complete primary structure of the molt-inhibiting hormone (mih) of the mexican crayfish procambarus...
TRANSCRIPT

Peptides, Vol. 17, No. 3, pp. 367-374, 1996 Copyright 8 1996 Elsevier Science Inc. printed in the USA. All rights reserved
0196-9781/96 $15.00 + .OO
ELSEVIER PII Sol%-9781(96)00010-l
Complete Primary Structure of the Molt-Inhibiting Hormone (MIH) of the Mexican Crayfish
Procambarus bouvieri (Ortmann)
MANUEL B. AGU:[LAR,* ROCCO FALCHETTO,t JEFFREY SHABANOWITZ,? DONALD F. HUNTt$ AND ALBERT0 HUBERMAN * ’
*Department of Biochemistry, Institute National de la Nutricidn “Salvador Zubirdn, ” Vasco de Quiroga 15, Tlalpan, Mexico City 14000, Mexico
?Department of Chemistry, Biological Mass Spectrometry Laboratory, and SDepartment of Pathology, University of Virginia, McCormick Road, Charlottesville, VA 22901
Received 21 September 1995
AGUILAR, M. B., R. FALCHETTO, J. SHABANOWITZ, D. F. HUNT AND A. HUBERMAN. Complete pn’mury structure of the molt-inhibiting hormone (MIH) of the Mexican cruyfih Procambarus bouvieri (Urtmann). PEPTIDES 17(3) 367-374, 1996.-The amino acid sequence of MIH was elucidated by means of digestions with specific proteases, manual Edman degra- dation, and mass spectrometry. MIH consists of a 72-residue peptide chain (molecular mass 8322 Da) with six cysteines forming three disulfide bridges that connect residues 7-43, 23-39, and 26-52. It has blocked N- and C-termini and lacks tryptophan, histidine, and methionine. MIH shows striking similarity to the crustacean hyperglycemic hormone (CHH) isomorphs of Procam- barus bouvieri (90% identity) and to the MM from Homarus americanus (79% identity) and Penaeus vannamei (46% identity). It is also related to the MIH from Carcinus maenas (28% identity) and Callinectes sapidus (28% identity).
Molt-inhibiting hormone Neuropeptide Crustacean hyperglycemic hormone
Sequence Procambarus bouvieri Crayfish
CRUSTACEANS possess a major neurosecretory system that consists of a group of cell somata that are clustered together in the medulla terminalis X-organ (MTXO) and their bulbous ax- onic terminals that constitute the sinus gland (SG). The SG is a neurohemal organ that releases into the hemolymph a number of peptide hormones involved in the regulation of many physiolog- ical processes, such as external color changes, gonadal devel- opment, light adaptation, glucose metabolism, molt cycle, and osmoregulation, among, others [for recent reviews see (7,16,31,37)].
(33) have been determined. They show a very high degree of sequence similarity to the crustacean hyperglycemic hormones from several crustaceans (2,25,29,30,34,43,47) and belong to the same neuropeptide family that includes the vitellogenesis-inhib- iting hormone ( 1,38), which has been found only in crustaceans.
Here we report the complete amino acid sequence of the MIH of the Mexican crayfish Procambarus bouvieri, and compare it with the CHH from the same organism and with other MIH.
METHOD
A clear correlation has been observed between increased cir- culating ecdysteroids, and proecdysis and molting. Crustacean ecdysteroids are synthesized in the Y-organs, mainly as o-ec- dysone, which is later converted to 20-OH-ecdysone (B-ecdy- sone) in the hemolymph and target tissues. The SG contains a factor (currently known as the molt-inhibiting hormone) that re- presses the rise in circulating ecdysteroids that follows eyestalk ablation in intermolt animals. Furthermore, it has been demon- strated that SG extracts will repress ecdysteroid biosynthesis by cultured Y-organs [for recent reviews see (8,9,20)].
Animals, Sinus Glands, Crude Extracts, Purij%ation Protocol, and Bioassay
Adult crayfish were obtained from local fishermen in Uru- apan, State of Michoacan The procedures for keeping them at the laboratory, and for eyestalk ablation (under cold anesthesia) and storage, SG dissection, crude extract preparation, and puri- fication of MIH by means of RP-HPLC on a PBondapak-Phenyl column (Waters, Mildford, MA) have been described previously (24).
