cyst–theca relationship of the arctic dinoflagellate cyst islandinium minutum (dinophyceae) and...
TRANSCRIPT

CYST–THECA RELATIONSHIP OF THE ARCTIC DINOFLAGELLATE CYSTISLANDINIUM MINUTUM (DINOPHYCEAE) AND PHYLOGENETIC POSITION BASED ON
SSU rDNA AND LSU rDNA1
�Eric Potvin2
School of Earth and Environmental Sciences, College of Natural Sciences, Seoul National University, Seoul 151-747, Korea
Andr�e Rochon
Institut des sciences de la mer de Rimouski, Universit�e du Qu�ebec �a Rimouski, Rimouski, QC, Canada G5L 3A1
and Connie Lovejoy
D�epartement de biologie, Universit�e Laval, Qu�ebec, QC, Canada G1V 0A6
Round brown spiny cysts constitute a morphologicalgroup common in high latitude dinoflagellate cystassemblages. The dinoflagellate cyst Islandiniumminutum (Harland et Reid) Head, Harlandet Matthiessen is the main paleoecological indicator ofseasonal sea-ice cover in the Arctic. Despite theimportance of this cyst in paleoceanographicalstudies, its biological affinity has so far beenunknown. The biological affinity of the speciesI. minutum and its phylogenetic position based on thesmall subunit ribosomal RNA gene (SSU rDNA) andthe large subunit ribosomal RNA gene (LSU rDNA)were established from cyst incubation experiments incontrolled conditions, optical and scanning electronmicroscopy, and single-cell PCR. The thecal motilecell obtained was undescribed. Although the motilecell was similar to Archaeperidinium minutum (Kofoid)J€orgensen, the motile cell of I. minutum lacked atransitional plate in the cingular series, which ispresent in Archaeperidinium spp. Islandinium minutumand Archaeperidinium spp. were paraphyletic in allphylogenetic analyses. Furthermore, Protoperidiniumtricingulatum, which also lacks a transitional plate, wasclosely related to I. minutum and transfered to thegenus Islandinium. Based on available data, it is clearthat Islandinium is distinct from Archae-peridinium. Therefore, we considered IslandiniumHead, Harland et Matthiessen as a non-fossil genusand emend its description, as well as the speciesI. minutum. This is the first description of a cyst–theca relationship and the first study that reportsmolecular data based on SSU rDNA and LSU rDNAon a species assigned to the genus Islandinium.
Key index words: Amphidiniopsis dragescoi; Archaepe-ridinium; Baffin Bay; Echinidinium; Herdmanialitoralis; Islandinium minutum; Protoperidiniaceae;
Protoperidinium minutum; round brown spiny cyst;single-cell PCR
Organic-walled dinoflagellate cysts, or dinocysts,are useful proxies for the reconstruction of past sea-surface conditions. Dinoflagellate populations mayproliferate under diverse conditions and producedinocysts that accumulate and are recovered fromsediments. In modern sediments of the NorthernHemisphere, relative abundance of round brownspiny cysts can account for 15%–40% of the totaldinocyst assemblages, notably in high-productivityareas such as polynyas, coastal upwelling zones, andestuaries (Hamel et al. 2002, Marret and Zonneveld2003, Pospelova et al. 2004, 2005, 2008, 2010,Radi et al. 2007, V�asquez-Bedoya et al. 2008,Bouimetarhan et al. 2009, Mertens et al. 2009,Krepakevich and Pospelova 2010, Limoges et al.2010, Pospelova and Kim 2010, Price and Pospelova2011). In cold environments, round brown spinycysts are often dominant in surface samples as wellas in sediment core sequences (Rochon et al. 1999,Head et al. 2001, Mudie and Rochon 2001, de Ver-nal et al. 2001, Hamel et al. 2002, Radi and de Ver-nal 2004, Matthiessen et al. 2005, Richerol et al.2008, Bonnet et al. 2010, 2012).Round brown spiny cysts are classified into two cyst
genera; Islandinium (Head et al. 2001) and Echinidi-nium (Zonneveld 1997). These two genera are distin-guished by their archeopyle. Islandinium has asaphopylic archeopyle reflecting three apical plateswhile Echinidinium has a theropylic archeopyle pro-duced by the separation along predetermined par-asutures on the epicyst (Head et al. 2001, Radi et al.2013).Several studies have recorded dinocyst taxa
throughout the Arctic and subarctic regions(Harland et al. 1980, Rochon et al. 1999, Solomonet al. 2000, Mudie and Rochon 2001, Hamel et al.2002, Richerol et al. 2008). The relationships
1Received 1 October 2012. Accepted 6 May 2013.2Author for correspondence: e-mail [email protected] Responsibility: D. Vaulot (Associate Editor)
J. Phycol. 49, 848–866 (2013)© 2013 Phycological Society of AmericaDOI: 10.1111/jpy.12089
848

between the cyst and its motile counterpart in theseregions have been confirmed for few species (Head1996), representing less than one third of therecorded cyst species (Rochon 2009). The biologicalaffinities of round brown spiny cysts occurring inthese regions are currently unknown. The determina-tion of cyst–theca relationships, by the recognition ofthe motile form producing the cyst, can provideinvaluable information on the distribution of speciesin relation to environmental factors. Furthermore, itsestablishment increases our knowledge of the taxon-omy and our understanding of the fossil record.
Incubation experiments investigating this mor-phological group have always produced motileforms associated to the genus Protoperidinium (e.g.,Lewis et al. 1984, Kawami et al. 2009), the subgenusArchaeperidinium (Wall and Dale 1968, Fukuyo et al.1977, Zonneveld and Dale 1994, Ribeiro et al.2010), which has been recently reinstated as genusby Yamaguchi et al. (2011), and the subfamily Dip-lopsalioideae (e.g., Kawami et al. 2006), all withinthe family Protoperidiniaceae.
The cyst species Islandinium minutum (Harlandet Reid) Head, Harland et Matthiessen was initiallydescribed as Multispinula minuta Harland et Reid(Harland et al. 1980) from modern sediment of theBeaufort Sea, Canadian Arctic. Matsuoka and Bujak(1988) thereafter assigned the species to the genusAlgidasphaeridium, implying an affinity with the orderGymnodiniales. In 2001, Head et al. (2001) againreclassified the species within the new cyst genusIslandinium as part of the family Protoperidiniaceae,which is considered to consist exclusively of non pho-tosynthetic marine species (Fensome et al. 1993).
The detailed taxonomic assignment of the dino-flagellate species I. minutum (Head et al. 2001) isimportant because of its relevance as a paleoecologi-cal indicator for the reconstruction of the durationof seasonal sea-ice cover (Rochon et al. 1999, Kunz-Pirrung 2001) and its distribution throughout theArctic and subarctic regions where it can accountfor up to 90% of the surface sediment assemblagesof northern Baffin Bay, the Canadian Arctic Archi-pelago, Fram Strait and the Laptev and Kara Seas(Head et al. 2001). The present work aims were firstto establish the relationship between the dinoflagel-late cyst I. minutum and its motile cell with incuba-tion experiments and detailed observations withoptical and scanning electron microscopy and sec-ond to determine the phylogenetic position of thespecies based on the small subunit ribosomal RNAgene (referred to as SSU rDNA) and the large sub-unit ribosomal RNA gene (referred to as LSUrDNA). This is the first study of this type on a cystassigned to the genus Islandinium.
MATERIALS AND METHODS
Germination. Isolation: Surface sediment samples from theCanadian Arctic were collected on September 18, 2006 at a
depth of 311 m in Baffin Bay (76°25′N, 77°18′W) on boardthe CCGS Amundsen when the surface temperature was 0°Cand salinity 31. The surface sediment was collected from aboxcore and stored in the dark at 4°C for further analyses.To concentrate potentially viable dinoflagellate cysts, between1 and 2 cc of sediment were sonicated for 5 min in filteredseawater and sieved through 100-lm and 20-lm Nytexmeshes. The >20 lm fraction was then transferred in a 50-mLbeaker with filtered seawater. A manual vortex was appliedand the suspended fraction was recovered. The cysts weresonicated once more for 5 min and rinsed on the 20-lm Ny-tex mesh with filtered seawater. The last step was repeatedtwice to remove particles from the cysts. Individual cysts ofI. minutum were identified at 4009 with an inverted transmit-ted light microscope Eclipse TE2000-U (Nikon, Tokyo, Japan)and transferred into 24-well plates (Corning Inc., Corning,New York, USA) using glass micropipettes. Each well con-tained ~2.0 mL of F/2-Si culture medium (Guillard and Ry-ther 1962) at a salinity of 32.
Incubation: Cysts were incubated up to 46 d in a plantgrowth chamber (model MLR-351H, Sanyo Electric Co Ltd.,Moriguchi, Japan) at 4°C under a 18:6 h light:dark cycle at alight intensity of 120 lmol photons � m�2 � s�1.
Morphology. Transmitted light microscopy: Specimens wereobserved with a transmitted light microscope Eclipse 80i(Nikon) equipped with differential interferential contrast.The cells were transferred individually into a drop of filteredseawater framed by vinyl tape on a glass microscope slide.The cells were sealed by applying silicon grease on the vinylframe and covering the latter with a coverslip (modified fromHoriguchi et al. 2000). The morphology of plates was deter-mined following the addition of commercial sodium hypo-chlorite (≤5.25%), which caused the plates to separate fromeach other. The morphology of the cysts was determined bymounting specimens in glycerin gel between slide and cover-slip. The specimens were repositioned by gently heating thegel. The coverslip was then moved until the specimens werein the desired position.
The motile cells and cysts were measured with an ocularruler in the eyepiece of a transmitted light or inverted lightmicroscope and images were acquired with a digital camera.
Scanning electron microscopy: For scanning electron micros-copy, cells were fixed with 2% formaldehyde and rinsed indistilled water. Cells were then put through a dehydrationseries in acetone (10, 20, 30, …, 100%). The cells were driedusing a critical point dryer (Polaron model E3000, QuorumTechnologies, Ashford, UK). Finally, the cells were mountedon stubs, sputter coated (Quorum Technologies SC7680 sput-ter coater; NewHaven, UK) with gold-palladium, andobserved with a scanning electron microscope (JEOL JSM-6460LV; Peabody, Massachusetts, USA).
The tabulation system developed by Kofoid (1909) was usedfor the description of the thecal organization. The sulcal platenomenclature followed Williams et al. (2000) for the genusProtoperidinium. The terminology used to describe plate modifi-cations from the usual plate pattern followed Lef�evre (1932).
Genetics. Polymerase chain reaction: To highlight the thecalplate arrangement and confirm the identity of the dinoflagel-late, several live motile cells were stained for 10 min with 1mg mL-1 calcofluor white (Fluorescent Brightener 28, Sigma-Aldrich, Oakville, Ontario, Canada; Fritz and Triemer 1985).The motile cells were then observed under epifluorescence(excitation: 360 nm, emission: 460 nm) with a microscopeOlympus BX51 (Olympus, Tokyo, Japan) to determine thearrangement of thecal plates. The live cells were then washedin three drops of Nanopure water (NANOpure Infinity UV,model D8971; Barnstead International, Dubuque, IA, USA)and broken with a glass micropipette. Drops containing
MORPHOLOGY AND PHYLOGENETIC POSITION OF ISLANDINIUM MINUTUM 849

