cytotoxicity of tio2 nanoparticles towards freshwater sediment microorganisms at low exposure...
TRANSCRIPT

Environmental Research 135 (2014) 333–345
Contents lists available at ScienceDirect
Environmental Research
http://d0013-93
n CorrE-m
journal homepage: www.elsevier.com/locate/envres
Cytotoxicity of TiO2 nanoparticles towards freshwater sedimentmicroorganisms at low exposure concentrations
Jyoti Kumari a, Deepak Kumar a, Ankita Mathur a, Arif Naseer a, Ravi Ranjan Kumar a,Prathna Thanjavur Chandrasekaran b, Gouri Chaudhuri a, Mrudula Pulimi a,Ashok M. Raichur b,c, S. Babu d, Natarajan Chandrasekaran a, R. Nagarajan e,Amitava Mukherjee a,n
a Centre for Nanobiotechnology, VIT University, Vellore 632014, Indiab Department of Materials Engineering, Indian Institute of Science, Bangalore, Indiac Department of Chemical Technology, University of Johannesburg, South Africad School of Bio Sciences and Technology, VIT University, Vellore, Indiae Department of Chemical Engineering, IIT Madras, Chennai, India
a r t i c l e i n f o
Article history:Received 4 August 2014Received in revised form23 September 2014Accepted 25 September 2014
Keywords:TiO2 NPsBacteriaVisible light and dark conditionsConsortium
x.doi.org/10.1016/j.envres.2014.09.02551/& Elsevier Inc. All rights reserved.
esponding author.ail addresses: [email protected], am
a b s t r a c t
There is a persistent need to assess the effects of TiO2 nanoparticles on the aquatic ecosystem owing totheir increasing usage in consumer products and risk of environmental release. The current study isfocused on TiO2 nanoparticle-induced acute toxicity at sub-ppm level (r1 ppm) on the three differentfreshwater sediment bacterial isolates and their consortium under two different irradiation (visible lightand dark) conditions. The consortium of the bacterial isolates was found to be less affected by theexposure to the nanoparticles compared to the individual cells. The oxidative stress contributedconsiderably towards the cytotoxicity under both light and dark conditions. A statistically significantincrease in membrane permeability was noted under the dark conditions as compared to the lightconditions. The optical and fluorescence microscopic images showed aggregation and chain formation ofthe bacterial cells, when exposed to the nanoparticles. The electron microscopic (SEM, TEM) observationssuggested considerable damage of cells and bio-uptake of nanoparticles. The exopolysaccrides (EPS)production and biofilm formation were noted to increase in the presence of the nanoparticles, andexpression of the key genes involved in biofilm formation was studied by RT-PCR.
& Elsevier Inc. All rights reserved.
1. Introduction
Titanium dioxide nanoparticles (TiO2 NPs) have a broad rangeof commercial applications, especially in consumer products(Newman et al., 2009). Nanotechnology-based products are esti-mated to achieve $3 trillion market with six million workers by2020 (Liu et al., 2013). Owing to the sharp increase in TiO2 NPusage in industry, an increase in their release into the environmentalong with industrial effluents is also anticipated. It has beenreported that TiO2 NPs can enter the aquatic environment eitherthrough direct or indirect release from nano-paints, sunscreenlotions, food additives, medical use, dismantling of batteries,recycling of plastic/glass/metal with nano-coating, and ground-water remediation (O’Brien and Cummins, 2010), which in turnmay cause toxicity to the living organisms in the environment.
[email protected] (A. Mukherjee).
The presence of microorganisms is essential for maintenance ofa sustainable ecosystem. Microorganisms being the chief agents ofearly transformation of organic matter, regeneration of nutrientsas well as food source for higher trophic level play an importantrole in ecosystem. Bacteria are being increasingly used as possibletest systems for evaluating nanoparticles toxicity. Not only theyare deemed as receptors necessary to study ecological nano-toxicology (Holden et al., 2012), but they also act as facile testsubjects that can be used in miniaturized toxicological screeningfor rapid hazard identification (Jin et al., 2010; Nel et al., 2013). Theconclusions from the bacterial toxicity assay can encourage thedevelopment of safer nanomaterial designs (Wang et al., 2012).
From a thorough literature review, the interaction between thenanoparticles and the bacterial cell leading to cytotoxicity hasbeen hypothesized to involve two steps. The first step is theoxidative damage by the nanoparticles attached to the cellmembrane, resulting in loss in membrane integrity withoutsignificant reduction in cell viability. The second step involvesleaking out of intracellular components, which is the leading cause

J. Kumari et al. / Environmental Research 135 (2014) 333–345334
of decreased viability and internalization of the nanoparticles,thereby causing damage to cell organelles including the nucleus(Klaine et al., 2008). Previous reports suggest that the adsorptionof TiO2 NPs onto the cell wall, interruption of the transmembraneelectron transfer, modification of the membrane potential, physicaldamage resulting in leakage of the cell contents, and the produc-tion of reactive oxygen species (ROS) were among the majorcontributors in the cell–nanoparticles cytotoxic interactions(Klaine et al., 2008; Li et al., 2008).
Generally the concentration of TiO2 NPs in various environ-mental compartments across several continents has been found tobe in the range of 0.7–16 μg/L (Mueller and Nowack, 2008). Thecytotoxic effects of the nanoparticles may be strongly influencedby their aggregation/agglomeration in the test system, with thesmaller/more stable particles having enhanced penetration andaccumulation in the cells, causing more damage of the cells thanthat of larger/agglomerated particles (Navarro et al., 2008). Sincethere is evidence showing free radical generation by TiO2 NPsunder both visible light and dark conditions (Sayes et al., 2006;Lipovsky et al., 2012) cytotoxicity under differing irradiationconditions requires attention. These influencing factors need tobe kept in mind while designing a complete toxicity assayinvolving environmental microorganisms and nanomaterials.
Most of the prior reports regarding environmental toxicity ofTiO2 NPs dealt with individual bacterial cells, which may notalways be the case in the natural environment. It can be hypothe-sized that the consortium being heterogeneous in nature mayexperience less toxic impact of the nanomaterial in the environ-ment than the individual bacterial cells, which are homogenous.The aim of the present study was to elucidate the phototoxiceffects of TiO2 NPs to different individual bacterial species isolatedfrom the freshwater sediments as well as their consortium (amixture of a number of different cultures) at low exposureconcentrations (0.25, 0.50, 0.75 and 1.00 μg/mL) in a freshwatersystem without any nutrient supplements (to mimic the chemicalmatrix of a freshwater aquatic environment) under visible lightand dark conditions.
2. Materials and methods
2.1. Materials
Titanium oxide NPs were procured from Sigma Aldrich (drytitanium (IV) dioxide nanopowder, 99.7% anatase, CAS no.:637254). Ethidium bromide (EtBr) was procured from MedoxBiotech India Pvt. Ltd. Acridine Orange (AO) was obtained fromHi-Media Pvt. Ltd. (Mumbai, India). 2′,7′-Dichloro fluoresceindiacetate (DCFH-DA) was purchased from Sigma Aldrich (St. Louis,MO, USA). All other chemicals used throughout the experimentwere of analytical reagent grade.
Lake water from VIT Lake, Vellore, India, was collected (12°58′10″N, 79°937″E) (Pakrashi et al., 2013). ICP-OES (Perkin ElmerOptima 5300 DV, USA) was carried out in order to quantify themetal ions (Al3þ ,Cu2þ , Zn2þ , Mn2þ) and other inorganic ions( − − − −SO , PO , NO , Cl4
24
3 3 ) present in lake water by titrimetricmethods (details are provided in our previous report) (Pakrashiet al., 2011). Lake water was filtered through Whattman No. 1 filterpaper to remove suspended dust particles, followed by steriliza-tion keeping the chemical composition of the water unperturbed.A secondary filtration, through a 0.22-μm membrane filter, wasdone to avoid the interference of large colloidal particles prior tohydrodynamic size measurement (Pakrashi et al., 2011). Thesterilized lake water was used as the experimental matrixthroughout the study (Dalai et al., 2014).
2.2. Stability analysis of nanoparticles dispersion
To study the colloidal stability of TiO2 NPs in the lake water atthe time periods of 0, 2, 4, 6, and 24 h mean hydrodynamic sizeanalysis by dynamic light scattering was carried out using 90 plusParticle Size Analyzer, Brookhaven Instruments Corporations, USA.A stock dispersion of TiO2 NPs (100 mg/mL) was prepared in MilliQwater and sonicated using an ultrasonic processor (Sonics, USA) of350 W for 10 min. A working concentration of 1 mg/mL of TiO2 NPswas prepared in filtered lake water by diluting the requiredvolume of stock solution. Hydrodynamic size of the particles inthe suspension was measured at time intervals of 0 h, 2 h, 4 h, 6 hand 24 h by dynamic light scattering method using a 90 plusParticle Size Analyzer (Brookhaven Instruments Corporations,USA) (Dalai et al., 2012).
The actual concentration of TiO2 NPs in the lake water mediumwas measured by ICP-OES (Perkin Elmer Optima 5300 DV, USA).Four different concentrations, i.e., 0.25, 0.50, 0.75, and 1 mg/mL,were prepared by adding desired volume of stock in 20 mLworking volume of lake water. The samples were then aciddigested and analysed by ICP-OES (Perkin-Elmer Optima 5300DV, USA).
2.3. Isolation and identification of bacteria
Sediment sample from the VIT Lake, VIT University, Vellore,India (conductance: 4.370.13 mS cm�1; pH: 7.8; DO:7.270.46 μg/mL; TDS: 800 774 μg/mL) was collected and storedin polypropylene bottles at 4 °C used for microbiological analysis.1 g of sediment sample as added to 100 mL of sterile water, stirredvigorously and then allowed to sit for the soil to settle. Serialdilution of the sample was carried out with sterile distilled water.This suspension was added to a nutrient agar plate and kept in anincubator at 30–37 °C. After 24 h the observed bacterial colonieswith different colony morphologies were isolated and labelled.The isolates were then plated again on a nutrient agar medium toobtain pure cultures.
2.4. Development of the ternary consortium
To examine the toxic effects of TiO2 NPs, three individualbacterial species and a bacterial consortium (a mixture of cultures)were prepared. Antagonistic/synergistic studies were carried outwith three bacterial isolates (Bacillus alitudinis, Bacillus subtilis andPseudomonas aeruginosa) for the development of the ternaryconsortium. One isolate (B. alitudinis) was taken and grown innutrient broth, after 4 h of incubation; 100 μL of culture broth waspoured onto the surface of nutrient agar and a loop full of cultureof the second isolate (B. subtilis) was streaked in the middle of theplate. Same step was followed in case of B. subtilis, P. aeruginosaand B. alitudinis, P. aeruginosa. After that the plates were incubatedat 30–37 °C for 24 h, and then the plate was inspected. The zone ofinhibition between the isolates was absent, which showed thatthey lacked competitive inhibition (Samuel et al., 2012). With thedeveloped ternary consortium, toxicity study of TiO2 NPs wascarried out.
