human periapical cyst mesenchymal stem cells differentiate into neuronal cells
TRANSCRIPT

Journal of Dental Research 1 –10© International & American Associations for Dental Research 2015Reprints and permissions: sagepub.com/journalsPermissions.navDOI: 10.1177/0022034515570316jdr.sagepub.com
Research Reports: Biological
IntroductionAlthough human neural stem cells and human embryonic stem cells could be used for cell therapies to treat neurodegenerative disorders, such as Parkinson and Alzheimer diseases, several technical and ethical difficulties have limited their use in human therapies (Martino and Pluchino 2006; Lees et al. 2009). Therefore, the improvement of different therapeutic strategies for the treatment of neurologic disease using adult stem cells, such as mesenchymal stem cells (MSCs), remains a practical alternative (Chamberlain et al. 2007; Uccelli et al. 2011). MSCs are multipotent stem cells that are capable of dif-ferentiating into multiple cell types (Herzog et al. 2003; Keating 2012). The in vitro growth of undifferentiated MSCs, followed by induction of neural cell differentiation and subse-quent transplantation, is one of the most relevant methods for cell therapy to treat neurodegenerative disease (Martino et al. 2010; Maltman et al. 2011).
Although MSCs can be derived from all postnatal tissues, human bone marrow (BM) has been used as the major source of MSCs to treat neurodegenerative disease (Pal et al. 2009; Yan K et al. 2013). However, there are some limitations to the clinical use of BM-MSCs; namely, cell harvesting requires an
invasive method of isolation, and the cells are not abundant. Recently, different laboratories have described the isolation of human MSCs with neural plasticity from many tissues, includ-ing umbilical cold matrix (Bieback et al. 2004), amniotic fluid (Yan ZJ et al. 2013), adipose tissue (Zuk et al. 2002), and den-tal pulp (hDP; Arthur et al. 2008).
Dental-derived MSCs, particularly dental pulp stem cells (DPSCs), have become an attractive source of MSCs because of their abundant availability and faster proliferation as com-pared with BM-MSCs (Gronthos et al. 2000). Dental-derived MSCs are related to BM-MSCs and are easily accessible, mak-ing them an interesting alternative for cell therapies (Morsczeck
570316 JDRXXX10.1177/0022034515570316Journal of Dental ResearchHuman Periapical Cyst–Mesenchymal Stem Cellsresearch-article2015
1Unit of Maxillofacial Surgery, Calabrodental, Crotone, Italy2Tecnologica Research Institute, Biomedical Section, Crotone, Italy*All authors equally contributed to this article.
A supplemental appendix to this article is published electronically only at http://jdr.sagepub.com/supplemental.
Corresponding Author:M. Tatullo, Scientific Director, Tecnologica Research Institute, St. E. Fermi, Crotone, Italy. Email: [email protected]
Human Periapical Cyst–Mesenchymal Stem Cells Differentiate Into Neuronal Cells
M. Marrelli1*, F. Paduano2*, and M. Tatullo2*
AbstractIt was recently reported that human periapical cysts (hPCys), a commonly occurring odontogenic cystic lesion of inflammatory origin, contain mesenchymal stem cells (MSCs) with the capacity for self-renewal and multilineage differentiation. In this study, periapical inflammatory cysts were compared with dental pulp to determine whether this tissue may be an alternative accessible tissue source of MSCs that retain the potential for neurogenic differentiation. Flow cytometry and immunofluorescence analysis indicated that hPCy-MSCs and dental pulp stem cells spontaneously expressed the neuron-specific protein β-III tubulin and the neural stem-/astrocyte-specific protein glial fibrillary acidic protein (GFAP) in their basal state before differentiation occurs. Furthermore, undifferentiated hPCy-MSCs showed a higher expression of transcripts for neuronal markers (β-III tubulin, NF-M, MAP2) and neural-related transcription factors (MSX-1, Foxa2, En-1) as compared with dental pulp stem cells. After exposure to neurogenic differentiation conditions (neural media containing epidermal growth factor [EGF], basic fibroblast growth factor [bFGF], and retinoic acid), the hPCy-MSCs showed enhanced expression of β-III tubulin and GFAP proteins, as well as increased expression of neurofilaments medium, neurofilaments heavy, and neuron-specific enolase at the transcript level. In addition, neurally differentiated hPCy-MSCs showed upregulated expression of the neural transcription factors Pitx3, Foxa2, Nurr1, and the dopamine-related genes tyrosine hydroxylase and dopamine transporter. The present study demonstrated for the first time that hPCy-MSCs have a predisposition toward the neural phenotype that is increased when exposed to neural differentiation cues, based on upregulation of a comprehensive set of proteins and genes that define neuronal cells. In conclusion, these results provide evidence that hPCy-MSCs might be another optimal source of neural/glial cells for cell-based therapies to treat neurologic diseases.
Keywords: dental pulp, periapical tissue, radicular cyst, regenerative medicine, stem cell research, neurogenesis
by guest on February 12, 2015 For personal use only. No other uses without permission.jdr.sagepub.comDownloaded from
© International & American Associations for Dental Research 2015

2 Journal of Dental Research
et al. 2008). hDP tissue is derived from neural crest cells during development (Peters and Balling, 1999) and contains multipo-tent stem progenitor cells. Several studies showed that human DPSCs and stem cells derived from exfoliated deciduous teeth are capable of differentiating into neurons and express neuro-nal markers when cultured in the appropriate media (Miura et al. 2003; Nosrat et al. 2004). In addition, in vivo studies dem-onstrated that DPSCs differentiate into active neurons under appropriate environmental cues (Arthur et al. 2008). However, dental-derived MSCs are not always available when needed. These multipotent MSCs are present in higher numbers in young individuals and are accessible mainly after extraction of unerupted third molar teeth. Therefore, additional easily avail-able tissue sources are desired. For this purpose, other tissues containing MSCs that may have neurogenic potential and that are easily obtained from the body are the inflamed periapical tissues and the inflamed periodontal ligament (Liao et al. 2011; Park et al. 2011). MSCs obtained from these pathologic dental tissues have numerous desirable properties, such as prolifera-tive and regenerative potential, that are similar to MSCs derived from healthy tissues. In fact, periapical inflammatory-derived cells (Liao et al. 2011), MSCs derived from periapical lesions (Dokic et al. 2012) and inflamed periodontal ligament stem cells (Park et al. 2011) have been shown to exhibit an MSC immunophenotype and the ability to differentiate toward osteogenic and adipogenic lineages.