The MIH primary structures of the lobster Homarus ameri- The average yield of pure MIH was 18 ng/SG as determined canus (lo), the crab Carcinus maenas (44), the white shrimp by means of the 2,4,6-trinitrobenzenesulfonic acid (TNBS, Penaeus vannamei (40), and the blue crab Callinectes sapidus Sigma Chemical Co., St. Louis, MO) method (19).
’ Requests for reprints should be addressed to Dr. Albert0 Huberman.
367

368 AGUILAR ET AL.
The bioassay consisted of the inhibition by MIH of ecdyster- oid biosynthesis in vitro by Y-organs of Orconectes limosus ( 15,27), as described in Huberman and Aguilar (24). The bio- assay for hyperglycemic activity was done as described in Hub- erman and Aguilar (2 1) .
Reagents and Glassware
All reagents were of analytical or sequencing grade. RP-HPLC solvents were from Fisher Scientific (Fair Lawn, NJ). Double- distilled water was further purified by passing through a Sybron- Barnstead (Boston, MA) NANOpure-A system, and only water with a resistivity above 10 Megohm/cm was used. 4-Vinylpyridine (VP, Sigma), N-ethylmorpholine (NEM, Pierce, Rockford, IL), and triethylamine (TEA, Fisher) were distilled. TNBS was re- crystallized. Glassware was calcined at 550°C for 4 h.
Chemical Modzjication of Residues
Cystine residues were reduced with dithiothreitol (Sigma) af- ter dissolving native MB-I in 0.1 M Tris(hydroxymethy1) ami- nomethane-HCl (Sigma), containing 1 mM ethylenediaminetet- raacetic acid, disodium salt (EDTA, Sigma), pH 8.3, or in 0.5 M NEM-acetate, pH 8.3, and alkylated with iodoacetic acid (Sigma) (3 ) or VP (4 1) , respectively. The carboxymethylated (CM-MM) and pyridylethylated (PE-MIH) derivatives were pu- rified by means of RP-HPLC on a Nova-Pak Cl8 column ( 150 X 3.9 mm i.d., Waters) as reported (24).
End-Group Analysis
The 1-dimethylaminonaphthalene-5-sulfonyl (dansyl) chlo- ride method (17) was used as described (3). The dansyl deriv- atives (DNS-aa) of the amino acids were identified by means of thin-layer chromatography (TLC) on 3 X 3 cm polyamide- coated aluminium sheets (Schleicher & Schuell GmbH, Keene, NH), employing solvent systems I (1.5%, v/v, aqueous formic acid), II (toluene:acetic acid, 10: 1, v/v), and III (ethyl ace- tate:methanol:acetic acid, 20: 1: 1, by vol.) (46). As the basic DNS-aa gave one coincident spot after the three previous steps, a fourth step consisting in solvent system IV (50 mM Na3P0,:ethanol, 3:1, v/v) (18) was applied to identify DNS- Arg, e-DNS-Lys and DNS-His.
Amino Acid Analysis
Qualitative amino acid composition was determined in frag- ments derived from enzymatic digestions as follows. Acid hy- drolysis was carried out under nitrogen at 145°C for 1 h in 6 M HCl containing 2% (v/v) thioglycolic acid (Sigma) (35). Iden- tification of the amino acid residues was performed by means of the dansyl chloride method and thin-layer chromatography (see End-Group Analysis). Quantitative amino acid analysis of tryp- tic and chymotryptic peptides was performed as described in leg- ends to Tables 1 and 2. Amino acid standard H mixture (Pierce) was processed identically. Precolumn derivatization with phen- ylisothiocyanate (Pierce), and RP-HPLC quantitation of the phenylthiocarbamoyl amino acid derivatives (PTC-aa) on two Ultrasphere-ODS columns (250 x 4.6 mm i.d., Beckman Instru- ments, San Ramon, CA) in tandem or one Nova-Pack C 18 (300 X 3.9 mm i.d., Waters) were conducted essentially as described (6), with only minor adjustments ( 1).