2.5. Toxicity assessment
2.5.1. Experimental setupThe bacterial isolates from fresh water sediment (VIT lake
water, Vellore, India) were identified as B. alitudinis (gram positiverods), B. subtilis (gram positive rods), and P. aeruginosa (gramnegative rods) through 16S rRNA analysis (length of the sequence,1462 bp; homology, 99%).

J. Kumari et al. / Environmental Research 135 (2014) 333–345 335
All the experiments were carried out on the bacterial speciesand consortium. The consortium was developed with the help ofthe three isolates B. alitudinis, B. subtilis, and P. aeruginosa withequal cell number i.e., 0.1 optical density. The bacterial cells werefirst inoculated in the NB and in the exponential phase, the cellswere harvested by centrifugation at 7000g for 10 min. Finally, thecells were washed twice with the sterilized lake water to removegrowth media components. From standard plate count assay, theinitial bacterial cell number was found to be 2.78�107. Tomaintain accuracy, all the experiments were performed in tripli-cates, and the standard error was calculated. Individual bacterialisolates and the consortium were treated with four differentconcentrations of TiO2 NP dispersion (0.25, 0.50, 0.75, and1 μg/mL). All the biological experiments were carried out undervisible light intensity of 0.000439239 W/cm2 (TL–D Super 80Linear fluorescent tube, Philips, India) and dark (no irradiation)conditions. For light experiments, continuous irradiation wasprovided, and for dark experiments, samples were covered withopaque sheets without intervention of any visible light. Controlsamples without NPs (for each condition-visible light and dark)were also kept.
2.5.2. Cytotoxic effects of TiO2 NPs on freshwater sediment bacteriaTo evaluate the toxic effect of TiO2 NPs on individual isolates as
well on the consortium, cell viability assessment was done. Thenumber of cells in the control was considered to be 100%. Adecrease in the cell viability in test samples was calculated withrespect to the control. After the interaction period (2, 4, 6 and24 h), the percentage in reduction of cellular viability was mea-sured. The decrease in cellular viability in treated samples wascalculated with respect to the control. Cell viability was deter-mined using the standard plate count assay on NB agar medium.
2.6. Oxidative stress assessment
2.6.1. Determination of reactive oxygen species (ROS)Reactive oxygen species (ROS) generation was quantified with
the help of a fluorescence probe dichloro-dihydro-fluoresceindiacetate (DCFH-DA). For ROS determination, a nonpolar dye istransformed into a hydrophilic derivative DCFH by cellular es-terases that are non-fluorescent, but later switched to becomeextremely fluorescent DCF when oxidized by peroxidases andintracellular ROS. Intracellular ROS generation was monitored inthe control and NP-interacted bacterial cells at 2, 4, 6, and 24 hfollowing the protocol, as explained by Wang and Joseph (1999),with minor modifications. 5 mL of cell pellet was incubated withDCFH-DA with an end most concentration of 100 μM at 37 °C for30 min. Measurement of fluorescence was done using a spectro-fluorometer (SL174, ELICO) with excitation and emission wave-lengths of 485 nm and 530 nm, respectively. To estimate the auto-fluorescence activity of TiO2 NPs that may interfere with theDCFDA dye, a negative control of NPs without cells were alsoanalysed.
2.6.2. Superoxide dismutase assay (SOD)The superoxide dismutase (SOD) activity of interacted cells was
evaluated by the method described by Wintherbourn et al. (1975)using riboflavin as the O2 generator. This method was based on thepotential of superoxide dismutase to hinder the reduction of nitro-blue tetrazolium by superoxide, generated by the reaction ofphoto-reduced riboflavin with oxygen (Hossain et al., 2007).Interacted cells were centrifuged at 7000g for 10 min, and thepellet was collected and washed with PBS. Then, the cells werevortexed, centrifuged again at 7000g for 10 min, and the super-natant was collected. 0.1 M EDTA and 1.5 mM Nitro blue tetra-zolium (NBT) were added and kept in uniform light intensity.
Riboflavin was added and kept for incubation for a period of15 min at room temperature. To determine the SOD activity, theabsorbance values at 530 nm was recorded by UV–vis spectro-scopy (Systronics, India Ltd.).
2.7. Membrane permeability assessment (lactate dehydrogenaseassay)
LDH released during cell damage is a common marker ofcytotoxicity and membrane permeability. Bacterial cells wereinteracted for 24 h and centrifuged at 7000g for 10 min. The LDHlevel in the supernatant was measured following the standardprotocol (Brown et al., 2001). To 100 μL of the supernatant, 100 μLof 30 mM sodium pyruvate and 2.8 mL of 0.2 M Tris–HCl wereadded. 100 mL of 6.6 mM NADH was added prior to use. Theabsorbance at 340 nm was recorded using UV–vis spectroscopyto determine the LDH activity.
2.8. Extraction and quantification of exopolysaccharides (EPS)
The EPS production ability of the bacterial isolates and con-sortium were examined. A loop full culture of each isolate wasinoculated into 100 mL of NB medium. The bacteria and consor-tium were allowed to grow for 24 h with temperature maintainedat 30 °C, and shaking at 180 rpm. The cultures were then centri-fuged at 10,000g for 10 min and the supernatant of each culturewas harvested. To the collected supernatant, an equal volume ofethanol was added and kept at 4 °C overnight to precipitate theEPS. The obtained mixture was then centrifuged at 10,000g for30 min, and the pellets were collected. The pellets were thenwashed twice with ethanol and dialyzed against de-ionized water.The EPS collected was estimated for each sample by the phenol–sulphuric acid method (Dubois et al. 1956).
2.9. Microtiter plate biofilm production assay
Overnight cultures of all the three bacteria and their consor-tium were diluted to 1:100 in fresh media. Addition of 100 μL ofthe bacterial suspension to each well was done in a 96-well platecontaining 1 mg/mL TiO2 NPs (sonicated for 10 min) and incubatedfor 24 h at 30–37 °C in static conditions. The controls included acolumn of wells, each with a concentration of NPs in the culturemedium and NB medium without inoculum. Following incubation,the 96 microtitre wells were washed five times with distilledwater to remove non-attached cells and kept for drying at 37 °C for1 h. After that, staining was done with 1% crystal violet for 30 minat room temperature. Stained wells were washed five times withsterile distilled water, and the remaining crystal violet was elutedby the introduction of 95% ethanol solution for 15 min. The biofilmbiomass was then assessed by measuring the absorbance at590 nm. Static biofilm assay was performed three times forindividual bacteria and bacterial consortium, and the averagesand standard error were calculated for the repetitions of all theexperiment. All values were recorded after subtraction of thebackground absorbance of wells containing NPs (Ammendoliaet al., 2014).
2.10. Microscopic analysis
2.10.1. Distinguishing between live and dead bacteria throughfluorescence microscopy
Membrane permeability of treated cells and untreated cells wasobserved by fluorescence microscopy (Leica, DM-2500) after 2 h ofTiO2 NP interaction. Cells were stained with AO and EtBr byfollowing the protocol described by Jakopec et al. (2006) withminor modifications. In 500 μL of bacterial suspension, 4 μL of AO

J. Kumari et al. / Environmental Research 135 (2014) 333–345336
(15 μg/mL in PBS) and 4 μL of EtBr (50 μg/mL in PBS) were added.The cell suspension was incubated for 5 min followed by centri-fugation, and the supernatant was discarded to remove anyunbound dye. The cell pellet was re-suspended in 500 μL PBS.Dark condition was maintained to avoid photo-bleaching of dyes.Through the BP 450–490, LP 590 filter fluorescence was detected;images were captured with a Leica-DFC-295 camera and processedusing Leica-Application Suite 3.8.
2.10.2. Scanning electron microscopy (SEM)The changes in surface elemental composition and the mor-
phology of consortium were analysed using SEM–EDX. Sampleswere collected after 2 h of incubation, washed, and fixed on glassslides using 2% glutaraldehyde for 1.5 h. Then, it was washed by aseries of dehydration (20%, 40%, 60%, 80%, and 100%) ethanol, andthen air-dried. With carbon tape, the glass slide was fixed on thespecimen mount. Gold sputtering was carried out in an argonatmosphere (Paul et al., 2012). The surface morphology of bacteriaand the surface elemental composition were observed under SEM–
EDX (S-400, HITACHI, and Tokyo, Japan; JEOL JSM-5510). The SEMimages and EDX spectra of bacterial consortium were obtained at40,000� magnification.
2.10.3. Scanning electron microscopy of the biofilmSamples for scanning electron microscopy were prepared as
follows: 24-well plates were inoculated with bacterial consortiumin NB medium containing or not containing TiO2 NPs at aconcentration of 1 μg/mL. After 24 h incubation, the samples werewashed three times with phosphate buffered saline (PBS) andfixed with 2%glutaraldehyde. Dehydratation was done in ethanol–water mixture with increasing ethanol concentrations (20%, 40%,60%, 80%, and 100%) and air-dried overnight (Ammendolia et al.,2014).
2.10.4. Transmission electron microscopy (TEM)Consortium of all three isolates was collected after 2 h of
incubation with TiO2 NPs and centrifuged; the pellet was collectedand a thin section of cells was taken on a copper grid for TEMexamination. Changes in cellular structure and internalization ofNPs in bacterial consortium were observed by transmissionelectron microscopy (TEM) (Philips CM12 Transmission ElectronMicroscope, Netherlands).
2.11. Surface interactions with NPs
2.11.1. Surface charge: zeta potential measurementSurface charge may play a key role in bacteria–NP interaction.
The charge distribution on bacterial cell as well as surfacepotential of NP dispersion in lake water (under both light anddark conditions, 2 h) was measured using 90 plus Particle SizeAnalyzer with Zeta facility (Brookhaven Instruments Corporations,USA).
2.11.2. Surface chemistry: FT-IR analysisFT-IR analysis was carried out to study the presence of surface
chemical characteristics of TiO2-interacted bacterial cells. After a2 h interaction period, the cell suspension was centrifuged for15 min at 7000g. The pellet was washed twice with 1 mL phos-phate-buffered saline and lyophilized to remove moisture. Thedried cells were then subjected to FT-IR analysis by the potassiumbromide technique (Samuel et al., 2012).
2.12. Gene expression study using reverse transcriptase PCR
2.12.1. Isolation of total RNATotal RNAwas isolated from individual bacteria and consortium
(treated and untreated) using a simple phenol method (Atshanet al., 2012). Cultures were centrifuged at 8000g for 3 min in arefrigerated centrifuge at 4 °C. The pellet was resuspended in100 mL of RNase-free water. The tube was vigorously vortexed for3 min and 100 mL of acid phenol was added with chloroform (1:1).Samples were vortexed for 1 min and incubated at 70 °C for30 min. The vortex process was repeated every 5 min periodically.Subsequently, the samples were centrifuged at 12,000g for 10 minand 100 mL from the aqueous phase was transferred into a freshtube. A lysis buffer (700 mL) was added to the aqueous phase,followed by the addition of 200 mL of isopropanol to precipitatethe RNA. The tubes were vigorously vortexed for 3 min andcentrifuged at 12,000g for 10 min. RNA pellets were washed with70% ethanol, air-dried, and dissolved in 25 mL of RNase-free water.RNA preparations were analysed in agarose gel and quantifiedspectrophotometrically.