The isolation and characterization of MSCs from human periapical cysts (hPCys) were recently described (Marrelli et al. 2013). These tissues are considered to be the most prevalent odontogenic cystic lesions of inflammatory origin and are located at the root apices of involved teeth.
hPCy-MSCs are highly proliferative and express antigens that are characteristic of MSCs. Furthermore, hPCy-MSCs are multipotent cells that are capable of differentiating into osteo-blasts and adipocytes (Marrelli et al. 2013). To evaluate the use of hPCy-MSCs in stem cell–based therapies for neurodegen-erative diseases, it is critical to determine whether these stem cells constitutively express neural markers. In the present investigation, quantitative reverse transcription polymerase chain reaction (qRT-PCR), flow cytometry, and immunofluo-rescence were used to characterize the basal expression levels of neuronal and glial markers in undifferentiated hPCy-MSCs, and undifferentiated DPSCs were used as a control. The evi-dence supporting the neural crest origin of DPSCs makes them ideal cells for comparison with hPCy-MSCs in terms of neural differentiation potential. Subsequently, the ability of hPCy-MSCs to differentiate into neurons was evaluated in neural dif-ferentiation medium. It was confirmed that hPCys contain
MSCs with a neurogenic potential similar to that of DPSCs. The present report is the first describing the production of neu-ronal cells from hPCy-MSCs by inducing the upregulation of neural markers and neural-related transcription factors. Therefore, because the neurogenic potential of hPCy-MSCs is similar to that of DPSCs, we propose a simple method for hPCy-MSCs’ differentiation into neural-like cells. Moreover, this study proposes that hPCys may serve as a new reservoir of MSCs for cell replacement therapies to treat several human diseases involving neural cell loss.
Materials and MethodsSee the Appendix.
Results
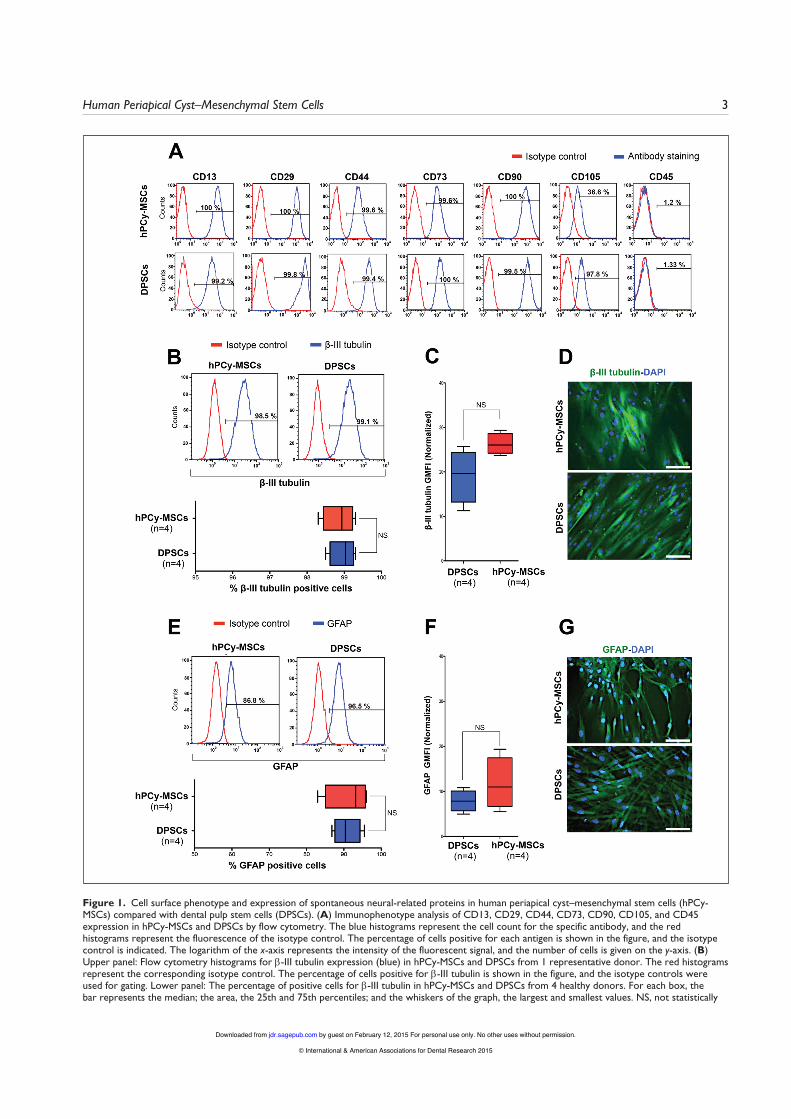
Isolation and Immunophenotypic Characterization of hPCy-MSCsWe established different cultures of MSCs isolated from hPCy and hDP, which were used as a control, according to the proper-ties of MSCs (Dominici et al. 2006; Calloni et al. 2013). The isolated cells displayed a fibroblast-like morphology and had the potential to differentiate into osteoblasts and adipocytes (Appendix Fig. 1A). The immunophenotype profiles of hPCy-MSCs and DPSCs included very high expression of the mesen-chymal surface antigens (Fig. 1A, Appendix Fig. 1B). The hematopoietic cell surface antigen CD45 was negative. Both types of MSCs had similar expression levels of CD13, CD29, CD44, CD73, and CD90, whereas CD105 expression was observed in 36.6% and 97.8% of the hPCy-MSCs and DPSCs, respectively.
Neural Marker Expression by Undifferentiated hPCy-MSCs in Normal Culture Conditions
To investigate the basal expression levels of neural-specific proteins and to evaluate the potential of hPCy-MSCs to dif-ferentiate toward the neural lineage, the cells were analyzed by flow cytometry and immunofluorescence. β-III tubulin is widely regarded as a neuronal marker in developmental neu-robiology and stem cell research. Glial fibrillary acidic protein (GFAP) is the primary component of intermediate filaments in astrocytes and provides a cell-specific marker to identify dif-ferentiated astrocytes from other glial cells (Sakai et al. 2012). To test the specificity of these markers, we evaluated their
significant. Flow cytometry data were analyzed using FlowJo software. (C) Expression levels of β-III tubulin in hPCy-MSCs and DPSCs from 4 healthy donors. The β-III tubulin expression levels were normalized by dividing the geometric mean fluorescence intensity (GMFI) of the experimental stains by that of the isotype control stains. (D) Representative immunofluorescence images (green) of undifferentiated hPCy-MSCs and DPSCs stained for β-III tubulin. The cell nuclei were stained with DAPI (blue). Scale bar = 100 μm. (E) Upper panel: Flow cytometry histograms for glial fibrillary acidic protein (GFAP) (blue) in hPCy-MSCs and DPSCs from 1 representative patient. The red histograms represent the corresponding isotype controls. The percentage of cells positive for GFAP is shown in the figure, and the isotype controls were used for gating. Lower panel: Percentage of cells positive for GFAP in hPCy-MSCs and DPSCs from 4 donors. (F) Box plots showing the expression of GFAP in hPCy-MSCs and DPSCs from 4 healthy donors. The results are presented as the GMFI of cells expressing GFAP normalized to the isotype control. (G) Representative immunofluorescence images (green) of undifferentiated hPCy-MSCs and DPSCs stained for GFAP. The cell nuclei were stained with DAPI (blue). Scale bar = 100 μm.