Enzymatic Cleavages
Trypsin [from bovine pancreas; N-tosyl-L-phenylalanyl chlo- romethyl ketone (TPCK)-treated, Sigma]. CM-MIH was sus-
pended (with the aid of sonication) in 200 ~1 of 0.1 M NHHC03, pH 8.3, and incubated, under nitrogen, at 37°C for 90 min with an enzyme:substrate ratio of 1:25 (w/w). The resulting tryptic pep- tides were purified by means of RP-HPLC on an Ultrasphere-ODS column (Beckman) as reported (23,24). Native MM was dis- solved in 0.1 M Tris-HCl containing 5% (v/v) acetonitrile (MeCN), and incubated at 37°C for 24 h at an enzyme:substrate ratio of 1:20 ( w/w ) . For the determination of the disulfide bridges, the tryptic peptides of native MIH were purified on a 150 X 3.9 mm i.d. Nova-Pak Cl8 column (see legend to Fig. 5).
a-chymotrypsin [from bovine pancreas; N-tosyl+lysyl chlo- romethyl ketone (TLCK)-treated, Sigma]. PE-MIH was dis- solved in 0.1 M NH4HC03, pH 8.0, containing 1 mM CaCl, and 0.01% (w/v) thiodiglycol (Pierce), and incubated at 30°C for 48 h with an enzyme:substrate ratio of 1:50 (w iw ) . The resulting chymotryptic peptides (QTPs) were purified as described for the tryptic peptides, on an Ultrasphere-ODS column (see legend to Fig. 3).
Protease V8 (from Staphylococcus aureus, Sigma). Tryptic peptide T2 + T5 + T7 from native MIH was dissolved in 50 mM KH,PO,, pH 7.8, and incubated at 37°C for 72 h at an en- zyme:substrate ratio of 1: 15 (w/w). The fragments produced were recovered as stated above for the tryptic peptides on an Ultrasphere-ODS column.
Pyroglutamate aminopeptidase (PGAP, from Bacillus amy- loliquefaciens, Sigma). Tryptic peptide T4 was dissolved in 0.1 M K,HPO,, 0.01 M EDTA.4Na, 0.01 M P-mercaptoethanol (Sigma) pH 8.0, and incubated at 37°C for 4 h with an en- zyme:substrate ratio of 1: 1 (w/w). The resulting peptide was purified on an Ultrasphere-ODS column as described previously.
Carboxypeptidase B [ CPB, from hog pancreas; diisopropyl- jluorophosphate (DFP) -treated, Sigma]. Tryptic peptide T3 was dissolved in 0.5 M NEM-acetate, pH 8.3, and incubated at 37°C for 1 h at an enzyme:substrate ratio of 1:25 (w/w). The released amino acids were identified by means of the dansyl chloride method and TLC as described in End-Group Analysis. An ex- traction with ethyl acetate was performed before TLC.
Carboxypeptidase Y (CPY, from baker’s yeast, Sigma). Chy- motryptic peptide ChyT4 was dissolved in 50 ,LL~ 0.1 M pyridine- acetate buffer solution, pH 5.5 (BS). Five microliters of 40 ng/ ~1 aqueous CPY was added and the mixture incubated at 37°C for 30 min. Ten microliters of concentrated acetic acid was used to stop the reaction and the hydrolysate was vacuum dried. Con- trol tubes were: BS, BS plus CPY, and human glucagon plus BS plus CPY. Amino acid residues released from all the samples were identified by means of dansylation and TLC.
Microsequencing
A film method with polybrene (4 1) for manual Edman mi- crosequencing was conducted with only minor adjustments. The phenylthiohydantoin derivatives (PTH-aa) were identified by means of RP-HPLC on two Ultrasphere-ODS columns (250 x 4.6 mm i.d., Beckman) in tandem, with an ammonium acetate/ acetonitrile elution system (36), or on one Nova-Pak C 18 col- umn (300 X 3.9 mm i.d., Waters) with solution A: 26 mM am- monium acetate, pH 4.8, and solution B: acetonitrile.