2.12.2. Primer designCoding sequences of epsG, ymcA (B. subtilis), and algF (P.
aeruginosa) were obtained from NCBI microbial genome database.Primers for PCR were designed using primer 3 software andvalidated using Primer-BLAST of NCBI. Primers were synthesizedat BioServe India Private Limited, Hyderabad, India. The sequencesof primers are given in Table S2.
2.12.3. Reverse transcriptase PCR analysisReaction mixture for cDNA synthesis contained RNase-free
water (5.5 mL), 5� reverse transcriptase buffer (4 mL), 10 mM dNTP4 mL [(1 mL each) (2 mM)], 10 mM DTT [1 mL (0.5 mM)], gene-specific reverse primer [4 mL (0.5 mM)], RNase inhibitor (0.5 mL),reverse transcriptase (AMV) [1 mL (5 Units)], and RNA template[5 mL (200 ng)] making up the total reaction volume of 20 mL. cDNAsynthesis was carried out at 42 °C for 1 h, after which the reactionwas stopped at 70 °C for 10 min and chilling on ice. The cDNAssynthesized were amplified with gene-specific forward and re-verse primers. The reaction mixture contained cDNA [6 mL(100 ng)], forward primer [2 mL (0.5 mM)], reverse primer [2 mL(0.5 mM)], PCR master mix (2� ) [10 mL (Taq polymerase 2 Units)],and sterile water [5 mL], making up a final volume of 25 mL. PCRwas carried out in a thermal cycler (Eppendorf, Germany) for 30cycles with an initial one-time denaturation at 95 °C for 2 min. Thesteps involved: Denaturation at 95 °C for 1 min, annealing at 55 °C(for epsG and ymcA) and 57 °C (for algF) for 1 min, and synthesis at72 °C for 1.5 min followed by final extension at 72 °C for 7 min. Theamplified products were analysed in 1% agarose gel electrophor-esis and visualized in gel documentation (Gel Doc™ EZ System, BioRad, USA).
Bacterial 16s rRNA was used as internal control and 200 ng oftotal RNA was used to perform the Reverse Transcriptase-PCRanalysis. Bacterial universal primers were used and the sequencesare given in Table S2. The denaturation and extension steps weresame as the test genes. The annealing temperature used forinternal control was 49 °C. Reverse-Transcriptase-PCR analysis ofinternal control gene was done separately for bacteria withoutnanoparticles treatment and the cultures with treatment. The PCRproducts were analysed separately in two agarose gels.
2.13. Statistical analysis
All in vitro toxicity tests were carried out in triplicates and thedata are given as mean7standard error. The data were processedusing one-way ANOVA, followed by Dunnett's post-hoc test with a
J. Kumari et al. / Environmental Research 135 (2014) 333–345 337
p value o0.05 for a standard plate count assay. The data for theROS, SOD, and LDH assay were processed through Student's t-testat po0.05.
3. Results
3.1. TiO2 NP stability in lake water
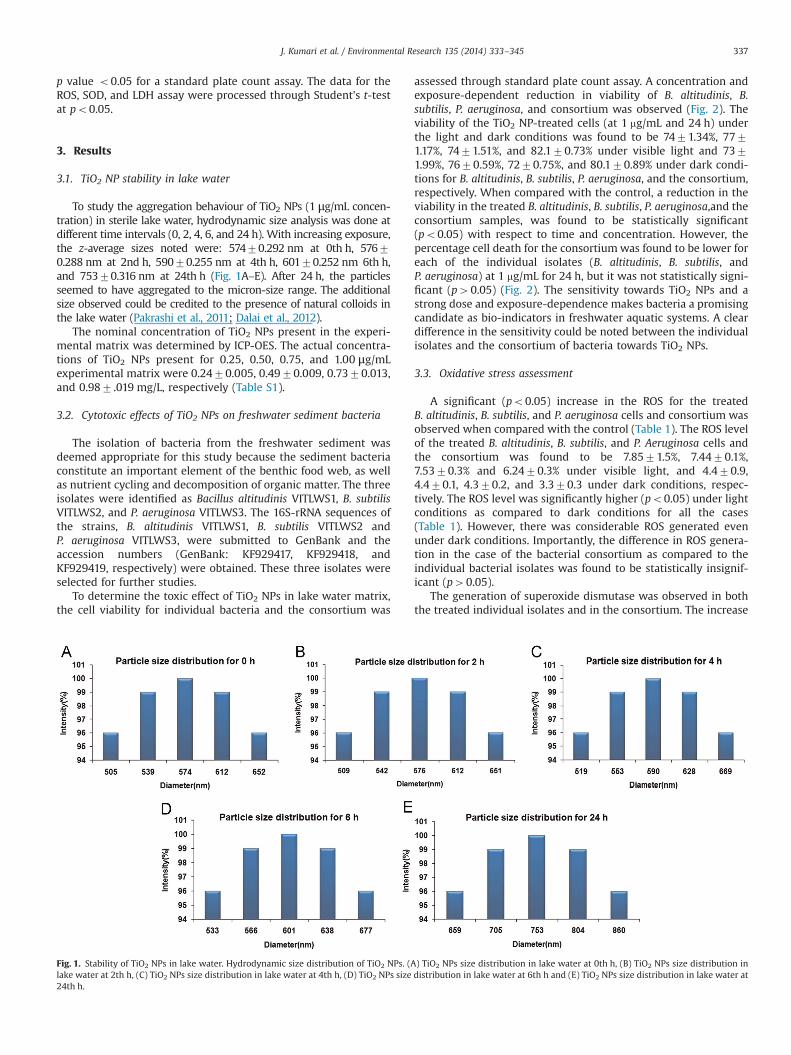
To study the aggregation behaviour of TiO2 NPs (1 μg/mL concen-tration) in sterile lake water, hydrodynamic size analysis was done atdifferent time intervals (0, 2, 4, 6, and 24 h). With increasing exposure,the z-average sizes noted were: 57470.292 nm at 0th h, 57670.288 nm at 2nd h, 59070.255 nm at 4th h, 60170.252 nm 6th h,and 75370.316 nm at 24th h (Fig. 1A–E). After 24 h, the particlesseemed to have aggregated to the micron-size range. The additionalsize observed could be credited to the presence of natural colloids inthe lake water (Pakrashi et al., 2011; Dalai et al., 2012).
The nominal concentration of TiO2 NPs present in the experi-mental matrix was determined by ICP-OES. The actual concentra-tions of TiO2 NPs present for 0.25, 0.50, 0.75, and 1.00 μg/mLexperimental matrix were 0.2470.005, 0.4970.009, 0.7370.013,and 0.987 .019 mg/L, respectively (Table S1).
3.2. Cytotoxic effects of TiO2 NPs on freshwater sediment bacteria
The isolation of bacteria from the freshwater sediment wasdeemed appropriate for this study because the sediment bacteriaconstitute an important element of the benthic food web, as wellas nutrient cycling and decomposition of organic matter. The threeisolates were identified as Bacillus altitudinis VITLWS1, B. subtilisVITLWS2, and P. aeruginosa VITLWS3. The 16S-rRNA sequences ofthe strains, B. altitudinis VITLWS1, B. subtilis VITLWS2 andP. aeruginosa VITLWS3, were submitted to GenBank and theaccession numbers (GenBank: KF929417, KF929418, andKF929419, respectively) were obtained. These three isolates wereselected for further studies.
To determine the toxic effect of TiO2 NPs in lake water matrix,the cell viability for individual bacteria and the consortium was
Fig. 1. Stability of TiO2 NPs in lake water. Hydrodynamic size distribution of TiO2 NPs. (Alake water at 2th h, (C) TiO2 NPs size distribution in lake water at 4th h, (D) TiO2 NPs size24th h.
assessed through standard plate count assay. A concentration andexposure-dependent reduction in viability of B. altitudinis, B.subtilis, P. aeruginosa, and consortium was observed (Fig. 2). Theviability of the TiO2 NP-treated cells (at 1 mg/mL and 24 h) underthe light and dark conditions was found to be 7471.34%, 7771.17%, 7471.51%, and 82.170.73% under visible light and 7371.99%, 7670.59%, 7270.75%, and 80.170.89% under dark condi-tions for B. altitudinis, B. subtilis, P. aeruginosa, and the consortium,respectively. When compared with the control, a reduction in theviability in the treated B. altitudinis, B. subtilis, P. aeruginosa,and theconsortium samples, was found to be statistically significant(po0.05) with respect to time and concentration. However, thepercentage cell death for the consortiumwas found to be lower foreach of the individual isolates (B. altitudinis, B. subtilis, andP. aeruginosa) at 1 mg/mL for 24 h, but it was not statistically signi-ficant (p40.05) (Fig. 2). The sensitivity towards TiO2 NPs and astrong dose and exposure-dependence makes bacteria a promisingcandidate as bio-indicators in freshwater aquatic systems. A cleardifference in the sensitivity could be noted between the individualisolates and the consortium of bacteria towards TiO2 NPs.
3.3. Oxidative stress assessment
A significant (po0.05) increase in the ROS for the treatedB. altitudinis, B. subtilis, and P. aeruginosa cells and consortium wasobserved when compared with the control (Table 1). The ROS levelof the treated B. altitudinis, B. subtilis, and P. Aeruginosa cells andthe consortium was found to be 7.8571.5%, 7.4470.1%,7.5370.3% and 6.2470.3% under visible light, and 4.470.9,4.470.1, 4.370.2, and 3.370.3 under dark conditions, respec-tively. The ROS level was significantly higher (po0.05) under lightconditions as compared to dark conditions for all the cases(Table 1). However, there was considerable ROS generated evenunder dark conditions. Importantly, the difference in ROS genera-tion in the case of the bacterial consortium as compared to theindividual bacterial isolates was found to be statistically insignif-icant (p40.05).