by guest on February 12, 2015 For personal use only. No other uses without permission.jdr.sagepub.comDownloaded from
© International & American Associations for Dental Research 2015
Human Periapical Cyst–Mesenchymal Stem Cells 3
Figure 1. Cell surface phenotype and expression of spontaneous neural-related proteins in human periapical cyst–mesenchymal stem cells (hPCy-MSCs) compared with dental pulp stem cells (DPSCs). (A) Immunophenotype analysis of CD13, CD29, CD44, CD73, CD90, CD105, and CD45 expression in hPCy-MSCs and DPSCs by flow cytometry. The blue histograms represent the cell count for the specific antibody, and the red histograms represent the fluorescence of the isotype control. The percentage of cells positive for each antigen is shown in the figure, and the isotype control is indicated. The logarithm of the x-axis represents the intensity of the fluorescent signal, and the number of cells is given on the y-axis. (B) Upper panel: Flow cytometry histograms for β-III tubulin expression (blue) in hPCy-MSCs and DPSCs from 1 representative donor. The red histograms represent the corresponding isotype control. The percentage of cells positive for β-III tubulin is shown in the figure, and the isotype controls were used for gating. Lower panel: The percentage of positive cells for β-III tubulin in hPCy-MSCs and DPSCs from 4 healthy donors. For each box, the bar represents the median; the area, the 25th and 75th percentiles; and the whiskers of the graph, the largest and smallest values. NS, not statistically
by guest on February 12, 2015 For personal use only. No other uses without permission.jdr.sagepub.comDownloaded from
© International & American Associations for Dental Research 2015
4 Journal of Dental Research
expression and distribution in undifferentiated hPCy-MSCs compared with DPSCs. The data revealed that both MSCs were strongly positive for β-III tubulin before induction (98.5% and 99.1%; Fig. 1B, upper panel) and were also posi-tive for GFAP (86.8 and 96.5%; Fig. 1E, upper panel). No significant differences were detected in the percentages of β-III tubulin–positive and GFAP-positive cells between the 2 types of undifferentiated MSCs from healthy donors (Fig. 1B, E, lower panels). These data indicated that a comparable per-centage of hPCy-MSCs and DPSCs express β-III tubulin and GFAP under normal conditions. Furthermore, the protein lev-els of β-III tubulin and GFAP in hPCy-MSCs were similar to those in DPSCs. These results were confirmed by determining
the expression profile for each protein in both MSCs isolated from donors using the comparable geometric mean fluorescence intensities for β-III tubulin and GFAP (Fig. 1C, F). Moreover, the immunofluorescence staining results for the hPCy-MSCs and DPSCs confirmed that under stan-dard culture conditions, both MSCs expressed β-III tubulin and GFAP (Fig. 1D, G).
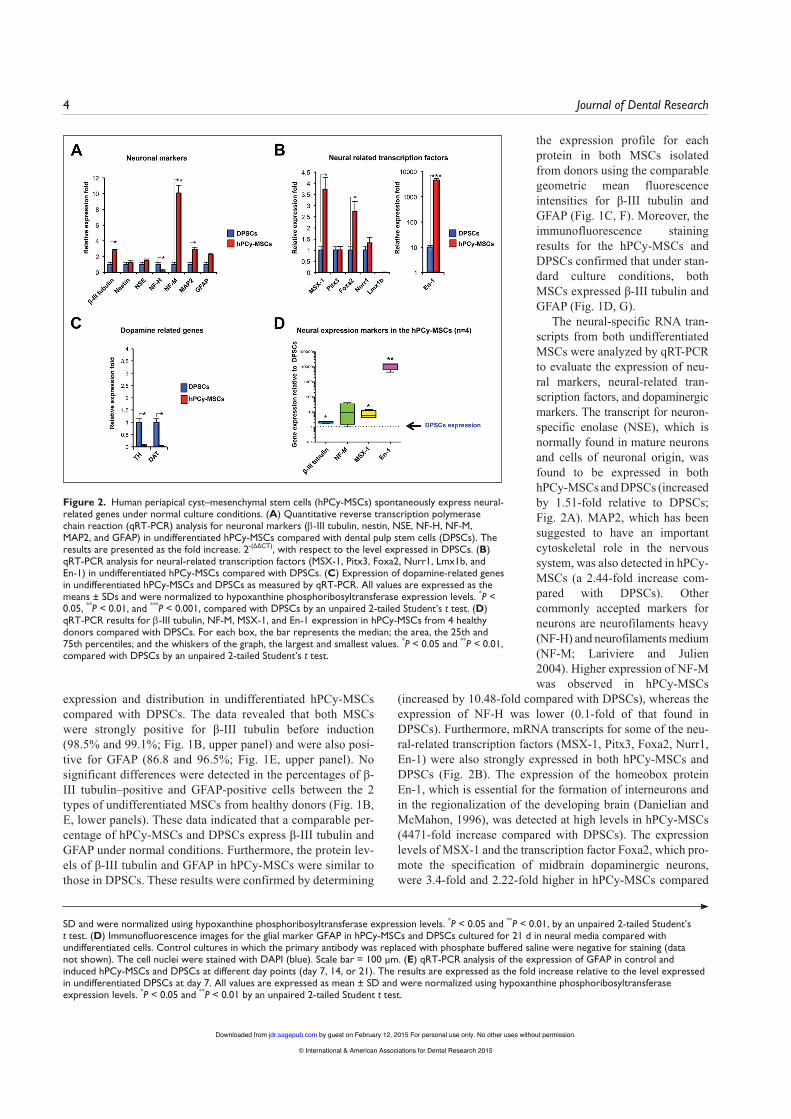
The neural-specific RNA tran-scripts from both undifferentiated MSCs were analyzed by qRT-PCR to evaluate the expression of neu-ral markers, neural-related tran-scription factors, and dopaminergic markers. The transcript for neuron-specific enolase (NSE), which is normally found in mature neurons and cells of neuronal origin, was found to be expressed in both hPCy-MSCs and DPSCs (increased by 1.51-fold relative to DPSCs; Fig. 2A). MAP2, which has been suggested to have an important cytoskeletal role in the nervous system, was also detected in hPCy-MSCs (a 2.44-fold increase com-pared with DPSCs). Other commonly accepted markers for neurons are neurofilaments heavy (NF-H) and neurofilaments medium (NF-M; Lariviere and Julien 2004). Higher expression of NF-M was observed in hPCy-MSCs
(increased by 10.48-fold compared with DPSCs), whereas the expression of NF-H was lower (0.1-fold of that found in DPSCs). Furthermore, mRNA transcripts for some of the neu-ral-related transcription factors (MSX-1, Pitx3, Foxa2, Nurr1, En-1) were also strongly expressed in both hPCy-MSCs and DPSCs (Fig. 2B). The expression of the homeobox protein En-1, which is essential for the formation of interneurons and in the regionalization of the developing brain (Danielian and McMahon, 1996), was detected at high levels in hPCy-MSCs (4471-fold increase compared with DPSCs). The expression levels of MSX-1 and the transcription factor Foxa2, which pro-mote the specification of midbrain dopaminergic neurons, were 3.4-fold and 2.22-fold higher in hPCy-MSCs compared
Figure 2. Human periapical cyst–mesenchymal stem cells (hPCy-MSCs) spontaneously express neural-related genes under normal culture conditions. (A) Quantitative reverse transcription polymerase chain reaction (qRT-PCR) analysis for neuronal markers (β-III tubulin, nestin, NSE, NF-H, NF-M, MAP2, and GFAP) in undifferentiated hPCy-MSCs compared with dental pulp stem cells (DPSCs). The results are presented as the fold increase. 2-(∆∆CT), with respect to the level expressed in DPSCs. (B) qRT-PCR analysis for neural-related transcription factors (MSX-1, Pitx3, Foxa2, Nurr1, Lmx1b, and En-1) in undifferentiated hPCy-MSCs compared with DPSCs. (C) Expression of dopamine-related genes in undifferentiated hPCy-MSCs and DPSCs as measured by qRT-PCR. All values are expressed as the means ± SDs and were normalized to hypoxanthine phosphoribosyltransferase expression levels. *P < 0.05, **P < 0.01, and ***P < 0.001, compared with DPSCs by an unpaired 2-tailed Student’s t test. (D) qRT-PCR results for β-III tubulin, NF-M, MSX-1, and En-1 expression in hPCy-MSCs from 4 healthy donors compared with DPSCs. For each box, the bar represents the median; the area, the 25th and 75th percentiles; and the whiskers of the graph, the largest and smallest values. *P < 0.05 and **P < 0.01, compared with DPSCs by an unpaired 2-tailed Student’s t test.