cellular contents were then placed in microtubes and ampli-fied directly.
Amplicons of the SSU rDNA and LSU rDNA were obtainedfollowing nested PCR protocols. The first PCR round finalmix concentrations were as follows: 19 of PCR buffer(Qiagen, Germantown, MD, USA), 0.2 mM of dNTP (dNTP-Mix; Qiagen), 0.5 lM of each primer, 0.1 U � lL�1 of Hot-StarTaq DNA polymerase (Qiagen), and 1.6 mM of MgCl2.SR1 and LSUR2 primers were used in the first round of PCRto amplify the genes coding for the SSU rRNA and LSUrRNA in a final volume of 25 lL. PCR was conducted usingEppendorf thermocyclor model 5341 (Brinkmann, Haup-pauge, New York) as follows: one activation step of 15 min at95°C, followed of 35 cycles at 94°C for 1 min, 53°C for1 min, and 72°C for 3 min, and a final elongation step at72°C for 10 min. A volume of 0.5 lL of a 1:1000 dilution ofthe first PCR step was used as template for the second PCRround using the same mix. Individual reactions in a final vol-ume of 35 lL were produced with these primer pairs: SR1(or SR1b) and SR5AY, SR4 and SR9p, SR8im and SR12b,25F1 and 25R1, and LSU D3A and LSU R2 (Table 1). Thesecond round of PCR was as follows: one activation step at95°C for 15 min, followed by 40 cycles at 94°C for 30 s, 53°Cfor 30 s, and 72°C for 30 s, and a final elongation step at72°C for 10 min.
Verification of PCR products and sequencing: Positive and neg-ative controls were used for all amplification reactions. Thesize of the amplicons was verified on a 1.0% agarose gel.Products were visualized under a UV lamp. PCR productswere sent to Centre de recherche du Centre hospitalier del’Universit�e Laval (CHUL, Qu�ebec, QC, Canada) where theywere purified with UniFilter 384-well microplates of Whatman(VWR, Mississauga, ON, Canada) and sequenced on aABI-3730xl system (Applied Biosystems, Foster City, CA, USA)with the primers used in the second round of PCR.
Phylogenetic analyses: The sequence fragments were assem-bled by manual alignment using BioEdit v7.0.9.0 (Hall 1999).Sequences of taxa used to construct the phylogenies wereobtained from NCBI GenBank (Table 2). The new and refer-ence sequences were aligned using CLUSTAL X v2.0 (Larkinet al. 2007). The alignment was inspected and refined manu-ally. Several Protoperidiniaceae sequences from GenBankcould not be aligned across the entire gene and these non-alignable regions were removed. The aligned matrix was thenanalyzed with PAUP v4.0b10 (Swofford 2002). Maximum like-lihood (ML) was used as phylogenetic method. Models ofnucleotide substitution were determined with Modeltest v7.3(Posada and Crandall 1998) based on the Akaike informationcriterion (Posada and Buckley 2004). A heuristic tree searchwas used to determine the optimal trees. The tree bisection-
reconnection algorithm was used with five random additionsof sequences. The characters were equally weighted andspaces in the alignment were treated as missing data. RAxMLv7.0.4 (Stamatakis 2006) was used to calculate bootstrap val-ues using the default algorithm with the general time revers-ible + Γ model of nucleotide substitution and 1,000replicates.
The matrix was also analyzed with MrBayes v3.1.2 (Huel-senbeck and Ronquist 2001) for Bayesian analyses. The mod-els previously selected by Modeltest 7.3 were used. Fourindependent Markov chain Monte Carlo simulationswere run simultaneously for 2,000,000 generations. Treeswere sampled every 1,000 generations and the first 800trees were deleted to ensure that the likelihood had reachedconvergence. A majority-rule consensus tree was created fromthe remaining 1,201 trees to examine the posterior probabili-ties of each clade.
RESULTS
Cyst. Generalities: The morphology of the cystsrecovered from sediments corresponded to the diag-nosis given for I. minutum by Head et al. (2001).The cysts were spherical, brown, spiny, proximate toproximochorate with a dark brown coloration whencell content was present (Fig. 1a) and slightly brownwhen empty (Fig. 1, b–f). The live cyst cell contentappeared slightly brown (Fig. 1a). Green and yellowglobular bodies were also present (Fig. 1a). The cystdiameter varied between 27 and 50 lm (mean39 lm, n = 102). The length of processes variedbetween 2 and 7 lm (mean 4 lm, n = 102, onemeasurement per cyst). The surface was orna-mented with granules (Fig. 1, b and c) and ran-domly distributed rings (Fig. 2, a–c). The straight orcurved processes (Fig. 1, d–f) were not tabular anddistributed evenly (Fig. 1b) or unevenly (Fig. 1c) onthe surface. Incomplete processes were observed ona minority of cysts (Fig. 2d). The wall was composedof two accolate membranes and never exceeded0.5 lm in thickness (Fig. 1f).Processes: Processes were acicular (Figs. 1 and 2).
A pericoel was present at the base of each processand occupied up to one third of the process length(Fig. 1, d–f). The remainder of each process wassolid (Fig. 1f). They were conical at the base and
TABLE 1. Primers used for the amplification and sequencing of SSU rDNA and LSU rDNA
Code SD Sequence (5′-3′) Position References
SR1 F TACCTGGTTGATCCTGCCAG 1–20 (a) Yamaguchi and Horiguchi (2005)SR1b F GATCCTGCCAGTAGTCATATGCTT 10–33 (a) Yamaguchi and Horiguchi (2005)SR4 F AGGGCAAGTCTGGTGCCAG 548–566 (a) Yamaguchi and Horiguchi (2005)SR5AY R ACTACGAGCTTTTTAACCGC 630–611 (a) Yamaguchi and Horiguchi (2005)SR8im F GGATTGACAGATTGATAGCT 1224–1243 (a) This studySR9p R AACTAAGAACRGCCATGCAC 1286–1267 (a) Yamaguchi and Horiguchi (2005)SR12b R CGGAAACCTTGTTACGACTTCTCC 1762–1739 (a) Yamaguchi and Horiguchi (2005)25F1 F CCGCTGAATTTAAGCATAT 27–45 (b) Kogame et al. (1999)25R1 R CTTGGTCCGTGTTTCAAGAC 732–713 (b) Yamaguchi and Horiguchi (2005)LSU D3A F GACCCGTCTTGAAACACGGA 708–727 (b) Takano and Horiguchi (2006)LSU R2 R ATTCGGCAGGTGAGTTGTTAC 1342–1322 (b) Takano and Horiguchi (2006)
The position of the primers is indicated based on the DNA sequence of (a) Volvox carteri (Rausch et al. 1989) coding for SSUrRNA and of (b) Prorocentrum micans (Lenaers et al. 1989) coding for LSU rRNA. SD: synthesis direction, F: forward, R: reverse.
850 �ERIC POTVIN ET AL.
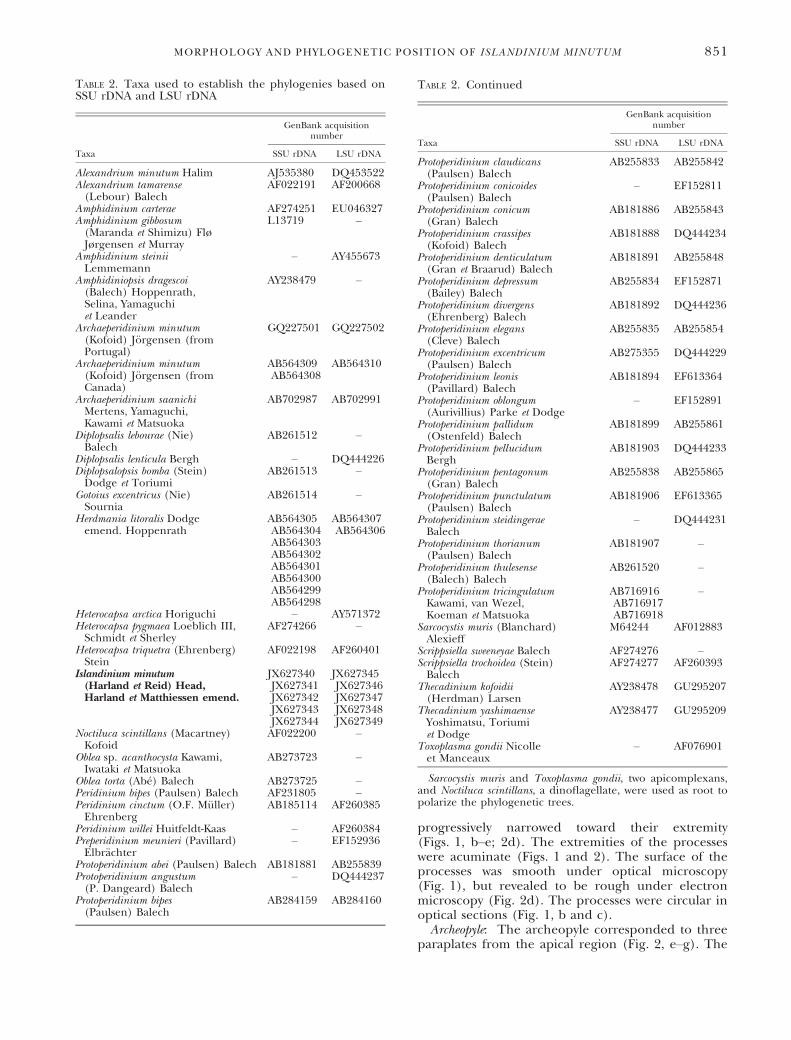
progressively narrowed toward their extremity(Figs. 1, b–e; 2d). The extremities of the processeswere acuminate (Figs. 1 and 2). The surface of theprocesses was smooth under optical microscopy(Fig. 1), but revealed to be rough under electronmicroscopy (Fig. 2d). The processes were circular inoptical sections (Fig. 1, b and c).Archeopyle: The archeopyle corresponded to three
paraplates from the apical region (Fig. 2, e–g). The
TABLE 2. Taxa used to establish the phylogenies based onSSU rDNA and LSU rDNA
Taxa
GenBank acquisitionnumber
SSU rDNA LSU rDNA
Alexandrium minutum Halim AJ535380 DQ453522Alexandrium tamarense(Lebour) Balech
AF022191 AF200668
Amphidinium carterae AF274251 EU046327Amphidinium gibbosum(Maranda et Shimizu) FløJørgensen et Murray
L13719 –
Amphidinium steiniiLemmemann
– AY455673
Amphidiniopsis dragescoi(Balech) Hoppenrath,Selina, Yamaguchiet Leander
AY238479 –
Archaeperidinium minutum(Kofoid) J€orgensen (fromPortugal)
GQ227501 GQ227502
Archaeperidinium minutum(Kofoid) J€orgensen (fromCanada)
AB564309AB564308
AB564310
Archaeperidinium saanichiMertens, Yamaguchi,Kawami et Matsuoka
AB702987 AB702991
Diplopsalis lebourae (Nie)Balech
AB261512 –
Diplopsalis lenticula Bergh – DQ444226Diplopsalopsis bomba (Stein)Dodge et Toriumi
AB261513 –
Gotoius excentricus (Nie)Sournia
AB261514 –
Herdmania litoralis Dodgeemend. Hoppenrath
AB564305AB564304AB564303AB564302AB564301AB564300AB564299AB564298
AB564307AB564306
Heterocapsa arctica Horiguchi – AY571372Heterocapsa pygmaea Loeblich III,Schmidt et Sherley
AF274266 –
Heterocapsa triquetra (Ehrenberg)Stein
AF022198 AF260401
Islandinium minutum(Harland et Reid) Head,Harland et Matthiessen emend.
JX627340JX627341JX627342JX627343JX627344
JX627345JX627346JX627347JX627348JX627349
Noctiluca scintillans (Macartney)Kofoid
AF022200 –
Oblea sp. acanthocysta Kawami,Iwataki et Matsuoka
AB273723 –
Oblea torta (Ab�e) Balech AB273725 –Peridinium bipes (Paulsen) Balech AF231805 –Peridinium cinctum (O.F. M€uller)Ehrenberg
AB185114 AF260385
Peridinium willei Huitfeldt-Kaas – AF260384Preperidinium meunieri (Pavillard)Elbr€achter
– EF152936
Protoperidinium abei (Paulsen) Balech AB181881 AB255839Protoperidinium angustum(P. Dangeard) Balech
– DQ444237
Protoperidinium bipes(Paulsen) Balech
AB284159 AB284160
TABLE 2. Continued
Taxa
GenBank acquisitionnumber
SSU rDNA LSU rDNA
Protoperidinium claudicans(Paulsen) Balech
AB255833 AB255842
Protoperidinium conicoides(Paulsen) Balech
– EF152811
Protoperidinium conicum(Gran) Balech
AB181886 AB255843
Protoperidinium crassipes(Kofoid) Balech
AB181888 DQ444234
Protoperidinium denticulatum(Gran et Braarud) Balech
AB181891 AB255848
Protoperidinium depressum(Bailey) Balech
AB255834 EF152871
Protoperidinium divergens(Ehrenberg) Balech
AB181892 DQ444236
Protoperidinium elegans(Cleve) Balech
AB255835 AB255854
Protoperidinium excentricum(Paulsen) Balech
AB275355 DQ444229
Protoperidinium leonis(Pavillard) Balech
AB181894 EF613364
Protoperidinium oblongum(Aurivillius) Parke et Dodge
– EF152891
Protoperidinium pallidum(Ostenfeld) Balech
AB181899 AB255861
Protoperidinium pellucidumBergh
AB181903 DQ444233
Protoperidinium pentagonum(Gran) Balech
AB255838 AB255865
Protoperidinium punctulatum(Paulsen) Balech
AB181906 EF613365
Protoperidinium steidingeraeBalech
– DQ444231
Protoperidinium thorianum(Paulsen) Balech
AB181907 –
Protoperidinium thulesense(Balech) Balech
AB261520 –
Protoperidinium tricingulatumKawami, van Wezel,Koeman et Matsuoka
AB716916AB716917AB716918
–
Sarcocystis muris (Blanchard)Alexieff
M64244 AF012883
Scrippsiella sweeneyae Balech AF274276 –Scrippsiella trochoidea (Stein)Balech
AF274277 AF260393
Thecadinium kofoidii(Herdman) Larsen
AY238478 GU295207
Thecadinium yashimaenseYoshimatsu, Toriumiet Dodge
AY238477 GU295209
Toxoplasma gondii Nicolleet Manceaux
– AF076901
Sarcocystis muris and Toxoplasma gondii, two apicomplexans,and Noctiluca scintillans, a dinoflagellate, were used as root topolarize the phylogenetic trees.
MORPHOLOGY AND PHYLOGENETIC POSITION OF ISLANDINIUM MINUTUM 851