The generation of superoxide dismutase was observed in boththe treated individual isolates and in the consortium. The increase
) TiO2 NPs size distribution in lake water at 0th h, (B) TiO2 NPs size distribution indistribution in lake water at 6th h and (E) TiO2 NPs size distribution in lake water at
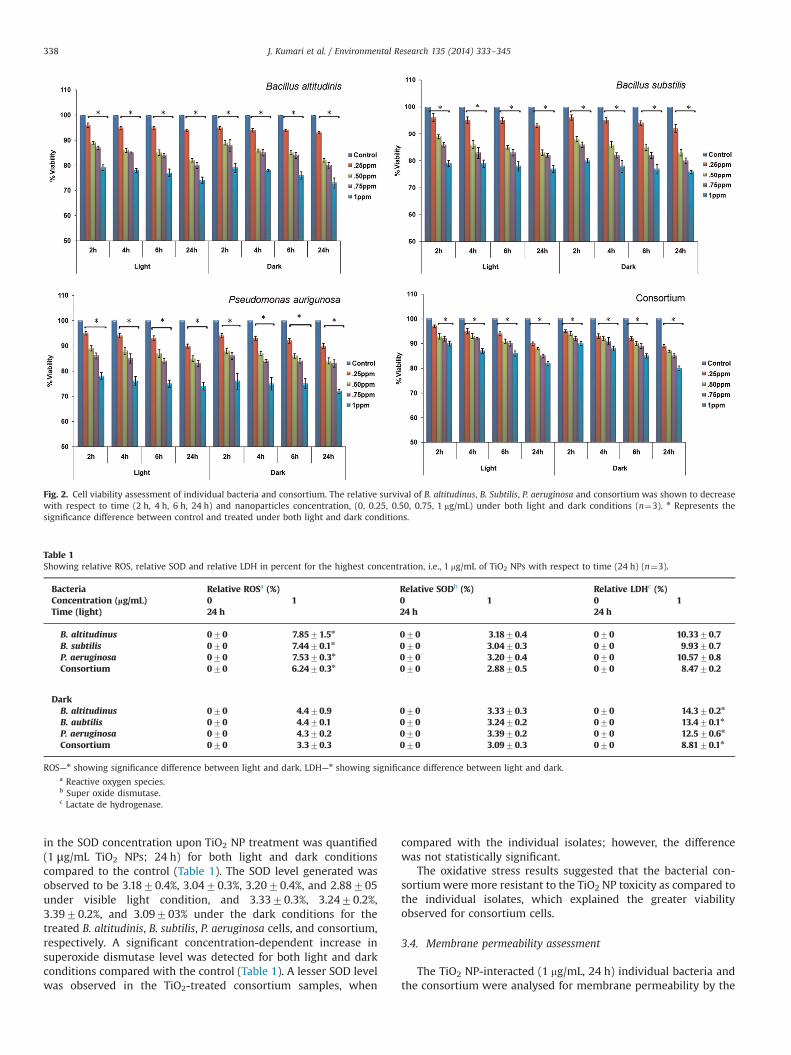
Fig. 2. Cell viability assessment of individual bacteria and consortium. The relative survival of B. altitudinus, B. Subtilis, P. aeruginosa and consortium was shown to decreasewith respect to time (2 h, 4 h, 6 h, 24 h) and nanoparticles concentration, (0, 0.25, 0.50, 0.75, 1 mg/mL) under both light and dark conditions (n¼3). n Represents thesignificance difference between control and treated under both light and dark conditions.
Table 1Showing relative ROS, relative SOD and relative LDH in percent for the highest concentration, i.e., 1 mg/mL of TiO2 NPs with respect to time (24 h) (n¼3).
Bacteria Relative ROSa (%) Relative SODb (%) Relative LDHc (%)Concentration (lg/mL) 0 1 0 1 0 1Time (light) 24 h 24 h 24 h
B. altitudinus 070 7.8571.5n 070 3.1870.4 070 10.3370.7B. subtilis 070 7.4470.1n 070 3.0470.3 070 9.9370.7P. aeruginosa 070 7.5370.3n 070 3.2070.4 070 10.5770.8Consortium 070 6.2470.3n 070 2.8870.5 070 8.4770.2
DarkB. altitudinus 070 4.470.9 070 3.3370.3 070 14.370.2n
B. aubtilis 070 4.470.1 070 3.2470.2 070 13.470.1n
P. aeruginosa 070 4.370.2 070 3.3970.2 070 12.570.6n
Consortium 070 3.370.3 070 3.0970.3 070 8.8170.1n
ROS—n showing significance difference between light and dark. LDH—n showing significance difference between light and dark.a Reactive oxygen species.b Super oxide dismutase.c Lactate de hydrogenase.
J. Kumari et al. / Environmental Research 135 (2014) 333–345338
in the SOD concentration upon TiO2 NP treatment was quantified(1 μg/mL TiO2 NPs; 24 h) for both light and dark conditionscompared to the control (Table 1). The SOD level generated wasobserved to be 3.1870.4%, 3.0470.3%, 3.2070.4%, and 2.88705under visible light condition, and 3.3370.3%, 3.2470.2%,3.3970.2%, and 3.09703% under the dark conditions for thetreated B. altitudinis, B. subtilis, P. aeruginosa cells, and consortium,respectively. A significant concentration-dependent increase insuperoxide dismutase level was detected for both light and darkconditions compared with the control (Table 1). A lesser SOD levelwas observed in the TiO2-treated consortium samples, when
compared with the individual isolates; however, the differencewas not statistically significant.
The oxidative stress results suggested that the bacterial con-sortiumwere more resistant to the TiO2 NP toxicity as compared tothe individual isolates, which explained the greater viabilityobserved for consortium cells.
3.4. Membrane permeability assessment
The TiO2 NP-interacted (1 mg/mL, 24 h) individual bacteria andthe consortium were analysed for membrane permeability by the
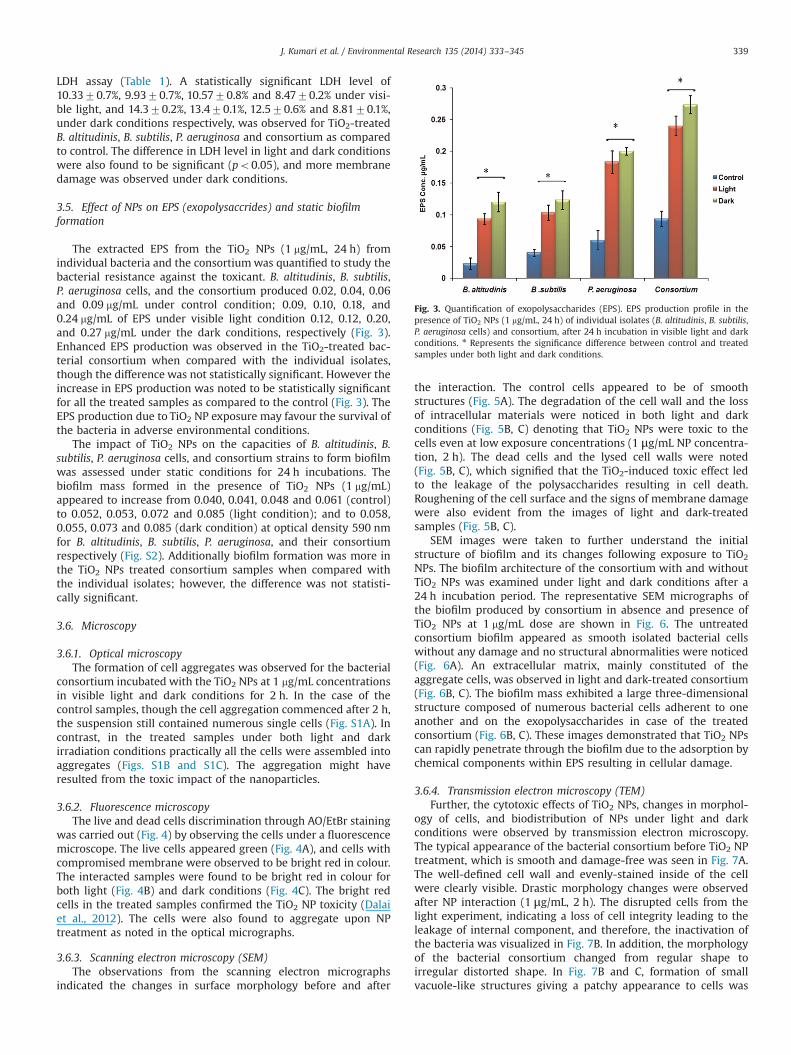
Fig. 3. Quantification of exopolysaccharides (EPS). EPS production profile in thepresence of TiO2 NPs (1 mg/mL, 24 h) of individual isolates (B. altitudinis, B. subtilis,P. aeruginosa cells) and consortium, after 24 h incubation in visible light and darkconditions. n Represents the significance difference between control and treatedsamples under both light and dark conditions.
J. Kumari et al. / Environmental Research 135 (2014) 333–345 339
LDH assay (Table 1). A statistically significant LDH level of10.3370.7%, 9.9370.7%, 10.5770.8% and 8.4770.2% under visi-ble light, and 14.370.2%, 13.470.1%, 12.570.6% and 8.8170.1%,under dark conditions respectively, was observed for TiO2-treatedB. altitudinis, B. subtilis, P. aeruginosa and consortium as comparedto control. The difference in LDH level in light and dark conditionswere also found to be significant (po0.05), and more membranedamage was observed under dark conditions.
3.5. Effect of NPs on EPS (exopolysaccrides) and static biofilmformation
The extracted EPS from the TiO2 NPs (1 mg/mL, 24 h) fromindividual bacteria and the consortiumwas quantified to study thebacterial resistance against the toxicant. B. altitudinis, B. subtilis,P. aeruginosa cells, and the consortium produced 0.02, 0.04, 0.06and 0.09 mg/mL under control condition; 0.09, 0.10, 0.18, and0.24 mg/mL of EPS under visible light condition 0.12, 0.12, 0.20,and 0.27 mg/mL under the dark conditions, respectively (Fig. 3).Enhanced EPS production was observed in the TiO2-treated bac-terial consortium when compared with the individual isolates,though the difference was not statistically significant. However theincrease in EPS production was noted to be statistically significantfor all the treated samples as compared to the control (Fig. 3). TheEPS production due to TiO2 NP exposure may favour the survival ofthe bacteria in adverse environmental conditions.
The impact of TiO2 NPs on the capacities of B. altitudinis, B.subtilis, P. aeruginosa cells, and consortium strains to form biofilmwas assessed under static conditions for 24 h incubations. Thebiofilm mass formed in the presence of TiO2 NPs (1 mg/mL)appeared to increase from 0.040, 0.041, 0.048 and 0.061 (control)to 0.052, 0.053, 0.072 and 0.085 (light condition); and to 0.058,0.055, 0.073 and 0.085 (dark condition) at optical density 590 nmfor B. altitudinis, B. subtilis, P. aeruginosa, and their consortiumrespectively (Fig. S2). Additionally biofilm formation was more inthe TiO2 NPs treated consortium samples when compared withthe individual isolates; however, the difference was not statisti-cally significant.
3.6. Microscopy
3.6.1. Optical microscopyThe formation of cell aggregates was observed for the bacterial
consortium incubated with the TiO2 NPs at 1 mg/mL concentrationsin visible light and dark conditions for 2 h. In the case of thecontrol samples, though the cell aggregation commenced after 2 h,the suspension still contained numerous single cells (Fig. S1A). Incontrast, in the treated samples under both light and darkirradiation conditions practically all the cells were assembled intoaggregates (Figs. S1B and S1C). The aggregation might haveresulted from the toxic impact of the nanoparticles.
3.6.2. Fluorescence microscopyThe live and dead cells discrimination through AO/EtBr staining
was carried out (Fig. 4) by observing the cells under a fluorescencemicroscope. The live cells appeared green (Fig. 4A), and cells withcompromised membrane were observed to be bright red in colour.The interacted samples were found to be bright red in colour forboth light (Fig. 4B) and dark conditions (Fig. 4C). The bright redcells in the treated samples confirmed the TiO2 NP toxicity (Dalaiet al., 2012). The cells were also found to aggregate upon NPtreatment as noted in the optical micrographs.
