a revised paleocene (teurian) dinoflagellate cyst zonation from eastern new zealand
TRANSCRIPT

This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/authorsrights

Author's personal copy
Research paper
A revised Paleocene (Teurian) dinoflagellate cyst zonation fromeastern New Zealand
Erica M. Crouch a,⁎, Pi Suhr Willumsen b,c, Denise K. Kulhanek a,d, Samantha J. Gibbs e
a Department of Paleontology, GNS Science, P.O. Box 30368, Lower Hutt 5040, New Zealandb MUSERUM, Natural History Division, Havnevej 14, 7800 Skive, Denmarkc Department of Geoscience, Aarhus University, Høegh-Guldbergs Gade 2, DK-8000 Aarhus C, Denmarkd Integrated Ocean Drilling Program, Texas A&M University, 1000 Discovery Drive, College Station, TX 77845-9547, USAe School of Ocean and Earth Sciences, National Oceanography Centre, University of Southampton, Southampton SO14 3ZH, UK
a b s t r a c ta r t i c l e i n f o
Article history:Received 12 November 2012Received in revised form 12 December 2013Accepted 20 December 2013Available online 11 January 2014
Keywords:dinoflagellate cystPaleoceneTeurianNew Zealandzonationsystematics
Organic-walled dinoflagellate cyst (dinocyst) assemblages are documented from Paleocene (New ZealandTeurian Stage) sediments in five sections from eastern New Zealand: Tawanui, Angora Road and Toi Flat-1 core in the East Coast Basin, mid-Waipara River in the Canterbury Basin, and ODP Site 1121 on the easternmargin of Campbell Plateau. Based on dinocyst results from these sections, along with published earliest Paleo-cene records from the East Coast, Canterbury and Great South Basins, a revised Paleocene (Teurian) dinocyst zo-nation is proposed. The zones are labelled as NZDP — New Zealand Dinocyst Paleocene — and are all intervalzones. The eight zones, NZDP1 to NZDP8, encompass the entire Paleocene, from the Cretaceous–Paleogeneboundary at 66.04 Ma to the Paleocene–Eocene boundary at 55.96 Ma. Correlation of the NZDP zones with theInternational and New Zealand Time Scales is provided, and is primarily based on correlation with calcareousnannofossil biostratigraphy. Three new dinocyst species are described: Leptodinium? pustulatum sp. nov.,Cerodinium angulatum sp. nov., and Vozzhennikovia tawanuiensis sp. nov.
© 2013 Elsevier B.V. All rights reserved.
1. Introduction
The Paleocene Epoch (66.04 to 55.96 Ma), or New ZealandTeurian Stage (Cooper, 2004), represents a globally significant tran-sition in climate systems and carbon cycles from cooler climates ofthe Late Cretaceous into the greenhouse world of the Early Eocene(e.g., Zachos et al., 2001, 2008; Westerhold et al., 2011; Hollis et al.,2012). In the New Zealand region, the Paleocene was a time of tectonicquiescence with the New Zealand microcontinent isolated fromAustralia and Antarctica (Fig. 1), and extensive passive margin trans-gressive sediment deposition along the margins of the microcontinent(Ballance, 1993; King et al., 1999; Sutherland and King, 2008).
Although foraminifera and calcareous nannofossils are the prima-rymicrofossil groups that underpin the New Zealand Paleogene TimeScale (Jenkins, 1966; Edwards, 1971; Jenkins, 1971; Hornibrooket al., 1989; Morgans et al., 2004), calcareous microfossils have apatchy distribution in Paleocene sediments. This is due to sedimentsoften having low levels of carbonate or being non-calcareous, poorpreservation of calcareous microfossil assemblages, and a lack oflow-latitude calcareous biostratigraphic marker taxa being recordedin New Zealand Paleocene sediments (Hornibrook and Harrington,1957; Hornibrook and Edwards, 1971; Moore, 1989; Morgans et al.,
2004). In contrast, diverse and well-preserved organic-walled dino-flagellate cyst (dinocyst) assemblages are found in almost all marinesettings of Paleocene age, fromnearshore to deepmarine environments(Strong et al., 1995; Crouch and Brinkhuis, 2005;Willumsen and Vajda,2010). Numerous dinocyst studies have been carried out in NewZealand, with long-established Jurassic to Eocene zonations (Wilson,1984, 1987, 1988), and more recent refinements of Late Cretaceousand Eocene zones (Schiøler and Wilson, 1998; Roncaglia et al., 1999;Willumsen, 2003, 2011; Morgans et al., 2004; Clowes, 2009).
The New Zealand Teurian Stage is currently encompassed bythree formal dinocyst interval zones (Fig. 2): the Trithyrodiniumevittii Zone, Palaeocystodinium golzowense Zone, and the lowermostpart of the Apectodinium homomorphum Zone (Wilson, 1984, 1987,1988; Cooper, 2004). There is scope, however, to establish a more de-tailed Paleocene dinocyst zonation (Wilson, 1988; Beggs et al., 1992;Willumsen, 2011), as already exist in other regions such as Australia(Partridge, 2001, 2004) and Northwest Europe (Heilmann-Clausen,1985; Powell, 1988; Mudge and Bujak, 1996). With an increasingfocus on the paleoclimatic importance of the Paleocene Epoch (e.g.,Petrizzo, 2005; Bernaola et al., 2007; Hollis et al., 2012) and petroleumpotential of Paleocene sediments in New Zealand (e.g., Moore et al.,1987; Killops et al., 2000; Schiøler and Roncaglia, 2007; Schiøler et al.,2010), it is timely to review Teurian dinocyst assemblages with theaim of establishing a high-resolution zonation.
In this paper, we examine dinocyst assemblages from Paleocenesections in eastern New Zealand, establish a revised Paleocene dinocyst
Review of Palaeobotany and Palynology 202 (2014) 47–79
⁎ Corresponding author. Tel.: +64 4 5704810; fax: +64 4 5704600.E-mail address: [email protected] (E.M. Crouch).
0034-6667/$ – see front matter © 2013 Elsevier B.V. All rights reserved.http://dx.doi.org/10.1016/j.revpalbo.2013.12.004
Contents lists available at ScienceDirect
Review of Palaeobotany and Palynology
j ourna l homepage: www.e lsev ie r .com/ locate / revpa lbo

Author's personal copy
zonation, and correlate this scheme to calcareous nannofossil zones andthe International Geological Time Scale of Gradstein et al. (2012). Weadopt the formal subdivisions of the Paleocene Epoch/Series establishedby Schmitz et al. (2011) and Gradstein et al. (2012): Early/Lower(Danian), Middle (Selandian), and Late/Upper (Thanetian).
2. Previous dinocyst work
Initial studies of dinocysts in New Zealand were completed by G.J.Wilson, who has published numerous studies incorporating biostra-tigraphy and taxonomy from Jurassic to Eocene sections (Wilson, 1967,1978, 1982, 1984, 1987, 1988; Strong et al., 1995;Wilson andMcMillan,1996; Schiøler and Wilson, 1998). Wilson (1984) established a NewZealand Late Jurassic to Eocene dinocyst zonation, and provided anamended Paleocene to Middle Eocene zonation based on records fromthe Waipawa section, southern Hawkes Bay (Wilson, 1988) and mid-Waipara River section, North Canterbury (Wilson, 1987; Fig. 1).
High-resolution documentation of dinocyst assemblages across theCretaceous–Paleogene (K–Pg) boundary (66.04 Ma) and Paleocene–Eocene (P–E) boundary (55.96 Ma) has recently been carried out onsections from eastern New Zealand. A detailed sequence of dinocystbioevents and acme intervals across the K–Pg boundary (Willumsen,2000, 2003, 2004, 2011) are recognised fromMead and Branch Streams,Clarence Valley, East Coast Basin (Fig. 1), and discussed in relation tothe existing Paleocene dinocyst zonation (Wilson, 1988) and inte-grated with foraminiferal datums and the South Pacific radiolarianzonation (Hollis, 1993, 2002). In addition, dinocyst records fromthe K–Pg boundary are detailed from mid-Waipara and Grey Rivers,Canterbury Basin, and Fairfield Quarry, Great South Basin (Fig. 1;Willumsen, 2003, 2006; Willumsen and Vajda, 2010; Ferrow et al.,2011; Willumsen, 2012).
Dinocyst zones have been established for the earliest Eocene(Crouch, 2001; Morgans et al., 2004), based mainly on the Tawanuisection, East Coast Basin (Fig. 1). The P–E boundary, or Paleocene–Eocene Thermal Maximum (PETM), is marked by the onset of a glob-ally recognised negative carbon isotope excursion (CIE) and benthicforaminiferal extinction event (e.g., Zachos et al., 2001; McInerney and
Wing, 2011). In New Zealand sections, the onset of the PETM is also coin-cident with the beginning of the lowest common occurrence (N~40%) ofthe Apectodinium genus (Crouch et al., 2001). This Apectodinium acmehas been recorded within the PETM worldwide (Bujak andBrinkhuis, 1998; Sluijs et al., 2007, 2008, 2011; Bijl et al., 2013).
3. Stratigraphy
The Paleocene sections examined in this study are located in theEast Coast and Canterbury sedimentary basins, and on the easternflank of Campbell Plateau (Fig. 1). Paleocene sediments in easternNew Zealand are part of an overall fining-upward transgressive se-quence that began in the latest Cretaceous and continued through tothe Eocene (Ballance, 1993; King et al., 1999).
In the East Coast Basin, samples were collected from the Tawanuiand Angora Road sections and Toi Flat-1 core, southern Hawkes Bay(Fig. 1). Early and Middle Paleocene sediments of the East Coast Basinare dominated by fine-grained, non-calcareous to calcareous siliceousmudstone of the Whangai Formation (Moore, 1988). This formation istypically overlain by the Waipawa Formation, a distinctive dark mud-stone found throughout the East Coast Basin and other New Zealandsedimentary basins (Moore, 1989; Killops et al., 2000; Schiøler et al.,2010). The Waipawa Formation is a poorly-bedded, bioturbated, darkmudstone (Moore, 1989), with distinctive geochemical markers suchas high total organic carbon (TOC), isotopically heavy δ13CTOC and abun-dant 24-n-propylcholestanes (Killops et al., 2000), that is dated lateMiddle to early Late Paleocene (calcareous nannofossil Zone NP6 tolower NP7) (Hollis et al., 2012). To the south, in Canterbury and GreatSouth Basins, the coeval facies-equivalent to the Waipawa Formationis the Tartan Formation (Cook et al., 1999; Schiøler and Roncaglia,2007; Schiøler et al., 2010). Above the Waipawa Formation in the EastCoast Basin, sediments comprise poorly-bedded, calcareous, smectite-rich mudstone of the Wanstead Formation (Moore and Morgans,1987). In some East Coast sections, the base of the Paleocene coincideswith the base of a distinctive Whangai Formation member, the glauco-nitic Te Uri Member. At Tawanui, the upper part of the Te Uri Memberis correlated with the Waipawa Formation, which is in turn overlain
160° E 170° E 180° 170° W
50° S
40° S
30° S
ODP Site 1121
NCampbell Is.
Chatham Is.
NorthlandBasin
TaranakiBasin
2000 m
Campbell Plateau
ChallengerPlateau
LordHoweRise
Chatham Rise
Hikurangi Plateau
MBFQ
MH
mWRKC GR
Md, Bh
Wp
AgTwTF
Present-day locationof Paleocene sections
EastCoastBasin
GreatSouthBasin
CanterburyBasin
Wellington
Christchurch
AB
CanterburyBasin
CanterburyBasin
TaranakiBasin
TaranakiBasin
West CoastBasin
West CoastBasin
Lord Howe RiseLord Howe Rise
EastTasman Plateau
EastTasman Plateau
Norfolk RidgeNorfolk Ridge
CampbellPlateau
CampbellPlateau
HikurangiPlateau
HikurangiPlateau
East CoastBasin
East CoastBasin
60°S
50°S
170 180 -160-170 -150 -140
Chatham RiseChatham Rise
Great SouthBasin
Great SouthBasin
KeyNon-deposition(probable land)
Non-marine
Shelf (neritic)
Bathyal
Abyssal
11211121
11721172
mWRmWR
TF, Tw,A gTF, Tw,A g
Md, BhMd, Bh
Fig. 1. Location of New Zealand sections, ODP Site 1121 and other localities mentioned in the text, along with sedimentary basins around New Zealand (A), and (B) paleogeographic re-construction of the New Zealand region ~60 Ma. Tw = Tawanui, TF = Toi Flat-1 core, Ag = Angora Road, Wp = Waipawa, Md = Mead Stream, Bh = Branch Stream, mWR = mid-Waipara River, GR = Grey River, KC = Kumara-2 core, MH = Moeraki–Hampden, MB = Measly Beach core, FQ = Fairfield Quarry.After King et al. (1999) and modified from Hollis et al. (2009).
48 E.M. Crouch et al. / Review of Palaeobotany and Palynology 202 (2014) 47–79

Author's personal copy
by the Wanstead Formation (Rogers et al., 2001). In other East Coastsections, the Waipawa Formation is absent and Paleocene sequencesconsist of only two formations — the Whangai and WansteadFormations.
The Paleocene sections sampled in the Canterbury Basin, at mid-Waipara River (Fig. 1), comprise the following units. The UpperCretaceous to Lower Paleocene is dominated by brownish-grey finesandstone of the Conway Formation (Browne and Field, 1985). TheK–Pg boundary is recorded in the uppermost part of the Conway For-mation (Hollis and Strong, 2003; Vajda and Raine, 2003; Ferrowet al., 2011; Willumsen, 2012). The Conway Formation is conformablyoverlain by the Loburn Formation (Lower to Middle Paleocene), abrown to grey sandy non-calcareous to calcareous mudstone (Browneand Field, 1985; Morgans et al., 2005). Conformably overlying theLoburn Formation is the Waipara Greensand (Middle and Late Paleo-cene), a green-grey bioturbated medium to fine sandstone (Browneand Field, 1985). While the basal part of the Waipara Greensand is cal-careous, most of the unit is non-calcareous. A recent study has correlat-ed the upper part of theWaipara Greensandwith the Tartan Formation(Hollis et al., 2012). The Waipara Greensand is overlain by the AshleyMudstone, a soft grey siltstone (Browne and Field, 1985).
Paleocene sediments at ODP Site 1121, on the eastern flanks of theCampbell Plateau (Fig. 1), consist primarily of lower bathyal diatomand nannofossil ooze (Carter et al., 1999).
4. Material and methods
4.1. Sections examined
Dinocyst assemblages from mid-Waipara River, Tawanui, Toi Flat-1core, Angora Road and ODP Site 1121 (Fig. 1) form the basis of thenew Paleocene dinocyst zonation, and more detail of these sections isgiven below. Calcareous nannofossil assemblages have also been exam-ined from selected samples in these sections.
Dinocyst results from other sections provide supplementary bio-stratigraphic data; however, these sections are not explained in de-tail here. Primarily, the dinocyst zones established in the lower partof the Early Paleocene utilise dinocyst biostratigraphy from Branchand Mead Stream, East Coast Basin; mid-Waipara River and GreyRiver, Canterbury Basin; and Fairfield Quarry, Great South Basin(Fig. 1; Willumsen, 2003, 2004, 2006; Willumsen and Vajda, 2010;Ferrow et al., 2011; Willumsen, 2011, 2012).
4.1.1. Mid-Waipara River sectionThe mid-Waipara River section is located ~13 km west of the
Waipara township, northern Canterbury (Fig. 1), and includes thearea downstream from Doctors Gorge to the top of the AmuriGroup in the ‘lower gorge’ (grid reference NZMS 260-M34/755 946to M34/789 944). Paleocene sediments are well-exposed along theriver bank, and a series of low-dipping sections have been logged,sampled and integrated into a single composite section (Morganset al., 2005). This study is based on two sub-sections, referred to asColumns 2 and 4.
From Column 2, ~83 m of section, from 49.44 m to 132.53 m, wassampled (Figs. 3 and 4). A total of 24 samples have been examined,five from the Loburn Formation and 19 from the Waipara Greensandand very basal Ashley Mudstone (Supplementary Data, Appendix B).From Column 4, ~116 m of section, from 0.74 m to 117 m, was ex-amined (Figs. 5 and 6), with five samples analysed from the ConwayFormation, seven samples from the Loburn Formation and eight sam-ples from the Waipara Greensand (Supplementary Data, AppendixB). The contacts between the Conway and Loburn Formations andWaipara Greensand are visible (Morgans et al., 2005).
In Column 2, an unconformity is recorded in the uppermostWaipara Greensand, at ~131 m, in the upper part of the interval cor-related with the Tartan Formation (Hollis et al., 2012). Based on bio-stratigraphy and geochemistry, primarily δ13CTOC, TOC (wt.%) andTEX86, the unconformity appears to span from early Late Paleoceneto close to the P–E boundary (Hollis et al., 2012).We assume that this un-conformity, with a similar temporal duration, is present in Column 4.Magnetostratigraphy from Column 2 correlates the middle part of theWaipara Greensand, from ~65 to 80 m, with Chron C26r in the MiddlePaleocene (Lurcock, 2011; Hollis et al., 2012). The position of the overly-ing Chron C26n or underlying C27n boundaries are not recognised(Lurcock, 2011).
Dinocyst data from the K–Pg boundary (Willumsen, 2003, 2006;Willumsen and Vajda, 2010; Ferrow et al., 2011) in a sub-section(Column 1; Morgans et al., 2005), upstream from Columns 2 and 4,are utilised in the lowermost Paleocene dinocyst zones establishedhere. There is a small temporal gap between the Paleocene sectionstudied by Willumsen and the lowermost interval examined in thisstudy (Column4). The uppermost sample in the K–Pg boundary section,Column 1, contains superabundant Trithyrodinium evittii (Willumsen,2003), while T. evittii is rare in the basal sample examined in Column4 and the highest occurrence is seen in the sample stratigraphicallyabove (Figs. 5 and 6).
4.1.2. Tawanui sectionThe Tawanui section is located on the upper Akitio River (grid ref-
erence NZMS 260-U24/967 863), on the southeastern margin of theWhangai Range, southern Hawkes Bay (Fig. 1). Twenty samples
T rith
yrod
iniu
mev
ittii
Zon
e
Din
ocys
t zon
e(W
ilson
198
7,19
88)
Pal
aeoc
ysto
dini
um g
olzo
wen
seZ
one
Ape
ctod
iniu
mho
mom
orph
umZ
one
Ma
55
56
57
58
59
60
61
62
63
64
65
66
C29
C28
C27
C26
C25
C24
Late Cretaceous
Eoc
ene
Ear
ly P
aleo
cene
Late
Pal
eoce
ne
Dan
ian
Sel
andi
anT
hane
tian
Ypr
esia
n
low
erTe
uria
nup
per
Teur
ian
Wai
paw
an
P0Pa
P1
P2
P3
P4
P5
P6
CC26
NP1
NP2
NP3
NP4
NP5
NP6
NP7
NP8
NP9
RP1
RP2
RP3
RP4
RP5
RP6
RP7
RP8NP10
Chr
on
Pol
arity
Epo
ch
Sta
ge
NZ
Sta
ge
For
am Z
one
Mid
Pal
eoce
nedl
e
Nan
nopl
ank.
Zon
eR
adio
laria
nZ
one
Fig. 2. The Paleocene Epoch and New Zealand Teurian Stage.The International Time Scale is from Gradstein et al. (2012) and New Zealand Stages fromRaine et al. (2013). Calcareous nannofossil (NP), foraminifera (P), and radiolarian (RP) zonesfrom Gradstein et al. (2012). The Paleocene (Teurian) dinocyst zonation of Wilson (1987,1988) is shown.
49E.M. Crouch et al. / Review of Palaeobotany and Palynology 202 (2014) 47–79

Author's personal copy
have been examined from the Te Uri Member and the overlyingWanstead Formation (Supplementary Data, Appendix B). Approxi-mately 8.5 m of Te Uri Member was sampled, with the upper ~3 mcomprising a bioturbated, sandy greensand that is correlated withthe Waipawa Formation (Morgans, 2001; Rogers et al., 2001). Thecontact between the Te Uri Member and Wanstead Formation is un-conformable. The Upper Paleocene part of the Wanstead Formation,~80 m of section, is sampled (Figs. 7 and 8).
Previous dinocyst assemblage data from across the P–E boundary atTawanui, with a detailed record from the PETM (Crouch, 2001; Crouchet al., 2001; Crouch and Brinkhuis, 2005), are used to define the upper-most Paleocene dinocyst zone established here.
4.1.3. Toi Flat-1 coreToi Flat-1 stratigraphic core was drilled to a total depth of 55.37 m
(Laing, 1991) and is located on the flank of the Toi Flat anticline (gridreference NZMS 260-U24/905 899), in southern Hawkes Bay (Fig. 1).Fourteen samples (Supplementary Data, Appendix B) have been exam-ined from mudstone of the Wanstead Formation, and seven samplesfrom green glauconitic silty sandstone of the Te Uri Member (~39 mto 48.5 m). Elevated TOC values and isotopically heavy organic δ13Cvalues suggest part of the upper portion of the glauconitic interval canbe correlated with the Waipawa Formation (Fig. 9). The P–E boundaryis tentatively placed between 19.37 m and 16.37 m, based on thedinocyst assemblage and organic δ13C record (Fig. 9). Samples havenot been examined from the grey siltstone (undifferentiated WhangaiFormation) below the Te Uri Member.
4.1.4. Angora RoadThe Angora Road section is located ~2 km west of the Wimbledon
township (grid reference NZMS 260-U24/050 780), in southernHawkesBay (Fig. 1). Approximately 90 m of Paleocene sediment was logged(Tayler, 2011), consisting of ~60 m of calcareous mudstone, the UpperCalcareous Member of the Whangai Formation, which is conformablyoverlain by ~33 m of non-calcareous mudstone with generally highTOC that is identified as Waipawa Formation (Figs. 11 and 12).The Waipawa Formation boundary is placed at the position wherelithologies change from dominantly calcareous to dominantlynon-calcareous (Tayler, 2011). For this study, four samples from theUpper Calcareous Member and five samples from theWaipawa Forma-tion were examined for dinocyst assemblages (Supplementary Data,Appendix B).
4.1.5. ODP Site 1121Paleocene sediments, consisting primarily of lower bathyal dia-
tom and nannofossil ooze (Carter et al., 1999), have been recoveredfrom ODP Site 1121 (Fig. 1). Radiolarian (Hollis, 2002) and calcareousnannofossil biostratigraphy, and magnetostratigraphic analyses (Weiet al., 2005), have been completed. Palynomorph assemblages are pre-served in part of the Paleocene section, from ~110 m to 57 m, andone sample at 134.4 m also contains palynomorphs (SupplementaryData, Appendix B). Fourteen samples from Middle Paleocene to earlyLate Paleocene have been examined, and are correlatedwith calcareousnannofossil Zones NP4 to NP6/7 (Fig. 13). A recent study has identifiedan interval in the Late Paleocene, within Zone NP6/7, that may be an
Wai
para
G
reen
sand
Lobu
rn F
m
C26
r
Ppy
roph
orum
alae
oper
idin
ium
Van
gula
taoz
zhen
niko
via
I. ci
ngul
atum
Dfo
veol
ata
efla
ndre
a
Tde
licat
aha
lass
ipho
ra
E. r
etic
ulat
agr
oup
E. m
arga
rita
grou
p
Lept
odin
ium
?pu
stul
atum
sp. n
ov.
Mro
tund
aan
umie
lla
Ape
ctod
iniu
mge
nus
Voz
zhen
niko
via
taw
anui
ensi
s sp
. nov
.
Ape
ntar
adia
tum
lterb
idin
ium
120
130
100
110
90
80
70
?
?
?
50
60
40
Mid-Waipara River, Column 2
Sin
orna
taen
onia
spha
eraUpp
er N
P4
Chr
on
Hei
ght(
m)
For
mat
ion
NZ
one
PLo
wer
NP
4N
P3
?
Dinoflagellate cysts
Tow
eius
per
tusu
s
Din
ocys
t zon
e
nannos stop
Crib
rope
ridin
ium
wet
zelii
Hys
tric
hoko
lpom
age
nus
Rot
tnes
tia b
orru
sica
NZ
DP
3N
ZD
P4
NZ
DP
5N
ZD
P6
NZ
DP
7N
ZD
P8
Cha
isso
litus
mol
ithus
Calcareousnannofossils
N. p
erfe
ctus
Lith
olog
y
Fig. 3. Stratigraphic distribution of selected dinocysts and calcareous nannofossils from Column 2, mid-Waipara River (Morgans et al., 2005). Summary lithological column and samplepoints shown. Calcareous nannofossil (NP) zones and magnetostratigraphy discussed in text. Dashed line marks the proposed New Zealand Dinocyst Paleocene (NZDP) zones. Wavyline at ~131 m denotes an unconformity. Black box shows the interval correlated with the Tartan Formation (Hollis et al., 2012).
50 E.M. Crouch et al. / Review of Palaeobotany and Palynology 202 (2014) 47–79