adnate operculum remained on the cyst after germi-nation. The paraplates composing the operculumwere not always free from each other, whichcan cause confusion. Morphological variations werenoted for the outline of the archeopyle (Fig. 2, e–g).The reflected inverted V shape of the upper partof the first apical plate on the cyst was a reliablecriterion to orient the cyst (Fig. 2, e–g). Thearcheopyle was the only indication of tabulation onthe cysts.Germination. A total of 142 cysts of I. minutum
were incubated at the same time following29 months of storage. The cysts were observed dailyfor up to 46 d. A total of 120 cysts, 85% of thosetested, germinated within 36 d. The majority ofexcystments (65%) occurred between 2 and 9 dafter being placed in the well plates. The motilecells survived without any prey for up to 12 d aftergermination. A single cell was able to form tempo-
rary cysts up to four times. In some cases, one ortwo divisions were observed, but no culture wasobtained.Morphology of the motile cell. Generalities: The
motile cells were thecal. The tabulation pattern(Figs. 3, a–e; 4, a–d) and the tabulation formula Po,X, 4′, 2a, 6-7″, 3C, 5S, 5‴, 2′′′′ were similar to thenewly reinstated genus Archaeperidinium (Yamaguchiet al. 2011). The cells were subspherical (Fig. 5, aand b) and had green and yellow globulous bodiesin the cytoplasm (Fig. 5b). The cells were 33–58 lmlong (mean 44 lm, n = 102) and 30–50 lm wide(mean 41 lm, n = 102). The epitheca and hypot-heca were similar in size (Fig. 5, a and b). Theepitheca had a dome shape and the hypothecawas round in ventral view (Fig. 5, a and b). Thetheca had a slight asymmetry in ventral opticalsection (Fig. 5b). The cell had no projection at the
a
e
dc
f
b
FIG. 1. Micrographs of the dinoflagellate cyst Islandinium min-utum taken using a transmitted light microscope showing generalfeatures. (a) Optical cut of a living cyst illustrating the cellularcontent. (b–c) Tangential view illustrating details from the cystsurface and the distribution of processes. (d–e) Tangential viewillustrating details of processes. (f) Optical section of an emptycyst illustrating details from wall and processes; scalebars = 10 lm.
a
ed
c
f
b
g
FIG. 2. Micrographs of the dinoflagellate cyst Islandinium minu-tum taken using a transmitted light and scanning electron micro-scopes. (a–c) General view of the cyst showing the granulatedsurface with randomly distributed ring. (d) Details of the processesshowing conical bases, the progressive reduction in width towardtheir extremity, the acuminate tips, the granules ornamenting thesurface, and incomplete processes. (e–g) Details of the archeopyleand operculum. Scale bars = 10 lm (a–c and e–f), 2 lm (d),5 lm (g).
852 �ERIC POTVIN ET AL.

apex and antapex (Fig. 5, a and b). The generalmorphology of the cell was consistent among theobserved cells. The cingulum was equatorial, notdisplaced, excavated, and with a reduced list(Fig. 5, a–c, m). The sulcus was not excavated andwas ornamented with a list located on the left side ofthe right sulcal plate (rs; Fig. 5c). The thecalplates were ornamented with faint reticulation and,except those of the apical pore complex (APC),were ornamented with pores surrounded by a circularrim (Fig. 5d). Spines also ornamented the surface ofthe theca (Fig. 5d). The pores were disposedrandomly on the plates with the exception of tworows of pores located on the hypotheca below thecingulum and in the middle of the cingulum(Fig. 5a).
Apical pore complex: The APC was oriented to theright of the theca making it slightly asymmetrical inrelation to the dorsoventral axis (Fig. 5e). The com-plex included a quadrangular pore plate (Po) and asmall square shape X plate (x; Fig. 5e). The porewas located at the center of the pore plate Po(Fig. 5e).Epitheca: The first apical plate (1′) had the aspect
of a truncated lozenge in its inferior and superiorparts and demonstrated an ortho arrangement con-tacting four major epithecal plates. The right infe-rior side of the 1′ plate was curved and longer thanthe left inferior side. This particularity made the 1′plate asymmetrical (Fig. 5f). Minor contacts linkedthe 1′ plate to the anterior sulcal plate (as; Fig. 5c)and the small X plate of the APC (Fig. 5e). Bothanterior intercalary plates (a) were of similar sizeand displaced to the left of the epitheca in lateral(1a) and dorsal (2a) positions (Fig. 5f). Variationsin plate pattern were observed on the left side ofthe epitheca implicating the contact between the 1″and 1a plates (Figs. 4; 5, g–j). These modificationsas defined by Lef�evre (1932) are qualified as fol-lows: conjunctum, when the previous plates fully con-tact (Fig. 5g); contactum, when plates contact eachother at one point (Fig. 5h); and remotum (Fig. 5i),when plates are not in contact. Furthermore, thefusion of the 1″ and 2″ plates was also observed andis defined as a simplex modification (Figs. 4; 5j;
a b
c d
e
rs ls
ps
msas
3’’ 4’’ 5’’ 6’’
1a 2a
3’
3’’’ 4’’’2’’’
1’’’’ 2’’’’
2c
1’’’’ 2’’’’2’’’
3’’’
4’’’
1’’’ 5’’’ps
1c
2c
3c
2’’
1’2’4’
1a
1’’7’’
1’’’5’’’
as
rs ls
ps
1c 2c3c
x
x
1c
3’
2a
1a
2’4’
1’1’’
2’’
3’’
4’’5’’
6’’
7’’
2c
3c
P0
FIG. 3. Schematize view of the theca of the motile cellobtained from the cyst of Islandinium minutum. (a) Ventral view.(b) Dorsal view. (c) Antapical view. (d) Apical view. (e) Sulcalview. Abbreviations: Po, pore plate; x, X plate; as, anterior sulcal;rs, right sulcal; ms, median sulcal; ls, left sulcal; ps, posteriorsulcal.
a b
c d
P0
x
1’
2’4’ 1a
1’’2’’
3’’
4’’5’’
6’’
7’’
2a
3’
x
1’
2’4’ 1a
1’’2’’
3’’
4’’5’’
6’’
7’’
2a
3’P0
P0
x
1’
2’4’ 1a
1’’2’’
3’’
4’’5’’
6’’
7’’
2a
3’
x
1’
2’4’
1a
1’’2’’
3’’
4’’5’’
6’’
7’’
2a
3’P0
FIG. 4. Schematize view of the epithecal variability of themotile form obtained from the cyst of Islandinium minutum. (a)Conjunctum tabulation showed by the complete junction betweenthe 1″ and 1a plates. (b) Remotum tabulation showed by theabsence of contact between the 1″ and 1a plates. (c) Contactumtabulation showed by the presence of only one point of contactbetween the 1″ and 1a plates. (d) Simplex modification involvingthe fusion of 1″ and 2″ plates. Abbreviations: Po, pore plate; x, Xplate.
MORPHOLOGY AND PHYLOGENETIC POSITION OF ISLANDINIUM MINUTUM 853
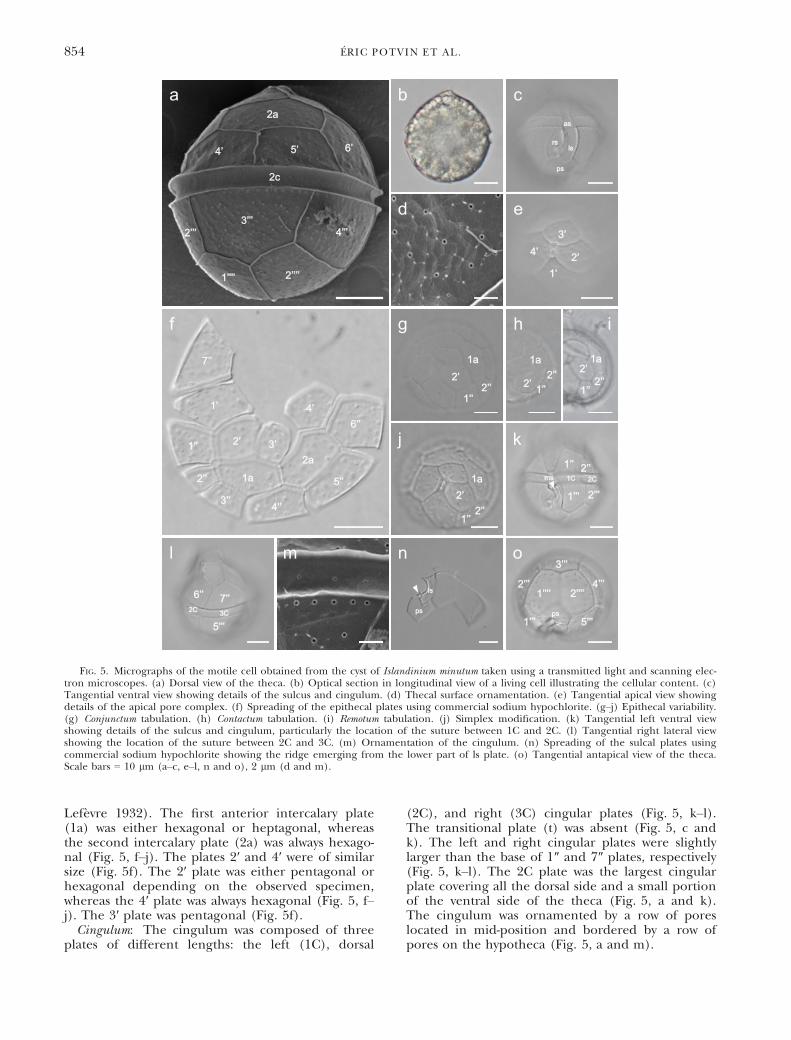
Lef�evre 1932). The first anterior intercalary plate(1a) was either hexagonal or heptagonal, whereasthe second intercalary plate (2a) was always hexago-nal (Fig. 5, f–j). The plates 2′ and 4′ were of similarsize (Fig. 5f). The 2′ plate was either pentagonal orhexagonal depending on the observed specimen,whereas the 4′ plate was always hexagonal (Fig. 5, f–j). The 3′ plate was pentagonal (Fig. 5f).Cingulum: The cingulum was composed of three
plates of different lengths: the left (1C), dorsal
(2C), and right (3C) cingular plates (Fig. 5, k–l).The transitional plate (t) was absent (Fig. 5, c andk). The left and right cingular plates were slightlylarger than the base of 1″ and 7″ plates, respectively(Fig. 5, k–l). The 2C plate was the largest cingularplate covering all the dorsal side and a small portionof the ventral side of the theca (Fig. 5, a and k).The cingulum was ornamented by a row of poreslocated in mid-position and bordered by a row ofpores on the hypotheca (Fig. 5, a and m).
6’’ 7’’
5’’’
2C 3C
1’’ 2’’2C1C
1’’’ 2’’’
ms
1’’2’’
2’1a
1’’2’’
2’
1a
1’’2’’
2’1a
1’’’’ 2’’’’
1’’’
2’’’
3’’’
4’’’
5’’’ps
b cas
rsls
ps
d
ps
ls
e
1’2’
3’4’
n
f
1’
2’ 3’
4’
1a
2a1’’
2’’
3’’ 4’’
5’’
6’’
7’’
g
1’’2’’
1a
2’
h
j
oml
a2a
4’ 5’ 6’
3’’’4’’’2’’’
1’’’’ 2’’’’
2c
i
k
FIG. 5. Micrographs of the motile cell obtained from the cyst of Islandinium minutum taken using a transmitted light and scanning elec-tron microscopes. (a) Dorsal view of the theca. (b) Optical section in longitudinal view of a living cell illustrating the cellular content. (c)Tangential ventral view showing details of the sulcus and cingulum. (d) Thecal surface ornamentation. (e) Tangential apical view showingdetails of the apical pore complex. (f) Spreading of the epithecal plates using commercial sodium hypochlorite. (g–j) Epithecal variability.(g) Conjunctum tabulation. (h) Contactum tabulation. (i) Remotum tabulation. (j) Simplex modification. (k) Tangential left ventral viewshowing details of the sulcus and cingulum, particularly the location of the suture between 1C and 2C. (l) Tangential right lateral viewshowing the location of the suture between 2C and 3C. (m) Ornamentation of the cingulum. (n) Spreading of the sulcal plates usingcommercial sodium hypochlorite showing the ridge emerging from the lower part of ls plate. (o) Tangential antapical view of the theca.Scale bars = 10 lm (a–c, e–l, n and o), 2 lm (d and m).
854 �ERIC POTVIN ET AL.