3.6.3. Scanning electron microscopy (SEM)The observations from the scanning electron micrographs
indicated the changes in surface morphology before and after
the interaction. The control cells appeared to be of smoothstructures (Fig. 5A). The degradation of the cell wall and the lossof intracellular materials were noticed in both light and darkconditions (Fig. 5B, C) denoting that TiO2 NPs were toxic to thecells even at low exposure concentrations (1 μg/mL NP concentra-tion, 2 h). The dead cells and the lysed cell walls were noted(Fig. 5B, C), which signified that the TiO2-induced toxic effect ledto the leakage of the polysaccharides resulting in cell death.Roughening of the cell surface and the signs of membrane damagewere also evident from the images of light and dark-treatedsamples (Fig. 5B, C).
SEM images were taken to further understand the initialstructure of biofilm and its changes following exposure to TiO2
NPs. The biofilm architecture of the consortium with and withoutTiO2 NPs was examined under light and dark conditions after a24 h incubation period. The representative SEM micrographs ofthe biofilm produced by consortium in absence and presence ofTiO2 NPs at 1 mg/mL dose are shown in Fig. 6. The untreatedconsortium biofilm appeared as smooth isolated bacterial cellswithout any damage and no structural abnormalities were noticed(Fig. 6A). An extracellular matrix, mainly constituted of theaggregate cells, was observed in light and dark-treated consortium(Fig. 6B, C). The biofilm mass exhibited a large three-dimensionalstructure composed of numerous bacterial cells adherent to oneanother and on the exopolysaccharides in case of the treatedconsortium (Fig. 6B, C). These images demonstrated that TiO2 NPscan rapidly penetrate through the biofilm due to the adsorption bychemical components within EPS resulting in cellular damage.
3.6.4. Transmission electron microscopy (TEM)Further, the cytotoxic effects of TiO2 NPs, changes in morphol-
ogy of cells, and biodistribution of NPs under light and darkconditions were observed by transmission electron microscopy.The typical appearance of the bacterial consortium before TiO2 NPtreatment, which is smooth and damage-free was seen in Fig. 7A.The well-defined cell wall and evenly-stained inside of the cellwere clearly visible. Drastic morphology changes were observedafter NP interaction (1 μg/mL, 2 h). The disrupted cells from thelight experiment, indicating a loss of cell integrity leading to theleakage of internal component, and therefore, the inactivation ofthe bacteria was visualized in Fig. 7B. In addition, the morphologyof the bacterial consortium changed from regular shape toirregular distorted shape. In Fig. 7B and C, formation of smallvacuole-like structures giving a patchy appearance to cells was
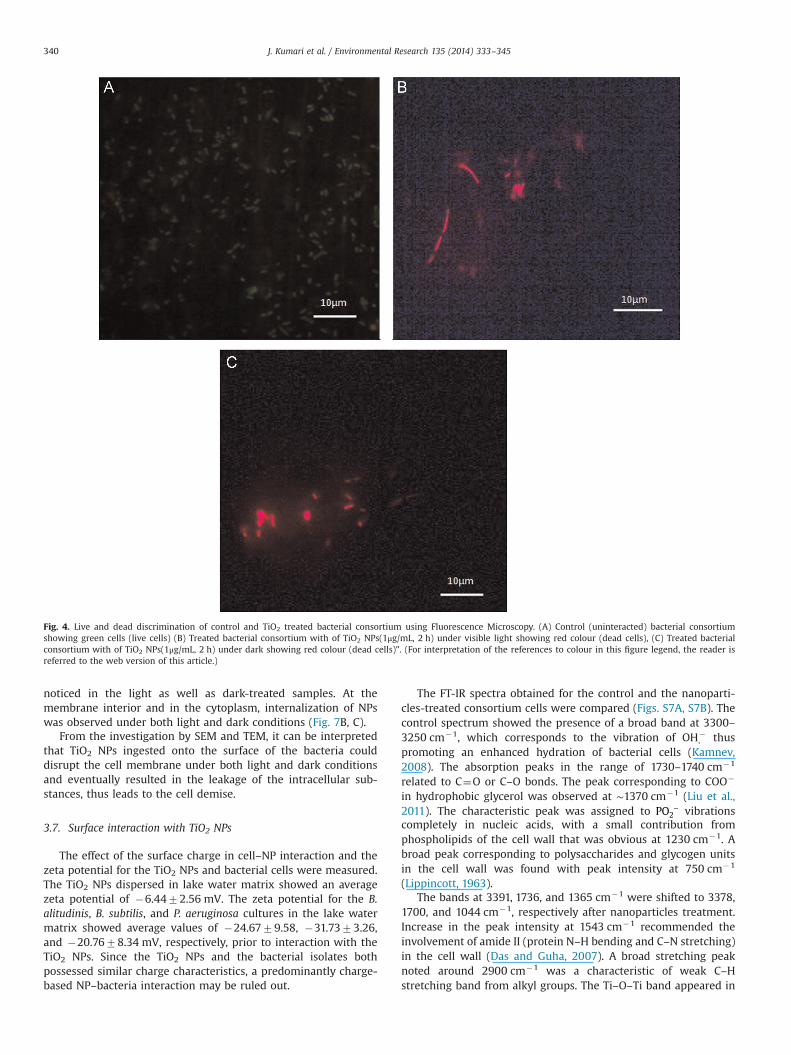
Fig. 4. Live and dead discrimination of control and TiO2 treated bacterial consortium using Fluorescence Microscopy. (A) Control (uninteracted) bacterial consortiumshowing green cells (live cells) (B) Treated bacterial consortium with of TiO2 NPs(1μg/mL, 2 h) under visible light showing red colour (dead cells), (C) Treated bacterialconsortium with of TiO2 NPs(1μg/mL, 2 h) under dark showing red colour (dead cells)". (For interpretation of the references to colour in this figure legend, the reader isreferred to the web version of this article.)
J. Kumari et al. / Environmental Research 135 (2014) 333–345340
noticed in the light as well as dark-treated samples. At themembrane interior and in the cytoplasm, internalization of NPswas observed under both light and dark conditions (Fig. 7B, C).
From the investigation by SEM and TEM, it can be interpretedthat TiO2 NPs ingested onto the surface of the bacteria coulddisrupt the cell membrane under both light and dark conditionsand eventually resulted in the leakage of the intracellular sub-stances, thus leads to the cell demise.
3.7. Surface interaction with TiO2 NPs
The effect of the surface charge in cell–NP interaction and thezeta potential for the TiO2 NPs and bacterial cells were measured.The TiO2 NPs dispersed in lake water matrix showed an averagezeta potential of �6.4472.56 mV. The zeta potential for the B.alitudinis, B. subtilis, and P. aeruginosa cultures in the lake watermatrix showed average values of �24.6779.58, �31.7373.26,and �20.7678.34 mV, respectively, prior to interaction with theTiO2 NPs. Since the TiO2 NPs and the bacterial isolates bothpossessed similar charge characteristics, a predominantly charge-based NP–bacteria interaction may be ruled out.
The FT-IR spectra obtained for the control and the nanoparti-cles-treated consortium cells were compared (Figs. S7A, S7B). Thecontrol spectrum showed the presence of a broad band at 3300–3250 cm�1, which corresponds to the vibration of OH�
, thuspromoting an enhanced hydration of bacterial cells (Kamnev,2008). The absorption peaks in the range of 1730–1740 cm�1
related to C¼O or C–O bonds. The peak corresponding to COO�
in hydrophobic glycerol was observed at ∼1370 cm�1 (Liu et al.,2011). The characteristic peak was assigned to −PO2 vibrationscompletely in nucleic acids, with a small contribution fromphospholipids of the cell wall that was obvious at 1230 cm�1. Abroad peak corresponding to polysaccharides and glycogen unitsin the cell wall was found with peak intensity at 750 cm�1
(Lippincott, 1963).The bands at 3391, 1736, and 1365 cm�1 were shifted to 3378,
1700, and 1044 cm�1, respectively after nanoparticles treatment.Increase in the peak intensity at 1543 cm�1 recommended theinvolvement of amide II (protein N–H bending and C–N stretching)in the cell wall (Das and Guha, 2007). A broad stretching peaknoted around 2900 cm�1 was a characteristic of weak C–Hstretching band from alkyl groups. The Ti–O–Ti band appeared in
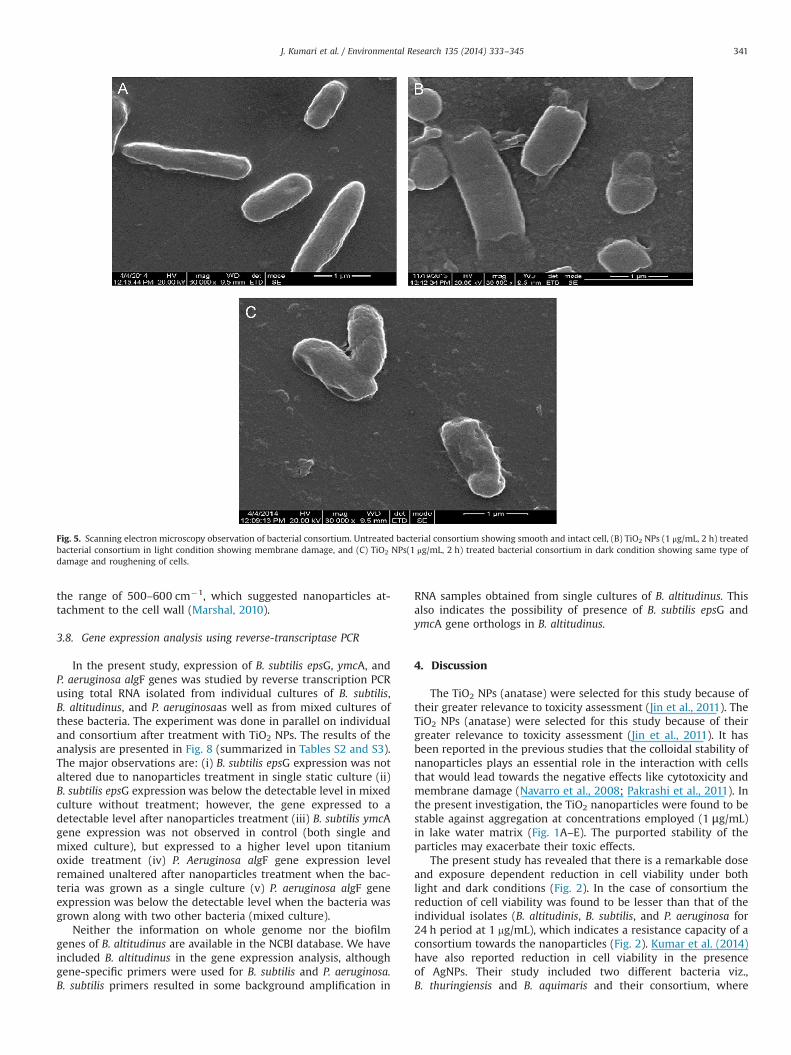
Fig. 5. Scanning electron microscopy observation of bacterial consortium. Untreated bacterial consortium showing smooth and intact cell, (B) TiO2 NPs (1 μg/mL, 2 h) treatedbacterial consortium in light condition showing membrane damage, and (C) TiO2 NPs(1 μg/mL, 2 h) treated bacterial consortium in dark condition showing same type ofdamage and roughening of cells.
J. Kumari et al. / Environmental Research 135 (2014) 333–345 341
the range of 500–600 cm�1, which suggested nanoparticles at-tachment to the cell wall (Marshal, 2010).