SD and were normalized using hypoxanthine phosphoribosyltransferase expression levels. *P < 0.05 and **P < 0.01, by an unpaired 2-tailed Student’s t test. (D) Immunofluorescence images for the glial marker GFAP in hPCy-MSCs and DPSCs cultured for 21 d in neural media compared with undifferentiated cells. Control cultures in which the primary antibody was replaced with phosphate buffered saline were negative for staining (data not shown). The cell nuclei were stained with DAPI (blue). Scale bar = 100 μm. (E) qRT-PCR analysis of the expression of GFAP in control and induced hPCy-MSCs and DPSCs at different day points (day 7, 14, or 21). The results are expressed as the fold increase relative to the level expressed in undifferentiated DPSCs at day 7. All values are expressed as mean ± SD and were normalized using hypoxanthine phosphoribosyltransferase expression levels. *P < 0.05 and **P < 0.01 by an unpaired 2-tailed Student t test.
by guest on February 12, 2015 For personal use only. No other uses without permission.jdr.sagepub.comDownloaded from
© International & American Associations for Dental Research 2015
Human Periapical Cyst–Mesenchymal Stem Cells 5
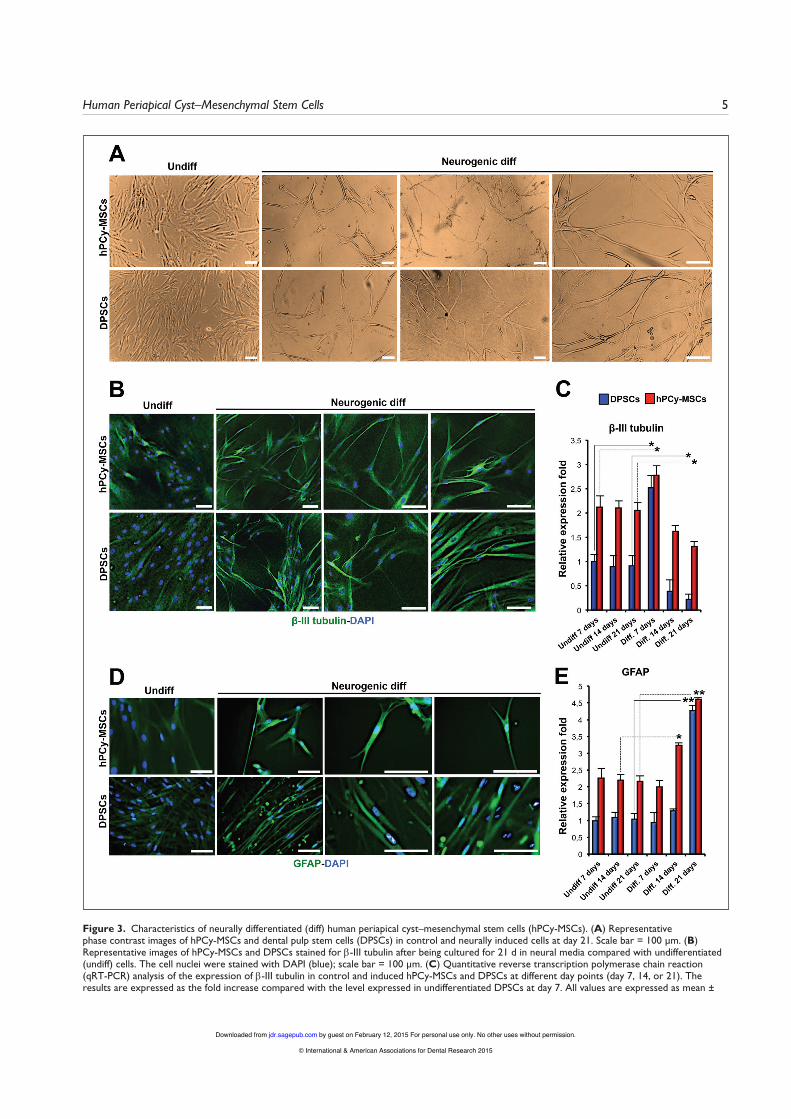
Figure 3. Characteristics of neurally differentiated (diff) human periapical cyst–mesenchymal stem cells (hPCy-MSCs). (A) Representative phase contrast images of hPCy-MSCs and dental pulp stem cells (DPSCs) in control and neurally induced cells at day 21. Scale bar = 100 μm. (B) Representative images of hPCy-MSCs and DPSCs stained for β-III tubulin after being cultured for 21 d in neural media compared with undifferentiated (undiff) cells. The cell nuclei were stained with DAPI (blue); scale bar = 100 μm. (C) Quantitative reverse transcription polymerase chain reaction (qRT-PCR) analysis of the expression of β-III tubulin in control and induced hPCy-MSCs and DPSCs at different day points (day 7, 14, or 21). The results are expressed as the fold increase compared with the level expressed in undifferentiated DPSCs at day 7. All values are expressed as mean ±
by guest on February 12, 2015 For personal use only. No other uses without permission.jdr.sagepub.comDownloaded from
© International & American Associations for Dental Research 2015

6 Journal of Dental Research
with DPSCs, respectively. The transcription factors Nurr1 and Pitx3, which are essential for the differentiation and mainte-nance of dopaminergic neurons, were expressed at similar lev-els in both types of MSCs, whereas Lmx1b was not detectable in either type of MSCs. The expression of tyrosine hydroxylase (TH), a neurotransmitter-related enzyme in catecholaminergic neurons, and dopamine transporter (DAT) was detected only at low levels in undifferentiated hPCy-MSCs compared with DPSCs (<0.1-fold that of DPSCs; Fig. 2C). These data demon-strate that both undifferentiated MSCs showed comparable expression of neural-related genes (nestin, NSE, GFAP) and neural-related transcription factors (Pitx3, Nurr1, Lmx1b). However, compared with undifferentiated DPSCs, hPCy-MSCs exhibited higher expression of β-III tubulin, NF-M, MAP2, MSX-1, Foxa2, and En-1. Moreover, when MSCs iso-lated from 4 healthy donors were compared, higher expression of the mRNAs for β-III tubulin, NF-M, MSX-1, and En-1 was observed in hPCy-MSCs compared with DPSCs (Fig. 2D).