Sulcus: The sulcus was composed of five plates:the anterior sulcal (as), the left (ls) and right (rs)sulcal, the median sulcal (ms), and the posteriorsulcal (ps) plates (Fig. 3e). The as plate slightlyinvaded the epitheca and contacted the 1′, 1″, and7″ plates (Fig. 5c). The ms plate was locatedbetween the ls and rs plates (Fig. 5k). The ls, ms,and rs plates were oriented toward the right side ofthe theca (Fig. 5c). The ls plate penetrated deeplyin the ps plate (Fig. 5, c and n). The rs platesupported a list on its left side. The list began itsdevelopment at the lower side of the anterior sulcalplate and increased in size as it developed towardthe antapex with a slight deviation to the right (5c).The base of the list was supported by a ridgeerected from the ls plate (Fig. 5n). The ps platewas large, pentagonal, and in contact with the1‴, 5‴, 1′′′′, and 2′′′′ plates (Fig. 5, n and o).Hypotheca: The 1‴ plate was taller than wide,
whereas the 5‴ plate was wider than tall (Fig. 5,k and o). The 5‴ plate was larger than the 1‴ plate(Fig. 5o). The 1‴, 3‴, and 5‴ plates were pentago-nal, whereas the 2‴ and 4‴ plates were quadrangularand similar in size (Fig. 5o). Both antapical plates(1′′′′ and 2′′′′) were large, pentagonal, and of similarsize (Fig. 5o).Genetics. Variability: Good quality sequences from
the genes coding for the SSU rRNA and the LSUrRNA were obtained from five motile cells that ger-minated from different cysts. The sequences werevariable at three loci.Phylogeny: Morphological results led us to include
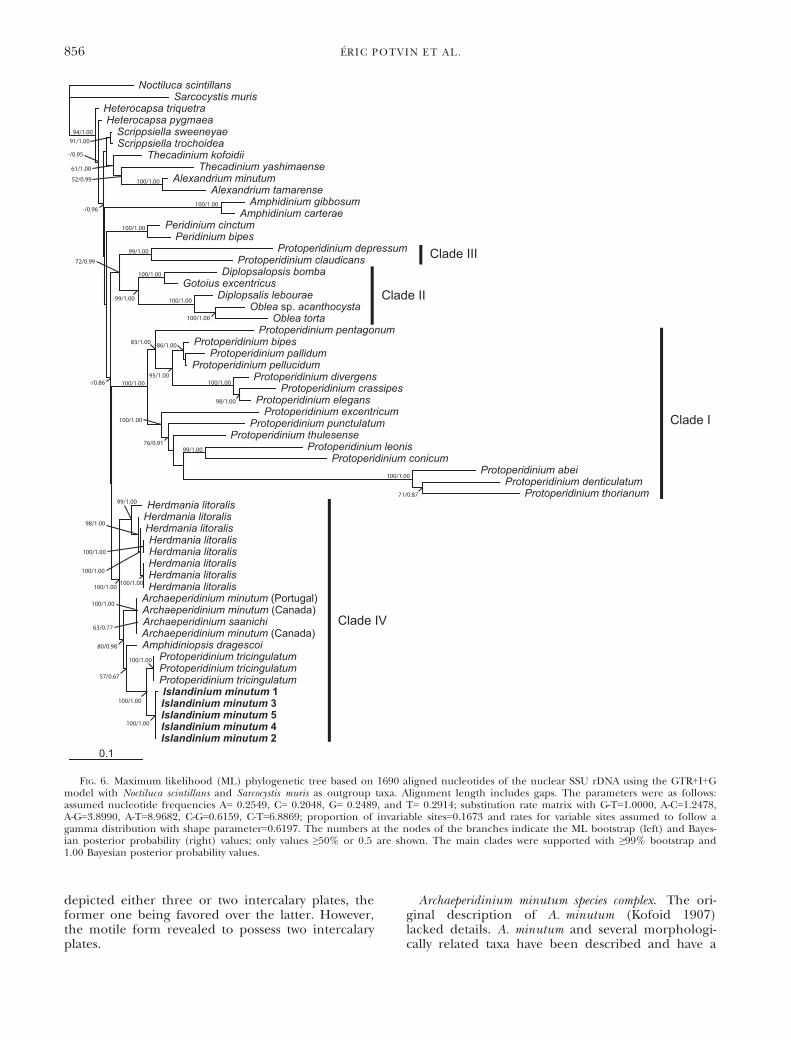
all sequences of Protoperidiniaceae available. TheProtoperidiniaceae clustered into four main cladesbased on SSU rDNA (Fig. 6). Three clades corre-sponded to clade I–III as defined by Ribeiro et al.(2010). Clade I was composed of the majority of thespecies belonging to the genus Protoperidinium fromwhich data are available, clade II was composed ofspecies of the genera Diplopsalopsis, Gotoius, Diplop-salis, and Oblea, and clade III was composed of thespecies of the genus Protoperidinium. The last cladecorresponded to clade IV named by Yamaguchiet al. (2011). Clade IV was composed of both ben-thic and pelagic species of the genera Herdmania,Amphidiniopsis, Protoperidinium, and Archaeperidinium.I. minutum grouped with clade IV as a sister taxonof the species Protoperidinium tricingulatum. Theclade produced by I. minutum and P. tricingulatumwas perfectly supported and reproduced in all phy-logenetic analyses. The Protoperidiniaceae clusteredin three main clades based on LSU rDNA (Fig. 7).Clades A and B, as defined by Ribeiro et al. (2010),were composed of the majority of the species belong-ing to the genus Protoperidinium. Clade D, named byYamaguchi et al. (2011), was composed, similar toclade IV, of both benthic and pelagic species of thegenus Herdmania and Archaeperidinium, respectively.I. minutum was included in clade D in basal positionto the Herdmania and Archaeperidinium clade. The
clades were well reproduced in all analyses and inmost cases well supported.
DISCUSSION
The germination experiments of distinct roundbrown spiny cyst morphotypes from several geo-graphic regions and contrasting environments pro-duced motile forms similar to Archaeperidiniumminutum (Table 3). Wall and Dale (1968) describedfor the first time an A. minutum cyst morphotype.Their results were based on the morphology ofhatched cells obtained by incubation experimentsof cysts isolated from Woods Hole, United States.Fukuyo et al. (1977), working with sediments fromJapan and through incubation experiments of adifferent cyst morphotype, also described a cyst–theca relationship for A. minutum. Ribeiro et al.(2010) described another morphotype similar toA. minutum from Portugal through cyst incubationexperiments. Then, Mertens et al. (2012) incu-bated cysts from the coast of British Columbia,Canada, that germinated to produce motile formsof A. minutum as described by Yamaguchi et al.(2011) and a new species closely related morpho-logically to A. minutum designated as A. saanichi.The motile form obtained from I. minutum wasclearly related morphologically to A. minutum.Detailed comparison with motile forms of thegenus Archaeperidinium and Protoperidinium (Table 4)established that the motile form of I. minutum hada unique set of features enabling its distinction.This is the first cyst–theca relationship formallyestablished for the cyst genus Islandinium.Cyst–theca comparison. The cysts used in incuba-
tion experiments did not show any paratabulationbesides the archeopyle, which was variable in shape.The archeopyle of the epitype of I. minutum is simi-lar to those observed in the excystment experimentsof this study (Fig. 8). The comparison of thesearcheopyles with the epitheca of the motile cellconfirmed the hypothesis of Head et al. (2001) onthe apical nature of the archeopyle reflecting the 2′,3′, and 4′ plates of the corresponding theca. Theasymmetrical arrangement of the paraplates noticedby Head et al. (2001) also reflected such asymmetryof the apical plates on the theca. While the disposi-tion of the plate producing this asymmetry was sta-ble on the theca, the disposition of the paraplateson the cyst was variable (Fig. 8). Furthermore, thethecal variability on the left side might reflect theuncertainty in the interpretation of the second api-cal plate (2′) by Head et al. (2001), which theyhypothesized having four or five sides based onobservation on cysts. An ortho-style first apical platewas assumed, and appears likely, but accessorysutures were not seen that would confirm this. Suchassumption has now been confirmed by observationson the epitheca of the motile cell. The hypotheticalepithecal tabulation proposed by Head et al. (2001)
MORPHOLOGY AND PHYLOGENETIC POSITION OF ISLANDINIUM MINUTUM 855
depicted either three or two intercalary plates, theformer one being favored over the latter. However,the motile form revealed to possess two intercalaryplates.
Archaeperidinium minutum species complex. The ori-ginal description of A. minutum (Kofoid 1907)lacked details. A. minutum and several morphologi-cally related taxa have been described and have a
0.1
Noctiluca scintillansSarcocystis muris
Heterocapsa triquetraHeterocapsa pygmaea
Scrippsiella sweeneyaeScrippsiella trochoidea
Thecadinium kofoidiiThecadinium yashimaense
Alexandrium minutumAlexandrium tamarense
Amphidinium gibbosumAmphidinium carterae
Peridinium cinctumPeridinium bipes
Protoperidinium depressumProtoperidinium claudicans
Diplopsalopsis bombaGotoius excentricus
Diplopsalis lebouraeOblea sp. acanthocysta
Oblea tortaProtoperidinium pentagonum
Protoperidinium bipesProtoperidinium pallidum
Protoperidinium pellucidumProtoperidinium divergens
Protoperidinium crassipesProtoperidinium elegans
Protoperidinium excentricumProtoperidinium punctulatum
Protoperidinium thulesenseProtoperidinium leonis
Protoperidinium conicumProtoperidinium abei
Protoperidinium denticulatumProtoperidinium thorianum
Herdmania litoralisHerdmania litoralisHerdmania litoralisHerdmania litoralisHerdmania litoralisHerdmania litoralisHerdmania litoralisHerdmania litoralis
Archaeperidinium minutum (Portugal)Archaeperidinium minutum (Canada)Archaeperidinium saanichiArchaeperidinium minutum (Canada)Amphidiniopsis dragescoi
Protoperidinium tricingulatumProtoperidinium tricingulatumProtoperidinium tricingulatumIslandinium minutum 1Islandinium minutum 3Islandinium minutum 5Islandinium minutum 4Islandinium minutum 2
Clade IV
Clade I
Clade II
Clade III
63/0.77
76/0.91
71/0.87
100/1.00
98/1.00
86/1.00
100/1.00
99/1.00
-/0.95
98/1.00
52/0.99
100/1.00
61/1.00
91/1.0094/1.00
100/1.00
100/1.00
100/1.00
99/1.00
100/1.00
100/1.00
100/1.0099/1.00
72/0.99
100/1.0095/1.00
83/1.00
100/1.00
99/1.00
100/1.00
100/1.00
100/1.00
57/0.67
80/0.98
100/1.00
100/1.00
100/1.00100/1.00
-/0.96
-/0.86
FIG. 6. Maximum likelihood (ML) phylogenetic tree based on 1690 aligned nucleotides of the nuclear SSU rDNA using the GTR+I+Gmodel with Noctiluca scintillans and Sarcocystis muris as outgroup taxa. Alignment length includes gaps. The parameters were as follows:assumed nucleotide frequencies A= 0.2549, C= 0.2048, G= 0.2489, and T= 0.2914; substitution rate matrix with G-T=1.0000, A-C=1.2478,A-G=3.8990, A-T=8.9682, C-G=0.6159, C-T=6.8869; proportion of invariable sites=0.1673 and rates for variable sites assumed to follow agamma distribution with shape parameter=0.6197. The numbers at the nodes of the branches indicate the ML bootstrap (left) and Bayes-ian posterior probability (right) values; only values ≥50% or 0.5 are shown. The main clades were supported with ≥99% bootstrap and1.00 Bayesian posterior probability values.
856 �ERIC POTVIN ET AL.