Mass Spectrometry
The molecular mass of MM was measured by matrix-assisted laser-desorption/ionization time-of-flight (MALDI-TOF) mass spectrometry (28) on a “home-made” instrument described by Amott et al. (4). The sample was dissolved in 30% acetonitrile, 0.1% trifluoroacetic acid in water containing an excess of matrix (a-cyano-4-hydroxycimramic acid), The mass ratio between pro-
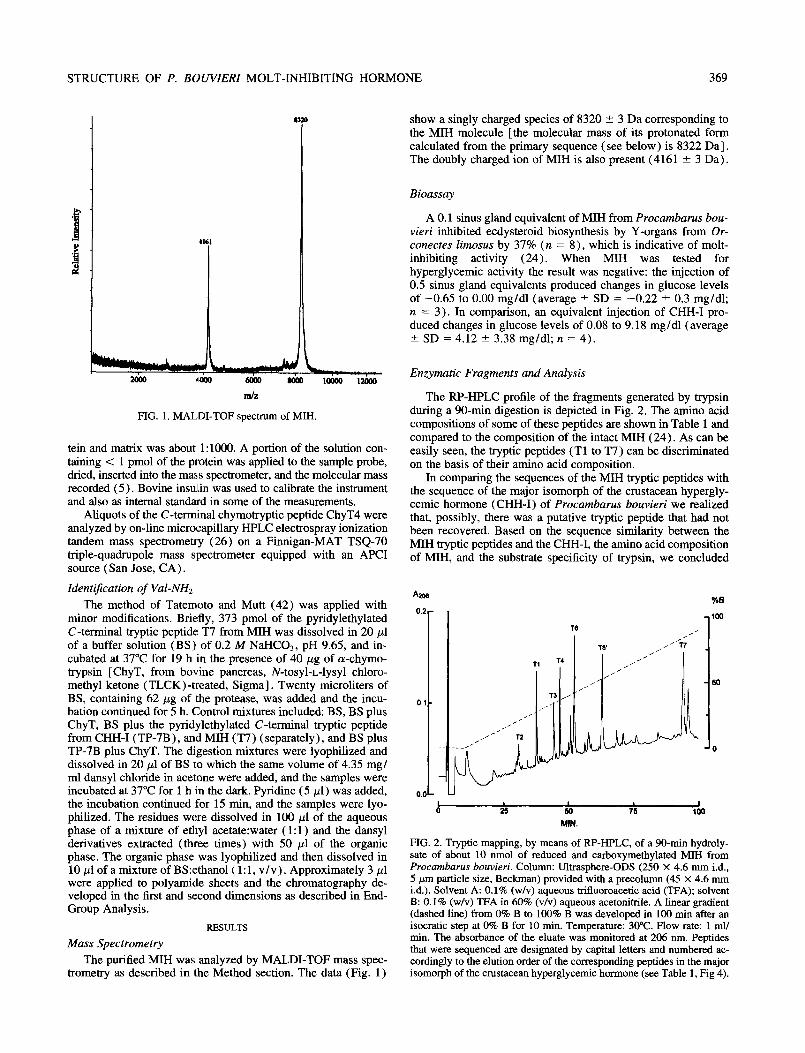
STRUCTURE OF P. BOUVZERZ MOLT-INHIBITING HORMONE 369
FIG. 1. MALDI-TOF spectrum of MIH.
tein and matrix was about 1:lOOO. A portion of the solution con- taining < 1 pmol of the protein was applied to the sample probe, dried, inserted into the mass spectrometer, and the molecular mass recorded (5). Bovine insulin was used to calibrate the instrument and also as internal standard in some of the measurements.
Aliquots of the C-terminal chymotryptic peptide ChyT4 were analyzed by on-line microcapillary HPLC electrospray ionization tandem mass spectrometry (26) on a Finmgan-MAT TSQ-70 triple-quadrupole mass spectrometer equipped with an APCI source (San Jose, CA).
Identijication of Val-NH2 The method of Tatemoto and Mutt (42) was applied with
minor modifications. Briefly, 373 pmol of the pyridylethylated C-terminal tryptic peptide T7 from MIH was dissolved in 20 1.11 of a buffer solution (BS) of 0.2 M NaHC03, pH 9.65, and in- cubated at 37°C for 19 h in the presence of 40 ,ug of cr-chymo- trypsin [ ChyT, from bovine pancreas, N-tosyl+lysyl chloro- methyl ketone (TLCK) -treated, Sigma]. Twenty microliters of BS, containing 62 hg of the protease, was added and the incu- bation continued for 5 h. Control mixtures included: BS, BS plus ChyT, BS plus the pyridylethylated C-terminal tryptic peptide from CHH-I (TP-7B), and MB-I (T7) (separately), and BS plus TP-7B plus ChyT. The digestion mixtures were lyophilized and dissolved in 20 ~1 of BS to which the same volume of 4.35 mg/ ml dansyl chloride in acetone were added, and the samples were incubated at 37°C for 1 h in the dark. Pyridine (5 ~1) was added, the incubation continued for 15 min, and the samples were lyo- philized. The residues were dissolved in 100 ~1 of the aqueous phase of a mixture of ethyl acetate:water ( 1:l) and the dansyl derivatives extracted (three times) with 50 ~1 of the organic phase. The organic phase was lyophilized and then dissolved in 10 ,ul of a mixture of BS:ethanol (l:l, v/v). Approximately 3 ~1 were applied to polyamide sheets and the chromatography de- veloped in the first and second dimensions as described in End- Group Analysis.
