cyclic postural behavior in the crayfish, procambarus clarkii: properties of the pattern-initiating...
TRANSCRIPT

THE JOURNAL OF EXPERIMENTAL ZOOLOGY 267:404-415 (1993)
Cyclic Postural Behavior in the Crayfish, Procambarws clarkii; Properties of the Pattern-Initiating Network
DARRELL MOORE AND JAMES L. LARIMER Department of Biological Sciences, East Tennessee State University, Johnson City, Tennessee 37614 (D.M.); and Department of Zoology, University of Texas, Austin, Texas 78712 (J.L.L.)
ABSTRACT Crayfish exhibit complex cyclical adjustments in abdominal posture during certain forms of backward walking. An isolated nerve cord preparation was used to investigate the proper- ties of the interneurons which direct this alternation of abdominal flexion and extension.
The command function for this cyclic postural behavior appears to be the domain of a distributed network of multiple pattern-initiating interneurons: each interneuron may be viewed as a command element within a command system. The cyclic pattern may be elicited by stimulation of small axon bundles pulled from the ventrolateral margins of any of the abdominal connectives. As few as one stimulus pulse to the axon bundle can elicit a single cycle of patterned output, although more pulses are generally necessary. This suggests some convergence or amplification step in the pattern-initiating interneurons. The amplification may be accomplished by several pattern-initiating interneurons that are coupled to one another and converge on pattern generating circuits in each ganglion. Evidence supporting this interpretation is presented.
Experiments involving resection of the cord reveal that the pattern-initiating signals transfer lat- erally across all of the abdominal ganglia, but the network contains a bias for descending signal conduction once a lateral transfer is made. This finding agrees with other results. For example, re- cordings from pattern-initiating axon bundles a t rostra1 and caudal locations in the abdominal nerve cord show several descending but only one ascending unit activated during cyclic pattern generation.
We also show that an isolated ganglion is capable of producing the cyclic motor program, although the outputs are much weaker than those elicited in the intact abdominal cord. Therefore, the pattern- initiating system is both central and distributed.
In decapod crustaceans, the last six body seg- ments constitute the abdomen. This structure, which is well developed in the crayfish, lobster, and shrimp, has a prominent role in the overall behav- ior of the animal. The abdomen participates in re- production, in normal locomotion as well as escape movements, and in a wide variety of postural ad- justments that often assist other behavioral pat- terns by shifting the body orientation. The present study is concerned with cyclical movements of the postural musculature that accompany backwards walking.
The motor system underlying postural move- ments of the crayfish abdomen has received much attention by neurophysiologists over the years (for reviews see Bowerman and Larimer, ’76; Page, ’82; Larimer ’88) and is a relatively simple model prep- aration for the study of motor control. Its primary advantage is a small set of identified, readily ac- cessible motoneurons (Kennedy and Takeda, ’65; Kennedy et al., ’66; Fields, ’66; Sokolove and Tatton, ’75) which innervate the superficial tonic
0 1993 WILEY-LISS, INC.
0 1993 Wiley-Liss, Inc.
flexor and extensor muscles in each abdominal seg- ment. Each flexor and extensor muscle receives five excitatory motoneurons and one peripheral inhibitor.
A large population of premotor interneurons con- trol the tonic (postural) flexion and extension mo- toneurons. Earlier studies, using extracellular stimulation of small axon bundles, showed that spe- cific postures could be elicited by activating certain “command neurons” found in the interganglionic connectives of the abdominal nerve cord (Kennedy et al., ’66, ’67; Evoy and Kennedy, ’67; Williams and Larimer, ’81). As a result of recent intracellu- lar work, many of these premotor interneurons are now identified cells, characterized according to mor- phological as well as physiological properties (Miall and Larimer, ’82b; Larimer and Jellies, ’83; Larimer and Moore, ’84). Contrary to the earlier view of these premotor elements as single “command neu- rons” for abdominal flexion or extension, the con-
Received March 19,1993; revision accepted July 14,1993. Address reprint requests to James L. Larimer, Dept. ofZoology, Univ.
ofTexas, Austin, TX 78712.

CYCLIC MOTOR PATTERN GENERATION IN CRAYFISH 405
trol of postural adjustments is accomplished by large groups of synaptically interacting premotor interneurons (Jellies and Larimer, '85, '86; Murphy et al., '89).
Besides the well-studied tonic positional adjust- ments of the crayfish abdomen, the postural moto- neurons also participate in a more complex, rhythmic behavior (Larimer and Kennedy, '69). This cycli- cal motor act is expressed during backward walk- ing (Kovac, '74a) and allows the tail to alternately come in contact with and then withdraw from the substrate. As with the tonic postural system, the behavior can be released by extracellular stimula- tion of axon bundles in the abdominal connectives. One normal route of activation for this behavior is by illumination of the paired caudal photoreceptor neurons of the last abdominal ganglion (Edwards, '84). Is is now known that the cyclic postural be- havior may be initiated by intracellular activation of any of a number of pattern-initiating (PI) inter- neurons. These PI interneurons form a tightly cou- pled interganglionic network; the entire network is activated when any single PI cell is driven by un- patterned stimulation (Moore and Larimer, '87). Di- rect stimulation of the caudal photoreceptor activates this PI network (Simon and Edwards, '90). Inter- neurons controlling the tonic postural motor pro- grams interact in complex ways with the cyclical motor pattern, but the two behavioral subsystems apparently do not share premotor interneurons (Moore and Larimer, '88).
According to Kovac ('74b), rhythmicity in this motor pattern probably does not arise from central pattern generators present in each abdominal gan- glion but through interganglionic loops formed by flexion-driving interneurons subordinate to a com- mand neuron. In this paper, we report additional properties of the interneurons responsible for ini- tiating the cyclical pattern and their functional pathways throughout the abdominal nervous sys- tem. Our results indicate that pattern generation is derived from discrete intraganglionic oscillators as well as from interactions among interganglionic elements.