complex taxonomic history. A. monospinum (Paul-sen) Zonneveld et Dale, A. aspidiotum Balech, andA. constrictum Ab�e, all have a globular shape, epit-heca and hypotheca of approximately the samesize, a short but pronounced apical horn, noantapical horns or spines, a well-developed list
covering the sulcal area and the plate formula Po,X, 4′, 2a, 7″, 4C (3+t), 6S, 5‴, 2′′′′. Meunier (1919),Lebour (1925), Ab�e (1936), and Schiller (1937),among others, mentioned these taxa, either doubt-ing their identity as individual species or regardingthem as synonyms of A. minutum.
0.1
Sarcocystis murisToxoplasma gondii
Amphidinium carteraeAmphidinium steinii
Heterocapsa arcticaHeterocapsa triquetra
Thecadinium kofoidiiThecadinium yashimaense
Alexandrium minutumAlexandrium tamarense
Scrippsiella trochoideaPeridinium cinctum
Peridinium willeiDiplopsalis lenticula
Herdmania litoralisHerdmania litoralis
Archaeperidinium minutum (Portugal)Archaeperidinium saanichiArchaeperidinium minutum (Canada)Islandinium minutum 1Islandinium minutum 3Islandinium minutum 5Islandinium minutum 2Islandinium minutum 4Preperidinium meunieri
Protoperidinium steidingeraeProtoperidinium claudicans
Protoperidinium depressumProtoperidinium oblongum
Protoperidinium conicoidesProtoperidinium denticulatum
Protoperidinium abeiProtoperidinium excentricum
Protoperidinium conicumProtoperidinium leonis
Protoperidinium punctulatumProtoperidinium pentagonum
Protoperidinium bipesProtoperidinium pallidum
Protoperidinium pellucidumProtoperidinium angustumProtoperidinium elegans
Protoperidinium divergensProtoperidinium crassipes
100/1.00
100/1.00
100/1.00
100/1.00
100/1.00
100/1.00
100/1.00
62/0.86
100/1.00
100/1.00
68/0.95
100/1.00
100/1.00
98/0.99
74/0.98
71/0.97
89/1.00
70/0.80
-/0.6495/1.00
50/0.68-/0.50
92/1.00 100/1.00
-/0.98
83/0.99
-/0.94
-/0.77
Clade A
Clade B
Clade D
56/0.99
100/1.00
FIG. 7. Maximum likelihood (ML) phylogenetic tree based on 717 aligned nucleotides of the nuclear LSU rDNA using the GTR+I+Gmodel with Sarcocystis muris and Toxoplasma gondii as outgroup taxa. Alignment length includes gaps. The parameters were as follows:assumed nucleotide frequencies A=0.2550, C=0.1829, G=0.2835, and T=0.2786; substitution rate matrix with G-T=1.0000, A-C=1.3456,A-G=3.4344, A-T=1.0364, C-G=0.4794, C-T=6.4692; proportion of invariable sites=0.1183 and rates for variable sites assumed to follow agamma distribution with shape parameter=1.1031. The numbers at the nodes of the branches indicate the ML bootstrap (left) and Bayes-ian posterior probability (right) values; only values ≥50% or 0.5 are shown. The main clades were supported with ≥92% bootstrap and1.00 Bayesian posterior probability values with the exception of clade B showing less support (50%, 0.68).
MORPHOLOGY AND PHYLOGENETIC POSITION OF ISLANDINIUM MINUTUM 857

TABLE3.
Morphologicalfeaturesofthecyst
ofIslandinium
minutum
andspeciesofthege
nusArcha
eperidinium
andProtoperidinium
Gen
us
Islandinium
Archaeperidinium
Protoperidinium
Species
I.minutum
A.minutum
A.minutuma
A.minutum
A.minutum
A.saan
ichi
A.monospinum
P.parthenopes
P.am
erican
um
P.tricingulatum
Locality
Arctic
(Baffin
Bay)
United
States
(Woods
Hole)
Japan
(Ham
ana
lake
)
Portugal
(Mondeg
oestuary)
Can
ada
(British
Columbia
Coast)
Can
ada
(British
Columbia
Coast)
Norw
ay(O
slo
fjord)
Japan
(Omura
bay)
Scotlan
d(W
est
coast)
The
Netherlands
(Wad
den
Sea,
Huibertsplaat)
Gen
erality
Brown
Spiny
Spherical
Pale-gree
norbrown
Spiny
Spherical
Brown
Spiny
Spherical
Lightbrown
Spiny
Subspherical
Lightbrown
Spiny
Spherical
tosubspherical
Hazelbrown
Spiny
Spherical
tosubspherical
Palebrown
Spiny
Subspherical
Palebrown
Without
spine
Spherical
Cap
sulated
Palebrown
Without
spine
Spherical
Cap
sulated
Lightbrown
Spiny
Spherical
Diameter
(lm)
27–5
041
–43
28–3
225
–34
24–3
436
–58
28–4
024
–34
35–5
227
–34
Wall
Granulated
Rings
Red
uced
spines
betwee
nregu
larspines
Minutely
corrugated
–Finely
gran
ulated
Smooth
Bumps
betwee
nprocesses
arrange
din
apolygo
nal
pattern
aroundthe
spines
Smooth
Bumpsor
reducedspines
betwee
nregu
larspines
–Granulated
Smooth
capsule,
thin,pale
brownish-
orange
,someparts
folded
sharply
Granulated
Smooth
capsule,
hollow,fold
onsome
specim
ens
Smooth
toslightly
gran
ulated
surface
Process
Len
gth:
2–7lm
Basal
pericoel,
solid,
extrem
ity
acuminate,
random
distribution
Len
gth:
7–9lm
Flat-topped
distal
extrem
ities
(pointed
spinulesat
theouter
rimsof
process
extrem
ities),
circular
bases,
hollow,~4
0processes
Len
gth:
3–5lm
Acu
minate
andcu
rved
Len
gth:
4–9lm
Tap
ered
stem
s,minutely
expen
ded
tips,
circularin
section,
hollow,
often
curved
,even
lydistributed
Len
gth:
2–8lm
Distally
solidwith
pericoels
extending
variab
lyalongthe
process,
acuminate
orcapitate
withdistal
expan
sions,
tapering
anderect,
never
fused
or
branch
edat
theen
d,
process
bases
circularin
section,
even
lydistributed
Len
gth:
3–8lm
Cap
itate,
endingwith
distal
expan
sions
andtapering,
hollowan
dge
nerally
erect,
canbeslightly
recu
rved
,never
fusedor
branch
edat
theen
d,
process
bases
are
circular
insection,
even
lydistributed
Len
gtha:
3–7lm
Len
gthb:
3lm
a:Large
,hollow,
capitatean
dbifurcated
or
double-
branch
edat
thedistalen
ds,
random
distribution;
b:So
lid,
acuminate,
andrandom
distribution
betwee
nlarge
processes
None
None
Len
gth:
5–7lm
Solidwith
hollow
proximal
base,
slen
der,
capitateor
cauliforate
without
branch
ing
atdistal
extrem
ities,
circularat
the
proximal
basein
opticalcu
t,random
distribution
858 �ERIC POTVIN ET AL.

Based on Balech (1964a,b), it is possible to distin-guish the previous taxa from each other. A. monospi-num has regularly shaped plates and plate 1a isconnected to 1′. Plates on the left side of the thecaare larger, and thus cells tend to be asymmetrical.A. constrictum is generally symmetrical, plates 1a and1′ are not connected, and the sulcal list is less devel-oped than other species. A. minutum has irregularlyshaped plates, plates 1a and 1″ are not connected,the plates on the right side of the theca tend to belarger than the plates on the opposite side, and thewell-developed sulcal list is ear shaped. In A. aspidio-tum, plates on the right side of the theca are largerthan those on the left side, the sulcal list iswell developed and has a pointed posterior end.A. constrictum was later considered a synonym ofA. monospinum (Ab�e 1981). Later, after furtherobservations, Balech et al. (1984) concluded thatA. minutum and A. aspidiotum were more difficult todifferentiate.The comparison of A. minutum described from
different localities indicates site-related morphologi-cal differences such as the presence or absence ofan apical horn, divergent plate ornamentations, andvariable number of cingular or sulcal plates(Table 4) bringing more confusion.This complex situation, revealed by the motile
cells similar to A. minutum as well as their cystsshowing morphological variability depending of thelocality, likely reflects a species complex. Knowledgeabout this species complex recently increased fol-lowing the incubation of round brown spiny cystmorphotypes from several geographic regions andthe subsequent description of the motile cell as dis-cussed above.Phylogenetic analyses. The molecular phylogenetic
analyses of the sequences within the Protoperidinia-ceae based on the SSU rDNA resulted in four well-supported clades (clades I–IV; Fig. 6). All of thespecies within clades I–III (Fig. 6) have a cingulumcomprised of three plates and a transitional plate,congruent with the diagnosis of the Protoperidinia-ceae (Fensome et al. 1993). In contrast, the fourthclade (clade IV; Fig. 6) contains A. minutum, whichhas a typical tabulation pattern of the Protoperidini-aceae, and sand-dwelling dinoflagellates such asH. litoralis and Amphidiniopsis dragescoi. P. tricingula-tum is the only species of the genus Protoperidiniumincluded in clade IV. Ribeiro et al. (2010) initiallyfound that A. minutum (previously P. minutum) wasnot closely related to other Protoperidinium species.In the study of Yamaguchi et al. (2011), P. minutumfrom Portugal and Canada branched with H. litoralisand Amphidiniopsis dragescoi (previously Thecadiniumdragescoi; Hoppenrath et al. 2012). P. minutum fromPortugal and Canada formed a strongly supportedmonophyletic clade closely related to Thecadiniumdragescoi. These species were only very distantlyrelated to the clade that contains the type species ofthe genus Protoperidinium, P. pellucidum (clade I;T
ABLE3.
Continued
Gen
us
Islandinium
Archaeperidinium
Protoperidinium
Species
I.minutum
A.minutum
A.minutuma
A.minutum
A.minutum
A.saan
ichi
A.monospinum
P.parthenopes
P.am
erican
um
P.tricingulatum
Archeo
pyle
Apical
Saphopylic
Intercalary
(1aor2a?)
–
Intercalary
Theropylic
Intercalary
Theropylic
Intercalary
Theropylic
Intercalary
Theropylic
Undetermined
Theropylic
Apical
Saphopylic
Apical
Saphopylic
Apical
Theropylic
Referen
ces
Thisstudy
Wallan
dDale(196
8)Fuku
yoet
al.
(197
7)
Ribeiro
etal.
(201
0)
Mertens
etal.
(201
2)
Mertenset
al.
(201
2)Zonneveldan
dDale(199
4)Kaw
amian
dMatsuoka
(200
9)
Lew
isan
dDodge
(198
7)
Kaw
ami
etal.
(200
9)
aLew
iset
al.(198
4)retrievedsimilar
morphotypes
intheloch
esofthewestco
astofScotlan
d.
MORPHOLOGY AND PHYLOGENETIC POSITION OF ISLANDINIUM MINUTUM 859
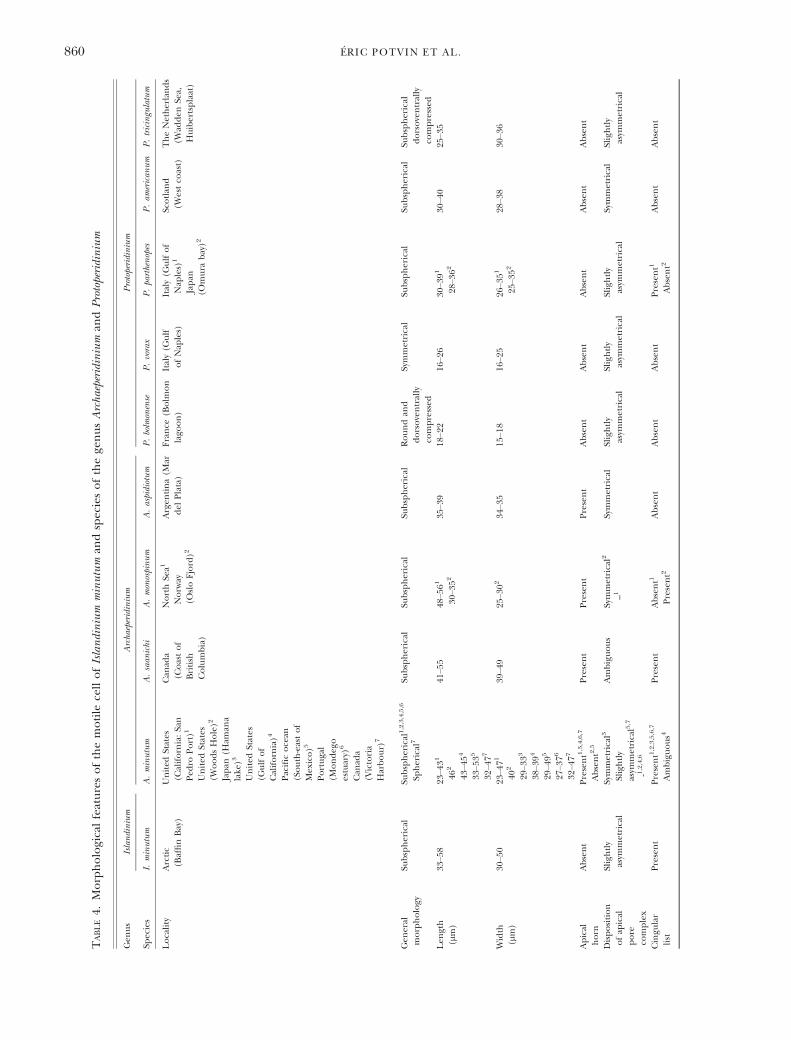
TABLE4.
Morphologicalfeaturesofthemotile
cellofIslandinium
minutum
andspeciesofthege
nusArcha
eperidinium
andProtoperidinium
Gen
us
Islandinium
Archa
eperidinium
Protoperidinium
Species
I.minutum
A.minutum
A.saan
ichi
A.monospinum
A.aspidiotum
P.bolmonense
P.vorax
P.parthenopes
P.am
erican
um
P.tricingulatum
Locality
Arctic
(BaffinBay)
United
States
(California:San
Ped
roPort)1
United
States
(WoodsHole)2
Japan
(Ham
ana
lake
)3
United
States
(Gulfof
California)4
Pacificocean
(South-eastof
Mex
ico)5
Portugal
(Mondeg
oestuary)
6
Can
ada
(Victoria
Harbour)
7
Can
ada
(Coastof
British
Columbia)
NorthSe
a1
Norw
ay(O
sloFjord)2
Argen
tina(M
ardel
Plata)
France
(Bolm
on
lago
on)
Italy(G
ulf
ofNap
les)
Italy(G
ulfof
Nap
les)
1
Japan
(Omura
bay)2
Scotlan
d(W
estco
ast)
TheNetherlands
(Wad
den
Sea,
Huibertsplaat)
Gen
eral
morphology
Subspherical
Subspherical
1,2,3,4,5,6
Spherical
7Su
bspherical
Subspherical
Subspherical
Roundan
ddorsoventrally
compressed
Symmetrical
Subspherical
Subspherical
Subspherical
dorsoventrally
compressed
Len
gth
(lm)
33–5
823
–431
462
43–4
54
33–5
35
32–4
77
41–5
548
–561
30–3
5235
–39
18–2
216
–26
30–3
91
28–3
62
30–4
025
–35
Width
(lm)
30–5
023
–471
402
29–3
33
38–3
94
29–4
95
27–3
76
32–4
77
39–4
925
–302
34–3
515
–18
16–2
526
–351
25–3
52
28–3
830
–36
Apical
horn
Absent
Present1,3,4,6,7
Absent2,5
Present
Present
Present
Absent
Absent
Absent
Absent
Absent
Disposition
ofap
ical
pore
complex
Slightly
asym
metrical
Symmetrical3
Slightly
asym
metrical5,7
-1,2,4,6
Ambiguous
Symmetrical2
–1Symmetrical
Slightly
asym
metrical
Slightly
asym
metrical
Slightly
asym
metrical
Symmetrical
Slightly
asym
metrical
Cingu
lar
list
Present
Present1,2,3,5,6,7
Ambiguous4
Present
Absent1
Present2
Absent
Absent
Absent
Present1
Absent2
Absent
Absent
860 �ERIC POTVIN ET AL.