Mass Spectrometry
RESULTS
The purified MM was analyzed by MALDI-TOF mass spec- trometry as described in the Method section. The data (Fig. 1)
show a singly charged species of 8320 2 3 Da corresponding to the MIH molecule [the molecular mass of its protonated form calculated from the primary sequence (see below) is 8322 Da]. The doubly charged ion of MIH is also present (4161 + 3 Da).
Bioassay
A 0.1 sinus gland equivalent of MIH from Procambarus bou- vieri inhibited ecdysteroid biosynthesis by Y-organs from Or- conectes limosus by 37% (n = 8), which is indicative of molt- inhibiting activity (24). When MM was tested for hyperglycemic activity the result was negative: the injection of 0.5 sinus gland equivalents produced changes in glucose levels of -0.65 to 0.00 mg/dl (average + SD = -0.22 + 0.3 mg/dl; n = 3). In comparison, an equivalent injection of CHH-I pro- duced changes in glucose levels of 0.08 to 9.18 mg/dl (average + SD = 4.12 ? 3.38 mg/dl; n = 4).
Enzymatic Fragments and Analysis
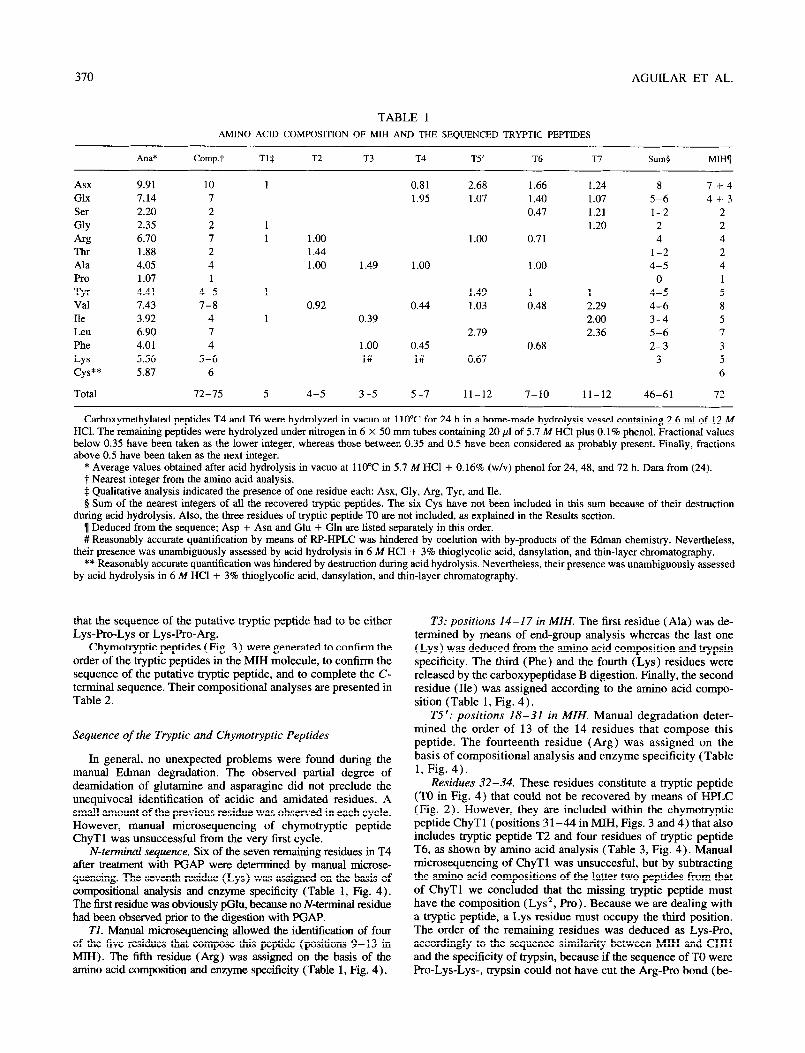
The RP-HPLC profile of the fragments generated by trypsin during a 90min digestion is depicted in Fig. 2. The amino acid compositions of some of these peptides are shown in Table 1 and compared to the composition of the intact MIH (24). As can be easily seen, the tryptic peptides (Tl to T7) can be discriminated on the basis of their amino acid composition.