MATERIALS AND METHODS Crayfish (Procambarus clarkii) were obtained
from Waubun Laboratories of Schriever, Louisiana and from Louisiana Procambarus of St. Martinville, Louisiana. Both sexes were used and the animals ranged from 8 to 12 cm in rostrum-telson length. All experiments were performed on isolated abdom- inal nerve cords which were pinned ventral surface up in a sylgard-bottomed dish filled with crayfish saline (van Harreveld, '36) at 20°C.
The cyclical motor program, which consists of flexion and extension postural motoneuron outputs and is normally associated with backward walking (Kovac, '74a), was elicited by electrical stimulation applied to a small, consistently localized bundle of fibers (usually less than 25 pm in diameter) stripped from the interganglionic connectives. The stimuli, isolated from ground via a RF-coupled stimulus iso- lation unit and in the form of square wave pulses of 0.3 msec duration, were delivered through con- ventional suction electrodes. The experiments of the present study were properties of the interganglionic pathways through which the pattern-initiating sig- nals are carried and not with the oscillatory mech- anism per se. A single complete cycle of the motor output was remarkably consistent from prepara- tion to preparation and provided a reliable indica- tion that the pattern had been successfully evoked.
Motor outputs were recorded by means of conven- tional suction electrodes placed on various super- ficial third roots, which innervate the postural flexor muscles, and second roots, which contain the postural extension motoneurons. Individual moto- neurons were identified in these extracellular re- cordings by their relative spike amplitudes, a well-established method for the crayfish (Kennedy and Takeda, '65; Kennedy et al., '66; Evoy, et al., '67; Fields, et al., '67; Miall and Larimer, '82a,b; Larimer and Jellies, '83; Larimer and Moore, '84). According to convention, the six superficial flexor motoneurons (SFMNs) and the six superficial ex- tensor motoneurons (SEMNs) found in their respec- tive motor roots are numbered in order of increasing spike size. In most preparations, the second larg- est motoneuron spike (SFMN 5 and SEMN 5 ) cor- responds with the peripheral inhibitor to the flexor and extensor muscles, respectively. All of the re- maining units are excitatory motoneurons.
A variety of experimental designs were employed for this study, but all were based upon the profiles of extension and flexion motor activity evoked dur- ing a single cycle of pattern generation (Moore and Larimer, '87, '88). All electrophysiological data were taken from stored oscilloscope sweeps and from Polaroid photographs of these sweeps. Each indi- vidual experiment was replicated at least five times.
RESULTS Pattern initiation
The characteristic cyclic motor program was evoked throughout the isolated abdominal cord by caudal-directed stimulation of a small bundle of neurons stripped from one of the rostra1 (typically G1-G2) hemiconnectives. This bundle was, without
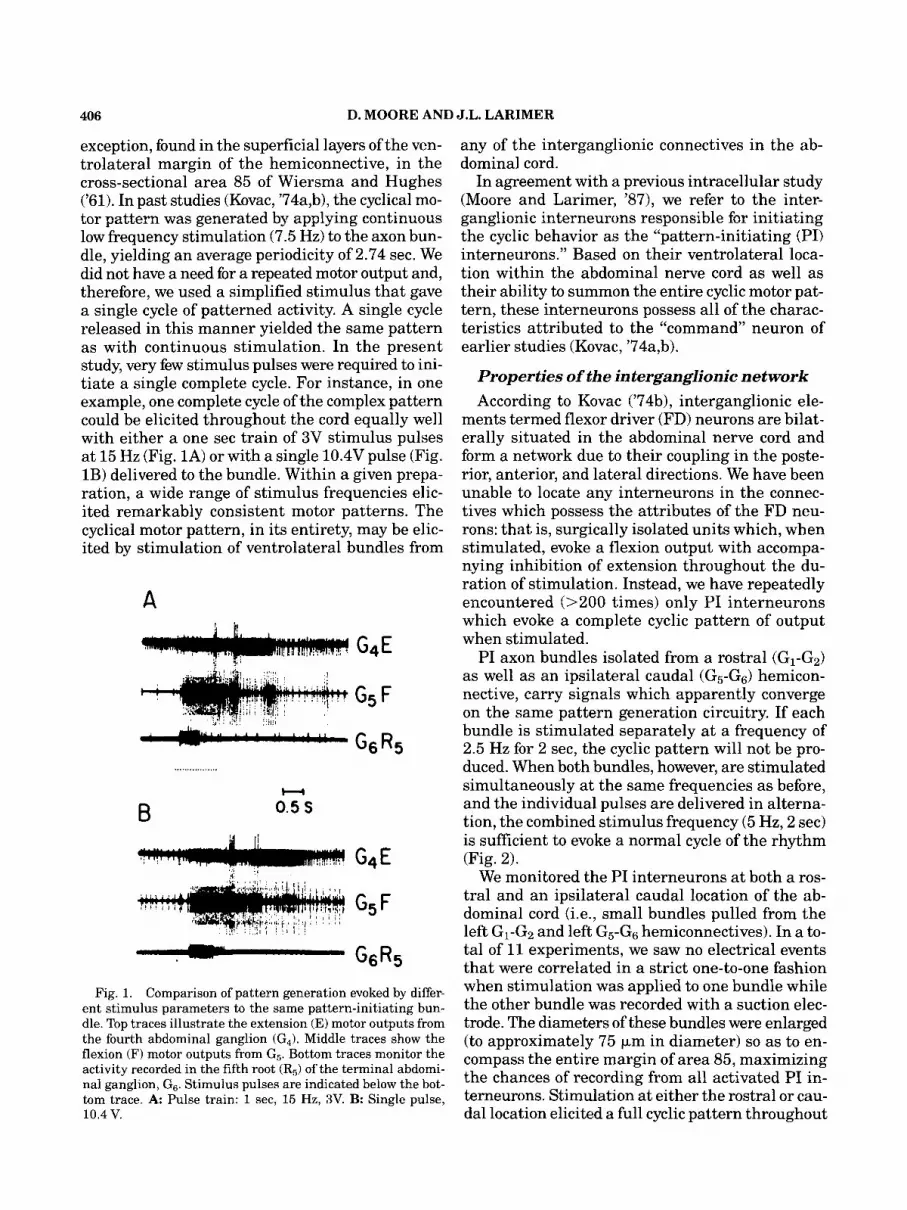
406 D. MOORE AND J.L. LARIMER
exception, found in the superficial layers of the ven- trolateral margin of the hemiconnective, in the cross-sectional area 85 of Wiersma and Hughes (’61). In past studies (Kovac, ’74a,b), the cyclical mo- tor pattern was generated by applying continuous low frequency stimulation (7.5 Hz) to the axon bun- dle, yielding an average periodicity of 2.74 sec. We did not have a need for a repeated motor output and, therefore, we used a simplified stimulus that gave a single cycle of patterned activity. A single cycle released in this manner yielded the same pattern as with continuous stimulation. In the present study, very few stimulus pulses were required to ini- tiate a single complete cycle. For instance, in one example, one complete cycle of the complex pattern could be elicited throughout the cord equally well with either a one sec train of 3V stimulus pulses at 15 Hz (Fig. 1A) or with a single 10.4V pulse (Fig. 1B) delivered to the bundle. Within a given prepa- ration, a wide range of stimulus frequencies elic- ited remarkably consistent motor patterns. The cyclical motor pattern, in its entirety, may be elic- ited by stimulation of ventrolateral bundles from
A
G6R5
w 0.5 s B