Differentiation of hPCy-MSCs Into Neuronal CellsUnder neurogenic differentiation conditions, neurosphere for-mation was observed in hPCy-MSCs after 7 d (Appendix Fig. 2). After plating neurosphere-derived hPCy-MSCs on polyor-nithine- and laminin-coated dishes at day 15, the extension of long, thin cytoplasmic processes was observed at day 16 (Appendix Fig. 2). The cells appeared to have morphologies consistent with neuron-like cells and demonstrated a stellar-like appearance (Fig. 3A, Appendix Fig. 2). Both MSCs that were not exposed to neuronal differentiation stimuli did not show any changes in morphology (Appendix Fig. 3A).
To verify this phenotype, immunofluorescence analysis and confocal imaging were used. It was observed that both types of MSCs were able to differentiate with similar efficiencies into cells that expressed higher levels of β-III tubulin compared with controls (Fig. 3B, Appendix Fig. 3B). Furthermore, the protein expression pattern of GFAP in hPCy-MSCs and DPSCs cultured in neural inductive conditions was determined by immunofluorescence. The results showed a progressive increase in GFAP expression in both types of MSCs compared with controls (Fig. 3D).
After inducing neurogenic differentiation, qRT-PCR was performed to detect the expression of neuron-specific markers at different time points. The mRNA expression of β-III tubulin was substantially upregulated at day 7 in both MSCs and then reduced at days 14 and 21 (Fig. 3C). As shown in Figure 3E, the GFAP transcript level was upregulated in differentiated hPCy-MSCs and DPSCs at days 14 and 21. qRT-PCR analysis indicated that the gene expression profiles of neurally differen-tiated hPCy-MSCs and DPSCs were similar to those of mature neuronal cells.
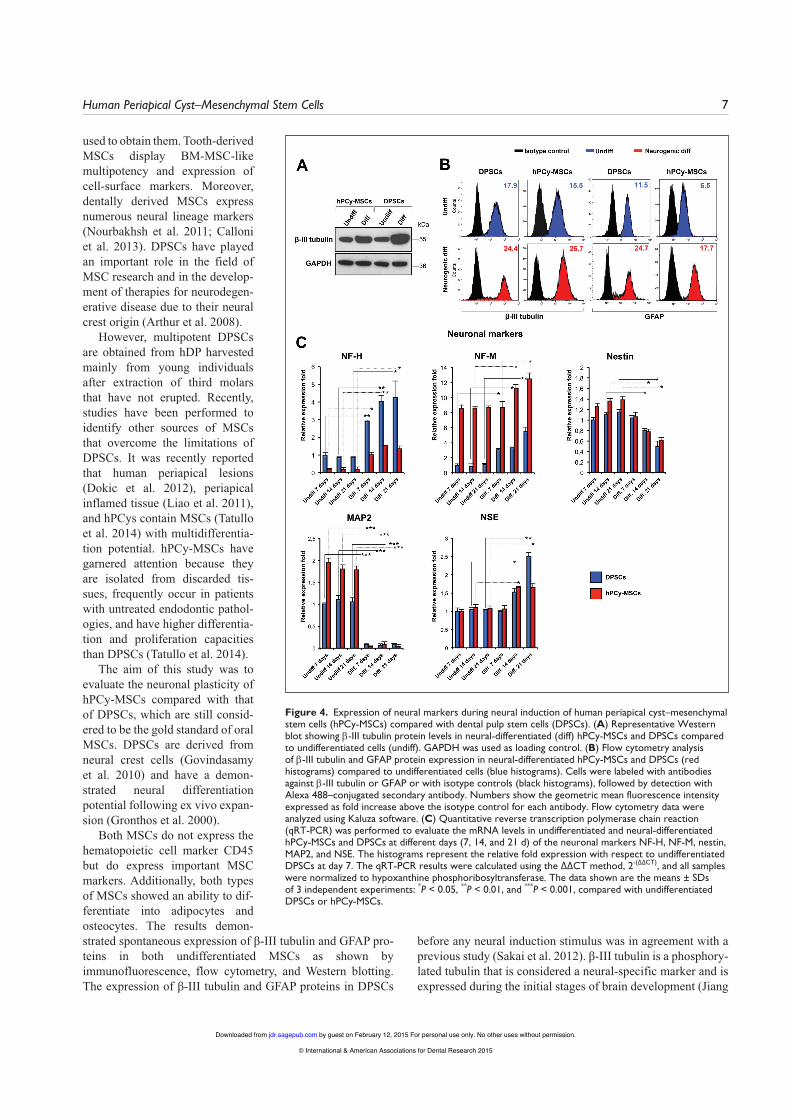
Changes in gene expression were also seen at the protein level, as shown by quantitative immunoblot and flow cytome-try analyses with antibodies targeting β-III tubulin and GFAP proteins. Upregulation of these proteins was observed for both
neuronal differentiated MSCs compared to undifferentiated cells (Fig. 4A, B). The results demonstrated that the protein expression levels of β-III tubulin and GFAP increased signifi-cantly after neurogenic stimulation in hPCy-MSCs and DPSCs.
When hPCy-MSCs and DPSCs were cultured in neuroin-ductive media, the transcript levels of the neural precursor marker nestin were decreased at days 14 and 21 compared with controls (Fig. 4C). Conversely, when hPCy-MSCs and DPSCs were cultured in neurogenic media, there was a significant increase in the levels of NF-H and NF-M transcripts compared with controls. The transcript level of NF-H was upregulated by 5.66-fold at day 7, by 8.66-fold at day 14, and by 7.5-fold at day 21 in differentiated hPCy-MSCs compared with undiffer-entiated hPCy-MSCs. The transcript level of NF-M, an inter-mediate neurofilament protein expressed in growing axons, was upregulated by 1.30-fold at day 14 and by 1.45-fold at day 21 in differentiated hPCy-MSCs compared with controls (Fig. 4C). The transcript level of MAP2, a rich neuronal cytoskeletal protein that binds to tubulin and stabilizes microtubules, was significantly downregulated by day 7 of induction compared with controls in both MSCs. The expression of NSE was increased in both neurally differentiated MSCs at day 14 and day 21 compared with controls (Fig. 4C).