Author's personal copy
Alterb
idiniu
m p
enta
radia
tum
Apecto
dinium
spp.
Cerod
inium
med
calfii
Cerod
inium
spp.
(par
s.)
Deflan
drea
deli
neat
atyp
e
Deflan
drea
fove
olata
20 40
Isabe
lidini
umsp
p.
Lejeu
necy
sta/P
helod
inium
Man
umiel
la ro
tund
a
Palaeo
cysto
dinium
golz
owen
se
Palaeo
perid
inium
pyr
opho
rum
20 40 60
Seneg
aliniu
msp
p.
20
Spinidi
nium
spp.
20 2040 40 60 80
Vozzh
ennik
ovia
angu
lata
V. tawan
uiens
issp
. nov
.
Mid-Waipara River, Column 2
Din
ocys
t zon
eN
ZD
P3
NZ
DP
4N
ZD
P5
NZ
DP
6N
ZD
P7
NZ
DP
8
Total peridinioddinocystsAch
omos
phae
ra cr
assip
llise
Eisena
ckia
spp.
Cordo
spha
eridi
umsp
p.
Lept
odini
um?
pustu
latum
sp. n
ov.
20
Fibroc
ysta
Cordo
spha
eridi
um
-
type
20
Fibroc
ysta
Turb
iosph
aera
-
type
Fibroc
ysta
spp.
(par
s.)
Glaphp
yroc
ysta
spp.
(par
s.)
20% 40
Glaphy
rocy
sta re
tiinte
xta
Glaphy
rocy
stagr
oup
Areoli
gera
seno
nens
is
Hystri
chok
olpom
asp
p.
Hystri
chos
phae
ridium
tubif
erum
Impa
gidini
umsp
p.
Lingu
lodini
um m
acha
erop
horu
m
20 40
Elytro
cysta
sp. B
20
Elytro
cysta
sp. C
Corru
dinium
incom
posit
um
cf.
20 40
Operc
ulodin
iumsp
p.
Cereb
rocy
stasp
p.
Senon
iasph
aera
inor
nata
20 40 60
Spinife
rites
spp.
Cleisto
spha
eridi
um
um
placa
cant
h
Thalas
sipho
ra d
elica
ta
Cribro
perid
inium
wet
zelii
Wai
para
G
nsd
Lobu
rn F
m
120
130
100
110
90
80
70
50 645
646
647
649650
651
652
653
655
656
657
658
660662664
667
669
678
665
60
40 gonyaulacoid dinocysts peridinioid dinocysts
C26
r
?
?
?
Upp
er N
P4
Chr
on
NZ
one
PLo
wer
NP
4N
P3
?
Hei
ght(
m)
For
mat
ion
Sam
ple
(M34
/f)
Lith
olog
y
Fig. 4. Abundance record of selected dinocyst species and genera, and total peridinioid dinocysts, from Column 2, mid-Waipara River (Morgans et al., 2005). Summary lithological column and sample points shown. Calcareous nannofossil (NP) zonesandmagnetostratigraphy discussed in text. Dashed linemarks the proposed New Zealand Dinocyst Paleocene (NZDP) zones. Wavy line at ~131 m denotes an unconformity. Black box shows the interval correlated with the Tartan Formation (Holliset al., 2012).
51E.M
.Crouchetal./Review
ofPalaeobotanyand
Palynology202
(2014)47
–79
Author's personal copy
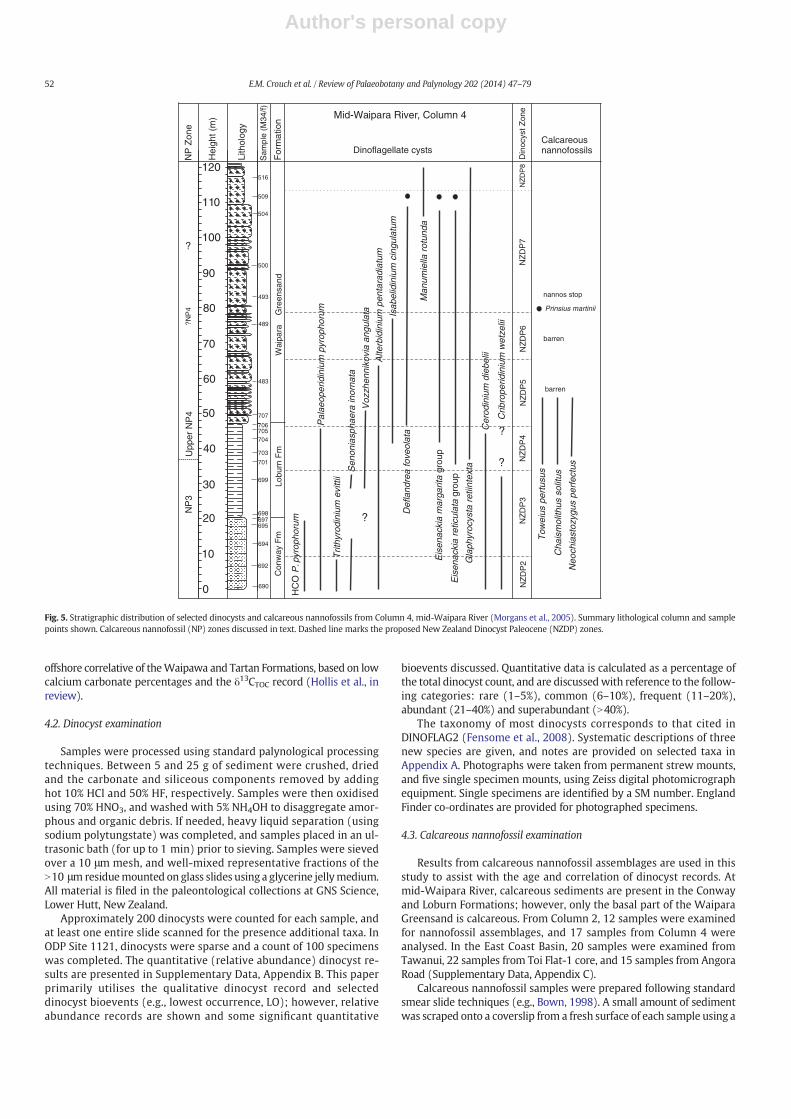
offshore correlative of theWaipawa and Tartan Formations, based on lowcalcium carbonate percentages and the δ13CTOC record (Hollis et al., inreview).
4.2. Dinocyst examination
Samples were processed using standard palynological processingtechniques. Between 5 and 25 g of sediment were crushed, driedand the carbonate and siliceous components removed by addinghot 10% HCl and 50% HF, respectively. Samples were then oxidisedusing 70% HNO3, and washed with 5% NH4OH to disaggregate amor-phous and organic debris. If needed, heavy liquid separation (usingsodium polytungstate) was completed, and samples placed in an ul-trasonic bath (for up to 1 min) prior to sieving. Samples were sievedover a 10 μm mesh, and well-mixed representative fractions of theN10 μm residuemounted on glass slides using a glycerine jellymedium.All material is filed in the paleontological collections at GNS Science,Lower Hutt, New Zealand.
Approximately 200 dinocysts were counted for each sample, andat least one entire slide scanned for the presence additional taxa. InODP Site 1121, dinocysts were sparse and a count of 100 specimenswas completed. The quantitative (relative abundance) dinocyst re-sults are presented in Supplementary Data, Appendix B. This paperprimarily utilises the qualitative dinocyst record and selecteddinocyst bioevents (e.g., lowest occurrence, LO); however, relativeabundance records are shown and some significant quantitative
bioevents discussed. Quantitative data is calculated as a percentage ofthe total dinocyst count, and are discussedwith reference to the follow-ing categories: rare (1–5%), common (6–10%), frequent (11–20%),abundant (21–40%) and superabundant (N40%).
The taxonomy of most dinocysts corresponds to that cited inDINOFLAG2 (Fensome et al., 2008). Systematic descriptions of threenew species are given, and notes are provided on selected taxa inAppendix A. Photographs were taken from permanent strew mounts,and five single specimen mounts, using Zeiss digital photomicrographequipment. Single specimens are identified by a SM number. EnglandFinder co-ordinates are provided for photographed specimens.
4.3. Calcareous nannofossil examination
Results from calcareous nannofossil assemblages are used in thisstudy to assist with the age and correlation of dinocyst records. Atmid-Waipara River, calcareous sediments are present in the Conwayand Loburn Formations; however, only the basal part of the WaiparaGreensand is calcareous. From Column 2, 12 samples were examinedfor nannofossil assemblages, and 17 samples from Column 4 wereanalysed. In the East Coast Basin, 20 samples were examined fromTawanui, 22 samples from Toi Flat-1 core, and 15 samples from AngoraRoad (Supplementary Data, Appendix C).
Calcareous nannofossil samples were prepared following standardsmear slide techniques (e.g., Bown, 1998). A small amount of sedimentwas scraped onto a coverslip from a fresh surface of each sample using a
Wai
para
G
reen
sand
Lobu
rn F
mC
onw
ay F
m
Mid-Waipara River, Column 4
120
100
110
90
80
70
50
40
30
20
10
0
60
nannos stop
barren
barren
Prinsius martinii
483
489
493
500
504
509
516
707
690
692
694
695697698
699
701
703
704705706 P
pyro
phor
umal
aeop
erid
iniu
m
Tev
ittii
rithy
rodi
nium
Sin
orna
taen
onia
spha
era
Cer
odin
ium
die
belii
Crib
rope
ridin
ium
wet
zelii
HC
OP
. pyr
opho
rum
Van
gula
taoz
zhen
niko
via
Ape
ntar
adia
tum
lterb
idin
ium I
cing
ulat
umsa
belid
iniu
m
Dfo
veol
ata
efla
ndre
a
Mro
tund
aan
umie
lla
Cha
isso
litus
mol
ithus
Tow
eius
per
tusu
s
?NP
4
Eis
enac
kia
mar
garit
a gr
oup
Ere
ticul
ata
isen
acki
agr
oup
Gla
phyr
ocys
ta r
etiin
text
a
?
?
?
NZ
DP
2N
ZD
P3
NZ
DP
4N
ZD
P5
NZ
DP
6N
ZD
P7
NZ
DP
8
Upp
er N
P4
NP
Zon
e
Hei
ght (
m)
Sam
ple
(M34
/f)
NP
3
?
For
mat
ion
Dinoflagellate cysts
Din
ocys
t Zon
e
Calcareousnannofossils
Npe
rfec
tus
eoch
iast
ozyg
us
Lith
olog
y
Fig. 5. Stratigraphic distribution of selected dinocysts and calcareous nannofossils from Column 4, mid-Waipara River (Morgans et al., 2005). Summary lithological column and samplepoints shown. Calcareous nannofossil (NP) zones discussed in text. Dashed line marks the proposed New Zealand Dinocyst Paleocene (NZDP) zones.
52 E.M. Crouch et al. / Review of Palaeobotany and Palynology 202 (2014) 47–79

Author's personal copy
Wai
para
G
reen
sand
Lobu
rn F
mC
onw
ayF
m
120
100
110
90
80
70
50
40
30
20
10
0
60 483
489
493
500
504
509
516
707
690
692
694
695697698
699
701
703
704705706
?NP
4U
pper
NP
4N
Zon
eP
Hei
ght (
m)
Sam
ple
(M34
/f)
NP
3
?
For
mat
ion
20
Cordo
spha
eridi
um fib
rosp
inosu
m
Lept
odini
um?
pustu
latum
sp. n
ov.
Cribro
perid
inium
wet
zelii
Spong
odini
um
deliti
ense
cf.
Eisena
ckia
spp.
Fibroc
ysta
spp.
(par
s.)
Fibroc
ysta
Tur
biosp
haer
a
-
type
Fibroc
ysta
bipo
laris
20 40 60 80
Glaphy
rocy
stagr
oup
20 40
Glaphy
rocy
sta re
tiinte
xta
Areoli
gera
seno
nens
is
Corru
dinium
incom
posit
um
cf.
Hystri
chos
phae
ridium
tubif
erum
Impa
gidini
umsp
p.
Kallos
phae
ridium
sp.
20
Elytro
cysta
sp. B
20
Elytro
cysta
sp. C
20
Operc
ulodin
iumsp
p.
Cereb
rocy
stasp
p.
Senon
iasph
aera
inor
nata
20 40
Spinife
rites
spp.
Cleisto
spha
eridi
um p
lacac
anth
um
Thal ss
phor
a de
licat
a
ai
gonyaulacoid dinocysts peridinioid dinocysts
Lith
olog
y
%
Din
ocys
t Zon
e
Alterb
idiniu
m p
enta
radia
tum
Cerod
inium
med
calfii
Cerod
inium
/Def
landr
ea
Deflan
drea
fove
olata
20
Isabe
lidini
umsp
p.
Lejeu
necy
sta/P
helod
inium
Man
umiel
la ro
tund
a
Palaeo
cysto
dinium
golz
owen
se
20 40
Palaeo
perid
inium
pyr
opho
rum
20 40 60
Seneg
aliniu
msp
p.
20 40
Spinidi
nium
spp.
Trithyr
odini
um e
vittii
20 40
Vozzh
ennik
ovia
angu
lata
Mid-Waipara River, Column 4
V. t
awan
uien
sis
sp. n
ov.
NZ
DP
2N
ZD
P3
NZ
DP
4N
ZD
P5
NZ
DP
6N
ZD
P7
NZ
DP
8
Fig. 6. Abundance record of selected dinocyst species and genera from Column 4, mid-Waipara River (Morgans et al., 2005). Summary lithological column and sample points shown. Calcareous nannofossil zones (NP) discussed in text. Dashed linemarks the proposed New Zealand Dinocyst Paleocene (NZDP) zones.
53E.M
.Crouchetal./Review
ofPalaeobotanyand
Palynology202
(2014)47
–79

Author's personal copy
razor blade. The sediment was mixed with a drop of water, spreadevenly over the coverslip and then dried on a hotplate. The coverslipwas affixed to a glass microscope slide using Norland Optical Adhe-sive 61 and cured under and ultraviolet light. Slides were examinedat magnifications of 1000× and 630× on a Leitz Ortholux II POL-BKand Olympus BX53 microscope using cross-polarised, phase-contrastand plain-transmitted light. Aminimum of four coverslip traverses (ap-proximately 800 fields of view (FOV))were observed for each slide, un-less no nannoplankton specimenswere found in the first two traverses.Presence/absence data were collected for Angora Road and mid-Waipara River, whereas counts to 400 specimens were conducted forToi Flat-1 core and Tawanui. In a few cases, where nannofossils wereparticularly rare, the number of specimens observed in 200 FOV(Tawanui) or 400 FOV (Toi Flat-1 core)was recorded. Results are corre-lated to the biostratigraphic zonation scheme of Martini (1971). Taxo-nomic concepts for species are those given in Perch-Nielsen (1985)and Bown (1998, 2005).
5. Biostratigraphy
The main dinocyst and calcareous nannofossil biostratigraphicresults from the five sections examined are discussed below. In general,samples yielded diverse and well-preserved dinocyst assemblages. Theoccurrence of calcareous nannofossils is variable, with Early Paleocenesamples often barren or containing fewer nannofossils than Middle to
Late Paleocene samples from the East Coast sections. In contrast,the Middle to Late Paleocene interval is barren in mid-WaiparaRiver. When present, nannofossil assemblages are generally moder-ately preserved, although there are some intervals of poorer preser-vation at Angora Road and Toi Flat-1 core.
5.1. Mid-Waipara River section
Dinocyst assemblages from Columns 2 and 4 are very similar (Figs. 3to 6), and combined results are discussed. Dinocysts are the dominantcomponent of palynomorph assemblages in the Conway and LoburnFormations, while spores/pollen become more abundant up-section inthe Waipara Greensand with the highest relative abundance, N80%,seen during the interval correlated with the Tartan Formation (Holliset al., in review). Peridinioid dinocysts are superabundant in the Con-way and Loburn Formations and the uppermost part of the WaiparaGreensand (Figs. 4 and 6).
The lowermost samples examined, in the Conway Formation,contain common to abundant peridinioid taxa of Palaeoperidiniumpyrophorum and various Spinidinium species, such as Spinidiniumdensispinatum (Fig. 6). The highest occurrence (HO) of Trithyrodiniumevittii is recorded at 6.32 m in Column 4 (Fig. 5). Spinidinium speciescontinue to be common to abundant in the Loburn Formation, alongwith Vozzhennikovia angulata (Figs. 4 and 6) that has a HO close to thecontact between the Loburn Formation and Waipara Greensand. In
50
60
70
80
90
100
110
120
130
140
Te
Uri
Mem
ber
931929
924925
922920
932
933934935936937
940
944
948
950
F?
952poorlyexposed
954
956957
958
959960
962
963964
966W
anst
ead
For
mat
ion
?
?
?
?NP6
NP
8N
P9
NZ
one
PN
P5
Dfo
veol
ata
efla
ndre
a
M. r
otun
da
F. t
ympa
nifo
rmis
Hel
iolit
hus
cant
abria
e Hel
iolit
hus
klei
npel
lii
Spe
nolit
hus
mor
iform
is
Dis
coas
ter
moh
eri
Dis
coas
ter
nobi
lis
Hel
iolit
hus
riede
lii
Dis
coas
ter
mul
tirad
iatu
s
T. d
elic
ata
V. t
awan
uien
sis
sp. n
ov.
C. a
ngul
atum
sp. n
ov.
L ?.
pust
ulat
umsp
. nov
.
Ere
ticul
ata
isen
acki
agr
oup
NZ
DP
7N
ZD
P8
For
mat
ion
Hei
ght (
m)
Lith
olog
y
Sam
ples
(U
24/f)
Dinoflagellate cysts Calcareous nannofossils
Tawanui Section
Din
ocys
t Zon
e
Fig. 7. Stratigraphic distribution of selected dinocysts and calcareous nannofossils from the Tawanui section. Summary lithological column (Morgans, 2001) and sample points shown.Calcareous nannofossil (NP) zones discussed in text. Dashed line marks the proposed New Zealand Dinocyst Paleocene (NZDP) zones. Wavy line at ~58 m denotes an unconformity.Black box shows the interval correlated with the Waipawa Formation (Rogers et al., 2001).
54 E.M. Crouch et al. / Review of Palaeobotany and Palynology 202 (2014) 47–79

Author's personal copy
50
60
70
80
90
100
110
120
130
140
Te
Uri
Mem
ber
931929
924922920
932
934
936
940
944
948
F?
952poorlyexposed
954
956
958
960
962
963964
966
Wan
stea
d F
orm
atio
n
NP
8N
P9
NZ
one
PN
P5
For
mat
ion
Hei
ght(
m)
Lith
olog
y
Sam
ples
(U
24/f)
NP6
Cordo
spha
eridi
umsp
p.
20%
L.?
pustu
latum
sp. n
ov.
Eisena
ckia
spp.
20
Elytro
cysta
sp. B
Fibroc
ysta
spp.
20
Glaphy
rocy
stasp
p.
H. tub
iferu
m
20
Impa
gidini
umsp
p.
L. m
acha
erop
horu
m
20
Operc
ulodin
iumsp
p.
20
Cereb
rocy
stasp
p.
20 40 60
Spinife
rites
spp.
Thalas
sipho
ra d
elica
taTawanuiSection
C. ang
ulatu
msp
. nov
.
Cerod
inium
spp.
(par
s.)
Deflan
drea
fove
olata
20
Man
umiel
la ro
tund
a
P. golz
owen
se
Phtha
nope
ridini
umsp
p.
20 40
Seneg
aliniu
msp
p.
20
Spinidi
nium
spp.
V. t
awan
uien
sis
sp. n
ov.
NZ
DP
8N
ZD
P7
Din
ocys
t Zon
e
gonyaulacoid dinocysts peridinioid dinocysts
Fig. 8. Abundance record of selected dinocyst species and genera from the Tawanui section. Summary lithological column (Morgans, 2001) and sample points shown. Calcareousnannofossil zones (NP) discussed in text. Dashed line marks the proposed New Zealand Dinocyst Paleocene (NZDP) zones. Wavy line at ~58 m denotes an unconformity. Black boxshows the interval correlated with the Waipawa Formation (Rogers et al., 2001).
10
0
30
20
50
40
Dep
th (
m)
Sam
ples
For
mat
ion
NZ
one
P
Din
ocys
t Zon
e
-30 -28 -26 -24 -22
Organic carbon isotope
TOC (%)0 0.4 0.8 1.2
NP5
Wha
ngai
For
mat
ion
NP9
?10NP
NP9
NP4
Isab
elid
iniu
m c
ingu
latu
m
Dfo
veol
ata
efla
ndre
a
Ere
ticul
ata
isen
acki
agr
oup
V. t
awan
uien
sis
sp. n
ov.
Lept
odin
ium
?pu
stul
atum
sp. n
ov.
Mro
tund
aan
umie
lla Ape
ctod
iniu
mge
nus
Hys
tric
hoko
lpom
age
nus
Ken
leyi
age
nus
Ape
ntar
adia
tum
lterb
idin
ium
Voz
zhen
niko
via
angu
lata
Eis
enac
kia
mar
garit
a gr
oup
Ape
ctod
iniu
mac
me
Dinoflagellate cysts
Toi Flat-1 core
NZDP5
NZDP/76
NZ
DP
8
Epo
chE
ocen
e
Wan
stea
d F
orm
atio
nT
e U
riM
embe
rPal
eoce
ne
F. t
ympa
nifo
rmis
Chi
asm
olith
us d
anic
us/b
iden
s
T. p
ertu
sus
Cam
plyo
spha
era
eo/d
ela
Eric
soni
a ro
bust
a
Dis
coas
ter
mul
tirad
iatu
s
Calcareousnannofossils
( )
Fig. 9. Stratigraphic distribution of selected dinocysts and calcareous nannofossils from the Toi Flat-1 core. Summary lithological column and sample points shown. Calcareous nannofossil(NP) zones discussed in text. Dashed linemarks the proposed New Zealand Dinocyst Paleocene (NZDP) zones. Black box shows the interval tentatively correlated with theWaipawa For-mation. TOC (%) and organic δ13C record provided.
55E.M. Crouch et al. / Review of Palaeobotany and Palynology 202 (2014) 47–79