Fig. 1. Comparison of pattern generation evoked by differ- ent stimulus parameters to the same pattern-initiating bun- dle. Top traces illustrate the extension (E) motor outputs from the fourth abdominal ganglion (GJ. Middle traces show the flexion (F) motor outputs from G5. Bottom traces monitor the activity recorded in the fifth root (R5) of the terminal abdomi- nal ganglion, G6. Stimulus pulses are indicated below the bot- tom trace. A: Pulse train: 1 sec, 15 Hz, 3V. B: Single pulse, 10.4 V.
any of the interganglionic connectives in the ab- dominal cord.
In agreement with a previous intracellular study (Moore and Larimer, ,871, we refer to the inter- ganglionic interneurons responsible for initiating the cyclic behavior as the “pattern-initiating (PI) interneurons.” Based on their ventrolateral loca- tion within the abdominal nerve cord as well as their ability to summon the entire cyclic motor pat- tern, these interneurons possess all of the charac- teristics attributed to the “command” neuron of earlier studies (Kovac, ’74a,b).
Properties of the interganglionic network According to Kovac (’74b), interganglionic ele-
ments termed flexor driver (FD) neurons are bilat- erally situated in the abdominal nerve cord and form a network due to their coupling in the poste- rior, anterior, and lateral directions. We have been unable to locate any interneurons in the connec- tives which possess the attributes of the FD neu- rons: that is, surgically isolated units which, when stimulated, evoke a flexion output with accompa- nying inhibition of extension throughout the du- ration of stimulation. Instead, we have repeatedly encountered (>200 times) only PI interneurons which evoke a complete cyclic pattern of output when stimulated.
PI axon bundles isolated from a rostral (G1-G2) as well as an ipsilateral caudal (G5-Gs) hemicon- nective, carry signals which apparently converge on the same pattern generation circuitry. If each bundle is stimulated separately at a frequency of 2.5 Hz for 2 sec, the cyclic pattern will not be pro- duced. When both bundles, however, are stimulated simultaneously at the same frequencies as before, and the individual pulses are delivered in alterna- tion, the combined stimulus frequency (5 Hz, 2 sec) is sufficient to evoke a normal cycle of the rhythm (Fig. 2).
We monitored the PI interneurons at both a ros- tral and an ipsilateral caudal location of the ab- dominal cord (i.e., small bundles pulled from the left G1-G2 and left G5-Gs hemiconnectives). In a to- tal of ll experiments, we saw no electrical events that were correlated in a strict one-to-one fashion when stimulation was applied to one bundle while the other bundle was recorded with a suction elec- trode. The diameters of these bundles were enlarged (to approximately 75 pm in diameter) so as to en- compass the entire margin of area 85, maximizing the chances of recording from all activated PI in- terneurons. Stimulation at either the rostral or cau- dal location elicited a full cyclic pattern throughout

CYCLIC MOTOR PATTERN GENERATION IN CRAYFISH 407
A
, . . . . . W
0.5 S B
. . . . . .
c
. . . . . . . . . . . . Fig. 2. Demonstration that signals originating from ipsi-
lateral rostral and caudal pattern-initiating bundles converge on the same pattern generating circuitry. A 2 sec, 2.5 Hz stim- ulus pulse train to caudal bundle fails to elicit the pattern. B: Identical stimulus fails to elicit pattern from rostral bundle. C: Same stimuli, but in alternation from the two separate bun- dles, combine to evoke the cyclic pattern.
the abdominal cord. When the rostral axon bun- dle was stimulated, a 600-800 msec burst of im- pulses (50-60 spikes) was consistently recorded in the caudal bundle (Fig. 3B1). The caudal burst did not occur unless the stimulation applied rostrally was at or above the voltage and frequency thresh- old necessary to release the cyclic pattern. This cau- dal burst always incorporated several different spike amplitudes, none of which could be correlated one- to-one with the rostral stimuli. When the ipsilateral pathway was interrupted by cutting hemiconnectives between the stimulus and recording bundles (Fig. 3Bz), the number of spikes present in the caudal burst as well as the number of spike amplitudes contributing to the burst (Fig. 3B3) were severely reduced. Because the caudal burst persisted, though somewhat diminished, when the ipsilateral path- way was cut, the remaining units in the burst were activated from the contralateral hemiconnectives. This, of course, requires that the bilateral PI in-
terneurons be coupled to one another through con- nections across the ganglia.