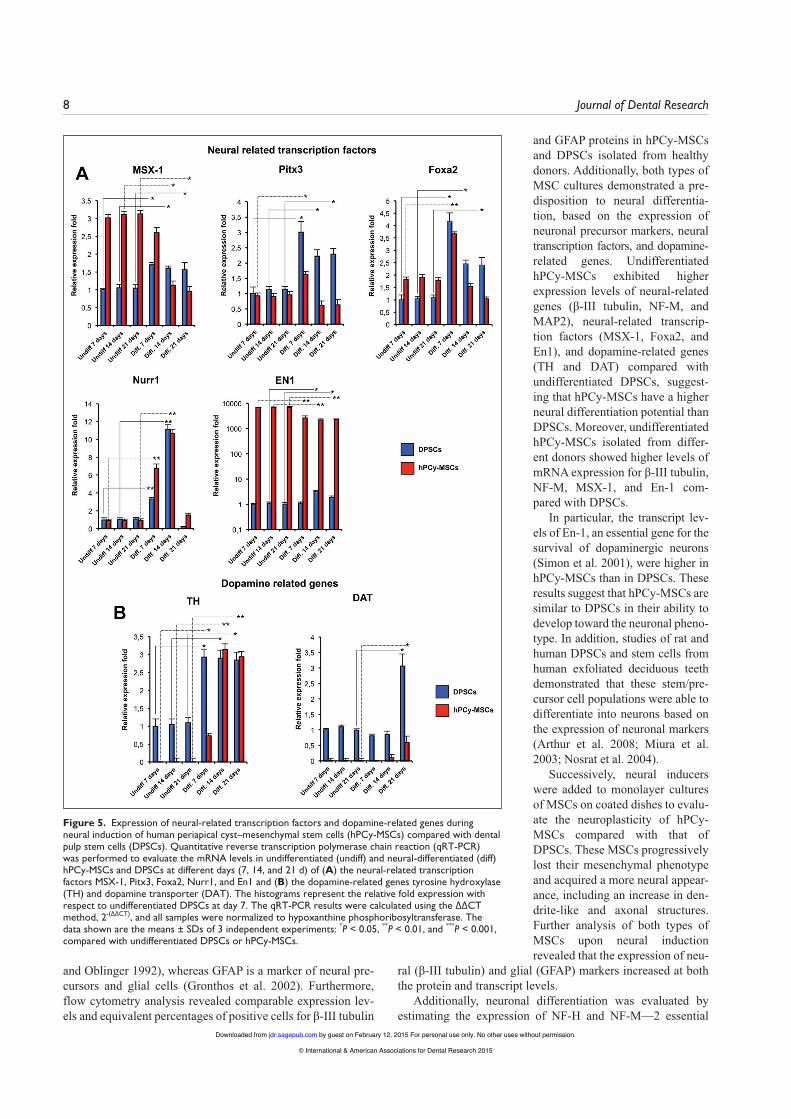
Furthermore, qRT-PCR was performed to detect the expres-sion of the neuron-specific transcription factors En1, Nurr1, Pitx3, Foxa2, and MSX-1 at different time points. As shown in Figure 5A, the transcript levels of Pitx3 and Foxa2 in hPCy-MSCs were strongly upregulated in the presence of neurogenic media at day 7, which was followed by a reduction in expres-sion at days 14 and 21, whereas Nurr1 was upregulated until day 14, followed by a strong reduction in expression on day 21. The transcription factors MSX-1 and En1 were downregulated in hPCy-MSCs and upregulated in DPSCs after neural differ-entiation (Fig. 5A). These results strongly suggest that hPCy-MSCs generate neuron-like cells under suitable conditions. The induced hPCy-MSCs and DPSCs were also screened for the expression of the dopamine-related genes TH and DAT using qRT-PCR. TH expression was strongly upregulated in neurally differentiated hPCy-MSCs starting from day 7, and TH expression was higher at day 14 than in neurally differenti-ated DPSCs (Fig. 5B). The upregulation of TH suggests that the cells had been induced to form dopaminergic neurons. The level of DAT was upregulated in differentiated hPCy-MSCs at days 14 and 21. Conversely, in DPSCs, the transcript level of DAT was upregulated by 3.05-fold only at day 21 (Fig. 5B).
DiscussionCurrently, the main tissue sources for isolating MSCs that could be used for cell-based therapies to treat neurologic dis-eases include the BM, umbilical cord, amniotic fluid, adipose tissue, and hDP (Zuk et al. 2002; Nosrat et al. 2004; Arthur et al. 2008; Pal et al. 2009; Yan ZJ et al. 2013). MSC isolation can be technically difficult and can produce severe discomfort for patients due to the invasiveness of most surgical procedures
by guest on February 12, 2015 For personal use only. No other uses without permission.jdr.sagepub.comDownloaded from
© International & American Associations for Dental Research 2015
Human Periapical Cyst–Mesenchymal Stem Cells 7
Figure 4. Expression of neural markers during neural induction of human periapical cyst–mesenchymal stem cells (hPCy-MSCs) compared with dental pulp stem cells (DPSCs). (A) Representative Western blot showing β-III tubulin protein levels in neural-differentiated (diff) hPCy-MSCs and DPSCs compared to undifferentiated cells (undiff). GAPDH was used as loading control. (B) Flow cytometry analysis of β-III tubulin and GFAP protein expression in neural-differentiated hPCy-MSCs and DPSCs (red histograms) compared to undifferentiated cells (blue histograms). Cells were labeled with antibodies against β-III tubulin or GFAP or with isotype controls (black histograms), followed by detection with Alexa 488–conjugated secondary antibody. Numbers show the geometric mean fluorescence intensity expressed as fold increase above the isotype control for each antibody. Flow cytometry data were analyzed using Kaluza software. (C) Quantitative reverse transcription polymerase chain reaction (qRT-PCR) was performed to evaluate the mRNA levels in undifferentiated and neural-differentiated hPCy-MSCs and DPSCs at different days (7, 14, and 21 d) of the neuronal markers NF-H, NF-M, nestin, MAP2, and NSE. The histograms represent the relative fold expression with respect to undifferentiated DPSCs at day 7. The qRT-PCR results were calculated using the ∆∆CT method, 2-(∆∆CT), and all samples were normalized to hypoxanthine phosphoribosyltransferase. The data shown are the means ± SDs of 3 independent experiments: *P < 0.05, **P < 0.01, and ***P < 0.001, compared with undifferentiated DPSCs or hPCy-MSCs.
used to obtain them. Tooth-derived MSCs display BM-MSC-like multipotency and expression of cell-surface markers. Moreover, dentally derived MSCs express numerous neural lineage markers (Nourbakhsh et al. 2011; Calloni et al. 2013). DPSCs have played an important role in the field of MSC research and in the develop-ment of therapies for neurodegen-erative disease due to their neural crest origin (Arthur et al. 2008).
However, multipotent DPSCs are obtained from hDP harvested mainly from young individuals after extraction of third molars that have not erupted. Recently, studies have been performed to identify other sources of MSCs that overcome the limitations of DPSCs. It was recently reported that human periapical lesions (Dokic et al. 2012), periapical inflamed tissue (Liao et al. 2011), and hPCys contain MSCs (Tatullo et al. 2014) with multidifferentia-tion potential. hPCy-MSCs have garnered attention because they are isolated from discarded tis-sues, frequently occur in patients with untreated endodontic pathol-ogies, and have higher differentia-tion and proliferation capacities than DPSCs (Tatullo et al. 2014).
The aim of this study was to evaluate the neuronal plasticity of hPCy-MSCs compared with that of DPSCs, which are still consid-ered to be the gold standard of oral MSCs. DPSCs are derived from neural crest cells (Govindasamy et al. 2010) and have a demon-strated neural differentiation potential following ex vivo expan-sion (Gronthos et al. 2000).
