Barbus balcanicus Bulgaria,South Europe
Black Sea D. crivelliusD. dykiD. petenyi
Barbus barbus Czech Republic,Central Europe
Black Sea D. carpathicusD. dykiD. malleus
Barbus cyclolepis Bulgaria,South Europe
Aegean Sea D. dyki
Barbus meridionalis Spain,West Europe
MediterraneanSea
D. carpathicusD. dyki
2222 A. S IMKOVA AND S. MORAND
# 2008 The Authors
Journal compilation # 2008 The Fisheries Society of the British Isles, Journal of Fish Biology 2008, 73, 2210–2227

however, showed differences between Dactylogyrus populations infecting thegeographically isolated Barbus species. This suggests an association betweenhost switching and the evolutionary history of Barbus species. A commonancestor of a parasite species (D. dyki in this case) colonizes an ancestral Barbusspecies or more Barbus species living in sympatry and subsequently, the isola-tion of Barbus leads to the morphological and molecular differentiation of par-asite species. Moreover, this observation also suggested the need to re-evaluatethe taxonomic status of the parasites investigated.The comparison of monogenean and host phylogenetic trees (Fig. 7) revealed
that Dactylogyrus species coexisting in one host species did not diverge byintra-host speciation. A few co-speciation events were estimated using the com-parison of host and parasite phylogenetic reconstructions. Host switching withconsequent speciation could explain the incongruent phylogenies. Two well-supported clades of Dactylogyrus species were recognized from the phylogeneticreconstruction differing in the morphology of their haptors (see Fig. 7 for thedifferences in morphology of the ventral connective bar), supporting thehypothesis that morphological evolution of the haptor is associated withDactylogyrus phylogeny as proposed by Simkova et al. (2006b).
CONCLUSIONS
Dactylogyrus spp., due to their high diversity and host restriction mainly tocyprinids, were selected as suitable models for the study of parasite speciationand diversification. It was predicted and confirmed that the biogeography anddistribution of these parasites is clearly linked to the evolutionary history ofcyprinids. Intra-host speciation was observed as an important process of Dac-tylogyrus diversification in sympatric cyprinid species, even if this mode of spe-ciation does not seem to be favoured in Dactylogyrus occurring on congenerichosts (Barbus in this case). Subsequent investigations including a wide range ofcongeneric host species parasitized by Dactylogyrus are needed; this parasitemodel system could also be utilized to test biogeographical hypotheses relatingto the origin, past dispersion and colonization of the congeneric cyprinids inthe future.Another finding in this host–parasite system is the association between intra-
host speciation and species coexistence. Parasite species, speciating within a sin-gle host species, tend to occupy niches differently in at least one variable. Thisconfirms the prediction that the coexistence of congeneric species is facilitatedby an increase in niche distances, which prevents interspecific competition. Thefacilitation of species coexistence is also confirmed by ecological studies demon-strating that the structure of Dactylogyrus communities is determined by intra-specific and positive interspecific interactions; therefore, competition is nota potential force shaping this structure of parasite communities.It was observed that the host specificity of Dactylogyrus evolved from special-
ists to intermediate states of host specificity or to generalists. Host specificity ispartially constrained by parasite phylogeny and determined by host body size,longevity and host ecology. These findings support: (1) the idea that the selectedmeasurements should be considered as the reliable variables of host predictabilityand (2) the hypothesis of parasite specialisation on predictable resources.
EVOLUTIONARY PATTERNS IN FISH MONOGENEANS 2223
# 2008 The Authors
Journal compilation # 2008 The Fisheries Society of the British Isles, Journal of Fish Biology 2008, 73, 2210–2227

FIG. 7. Tanglegram of Dactylogyrus species and Barbus species deduced from comparison of the parasite
tree topology and fish tree (modified from Simkova et al., 2007, with permission from Cambridge
University Press). The different shape of ventral connective bars (a sclerotized part of the haptor) are
indicated by a, b, c and d.