When the caudal axon bundle was stimulated (with the ipsilateral pathway intact), however, a burst was never observed in the rostral bundle. In- stead, one unit typically responded with a constant latency to the caudal stimulus pulses except the first few pulses in the stimulus pulse train were not followed. For example, in the experiment depicted in Figure 3A, 19 pulses were delivered to the left G5-Gs bundle and the unit recorded in the left G1-GB responded, though at constant latency, only to the last 15 pulses. This result suggests that some fa- cilitation of signals must occur before the unit can fire in response to the caudal stimulus pulses. Sig- nificantly, the unit did not appear unless the cau- dal stimulus was of sufficient voltage and frequency to evoke pattern generation throughout the abdom- inal cord.
Network organization The neuronal elements underlying pattern ini-
tiation form a distributed network. The organiza- tion of these PI elements within the abdominal nerve cord was investigated by cutting the presump- tive pathways for signal conduction at various points. The results of the next experiment (Fig. 4) illustrate in detail the effects of ascending, descend- ing, and lateral pattern-initiating signal conduc- tion on the cyclic motor program. Extension and flexion motor outputs were monitored at G3 and G5 in the isolated abdominal nerve cord. When rostral- directed stimulation (10 Hz, 1 sec) was applied to a ventrolateral PI bundle stripped from the G4-G5 hemiconnective (Fig. 4A), the cyclic pattern was evoked at both G3 and G5. To reach G3, the signals must have ascended the cord, presumably through both hemiconnectives. To reach G5, the signals first must have crossed G4 and then descended the cord along the contralateral G4-G5 hemiconnective. These motor pattern recordings were not distin- guishable from those generated in nerve cords with- out the G4-G5 hemiconnective interrupted (data not shown; however, the G5F traces in Figure 4 may be compared with the G5F traces in Figure 1 which illustrates the motor patterns evoked in a cord with- out severed hemiconnectives).
Next, when caudal-directed stimulation was ap- plied to a PI bundle in the G1-G2 hemiconnective, with the ipsilateral G3-G4 and G4-G5 hemiconnectives severed, the cyclic pattern again occurred at both G3 and G5 (Fig. 4B). To reach G3, the signals pre- sumably descended along both hemiconnectives. However, to reach G5, the signals must have been

408
A D. MOORE AND J.L. LARIMER
IIII!IIIIIIIIII
. . . . . . . . . . . . . . . . . . _ 0.5’s
BI -I>-
i r - I j , . -- . I - ,,..,, . .- . 0.2s . . . . . * . . . .
8 3
. . . . . . . . . .
4
Fig. 3. Responses recorded in rostral and caudal pattern- initiating bundles during the cyclic postural program. Pres- ence or absence of the pattern is monitored with a suction electrode placed on an extension root of G4 (not shown). A Sin- gle unit in rostral (G,-G2) bundle is activated by stimulation of ipsilateral caudal (G,-G,J bundle. Spikes follow stimulus
conducted via the contralateral hemiconnective be- cause the ipsilateral pathways were interrupted. The motor pattern recordings in Figure 4A,B were remarkably similar.
Finally, when rostral-directed stimulation was applied to the G4-G5 PI bundle (Fig. 4 0 , with the cord cuts unchanged from the conditions of Figure 4B, the cyclic patterns again appeared at both G3 and G5. Interestingly, the motor output in G5 showed little or no change from the previous manipulation whereas the G3 motor pattern was severely reduced (compare with Fig. 4B). To reach G3, the signals had to ascend the cord in the contralateral hemi- connective after transferring laterally across G4. Similarly, to reach G5, the signals had to transfer laterally across G4 and descend the cord in the con- tralateral hemiconnective. Thus, there was only one route of access (i.e., a single hemiconnective) avail- able to conduct the pattern-initiating signals to each ganglion. Because the cyclic motor program evoked by the ascending signals in G3 was greatly diminished but the motor program elicited by the descending signals in G5 showed little or no reduc- tion, we conclude that a bias exists within the pattern-initiating network favoring descending over
pulses a t constant latency after missing first four pulses. B,: Burst recorded in caudal bundle is initiated from rostral bun- dle. B2: Arrangement used to block pathway between the ros- tral and caudal bundles BS: There is a reduction in the number of spikes in the caudal burst when the rostral-to-caudal path- way is blocked.
ascending signal conduction after lateral transfers across ganglia.
The signal required for pattern generation was found to travel through all of the interganglionic hemiconnectives and was capable of “transferring” from one side to the other at all of the abdominal ganglia, including G6 (Fig. 5). For example, when PI axons were stimulated in the left thoraco-ab- dominal hemiconnective and the ipsilateral (left) G1-G2 was cut, the pattern was still recorded in the nerve cord (Fig. 5A). Therefore, the pattern-initiating signal was conducted laterally across G1 and a de- scending signal was transmitted through the con- tralateral G1-G2 hemiconnective. Both ascending (Fig. 5C,F) and descending (Fig. 5A,B,D,E,G) sig- nals were conducted laterally across ganglia. Once the signal had crossed a ganglion, pattern genera- tion occurred in ganglia both rostral and caudal to the cross-over location (Fig. 5C). This indicated that the coupling mechanism between the right and left sides gives rise to both ascending and descending pattern-initiating signals. Patterned outputs were obtained even after cutting hemiconnectives such that the only available pathways for signal conduc- tion involved lateral transfers across two ganglia

CYCLIC MOTOR PATTERN GENERATION IN CRAYFISH 409
H
0.5 S
,.. , ' , ' '
,,;.. : , . , . i i G5 F
. . . , . . . .
..........
...,......