Author's personal copy
Column 4, a notable peak (78%) in a Glaphyrocysta-group is recorded at36.19 m in the lower part of the Loburn Formation (Fig. 6). Within theLoburn Formation several lowest occurrences (LOs) are seen, includingIsabelidinium cingulatum andDeflandrea foveolata, alongwith theHOs ofP. pyrophorum and Senoniasphaera inornata (Figs. 3 and 5).
In the lowermost part of theWaipara Greensand, the most abundantperidinioid species is Isabelidinium cingulatum and the gonyaulacoidspecies Glaphyrocysta retiintexta is common to abundant. Othergonyaulacoid species, such as Spiniferites spp., Elytrocysta sp. B andOperculodinium spp., become common to abundant in the middle partof theWaipara Greensand, prior to another phase of abundant to super-abundant peridinioid taxa, primarily Senegalinium and Spinidiniumspecies, in the upper part of theWaipara Greensand (Figs. 4 and 6). Sev-eral important bioevents are recorded in theWaipara Greensand, includ-ing the HO of Alterbidinium pentaradiatum, I. cingulatum, Deflandreafoveolata, Eisenackia margarita group and Eisenackia reticulata group,along with the LO of Manumiella rotunda and Leptodinium? pustulatumsp. nov. (Figs. 3 and 5).
In Column 2, the dinocyst assemblage above the unconformity(~131 m) at the top of the Waipara Greensand is different to the un-derlying assemblage (Fig. 4). The assemblage is dominated bygonyaulacoid taxa, such as Operculodinium spp. and Spiniferitesspp., and the LO of the genera Apectodinium and Hystrichokolpomaare recorded. Carbon isotopes (δ13CTOC) and TEX86 analyses suggestthis interval can be correlated with the PETM (Hollis et al., 2012);however, it is notable that while the Apectodinium genus is common(~7%) no acme is recognised.
While calcareous nannofossil preservation was variable, age assess-ment was possible from Conway and Loburn Formations and basalWaipara Greensand. In the Waipara Greensand, samples were barrenof nannofossils above ~71 m in Column 2 and ~80 m in Column 4.The LO of Toweius pertusus occurs in the Loburn Formation in both Col-umns 2 and 4 (Figs. 3 and 5), and is used as a secondary bioevent tomark the base of Zone NP4, as the zonal marker Ellipsolithus macellushas a delayed first appearance in the New Zealand region (Edwards,1971). The interval below this is assigned to Zone NP3, and the NP2/NP3 zonal boundary was not recorded. The LO of Chiasmolithussolitus (=Chiasmolithus bidens), used here to subdivide Zone NP4,is recognised at ~70.41 m in Column 2, in the uppermost Loburn For-mation (Fig. 3). In Column 4, the LO of C. solitus is coeval with the LOof T. pertusus at ~36.19 m and the interval stratigraphically abovethis is assigned to upper NP4 zone (Fig. 5).
5.2. Tawanui section
In the lower part of the Te Uri Member, assemblages are domi-nated by gonyaulacoid dinocysts, particularly Cerebrocysta spp.,Operculodinium spp., and Spiniferites spp., while peridinioid dinocysts(mainly Senegalinium spp. and Spinidinium spp.) are superabundant inthe upper part of the Te Uri Member, correlated with theWaipawa For-mation (Fig. 8). In the overlying section, in theWanstead Formation, as-semblages are relatively uniform and gonyaulacoid dinocysts are moreabundant than peridinioid taxa. Important bioevents are recorded inthe upper part of the Te Uri Member, with the HO of Deflandreafoveolata and the LO of Manumiella rotunda (Fig. 7). In the lowermostWanstead Formation, the LOs of Vozzhennikovia tawanuiensis sp. nov.and Cerodinium angulatum sp. nov. are recorded.
Calcareous nannofossils are reasonably well preserved, and semi-quantitative counts to 400 specimens were possible on all but two ofthe 20 samples examined. The presence of Fasciculithus tympaniformisin the lowermost sample indicates the base of this section is withinZone NP5 (Fig. 7). The marker species for Zone NP6, Heliolithuskleinpellii, has a LO within the lower part of the Te Uri Member, andthe NP5/NP6 zonal boundary is placed at ~54.0 m, between samplesU24/f924 and U24/f925 (Fig. 7). The upper part of the Te Uri Member,correlated with the Waipawa Formation, is assigned to Zone NP6,
although the uppermost sample examined (U24/f931) was barren.The LO of several marker species occurs in the lower part of theWanstead Formation, suggesting an unconformity between it and theunderlying TeUriMember. The lowermost sample in theWanstead For-mation, at 60.5 m, contains Discoaster mohleri, Heliolithus riedelii andDiscoaster nobilis, and is assigned to upper Zone NP8. The NP8/NP9zonal boundary, marked by the LO of Discoaster multiradiatus, is placedat ~100.0 m, between samples U24/f948 and U24/f950 (Fig. 7). Thestratigraphic interval above this is assigned to Zone NP9.
5.3. Toi Flat-1 core
Dinocyst assemblages are reasonablywell-preserved, although sam-ples from the Te Uri Member had variable preservation. In the lower-most sample examined, at 47.54 m, peridinioid taxa (Spinidinium,Vozzhennikovia angulata) are superabundant and the biostratigraphicallyuseful taxon Alterbidinium pentaradiatum is recorded. Peridinioiddinocysts continue to dominate in the Te Uri Member, with Isabelidiniumcingulatum abundant at 45.56 m and Senegalinium spp. superabundant inthe upper part of the Te Uri Member, in an interval tentatively correlatedwith the Waipawa Formation (Fig. 10). In the lower part of the Te UriMember, at 46.49 m, the gonyaulacoid species Spongodinium cf. delitiensedominates (N60%) the assemblage. The HO of A. pentaradiatum is record-ed at 45.56 m, and the HO of I. cingulatum and Deflandrea foveolata areseen at 42.89 m (Fig. 9).
In the lowermost sample examined from the overlying WansteadFormation, at 38.66 m, the LO ofManumiella rotunda and Vozzhennikoviatawanuiensis sp. nov. is recorded. Gonyaulacoid dinocysts, particularlyCerebrocysta spp., Glaphyrocysta retiintexta, Operculodinium spp.and Spiniferites spp., are superabundant in the Wanstead Formation(Fig. 10). The LO of the genus Apectodinium is recorded at 31.37 m.Combined palynological and geochemical results suggest the P–Eboundary can tentatively be placed between samples at 19.37 mand 16.37 m (Fig. 9). At 16.37 m, the Apectodinium genus is super-abundant and the LO of the genus Hystrichokolpoma is recorded(Fig. 10). Moreover, taxa associated with warm-water conditionsand other PETM sections in New Zealand (e.g., Tawanui, Kumara-2core), such as the genus Kenleyia and various morphotypes assignedto the genus Fibrocysta, are recorded for the first time. High-resolutionanalyses have not yet been completed across the P–E boundary.
Preservation of calcareous nannofossils is variable and barrensamples are present throughout the section. More abundant and bet-ter preserved assemblages were recovered from the mudstone, andassemblages are generally poor from samples within the greensand.Samples from the lower part of the Te Uri Member, at 46.5 m and45.6 m, contain Chiasmolithus bidens and Chiasmolithus danicus,which suggest this interval lies within Zone NP4 (Fig. 9). The pres-ence of Fasciculithus tympaniformis at 43.5 m indicates Zone NP5.The lowermost sample examined in the Wanstead Formation, at38.66 m, contains Discoaster multiradiatus, which is the marker spe-cies for Zone NP9. Currently, zones NP6 to NP8 are not recognised,and it may be that nannofossil assemblages are not preserved orthere is an unconformity between the Te Uri Member and WansteadFormation, as is seen in the nearby Tawanui section. Samples fromthe Wanstead Formation are assigned to Zone NP9. The HO ofEricsonia robusta at 17.32 m may suggest Zone NP10, as this taxonappears to have a HO close to the NP9/NP10 boundary in the NewZealand region.
5.4. Angora Road
Dinocyst assemblages from theWaipawa Formation were less well-preserved and not as diverse as those examined from the Upper Calcar-eous Member of the Whangai Formation. Peridinioid dinocysts are su-perabundant in the lower part of the Upper Calcareous Member andalso in the Waipawa Formation (Fig. 12).
56 E.M. Crouch et al. / Review of Palaeobotany and Palynology 202 (2014) 47–79

Author's personal copy
10
0
30
20
50
40
Dep
th (
m)
Sam
ples
NZ
one
P
NP9
Epo
chE
ocen
eP
aleo
cene
Cordo
spha
eridi
umsp
p.
Lept
odini
um?
pustu
latum
sp. n
ov.
20% 40 60
Spong
odini
um
deliti
ense
cf.
Eisena
ckia
spp.
Fibroc
ysta
spp.
(par
s.)
20
Fibroc
ysta
grou
p
Kenley
ia-Fibr
ocys
tatyp
e
20 40
Glaphy
rocy
sta re
tiinte
xta
Hystri
chok
olpom
asp
p.
Hystri
chos
phae
ridium
tubif
erum
Impa
gidini
umsp
p.
Kallos
phae
ridium
sp.
20
Elytro
cysta
sp. B
20 40
Operc
ulodin
iumsp
p.
20
Cereb
rocy
stasp
p.
20 40
Spinife
rites
spp.
Cleisto
spha
eridi
um p
lacac
anth
um
Thalas
sipho
ra d
elica
ta
Turbios
phae
rasp
p.
gonyaulacoid dinocysts peridinioid dinocysts
Toi Flat-1 core
NP5
NP9
NP4
?NP10
Alterb
idiniu
m p
enta
radia
tum
20 40 60
Apecto
dinium
spp.
20
Cerod
inium
med
calfii
Cerod
inium
/Def
landr
ea
Deflan
drea
deli
neat
atyp
e
Deflan
drea
fove
olata
20
Deflan
drea
robu
stagr
oup
20 40
Isabe
lidini
umsp
p.
20
Man
umiel
la ro
tund
a
Palaeo
cysto
dinium
golz
owen
se
20 40
Seneg
aliniu
msp
p.
20 40
Spinidi
nium
spp.
V. ang
ulata
V. t
awan
uien
sis
sp. n
ov.
NZDP5
NZDP/76
NZ
DP
8D
inoc
yst Z
one
Fig. 10. Abundance record of selected dinocyst species and genera from the Toi Flat-1 core. Summary lithological column and sample points shown. Calcareous nannofossil (NP) zones discussed in text. Dashed line marks the proposed New ZealandDinocyst Paleocene (NZDP) zones. Black box shows the interval tentatively correlated with the Waipawa Formation.
57E.M
.Crouchetal./Review
ofPalaeobotanyand
Palynology202
(2014)47
–79

Author's personal copy
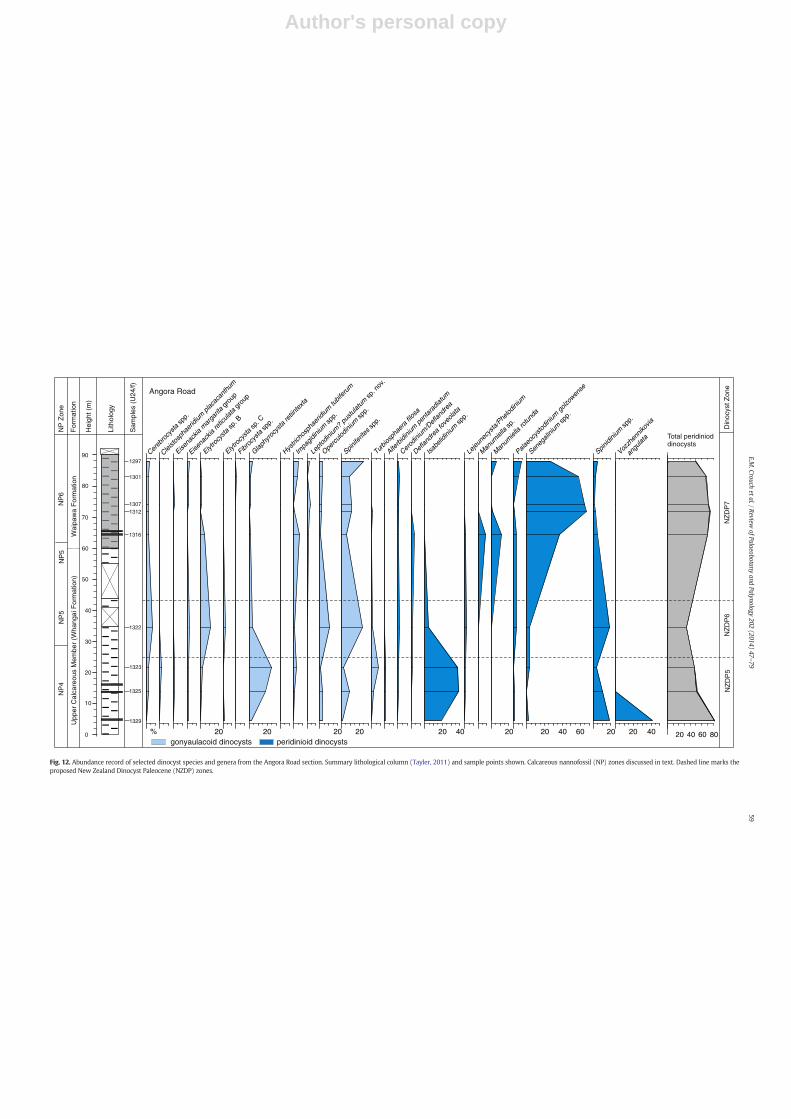
In the section examined from the Upper Calcareous Member, theperidinioid taxon Vozzhennikovia angulata is superabundant in thelowermost sample, at 4.55 m. Stratigraphically above this, theperidinioid genus Isabelidinium (primarily Isabelidinium cingulatum)is abundant, and the gonyaulacoid species Glaphyrocysta retiintextais frequent to abundant (Fig. 12). A similar pattern is also recordedin Columns 2 and 4, mid-Waipara River, with abundant V. angulata re-placed by abundant Isabelidinium species and frequent G. retiintexta(Figs. 4 and 6). The HOs of V. angulata, I. cingulatum and Alterbidiniumpentaradiatum, along with the LO of Deflandrea foveolata, are recordedin the Upper Calcareous Member (Fig. 11). Peridinioid dinocysts areabundant to superabundant in samples from the Waipawa Formation,and are dominated by the genus Senegalinium (Fig. 12). Deflandreafoveolata is present throughout the Waipawa Formation, although it israre and preservation is poor, particularly in the uppermost samplesexamined. In the lower part of the Waipawa Formation, the LO ofManumiella rotunda is seen (Fig. 11).
Calcareous nannofossils are well-preserved, and interestingly this isthefirst timenannofossils have been recovered from sediment typical ofthe Waipawa Formation, which are usually non-calcareous (Kulhaneket al., 2011). The base of the section is assigned to Zone NP4, based onthe presence of the secondary marker taxon Toweius pertusus(Fig. 11). The LO of Chiasmolithus bidens in the lower part of the UpperCalcareous Member, at 13.9 m, indicates upper Zone NP4. The base ofZone NP5, based on the LO of Fasciculithus tympaniformis, is seen at
34.55 m in the Upper Calcareous Member. The uppermost sample ex-amined in the Upper Calcareous Member, at 58.85 m, is also assignedto Zone NP5. The lowermost sample examined in the Waipawa Forma-tion, at 73.58 m, contains the LO of Heliolithus kleinpellii and is assignedto Zone NP6 (Fig. 11). The LO of Discoaster mohleri, themarker taxon forZoneNP7, is recorded in the uppermost part of theWaipawa Formation,~3 m below the base of theWanstead Formation (Supplementary Data,Appendix C).
5.5. ODP Site 1121
Dinocyst assemblages are preserved in an interval from ~110 mto ~56 m (Fig. 13), spanning zones NP5 to NP6/7 (Carter et al.,1999; Wei et al., 2005) and RP5 to RP6 (Hollis, 2002). One sampleat 134.4 m, within Zone NP4, also contains a sparse and low diversitydinocyst assemblage. Palynomorph assemblages are dominated bydinocysts, with spores/pollen and other organic matter virtuallyabsent.
ThePaleocene lower bathyal depositional setting is seen in thedinocystassemblage, with frequent to abundant occurrences of the gonyaulacoidgenus Impagidinium, and common to abundant Cerebrocysta spp. andSpiniferites spp. (Fig. 13). The dominance of these three gonyaulacoidgenera decreases in the upper part of the sequence, in Zone NP6/7,where the peridinioid species Deflandrea foveolata, Senegalinium spp.and taxa tentatively assigned to the genus Eurydinium become a more
0
10
20
30
40
50
60
70
80
90
1326
1329
13251324
1323
1322
1321
1320
1319
1316
13121310
13081307
1306
1301
12981297
noitamro
F awapia
W
6P
N
)noitamro
F iag n ahW( reb
meM suo er acla
C rep pU
enoZ
NP
5P
N5
PN
4P
N
noi tamro
F
)m( thgie
H
ygolohtiL
)f/42U( selp
maS
ata loev of .D
muta ida ra tne pA
mu ini dibretl
mutalugnicI
mu inidilebas
atalugnaV
aivo kinneh zzo
?muin idot peL
muta lu ts up.v on .ps
ad nutorallei
munaM
ataluc iter .E
grou
p
at iragr am .
E grou
p
5P
DZ
N6
PD
ZN
7P
DZ
N
Dinoflagellate cysts
Angora Road
Calcareousnannofossils
simr of inap
myt .F
snedi b .C
s usut rep .T
suc inad .C
eair batnac suh til oi leH
i illepniel k s uhtiloileH
?
enoZ tsyc oni
D
Fig. 11. Stratigraphic distribution of selected dinocysts and calcareous nannofossils from the Angora Road section. Summary lithological column (Tayler, 2011) and sample points shown.Calcareous nannofossil (NP) zones discussed in text. Dashed line marks the proposed New Zealand Dinocyst Paleocene (NZDP) zones.
58 E.M. Crouch et al. / Review of Palaeobotany and Palynology 202 (2014) 47–79
Author's personal copy
0
10
20
30
40
50
60
70
80
90
1329
1325
1323
1322
1316
13121307
1301
1297
Wai
paw
a F
orm
atio
n
NP
6
Upp
er C
alca
reou
s M
embe
r (W
hang
ai F
orm
atio
n)
NZ
one
PN
P5
NP
5N
P4
For
mat
ion
Hei
ght (
m)
Lith
olog
y
Sam
ples
(U
24/f)
Lept
odini
um?
pustu
latum
sp. n
ov.
Eisena
ckia
mar
garit
agr
oup
Eisena
ckia
retic
ulata
grou
p
Fibroc
ysta
spp.
20%
Glaphy
rocy
sta re
tiinte
xta
Hystri
chos
phae
ridium
tubif
erum
Impa
gidini
umsp
p.
20
Elytro
cysta
sp. B
Elytro
cysta
sp. C
20
Operc
ulodin
iumsp
p.
Cereb
rocy
stasp
p.
20
Spinife
rites
spp.
Cleisto
spha
eridi
um
um
placa
cant
h
Turbios
phae
ra fil
osa
gonyaulacoid dinocysts peridinioid dinocysts
Angora Road
Din
ocys
t Zon
e
Alterb
idiniu
mpe
ntar
adiat
um
Cerod
inium
/Def
landr
ea
Deflan
drea
fove
olata
20 40
Isabe
lidini
umsp
p.
Lejeu
necy
sta/P
helod
inium
Man
umiel
lasp
.
20
Man
umiel
la ro
tund
a
Palaeo
cysto
dinium
golzo
wense
20 40 60
Seneg
aliniu
msp
p.
20
Spinidi
nium
spp.
20 40
Vozzh
ennik
ovia
angu
lata
20 40 60 80
Total peridinioddinocysts
NZ
DP
5N
ZD
P6
NZ
DP
7
Fig. 12. Abundance record of selected dinocyst species and genera from the Angora Road section. Summary lithological column (Tayler, 2011) and sample points shown. Calcareous nannofossil (NP) zones discussed in text. Dashed line marks theproposed New Zealand Dinocyst Paleocene (NZDP) zones.
59E.M
.Crouchetal./Review
ofPalaeobotanyand
Palynology202
(2014)47
–79