TABLE4.
Continued
Gen
us
Islandinium
Archa
eperidinium
Protoperidinium
Species
I.minutum
A.minutum
A.saan
ichi
A.monospinum
A.aspidiotum
P.bolmonense
P.vorax
P.parthenopes
P.am
erican
um
P.tricingulatum
Ornam
entation
ofplates
Scattered
pores
Faintreticu
lation
Spines
Arowofporeson
andbelowthe
cingu
lum
Scatteredpores,
noreticu
lation1
–2,4,5
Scatteredpores
andmicrogran
ules3
Inco
mplete
polygo
nal
reticu
lations6
Scatteredpores7
Microreticu
late
Scatteredpores
Granulatedsurface1
Finereticu
lation2
–Scatteredpores,
poresat
the
margins
Poresat
the
margins
Pores,
protuberan
cean
dfaint
reticu
lations1
Poresaligned
atthe
margins1
,2
Arowofpores
onthecingu
lum
1
Smooth
with
scattered
pores2
Undulations
andpores
aligned
atthemargins
Smooth
with
scattered
pores
Dispositionof
intercalary
plates
Asymmetrical
Symmetrical1,2,3,7
–4,6
Slightly
asym
metrical5
Asymmetrical
Ambiguous1
Symmetrical2
Slightly
asym
metrical
Asymmetrical
Asymmetrical
Asymmetrical
Asymmetrical
Asymmetrical
Intercalaryplates
2a2a
2a2a
2a3a
3a3a
4a2a
Cingu
larplates
3c4(t+3)c3
,6,7
–2,4
Ambiguous1
,5
4(t+3)c
4(t+3)c2
–14(t+3)c
3c3c
4c4c
3c
Contact
betwee
n1a
and1″
Present
Absent
Absent1,2,3,5,6,7
–4Absent
Present1
Absent2
Absent
Present
Present
Present
Present
Present
Dispositionof
thesuturesat
thejunction
ofcingu
lar
plates
Ven
tral
Ven
tral
3,6,7
–2,4
Ambiguous1
,5
Ven
tral
Ven
tral
2
–1Ven
tral
Ven
tral
Dorsal
Ven
tral
Dorsal
Ven
tral
Dorsal
Ven
tral
Dorsal
Ven
tral
Sulcal
plates
5s4s
56s
3,6
–1,2,4,7
6s5s
2 (withoutt)
2
–1
6s4(?)s
5s6s
7s6s?
Cyst
production
Yes
Yes2
,3,6,7
–1,4,5
Yes
Yes2 –1
––
– (Notobserved
)Ye
s2 –1Ye
sYe
s
Referen
ces
Thisstudy
Kofoid
(190
7)1,
Wallan
dDale
(196
8)2,
Fuku
yoet
al.
(197
7)3,
Hern� an
dez-Becerril
(199
1)4,
Oko
lodko
v(200
5)5,
Ribeiro
etal.(201
0)6,
Yamaguch
iet
al.(201
1)7
Mertenset
al.
(201
2)Pau
lsen
(190
8)1
Zonneveldan
dDale(199
4)2
Balech(198
8)Chom� erat
and
Cout�e
(200
8)Sian
oan
dMontresor
(200
5)
Zingo
nean
dMontresor
(198
8)1
Kaw
amian
dMatsuoka
(200
9)2
Lew
isan
dDodge
(198
7)
Kaw
ami
etal.
(200
9)
MORPHOLOGY AND PHYLOGENETIC POSITION OF ISLANDINIUM MINUTUM 861

Fig. 6). This situation led to the re-introduction ofArchaeperidinium as a genus (Yamaguchi et al. 2011).Following this, Mertens et al. (2012) described anew species in the genus Archaeperidinium, A. saani-chi. The latter clustered with A. minutum. Species ofthe genus Archaeperidinium still formed a stronglysupported monophyletic clade with the addition ofI. minutum. However, P. tricingulatum is more closelyrelated to I. minutum. The closer relationshipbetween I. minutum and P. tricingulatum as well asthe closer relatedness of Amphidiniopsis dragescoi tothe previous species made I. minutum and Archaepe-ridinium spp. paraphyletic.
The molecular phylogenetic analyses of thespecies classified in the Protoperidiniaceae based onLSU rDNA formed three supported clades (cladesA, B, D; Fig. 7). Clades A and B consisted of similarspecies to clades I and III in the SSU rDNA ana-lyses, respectively (Fig. 7). Clade C described byYamaguchi et al. (2011) was made of two diplopsa-lid species (Diplopsalis lenticula and Preperidiniummeunieri) and was not reproduced as in Ribeiroet al. (2010) and Mertens et al. (2012). Clade Dcontaining I. minutum also included H. litoraliswhich rendered the unification of I. minutum andArchaeperidinium spp. and made these species para-phyletic, suggesting that Islandinium might be a sep-arate genus from Archaeperidinium.Ecology. The dinoflagellate cyst I. minutum is a
major paleoecological indicator of the duration ofthe seasonal sea-ice cover in the Arctic. However,direct proof of the association of the species to thesea-ice cover was never established. While the distri-bution of the cyst is well known in surface sedimentsamples, the motile cell germinated from the cyst ofI. minutum has never been reported in the CanadianArctic. However, sequences ≥99% similar to the SSUrDNA of I. minutum have been retrieved as part of arecent study using high throughput ampliconsequencing on sea-ice samples collected in theBeaufort Sea in April (Comeau et al. 2013). Boththe lack of access to the high Arctic in late winterand early spring and the fragility of motile cells mayexplain the lack of previous records. The motile cellof I. minutum, like Polarella glacialis (Montresor et al.2003), could live in association with sea-ice andform resting cysts when first year sea-ice melts.Motile cells of Polarella are rarely reported from Arc-tic sea ice but are common in Antarctic sea ice(Thomson et al. 2004), and are readily isolatedfrom either sea ice or the surface water of the polaroceans. Since this species is photosynthetic it islikely much easier to bring and maintain in culturecompared to the heterotrophic species I. minutum.Systematics. Although the motile cell of I. minutum
was related morphologically to the genus Archaepe-ridinium, the paraphyly of both SSU rDNA and LSUrDNA phylogenetic analyses suggest that thesespecies belong to different genera.
Most species in the genus Archaeperidinium possessan apical horn, all have four cingular plates includingthe transitional plate, and sutures at the junction ofcingular plates all disposed ventrally. The sutures atthe junction of the cingular plates of I. minutum arealso located ventrally. Therefore, this feature cannotbe used to distinguish I. minutum from Archaeperidini-um spp. at the genus level. However, the motile cell ofI. minutum does not have an apical horn and possessesonly three cingular plates lacking the transitional one.These features may therefore be useful to distinguishthe motile cells of the genus Islandinium from those ofthe genus Archaeperidinium. Only species of the genusArchaeperidinium described by Wall and Dale (1968)and Okolodkov (2005) designated as Peridinium minu-tum and Protoperidinium minutum, respectively, weredescribed without an apical horn (Table 4). Thesespecies might therefore be more closely related toI. minutum. Other species such as Protoperidinium bol-monense, P. vorax, P. parthenopes, and P. americanumcould also be related to I. minutum since they sharemultiple features such as the absence of apical horn,the contact between the first precingular (1′) andanterior intercalary (1a) plates, and the asymmetricaldisposition of the anterior intercalary plates. How-ever, the ventral and dorsal dispositions of the suturesat the junction of cingular plates seem to distinguishthem from the motile cells of I. minutum. The fourcingular plates of P. parthenopes and P. americanum, asopposed to three for P. bolmonense and P. vorax, seemto distinguish them further.Cysts produced by motile cells of the genus Archae-
peridinium are all brown, spherical to subspherical,and spiny, and appear to have in most cases an inter-calary and theropylic archeopyle. The cyst of I. minu-tum is brown, spherical, and spiny. Therefore, thesefeatures cannot be used to distinguish these speciesat the genus level. However, the archeopyle of I. min-utum is distinctive by being apical and saphopylic andcan therefore have some value at the genus level. Thecyst from Wall and Dale (1968) produced a motileform without horn following incubation, which is afeature highlighted here as distinctive of the motilecells of the genus Islandinium. The archeopyle of thecyst of Wall and Dale (1968) was described as interca-lary with some uncertainty. The operculum ofrecently germinated I. minutum is adnate. Paraplatesof their operculum can still be in place on the cystand paraplates are not always free from each other.This can bring confusion in the interpretation of thearcheopyle if not looked at closely. Cysts from WoodsHole (United States) described by Wall and Dale(1968) should be reassessed to fully document thearcheopyle.Protoperidinium americanum and P. parthenopes also
produce cysts with apical archeopyles as I. minutum,but their cysts lack processes and are capsulate, andtherefore rather different from the round brownspiny cysts.
862 �ERIC POTVIN ET AL.