Fig. 4. Experiment demonstrating a directional bias for sig- nal conduction within the pattern-initiating network after lat- era1 signal transfer across a ganglion. The cyclic pattern is monitored for both extension and flexion at G3 and G5, as shown. Pattern-initiating bundles were pulled from a rostral (Gl-Gz) and an ipsilateral caudal (G,-G,) hemiconnective. Stimulus pulses (10 Hz, 1 sec) are indicated below each pair of oscillo-
scope traces. A Pattern-initiating stimulus is delivered to cau- dal bundle. Note cut to G4-G5 hemiconnective below bundle. B The G3-G4 hemiconnective is severed ipsilateral to the pattern- initiating bundles. Stimulus is applied to the rostral bundle. Compare motor outputs with those in A. C Nerve cord under same conditions as in B, except stimulus is delivered to caudal pattern-initiating bundle. Compare motor outputs with those in B.
(Fig. 5E). That the signals for pattern initiation need not travel simultaneously through both hemi- connectives for the pattern to occur was shown by
severing all the hemiconnectives on one side of the abdominal nerve cord (Fig. 5G). In all of the above experiments, the motor patterns were robust, though

410 D. MOORE AND J.L. LARIMER
.n
E
"h i "4
Fig. 5. Pattern-initiating signals transfer laterally across all of the ganglia. Positions of stimulating and recording elec- trodes, as well as locations of cuts made on various connectives, are shown for each experiment. A Descending signal trans- fers laterally across GI. B: Descending signal laterally tra- verses GS. C: Ascending signal transfers across G,, then both ascending and descending signals occur in contralateral hemi-
slightly reduced in some cases, after the mani- pulations.
The cyclic motor pattern at each abdominal gan- glion is bilaterally symmetrical. This symmetry is robust, as demonstrated by its persistence after a variety of pathway interruption experiments. For example, the right and left extension outputs from
connectives as evidenced by the presence of patterned outputs at the indicated locations. D: Descending signal crosses G,, then signal ascends contralateral hemiconnective. E: Descending sig- nal crosses G2 and Gq. F: Ascending signal crosses G2, then descends nerve cord. G Pattern generation still occurs if all hemiconnectives on one side of cord are cut.
G4 are remarkably similar in response to caudal- directed stimulation applied to the left G I G B PI bundle (Fig. 6A). When the left G4-G5 hemicon- nective is severed, both the right and left exten- sion motor patterns show a similar diminution in activity (Fig. 6B). Further bilateral reduction in the motor pattern occurs when the left G3-G4

CYCLIC MOTOR PATTERN GENERATION IN CRAYFISH 41 1
A 8 C
L ....._._..
H 0.5 S
R
. I . .
..........
Fig. 6. Cyclic motor patterns are bilaterally symmetrical. The cyclic pattern is monitored from both the right (R) and left (L) extension roots of G4, as shown. G6 is severed from the rest of the abdominal nerve cord in all of these preparations. A: Control: note similarity of R and L motor patterns. B. Motor
hemiconnective is cut and the abdominal cord is completely severed between G4 and G5 (Fig. 6C).
Isolated ganglia exhibit cyclic motor outputs Figure 7A shows a single cycle of the normal ex-
tension pattern produced at G4 by rostral-directed stimulation of a PI bundle teased from the left G4-G5 hemiconnective. Except for the separation of this bundle, the abdominal nerve cord was other- wise intact. When all of the hemiconnectives to this ganglion were cut, it retained its ability to produce cyclical motor outputs under stimulation from the teased-out PI bundle. Note that a diminution of re- sponse is apparent after the hemiconnectives were cut (Fig. 7B). Isolated ganglia, therefore, possess an oscillatory mechanism which can be activated by input signals from a single PI axon bundle. Con- sequently, this oscillation is not totally dependent upon interactions with the other segmental gan- glia. These interganglionic interactions, however,
L
patterns are reduced, but retain their symmetry, after sev- erence of the left G4-G5 hemiconnective. C: Motor patterns are severely reduced, but still symmetrical, after complete severence of the G4-G5 connective and section of the left G3-G4 hemiconnective.
may serve to intensify and shape the oscillatory patterns at each ganglion.
DISCUSSION The neural basis of pattern generation in this
sytem is apparently more complicated than was pre- viously thought (Larimer and Kennedy, ’69; Kovac, ’74a,b). However, the model proposed by Kovac (’74b) to account for the production of cyclical activity has proved to be a valuable first approximation of the neural substrate for the behavior and, accordingly, will serve as a point of departure for our work.
The early model for cyclic pattern generation The Kovac (’74b) model contained several impor-
tant hypothetical components. First, Kovac noted that the behavior could be initiated by stimulation of a single “command” interneuron found in the ven- trolateral margin (area 85 of Wiersma and Hughes, ’61) of the abdominal nerve cord. Second, Kovac pro-

412 D. MOORE AND J.L. LARIMER
H
, ...............................*............................ I .......... .. Fig. 7. Oscillatory motor output in a n isolated ganglion. A:
Control: extension pattern from G4 elicited by stimulation of left G4-G5 pattern-initiating bundle. Abdominal nerve cord is otherwise intact. Arrow indicates onset of burst from excitatory motoneurons. B: G4 isolated: cyclical activity resulting from continuous stimulation of left G3-G4 bundle. Arrows indicate onsets of two consecutive cycles.
posed tha t the cyclic alternation of flexion and extension at each ganglion was produced by inter- ganglionic elements, termed flexor driver (FD) units, which were believed to function by simulta- neously activating the flexor excitor motoneurons and inhibiting the extensors. According to Kovac, termination of the activity burst in the FD unit dis- continued flexor motoneuron excitation and allowed activation of the extensor motoneurons by post- inhibitory rebound. Evidence favoring the existence of such FD neurons was provided from extracellu- lar recordings in the G4-G5 connective showing ap- proximately 600 msec activity bursts always leading the bursts of flexor excitor motoneurons in G by about 100 msec during continuous, unpatterned "command" fiber stimulation. Furthermore, stim- ulation of the FD units during command-driven ac- tivity succeed in resetting the phase of the cycle. Thirdly, according to the model, the FD units in the bilateral connectives were linked together with other FD units in the anterior, posterior, and lat- eral directions, thus forming a "mutually reinforcing" network. Evidence in support of this interganglionic- oscillatory network was provided by experiments showing that stimulation of any FD unit produced activity in each of the three other FD units (one per hemiconnective) associated with an abdom- inal ganglion. Also, cutting any one of the four
hemiconnectives to a ganglion resulted in a dimi- nution of the cyclic output at that ganglion. Nor- mal rhythmicity was not observed unless at least two ganglia wpre connected, with the exception of the terminal ganglion, G6, which has evolved as a fusion of two ganglia (Wine, '84). A mechanism in- volving a discrete oscillator in each ganglion was ruled out because individual, isolated ganglia did not appear capable of generating rhythmic outputs (Kovac, '74a,b).