Author's personal copy
50
60
70
80
90
100
110
120
130
140
NP
4N
P5
NP
6/7
C25
rC
26r
NP
8N
P Z
one
Rad
iola
ria Z
one
Lith
olog
y
Dep
th (
mbs
f)
Box
cor
e
Epo
ch
Chr
on
RP
6 (
)B
urye
lla p
enta
dica
RP
4R
P5
Ear
ly P
aleo
cene
Late
Pal
eoce
neM
iddl
e P
aleo
cene
Sam
ples
7
8
9
10
11
12
15
16
17
13
14
barren
barren
barren of dinocysts
Dinoflagellate cysts
ODP Site 1121
E. m
arga
rita
grou
p
E. r
etic
ulat
agr
oup
Spo
ngod
iniu
mde
litie
nse
cf.
Lept
odin
ium
?pu
stul
atum
sp. n
ov.
Def
land
rea
fove
olat
a
Isab
elid
iniu
m c
ingu
latu
m
Eur
ydin
ium
?gr
oup
Tha
lss
phor
a de
licat
aai
?
?
C26
n?
C26
r??
Spong
odini
um
deliti
ense
cf.
20%
Lept
odini
um?
pustu
latum
sp. n
ov.
Eisena
ckia
mar
garit
agr
oup
Eisena
ckia
retic
ulata
grou
p
Hystri
chos
phae
ridium
tubif
erum
20 40
Impa
gidini
umsp
p.
Elytro
cysta
sp. B
Elytro
cysta
sp. C
20
Operc
ulodin
iumsp
p.
20
Cereb
rocy
stasp
p.
20
Spinife
rites
spp.
Thal ss
phor
a de
licat
a
ai
Turb
iosph
aera
sp.p
gonyaulacoid dinocysts peridinioid dinocysts
Din
ocys
t Zon
eN
ZD
P6
NZ
DP
7
Deflan
drea
/Cer
odini
um
20 40
Deflan
drea
fove
olata
Isabe
lidini
um ci
ngula
tum
20
Euryd
inium
?gr
oup
Palaeo
cysto
dinium
golz
owen
se
20
Seneg
aliniu
msp
p.
Spinidi
nium
spp.
Fig. 13. Stratigraphic distribution and abundance record of selected dinocyst species and genera fromODP Site 1121. Summary lithological column and sample points shown. Black box in the Lithology column shows the interval tentatively correlatedwith the Waipawa and Tartan Formations (Hollis et al., in review). Calcareous nannofossil (NP) zones from Carter et al. (1999) and Wei et al. (2005), and radiolarian (RP) zones from Hollis (2002).
60E.M
.Crouchetal./Review
ofPalaeobotanyand
Palynology202
(2014)47
–79

Author's personal copy
abundant component of the assemblage. Interestingly, this upper se-quence is correlated with the Waipawa and Tartan Formations (Holliset al., in review), and a similar pattern is also noted at mid-WaiparaRiver, Tawanui, Toi Flat-1 core and Angora Road where peridinioiddinocysts are abundant to superabundant during deposition ofWaipawa and Tartan Formation sediments (Figs. 4, 6, 8, 10 and 12).
Deflandrea foveolata is present throughout the interval examined,along with Eisenackia reticulata group, and Leptodinium? pustulatumsp. nov. is present from 100.3 m (Fig. 13). The HO of Isabelidiniumcingulatum is recognised at 87.12 m, in the upper part of Zone NP5.Taxa tentatively assigned to the genus Eurydinium are only recordedin the upper part of the section examined.
6. Paleocene (Teurian) dinocyst zonation
Based on dinocyst assemblage results from mid-Waipara River,Tawanui, Toi Flat-1 core, Angora Road and ODP Site 1121, along withearliest Paleocene records from Clarence Valley sections, mid-WaiparaRiver and Fairfield Quarry (Willumsen, 2003, 2004, 2006; Willumsenand Vajda, 2010; Ferrow et al., 2011;Willumsen, 2011, 2012), a revisedPaleocene (Teurian) dinocyst zonation is proposed. Two importantcriteria for inclusion of selected dinocyst taxa in the zonation is thatthey are a common component of the dinocyst assemblage, and are rou-tinely recognised in eastern New Zealand sedimentary basins.
The dinocyst zones are labelled as NZDP— New Zealand DinocystPaleocene — and are all interval zones. A summary diagram (Fig. 14)shows zones NZDP1 to NZDP8, along with primary and secondarydinocyst bioevents, and current correlation with the InternationalGeological Time Scale (Gradstein et al., 2012). The Paleocenedinocyst zones of Wilson (1987, 1988) are also shown. Where possi-ble, NZDP zones are correlated with calcareous nannofossil zones, ra-diolarian zones, magnetostratigraphic chrons, and carbon isotopes.
6.1. NZDP1 zone
Definition: The interval from the lowest occurrence (LO) of Trithyrodiniumevittii to the LO of Cerodinium striatum.Type section: Mead Stream section, Clarence Valley (Willumsen, 2011),from sample P30/f965 (−0.05 m) to P30/f997 (2.37 m).Age range and correlation: Earliest Paleocene (earliest Teurian). Thelower boundary is correlated with the Cretaceous–Paleogene (K–Pg)boundary, at 66.04 Ma (Gradstein et al., 2012). The NZDP1 zone iscorrelated with radiolarian Zones RP1 to uppermost RP2 (66.04 to~65.0 Ma).Other useful bioevents: The LO of Senoniasphaera inornata is recorded(Willumsen, 2011) at the base of the NZDP1 zone, and the HO ofTrichodinium hirsutum is recorded (Willumsen, 2011) close to the topof the NZDP1 zone. The HO of Manumiella druggii occurs (Willumsen,2003, 2011) within this zone.Remarks: The definition and age range of the NZDP1 zone is based onWillumsen (2011). AtMeadStream, the LOof Trithyrodiniumevittii is re-corded−0.05 mbelow theK–Pg boundary; however, this occurrence ismost likely due to burrow fills. This zone is also based on observationsfrom the mid-Waipara River K–Pg boundary section in Column 1(Willumsen, 2003; Morgans et al., 2005; Willumsen, 2006).
6.2. NZDP2 zone
Definition: The interval from the LOof Cerodinium striatum to thehighestoccurrence (HO) of Trithyrodinium evittii.Type section: Mead Stream section, Clarence Valley (Willumsen, 2011),from sample P30/f998 (2.39 m) to P30/f1018 (18.26 m).Age range and correlation: Early Paleocene (early Teurian). The NZDP2zone is correlated with the uppermost radiolarian Zone RP2 to upper-most RP3 Zone (~65.0 to ~64.0 Ma).
Other useful bioevents: The LO of Alterbidinium pentaradiatum is record-ed within zone NZDP2.Remarks: The definition and age range of the NZDP2 zone is based onWillumsen (2003, 2011). The HO of Trithyrodinium evittii is also record-ed in the Mid-Waipara River section, Column 4, in sample M34/f692(6.32 m) and within calcareous nannofossil Zone NP3.
6.3. NZDP3 zone
Definition: The interval from theHOof Trithyrodinium evittii to theHOofSenoniasphaera inornata.Type section: Mid-Waipara River section, Column 4, from sample M34/f692 (6.32 m) to M34/f699 (31.12 m).Age range and correlation: Early Paleocene (middle early Teurian). Thelower boundary is correlated with uppermost RP3 zone (Willumsen,2011). The NZDP3 zone is correlated with mid to uppermost calcareousnannofossil Zone NP3 (~64.0 to ~63.4 Ma).Other useful bioevents: Palaeoperidinium pyrophorum is frequent toabundantwithin this zone, and the transition between the highest com-mon occurrence (HCO; suggested abundance N10% of assemblage) ofP. pyrophorum and rare P. pyrophorum occurs within this zone. The LOof Vozzhennikovia angulata is recorded within zone NZDP3.Remarks: This zone is based on observations from the mid-WaiparaRiver section, primarily Column 4.
6.4. NZDP4 zone
Definition: The interval from theHOof Senoniasphaera inornata to the LOof Deflandrea foveolata.Type section: Mid-Waipara River section, Column 4, from sample M34/f701 (36.19 m) to M34/f705 (45.46 m).Age range and correlation: Early Paleocene (late early Teurian). TheNZDP4 zone is correlated with uppermost Zone NP3 to mid Zone NP4(~63.4 to ~62.5 Ma).Other useful bioevents: The HO of Palaeoperidinium pyrophorum is re-corded in the uppermost part of the zone, and may offer an alternativebiostratigraphic marker for the top of Zone NZDP4 if Deflandreafoveolata is absent (e.g., D. foveolata is not currently recorded in situfrom the Great South Basin). The LO of Isabelidinium cingulatum, alongwith Isabelidinium bakeri, is recorded within Zone NZDP4. The HO ofCerodinium diebelii and Cribroperidinium wetzelii are recorded withinzone NZDP4.Remarks: This zone is based on observations fromColumns 2 and 4,mid-Waipara River section.
6.5. NZDP5 zone
Definition: The interval from the LO of Deflandrea foveolata to the HO ofAlterbidinium pentaradiatum.Type section: Mid-Waipara River section, Column 2, from sample M34/f650 (63.83 m) to M34/f653 (79.65 m).Age range and correlation: late Early Paleocene (late early Teurian). TheNZDP5 zone is correlated with the upper part of calcareous nannofossilZone NP4 (~62.5 to ~61.7 Ma).Other useful bioevents: The HO of Vozzhennikovia angulata is recorded inthe upper part of this zone.Remarks: This zone is based on observations fromColumns 2 and 4,mid-Waipara River section, Angora Road section and Toi Flat-1 core.
6.6. NZDP6 zone
Definition: The interval from the HO of Alterbidinium pentaradiatum tothe HO of Isabelidinium cingulatum.Type section: Mid-Waipara River section, Column 2, from sample M34/f655 (88.67 m) to M34/f656 (93.69 m).
61E.M. Crouch et al. / Review of Palaeobotany and Palynology 202 (2014) 47–79

Author's personal copy
NZDP3
NZDP4
NZDP5
NZDP6
NZDP7
NZDP8
NZDP2
NZDP1
Trithyrodinium evittii
Trith
yrod
iniu
mev
ittii
Zon
e
Trithyrodinium evittii
Senoniasphaera inornata
Senoniasphaera inornata
Trichodinium hirsutum
Deflandrea foveolata
Deflandrea foveolata
Alterbidinium pentaradiatum
Alterbidinium pentaradiatum
Isabelidinium cingulatum
Isabelidinium cingulatum
Eisenackia reticulata groupEisenackia margarita group
LCO (>40%) ApectodiniumApectodinium genus Hystrichokolpoma genus
Cerodinium striatum
Cerodinium diebeliiCribroperidinium wetzelii
Vozzhennikovia angulata
Manumiella druggii
Vozzhennikovia angulata
Vozzhennikovia tawanuiensis sp. nov.
Manumiella rotunda
Leptodinium? pustulatum sp. nov.
HCO P. pyrophorum
Palaeoperidinium pyrophorum
Primary bioevents
NZ
Pal
eoce
ne d
inoc
yst
(NZ
DP
) zo
ne (
this
stu
dy)
Din
ocys
t zon
e(W
ilson
198
7,19
88)
Secondary bioevents
Pal
aeoc
ysto
dini
um g
olzo
wen
seZ
one
Ape
ctod
iniu
mho
mom
orph
umZ
one
Ma
55
56
57
58
59
60
61
62
63
64
65
66
C29
C28
C27
C26
C25
C24
Late Cretaceous
Eoc
ene
Ear
ly P
aleo
cene
Late
Pal
eoce
ne
Dan
ian
Sel
andi
anT
hane
tian
Ypr
esia
n
low
erTe
uria
nup
per
Teur
ian
Wai
paw
an
P0Pa
P1
P2
P3
P4
P5
P6
CC26
NP1
NP2
NP3
NP4
NP5
NP6
NP7
NP8
NP9
RP1
RP2
RP3
RP4
RP5
RP6
RP7
RP8NP10
Chr
on
Pol
arity
Epo
ch
Sta
ge
NZ
Sta
ge
For
am Z
one
Mid
Pal
eoce
nedl
e
Nan
nopl
ank.
Zon
eR
adio
laria
nZ
one
Fig. 14. Proposed New Zealand Dinocyst Paleocene (NZDP) zones, along with the primary index bioevents. Correlation of the NZDP zones is shownwith the International Time Scale (Gradstein et al., 2012) and New Zealand Stages (from Raine et al.,2013). A selection of secondary dinocyst bioevents considered to be stratigraphically important in the Paleocene is shown. LCO = lowest common occurrence. HCO = highest common occurrence.The Paleocene (Teurian) dinocyst zonation of Wilson (1987, 1988) is provided. Calcareous nannofossil (NP), foraminifera (P), and radiolarian (RP) zones from Gradstein et al. (2012).
62E.M
.Crouchetal./Review
ofPalaeobotanyand
Palynology202
(2014)47
–79

Author's personal copy
Age range and correlation: late Early Paleocene–Middle Paleocene(late early Teurian–early late Teurian). The NZDP6 zone is correlatedwith the upper part of Zone NP4 to upper Zone NP5 (~61.7 to~60.0 Ma).Other useful bioevents: The LO of Leptodinium? pustulatum sp. nov. is re-corded within this zone.Remarks: This zone is based on observations fromColumns 2 and 4,mid-Waipara River section, along with the Angora Road section, Toi Flat-1core, and ODP Site 1121.
6.7. NZDP7 zone
Definition: The interval from the HO of Isabelidinium cingulatum to theHO of Deflandrea foveolata.Type section: Mid-Waipara River section, Column 2, from sample M34/f657 (101.16 m) to M34/f667 (125.23 m).Age range and correlation: Middle Paleocene–early Late Paleocene(late Teurian). The NZDP7 zone is correlated with upper calcareousnannofossil Zone NP5 to Zone NP7 (~60.0 to ~58.8 Ma).Other useful bioevents: The LO of Manumiella rotunda occursstratigraphically below the HO of Deflandrea foveolata. The HO ofEisenackia margarita group and Eisenackia reticulata group occur inthe upper part of Zone NZDP7.Remarks: This zone is based on observations from Columns 2 and 4,mid-Waipara River section, along with the Tawanui section, AngoraRoad section, Toi Flat-1 core, and ODP Site 1121. The age of the topof zone NZDP7 is correlated with calcareous nannofossils; however,it should be noted that these datums are based on sediments associ-ated with the Waipawa and Tartan Formations, and that in the sec-tions examined in this study the top of the Waipawa and TartanFormations appear to be an erosional unconformity (Schiøler et al.,2010; Hollis et al., in review).
6.8. NZDP8 zone
Definition: The interval from the HO of Deflandrea foveolata to lowestcommon occurrence (LCO; N~40%) of the Apectodinium genus.Type section: Tawanui section, from sample U24/f932 (60.5 m, thisstudy) to U24/f1076 (0.03 m; Crouch, 2001).Age range and correlation: Late Paleocene (late Teurian). The base ofzone NZDP8 is correlated with calcareous nannofossil Zone NP7(~58.8 Ma), and the top of the zone is correlated with the Paleocene–Eocene boundary, at 55.96 Ma (Gradstein et al., 2012).Other useful bioevents: The LO of Vozzhennikovia tawanuiensis sp. nov.occurs in the lower part of Zone NZDP8. The LO of the generaApectodinium and Hystrichokolpoma are recorded in the uppermostpart of this zone. The speciesManumiella rotunda is present throughoutZone NZDP8.Remarks: This zone is based on observations fromTawanui section,mid-Waipara River section and Toi Flat-1 core. The base of zoneNZDP8 is ob-served inmid-Waipara River, although currently the top of the zone hasnot been identified. The occurrence and age correlation of the LCO of theApectodinium genus is based on observations from Tawanui (Crouchet al., 2001) and Kumara-2 core, Westland (Sluijs et al., 2008; Handleyet al., 2011). At present, the beginning of the LCO of the Apectodiniumgenus in New Zealand sections does not appear to precede the onsetof the negative CIE, associated with the P–E boundary and PETM, asnoted in other regions, such as New Jersey, North Sea and East TasmanPlateau (Sluijs et al., 2007; Bijl et al., 2013).
7. Discussion
7.1. A New Zealand-wide Paleocene zonation
Dinocyst results from sections in eastern New Zealand sedimen-tary basins underpin the eight Paleocene dinocyst zones proposed
in this paper. However, providing improved biostratigraphic agecontrol in other sedimentary basins around New Zealand, such asthe Taranaki and Great South Basins (Fig. 1) for petroleum explora-tion, is an important application of the proposed Paleocene dinocystzonation.
The primary dinocyst bioevents in the zonation (Fig. 14) are record-ed from Paleocene sections throughout New Zealand, and severalspecies (e.g., Alterbidinium pentaradiatum, Isabelidinium cingulatum)previously have been identified as useful for Paleocene biostratigraphy(Wilson, 1988; Beggs et al., 1992; Schiøler and Roncaglia, 2007; Schiøleret al., 2010;Willumsen, 2003, 2011;Willumsen and Vajda, 2010). In theGreat South Basin (Fig. 1), onshore locations (e.g., Measly Beach core,Fairfield Quarry, Fairfield Estate core) and offshore drillholes (e.g.,Hoiho-1C, Kawau-1A) have good records of these primary bioevents,along with many secondary bioevents such as Palaeoperidiniumpyrophorum, Vozzhennikovia angulata, Eisenackia margarita group andManumiella rotunda (Wilson and McMillan, 1996; McMillan andWilson, 1997; Crouch, 2001; Willumsen, 2003, 2006, 2012; Schiølerand Roncaglia, 2007; Schiøler et al., 2010; Willumsen and Vajda,2010). At present, the peridinioid taxon Deflandrea foveolata has notbeen recorded in situ in the Great South Basin, and has a sporadic occur-rence in western basins such as Taranaki and Northland Basins. Basedon current research, the HO of P. pyrophorum occurs at a similar strati-graphic level as the LO of D. foveolata (Fig. 14), andmay be a good alter-native marker for the top of NZDP4. Likewise, the LO of Manumiellarotunda, and also the HO of E. margarita group and Eisenackia reticulatagroup, occur close to the HO of D. foveolata and may help define the topof NZDP7.
Drillholes in offshore Taranaki (e.g., Tane-1, Witiora-1) and North-land (e.g., Waka Nui-1) Basins are the primary source of Paleocenedinocyst data in western New Zealand sedimentary basins (e.g., Beggset al., 1992; Strong et al., 1999; Cooper et al., 2001; Crampton et al.,2012), although detailed quantitative studies are sparse. As in theGreat South Basin, dinocyst taxa that define the NZDP zones, otherthan Deflandrea foveolata, are recorded in western New Zealand basinsand it appears that the revised Paleocene dinocyst zonation will be ap-plicable to this region.
We have not attempted to correlate with Paleocene sections inAustralia; however, many of the same dinocyst bioevents (e.g., HOTrithyrodinium evittii, HO Palaeoperidinium pyrophorum, LO Apectodiniumgenus) are recorded in Australian regions, such as the Gippsland andOtway Basins (Partridge, 2001, 2004; Mantle et al., 2010). In addition, arecent dinocyst study of Paleocene sediments from ODP Site 1172,offshore Tasmania (Fig. 1), has recorded many of the same taxa (Bijl,2011) and magnetostratigraphic analyses have been completed (Röhlet al., 2004; Bijl et al., 2009, 2013). More detailed comparison of dinocystassemblages and correlation with other forms of age control, such asmagnetostratigraphy, is needed to identify coeval dinocyst bioevents inthe Australasian region.
Paleocene dinocyst events further afield, such as NorthwestEurope (Heilmann-Clausen, 1985; Powell, 1988; Mudge and Bujak,1996; Gradstein et al., 2012), are different enough to warrant estab-lishment of a New Zealand regional zonation. Some dinocyst taxa,such as Palaeoperidinium pyrophorum, Senoniasphaera inornata,Eisenackia margarita and the Apectodinium genus, are recorded inSouthern and Northern Hemisphere sections. However, the strati-graphic range of taxa similar to both hemispheres is often different;for example, the HO of P. pyrophorum in Northwest Europe is seen incalcareous nannofossil Zone NP6 (Mudge and Bujak, 1996; Bujak andBrinkhuis, 1998; Schiøler et al., 2007), while in New Zealand it ap-pears to occur lower in Zone NP4 (Fig. 14). More discussion of thestratigraphic range of selected taxa in New Zealand and records inAustralia and the Northern Hemisphere is provided in Appendix A. In ad-dition, several key Paleocene taxa in New Zealand, such as Alterbidiniumpentaradiatum and Deflandrea foveolata, are not recognised in NorthernHemisphere sections.
63E.M. Crouch et al. / Review of Palaeobotany and Palynology 202 (2014) 47–79