3.8. Gene expression analysis using reverse-transcriptase PCR
In the present study, expression of B. subtilis epsG, ymcA, andP. aeruginosa algF genes was studied by reverse transcription PCRusing total RNA isolated from individual cultures of B. subtilis,B. altitudinus, and P. aeruginosaas well as from mixed cultures ofthese bacteria. The experiment was done in parallel on individualand consortium after treatment with TiO2 NPs. The results of theanalysis are presented in Fig. 8 (summarized in Tables S2 and S3).The major observations are: (i) B. subtilis epsG expression was notaltered due to nanoparticles treatment in single static culture (ii)B. subtilis epsG expression was below the detectable level in mixedculture without treatment; however, the gene expressed to adetectable level after nanoparticles treatment (iii) B. subtilis ymcAgene expression was not observed in control (both single andmixed culture), but expressed to a higher level upon titaniumoxide treatment (iv) P. Aeruginosa algF gene expression levelremained unaltered after nanoparticles treatment when the bac-teria was grown as a single culture (v) P. aeruginosa algF geneexpression was below the detectable level when the bacteria wasgrown along with two other bacteria (mixed culture).
Neither the information on whole genome nor the biofilmgenes of B. altitudinus are available in the NCBI database. We haveincluded B. altitudinus in the gene expression analysis, althoughgene-specific primers were used for B. subtilis and P. aeruginosa.B. subtilis primers resulted in some background amplification in
RNA samples obtained from single cultures of B. altitudinus. Thisalso indicates the possibility of presence of B. subtilis epsG andymcA gene orthologs in B. altitudinus.
4. Discussion
The TiO2 NPs (anatase) were selected for this study because oftheir greater relevance to toxicity assessment (Jin et al., 2011). TheTiO2 NPs (anatase) were selected for this study because of theirgreater relevance to toxicity assessment (Jin et al., 2011). It hasbeen reported in the previous studies that the colloidal stability ofnanoparticles plays an essential role in the interaction with cellsthat would lead towards the negative effects like cytotoxicity andmembrane damage (Navarro et al., 2008; Pakrashi et al., 2011). Inthe present investigation, the TiO2 nanoparticles were found to bestable against aggregation at concentrations employed (1 μg/mL)in lake water matrix (Fig. 1A–E). The purported stability of theparticles may exacerbate their toxic effects.
The present study has revealed that there is a remarkable doseand exposure dependent reduction in cell viability under bothlight and dark conditions (Fig. 2). In the case of consortium thereduction of cell viability was found to be lesser than that of theindividual isolates (B. altitudinis, B. subtilis, and P. aeruginosa for24 h period at 1 mg/mL), which indicates a resistance capacity of aconsortium towards the nanoparticles (Fig. 2). Kumar et al. (2014)have also reported reduction in cell viability in the presenceof AgNPs. Their study included two different bacteria viz.,B. thuringiensis and B. aquimaris and their consortium, where
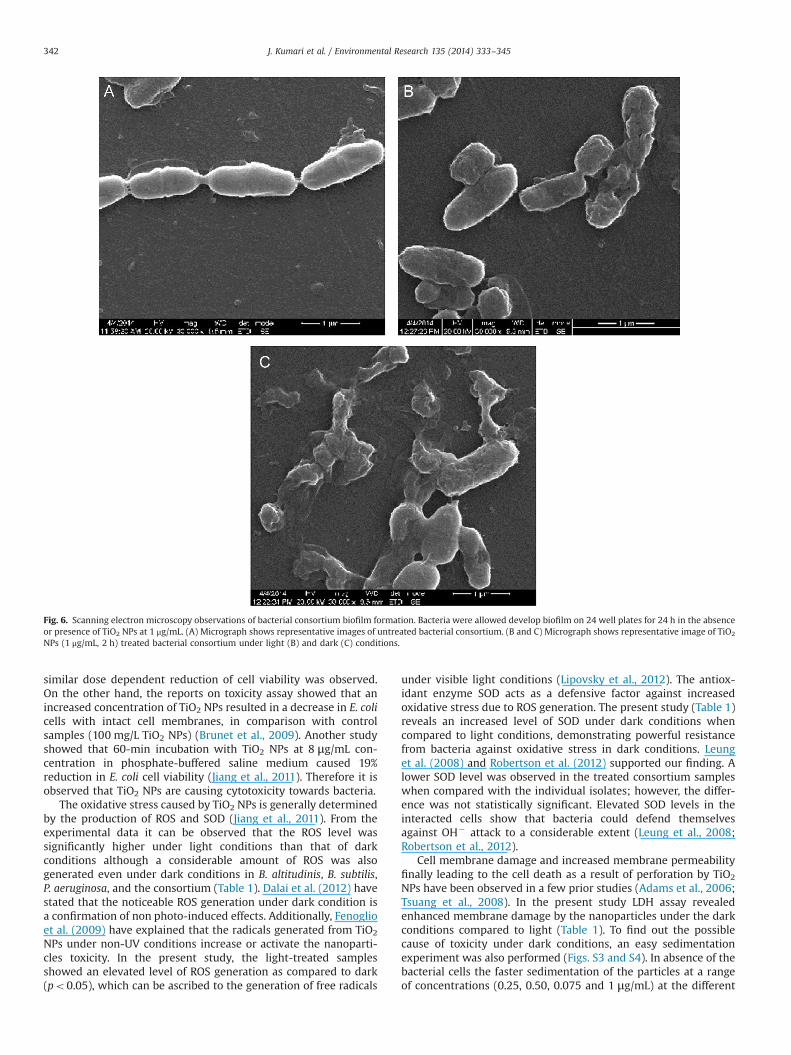
Fig. 6. Scanning electron microscopy observations of bacterial consortium biofilm formation. Bacteria were allowed develop biofilm on 24 well plates for 24 h in the absenceor presence of TiO2 NPs at 1 μg/mL. (A) Micrograph shows representative images of untreated bacterial consortium. (B and C) Micrograph shows representative image of TiO2
NPs (1 μg/mL, 2 h) treated bacterial consortium under light (B) and dark (C) conditions.
J. Kumari et al. / Environmental Research 135 (2014) 333–345342
similar dose dependent reduction of cell viability was observed.On the other hand, the reports on toxicity assay showed that anincreased concentration of TiO2 NPs resulted in a decrease in E. colicells with intact cell membranes, in comparison with controlsamples (100 mg/L TiO2 NPs) (Brunet et al., 2009). Another studyshowed that 60-min incubation with TiO2 NPs at 8 μg/mL con-centration in phosphate-buffered saline medium caused 19%reduction in E. coli cell viability (Jiang et al., 2011). Therefore it isobserved that TiO2 NPs are causing cytotoxicity towards bacteria.
The oxidative stress caused by TiO2 NPs is generally determinedby the production of ROS and SOD (Jiang et al., 2011). From theexperimental data it can be observed that the ROS level wassignificantly higher under light conditions than that of darkconditions although a considerable amount of ROS was alsogenerated even under dark conditions in B. altitudinis, B. subtilis,P. aeruginosa, and the consortium (Table 1). Dalai et al. (2012) havestated that the noticeable ROS generation under dark condition isa confirmation of non photo-induced effects. Additionally, Fenoglioet al. (2009) have explained that the radicals generated from TiO2
NPs under non-UV conditions increase or activate the nanoparti-cles toxicity. In the present study, the light-treated samplesshowed an elevated level of ROS generation as compared to dark(po0.05), which can be ascribed to the generation of free radicals
under visible light conditions (Lipovsky et al., 2012). The antiox-idant enzyme SOD acts as a defensive factor against increasedoxidative stress due to ROS generation. The present study (Table 1)reveals an increased level of SOD under dark conditions whencompared to light conditions, demonstrating powerful resistancefrom bacteria against oxidative stress in dark conditions. Leunget al. (2008) and Robertson et al. (2012) supported our finding. Alower SOD level was observed in the treated consortium sampleswhen compared with the individual isolates; however, the differ-ence was not statistically significant. Elevated SOD levels in theinteracted cells show that bacteria could defend themselvesagainst OH� attack to a considerable extent (Leung et al., 2008;Robertson et al., 2012).
Cell membrane damage and increased membrane permeabilityfinally leading to the cell death as a result of perforation by TiO2
NPs have been observed in a few prior studies (Adams et al., 2006;Tsuang et al., 2008). In the present study LDH assay revealedenhanced membrane damage by the nanoparticles under the darkconditions compared to light (Table 1). To find out the possiblecause of toxicity under dark conditions, an easy sedimentationexperiment was also performed (Figs. S3 and S4). In absence of thebacterial cells the faster sedimentation of the particles at a rangeof concentrations (0.25, 0.50, 0.075 and 1 μg/mL) at the different
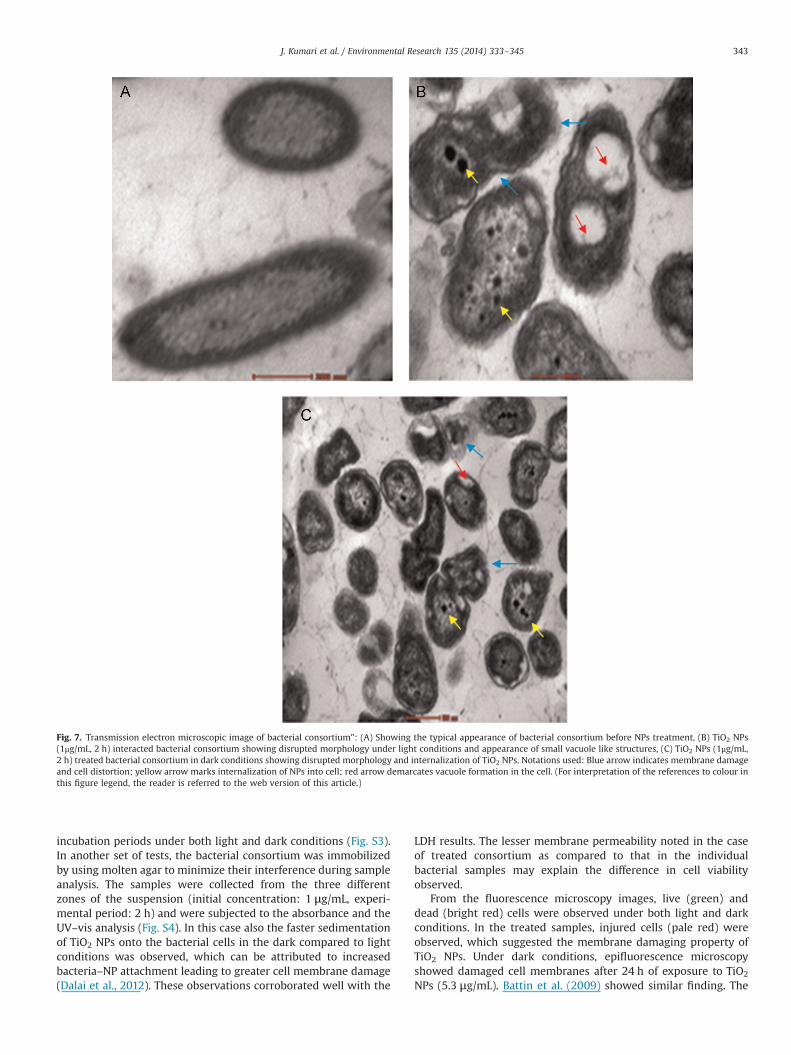
Fig. 7. Transmission electron microscopic image of bacterial consortium": (A) Showing the typical appearance of bacterial consortium before NPs treatment, (B) TiO2 NPs(1μg/mL, 2 h) interacted bacterial consortium showing disrupted morphology under light conditions and appearance of small vacuole like structures, (C) TiO2 NPs (1μg/mL,2 h) treated bacterial consortium in dark conditions showing disrupted morphology and internalization of TiO2 NPs. Notations used: Blue arrow indicates membrane damageand cell distortion; yellow arrow marks internalization of NPs into cell; red arrow demarcates vacuole formation in the cell. (For interpretation of the references to colour inthis figure legend, the reader is referred to the web version of this article.)