Cyclic pattern genera tion: an alternative interpretation
The incomplete separation and, indeed, extensive overlap of flexion and extension in the oscillatory pattern at each abdominal ganglion (Moore and Larimer, '87, '88; Fig. 4 in this study) would seem to argue against the mechanism proposed by Kovac ('74b) to explain the cyclic patterning of the two antagonistic motor outputs. The Kovac mechanism requires simultaneous flexor motoneuron activa- tion and extensor motoneuron inhibition. Another difficulty with the Kovac model stems from the ab- sence of a mechanism which would inactivate the hypothetical FD units so that extension outputs could be released (i.e., a recurrent cyclic inhibitory loop or some other negative feedback mechanism was not included).
We have isolated small bundles of axons in the interganglionic connectives which burst in a cycli- cal fashion as found by Kovac ('74b) for the FD units (Fig. 3). However, contrary to the earlier work, there appear to be several different units activated in the bursts. This is in agreement with a recent intracel- lular study (Moore and Larimer, '87), which re- vealed several different morphological cell types functioning as pattern-initiating elements. Further- more, extensive dye-coupling among these cells sug- gested that they formed an electrically coupled network. All of the neurons filled by Moore and Larimer ('87) possessed axons coursing through the same ventrolateral margins of the connectives, where the PI axon bundles are always found. With continuous, unpatterned stimulation applied intra- cellularly to the PI interneurons or extracellularly to the PI bundles, the motor pattern cycles and is indistinguishable from that elicited by stimulation of the command neuron isolated from the anterior connectives. Because a patterned input is not re- quired for these cells to generate a patterned out- put, the interganglionic PI cells (Moore and Larimer, '87) are functionally equivalent to the cell that was earlier called the command neuron (Kovac, '74b). We propose that the command neuron is one mem- ber of an assembly of PI interneurons.

CYCLIC MOTOR PATTERN GENERATION IN CRAYFISH 413
It is not known which (or how many) of the PI interneurons are recruited in the intact animal dur- ing normal behavior. One interesting possibility is that the entire network is activated when any one PI neuron is stimulated to a certain level, as a re- sult of the excitatory coupling within this system. Kovac ('74a) has compared the abdominal motor programs elicited during normal backward walk- ing behavior and from electrical stimulation of the connectives and found them to be identical. Al- though any one of the PI interneurons is sufficient, when stimulated, to elicit the behavior, no single one also satisfies the criterion of necessity ascribed to "command neurons" (Kupfermann and Weiss, '78). These cells, thus, constitute a command sys- tem, with each PI interneuron capable of operat- ing as a command element within the system. One natural trigger for the behavior is the caudal pho- toreceptor (Edwards, '84). In response to bright light, the caudal photoreceptor discharges a high- frequency burst of spikes. If the frequency burst ex- ceeds 60 Hz, the stimulus is adequate to trigger the cyclic motor pattern (Simon and Edwards, '90). The output site for caudal photoreceptor activation of the pattern-initiating network most likely is ros- tral to the abdominal nerve cord (Edwards, '84) but below the brain (Simon and Edwards, '90). It is not known whether this rostral activation of the ab- dominal PI network is related to the network bias for descending signals observed in this study (Figs. 3,4).
One possible function of a multiple PI interneu- ron network might be to amplify a relatively weak input signal into a much stronger output signal. Supporting this hypothesis is the finding that the patterned motor program was elicited throughout the abdominal nerve cord by low frequency stim- uli or even by a single stimulus pulse (Fig. 1B). Amplification in the cyclic postural system pre- sumably could be the result of having multiple PI interneurons that are coupled to one another and converge on the ganglionic oscillators. For exam- ple, below-threshold stimuli from rostral and cau- dal sources along the cord were shown to converge on intermediate ganglia to trigger pattern genera- tion (Fig. 2).
Despite examining over 200 different isolated nerve cord preparations, we have found no inter- neurons possessing the characteristics of Kovac's ('74b) FD neurons. There are at least two possibil- ities that may explain the differences in our obser- vations and those of Kovac. First, a PI axon bundle (e.g., in the G4-G5 connective) will exhibit the prop- erties of an FD unit if stimulated at high frequency
(50 Hz) for a short duration: the major response will be an activation of flexion and a reduction of ex- tension in G5. However, if the stimulation is applied continuously, both flexion and extension outputs will cycle and, thus, the interneuron will no longer behave like an FD neuron. Second, there could have been co-activation of other premotor interneurons in the axon bundle. The axons of many premotor interneurons controlling flexion and extension mo- toneurons are found in the interganglionic connec- tives, including the ventrolateral area 85. Among them are pure flexion-producing and extension- producing interneurons (Miall and Larimer, '82b; Larimer and Jellies, '83; Larimer and Moore, '84; Jellies and Larimer, '85, '86) as well as flexion- suppressing and extension-suppressing interneu- rons (Larimer and Moore, '84; Moore and Larimer, '88) and general inhibitors which suppress both tonic extension and flexion simultaneously (Moore and Larimer, '88). All of these interneurons have been recorded intracellularly and many of them are identified cells. Various combinations of effects are possible if two or more of these interneurons are activated together. One possible way of duplicating the motor output of an FD neuron would be to si- multaneously activate a pure flexion-producing in- terneuron and a general inhibitor. The result would be a reduction in spontaneous extension activity but an enhancement of flexion activity because the flexion-producing interneuron would be able to at least partially override the suppressing effect of the general inhibitor. Once the extracellular stimula- tion stops, extension activity presumably could re- bound from inhibition and flexion could return to pre-stimulus, spontaneous levels. The same output pattern could theoretically occur if a flexion- producing and an extension-suppressing interneu- ron were co-activated.