2224 A. S IMKOVA AND S. MORAND
# 2008 The Authors
Journal compilation # 2008 The Fisheries Society of the British Isles, Journal of Fish Biology 2008, 73, 2210–2227

This study was funded by the Research Project of the Masaryk University, Brno, Pro-ject No.: MSM 0021 622 416. A.S. was supported by the Ichthyoparasitology, Centre ofExcellence, Project No. LC 522 funded by Ministry of Education, Youth and Sports ofthe Czech Republic. We are very grateful to G. Speas for the revision of the English inthe final version.
References
Brooks, D. R. & McLennan, D. A. (1993). Parascript: Parasites and the Language ofEvolution. Washington, DC: Smithsonian Institution Press.
Bush, A. O. & Holmes, J. C. (1986). Intestinal helminths of lesser scaup ducks: aninteractive community. Canadian Journal of Zoology 64, 142–152.
Caira, J. N., Jensen, K. & Holsinger, K. E. (2003). On a new index of host specificity. InTaxonomy, Ecology and Evolution of Metazoan Parasites (Combes, C. & Jourdane, J.,eds), pp. 161–181. Perpignan: Presses Universitaires de Perpignan.
Cone, D., Eurell, T. & Beasley, V. (1994). A report of Dactylogyrus amphibothrium(Monogenea) on the gills of European ruffe in western Lake Superior. Journal ofParasitology 80, 476–478.
Cunha, C., Mesquita, N., Dowling, T. E., Gilles, A. & Coelho, M. M. (2002).Phylogenetic relationships of Eurasian and American cyprinids using cytochromeb sequences. Journal of Fish Biology 61, 929–944. doi: 10.1006/jfbi.2002.2105
Desdevises, Y., Morand, S. & Legendre, P. (2002). Evolution and determinants of hostspecificity in the genus Lamellodiscus (Monogenea). Biological Journal of theLinnean Society 77, 431–443.
Durand, J.-D., Tsigenopoulos, C. S., Unlu, E. & Berrebi, P. (2002). Phylogeny andbiogeography of the family Cyprinidae in the Middle East inferred fromcytochrome b DNA – evolutionary significance of this region. Molecular Phyloge-netics and Evolution 22, 91–100. doi: 10.1006/mpev.2001.1040
El Gharbi, S., Renaud, F. & Lambert, A. (1992). Dactylogyrids (Platyhelminthes:Monogenea) of Barbus spp. (Teleostei: Cyrpinidae) from the Iberian Peninsula.Research and Reviews in Parasitology 52, 103–116.
Fox, L. R. & Morrow, P. A. (1981). Specialization: species property of local pheno-menon? Science 211, 887–893.
Futuyma, D. J. & Moreno, G. (1988). The evolution of ecological specialization. AnnualReview of Ecology and Systematics 19, 207–233.
Gibson, D. I., Timofeeva, T. A. & Gerasev, P. I. (1996). A catalogue of the nominalspecies of the monogenean genus Dactylogyrus Diesing, 1850 and their host genera.Systematic Parasitology 35, 3–48.
Gilles, A., Lecointre, G., Faure, E., Chappaz, R. & Brun, G. (1998). Mitochondrialphylogeny of the European cyprinids: implications for their systematics, reticulateevolution, and colonization time. Molecular Phylogenetics and Evolution 10, 132–143. doi: 10.1006/mpev.1997.0480
Gilles, A., Lecointre, G., Miquelis, A., Loerstcher, M., Chappaz, R. & Brun, G. (2001).Partial combination applied to phylogeny of European cyprinids using themitochondrial control region. Molecular Phylogenetics and Evolution 19, 22–33.doi: 10.1006/mpev.2000.0916
Gu�egan, J.-F. & Agnese, J.-F. (1991). Parasite evolutionary events inferred from hostphylogeny: the case of Labeo species (Teleostei, Cyprinidae) and their dactylogyridparasites (Monogenea, Dactylogyridae). Canadian Journal of Zoology 69, 595–603.
Gu�egan, J.-F. & Lambert, A. (1990). Twelve new species of dactylogyrids (Platyhel-minthes, Monogenea) from West African barbels (Teleostei, Cyprinidae), withsome biogeographical implications. Systematic Parasitology 17, 153–181.