The motile form of P. tricingulatum is very pecu-liar in the genus Protoperidinium by, as its name sug-gests, having only three cingular plates lacking thetransitional plate and two anterior intercalary plates(Kawami et al. 2009). The absence of transitionalplate is the only feature that prevents its assignmentto the genus Archaeperidinium. Furthermore, this spe-cies does not have an apical horn which is a featurethat distinguishes it from species of the genusArchaeperidinium. These features that distinguish thisspecies from both Protoperidinium spp. and Archaepe-ridinium spp. are however common to the motileform of I. minutum further stressing the close rela-tionship between both species. The similarity ofP. tricingulatum and I. minutum is also reflected withthe apical nature of the archeopyle which is peculiaramong round brown spiny cyst. This suggests that aset of features regarding the motile form and thecyst unify and distinguish them from Protoperidiniumspp. and Archaeperidinium spp. The close relation-ship between I. minutum and P. tricingulatum is fur-ther supported by the phylogenetic analyses basedon SSU rDNA. Therefore, we transfer P. tricingula-tum to the genus Islandinium. Furthermore, we treatIslandinium as a non-fossil genus and emend itsdescription as well as the species I. minutum toinclude the attributes of the motile form.
The motile cells included in the genus Islandini-um have the plate formula Po, X, 4′, 2a, 7″, 3C,4-7S, 5‴, 2′′′′. While the absence of an apical hornappears to have a possible value to distinguish spe-cies of the genera Islandinium and Archaeperidinium,the absence of transitional plate leading to theoccurrence of only three cingular plates is enoughto clearly distinguish both genera. Therefore, onlythe number of cingular plate was used at the genuslevel. Thecal plasticity from the main pattern canoccur. They differed from species of sections Avell-ana and Excentrica of the genus Protoperidinium by aflat sulcus, a sulcal list covering the sulcal area, andan equatorial cingulum lacking displacement.
The cyst genus Islandinium was defined to includebrown cysts having spheroidal central body whosewall is smooth to finely ornamented with numerous
processes that are non-tabular, and have acuminate,expanded, or branched tips. The definition of thegenus also characterizes the archeopyle as onlysaphopylic. However, the newly transferred speciesP. tricingulatum has a theropylic archeopyle and thisfeature needs to be added at the genus level. Thecyst of I. minutum also revealed to be proximoch-orate in the material collected from Baffin Bay. Fur-thermore, scanning electron microscopy revealedthat the spines of I. minutum were rough and notsmooth. These details were added or corrected inthe description of the cyst at both the genus andspecies levels.Genus Islandinium Head, Harland et Matthiessen
emend.Type of genus: Islandinium minutumMotile cellArmoured dinoflagellate. Plate formula Po, X, 4′,
2a, 7″, 3C, 4-7S, 5‴, 2′′′′. Thecal plasticity from themain pattern can occur. Chloroplasts absent. Differ-ing from species of sections Avellana and Excentricaof the genus Protoperidinium by the flat sulcus, thesulcal list covering the sulcal area, and the equato-rial cingulum lacking displacement.CystCysts are brown in color, having spheroidal
central body and whose wall is smooth to finelyornamented. Processes numerous, non-tabular, ter-minating distally in acuminate or expanded, includ-ing branched, tips. Principal and occasionallyaccessory archeopyle sutures are only clear expres-sion of tabulation. Archeopyle saphopylic or thero-pylic generally restricted to the apical series, butmay include the Po and X plates. Operculum freeor not; opercular pieces lost separately or not.Islandinium minutum (Harland et Reid) Head,
Harland et Matthiessen emend.Motile cellThecal cells. Length 33–58 (mean 44) lm and
width 30–50 (mean 41) lm. Epitheca is in domeshape and hypotheca is rounded. Cell has multipleglobular bodies in the cytoplasm of green and yel-low colors. Apical horn is absent. Thecal plates areornamented with faint reticulation and, with the
a b c d
2'3'
4'1'
2'3'
4'1'2'
3'4'
1'
3’
2a
1a2’
4’
1’ 1’’2’’
3’’
4’’5’’
6’’
7’’
FIG. 8. Comparison between the most common variation (conjunctum) of the epitheca observed and the archeopyles of the dinoflagel-late cyst Islandinium minutum. (a) Epitheca of the motile cell obtained from the dinoflagellate cyst I. minutum. The dash line representsthe outline of plates reflected by the cyst’s archeopyle. (b) Archeopyle of the epitype of I. minutum as in Head et al. (2001). (c–d) Archeo-pyles of I. minutum obtained from this study.
MORPHOLOGY AND PHYLOGENETIC POSITION OF ISLANDINIUM MINUTUM 863

exception of those of the APC, have pores sur-rounded by a circular rim and spines. The cingulumis excavated, not displaced and located in the equa-torial part of the cell. The sulcus is not excavated.The tabular formula is Po, X, 4′, 2a, 6–7″, 3C, 5S,5‴, 2′′′′. The first apical plate (1′) is ortho. Theintercalary plates are displaced toward the left sideof the epitheca. The first intercalary plate (1a) ishexagonal or heptagonal, the second (2a) is hexag-onal. The cingulum is composed of three plates ofdifferent length. The transitional plate is absent.The rs plate carries a list on its left side. Variationsof the plate pattern were observed on the left sideof the epitheca including contacts between theplates 2′, 1a, 1″ and 2″ and the fusion of the plates1″ and 2″. The plate 1a can be in contact with thefirst precingular plate (1″).
CystCysts are brown in color, proximochorate to cho-
rate, having spheroidal central body whose wall isgranulate. Processes numerous, erect, non-tabular,mostly solid to apiculocavate, tapering distally, termi-nating in acuminate tips, and having circular trans-verse section and rough surface. Principal and rareaccessory archeopyle sutures are only clear expressionof tabulation. Archeopyle apical and saphopylic, TypeA3, formed by loss of three apical plates, 2′, 3′, and 4′,and usually also canal plate and APC. Fourth apicalplate (4′) is large, occupying most of the right side ofthe apical region, and almost square in shape; apicalplates 2′ and 3′ occupy most of the left side of theapical region. Operculum free; opercular piecesreleased separately.
Islandinium tricingulatum (Kawami, van Wezel,Koeman et Matsuoka) comb. nov.
Basionym: Protoperidinium tricingulatum Kawamiet al. 2009
Motile cellMotile cell is subspherical without spine or horn,
25.0–35.0 lm long and 30.0–36.0 lm wide. Plate for-mula: Po, X, 4′, 2a, 7″, 3C, 6S?, 5′″, 2″″ (Figs. 3, e–p;4, a–c, Kawami et al. 2009). The first apical plate is ameta type. The first antapical plate is extremelysmall, and does not connect with the thirdpostcingular plate. The cingulum consists of threeplates.
CystCyst is round and brown. Cyst is smooth on the
surface and bears many solid and slender spineswith capitate distal tips randomly distributed on thesurface. Archeopyle theropylic. Parasutures areformed at the plate boundaries of 2′/1a (com-plete), 2′/2a (complete), 3′/2a (complete), 4′/2a(complete), 4′/6″ (incomplete) and 4′/7″ (incom-plete?).
We thank Dr Martin J. Head, Dr John Dodge and Dr YuriB. Okolodkov for providing relevant comments andliterature as well as the anonymous reviewers for valuablesuggestions. We thank the scientists, crew, and officers from
the CCGS Amundsen for there help during sampling. Thisarticle was funded by the MSc scholarship from theFonds de Recherche du Qu�ebec – Nature et Technologiesawarded to �Eric Potvin, the Natural Sciences and Engineer-ing Research Council of Canada Discovery Grants awardedto Andr�e Rochon and Connie Lovejoy, the NorthernScientific Training Program from the Department of IndianAffairs and Northern Development and ArcticNet.
REFERENCES
Abe, T. H. 1936. Report of the biological survey of Mutsu Bay. 29.Notes on the protozoan fauna of Mutsu Bay. II. Genus Peridi-nium; subgenus Archaeperidinium. Science Reports of TohokuUniversity, 4th Series. Biology 10:639–86.
Ab�e, T. H. 1981. Studies on the family Peridinidae. An unfinishedmonograph of armoured Dinoflagellata. Pub. Seto Mar. Biol.Lab. Special Pub. Ser. 6:1–413.
Balech, E. 1964a. El Plankton de Mar del Plata durante el period-o 1961–62. Bolet�ın del Instituto de Biolog�ıa Marina Mar delPlata. 4:1–49.
Balech, E. 1964b. Tercera contribuci�on al conocimiento delg�enero Peridinium. Revista del Museo Argentino de Cien-cias Naturales Bernardino Rivadavia. Hidrobiolog�ıa I:179–201.
Balech, E. 1988. Los Dinoflagellados del Atlantico Sudoccidental. Pub-licaciones Especiales Instituto Espanol de Oceanografia,Madrid, 1: 310 pp.
Balech, E., Akselman, R., Benavides, H. R. & Negri, R. M.1984. Suplemento a «Los dinoflagelados del Atl�anticoSudoccidental». Rev. Invest. Des. Pesq. INIDEP. Mar del Plata.4:5–20.
Bonnet, S., de Vernal, A., Gersonde, R. & Lembke-Jene, L. 2012.Modern distribution of dinocysts from the North PacificOcean (37–64°N, 144°E–148°W) in relation to hydrographicconditions, sea-ice and productivity. Mar. Micropaleontol.84–85:87–113.
Bonnet, S., de Vernal, A., Hillaire-Marcel, C., Radi, T. & Husum,K. 2010. Variability of sea-surface temperature and sea-icecover in the Fram Strait over the last two millennia. Mar.Micropaleontol. 74:59–74.
Bouimetarhan, I., Marret, F., Dupont, L. & Zonneveld, K. 2009.Dinoflagellate cyst distribution in marine surface sedimentsoff West Africa (17–6°N) in relation to sea-surface condi-tions, freshwater input and seasonal coastal upwelling. Mar.Micropaleontol. 71:113–30.
Chom�erat, N. & Cout�e, A. 2008. Protoperidinium bolmonensesp. nov. (Peridiniales, Dinophyceae), a small dinoflagellatefrom a brackish hypereutrophic lagoon (South of France).Phycologia 47:392–403.
Comeau, A. M., Philippe, B., Thaler, M., Gosselin, M., Poulin, M.& Lovejoy, C. 2013. Protists in Arctic drift and land-fast seaice. J. Phycol. 49:229–40.
Fensome, R. A., Taylor, F. J. R., Norris, G., Sarjeant, W. A. S.,Wharton, D. I. & Williams, G. L. 1993. A Classification ofLiving and Fossil Dinoflagellates. Micropaleontology specialpublication number 7, Micropaleontology Press, Massachu-setts, 351 pp.
Fritz, L. & Triemer, R. E. 1985. A rapid simple technique utilizingcalcofluor white M2R for the visualization of dinoflagellatethecal plates. J. Phycol. 21:662–4.
Fukuyo, Y., Kittaka, J. & Hirano, R. 1977. Studies on the cysts ofmarine dinoflagellates-1. Bull. Plankt. Soc. Jpn. 24:11–8.
Guillard, R. R. L. & Ryther, J. H. 1962. Studies of marine plank-tonic diatoms. I. Cyclotella nana Hustedt, and Detonula conferv-acea (Cleve) Gran. Can. J. Microbiol. 8:229–39.
Hall, T. A. 1999. BioEdit: a user-friendly biological sequencealignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 41:95–8.
Hamel, D., de Vernal, A., Gosselin, M. & Hillaire-Marcel, C. 2002.Organic-walled microfossils and geochemical tracers: sedi-mentary indicators of productivity changes in the North
864 �ERIC POTVIN ET AL.