Our experimental design was structured so as to eliminate the possible interfering effects of the tonic postural interneurons within the bundle or in ad- jacent areas. These interfering cells have a thresh- old frequency of 25-30 Hz for a detectable motor effect (Jellies and Larimer, '85) and our stimulus pulse trains were delivered at 10-15 Hz to the pattern-initiating bundles.
Properties of the pattern-initiating network The present study has revealed several proper-
ties of the cyclic pattern-initiating system that were not detected from intracellular recordings of indi- vidual PI interneurons (Moore and Larimer, '87). First, we now know that there are PI axons in ev- ery abdominal hemiconnective because unpatterned

414 D. MOORE AND J.L. LARIMER
stimulation to the ventrolateral portion of any hemiconnective will generate a normal cyclic pat- tern throughout the abdominal cord. Second, we found that no single PI axon projects the entire length of the abdominal nerve cord, since complete one-to-one, phase-locked impulses do not occur be- tween rostral and caudal PI bundles (Fig. 3). How- ever, we cannot discount the possible existence of PI-like interneurons in other (non-ventrolateral) portions of the abdominal nerve cord, although none have yet been discovered. Third, as demonstrated by the various hemiconnective cutting experiments (Figs. 4, 5 ) , the bilaterally organized PI interneu- rons are functionally coupled across all of the ab- dominal ganglia. This particular finding may not be significantly different from Kovac's ('74b) idea of a mutually reinforcing network. In the present scenario, the signals are carried by PI interneurons rather than by Kovac's hypothetical FD neurons, as noted earlier in the Discussion. Fourth, there exists a bias within the PI network for signal con- duction in the descending (caudal) direction. Sup- port for this conclusion is two-fold: (1) the presence of multiple spike amplitudes from descending sig- nals within PI bundles (Fig. 3), and (2) after lat- eral signal transfers across a ganglion, the cyclic motor pattern in the next rostral ganglion demon- strates a significant deficit in motor activity com- pared to normal, whereas the pattern in the next caudal ganglion shows little or no effect (Fig. 4). Fifth, this study provides evidence that the cyclic pattern, though weak, may be generated within an individual isolated abdominal ganglion (Fig. 71, con- trary to Kovac's ('74b) results.
Comparative aspects Because pattern generation in the cyclic postural
system arises partially from an interganglionic net- work of PI elements and partially from the activa- tion of individual ganglionic oscillators, the cyclic postural system is both central and distributed, much like the swimmeret neuronal network (Stein, '71). There are two swimmeret oscillators in each abdominal ganglion, each governing the activity of its own swimmeret (Hughes and Wiersma, '60). In contrast, the patterned outputs of the cyclic pos- tural system are bilaterally symmetrical, and persist as such after a variety of experimental manipula- tions (Fig. 61, suggesting that there may be a sin- gle oscillator controlling the postural pattern from each ganglion. If there are two oscillators per gan- glion, as in the swimmeret system, then the two are tightly coupled.
The functional pathways for the command sig-
nals in the swimmeret system descend bilaterally in the connectives and may transfer laterally across GI, G2, and G3 but not G4 and G5 (Wiersma and Ikeda, '64). In contrast, pattern-initiating signals in the cyclic postural system may cross any of the abdominal ganglia (Fig. 5).
Interganglionic command elements initiate both the swimmeret and cyclic postural behaviors. In the swimmeret system, there are several different "com- mand" neurons which are found in various, widely separate locations in the abdominal cord cross- section (Wiersma and Ikeda, '64; Davis and Ken- nedy, '72a). Similarly in the cyclic postural system, there are apparently several command elements (pattern-initiating interneurons) but, in contrast to the swimmeret system, these elements are all confined to a rather restricted ventrolateral mar- gin of the nerve cord. No single swimmeret command element produces the entire range of outputs ob- served, indicating that a group of these command elements must operate in concert to elicit normal swimmeret behavior (Davis and Kennedy, '72b). Likewise, tonic abdominal postures in crayfish are derived from the activation of constellation^^^ of flexion-producing and extension-producing inter- neurons (Jellies and Larimer, '86). In contrast, each pattern-initiating element in the cyclic postural sys- tem is apparently capable of generating the com- plete motor pattern. This capability may arise by virtue of the extensive coupling among the PI cells. Unlike central pattern generation in many other systems, the cyclic postural behavior requires con- tinuous stimulation to maintain rhythmicity. This may allow the behavior to take advantage of the oscillatory circuitry but at the same time retain the ability to exert more direct control over the initia- tion and termination of the motor act.
ACKNOWLEDGMENTS We thank J. Weeks, D.H. Edwards, J. Burdohan,
and L. Brewer for commenting on an earlier draft of this manuscript. We also appreciate the discus- sions of J. Jellies, W. Thompson, and D. Murchi- son as well as technical assistance from C-L. Hsieh. This project was supported by NIH Grant NS 05423 (J.L.L.) and by a grant from the University Re- search Institute of The University of Texas.
LITERATURE CITED Bowerman, R.F., and J.L. Larimer (1976) Command neurons
in crustaceans. Comp. Biochem. Physiol., 54:1-5. Davis, W.J., and D. Kennedy (1972a) Command interneurons
controlling swimmeret movements in the lobster. I. Types of effects on motoneurons. J. Neurophysiol., 35:13-19.

CYCLIC MOTOR PATTERN GENERATION IN CRAYFISH 415
Davis, W.J., and D. Kennedy (1972b) Command interneurons controlling swimmeret movements in the lobster. 11. Interac- tion of effects on motoneurons. J . Neurophysiol., 35:20-29.