Author's personal copy
7.2. Dinocyst assemblage variations
The dinocyst assemblage records notable relative abundance var-iations in genera and in peridinioid (P) and gonyaulacoid (G) groupsthrough the Paleocene. Peridinioid dinocysts are thought to predom-inantly represent heterotrophic dinoflagellates feeding on diatoms,organic detritus and other phytoplankton, whereas gonyaulacoiddinocysts mainly represent autotrophic dinoflagellates (e.g., Dale,1996; Sluijs et al., 2005). Hence, notable and coeval variations in P/Gratios in multiple locations can provide important informationabout paleoenvironmental conditions in the Paleocene, along withanother potential tool for age correlation between regions withinNew Zealand.
In the Paleocene strata examined, two notable phases ofperidinioid dominated assemblages are recorded. In the Early Paleo-cene, peridinioid dinocysts are superabundant from Zone NZDP2 tolower Zone NZDP5 at mid-Waipara River (Figs. 4 and 6), and also inZone NZDP5 at Angora Road (Fig. 12) and Toi Flat-1 core (Fig. 10).This interval correlates with calcareous nannofossil zones NP3 andNP4 (Fig. 14). At mid-Waipara River, several peridinioid genera areabundant at different times, with Palaeoperidinium pyrophorumdominant in NZDP2 and part of NZDP3, peaks in Spinidinium andVozzhennikovia angulata in NZDP3 and NZDP4, and abundantIsabelidinium in upper NZDP4 and NZDP5. At the base of the sectionexamined in Angora Road and Toi Flat-1 core, V. angulata andSpinidinium are dominant, which is subsequently replaced by abundantIsabelidinium. Acme intervals of P. pyrophorum and Trithyrodinium evittiihave also been seen in mid-Waipara River and Branch and MeadStream, Clarence Valley, in the earliest Paleocene and correlated withradiolarian zones RP1 to RP3 (Willumsen, 2003, 2011). This interval ofsuperabundant peridinioid dinocysts is coincident with a phase of in-creased biogenic silica deposition in Clarence Valley sections, thoughtto be related to a relatively cool and nutrient rich upwelling regime inthe Marlborough region in the earliest Paleocene (Hollis et al., 1995,2003).
The other notable phase of peridinioid dominated assemblages isin the latest Middle to early Late Paleocene, in upper NZDP7 and low-ermost NZDP8, and in an interval that correlates with deposition ofthe Waipawa and Tartan Formation. At mid-Waipara River, speciesof the genus Senegalinium are superabundant in the upper part ofthe Waipara Greensand, which in Column 2 (Fig. 4) is correlatedwith the Tartan Formation (Hollis et al., 2012). This pattern is seenat Tawanui, with abundant Senegalinium species recorded in the up-permost part of the Te Uri Member and calcareous nannofossil ZoneNP6 (Fig. 8), and also in Toi Flat-1 core where Senegalinium is super-abundant in the interval tentatively correlated with the WaipawaFormation (Fig. 10). In samples from the Waipawa Formation at An-gora Road Senegalinium is again the most dominant peridinioidgenus (Fig. 12), and in ODP Site 1121 peridinioid dinocysts aremost abundant (28–43%) from 71.2 m to 56.97 m (Fig. 13), which isassigned to zones NP6/7 and tentatively correlated with the Waipawaand Tartan Formations (Hollis et al., in review). Hence, there is a wide-spread pattern of abundant to superabundant peridinioid assemblages,dominated by the genus Senegalinium, during calcareous nannofossilzones NP6/7 and in sediments characteristic of Waipawa and TartanFormation deposition.
8. Systematic palynology
Systematic descriptions of three species are provided below andillustrations given in Plates I to III. Descriptions are ordered accord-ing to the suprageneric classification of Fensome et al. (1993), andthe descriptive terminology largely follows that in Evitt (1985) andFensome et al. (1993) after Kofoid (1909). Dinocysts recorded inthis study, along with notes on selected taxa, are listed in AppendixA. Selected dinocysts are illustrated in Plates IV to VIII. Full references
for all species mentioned in the text can be found in DINOFLAG2(Fensome et al., 2008).
Division Dinoflagellata (Bütschli, 1885) Fensome et al., 1993Subdivision Dinokaryota Fensome et al., 1993Class Dinophyceae Pascher, 1914Subclass Peridiniphycidae Fensome et al., 1993Order Gonyaulacales Taylor, 1980Family Gonyaulacaceae Lindemann, 1928Genus Leptodinium (Klement, 1960) Stover and Evitt, 1978Type Leptodinium subtile (Klement, 1960)
Leptodinium? pustulatum Crouch et al., sp. nov.Plate I
Holotype: Sample U24/f1026 (Slide SM4836), England Finder co-ordinate H36/1; Plate I, 1–3.Paratype: Sample U24/f1029 (Slide 1), England Finder co-ordinateM49/0; Plate I, 4–6.Repository: The palynological type collection, GNS Science, Lower Hutt,New Zealand.Type locality: Wanstead Formation; Tawanui section, Akitio River,southern Hawkes Bay, New Zealand; grid reference NZMS 260-U24/967 863.Derivation of name: With reference to the presence of pustules on theautophragm.Previous illustrations: 2001, Acanthaulax? sp. A, Crouch, p. 43, Plate I, 1–2.Description: A proximate cyst of small to intermediate size consisting ofan autophragm. The surface of the autophragm has low ridges that forma parasutural ornamentation. The parasutural ridges are discernible bycomplete to incomplete single rows of almost connected low pustules.The ridges can be up to ~3 μm in height and the distal margins of theridges are crenulate. Intratabular ornamentation consists of pustulesor tubercles, which can sometimes be aligned or almost connectedinto secondary ridges. The cyst wall is smooth to finely granularwhere the surface is not ornamented. The paracingulum is generally dis-cernible and the parasulcal region is visible, although identifying all in-dividual paraplates is difficult. Ornamentation is sparse but presentwithin the paracingulum and parasulcus. The cyst has an L-type ventralorganisation, and appears to have neutral to very slight dextral torsion(position of the boundary between the postcingular 4‴/5‴ plates rela-tive to the boundary between the precingular 3″/4″ plates). Apical andantapical horns or protrusions are absent. The archeopyle is a single-plate precingular, type P(3), and the operculum is free inmost observedspecimens. Paratabulation is expressed by parasutural ornamentation,but can be difficult to ascertain. Inferred paratabulation is: 3 to 4′, 5 to6″, 6c, 1ps, ?s, 5 to 6‴, 1p, 1⁗.
Dimensions (in μm) Holotype Range
Overall length 56 49 (56) 66Overall width 58 44 (52) 60
Number of specimens measured: 14.Stratigraphic occurrence: Middle Paleocene (late Teurian)–earliestEocene (early Waipawan). The LO of Leptodinium? pustulatum sp. nov.is inNZDP6 and calcareousnannofossil ZoneNP5, based on observationsat mid-Waipara River and ODP Site 1121. The LO is stratigraphicallybelow the HO of Isabelidinium cingulatum. The HO of Leptodinium?pustulatum sp. nov. is very close to the Paleocene–Eocene boundary.Currently, the only observation is in Tawanui section where the HO isat 0.08 m (U24/f1081) above the P–E boundary (Crouch, 2001).Observed in mid-Waipara River, Tawanui, Toi-Flat-1 core, AngoraRoad, ODP Site 1121, and Moeraki–Hampden in North Otago (Crouch,2001). Rare to common in sections examined.Remarks: When first recorded, Leptodinium? pustulatum sp. nov. wastentatively placed in the genus Acanthaulax (Sarjeant, 1968) Brenner,
64 E.M. Crouch et al. / Review of Palaeobotany and Palynology 202 (2014) 47–79

Author's personal copy
1988, based on the character of the parasutural ridges, the presence ofintratabular ornamentation and the lack of dextral torsion (Crouch,2001). Poulsen (1996) considered Acanthaulax to be a taxonomic juniorsynonym of the genus Cribroperidinium (Neale and Sarjeant, 1962)Helenes, 1984. However, Leptodinium? pustulatum sp. nov. lacks distinc-tive Cribroperidinium characteristics such as marked dextral torsion,parallel accessory septa (‘growth bands’) and an apical protrusion.The species is tentatively included in the genus Leptodinium, basedon the presence of an L-type ventral organisation, neutral torsion,and lack of apical protrusion; however, species of Leptodinium onlypossess parasutural septa and intratabular ornamentation is notpresent. Leptodinium? pustulatum sp. nov. is distinguished from thegenus Corrudinium Stover and Evitt, 1978 by having an L-type ventralorganisation, rather than a S-type organisation. Species of Corrudinium,however, do possess accessory ridges or septa. Leptodinium? pustulatumsp. nov. differs from species of Impagidinium Stover and Evitt, 1978 inpossessing an L-type ventral organisation and accessory ridges. Thecyst is generally seen in lateral view, rather than dorsal-ventral view.
Order Peridiniales Haeckel, 1894Family Peridiniaceae Ehrenberg, 1831Genus Cerodinium (Vozzhennikova, 1963) Lentin and Williams, 1987Type species Cerodinium sibiricum (Vozzhennikova, 1963) Lentin andVozzhennikova, 1990
Cerodinium angulatum Crouch et al., sp. nov.Plate II
Holotype: Sample U24/f1037 (Slide 1), England Finder co-ordinate R38/4; Plate II, 1–2.Paratype: Sample U24/f1032 (Slide SM4838), England Finder co-ordinate J39/3; Plate II, 4–5.Repository: The palynological type collection, GNS Science, Lower Hutt,New Zealand.Type locality: Wanstead Formation; Tawanui section, Akitio River,southern Hawkes Bay, New Zealand; grid reference NZMS 260-U24/967 863.Derivation of name: In reference to the angular shape of the pericyst.Previous illustrations: 2001, Cerodinium sp. cf. Cerodinium depressum,Crouch, p. 43, Plate II, 6.Description: A cornucavate peridinioid cyst of intermediate size, withone apical horn and two antapical horns that are approximately ofequal length. The base of the apical horn is quite narrow and the horngently tapers to a rounded tip. The pericyst outline in dorsal–ventralview is typically elongate and the central part of the pericyst is some-what angular. The periphragm is generally brown, thin, often foldedand can range from a smooth to fine granulate–faintly striate appear-ance. The endocyst is oval to pentagonal in most specimens, with thelength of the endocyst almost equal to or slightly greater than thewidth. Paratabulation is indicated by the paracingulum and sometimesby the archeopyle and the parasulcal region. The paracingulum is dis-cernible by thickened parallel folds in the periphragm and it protrudesslightly in many specimens. The archeopyle is intercalary Type I (2aparaplate). The operculum is free, although it often remains in positionon many specimens. On these, the archeopyle outline can be difficult todetermine.
Dimensions (in μm) Holotype Range
Overall length 97 72 (90) 101Overall width 42 35 (42) 51Endocyst length 44 36 (42) 51Length of apical horn 30 14 (25) 30Length of antapical horns 26 13 (23) 35
Number of specimens measured: 11.Stratigraphic occurrence: Late Paleocene (late Teurian)–Early Eocene(Mangaorapan). The LO of Cerodinium angulatum sp. nov. is in NZDP8
and the lower part of calcareous nannofossil Zone NP8, based on obser-vations at the Tawanui section. The HO is observed in early Eocene sed-iments correlated with the Wilsonidium ornatum Zone (Wilson, 1988)and the NZE4 zone (Crouch, 2001; Morgans et al., 2004), based on ob-servations at the Moeraki–Hampden section. Observed in Tawanui,Moeraki–Hampden and the Measly Beach core (Crouch, 2001). Rare insections examined, but consistently present.Remarks: Cerodinium angulatum sp. nov. differs from Cerodiniumdepressum (Morgenroth, 1966) Lentin and Williams, 1987 in havingan endocyst that is slightly elongated (rather than the width being no-tably greater than the length), by not possessing a conical apical horn,by the central part of the pericyst being angular, and by lacking spineson the paracingulum. Cerodinium angulatum sp. nov. is smaller thanCerodinium diebelii (Alberti, 1959) Lentin and Williams, 1987 and theapical and antapical horns are not as long. Cerodinium angulatum sp.nov. differs from Cerodinium striatum (Drugg, 1967) Lentin and Wil-liams, 1987 in being significantly smaller, having a more angularshape in the central part of the pericyst, and lacking longitudinal rowsof grana on the pericyst wall. Cerodinium angulatum sp. nov. differsfrom Cerodinium speciosum (Alberti, 1959) Lentin and Williams, 1987in being smaller and lacking longitudinal or tubercle ornamentationon the pericyst wall. Cerodinium angulatum sp. nov. is smaller thanCerodinium nielsii (Willumsen, 2011), the pericyst outline is more elon-gate and the apical and antapical horns are longer.
Genus Vozzhennikovia (Lentin and Williams, 1976) Sluijs et al., 2009Type Vozzhennikovia apertura (Wilson, 1967) Lentin andWilliams, 1976
Vozzhennikovia tawanuiensis Crouch et al., sp. nov.Plate III
Holotype: Sample U24/f960 (Slide SM4839), England Finder co-ordinateL36/0; Plate III, 1–2.Paratype: Sample U24/f960 (Slide SM4840), England Finder co-ordinateH36/0; Plate III, 3.Repository: The palynological type collection, GNS Science, Lower Hutt,New Zealand.Type locality: Wanstead Formation; Tawanui section, Akitio River,southern Hawkes Bay, New Zealand; grid reference NZMS 260-U24/967 863.Derivation of name: After Tawanui farm, where the type section islocated.Previous illustrations: 2001, Spinidinium sp. cf. Spinidinium densispinatum,Crouch, p. 47, Plate X, 3, 4.Description: A proximate cyst of intermediate size, with a broad roundedapex and two very low antapical protrusions that taper into short horns.The left antapical horn can be slightlymore developed than the right, al-though both antapical horns are short (up to 9 μm in specimens record-ed). The apex lacks protrusion and horn. The cyst is cornucavate and thecavation is generally most obvious in the hypocyst near the base of theantapical horns. The epicyst and hypocyst are approximately the samesize. The periphragm and endophragm are both thin and rounded topentagonal in outline in dorso-ventral view. The periphragm isornamented with short (up to 5 μm in specimens recorded), rod-likecapitate processes that are closed distally. The processes are not denselyarranged and ornamentation is non-tabular. The processes tend to bemore common in the anterior and posterior regions, and also on themargins. The endophragm is difficult to delineate in most specimens.The archeopyle is an intercalary, Type I (2a paraplate), that can be ob-scure. The operculum is free but remains in position in some specimens.
Dimensions (in μm) Holotype Range
Overall length 58 52 (58) 71Overall width 61 50 (58) 70Endocyst length 51 47 (52) 65Endocyst width 56 46 (52) 64
65E.M. Crouch et al. / Review of Palaeobotany and Palynology 202 (2014) 47–79

Author's personal copy
Plate I. Leptodinium? pustulatum Crouch et al. sp. nov. Scale bar = 20 μm. All specimens are from the Tawanui section (NZMS 260-U24). The sample number, GNS Science palynology Lnumber, and/or slide/single mount (SM) number, and England Finder co-ordinate are listed.
1–3. Holotype, U24/f1026 (SM4836), H36/1. 1 — dorsal view, low focus; 2–3 — ventral view, high focus.4–6. Paratype, U24/f1029 (Slide 1), M49/0. 4 — dorsal view, low focus; 5–6 — ventral view, high focus.7. U24/f1010 (Slide 1), E33/1. Dorsal view, low focus.8–9. U24/f0153 (Slide 1), R34/0. 8 — left lateral view, high focus; 9 — left lateral view, low focus.10. U24/f1051 (Slide 1), Q33/4. Dorsal view, high focus.11–12. U24/f1029 (Slide SM4837), P41/0. 11— dorsal view, low focus; 12— lateral view, high focus.
Plate II. Cerodinium angulatum Crouch et al. sp. nov. Scale bar = 20 μm. All specimens are from the Tawanui section (NZMS 260-U24). The sample number, GNS Science palynology Lnumber, and/or slide/single mount (SM) number, and England Finder co-ordinate are listed. (see on page 68)
1–2. Holotype, U24/f1037 (Slide 1), R38/4. Dorso-ventral view.3. U24/f1050 (Slide 1), O40/4. Dorso-ventral view.4–5. Paratype, U24/f1032 (Slide SM4838), J39/3. Dorso-ventral view.6. U24/f1029 (Slide 1), H41/2. Lateral view.7–8. U24/f1023 (Slide 1), T36/3. Dorso-ventral view.9. U24/f960 (Slide 3), L33/2. Dorso-ventral view.
Plate III. Vozzhennikovia tawanuiensis Crouch et al. sp. nov. Scale bar = 20 μm. The sample number, GNS Science palynology L number, and/or slide/single mount (SM) number, andEngland Finder co-ordinate are listed. (see on page 69)
1–2 Holotype, Tawanui section, U24/f960 (Slide SM4839), L36/0. 1 — dorso-ventral view, low focus; 2 — dorso-ventral view, high focus.3. Paratype, Tawanui section, U24/f960 (Slide SM4840), H36/0. Dorso-ventral view.4–5. Tawanui section, U24/f1004 (Slide 1), C41/4. 4 — dorso-ventral view, high focus; 5 — dorso-ventral view, low focus.6 and 9. Tawanui section, U24/f1007 (Slide 1), S37/0. 6 — dorso-ventral view, high focus; 9 — dorsal-ventral view, low focus.7–8. Tawanui section, U24/f1000 (Slide 2), O36/1. 7 — dorso-ventral view, low focus; 8 — dorso-ventral view, high focus.10–11. Moeraki–Hampden section, J42/f386 (Slide 1), H46/0. 10 — dorso-ventral view, high focus; 11 — dorso-ventral view, medium focus.12. Tawanui section, U24/f962 (Slide 1), F39/3. Dorso-ventral view.
Plate IV. Scale bar = 20 μm. Section, sample number, GNS Science palynology L number and slide number, and England Finder co-ordinate are listed. (see on page 70)
1. Trithyrodinium evittii Mid-Waipara River section, M34/f690, L21650/1, L43/0.2. Trithyrodinium evittii Mid-Waipara River section, M34/f690, L21650/4, K33/1.3. Senoniasphaera inornataMid-Waipara River section, M34/f645, L21684/2, T32/0.4. Alterbidinium pentaradiatummid-Waipara River section, M34/f695, L21653/2, K37/45. Alterbidinium pentaradiatumMid-Waipara River section, M34/f653, L21689/1, U52/0.6. Senoniasphaera inornataMid-Waipara River section, M34/f645, L21684/2, M38/0.7. Cerodinium striatum Mead Stream section, P30/f1010, L18518/4, N36/0.8. Deflandrea foveolata Mid-Waipara River section, M34/f706, L21161/2, Q49/49. Deflandrea foveolata Angora Road section, U24/f1316, L25746/1, R37/110. Deflandrea foveolata ODP Site 1121B-9X-CC (71.2 m), L18756/1, O37/3.11. Isabelidinium cingulatumMid-Waipara River section, M34/f649, L21686/2, S52/2.12. Isabelidinium cingulatumMid-Waipara River section, M34/f706, L21161/2, Q37/0.13. Apectodinium round group Toi Flat-1 core, 16.37 m, L21703/2, W32/4.14. Apectodinium angular group Toi Flat-1 core, 28.37 m, L24187/1, P41/2.
Plate V. Scale bar = 20 μm. Section, sample number, GNS Science palynology L number and slide number, and England Finder co-ordinate are listed. (see on page 71)
1. Cerebrocysta waipawaensis Toi Flat-1 core, 28.37 m, L24187/1, H34/0.2. Cerebrocysta sp. A Toi Flat-1 core, 19.37 m, L21704/1, E41/3.3. Cerodinium diebelii Mid-Waipara River section, M34/f697, L21654/2, V42/2.4. Corrudinium cf. incompositum Mid-Waipara River section, M34/f656, L21155/2, H32/0.5. Corrudinium cf. incompositum Mid-Waipara River section, M34/f656, L21155/2, C44/4.6. Elytrocysta sp. B Toi Flat-1 core, 38.38–38.46 m, L26787/2, K40/4.7. Elytrocysta sp. C Mid-Waipara River section, M34/f704, L21658/1, J45/0.8. Cribroperidinium wetzeliiMid-Waipara River section, M34/f694, L21652/1, F42/0.9. Cribroperidinium wetzelii Angora Road section, MT3.03, L25876/2, Y39/0.10. Cleistosphaeridium placacanthumMid-Waipara River section, M34/f652, L21688/1, R38/2.11. Eurydinium? group ODP Site 1121B-8X-4, 46.5–48.5 cm (56.97 m), L26457/1, Q32/0.12. Eurydinium? group ODP Site 1121B-8X-4, 46.5–48.5 cm (56.97 m), L26457/1, M29/2.13. Eurydinium? group ODP Site 1121B-9X-2, 45.5–47.5 cm (63.56 m), L26461/1, Q38/0.14. Eurydinium? group ODP Site 1121B-8X-4, 46.5–48.5 cm (56.97 m), L26457/1, U42/3.
Plate VI. Scale bar = 20 μm. Section, sample number, GNS Science palynology L number and slide number, and England Finder co-ordinate are listed. (see on page 72)
1. Eisenackia margarita group Angora Road section, U24/f1316, L25746/1, W29/4.2. Eisenackia margarita group Mid-Waipara River section, M34/f660, L21693/1, R37/0.3. Eisenackia margarita group Mid-Waipara River section, M34/f660, L21693/2, U53/3.4. Eisenackia reticulata group ODP Site 1121B-17X-CC (134.4 m), L18765/2, O30/3.5. Eisenackia reticulata group Mid-Waipara River section, M34/f705, L21659/2, H40/4.6. Eisenackia margarita group ODP Site 1121B-17X-CC (134.4 m), L18765/1, H30/3.7. Eisenackia reticulata group Mid-Waipara River section, M34/f656, L21154/2, P41/0.8. Eisenackia reticulata group Mid-Waipara River section, M34/f701, L21657/2, X48/1.9. Hystrichokolpoma cf. cinctum Mid-Waipara River section, M34/f678, L21153/3, Q50/4.10. Glaphyrocysta retiintexta Angora Road section, U24/f1325, L25878/2, J29/4.11. Impagidinium sp. A Mid-Waipara River section, M34/f678, L21153/3, L38/2.12. Lingulodinium machaerophorum Toi Flat-1 core, 28.37 m, L24187/1, G44/4.
66 E.M. Crouch et al. / Review of Palaeobotany and Palynology 202 (2014) 47–79
Author's personal copy
21 3
4 5 6
11 12
7 8 9
10
67E.M. Crouch et al. / Review of Palaeobotany and Palynology 202 (2014) 47–79
Author's personal copy
1 2 3
4 5 6
87 9Plate II (caption on page 66).
68 E.M. Crouch et al. / Review of Palaeobotany and Palynology 202 (2014) 47–79
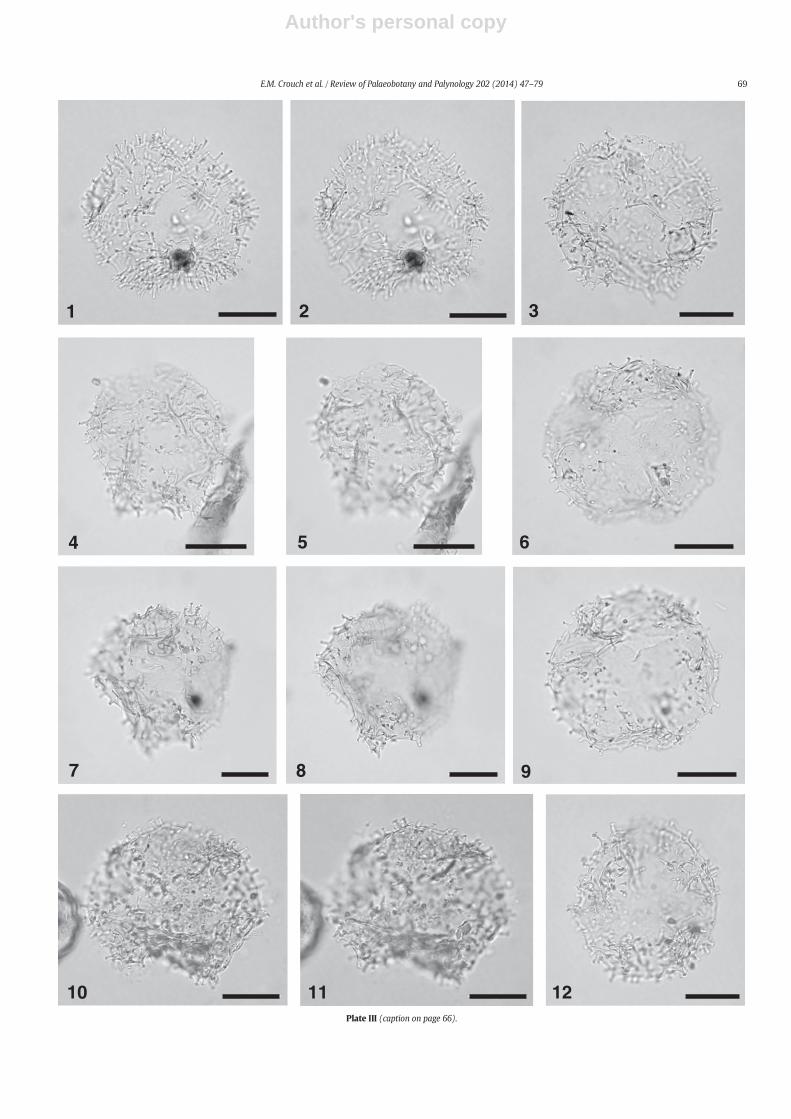
Author's personal copy
1 2 3
4 5 6
7 8 9
10 11 12Plate III (caption on page 66).
69E.M. Crouch et al. / Review of Palaeobotany and Palynology 202 (2014) 47–79
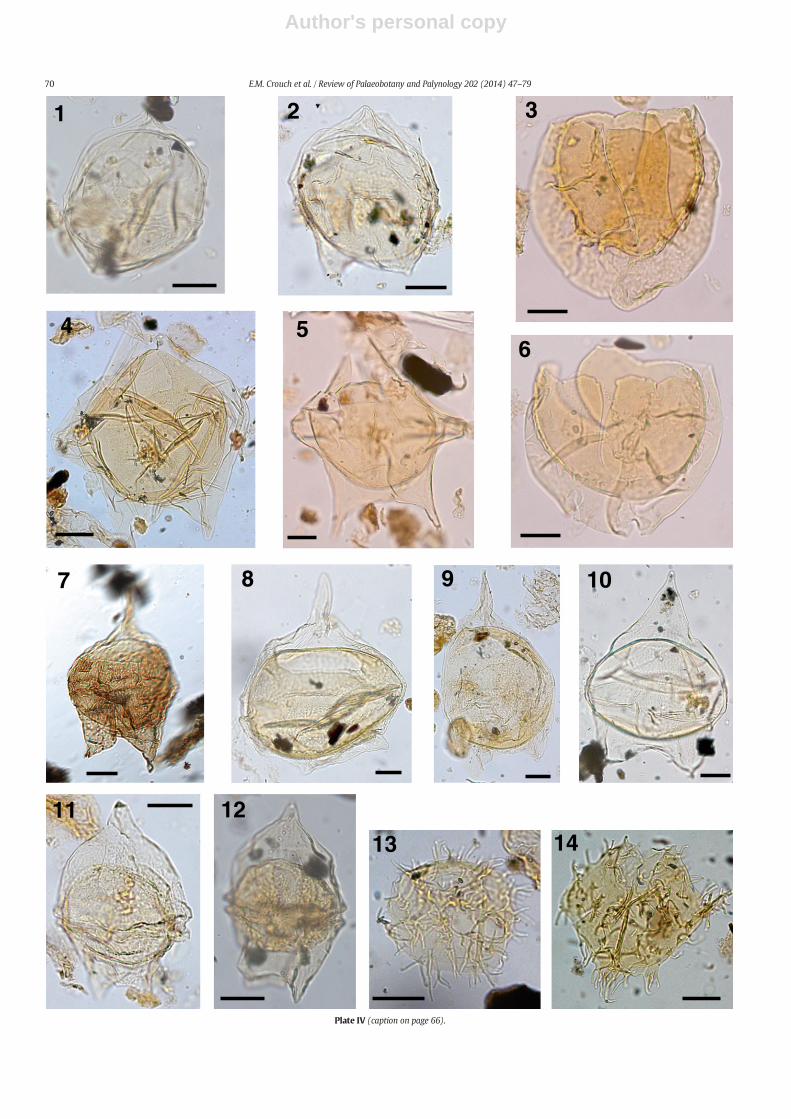
Author's personal copy
1 2
4 5
8 9 10
11 12
13 14
3
7
6
Plate IV (caption on page 66).
70 E.M. Crouch et al. / Review of Palaeobotany and Palynology 202 (2014) 47–79
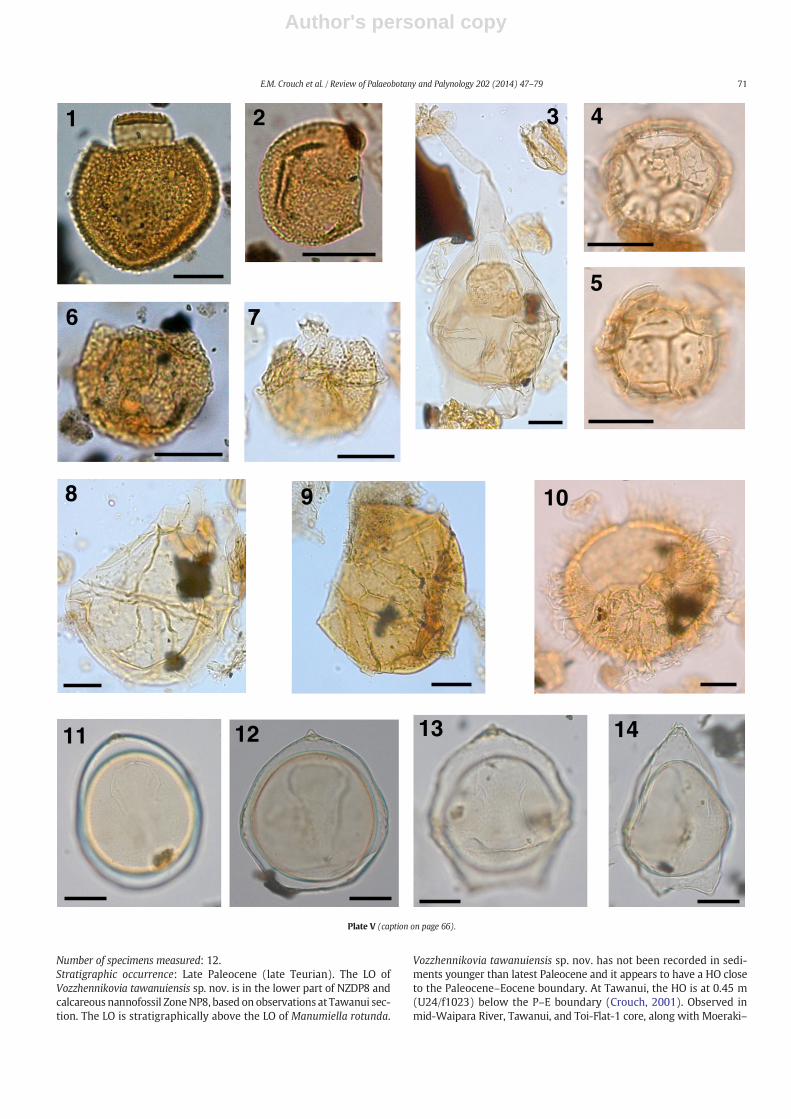
Author's personal copy
Number of specimens measured: 12.Stratigraphic occurrence: Late Paleocene (late Teurian). The LO ofVozzhennikovia tawanuiensis sp. nov. is in the lower part of NZDP8 andcalcareous nannofossil ZoneNP8, based on observations at Tawanui sec-tion. The LO is stratigraphically above the LO of Manumiella rotunda.
Vozzhennikovia tawanuiensis sp. nov. has not been recorded in sedi-ments younger than latest Paleocene and it appears to have a HO closeto the Paleocene–Eocene boundary. At Tawanui, the HO is at 0.45 m(U24/f1023) below the P–E boundary (Crouch, 2001). Observed inmid-Waipara River, Tawanui, and Toi-Flat-1 core, along with Moeraki–
1 2
6
3
9
11
7
12 1413
4
5
108
Plate V (caption on page 66).
71E.M. Crouch et al. / Review of Palaeobotany and Palynology 202 (2014) 47–79
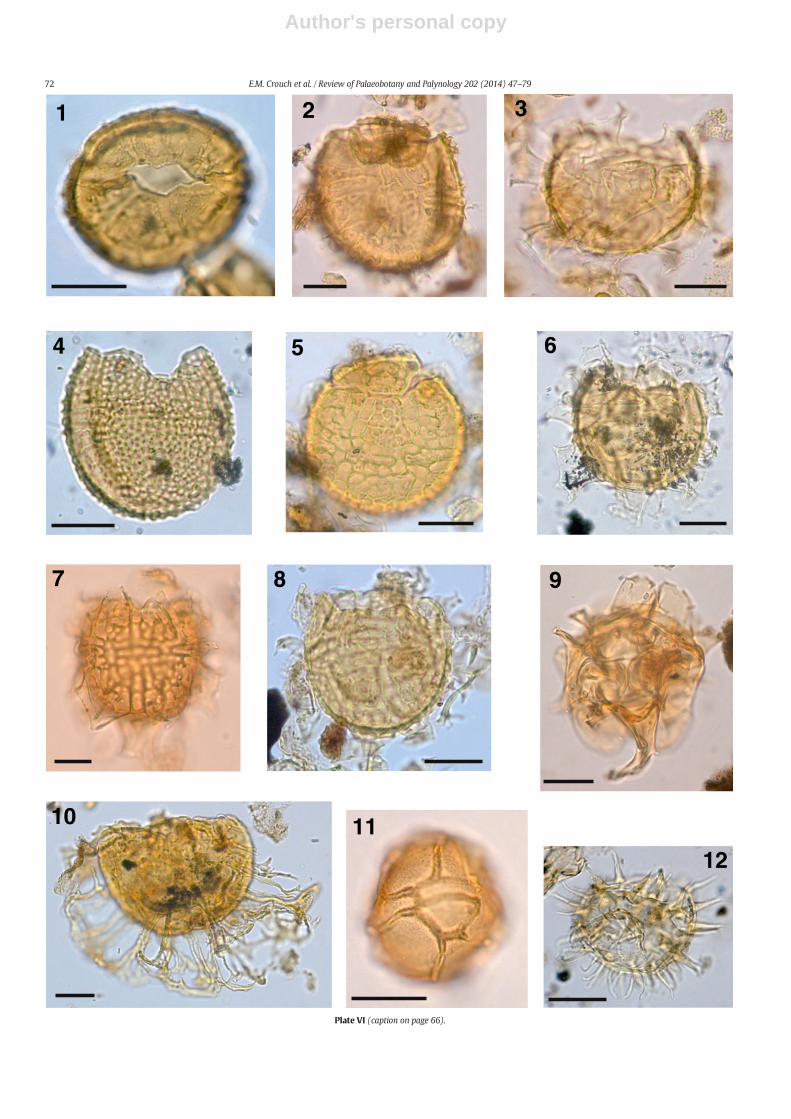
Author's personal copy
7
4 6
2
11
9
3
8
10
1
12
5
Plate VI (caption on page 66).
72 E.M. Crouch et al. / Review of Palaeobotany and Palynology 202 (2014) 47–79