J. Kumari et al. / Environmental Research 135 (2014) 333–345 343
incubation periods under both light and dark conditions (Fig. S3).In another set of tests, the bacterial consortium was immobilizedby using molten agar to minimize their interference during sampleanalysis. The samples were collected from the three differentzones of the suspension (initial concentration: 1 μg/mL, experi-mental period: 2 h) and were subjected to the absorbance and theUV–vis analysis (Fig. S4). In this case also the faster sedimentationof TiO2 NPs onto the bacterial cells in the dark compared to lightconditions was observed, which can be attributed to increasedbacteria–NP attachment leading to greater cell membrane damage(Dalai et al., 2012). These observations corroborated well with the
LDH results. The lesser membrane permeability noted in the caseof treated consortium as compared to that in the individualbacterial samples may explain the difference in cell viabilityobserved.
From the fluorescence microscopy images, live (green) anddead (bright red) cells were observed under both light and darkconditions. In the treated samples, injured cells (pale red) wereobserved, which suggested the membrane damaging property ofTiO2 NPs. Under dark conditions, epifluorescence microscopyshowed damaged cell membranes after 24 h of exposure to TiO2
NPs (5.3 μg/mL). Battin et al. (2009) showed similar finding. The

Fig. 8. Reverse transcriptase-PCR analysis of epsG, ymcA and algF gene expressionbefore and after titanium oxide treatment in single and mixed cultures of bacteria.Lanes 1: Bacillus altitudinis-control; 2: Bacillus subtilis-control; 3: Pseudomonasaeruginosa-control; 4: consortium-control; 5: Bacillus altitudinis-treated; 6: Bacillussubtilis-treated; 7: Pseudomonas aeruginosa-treated; 8: Consortium-treated.
J. Kumari et al. / Environmental Research 135 (2014) 333–345344
alteration in surface morphology of the bacterial cells whenexposed to TiO2 NPs was studied through scanning electronmicroscopy. The evenness of the membrane was decreased afterthe NP interaction, and the cells were found to form agglomerates(Fig. 5). Previous report also mentioned about the membranelocalization of TiO2 NPs on bacteria being responsible for thecellular strain (Morones et al., 2005).
In our present investigation the internalization and distributionof NPs could be confirmed from the TEM images (Fig. 7). The totalNP uptake by bacterial consortium under light and dark conditionsafter 24 h incubation at concentrations 0.25, 0.50, 0.75, 1 mg/mLwas also quantified by ICP-OES. Comparing the uptake data (Figs.S5) with the toxicity results of bacterial consortium (Fig. 2) forvisible light and dark conditions, it can be inferred that the totaluptake of NPs have directly contributed to the toxicity observed.Similar morphological damage caused to E. coli by irradiating TiO2
was observed using TEM by Pigeot-Rémy et al., 2011. Our previousstudy (Pakrashi et al., 2011) on aluminium oxide nanoparticlestoxicity on B. licheniformis also supports the present findings. Theinternalization of the nanoparticles may have been facilitated bythe oxidative stress generation and the membrane damage asdiscussed in the preceding section.
The zeta potential measurements demonstrated no involve-ment of charge-based interaction between nanoparticles andbacteria. Jiang et al. (2009) also reported the similar findings.Nevertheless the surface attachment of the nanoparticles onto thebacterial cell membrane was also noted in SEM–EDX analysis ofthe TiO2 interacted bacterial consortium (Figs. S6). The FT-IRanalysis was done to find out the contribution of surface chemicalgroups in bacteria–NP interaction. Attachment of TiO2 NPs to thecell wall was noticeable from the characteristic Ti–O–Ti bandappearing in the range of 500–600 cm�1 (Marshal, 2010). Previousfindings on TiO2 NP toxicity also suggested participation of surfacefunctional groups in damage of cellular membrane after NPinteraction (Sadiq et al., 2010, 2011). The surface interactions ofthe nanoparticles with the bacterial cells may exacerbate themembrane damage and thus cellular uptake of these particles.
Moving beyond viability and oxidative stress, functional assess-ment of bacteria was performed by examining both the bacteriaand the consortium. EPS was commonly considered to be thestructural support for biofilm and is a mixture of macromolecules,including secreted proteins and polysaccharides. EPS productionand biofilm formation in the case of the bacterial consortium washigher as compared to the individual bacterial isolates in both lightand dark conditions, though the difference was found to bestatistically insignificant (p40.05). A previous report showed thatthe biofilm mass formed in the presence of TiO2 NPs appeared toincrease in LM2 cultures at all nanoparticles doses utilized in a
dose-dependent manner during 24 h incubations with significantvalues for 80, 8, and 0.8 mg/mL (Ammendolia et al., 2014). It hasbeen shown that the presence of iron, especially FeCl3, enhancesbiofilm formation in P. aeruginosa (Patriquin et al., 2008;Borcherding et al., 2014). Our previous study also reported thatEPS production increased with increasing Cr(VI) concentration inthe growth medium (Harish et al., 2012). Biofilm growth and EPSproduction may protect the bacteria against host defences and theactivity of antimicrobial agents. The gene epsG of B. subtilis is a keygene in the eps operon involved in exopolysaccharide productionfor the building of submerged biofilm (Vlamakis et al., 2013). Thegene ymcA is a part of a complex of the genes including yibF–ymcA–yaaT, which is the master regulator of sporulation, compe-tence, and biofilm formation of B. subtilis (Franklin et al., 2004).P. aeruginosa algF gene is known to involve in the architecture ofbiofilms and its biochemical function is to perform O-acetylationof alginate polysaccharide. This process is known to play animportant role in mucoid phenotype of the bacteria and furtherto form biofilms (Carabetta et al., 2013). The O-acetylation processis complex, requiring a periplasmic protein coded by aglF, atransmembrane protein coded by agll, and a type-II membraneprotein coded by algJ. Hence, the role of aglF in P. aeruginosabiofilm formation is indispensable. Both changes in biofilm for-mation and EPS production are supported by changes in geneexpression in TiO2 NP-exposed bacteria and consortium.
5. Conclusion
A remarkable toxicity effect of TiO2 NPs on the three freshwatersediment isolates viz., B. altitudinis, B. subtilis, P. aeruginosa as wellas their consortium was observed at low exposure concentrations(0.25, 0.50, 0.75 and 1.00 μg/mL). The present study clearlysupports the hypothesis that the consortium/ mixture of thedifferent bacterial species may be less affected than that ofindividual species in the presence of TiO2 NPs in the environment.The results also suggest that toxic effect of TiO2 NPs was stronglydependent on dose, duration, and the irradiation conditions.Enhanced EPS secretion and biofilm formation may protect thebacterial consortium against the nanoparticles generated stressconditions. However, further detailed studies need to be con-ducted to understand the range of physicochemical behaviour andtoxicity responses of environmental microbes with the differentenvironmentally relevant ultra-fine particles.
Acknowledgments
Wewould like to thank sophisticated instrument analysis (SAIF,IITM) for scanning electron microscope analysis, Indian Institute ofSciences, Bangalore for ICP-OES and Christian Medical College forTransmission electron microscopy. We acknowledge the researchgrant provided by LSRB-DRDO (Grant no. DLS/81/48222/LSRB-262/BTB/2012), Government of India for executing the current study.We are grateful to the anonymous reviewers for their commentsand suggestions to substantially improve the technical quality ofthe manuscript.
Appendix A. Supplementary information
Supplementary data associated with this article can be found inthe online version at http://dx.doi.org/10.1016/j.envres.2014.09.025.

J. Kumari et al. / Environmental Research 135 (2014) 333–345 345
References
Adams, L.K., Lyon, D.Y., Alvarez, P.J., 2006. Comparative eco-toxicity of nanoscaleTiO2, SiO2, and ZnO water suspensions. Water Res. 40 (19), 3527–3532.
Ammendolia, M.G., Iosi, F., De Berardis, B., Guccione, G., Superti, F., Conte, M.P.,Longhi, C., 2014. Listeria monocytogenes behaviour in presence of non-UV-irradiated titanium dioxide nanoparticles. PloS One 9 (1), e84986.
Atshan, S.S., Nor Shamsudin, M., Sekawi, Z., Lung, L.T.T., Hamat, R.A., Karunanidhi,A., Pei Pei, C., 2012. Prevalence of adhesion and regulation of biofilm-relatedgenes in different clones of Staphylococcus aureus. BioMed. Res. Int. 1, 1–4.
Battin, T.J., Kammer, F.V., Weilhartner, A., Ottofuelling, S., Hofmann, T., 2009.Nanostructured TiO2: transport behavior and effects on aquatic microbialcommunities under environmental conditions. Environ. Sci. Technol. 43 (21),8098–8104.
Borcherding, J., Baltrusaitis, J., Chen, H., Stebounova, L., Wu, C.M., Rubasinghege, G.,Comellas, A.P., 2014. Iron oxide nanoparticles induce Pseudomonas aeruginosagrowth, induce biofilm formation, and inhibit antimicrobial peptide function.Environ. Sci.: Nano 1 (2), 123–132.
Brown, D.M., Wilson, M.R., MacNee, W., Stone, V., Donaldson, K., 2001. Size-dependent proinflammatory effects of ultrafine polystyrene particles: a rolefor surface area and oxidative stress in the enhanced activity of ultrafines.Toxicol. Appl. Pharmacol. 175 (3), 191–199.
Brunet, L., Lyon, D.Y., Hotze, E.M., Alvarez, P.J., Wiesner, M.R., 2009. Comparativephotoactivity and antibacterial properties of C60 fullerenes and titaniumdioxide nanoparticles. Environ. Sci. Technol. 43 (12), 4355–4360.
Carabetta, V., Tanner, A.W., Greco, T.M., Defrancesco, M., Cristea, I.M., Dubnau, D.,2013. A complex of YibF, YmcA and YaaT regulates sporulation, competence andbiofilm formation by accelerating the phosphorylation of Spo0A. Mol. Micro-biol. 88, 283–300.
Dalai, S., Pakrashi, S., Kumar, R.S., Chandrasekaran, N., Mukherjee, A., 2012. Acomparative cytotoxicity study of TiO2 nanoparticles under light and darkconditions at low exposure concentrations. Toxicol. Res. 1 (2), 116–130.