Edwards, D.H. (1984) Crayfish extraretinal photoreception. I. Behavioural and motoneuronal responses to abdominal illu- mination. J. Exp. Biol., 165~223-238.
Evoy, W.H., and D. Kennedy (1967) The central nervous orga- nization underlying control of antagonistic muscles in the crayfish. I. Typesofcommand fibers. J. Exp. Zool., 165:223-238.
Evoy, W.H., D. Kennedy, and D.M. Wilson (1967) Discharge pat- terns of neurons supplying tonic abdominal flexor muscles in the crayfish. J . Exp. Biol.,46:393-411.
Fields, H.L. (1966) Proprioceptive control of posture in the cray- fish abdomen. J . Exp. Biol., 44:455-468.
Fields, H.L., W.H. Evoy, and D. Kennedy (1967) Reflex role played by efferent control of an invertebrate stretch receptor. J . Neurophysiol., 30:859-875.
Hughes, G.M., and C.A.G. Wiersma (1960) The coordination of swimmeret movements in the crayfish, Procambarus clarkii. J . Exp. Biol., 37:657-670.
Jellies, J., and J.L. Larimer (1985) Synaptic interactions be- tween neurons involved in the production of abdominal pos- ture in crayfish. J . Comp. Physiol. [A], 1562361-873.
Jellies, J . , and J.L. Larimer (1986) Activity of crayfish ab- dominal-positioning interneurons during spontaneous and sensory-evoked movements. J . Exp. Biol., 120:173-188.
Kennedy, D., and I. Takeda (1965) Reflex control of abdomi- nal flexor muscles in the crayfish. 11. The tonic system. J. Exp. Biol., 43:229-246.
Kennedy, D., W.H. Evoy, and J.T. Hanawalt (1966) Release of coordinated behavior in crayfish by single central neurons. Science, 154 :9 17-9 19.
Kennedy, D., W.H. Evoy, B. Dane, and J.T. Hanawalt (1967) The central nervous organization underlying control of antagonistic muscles in the crayfish. 11. Coding of position by command fibers. J. Exp. Zool., 165:239-248.
Kovac, M. (1974a) Abdominal movements during backward walking in crayfish. I. Properties of the motor program. J . Comp. Physiol., 95:79-94.
Kovac, M. (1974b) Abdominal movements during backward walking in crayfish. 11. The neuronal basis. J . Comp. Physiol., 95:79-94.
Kupfermann, I., and K.R. Weiss (1978) The command neuron concept. Behav. Brain Sci., 1:3-39.
Larimer, J.L. (1988) The command hypothesis: a new look us- ing an old example. Trends Neurosci., 11:506-510.
Larimer, J.L., and D. Kennedy (1969) The central nervous con- trol of complex movements in the uropods of crayfish. J. Exp. Biol., 51:135-150.
Larimer, J.L., and J . Jellies (1983) The organization of flexion- evoking interneurons in the abdominal nerve cord of the cray- fish, Procambarus clarkii. J . Exp. Zool., 226:341-351.
Larimer, J.L., and D. Moore (1984) Abdominal positioning in- terneurons in crayfish: projections to and activation by higher CNS centers. J. Exp. Zool., 23O:l-10.
Miall, R.C., and J.L. Larimer (1982a) Central organization of abdominal posture motoneurons: connectivity and command fiber inputs. J. Exp. Zool., 224:45-56.
Miall, R.C., and J.L. Larimer (198223) Interneurons involved in abdominal posture in crayfish structure, function, and com- mand fiber responses. J . Comp. Physiol., 148:159-173.
Moore, D., and J.L. Larimer (1987) Neural control of a cyclic postural behavior in the crayfish, Procambarus clarkii: the pattern-initiating interneurons. J. Comp. Physiol. [A], 160: 169-179.
Moore, D., and J.L. Larimer (1988) Interactions between the tonic cyclic postural motor programs in the crayfish abdomen. J. Comp. Physiol. [A], 163:187-199.
Murphy, B.F., McAnelly, M.L., and J.L. Larimer (1989) Abdom- inal positioning interneurons in crayfish: participation in be- havioral acts. J . Comp. Physiol. [A], 165:461-470.
Page, C.H. (1982) Control of posture. In: The Biology of Crus- tacea. D.C. Sandeman and H.L. Atwood, eds. Academic Press, New York, vol. 4, pp. 33-59.
Simon, T.W., and D.H. Edwards (1990) Light-evoked walking in crayfish: behavioral and neuronal responses triggered by thecaudal photoreceptor. J . Comp. Physiol. [A], 166:745-755.
Sokolove, P.G., and W.G. Tatton (1975) Analysis of postural mo- toneuron activity in crayfish abdomen. I. Coordination by premotor connections. J. Neurophysiol. 38:313-331.
Stein, P.S.G. (1971) Intersegmental coordination of swim- meret motoneuron activity in crayfish. J . Neurophysiol., 34: 3 10-3 18.
van Harreveld, A. (1936) A physiological solution for freshwa- ter crustaceans. Proc. SOC. Exp. Biol., 34:428-432.
Wiersma, C.A.G., and G.M. Hughes (1961) On the functional anatomy of neuronal circuits in the abdominal cord of the crayfish, Procambarus clarkii (Girard). Comp. Biochem. Physiol., 12:509-525.
Wiersma, C.A.G., and K. Ikeda (1964) Interneurons com- manding swimmeret movements in the crayfish, Pro- cambarus clarkii (Girard). Comp. Biochem. Physiol., 12: 509-525.
Williams, B.J., and J.L. Larimer (1981) Neuroal pathways of reflex-evoked behaviors and command systems in the abdomen of the crayfish. J . Comp. Physiol., 143:27-42.
Wine, J.J. (1984) The structural basis of an innate behavioural pattern. J . Exp. Biol., 112:283-319.