Author's personal copy
Hampden andMeasly Beach core inNorthOtago (Crouch, 2001). Rare tocommon in sections examined.Remarks: Vozzhennikovia tawanuiensis sp. nov. is placed in the genusVozzhennikovia Lentin and Williams, 1976, following discussion bySluijs et al. (2009). They “retain Vozzhennikovia for peridiniaceandinocysts with an isodeltaform to isothetaform 2a, an archeopyle thatcan result from the loss of the 2a only or all three intercalary platesand a more or less uniform covering of spines of coni on the cyst thatmay or may not reflect tabulation”. Vozzhennikovia tawanuiensis sp.nov. differs from Vozzhennikovia angulata Wilson, 1988 in lacking aperiphragm that is sharply angular in outline, in not having densely ar-ranged processes, and in lacking an easily discernible paracingulum. Itdiffers from Vozzhennikovia rotunda (Wilson, 1967) Lentin and Wil-liams, 1976 in lacking development of an apical and left antapicalhorn, in having a less globular shape, and in lacking spinules in theparacingulum area. Vozzhennikovia tawanuiensis sp. nov. is distin-guished from Spinidinium densispinatum Stanley, 1965 by lacking a dis-tinct paracingulum, by having no evidence of sutural spines and bylacking a well-developed left antapical horn and apical horn. Somespecimens of Vozzhennikovia tawanuiensis sp. nov. resemble roundedforms of Apectodinium homomorphum (Deflandre and Cookson,1955) Lentin and Williams, 1977; however, the short rod-like capitateprocesses are distinctive for V. tawanuiensis sp. nov. and, furthermore,A. homomorphum has a more uniform distribution of processes and ev-idence of intratabular ornamentation.
9. Conclusions
Diverse and well-preserved organic-walled dinocyst assemblagesare found in almost all marine settings of Paleocene age throughoutNew Zealand. This paper documents dinocyst assemblages fromPaleocene, or New Zealand Teurian Stage, sediments in easternNew Zealand and provides a revised Paleocene dinocyst zonation.
The Paleocene sections come from onshore locations in the EastCoast (Tawanui, Angora Road, Toi Flat-1 core) and Canterbury(mid-Waipara River) sedimentary basins, and ODP Site 1121 on theeastern flank of the Campbell Plateau. Calcareous nannofossil assem-blages were also examined from these sections to assist with the ageand correlation of dinocyst records. In addition, published dinocystdata from earliest Paleocene sections in East Coast, Canterbury andGreat South Basins were utilised in the lowermost dinocyst zones.
The Paleocene (Teurian) dinocyst zones are labelled as NZDP—NewZealand Dinocyst Paleocene — and are all interval zones. The eightzones, NZDP1 to NZDP8, encompass the entire Paleocene, from the Cre-taceous–Paleogene boundary at 66.04 Ma to the Paleocene–Eoceneboundary at 55.96 Ma. The stratigraphic distribution of the primarydinocyst bioevents, on which the zones are based, is shown, alongwith secondary bioevents that are considered to be stratigraphically im-portant in the Paleocene. The NZDP zones are correlated with the 2012International Geological Time Scale (Gradstein et al., 2012), and are pri-marily based on correlationwith calcareous nannofossil biostratigraphy(NP zones).While Zones NZDP1 to NZPD8 are based on sections in east-ern New Zealand, it appears that the zonation will be applicablethroughout the New Zealand region, and perhaps wider afield in thesouthwest Pacific.
In the Paleocene, dinocyst assemblages record prominent relativeabundance variations in peridinioid and gonyaulacoid groups.Peridinioid dinocysts, thought to represent primarily heterotrophicdinoflagellates, are abundant in two intervals. Several peridiniodgenera dominate assemblages through the Early Paleocene, whichis coeval with a record in Marlborough, East Coast Basin, of increasedbiogenic silica deposition due to a high productivity upwelling zone.Peridinioid dinocysts, primarily species of the genus Senegalinium,are also abundant to superabundant in the late Middle to early LatePaleocene and in an interval that correlates with deposition of an
organic-rich dark mudstone, the Waipawa and Tartan Formations,which is present throughout the New Zealand region.
The following three new dinocyst species are described: Leptodinium?pustulatum sp. nov., Cerodinium angulatum sp. nov., and Vozzhennikoviatawanuiensis sp. nov.
Acknowledgements
We thank Brad Field, Chris Hollis, Hugh Morgans, Ian Raine, PercyStrong, and Michael Tayler for sample collection. Sonja Fry, RandallMcDonnell and Roger Tremain are thanked for sample preparationand palynology and nannofossil processing. Assistance with dinocystsystematics from Henk Brinkhuis, Poul Schiøler and Graeme Wilsonwas greatly appreciated. Peter Bijl is thanked for access to ODP Site1172 dinocyst data and on-going discussions. Discussions and com-ments by Chris Hollis and Poul Schiøler greatly improved the manu-script. EMC and DKK acknowledge support from GNS Science researchprogrammes Global Change with Time, and Petroleum Basin Research.PSW acknowledges financial support by MUSERUM, Skive, Denmark.We thank Peter Bijl and Michael Hannah for constructive reviews.
Appendix A
List of dinocysts recorded from Paleocene sections examined inthis study. The references for the author citations listed below aregiven in DINOFLAG2 (Fensome et al., 2008). Notes on selected generaand species are provided. Selected dinocysts are illustrated in PlatesIV to VIII.Achomosphaera crassipellis (Deflandre and Cookson, 1955) Stover andEvitt, 1978Alterbidinium pentaradiatum (Cookson and Eisenack, 1965) Lentin andWilliams, 1985 (Plate IV, 4, 5)Remarks: Both Alterbidinium pentaradiatum subsp. pentaradiatum(Cookson and Eisenack, 1965) Stover, 1974 and Alterbidiniumpentaradiatum subsp. precedum (Cookson and Eisenack, 1974) LentinandWilliams, 1985 are seen inmid-Waipara River, Toi Flat-1 core andAn-gora Road. In general, A. pentaradiatum ssp. precedum is more common,and specimens are often folded or partly broken. The stratigraphic rangeis similar and the occurrences are shown as Alterbidinium pentaradiatum,which includes both A. pentaradiatum subsp. pentaradiatum andA. pentaradiatum subsp. precedum.Apectodinium round group (Plate IV, 13)Remarks: A variety of Apectodinium morphotypes are recognised. Thisgroup includes round forms such as Apectodinium homomorphum(Deflandre and Cookson, 1955) Lentin and Williams, 1977, andApectodinium parvum (Alberti, 1971) Lentin and Williams, 1977.Apectodinium angular group (Plate IV, 14)Remarks: A variety of Apectodinium morphotypes are recognised. Thisgroup includes angular forms such as Apectodinium hyperacanthum(Cookson and Eisenack, 1965) Lentin and Williams, 1977 andA. quinquelatum (Williams and Downie, 1966) Costa and Downie, 1979.Apectodinium spp. (pars.)Remarks: In this study, the LO of the genus Apectodinium is in calcareousnannofossil Zone NP9, and close to the Paleocene–Eocene boundary.This record differs to Northwest Europe, where the LO of Apectodiniumhomomorphum is in Zone NP8 (Powell et al., 1996; Bujak andBrinkhuis, 1998) and in lower latitudes where the genus first appearsin the Middle Paleocene (Bujak and Brinkhuis, 1998; Crouch et al.,2003).Areoligera senonensis Lejeune-Carpentier, 1938Areoligera spp. (pars.)Cerebrocysta waipawaensis (Wilson, 1988) Fensome et al., 2009(Plate V, 1)Cerebrocysta cf. waipawaensisRemarks: This taxon is similar to Cerebrocysta waipawaensis, but dif-fers primarily in the degree of reticulation of the autophragm. The
73E.M. Crouch et al. / Review of Palaeobotany and Palynology 202 (2014) 47–79