Both MSCs do not express the hematopoietic cell marker CD45 but do express important MSC markers. Additionally, both types of MSCs showed an ability to dif-ferentiate into adipocytes and osteocytes. The results demon-strated spontaneous expression of β-III tubulin and GFAP pro-teins in both undifferentiated MSCs as shown by immunofluorescence, flow cytometry, and Western blotting. The expression of β-III tubulin and GFAP proteins in DPSCs
before any neural induction stimulus was in agreement with a previous study (Sakai et al. 2012). β-III tubulin is a phosphory-lated tubulin that is considered a neural-specific marker and is expressed during the initial stages of brain development (Jiang
by guest on February 12, 2015 For personal use only. No other uses without permission.jdr.sagepub.comDownloaded from
© International & American Associations for Dental Research 2015
8 Journal of Dental Research
Figure 5. Expression of neural-related transcription factors and dopamine-related genes during neural induction of human periapical cyst–mesenchymal stem cells (hPCy-MSCs) compared with dental pulp stem cells (DPSCs). Quantitative reverse transcription polymerase chain reaction (qRT-PCR) was performed to evaluate the mRNA levels in undifferentiated (undiff) and neural-differentiated (diff) hPCy-MSCs and DPSCs at different days (7, 14, and 21 d) of (A) the neural-related transcription factors MSX-1, Pitx3, Foxa2, Nurr1, and En1 and (B) the dopamine-related genes tyrosine hydroxylase (TH) and dopamine transporter (DAT). The histograms represent the relative fold expression with respect to undifferentiated DPSCs at day 7. The qRT-PCR results were calculated using the ∆∆CT method, 2-(∆∆CT), and all samples were normalized to hypoxanthine phosphoribosyltransferase. The data shown are the means ± SDs of 3 independent experiments: *P < 0.05, **P < 0.01, and ***P < 0.001, compared with undifferentiated DPSCs or hPCy-MSCs.
and Oblinger 1992), whereas GFAP is a marker of neural pre-cursors and glial cells (Gronthos et al. 2002). Furthermore, flow cytometry analysis revealed comparable expression lev-els and equivalent percentages of positive cells for β-III tubulin
and GFAP proteins in hPCy-MSCs and DPSCs isolated from healthy donors. Additionally, both types of MSC cultures demonstrated a pre-disposition to neural differentia-tion, based on the expression of neuronal precursor markers, neural transcription factors, and dopamine-related genes. Undifferentiated hPCy-MSCs exhibited higher expression levels of neural-related genes (β-III tubulin, NF-M, and MAP2), neural-related transcrip-tion factors (MSX-1, Foxa2, and En1), and dopamine-related genes (TH and DAT) compared with undifferentiated DPSCs, suggest-ing that hPCy-MSCs have a higher neural differentiation potential than DPSCs. Moreover, undifferentiated hPCy-MSCs isolated from differ-ent donors showed higher levels of mRNA expression for β-III tubulin, NF-M, MSX-1, and En-1 com-pared with DPSCs.
In particular, the transcript lev-els of En-1, an essential gene for the survival of dopaminergic neurons (Simon et al. 2001), were higher in hPCy-MSCs than in DPSCs. These results suggest that hPCy-MSCs are similar to DPSCs in their ability to develop toward the neuronal pheno-type. In addition, studies of rat and human DPSCs and stem cells from human exfoliated deciduous teeth demonstrated that these stem/pre-cursor cell populations were able to differentiate into neurons based on the expression of neuronal markers (Arthur et al. 2008; Miura et al. 2003; Nosrat et al. 2004).
Successively, neural inducers were added to monolayer cultures of MSCs on coated dishes to evalu-ate the neuroplasticity of hPCy-MSCs compared with that of DPSCs. These MSCs progressively lost their mesenchymal phenotype and acquired a more neural appear-ance, including an increase in den-drite-like and axonal structures. Further analysis of both types of MSCs upon neural induction revealed that the expression of neu-
ral (β-III tubulin) and glial (GFAP) markers increased at both the protein and transcript levels.
Additionally, neuronal differentiation was evaluated by estimating the expression of NF-H and NF-M—2 essential
by guest on February 12, 2015 For personal use only. No other uses without permission.jdr.sagepub.comDownloaded from
© International & American Associations for Dental Research 2015

Human Periapical Cyst–Mesenchymal Stem Cells 9
structural components of the axon cytoskeleton. NF-M is expressed prior to NF-H during neural differentiation. After neural induction, qRT-PCR results demonstrated that the tran-script levels of NF-H and NF-M were upregulated in both types of MSCs. In addition, qRT-PCR analysis showed upregu-lation of NSE, neural-related transcription factors (Pitx3, Foxa1, Nurr1), and dopamine-related genes (TH, DAT) in both hPCy-MSCs and DPSCs. However, the expression levels of neural markers MAP2 and nestin in both types of differentiated MSCs were decreased by day 7.
MAP2 is a mature neuron marker, and normally it is increased in association with neural differentiation. However, the MAP2 expression levels were downregulated in neurally differentiated dental-derived MSCs (Vollner et al. 2009; Yang et al. 2014).
Nestin is a neural stem/precursor marker and is supposed to be decreased when MSCs are induced into neurogenic differ-entiation (Arthur et al. 2008). The expression levels of neural transcription factors MSX-1 and En-1 were decreased by day 7 in differentiated hPCy-MSCs and were upregulated in differ-entiated DPSCs at day 7, 14, and 21.
In summary, this study showed that hPCy-MSCs, in a man-ner similar to DPSCs, have neural progenitor-like properties and express neuron-specific proteins and genes, spontaneously and after neural induction. hPCy-MSCs attained a neural mor-phology and significantly overexpressed several neural mark-ers under appropriate neural stimulation conditions. hPCy-MSCs can be easily obtained from patients without the need to sacrifice biologically healthy tissues and without the need to perform invasive surgical procedures. Thus, these MSCs could become one of the most promising cell sources for the treatment of neurodegenerative diseases.
Author ContributionsM. Marrelli, F. Paduano, M. Tatullo, contributed to conception, design, data acquisition, analysis, and interpretation, drafted and critically revised the manuscript. All authors gave final approval and agree to be accountable for all aspects of the work.
AcknowledgmentsThe present study was supported by the “PROMETEO Project–Progettazione e Sviluppo di piattaforme tecnologiche innovative ed ottimizzazione di PROcessi per applicazioni in MEdicina rigenerativa in ambito oromaxillofaciale, emaTologico, nEuro-logico e cardiOlogico” (PON01_02834). We are grateful to Dr. Biamonte Filippo for providing the GFAP antibody. The authors declare no potential conflicts of interest with respect to the authorship and/or publication of this article.
ReferencesArthur A, Rychkov G, Shi S, Koblar SA, Gronthos S. 2008. Adult human den-
tal pulp stem cells differentiate toward functionally active neurons under appropriate environmental cues. Stem Cells (Dayton, Ohio). 26:1787–1795.
Bieback K, Kern S, Kluter H, Eichler H. 2004. Critical parameters for the iso-lation of mesenchymal stem cells from umbilical cord blood. Stem Cells. 22:625–634.
Calloni R, Cordero EA, Henriques JA, Bonatto D. 2013. Reviewing and updat-ing the major molecular markers for stem cells. Stem Cells Dev. 22:1455–1476.
Chamberlain G, Fox J, Ashton B, Middleton J. 2007. Concise review: mesen-chymal stem cells: their phenotype, differentiation capacity, immunological features, and potential for homing. Stem Cells. 25:2739–2749.
Danielian PS, McMahon AP. 1996. Engrailed-1 as a target of the Wnt-1 signal-ling pathway in vertebrate midbrain development. Nature. 383(6598):332–334.
Dokic J, Tomić S, Cerović S, Todorović V, Rudolf R, Colić M. 2012. Characterization and immunosuppressive properties of mesenchymal stem cells from periapical lesions. J Clin Periodontol. 39:807–816.