Dalai, S., Pakrashi, S., Chakravarty, S., Hussain, S., Chandrasekaran, N., Mukherjee, A.,2014. Studies on interfacial interactions of TiO2 nanoparticles with bacterialcells under light and dark conditions. Bull. Mater. Sci. 37 (3), 371–381.
Das, S.K., Guha, A.K., 2007. Biosorption of chromium by Termitomyces clypeatus.Colloids Surf. B 60 (1), 46–54.
Dubois, M., Gilles, K.A., Hamilton, J.K., Rebers, P., Smith, F., 1956. Colorimetricmethod for determination of sugars and related substances. Anal. Chem. 28 (3),350–356.
Fenoglio, I., Greco, G., Livraghi, S., Fubini, B., 2009. Non‐UV‐induced radicalreactions at the surface of TiO2 nanoparticles that may trigger toxic responses.Chem. Eur. J. 15 (18), 4614–4621.
Franklin, M.J., Douthit, S.A., McClure, M.A., 2004. Evidence that the algI/algJ genecassette, required for O acetylation of Pseudomonas aeruginosa alginate, evolvedby lateral gene transfer. J. Bacteriol. 186 (14), 4759–4773.
Harish, R., Samuel, J., Mishra, R., Chandrasekaran, N., Mukherjee, A., 2012. Bio-reduction of Cr (VI) by exopolysaccharides (EPS) from indigenous bacterialspecies of Sukinda chromite mine, India. Biodegradation 23 (4), 487–496.
Holden, P.A., Nisbet, R.M., Lenihan, H.S., Miller, R.J., Cherr, G.N., Schimel, J.P., Gardea-Torresdey, J.L., 2012. Ecological nanotoxicology: integrating nanomaterialhazard considerations across the subcellular, population, community, andecosystems levels. Acc. Chem. Res. 46 (3), 813–822.
Hossain, M.S., Mohamad, A.U., Alam, M.S., Hossain, M.A., Rahman, M.S., Haque, M.E.,2007. Antioxidant effect of Spirulina in long evans rat. J. Pharm. Sci. 6 (1),43–45.
Jakopec, S., Dubravcic, K., Polanc, S., Kosmrlj, J., Osmak, M., 2006. Diazene JK-279induces apoptosis-like cell death in human cervical carcinoma cells. Toxicol. InVitro 20 (2), 217–226.
Jiang, G., Shen, Z., Niu, J., Bao, Y., Chen, J., He, T., 2011. Toxicological assessment ofTiO2 nanoparticles by recombinant Escherichia coli bacteria. J. Environ. Monit.13 (1), 42–48.
Jiang, W., Mashayekhi, H., Xing, B., 2009. Bacterial toxicity comparison betweennano- and micro-scaled oxide particles. Environ. Pollut. 157 (5), 1619–1625.
Jin, C., Tang, Y., Yang, F.G., Li, X.L., Xu, S., Fan, X.Y., Yang, Y.J., 2011. Cellular toxicity ofTiO2 nanoparticles in anatase and rutile crystal phase. Biol. Trace Elem. Res. 141(1–3), 3–15.
Jin, X., Li, M., Wang, J., Marambio-Jones, C., Peng, F., Huang, X., Hoek, E.M., 2010.High-throughput screening of silver nanoparticle stability and bacterial in-activation in aquatic media: influence of specific ions. Environ. Sci. Technol. 44(19), 7321–7328.
Kamnev, A.A., 2008. FTIR spectroscopic studies of bacterial cellular responses toenvironmental factors, plant-bacterial interactions and signalling. Spectro-scopy: An International Journal, 22 (2), 83–95.
Klaine, S.J., Alvarez, P.J., Batley, G.E., Fernandes, T.F., Handy, R.D., Lyon, D.Y., Lead, J.R.,2008. Nanomaterials in the environment: behaviour, fate, bioavailability, andeffects. Environ. Toxicol. Chem. 27 (9), 1825–1851.
Kumar, D., Kumari, J., Pakrashi, S., Dalai, S., Raichur, A.M., Sastry, T.P., Mukherjee, A.,2014. Qualitative toxicity assessment of AgNPs on the fresh water bacterialisolates and consortium at low level of exposure concentration. Ecotoxicol.Environ. Saf. 108, 152–160.
Leung, T.Y., Chan, C.Y., Hu, C., Yu, J.C., Wong, P.K., 2008. Photocatalytic disinfection ofmarine bacteria using fluorescent light. Water Res. 42, 4827–4837.
Li, Y., MA, M., Wang, X., Wang, X., 2008. Inactivated properties of activated carbon-supported TiO2 nanoparticles for bacteria and kinetic study. J. Environ. Sci. 20(12), 1527–1533.
Lipovsky, A., Ievitski, L., Tzitrinovich, Z., Gedanken, A., Lubart, R., 2012. Thedifferential behaviour of rutile and anatase nanoparticles in forming oxyradicals upon illumination with visible light: An EPR study. Photochem.Photobiol. 88, 14–20.
Lippincott, E.R., 1963. Infrared spectra of inorganic and coordination compounds. J.Am. Chem. Soc. 85 (21)3532.
Liu, X., Chen, G., Keller, A.A., Su, C., 2013. Effects of dominant material properties onthe stability and transport of TiO2 nanoparticles and carbon nanotubes inaquatic environments: from synthesis to fate. Environ. Sci. Process. Impact 15(1), 169–189.
Liu, Z., Yang, S., Bai, Y., Xiu, J., Yan, H., Huang, J., Wang, L., Zhang, H., Liu., Y., 2011.Miner. Eng. 24, 839–944.
Marshal, D., 2010. Using nano-arrayed structures in sol–gel derived Mn2þ dopedTiO2 for high sensitivity urea biosensor. J. Biosens. Bioelectron. 2012, 1–10.
Morones, J.R., Elechiguerra, J.L., Camacho, A., Holt, K., Kouri, J.B., Ramírez, J.T.,Yacaman, M.J., 2005. The bactericidal effect of silver nanoparticles. Nanotech-nology 16 (10), 2346.
Mueller, N.C., Nowack, B., 2008. Exposure modeling of engineered nanoparticles inthe environment. Environ. Sci. Technol. 42 (12), 4447–4453.
Navarro, E., Baun, A., Behra, R., Hartmann, N.B., Filser, J., Miao, A.J., Sigg, L., 2008.Environmental behavior and ecotoxicity of engineered nanoparticles to algae,plants, and fungi. Ecotoxicology 17 (5), 372–386.
Nel, A.E., Nasser, E., Godwin, H., Avery, D., Bahadori, T., Bergeson, L., Wong, J., 2013.A multi-stakeholder perspective on the use of alternative test strategies fornanomaterial safety assessment. ACS Nano 7 (8), 6422–6433.
Newman, M.D., Stotland, M., Ellis, J.I., 2009. The safety of nanosized particles intitanium dioxide- and zinc oxide-based sunscreens. J. Am. Acad. Dermatol. 61(4), 685–692.
O’Brien, N., Cummins, E., 2010. Ranking initial environmental and human healthrisk resulting from environmentally relevant nanomaterials. J. Environ. Sci.Health A 45 (8), 992–1007.
Pakrashi, S., Dalai, S., Sabat, D., Singh, S., Chandrasekaran, N., Mukherjee, A., 2011.Cytotoxicity of Al2O3 nanoparticles at low exposure levels to a freshwaterbacterial isolate. Chem. Res. Toxicol. 24 (11), 1899–1904.
Pakrashi, S., Dalai, S., Humayun, A., Chakravarty, S., Chandrasekaran, N., Mukherjee,A., 2013. Ceriodaphniadubia as a potential bio-indicator for assessing acutealuminum oxide nanoparticle toxicity in fresh water environment. PloS One 8(9), e74003.
Patriquin, G.M., Banin, E., Gilmour, C., Tuchman, R., Greenberg, E.P., Poole, K., 2008.Influence of quorum sensing and iron on twitching motility and biofilmformation in Pseudomonas aeruginosa. J. Bacteriol. 190 (2), 662–671.
Paul, M.L., Samuel, J., Chandrasekaran, N., Mukherjee, A., 2012. Comparativekinetics, equilibrium, thermodynamic and mechanistic studies on biosorptionof hexavalent chromium by live and heat killed biomass of AcinetobacterjuniiVITSUKMW2, an indigenous chromite mine isolate. Chem. Eng. J. 187, 104–113.
Pigeot-Rémy, S., Simonet, F., Errazuriz-Cerda, E., Lazzaroni, J.C., Atlan, D., Guillard,C., 2011. Photocatalysis and disinfection of water: identification of potentialbacterial targets. Appl. Catal. B 104 (3), 390–398.
Robertson, P.K.J., Robertson, J.M.C., Bahnemann, D.W., 2012. Removal of micro-organisms and their chemical metabolites from water using semiconductorphotocatalysis. J. Hazard. Mater. 211, 161–171.
Sadiq, I.M., Chandrasekaran, N., Mukherjee, A., 2010. Studies on effect of TiO2
nanoparticles on growth and membrane permeability of Escherichia coli,Pseudomonas aeruginosa, and Bacillus subtilis. Curr. Nanosci. 6 (4), 381–387.
Sadiq, I.M., Dalai, S., Chandrasekaran, N., Mukherjee, A., 2011. Ecotoxicity study oftitania (TiO2) NPs on two microalgae species: Scenedesmus sp. and Chlorella sp.Ecotoxicol. Environ. Saf. 74 (5), 1180–1187.
Samuel, J., Paul, M.L., Pulimi, M., Nirmala, M.J., Chandrasekaran, N., Mukherjee, A.,2012. Hexavalent chromium bioremoval through adaptation and consortiadevelopment from Sukinda chromite mine isolates. Ind. Eng. Chem. Res. 51(9), 3740–3749.
Sayes, C.M., Wahi, R., Kurian, P.A., Liu, Y., West, J.L., Ausman, K.D., Colvin, V.L., 2006.Correlating nanoscaletitania structure with toxicity: a cytotoxicity and inflam-matory response study with human dermal fibroblasts and human lungepithelial cells. Toxicol. Sci. 92 (1), 174–185.
Tsuang, Y.H., Sun, J.S., Huang, Y.C., Lu, C.H., Chang, W.H.S., Wang, C.C., 2008. Studiesof photokilling of bacteria using titanium dioxide nanoparticles. Artif. Organs32 (2), 167–174.
Vlamakis, H., Chai, Y., Beauregard, P., Losick, R., Kolter, R., 2013. Sticking together:building a biofilm the Bacillus subtilis way. Nat. Rev. Microbiol. 11 (3), 157–168.
Wang, H., Joseph, J.A., 1999. Quantifying cellular oxidative stress by dichlorofluor-escein assay using microplate reader. Free Radic. Biol. Med. 27 (5), 612–616.
Wang, X., Xia, T., Duch, M.C., Ji, Z., Zhang, H., Li, R., Nel, A.E., 2012. Pluronic F108coating decreases the lung fibrosis potential of multiwall carbon nanotubes byreducing lysosomal injury. Nano Lett. 12 (6), 3050–3061.
Wintherbourn, C., Hawkins, R., Brian, M., Carrell., R., 1975. J. Lab. Clin. Med. 85,337–341.