Author's personal copy
reticulation pattern is less defined; the projecting ridges that formthe muri are higher and more numerous, and this gives a lessorganised appearance. The size is similar to C. waipawaensis.Cerebrocysta sp. A (Plate V, 2)Remarks: These specimens are proximate and subspherical in outline,with a precingular (3″) archeopyle. The autophragm is sharply definedby very short ridges that form a reticulate surface, which is quite evenlydistributed and the pattern is small and dense. Paratabulation is not dis-cernible. The cyst size is small (average ~30 μm). The taxon differs toCerebrocysta waipawaensis in having a smaller cyst size, thinner walland a smaller reticulate pattern.Cerebrocysta baculate groupRemarks: Includes specimens in which the autophragm has a baculatesurface. A variety in the length and the number of baculae is observed.Cerebrocysta spp. (pars.)Remarks: All specimens previously assigned to the genus Pyxidinopsisare now assigned to the genus Cerebrocysta, following Fensome et al.(2009). They state the Cerebrocysta “genus is very difficult to differenti-ate from Pyxidinopsis” and that “weprefer to retain Cerebrocysta for spe-cies of mainly Cenozoic grouping and Pyxidinopsis for species of theJurassic–Neocomian grouping”.Cerodinium diebelii (Alberti, 1959) Lentin and Williams, 1987 (PlateV, 3)Cerodinium medcalfii (Stover, 1974) Lentin and Williams, 1987Cerodinium striatum (Drugg, 1967) Lentin andWilliams, 1987 (Plate IV,10)Cerodinium spp. (pars.)Cleistosphaeridium placacanthum (Deflandre and Cookson, 1955) Daveyet al., 1969; Islam, 1993 (Plate V, 10)Cleistosphaeridium spp. (pars.)Cordosphaeridium fibrospinosum (Davey and Williams, 1966) Davey1969Cordosphaeridium spp. (pars.)Corrudinium cf. incompositum (Drugg, 1970) Stover and Evitt, 1978(Plate V, 4, 5)Cribroperidinium wetzelii (Lejeune-Carpentier, 1939) Helenes, 1984(Plate V, 8, 9)Cribroperidinium spp. (pars.)Deflandrea delineata Cookson and Eisenack, 1965Deflandrea foveolata Wilson, 1984 (Plate IV, 7–9)Deflandrea galeata (Lejeune-Carpentier, 1942) Lentin and Williams,1973Deflandrea cf. phosphoritica Eisenack, 1938Deflandrea robusta groupRemarks: This includes species Deflandrea robusta Deflandre andCookson, 1955 and D. truncata Stover, 1974.Deflandrea spp. (pars.)Eisenackia circumtabulata Drugg, 1967Eisenackia margarita group (Plate VI, 1, 2, 3, 6)
Remarks: This group includes Eisenackia margarita (Harland, 1979)Quattrocchio and Sarjeant, 2003, which has penitabular membranesthat havefinely serrate or smooth distalmargins andhave a “beaded ap-pearance”. As Harland (1979)mentions there is variability in the height,from 3 to 10 μm, of the penitabular membranes and this variability isnoted in this study. In addition, this group also includes forms thathave high membranous processes, similar to that illustrated byHeilmann-Clausen (1985). It appears that E. margarita has a similarHO in New Zealand and Northwest Europe, with the HO seen in calcar-eous nannofossil zoneNP6/7 in this study (Fig. 14) and close to theNP7/NP8 zonal boundary in Northwest Europe (e.g., Mudge and Bujak, 1996;Bujak and Brinkhuis, 1998).Eisenackia reticulata group (Plate VI, 4, 5, 7, 8)Remarks: This group includes Eisenackia reticulata (Damassa, 1979)Quattrocchio and Sarjeant, 2003, with low penitabular ridges and anintratabular ornamentation resembling a low reticulum. This groupalso includes specimens that Wilson (1988) described and illustratedas Cassidium filosum, which are formswith intratabular processes. Vari-ability in the height of the processes is seen in this study. In addition,specimens illustrated byWilson (1988) as Cassidium fragile are includedin the Eisenackia reticulata group. However, forms assigned to C. fragile,with no penitabular ridges, have been recorded in New Zealand and ap-pear to be restricted to the earliest Paleocene (Willumsen, 2003, 2011).Eisenackia spp.Remarks: Quattrocchio and Sarjeant (2003) “consider that Eisenackiamust be treated as a senior synonym of Alisocysta and that the latternames cease to be used”. This diagnosis is adopted in this study.Elytrocysta sp. B (Plate V, 6)Remarks: Specimens previously illustrated (Crouch, 2001) asMembranosphaera sp. B de Connick, 1975, and also illustrated inHeilmann-Clausen (1985). These specimens are now included inthe Elytrocysta genus, following Fensome et al. (2008, 2009) whoplaced the type species, Membranosphaera maastrichtia, in thegenus Elytrocysta.Elytrocysta sp. C (Plate V, 7)Remarks: Specimens very closely resemble Elytrocysta sp. B; however,there is a discernible paratabulation present.Eurydinium? group (Plate V, 11–14)Remarks: This group of dinocysts were seen in ODP Site 1121 and avariety of morphotypes are recognised. Specimens range fromround circumcavate forms with one apical prominence, no antapicalprominence and no discernible cingulum (Plate V, 11), to forms thatare lenticular in outline, bicavate or circumcavate, cingulumdiscernible,one apical prominence and two symmetrical antapical horns (Plate V,13–14). In general, the operculum is attached anteriorly. Themorphotypes are tentatively placed in the genus Eurydinium based ona more elliptical shaped endocyst than is seen in the Isabelidinium andManumiella genera, which tend to have a more rounded endocyst orand endocyst where the width is greater than the length. However,
Plate VII. Scale bar = 20 μm. Section, sample number, GNS Science palynology L number and slide number, and England Finder co-ordinate are listed.
1. Isabelidinium bakeriMid-Waipara River section, M34/f706, L21161/2, X49/2.2. Isabelidinium bakeriMid-Waipara River section, M34/f651, L21155/4, M30/0.3. Isabelidinium cf. pellucidumMid-Waipara River section, M34/f652, L21688/1, W49/2.4. Isabelidinium cf. pellucidumMid-Waipara River section, M34/f706, L21161/2, G33/3.5. Manumiella rotundaMid-Waipara River section, M34/f667, L21696/2, M33/4.6. Manumiella rotunda Tawanui section, U24/f929, L21781/2, O48/1.7. Phthanoperidinium? sp. A Mid-Waipara River section, M34/f675, L21699/2, Q43/0.8. Palaeocystodinium cf. australinumMid-Waipara River section, M34/f698, L21159/2, O44/1.9. Senegalinium dilwynenseMid-Waipara River section, M34/f695, L21653/2, Q38/2.10. Palaeoperidinium pyrophorumMid-Waipara River section, M34/f645, L21684/2, T44/0.11. Phthanoperidinium? sp. A Mid-Waipara River section, M34/f675, L21699/2, H51/1.12. Senegalinium dilwynense ODP Site 1121B-9X-CC (71.2 m), L18756/1, Y47/2.13. Senegalinium cf. dilwynenseMid-Waipara River section, M34/f667, L21696/2, D49/0.14. Senegalinium cf. dilwynense Toi Flat-1 core, 40.55–40.60 m, L26791/1, K31/0.15. Palaeocystodinium golzowenseMid-Waipara River section, M34/f703, L21160/1, X43/4.
74 E.M. Crouch et al. / Review of Palaeobotany and Palynology 202 (2014) 47–79
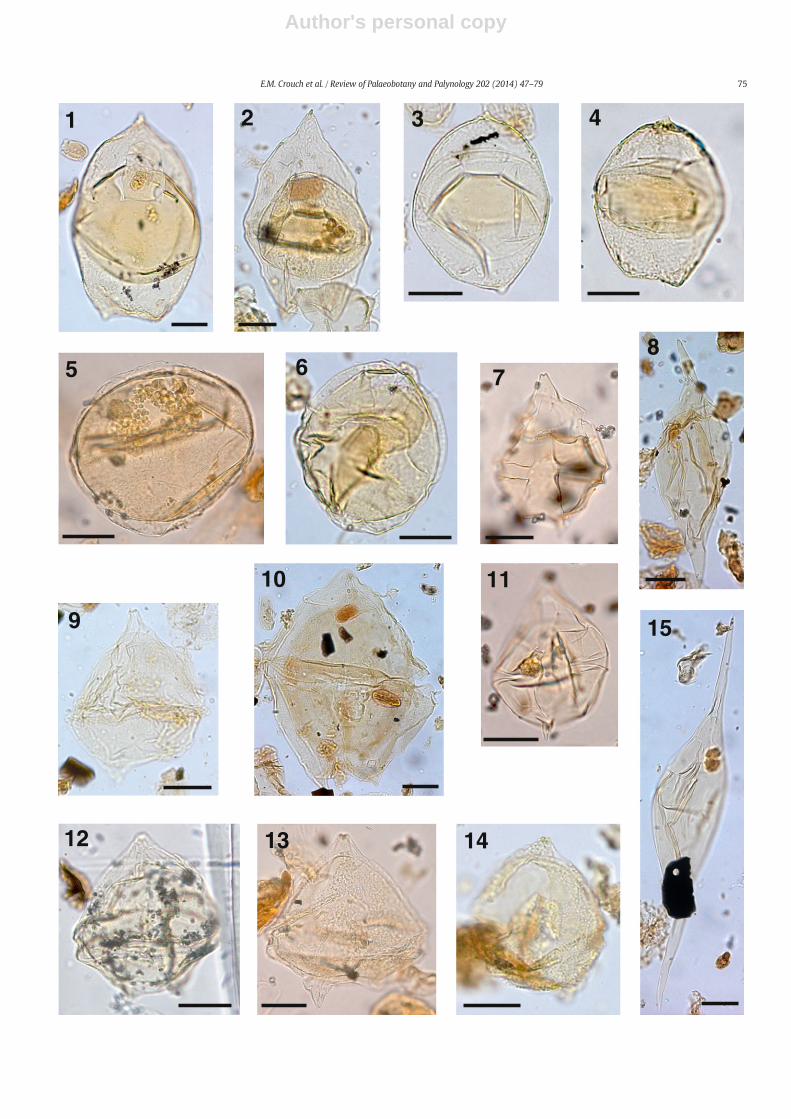
Author's personal copy
12
5
11
7
13
2
10
9
41
15
6
3
8
14
75E.M. Crouch et al. / Review of Palaeobotany and Palynology 202 (2014) 47–79
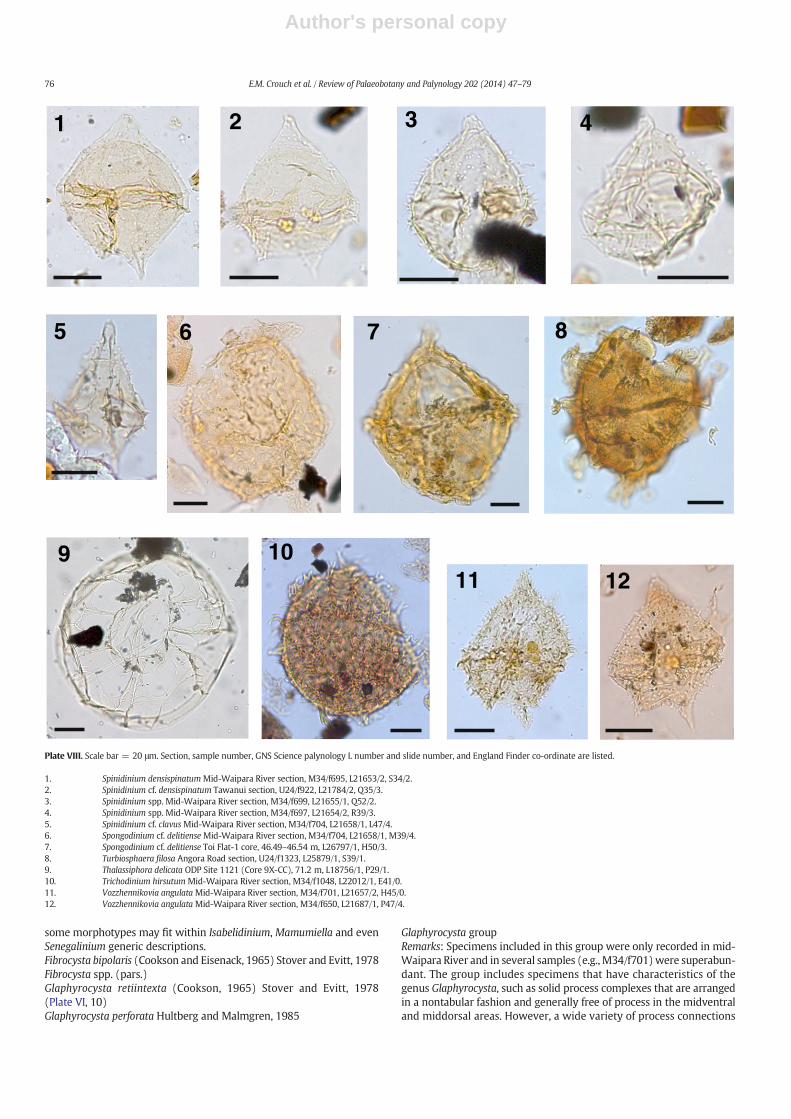
Author's personal copy
some morphotypes may fit within Isabelidinium, Mamumiella and evenSenegalinium generic descriptions.Fibrocysta bipolaris (Cookson and Eisenack, 1965) Stover and Evitt, 1978Fibrocysta spp. (pars.)Glaphyrocysta retiintexta (Cookson, 1965) Stover and Evitt, 1978(Plate VI, 10)Glaphyrocysta perforata Hultberg and Malmgren, 1985
Glaphyrocysta groupRemarks: Specimens included in this group were only recorded in mid-Waipara River and in several samples (e.g., M34/f701)were superabun-dant. The group includes specimens that have characteristics of thegenus Glaphyrocysta, such as solid process complexes that are arrangedin a nontabular fashion and generally free of process in the midventraland middorsal areas. However, a wide variety of process connections
7
12
31
5
11
4
8
9
2
6
10
Plate VIII. Scale bar = 20 μm. Section, sample number, GNS Science palynology L number and slide number, and England Finder co-ordinate are listed.
1. Spinidinium densispinatumMid-Waipara River section, M34/f695, L21653/2, S34/2.2. Spinidinium cf. densispinatum Tawanui section, U24/f922, L21784/2, Q35/3.3. Spinidinium spp. Mid-Waipara River section, M34/f699, L21655/1, Q52/2.4. Spinidinium spp. Mid-Waipara River section, M34/f697, L21654/2, R39/3.5. Spinidinium cf. clavusMid-Waipara River section, M34/f704, L21658/1, L47/4.6. Spongodinium cf. delitiense Mid-Waipara River section, M34/f704, L21658/1, M39/4.7. Spongodinium cf. delitiense Toi Flat-1 core, 46.49–46.54 m, L26797/1, H50/3.8. Turbiosphaera filosa Angora Road section, U24/f1323, L25879/1, S39/1.9. Thalassiphora delicata ODP Site 1121 (Core 9X-CC), 71.2 m, L18756/1, P29/1.10. Trichodinium hirsutum Mid-Waipara River section, M34/f1048, L22012/1, E41/0.11. Vozzhennikovia angulata Mid-Waipara River section, M34/f701, L21657/2, H45/0.12. Vozzhennikovia angulata Mid-Waipara River section, M34/f650, L21687/1, P47/4.
76 E.M. Crouch et al. / Review of Palaeobotany and Palynology 202 (2014) 47–79

Author's personal copy
were recognised, from singular processes to distal process ends thatranged from being connected in a simple manner to an intricate systemof trabeculae.Glaphyrocysta spp. (pars.)Hystrichokolpoma cf. cinctum Klump, 1953 (Plate VI, 9)Hystrichokolpoma spp. (pars.)Hystrichosphaeridium tubiferum (Ehrenberg, 1838) Deflandre, 1937;Davey and Williams, 1966Hystrichosphaeridium tubiferum subsp. brevispinum (Davey andWilliams, 1966) Lentin and Williams, 1973Impagidinium dispertitum (Cookson and Eisenack, 1965) Stover andEvitt, 1978Impagidinium maculatum (Cookson and Eisenack, 1961) Stover andEvitt, 1978Impagidinium cf. strialatum (Wall, 1967) Stover and Evitt, 1978Impagidinium sp. A (Plate VI, 11)Remarks: Specimenshave a thickened autophragmwith a granulate sur-face. Paratabulation is expressed by thickened fibrous ridges.Impagidinium spp. (pars.)Isabelidinium bakeri (Deflandre and Cookson, 1955) Lentin andWilliams, 1977 (Plate VII, 1, 2)Isabelidinium cingulatumWilson, 1988 (Plate IV, 11, 12)Remarks: This species has a clear paracingulum andmany examples likethe type specimens illustrated by Wilson (1988) are seen. Some formsgrade towards Isabelidinium bakeri but a clear paracingulum is still seen.Isabelidinium cf. pellucidum (Deflandre and Cookson, 1955) Lentin andWilliams, 1977 (Plate VII, 3, 4)Remarks: These specimens are smaller (length b90 μm, width b60 μm)than the size range given for Isabelidinium pellucidum. In this study,forms were observed that grade towards Isabelidinium bakeri. In addi-tion, I. pellucidum has commonly been recorded in the upper part ofthe Cretaceous from New Zealand and Australia (e.g., Helby et al.,1987; Roncaglia et al., 1999).Kallosphaeridium spp.Kenleyia spp.Lejeunecysta spp.Lingulodinium machaerophorum (Deflandre and Cookson, 1955) Wall,1967 (Plate VI, 12)Manumiella rotundaWilson, 1988 (Plate VII, 5, 6)Remarks: Specimens seen are very similar to the original description;however, themorphological variation is larger than originally described.The development of the periphragm can vary from bicavate tocircumcavate, while the length of the apical horn may vary. Develop-ment of the antapical prominence is minimal and the paracingulumcan be expressed.Manumiella spp. (pars.)Operculodinium tiara (Klumpp, 1953) Stover and Evitt, 1978Operculodinium spp. (pars.)Palaeocystodinium golzowense Alberti, 1961 (Plate VII, 15)Palaeocystodinium cf. australinum (Cookson, 1965) Lentin andWilliams,1976 (Plate VII, 8)Remarks: These specimens do not have an accessory horn or spike onthe antapical horn. The endophragm may be clear or dark brown incolour.Palaeoperidinium pyrophorum (Ehrenberg, 1838) Sarjeant, 1967 (PlateVII, 10)Remarks: At mid-Waipara River, Palaeoperidinium pyrophorum is fre-quent (N10%) in the lower part of the section, the abundance thendrops to b10%, and trace amounts are then seen prior to the HO. InColumn 4, the HCO (N10% of assemblage) is shown, along with theHO (Fig. 5). The HO of P. pyrophorum appears to be earlier in NewZealand than in Northwest Europe, offshore eastern Canada and pos-sibly Australia. In this study, the HO of P. pyrophorum is recorded incalcareous nannofossil Zone NP4 (Fig. 14) and correlated with the lowerTeurian (late Danian). In Northwest Europe, the HO of P. pyrophorum iscorrelated with Zone NP6 and lies close to the Selandian–Thanetian
boundary (Mudge and Bujak, 1996; Bujak and Brinkhuis, 1998; Schiøleret al., 2007), and in offshore eastern Canada the HO is recorded in theLate Selandian (Fensome et al., 2009). In the Gippsland Basin, Australia,the HO of P. pyrophorum is recorded just below the Danian–Selandianboundary (Partridge, 2004; Mantle et al., 2010), similar to this study,whereas at ODP Site 1172, the HO of P. pyrophorum lies close to theSelandian–Thanetian boundary (Bijl et al., 2013).Phelodinium spp.Phthanoperidinium? sp. A (Plate VII, 7, 11)Remarks: These specimens are tentatively placed in the genusPhthanoperidinium based on the presence of a clear paratabulation,a small antapical horn, reduced antapical horns and intercalary (2a)archeopyle. Specimens are thin-walled with a psilate autophragm, andthe parasutural lines are thin and not ornamented.Rottnestia borrusica (Eisenack, 1954) Cookson and Eisenack, 1961Senegalinium dilwynense (Cookson and Eisenack, 1965) Stover and Evitt,1978 (Plate VII, 9, 12)Senegalinium cf. dilwynense (Plate VII, 13, 14)Remarks: These specimens closely resemble Senegalinium dilwynense;however, the periphragm is granulate. This form was recognised inmid-Waipara River, Tawanui, Toi Flat-1 core and Angora Road.Senegalinium spp. (pars.)Remarks: Includes all other taxa assignable to the genus Senegalinium.Dinocyst preservation is variable and often it was only possible to assignspecimens to the genus level.Senoniasphaera inornata (Drugg, 1970) Stover and Evitt, 1978(Plate IV, 3, 6)Remarks: It appears that some forms grade towards the speciesGlaphyrocysta perforata Hultberg and Malmgren, 1985; specimensincluded as Senoniasphaera inornata in this study have a weaklyexpressed antapical lobe and some supporting processes in a proxi-mal position on the periphragm, but the periphragm does not havea ragged margin and is not strongly perforate. The HO of S. inornatain this study is in the uppermost part of calcareous nannofossilZone NP3, which correlates with the mid Danian and early Teurian.This HO is at a similar temporal level as that recorded in NorthwestEurope (Mudge and Bujak, 1996; Schiøler et al., 2007).Spinidinium cf. clavus (Harland, 1973) Sluijs et al., 2009 (PlateVIII, 5)Spinidinium essoi (Cookson and Eisenack, 1967) Sluijs et al., 2009Spinidinium densispinatum (Stanley, 1965) Sluijs et al., 2009 (PlateVIII, 1)Spinidinium cf. densispinatum (Plate VIII, 2)Remarks: These specimens resemble Spinidinium densispinatum in out-line; however, spinules are sparse on the periphragm. The cingulum isdiscernible. This form was most common in the Tawanui section.Spinidinium spp. (pars.) (Plate VIII, 3, 4)Remarks: Includes all other taxa assignable to the genus Spinidinium.Dinocyst preservation is variable and often it was only possible to assignspecimens to the genus level.Spiniferella cornutus (Gerlach, 1961) Sarjeant, 1970Spiniferites septatus (Cookson and Eisenack, 1967) McLean, 1971Spiniferites spp. (pars.)Spongodinium cf. delitiense (Plate VIII, 6, 7)Remarks: These specimens have a large cyst (length and widthN100 μm), with a precingular archeopyle and a broad apical horn.The ectophragm is partly perforate and has a finer appearancethan Spongodinium delitiense. The paracingulum is discernible.Thalassiphora delicata (Williams and Downie, 1966) Eaton, 1976 (PlateVIII, 9)Thalassiphora cf. velata (Deflandre and Cookson, 1955) Eisenack andGocht, 1960Trichodinium hirsutum Cookson, 1965 (Plate VIII, 10)Trithyrodinium evittii Drugg, 1967 (Plate IV, 1, 2)Remarks: In this study, the HO of Trithyrodinium evittii is observed in cal-careous nannofossil ZoneNP3 anduppermost radiolarian ZoneRP3. In the
77E.M. Crouch et al. / Review of Palaeobotany and Palynology 202 (2014) 47–79