Water and northern Baffin Bay during the last centuries.Deep-Sea Res. II 49:5277–95.
Harland, R., Reid, P. C., Dobell, P. & Norris, G. 1980. Recent andsub-Recent dinoflagellate cysts from the Beaufort Sea, Cana-dian Arctic. Grana 19:211–25.
Head, M. J. 1996. Modern dinoflagellate cysts and their biologicalaffinities. In Jansonius, J. & McGregor, D. C. [Eds.] Palynol-ogy: Principles and Applications, volume 3. American Associa-tion of Stratigraphic Palynologists Foundation, Texas, UnitedStates, pp. 1197–248.
Head, M. J., Harland, R. & Matthiessen, J. 2001. Cold marineindicators of the late Quaternary: the new dinoflagellate cystgenus Islandinium and related morphotypes. J. Quaternary Sci.16:621–36.
Hern�andez-Becerril, D. U. 1991. Protoperidinium (Dinophyceae)species in the gulf of California and off the coast of Baja Cal-ifornia. Anales del Instituto de Ciencias del Mar y Limnolog�ıa,UNAM. 18:77–88.
Hoppenrath, M., Selina, M., Yamaguchi, A. & Leander, B. 2012.Morphology and molecular phylogeny of Amphidiniopsisrotundata sp. nov. (Peridiniales, Dinophyceae), a benthicmarine dinoflagellate. Phycologia 51:157–67.
Horiguchi, T., Yoshizawa-Ebata, J. & Nakayama, T. 2000. Halosty-lodinium arenarium, gen. et sp. nov. (Dinophyceae), a coccoidsand-dwelling dinoflagellate from subtropical Japan. J. Phycol.36:960–71.
Huelsenbeck, J. P. & Ronquist, F. 2001. MRBAYES: Bayesian infer-ence of phylogenetic trees. Bioinformatics 17:754–5.
Kawami, H., Iwataki, M. & Matsuoka, K. 2006. A new diplopsalidspecies Oblea acanthocysta sp. nov. (Peridiniales, Dinophy-ceae). Plankt. Benthos Res. 1:183–90.
Kawami, H. & Matsuoka, K. 2009. A new cyst-theca relationshipfor Protoperidinium parthenopes Zingone & Montresor 1988(Peridiniales, Dinophyceae). Palynology 33:11–8.
Kawami, H., van Wezel, R., Koeman, R. P. T. & Matsuoka, K.2009. Protoperidinium tricingulatum sp. nov. (Dinophyceae), anew motile form of a round, brown, and spiny dinoflagellatecyst. Phycol. Res. 57:259–67.
Kofoid, C. A. 1907. Dinoflagellata of the San Diego region, III.Descriptions of new species. Univ. Calif. Publ. Zool.3:299–340.
Kofoid, C. A. 1909. On Peridinium steinii J€orgensen, with a noteon the nomenclature of the skeleton of the Peridinidae.Arch. Protistenk. 16:25–47.
Kogame, K., Horiguchi, T. & Masuda, M. 1999. Phylogeny of theorder Scytosiphonales (Phaeophyceae) based on DNAsequences of rbcL, partial rbcS, and partial LSU nrDNA.Phycologia 38:496–502.
Krepakevich, A. & Pospelova, V. 2010. Tracing the influence ofsewage discharge on coastal bays of Southern VancouverIsland (BC, Canada) using sedimentary records of phyto-plankton. Cont. Shelf Res. 30:1924–40.
Kunz-Pirrung, M. 2001. Dinoflagellate cyst assemblages in surfacesediments of the Laptev Sea region (Arctic Ocean) and theirrelationship to hydrographic conditions. J. Quaternary Sci.16:637–49.
Larkin, M. A., Blackshields, G., Brown, N. P., Chenna, R., McGett-igan, P. A., McWilliam, H., Valentin, F. et al. 2007. Clustal Wand Clustal X version 2.0. Bioinformatics 23:2947–8.
Lebour, M. V. 1925. The Dinoflagellates of Northern Seas. The Mar-ine biological association of the United Kingdom, Plymouth,250 pp.
Lef�evre, M. 1932. Monographie des esp�eces d’eau douce dugenre Peridinium. Arch. Bot. Mem. Caen. 2:1–208.
Lenaers, G., Maroteaux, L., Michot, B. & Herzog, M. 1989. Dino-flagellates in evolution. A molecular phylogenetic analysis oflarge subunit ribosomal RNA. J. Mol. Evol. 29:40–51.
Lewis, J. & Dodge, J. D. 1987. The cyst-theca relationship of Pro-toperidinium americanum (Gran & Braarud) Balech. J. Micropa-laeontol. 6:113–21.
Lewis, J., Dodge, J. D. & Tett, P. 1984. Cyst-theca relationships insome Protoperidinium species (Peridiniales) from Scottish sealochs. J. Micropalaeontol. 3:25–34.
Limoges, A., Kielt, J. F., Radi, T., Ru�ız-Fernandez, A. C. & deVernal, A. 2010. Dinoflagellate cyst distribution in surfacesediments along the south-western Mexican coast (14.76°Nto 24.75°N). Mar. Micropaleontol. 76:104–23.
Marret, F. & Zonneveld, K. A. F. 2003. Atlas of modern organic-walled dinoflagellate cyst distribution. Rev. Palaeobot. Palyno.125:1–200.
Matsuoka, K. & Bujak, J. P. 1988. Cenozoic dinoflagellate cystsfrom the Navarin Basin, Norton Sound and St. George Basin,Bering Sea. Bull. Fac. Liberal Arts Nagasaki Univ. 29:1–147.
Matthiessen, J., de Vernal, A., Head, M., Okolodkov, Y., Zonne-veld, K. & Harland, R. 2005. Modern organic-walleddinoflagellate cysts in Arctic marine environments and their(paleo-) environmental significance. Pal€aontologische Zeitsch-rift. 79:3–51.
Mertens, K. N., Verhoeven, K., Verleye, T., Louwye, S., Amorim,A., Ribeiro, S., Deaf, A. S. et al. 2009. Determining the abso-lute abundance of dinoflagellate cysts in recent marine sedi-ments: the Lycopodium marker-grain method put to the test.Rev. Palaeobot. Palyno. 157:238–52.
Mertens, K. N., Yamaguchi, A., Kawami, H., Ribeiro, S., Leander,B. S., Price, A. M., Pospelova, V., Ellegaard, M. & Matsuoka,K. 2012. Archaeperidinium saanichi sp. nov.: a new speciesbased on morphological variation of cyst and theca withinthe Archaeperidinium minutum J€orgensen 1912 species com-plex. Mar. Micropaleontol. 96–97:48–62.
Meunier, A. 1919. Microplancton de la mer Flamande. Partie 3.Les P�eridiniens. M�em. Mus. R. Hist. Nat. Belg. 8:1–116.
Montresor, M., Lovejoy, C., Orsini, L., Procaccini, G. & Roy, S.2003. Bipolar distribution of the cyst-forming dinoflagellatePolarella glacialis. Polar Biol. 26:186–94.
Mudie, P. J. & Rochon, A. 2001. Distribution of dinoflagellatecysts in the Canadian Arctic marine region. J. Quaternary Sci.16:603–20.
Okolodkov, Y. B. 2005. Protoperidinium Bergh (Dinoflagellata) inthe southeastern Mexican Pacific Ocean: part 1. Bot. Mar.48:284–96.
Paulsen, O. 1908. Peridiniales. In Brandt, K. & Apstein, C. [Eds.]Nordisches Plankton. Botanischer Teil, 18. Lipsius and Tischer,Kiel and Leipzig. pp. 1–124.
Posada, D. & Buckley, T. R. 2004. Model selection and modelaveraging in phylogenetics: advantages of akaike informationcriterion and bayesian approaches over likelihood ratio tests.Syst. Biol. 53:793–808.
Posada, D. & Crandall, K. A. 1998. MODELTEST: testing themodel of DNA substitution. Bioinformatics 14:817–8.
Pospelova, V., Chmura, G. L., Boothman, W. S. & Latimer, J. S.2005. Spatial distribution of modern dinoflagellate cysts inpolluted estuarine sediments from Buzzards Bay (Massachu-setts, USA) embayments. Mar. Ecol. Prog. Ser. 292:23–40.
Pospelova, V., Chmura, G. L. & Walker, H. A. 2004. Environmen-tal factors influencing the spatial distribution of dinoflagel-late cyst assemblages in shallow lagoons of southern NewEngland (USA). Rev. Palaeobot. Palyno. 128:7–34.
Pospelova, V., de Vernal, A. & Pedersen, T. F. 2008. Distributionof dinoflagellate cysts in surface sediments from the north-eastern Pacific Ocean (43–25°N) in relation to sea-surfacetemperature, salinity, productivity and coastal upwelling.Mar. Micropaleontol. 68:21–48.
Pospelova, V., Esenkulova, S., Johannessen, S. C., O’Brien, M. C.& Macdonald, R. W. 2010. Organic-walled dinoflagellate cystproduction, composition and flux from 1996 to 1998 in thecentral Strait of Georgia (BC, Canada): a sediment trapstudy. Mar. Micropaleontol. 75:17–37.
Pospelova, V. & Kim, S.-J. 2010. Dinoflagellate cysts in recent estu-arine sediments from aquaculture sites of southern SouthKorea. Mar. Micropaleontol. 76:37–51.
Price, A. M. & Pospelova, V. 2011. High-resolution sediment trapstudy of organic-walled dinoflagellate cyst production andbiogenic silica flux in Saanich Inlet (BC, Canada). Mar. Mi-cropaleontol. 80:18–43.
Radi, T., Bonnet, S., Cormier, M. A., de Vernal, A., Durantou, L.,Faubert, E., Head, M. J., Henry, M., Pospelova, V., Rochon,
MORPHOLOGY AND PHYLOGENETIC POSITION OF ISLANDINIUM MINUTUM 865

A. & Van Nieuwenhove, N. 2013. Operational taxonomy forround, brown, spiny dinocysts from high latitudes of theNorthern Hemisphere. Mar. Micropaleontol. 98:41–57.
Radi, T. & de Vernal, A. 2004. Dinocyst distribution in surfacesediments from the northeastern Pacific margin (40–60°N)in relation to hydrographic conditions, productivity andupwelling. Rev. Palaeobot. Palyno. 128:169–93.
Radi, T., Pospelova, V., de Vernal, A. & Barrie, J. V. 2007. Dinofla-gellate cysts as indicators of water quality and productivity inBritish Columbia estuarine environments. Mar. Micropaleontol.62:269–97.
Rausch, H., Larsen, N. & Schmitt, R. 1989. Phylogeneticrelationships of green algae Volvox carteri deduced fromsmall-subunit ribosomal RNA comparisons. J. Mol. Evol.29:255–65.
Ribeiro, S., Lundholm, N., Amorim, A. & Ellegaard, M. 2010. Pro-toperidinium minutum (Dinophyceae) from Portugal: cyst-thecarelationship and phylogenetic position on the basis of single-cell SSU and LSU rDNA sequencing. Phycologia 49:48–63.
Richerol, T., Rochon, A., Blasco, S., Scott, D. B., Schell, T. M. &Bennett, R. J. 2008. Distribution of dinoflagellate cysts in sur-face sediments of the Mackenzie Shelf and Amundsen Gulf,Beaufort Sea (Canada). J. Mar. Syst. 74:825–39.
Rochon, A. 2009. The ecology and biological affinity of Arcticdinoflagellates and their paleoceanographical significance inthe Canadian High Arctic. IOP Confer. Ser. 5:1–6.
Rochon, A., de Vernal, A., Turon, J. L., Matthiessen, J. & Head,M. J. 1999. Distribution of Recent Dinoflagellate Cysts in SurfaceSediments from the North Atlantic Ocean and Adjacent Seas inRelation to Sea-surface Parameters. Special Contribution Series35. American Association of Stratigraphic Palynologists Foun-dation, Dallas, Texas, United States, pp. 1–146.
Schiller, J. 1937. Dinoflagellatae (Peridineae). In Rabenhorst, L.[Ed.] Kryptogamen-Flora, vol. 10, 3(2). Akademische Verlagsge-sellschaft, Leipzig, 1–589.
Siano, R. & Montresor, M. 2005. Morphology, ultrastructure andfeeding behaviour of Protoperidinium vorax sp. nov. (Dinophy-ceae, Peridiniales). Eur. J. Phycol. 40:221–32.
Solomon, S., Mudie, P. J., Cranston, R., Hamilton, T., Thibau-deau, S. A. & Collins, E. S. 2000. Characterisation of marineand lacustrine sediments in a drowned thermokarst embay-ment, Richards Island, Beaufort Sea, Canada. Int. J. EarthSciences 89:503–21.
Stamatakis, A. 2006. RAxML-VI-HPC: maximum likelihood-basedphylogenetic analyses with thousands of taxa and mixedmodels. Bioinformatics 22:2688–90.
Swofford, D. L. 2002. PAUP*. Phylogenetic Analysis Using Parsimony(*and Other Methods). Version 4. Sinauer Associates, Sunder-land, Massachusetts.
Takano, Y. & Horiguchi, T. 2006. Acquiring scanning electronmicroscopical, light microscopical and multiple genesequence data from a single dinoflagellate cell. J. Phycol.42:251–6.
Thomson, P. G., Wright, S. W., Bolch, C. J. S., Nichols, P. D.,Skerratt, J. H. & McMinn, A. 2004. Antarctic distribution,pigment and lipid composition, and molecular identificationof the brine dinoflagellate Polarella glacialis (Dinophyceae).J. Phycol. 40:867–73.
V�asquez-Bedoya, L. F., Radi, T., Ruiz-Fern�andez, A. C., de Vernal,A., Machain-Castillo, M. L., Kielt, J. F. & Hillaire-Marcel, C.2008. Organic-walled dinoflagellate cysts and benthic forami-nifera in coastal sediments of the last century from the Gulfof Tehuantepec, South Pacific Coast of Mexico. Mar. Micro-paleontol. 68:49–65.
de Vernal, A., Henry, M., Matthiessen, J., Mudie, P. J., Rochon,A., Boessenkool, K. P., Eynaud, F. et al. 2001. Dinoflagel-late cyst assemblages as tracers of sea-surface conditions inthe northern North Atlantic, Arctic and sub-Arctic seas: thenew “n=677” data base and its application for quantitativepalaeoceanographic reconstruction. J. Quaternary Sci. 16:681–98.
Wall, D. & Dale, B. 1968. Modern dinoflagellate cysts and evolu-tion of the Peridiniales. Micropaleontology 14:265–304.
Williams, G. L., Fensome, R. A., Miller, M. A. & Sarjeant, W. A. S.2000. A Glossary of the Terminology Applied to Dinoflagellates,Acritarchs and Prasinophytes, with Emphasis on Fossils. 3rd ed.,Contributions Series number 37. American Association ofStratigraphic Palynologists Foundation, Texas, 370 pp.
Yamaguchi, A., Hoppenrath, M., Pospelova, V., Horiguchi, T. &Leander, B. S. 2011. Molecular phylogeny of the marinesand-dwelling dinoflagellate Herdmania litoralis and anemended description of the closely related planktonic genusArchaeperidinium J€orgensen. Eur. J. Phycol. 46:98–112.
Yamaguchi, A. & Horiguchi, T. 2005. Molecular phylogeneticstudy of the heterotrophic dinoflagellate genus Protoperidini-um (Dinophyceae) inferred from small subunit rRNA genesequences. Phycol. Res. 53:30–42.
Zingone, A. & Montresor, M. 1988. Protoperidinium parthenopessp. nov. (Dinophyceae), an intriguing dinoflagellate fromthe Gulf of Naples. Cryptogam. Algol. 9:117–25.
Zonneveld, K. A. F. 1997. New species of organic walled dinofla-gellate cysts from modern sediments of the Arabian Sea(Indian Ocean). Rev. Palaeobot. Palyno. 97:319–37.
Zonneveld, K. A. & Dale, B. 1994. The cyst-motile stage relation-ships of Protoperidinium monospinum (Paulsen) Zonneveld etDale comb. nov. and Gonyaulax verior (Dinophyta, Dino-phyceae) from the Oslo Fjord (Norway). Phycologia 33:359–68.
866 �ERIC POTVIN ET AL.