Gusev, A. V. (1985). Metazoa parasites. Part I. In Identification Key to Parasites ofFreshwater Fish, Vol. 2 (Bauer, O. N., ed.), pp. 1–424. Leningrad: Nauka (inRussian).
Hafner, M. S. & Nadler, S. A. (1988). Phylogenetic trees support the coevolution ofparasites and their hosts. Nature 332, 258–259.
EVOLUTIONARY PATTERNS IN FISH MONOGENEANS 2225
# 2008 The Authors
Journal compilation # 2008 The Fisheries Society of the British Isles, Journal of Fish Biology 2008, 73, 2210–2227

Holmes, J. C. (1973). Site selection by parasitic helminths: interspecific interactions, sitesegregation, and their importance to the development of helminth communities.Canadian Journal of Zoology 51, 333–347.
Holmes, J. C. (1990). Competion, contacts and other factors restricting niches of parasitehelminths. Annales de Parasitologie Humaine et Comparee 65, 69–72.
Humphery-Smith, I. (1989). The evolution of phylogenetic specificity among parasiticorganisms. Parasitology Today 5, 385–387.
Ives, A. R. (1988). Aggregation and the coexistence of competitors. Annales ZoologiciFennici 25, 75–88.
Ives, A. R. (1991). Aggregation and coexistence in a carrion fly community. EcologicalMonographs 61, 75–94.
Ives, A. R. & May, R. M. (1985). Competition within and between species in a patchyenvironment: relations between microscopic and macroscopic models. Journal ofTheoretical Biology 133, 65–92.
Jaenike, J. & James, A. C. (1991). Aggregation and the coexistence of mycophagousDrosophila. Journal of Animal Ecology 60, 913–928.
Nelson, J. S. (1994). Fishes of the World. New York: John Wiley & Sons Inc.Norton, D. A. & Carpenter, M. A. (1998). Mistletoes as parasites: host specificity and
speciation. Trends in Ecology and Evolution 13, 101–105.Paterson, A. M. & Banks, J. (2001). Analytical approaches to measuring cospeciation of
host and parasites: through a glass, darkly. International Journal for Parasitology31, 1012–1022. doi: 10.1016/S0020-7519(01)00199-0
Paterson, A. M. & Gray, R. D. (1997). Host-parasite co-speciation, host switching andmissing the boat. In Host-parasite Evolution: General Principles and Avian Models(Clayton, D. H. & Moore, J., eds), pp. 236–250. Oxford: Oxford University Press.
Paterson, A. M. & Poulin, R. (1999). Have chondracanthid copepods co-speciated withtheir teleost hosts? Systematic Parasitology 44, 79–85.
Poulin, R. (1997). Parasite faunas of freshwater fish: the relationship between richnessand the specificity of parasites. International Journal for Parasitology 27, 1091–1098.doi: 10.1016/S0020-7519(97)00070-2
Poulin, R. & Mouillot, D. (2003). Parasite specialization from a phylogenetic perspective:a new index of host specificity. Parasitology 126, 473–480. doi: 10.1017/S0031182003002993
Rohde, K. (1977). A non-competitive mechanism responsible for restricting niches inparasites. Zoologisher Anzeiger 199, 164–172.
Rohde, K. (1979). A critical evaluation of intrinsic and extrinsic factors responsible forniche restriction in parasites. American Naturalist 114, 648–667.
Rohde, K. (1989). Simple ecological systems, simple solution to complex problems?Evolutionary Theory 8, 305–350.
Rohde, K. (1991). Intra- and interspecific interactions in low density populations inresource-rich habitats. Oikos 60, 91–104.
Rohde, K. & Hobbs, R. (1986). Species segregation: competition or reinforcement ofreproductive barriers? In Parasite Lives. Papers on Parasites, their Hosts and theirAssociation to Honour J. F. A. Sprent (Cremin, M., Dobson, C. & Noorhouse, E.,eds), pp. 189–199. St Lucia: University of Queensland Press.
Shorrocks, B. (1996). Local diversity: a problem with too many solutions. In Aspects ofthe Genesis and Maintenance of Biological Diversity (Hochberg, M. E., Clobert, J. &Barbault, R., eds), pp. 104–122. Oxford: Oxford University Press.