Author's personal copy
GippslandBasin, Australia, theHOof T. evittii is also correlatedwith calcar-eous nannofossil Zone NP3 (Partridge, 2004).Turbiosphaera filosa Wilson, 1967 (Plate VIII, 8)Turbiosphaera galatea Eaton, 1976Turbiosphaera spp. (pars.)Vozzhennikovia angulata (Wilson, 1988) Sluijs et al., 2009 (Plate VIII,11, 12)Remarks: Many examples like the type forms illustrated by Wilson(1988) are observed in this study and high abundances are recorded.There can be a gradation towards forms that lookmore like Spinidiniumdensispinatum (Stanley, 1965) Sluijs et al., 2009.
Appendix B. Supplementary data
Supplementary data to this article can be found online at http://dx.doi.org/10.1016/j.revpalbo.2013.12.004.
References
Ballance, P.F., 1993. The paleo-Pacific, post-subduction, passive margin thermal relaxationsequence (Late Cretaceous–Paleogene) of the drifting New Zealand continent. In:Ballance, P.F. (Ed.), South Pacific Sedimentary Basins of the World 2. Elsevier SciencePublishers, pp. 93–110.
Beggs, J.M., Wilson, G.J., Raine, J.I., Pocknall, D.T., 1992. Biostratigraphic review of Creta-ceous and Early Cenozoic sections in Taranaki Basin exploration wells. DSIR Geologyand Geophysics Client Report 1992/38.
Bernaola, G., Baceta, J., Orue-Etxebarria, X., Alegret, L., Martin-Rubio, M., Arostegui, J.,Dinarès-Turell, J., 2007. Evidence of an abrupt environmental disruption during themid-Paleocene biotic event (Zumaia section, western Pyrenees). Geol. Soc. Am. Bull.199, 785–795.
Bijl, P.K., 2011. Environmental and Climatological Evolution of the early Paleogene South-ern Ocean. (PhD Thesis) LPP Contribution Series, 34. Utrecht University, Utrecht978-90-39356548.
Bijl, P.K., Schouten, S., Brinkhuis, H., Sluijs, A., Reichart, G.-J., Zachos, J., 2009. Early Paleo-gene temperature evolution of the Southwest Pacific ocean. Nature 461, 776–779.
Bijl, P.K., Sluijs, A., Brinkhuis, H., 2013. A magneto- and chemostratigraphically calibrateddinoflagellate cyst zonation of the early Paleogene South Pacific Ocean. Earth Sci. Rev.124, 1–31.
Bown, P.R. (Ed.), 1998. Calcareous Nannofossil Biostratigraphy. Cambridge Univ. Press,Cambridge.
Bown, P.R., 2005. Paleogene calcareous nannofossils from the Kilwa and Lindi areasof coastal Tanzania (Tanzania Drilling Project 2003-4). J. Nannoplankton Res.27, 21–95.
Browne, G.H., Field, B.D., 1985. The lithostratigraphy of Late Cretaceous to Early Pleisto-cene rocks or northern Canterbury, New Zealand. NZ Geol. Surv. Rec. 6.
Bujak, J.P., Brinkhuis, H., 1998. Global warming and dinocyst changes across the Paleo-cene/Eocene Epoch boundary. In: Aubry, M.P., Lucas, S., Berggren, W.A. (Eds.), LatePaleocene–Early Eocene Climatic and Biotic Events in the Marine and Terrestrial Re-cords. Colombia Univ Press, pp. 277–295.
Carter, R.M., et al. (Eds.), 1999. Proceedings of the Ocean Drilling Program, initial reports,volume 181. Southwest Pacific Gateways, Sites, pp. 1119–1125.
Clowes, C.D., 2009. Dinoflagellate Taxonomy and Biostratigraphy of the Mid- to Late Eo-cene and Early Oligocene of New Zealand. (PhD Thesis) Victoria University ofWellington, New Zealand.
Cook, R., Sutherland, R., Zhu, H., et al., 1999. Cretaceous–Cenozoic geology and petroleumsystems of the Great South Basin, New Zealand. Inst. Geol. Nucl. Sci. Ltd. Monogr. 20.
Cooper, R.A., 2004. The New Zealand Geological Timescale. Inst. Geol. Nucl. Sci. Ltd.Monogr. 22.
Cooper, R.A., Crampton, J.S., Raine, J.I., Gradstein, F.M., Morgans, H.E.G., Sadler, P.M.,Strong, C.P., Waghorn, D., Wilson, G.J., 2001. Quantitative biostratigraphy of the Tara-naki Basin, New Zealand: a deterministic and probabilistic approach. AAPG Bull. 85,1469–1498.
Crampton, J.S., Schiøler, P., Morgans, H.E.G., Raine, J.I., Roncaglia, L., Fohrmann, M., Reid, E.,2012. Quantitative stratigraphy of the Taranaki Basin. GNS Sci. Data Ser. 17a.
Crouch, E.M., 2001. Environmental Change at the Time of the Paleocene–Eocene BioticTurnover. (PhD thesis) LPP contribution series, 14. Utrecht University, Utrecht90-393-2828-5.
Crouch, E.M., Brinkhuis, H., 2005. Environmental change across the Paleocene–Eocenetransition from the eastern margin of New Zealand: a marine palynological approach.Mar. Micropaleontol. 56, 138–160.
Crouch, E.M., Heilmann-Clausen, C., Brinkhuis, H., Morgans, H.E.G., Rogers, K.M., Egger, H.,Schmitz, B., 2001. Global dinoflagellate event associated with the late Paleocene ther-mal maximum. Geology 29, 315–318.
Crouch, E.M., Brinkhuis, H., Visscher, H., Adatte, T., Bolle, M.-P., 2003. Late Paleocene–earlyEocene dinoflagellate cyst records from the Tethys: further observations on the globaldistribution of Apectodinium. Geol. Soc. Am. Spec. Pap. 369, 113–131.
Dale, B., 1996. Dinoflagellate cyst ecology: modelling and geological applications. In:Jansonius, J., McGregor, D.C. (Eds.), Palynology: Principles and Applications. AASPFoundation, pp. 1249–1275.
Edwards, A.R., 1971. A calcareous nannoplankton zonation of the New Zealand Paleogene.In: Farinacci, A. (Ed.), Proceedings of the II Planktonic Conference. EdizioniTecnoscienza, Rome, pp. 381–419.
Evitt, W.R., 1985. Sporopollenin Dinoflagellate Cysts: Their Morphology and Interpreta-tion. American Association of Stratigraphic Palynologists, Texas.
Fensome, R.A., Taylor, F.J.R., Norris, G., Sarjeant, W.A.S., Wharton, D.I., Williams, G.L., 1993.A classification of living and fossil dinoflagellates. Micropaleontol. Spec. Publ. 7.
Fensome, R.A., MacRae, R.A., Williams, G.L., 2008. DINOFLAG2. Version 1. American Asso-ciation of Stratigraphic Palynologists, Data Series No. 1.
Fensome, R.A., Williams, G.L., MacRae, R.A., 2009. Late Cretaceous and Cenozoic fossil di-noflagellates and other palynomorphs from the Scotian Margin, offshore easternCanada. J. Syst. Palaeontol. 7, 1–79.
Ferrow, E., Vajda, V., Koch, C.B., Peucker-Ehrenbrink, B., Willumsen, P.S., 2011. Multiproxyanalysis of a new terrestrial and a marine Cretaceous–Paleogene (K–Pg) boundarysite from New Zealand. Geochim. Cosmochim. Acta 75, 657–672.
Gradstein, F.M., Ogg, J., Schmitz, M., Ogg, G., 2012. The Geologic Time Scale 2012. ElsevierBV.
Handley, L., Crouch, E.M., Pancost, R.D., 2011. A New Zealand record of sea level rise andenvironmental change during the Paleocene–Eocene Thermal Maximum.Palaeogeogr. Palaeoclimatol. Palaeoecol. 305, 185–200.
Harland, R., 1979. Agerasphaera nov. gen., and ‘Eisenackia’ like dinoflagellate cysts fromthe Thanet Sands (Paleocene) of southeast England. Rev. Palaeobot. Palynol. 28,27–35.
Heilmann-Clausen, C., 1985. Dinoflagellate stratigraphy of the upper-most Danian toYpresian in the Viborg-1 borehole, central Jylland, Denmark. Dan. Geol. Unders. Ser.A 7.
Helby, R., Morgan, R., Partridge, A., 1987. A palynological zonation of the Australian Meso-zoic. Mem. Assoc. Australas. Palaeontol. 4, 1–94.
Hollis, C.J., 1993. Latest Cretaceous to Late Paleocene radiolarian biostratigraphy: a newzonation from the New Zealand region. Mar. Micropaleontol. 21, 295–327.
Hollis, C.J., 2002. Biostratigraphy and paleoceanographic significance of Paleoceneradiolarians from offshore eastern New Zealand. Mar. Micropaleontol. 46,265–316.
Hollis, C.J., Strong, C.P., 2003. Biostratigraphic review of the Cretaceous/Tertiary boundarytransition, mid-Waipara River section, North Canterbury, New Zealand. N. Z. J. Geol.Geophys. 46, 243–253.
Hollis, C.J., Rodgers, K.A., Parker, R.J., 1995. Siliceous plankton bloom in the earliest Tertia-ry of Marlborough, New Zealand. Geology 23, 835–838.
Hollis, C.J., Rodgers, K.A., Strong, C.P., Field, B.D., Rogers, K.M., 2003. Paleoenvironmentalchanges across the Cretaceous/Tertiary boundary in the northern Clarence Valley,southeastern Marlborough, New Zealand. N. Z. J. Geol. Geophys. 46, 209–234.
Hollis, C.J., Handley, L., Crouch, E.M., Morgans, H.E.G., Baker, J., Creech, J., Collins, K.S.,Gibbs, S., Huber, M., Schouten, S., Zachos, J.C., Pancost, R.D., 2009. Tropical sea temper-atures in the high-latitude South Pacific. Geology 37, 99–102.
Hollis, C.J., Taylor, K.W.R., Handley, L., Pancost, R.D., Huber, M., Creech, J.B., Hines, B.,Crouch, E.M., Morgans, H.E.G., Crampton, J.S., Gibbs, S., Pearson, P.N., Zachos, J.C.,2012. Early Paleogene temperature history of the Southwest Pacific Ocean: reconcil-ing proxies and models. Earth Planet. Sci. Lett. 349–350, 53–66.
Hollis, C.J., Tayler, M., Andrew, B., Lurcock, P., Creech, J.B., Kulhanek, D., Crouch, E.M.,Strong, C.P., Nelson, C., Pancost, R.D., Wilson, G., Crampton, J.S., Schiøler, P., 2014.Organic-rich sedimentation in the southwest Pacific Ocean associated with Late Pa-leocene climatic cooling. Earth Sci. Rev. (in review).
Hornibrook, N.de B., Edwards, A.R., 1971. Integrated planktonic foraminiferal and calcar-eous nannoplankton datum levels in the New Zealand Cenozoic. In: Farinacci, A.(Ed.), Proceedings of the II Planktonic Conference, Roma. Edizioni Tecnoscienza,Rome, pp. 649–657.
Hornibrook, N.de B., Harrington, H.J., 1957. The status of the Wangaloan Stage. N. Z. J. Sci.Technol. 38, 655–670.
Hornibrook, N.de B., Brazier, R., Strong, C.P., 1989. Manual of New Zealand Permian toPleistocene formainiferal biostratigraphy. N. Z. Geol. Surv. Paleontol. Bull. 56.
Jenkins, D.G., 1966. Planktonic foraminiferal zones and new taxa from the Danian toLower Miocene of New Zealand. N. Z. J. Geol. Geophys. 8, 1088–1126.
Jenkins, D.G., 1971. New Zealand Cenozoic planktonic foraminifera. N. Z. Geol. Surv.Paleontol. Bull. 42.
Killops, S.D., Hollis, C.J., Morgans, H.E.G., Sutherland, R., Field, B.D., Leckie, D.A., 2000.Paleoceanographic significance of Late Paleocene dysaerobia at the shelf/slopebreak around New Zealand. Palaeogeogr. Palaeoclimatol. Palaeoecol. 156, 51–70.
King, P., Naish, T., Browne, G., Field, B., Edbrooke, S., 1999. Cretaceous to Recent sed-imentary patterns in New Zealand. Inst. Geol. Nucl. Sci. Folio Ser. 1 (version1999.1).
Kofoid, C.A., 1909. On Peridinium steini Jörgensen, with a note on the nomenclature of theskeleton of the Peridinidae. Arch. Protistenkd. 16, 25–47.
Kulhanek, D.K., Tayler, M.J.S., Hollis, C.J., Nelson, C.S., 2011. Revised age of the WaipawaFormation based on calcareous nannofossils. Geol. Soc. NZ Misc. Publ. 130A, 63.
Laing, A.C.M., 1991. Weber Stratigraphic Drilling Report PPL 38317. Petroleum Report Se-ries PR1676Ministry of Economic Development, New Zealand.
Lurcock, P., 2011. Paleomagnetism of Paleogene Strata from Southern Zealandia: Implica-tions for Ice in the Greenhouse. (PhD Thesis) University of Otago, New Zealand.
Mantle, D.J., Kelman, A.P., Nicoll, R.S., Laurie, J.R., 2010. Australian Biozonation Chart 2010.Geoscience Australia Publication Chart.
Martini, E., 1971. Standard Tertiary and Quaternary calcareous nannoplankton zonation.In: Farinacci, A. (Ed.), Proceedings of the II Planktonic Conference, Roma. EdizioniTecnoscienza, Rome, pp. 739–785.
McInerney, F.A., Wing, S.L., 2011. The Paleocene–Eocene Thermal Maximum: a perturba-tion of carbon cycle, climate and biosphere with implications for the future. Ann. Rev.Earth Planet. Sci. 2011 (39), 489–516.
78 E.M. Crouch et al. / Review of Palaeobotany and Palynology 202 (2014) 47–79

Author's personal copy
McMillan, S.G.,Wilson, G.J., 1997. Allostratigraphy of coastal south and east Otago: a strat-igraphic framework for interpretation of the Great South Basin, New Zealand. N. Z.J. Geol. Geophys. 40, 91–107.
Moore, P.R., 1988. Stratigraphy, composition and environment of deposition of theWhangai Formation and associated Late Cretaceous–Paleocene rocks, eastern NorthIsland, New Zealand. N. Z. Geol. Surv. Bull. 100.
Moore, P.R., 1989. Stratigraphy of theWaipawa Black Shale (Paleocene), eastern North Is-land, New Zealand. NZ Geol. Surv. Rec. 38.
Moore, P.R., Morgans, H.E.G., 1987. Two new reference sections from the Wanstead For-mation (Paleocene–Eocene) in southern Hawkes Bay. NZ Geol. Surv. Rec. 20, 81–87.
Moore, P.R., Snowdon, L.R., Osadetz, K.G., 1987. Maturation and petroleum source rockpotential of the Whangai and Waipawa Formations (Late Cretaceous–Paleocene),Eastern North Island, New Zealand. NZ Geol. Surv. Rec. 20, 17–23.
Morgans, H.E.G., 2001. Sample collections (1982–2000) from Tawanui Section, AkitioRiver, Southern Hawkes Bay. Institute of Geological and Nuclear Sciences Limited Sci-ence Report 2001/5.
Morgans, H.E.G., Beu, A.G., Cooper, R.A., Crouch, E.M., Hollis, C.J., Jones, C.M., Raine, J.I.,Strong, C.P., Wilson, G.J., Wilson, G.S., 2004. Chapter 11, Paleogene (Dannevirke, Ar-nold and Landon Series). In: Cooper, R.A. (Ed.), The New Zealand Geological Time-scale. Institute of Geological and Nuclear Sciences Monograph, 22, pp. 125–163.
Morgans, H.E.G., Jones, C.M., Crouch, E.M., Field, B.D., Hollis, C.J., Raine, J.I., Strong, C.P.,Wilson, G.J., 2005. Upper Cretaceous to Eocene stratigraphy and sample collections,mid-Waipara River section, North Canterbury. Institute of Geological and Nuclear Sci-ences Limited Science Report 2003/08.
Mudge, D.C., Bujak, J.P., 1996. Paleocene biostratigraphy and sequence stratigraphy of theUK central North Sea. Mar. Pet. Geol. 11, 166–181.
Partridge, A.D., 2001. Revised stratigraphy of the Sherbrook Group, Otway Basin. PESAEastern Australasian Basins Symposium, pp. 455–464.
Partridge, A.D., 2004. Late Cretaceous–Cenozoic Palynology Zonations, Gippsland Basin.Geosciences Australia publication1-921236051 (chart 4 of 4).
Perch-Nielsen, K., 1985. Cenozoic calcareous nannofossils. In: Bolli, H., Perch-Nielsen, K.,Saunders, J.B. (Eds.), Plankton Stratigraphy. Cambridge University Press, NY,pp. 427–554.
Petrizzo, M., 2005. An early late Paleocene event on Shatsky Rise, northwest Pacific Ocean(ODP leg 198), evidence from planktonic foraminiferal assemblages. Proc. Ocean Drill.Program Sci. Results 198. http://dx.doi.org/10.2973/odp.proc.sr.198.110.2005 (29 pp.).
Powell, A.J., 1988. A modified dinoflagellate biozonation for the latest Palaeocene and ear-liest Eocene sediments from the central North Sea. Rev. Palaeobot. Palynol. 56,327–344.
Powell, A.J., Brinkhuis, H., Bujak, J.P., 1996. Upper Paleocene–Lower Eocene dinoflagellatecyst sequence biostratigraphy of southeast England. In: Knox, R., et al. (Eds.), Correla-tion of the Early Paleogene in Northwest Europe. Geological Society Special Publica-tion, 101, pp. 145–183.
Quattrocchio, M.E., Sarjeant, W.A.S., 2003. Dinoflagellates from the Chorrillo Chico Forma-tion (Paleocene) of southern Chile. Ameghiniana 40, 129–153.
Raine, J.I., Beu, A.G., Boyes, A.F., Campbell, H.J., Cooper, R.A., Crampton, J.S., Crundwell,M.P., Hollis, C.J., Morgans, H.E.G., 2013. New Zealand Geological Timescale v. 2012/1. Available at www.gns.cri.nz/Home/Our-Science/Earth-Science/Fossils/Online-Resources/New-Zealand-s-Geological-Timescale (NZGT2012_Wall_chart.pdf).
Rogers, K.M., Morgans, H.E.G., Wilson, G.S., 2001. Identification of a Waipawa Formationequivalent in the upper Te Uri Member of the Whangai Formation — implicationsfor depositional history and age. N. Z. J. Geol. Geophys. 44, 347–354.
Röhl, U., Brinkhuis, H., Sluijs, A., Fuller, M., 2004. On the search for the Paleocene/Eoceneboundary in the Southern Ocean: exploring ODP Leg 189 Holes 1171D and 1172D,Tasman Sea. Geophys. Monogr. Ser. 151, 113–124.
Roncaglia, L., Field, B.D., Raine, J.I., Schiøler, P., Wilson, G.J., 1999. Dinoflagellate biostratig-raphy of Piripauan–Haumurian (Upper Cretaceous) section from northeast South Is-land, New Zealand. Cretac. Res. 20, 271–314.
Schiøler, P., Roncaglia, L., 2007. Great South Basin and East Coast hydrocarbon potential:the Tartan and Waipawa Formations as age and facies equivalent source rocks.Geol. Soc. NZ Misc. Publ. 123A, 154.
Schiøler, P., Wilson, G.J., 1998. Dinoflagellate biostratigraphy of the middle Coniacian–lower Campanian (Upper Cretaceous) in south Marlborough, New Zealand. Micropa-leontology 44, 313–349.
Schiøler, P., Andsbjerg, J., Clausen, O.R., Dam, G., Dybkjaer, K., Hamberg, L., Heilmann-Clausen, C., Johannessen, E.P., Kristensen, L.E., Prince, I., Rasmussen, J.A., 2007. Litho-stratigraphy of the Palaeogene–Lower Neogene succession of the Danish North Sea.Geol. Surv. Den. Greenl. Bull. 12.
Schiøler, P., Rogers, K., Sykes, R., Hollis, C.J., Ilg, B., Meadows, D., Roncaglia, L., Uruski, C.,2010. Palynofacies, organic geochemistry and depositional environment of the TartanFormation (Late Paleocene), a potential source rock in the Great South Basin, NewZealand. Mar. Pet. Geol. 27, 351–369.
Schmitz, B., Pujalte, V., Molina, E., Monechi, S., Orue-Etxebarria, Speijer, R., et al., 2011. Theglobal stratotype sections and points for the bases of the Selandian (Middle Paleocene)and Thanetian (Upper Paleocene) stages at Zumaia, Spain. Episodes 34, 220–243.
Sluijs, A., Pross, J., Brinkhuis, H., 2005. From greenhouse to icehouse; organic-walled dino-flagellate cysts as paleoenvironmental indicators in the Paleogene. Earth-Sci. Rev. 68,281–315.
Sluijs, A., Bowen, G., Brinkhuis, H., Lourens, L., Thomas, E., 2007. The Paleocene–EoceneThermal Maximum super greenhouse: biotic and geochemical signatures, agemodelsand mechanisms of global change. In: Williams, M., et al. (Eds.), Deep–Time Perspec-tives on Climate Change: Marrying the Signal from Computer Models and BiologicalProxies, pp. 323–349.
Sluijs, A., Brinkhuis, H., Crouch, E.M., John, C.M., Handley, L., Munstermann, D.,Boharty, S.M., Zachos, J.C., Reichart, G.J., Schouten, S., Pancost, R.D., SinningheDamsté, J.S., Welters, N., Lotter, A., Dicken, G.R., 2008. Eustatic variations duringthe Paleocene–Eocene greenhouse world. Paleoceanography 23. http://dx.doi.org/10.1029/2008PA001615.
Sluijs, A., Brinkhuis, H., Williams, G.L., Fensome, R.A., 2009. Taxonomic revision of someCretaceous–Cenozoic spiny organic-walled peridiniacean dinoflagellate cysts. Rev.Palaeobot. Palynol. 154, 34–53.
Sluijs, A., Bijl, P., Schouten, S., Röhl, U., Reichart, G.J., Brinkhuis, H., 2011. Southern Oceanwarming and hydrological change during the Paleocene–Eocene Thermal Maximum.Clim. Past 7, 47–61.
Strong, C.P., Hollis, C.J., Wilson, G.J., 1995. Foraminiferal, radiolarian, and dinoflagel-late biostratigraphy of Late Cretaceous to middle Eocene pelagic sediments(Muzzle group), Mead Stream, Marlborough, New Zealand. N. Z. J. Geol. Geophys.38, 171–209.
Strong, C.P., Mildenhall, D.C., Raine, J.I., Wilson, G.J., Edwards, A.R., 1999. Biostratigraphy ofWaka Nui-1 offshore petroleum exploration well, Northland Basin, New Zealand. In-stitute of Geological and Nuclear Sciences Limited Client Report 1999/123.
Sutherland, R., King, P., 2008. Leaving Gondwana behind. In: Graham, I.J. (Ed.), AContinent on the Move: New Zealand Geoscience into the 21st Century,pp. 128–129.
Tayler, M.J.S., 2011. Investigating Stratigraphic Evidence for Antarctic Glaciation in theGreenhouseWorld of the Paleocene, Eastern North Island, New Zealand. (MSc Thesis)University of Waikato, New Zealand.
Vajda, V., Raine, J.I., 2003. Pollen and spores in marine Cretaceous/Tertiary boundary sed-iments at mid-Waipara River, North Canterbury, New Zealand. N. Z. J. Geol. Geophys.46, 255–273.
Wei, K., Mii, H., Shu, I., Lin, Y., 2005. Uppermost Cretaceous to middle Oligocene carbonand oxygen isotope stratigraphy of southwest Pacific: Holes 1121B and 1124C, ODPLeg 181. N. Z. J. Geol. Geophys. 48, 15–26.
Westerhold, T., Röhl, U., Donner, B., McCarren, H., Zachos, J.C., 2011. A complete high-resolution Paleocene benthis stable isotope record for the central Pacific (ODP Site1209). Paleoceanography 26, PA2216. http://dx.doi.org/10.1029/2010PA002092.
Willumsen, P.S., 2000. Late Cretaceous to early Paleocene palynological changes in mid-latitude Southern Hemisphere, New Zealand. In: Schmitz, B., et al. (Eds.), Early Paleo-gene Warm Climates and Biosphere Dynamics. GFF, 122, pp. 180–181.
Willumsen, P.S., 2003. Marine Palynology Across the Cretaceous–Tertiary Boundary inNew Zealand. (PhD Thesis) Victoria University of Wellington, New Zealand.
Willumsen, P.S., 2004. Two new species of the dinoflagellate cyst genus CarpatellaGrigorovich 1969 from the Cretaceous–Tertiary transition in New Zealand.J. Micropalaeontol. 23, 119–125.
Willumsen, P.S., 2006. Palynodinium minus sp. nov., a new dinoflagellate cyst from theCretaceous–Paleogene transition in New Zealand; its significance and palaeoecology.Cretac. Res. 27, 954–963.
Willumsen, P.S., 2011. Maastrichtian to Paleocene dinocysts from the Clarence Valley,South Island, New Zealand. Alcheringa 35, 199–240.
Willumsen, P.S., 2012. Three new species of dinoflagellate cyst from the Cretaceous–Paleogene sections at mid-Waipara River and Farifield Quarry, South Island, NewZealand. Palynology 36, 48–62.
Willumsen, P.S., Vajda, V., 2010. A new early Paleocene dinoflagellate cyst species,Trithyrodinium partridgei: its biogeographic significance and palaeoecology.Alcheringa 34, 523–538.
Wilson, G.J., 1967. Some species ofWetzeliella Eisenack (Dinophyceae) from New ZealandEocene and Paleocene strata. N. Z. J. Bot. 5, 469–497.
Wilson, G.J., 1978. The dinoflagellate species Isabelia druggii (Stover) and I. seelandica(Lange): their association in the Teurian of Woodside Creek, Marlborough, NewZealand. N. Z. J. Geol. Geophys. 21, 75–80.
Wilson, G.J., 1982. Eocene and Oligocene dinoflagellate assemblages from the Oamaruarea, North Otago, New Zealand. NZ Geol. Surv. Rep. PAL 60, 1–20.
Wilson, G.J., 1984. New Zealand Late Jurassic to Eocene dinoflagellate biostratigraphy — asummary. Newsl. Stratigr. 13, 104–117.
Wilson, G.J., 1987. Dinoflagellate biostratigraphy of the Cretaceous–Tertiary boundary,mid-Waipara River section, North Canterbury. NZ Geol. Surv. Rec. 20, 8–15.
Wilson, G.J., 1988. Paleocene and Eocene dinoflagellate cysts fromWaipawa, Hawkes Bay,New Zealand. N. Z. Geol. Surv. Paleontol. Bull. 57.
Wilson, G.J., McMillan, S.G., 1996. Late Cretaceous–Tertiary stratigraphic sections ofcoastal Otago, South Island, New Zealand: a summary of biostratigaphic andlithostratigraphic data. Institute of Geological and Nuclear Sciences Limited Sci-ence Report 96/39.
Zachos, J.C., Pagani, M., Sloan, L., Thomas, E., Billups, K., 2001. Trends, rhythms, andabberations in global climate 65 Ma to present. Science 292, 686–693.
Zachos, J.C., Dickens, G.R., Zeebe, R.E., 2008. An early Cenozoic perspective on greenhousewarming and carbon-cycle dynamics. Nature 451, 279–283.
79E.M. Crouch et al. / Review of Palaeobotany and Palynology 202 (2014) 47–79