Dominici M, Le Blanc K, Mueller I, Slaper-Cortenbach I, Marini F, Krause D, Deans R, Keating A, Prockop DJ, Horwitz E. 2006. Minimal criteria for defining multipotent mesenchymal stromal cells: the International Society for Cellular Therapy position statement. Cytotherapy. 8:315–317.
Govindasamy V, Abdullah AN, Ronald VS, Musa S, Ab Aziz ZA, Zain RB, Totey S, Bhonde RR, Abu Kasim NH. 2010. Inherent differential propen-sity of dental pulp stem cells derived from human deciduous and permanent teeth. J Endod. 36:1504–1515.
Gronthos S, Mankani M, Brahim J, Robey PG, Shi S. 2000. Postnatal human dental pulp stem cells (DPSCs) in vitro and in vivo. Proc Natl Acad Sci U S A. 97:13625–13630.
Gronthos S, Brahim J, Li W, Fisher LW, Cherman N, Boyde A, DenBesten P, Robey PG, Shi S. 2002. Stem cell properties of human dental pulp stem cells. J Dent Res. 81:531–535.
Herzog EL, Chai L, Krause DS. 2003. Plasticity of marrow-derived stem cells. Blood. 102:3483–3493.
Jiang YQ, Oblinger MM. 1992. Differential regulation of beta III and other tubulin genes during peripheral and central neuron development. J Cell Sci. 103 Pt 3:643–651.
Keating A. 2012. Mesenchymal stromal cells: new directions. Cell Stem Cell. 10:709–716.
Lariviere RC, Julien JP. 2004. Functions of intermediate filaments in neuronal development and disease. J Neurobiol. 58:131–148.
Lees AJ, Hardy J, Revesz T. 2009. Parkinson’s disease. Lancet. 373(9680):2055–2066.
Liao J, Al Shahrani M, Al-Habib M, Tanaka T, Huang GT. 2011. Cells isolated from inflamed periapical tissue express mesenchymal stem cell markers and are highly osteogenic. J Endod. 37:1217–1224.
Maltman DJ, Hardy SA, Przyborski SA. 2011. Role of mesenchymal stem cells in neurogenesis and nervous system repair. Neurochem Int. 59:347–356.
Marrelli M, Paduano F, Tatullo M. 2013. Cells isolated from human periapi-cal cysts express mesenchymal stem cell-like properties. Int J Biol Sci. 9:1070–1078.
Martino G, Pluchino S. 2006. The therapeutic potential of neural stem cells. Nature reviews Neuroscience. 7:395–406.
Martino G, Franklin RJ, Baron Van Evercooren A, Kerr DA; Stem Cells in Multiple Sclerosis (STEMS) Consensus Group. 2010. Stem cell transplan-tation in multiple sclerosis: current status and future prospects. Nat Rev Neurol. 6:247–255.
Miura M, Gronthos S, Zhao M, Lu B, Fisher LW, Robey PG, Shi S. 2003. SHED: stem cells from human exfoliated deciduous teeth. Proc Natl Acad Sci U S A. 100:5807–5812.
Morsczeck C, Schmalz G, Reichert TE, Völlner F, Galler K, Driemel O. 2008. Somatic stem cells for regenerative dentistry. Clin Oral Investig. 12:113–118.
Nosrat IV, Smith CA, Mullally P, Olson L, Nosrat CA. 2004. Dental pulp cells provide neurotrophic support for dopaminergic neurons and differentiate into neurons in vitro; implications for tissue engineering and repair in the nervous system. Eur J Neurosci. 19:2388–2398.
Nourbakhsh N, Soleimani M, Taghipour Z, Karbalaie K, Mousavi SB, Talebi A, Nadali F, Tanhaei S, Kiyani GA, Nematollahi M, et al. 2011. Induced in vitro differentiation of neural-like cells from human exfoliated deciduous teeth-derived stem cells. Int J Dev Biol. 55:189–195.
Pal R, Venkataramana NK, Bansal A, Balaraju S, Jan M, Chandra R, Dixit A, Rauthan A, Murgod U, Totey S. 2009. Ex vivo-expanded autologous bone marrow-derived mesenchymal stromal cells in human spinal cord injury/paraplegia: a pilot clinical study. Cytotherapy. 11:897–911.
Park JC, Kim JM, Jung IH, Kim JC, Choi SH, Cho KS, Kim CS. 2011. Isolation and characterization of human periodontal ligament (PDL) stem cells (PDLSCs) from the inflamed PDL tissue: in vitro and in vivo evaluations. J Clin Periodontol. 38:721–731.
by guest on February 12, 2015 For personal use only. No other uses without permission.jdr.sagepub.comDownloaded from
© International & American Associations for Dental Research 2015

10 Journal of Dental Research
Peters H, Balling R. 1999. Teeth: where and how to make them. Trends Genet. 15:59–65.
Sakai K, Yamamoto A, Matsubara K, Nakamura S, Naruse M, Yamagata M, Sakamoto K, Tauchi R, Wakao N, Imagama S, et al. 2012. Human dental pulp-derived stem cells promote locomotor recovery after complete tran-section of the rat spinal cord by multiple neuro-regenerative mechanisms. J Clin Invest. 122:80–90.
Simon HH, Saueressig H, Wurst W, Goulding MD, O’Leary DD. 2001. Fate of midbrain dopaminergic neurons controlled by the engrailed genes. J Neurosci. 21:3126–3134.
Tatullo M, Marrelli M, Shakesheff KM, White LJ. 2014. Dental pulp stem cells: function, isolation and applications in regenerative medicine. J Tissue Eng Regen Med. 2014 May 21 [Epub ahead of print] in press. doi:10.1002/term.1899
Uccelli A, Laroni A, Freedman MS. 2011. Mesenchymal stem cells for the treatment of multiple sclerosis and other neurological diseases. Lancet Neurol. 10:649–656.
Vollner F, Ernst W, Driemel O, Morsczeck C. 2009. A two-step strategy for neu-ronal differentiation in vitro of human dental follicle cells. Differentiation. 77:433–441.
Yan K, Zhang R, Sun C, Chen L, Li P, Liu Y, Peng L, Sun H, Qin K, Chen F, et al. 2013. Bone marrow-derived mesenchymal stem cells maintain the resting phenotype of microglia and inhibit microglial activation. PloS One. 8:e84116.
Yan ZJ, Hu YQ, Zhang HT, Zhang P, Xiao ZY, Sun XL, Cai YQ, Hu CC, Xu RX. 2013. Comparison of the neural differentiation potential of human mesenchymal stem cells from amniotic fluid and adult bone marrow. Cell Mol Neurobiol. 33:465–475.
Yang C, Sun L, Li X, Xie L, Yu M, Feng L, Jiang Z, Guo W, Tian W. 2014. The potential of dental stem cells differentiating into neurogenic cell lineage after cultivation in different modes in vitro. Cell Reprogram. 16:379–391.
Zuk PA, Zhu M, Ashjian P, De Ugarte DA, Huang JI, Mizuno H, Alfonso ZC, Fraser JK, Benhaim P, Hedrick MH. 2002. Human adipose tissue is a source of multipotent stem cells. Mol Biol Cell. 13:4279–4295.
by guest on February 12, 2015 For personal use only. No other uses without permission.jdr.sagepub.comDownloaded from
© International & American Associations for Dental Research 2015