Shorrocks, B. & Rosewell, J. (1986). Guild size in drosophilids: a simulation model.Journal of Animal Ecology 55, 527–541.
Shorrocks, B., Athenson, W. D. & Charlesworth, P. (1979). Competition on a dividedand ephemeral resource. Journal of Animal Ecology 48, 899–908.
Simkova, A. (2001). The analysis of the genus Dactylogyrus (Monogenea): evolutionary-ecological aspects in host-parasite system. PhD Thesis. University of Komensky,Bratislava, Slovakia.
Simkova, A., Desdevises, Y., Gelnar, M. & Morand, S. (2000). Coexistence of nine gillectoparasites (Dactylogyrus: Monogenea) parasitising the roach (Rutilus rutilus L.):
2226 A. S IMKOVA AND S. MORAND
# 2008 The Authors
Journal compilation # 2008 The Fisheries Society of the British Isles, Journal of Fish Biology 2008, 73, 2210–2227

history and present ecology. International Journal for Parasitology 30, 1077–1088.doi: 10.1016/S0020-7519(00)00098-9
Simkova, A., Desdevises, Y., Morand, S. & Gelnar, M. (2001a). Morphometric correlatesof host specificity in Dactylogyrus species (Monogenea) parasites of Europeancyprinid fish. Parasitology 123, 169–177. doi: 10.1017/S0031182001008241
Simkova, A., Gelnar, M. & Morand, S. (2001b). Order and disorder in ectoparasitecommunities: the case of congeneric gill monogeneans (Dactylogyrus spp.). Inter-national Journal for Parasitology 31, 1205–1210. doi: 10.1016/S0020-7519(01)00245-4
Simkova, A., Gelnar, M. & Sasal, P. (2001c). Aggregation of congeneric parasites(Monogenea: Dactylogyrus) among gill microhabitats within one host species(Rutilus rutilus L.). Parasitology 123, 599–607. doi: 10.1017/S0031182001008782
Simkova, A., Ondrackova, M., Gelnar, M. & Morand, S. (2002). Morphology andcoexistence of congeneric ectoparasite species: reinforcement of reproductiveisolation? Biological Journal of the Linnean Society 76, 125–135.
Simkova, A., Morand, S., Jobet, E., Gelnar, M. & Verneau, O. (2004). Molecularphylogeny of congeneric monogenean parasites (Dactylogyrus): a case of intra-hostspeciation. Evolution 58, 1001–1018.
Simkova, A., Matejusova, I. & Cunningham, C. O. (2006a). A molecular phylogeny of theDactylogyridae sensu Kritsky & Boeger (1989) (Monogenea) based on the D1-D3domains of large subunit rDNA. Parasitology 133, 43–53. doi: 10.1017/S0031182006009942
Simkova, A., Verneau, O., Gelnar, M. & Morand, S. (2006b). Specificity and speciali-sation of congeneric monogeneans parasitizing cyprinid fish. Evolution 60, 1023–1037.
Simkova, A., Pecınkova, M., Rehulkova, E., Vyskocilova, M. & Ondrackova, M. (2007).Dactylogyrus species parasitizing European Barbus species: morphometric andmolecular variability. Parasitology 134, 1751–1765. doi: 10.1017/S0031182007003265
Ward, S. A. (1992). Assessing functional explanations of host specificity. AmericanNaturalist 139, 883–891.
Zardoya, R. & Doadrio, I. (1999). Molecular evidence on the evolutionary andbiogeographical patterns of European cyprinids. Journal of Molecular Evolution49, 227–237.
Zardoya, R., Economidis, P. S. & Doadrio, I. (1999). Phylogenetic relationships of GreekCyprinidae: molecular evidence for at least two origins of the Greek cyprinidfauna. Molecular Phylogenetics and Evolution 13, 122–131. doi: 10.1006/mpev.1999.0630
EVOLUTIONARY PATTERNS IN FISH MONOGENEANS 2227
# 2008 The Authors
Journal compilation # 2008 The Fisheries Society of the British Isles, Journal of Fish Biology 2008, 73, 2210–2227