In comparing the sequences of the MIH tryptic peptides with the sequence of the major isomorph of the crustacean hypergly- cemic hormone (CHH-I) of Procambanrs bouvieri we realized that, possibly, there was a putative tryptic peptide that had not been recovered. Based on the sequence similarity between the MIH tryptic peptides and the CHH-I, the amino acid composition of MIH, and the substrate specificity of trypsin, we concluded
0 .l -
%B
-100 TO
/,’ ,’
TO4 ,/’ n
C’ /’
/-’ ,’ -50
u: 0
I I I 0 26 w 76 100
MIN.
PIG. 2. Tryptic mapping, by means of RP-HPLC, of a 90-min hydroly- sate of about 10 nmol of reduced and carboxymethylated MM from Procambarns bouvieri. Column: Ultrasphere-ODS (250 X 4.6 mm id., 5 pm particle size, Beckman) provided with a precolumn (45 x 4.6 mm i.d.). Solvent A: 0.1% (w/v) aqueous trifluoroacetic acid (TFA); solvent B: 0.1% (w/v) TFA in 60% (v/v) aqueous acetonitrile. A linear gradient (dashed line) from 0% B to 100% B was developed in 100 min after an isocratic step at 0% B for 10 min. Temperature: 30°C. Flow rate: 1 ml/ min. The absorbance of the eluate was monitored at 206 nm. Peptides that were sequenced are designated by capital letters and numbered ac- cordingly to the elution order of the corresponding peptides in the major isomorph of the crustacean hyperglycemic hormone (see Table 1, Fig 4).
370
TABLE 1
AMINO ACID COMPOSITION OF MIH AND THE SEQUENCED TRYPTIC PEPTIDES
AGUILAR ET AL.
A& C0mp.t TlS T2 T3 T4 T5’ T6 T7 SWIG MIHT
Asx 9.91 10 1 0.81 2.68 1.66 1.24 8 7-k-4
Glx 7.14 7 1.95 1.07 1.40 1.07 5-6 4+3
Ser 2.20 2 0.47 1.21 l-2 2
Gly 2.35 2 1 1.20 2 2
Arg 6.70 7 1 1 .oo 1.00 0.71 4 4
TlU 1.88 2 1.44 l-2 2
Ala 4.05 4 1.00 1.49 1 .oo 1 .oo 4-5 4
Pro 1.07 1 0 1
Tyr 4.41 4-5 1 1.49 1 1 4-5 5
Val 7.43 7-8 0.92 0.44 1.03 0.48 2.29 4-6 8
Ile 3.92 4 1 0.39 2.00 3-4 5
Leu 6.90 7 2.79 2.36 5-6 7
Phe 4.01 4 1 .oo 0.45 0.68 2-3 3
LYS 5.56 5-6 l# 1# 0.67 3 5
cys** 5.87 6 6
Total 72-75 5 4-5 3-5 5-7 11-12 7-10 11-12 46-61 72
Carboxymethylated peptides T4 and T6 were hydrolyzed in vacua at 110°C for 24 h in a home-made hydrolysis vessel containing 2.6 ml of 12 M HCl. The remaining peptides were hydrolyzed under nitrogen in 6 X 50 mm tubes containing 20 ~1 of 5.7 M HCl plus 0.1% phenol. Fractional values below 0.35 have been taken as the lower integer, whereas those between 0.35 and 0.5 have been considered as probably present. Finally, fractions above 0.5 have been taken as the next integer.
* Average values obtained after acid hydrolysis in vacua at 110°C in 5.7 M HCl + 0.16% (w/v) phenol for 24,48, and 72 h. Data from (24). t Nearest integer from the amino acid analysis. $ Qualitative analysis indicated the presence of one residue each: Asx, Gly, Arg, Tyr, and Be. 8 Sum of the nearest integers of al1 the recovered tryptic peptides. The six Cys have not been included in this sum because of their destruction
during acid hydrolysis. Also, the three residues of tryptic peptide TO are not included, as explained in the Results section. 1 Deduced from the sequence; Asp + Asn and Glu + Gln are listed separately in this order. # Reasonably accurate quantification by means of RP-HPLC was hindered by coelution with by-products of the Edrnan chemistry. Nevertheless,
their presence was unambiguously assessed by acid hydrolysis in 6 M HCI + 3% thioglycolic acid, dansylation, and thin-layer chromatography. ** Reasonably accurate quantification was hindered by destruction during acid hydrolysis. Nevertheless, their presence was unambiguously assessed
by acid hydrolysis in 6 M HCl + 3% thioglycolic acid, dansylation, and thin-layer chromatography.
that the sequence of the putative tryptic peptide had to be either Lys-Pro-Lys or Lys-Pro-Arg.
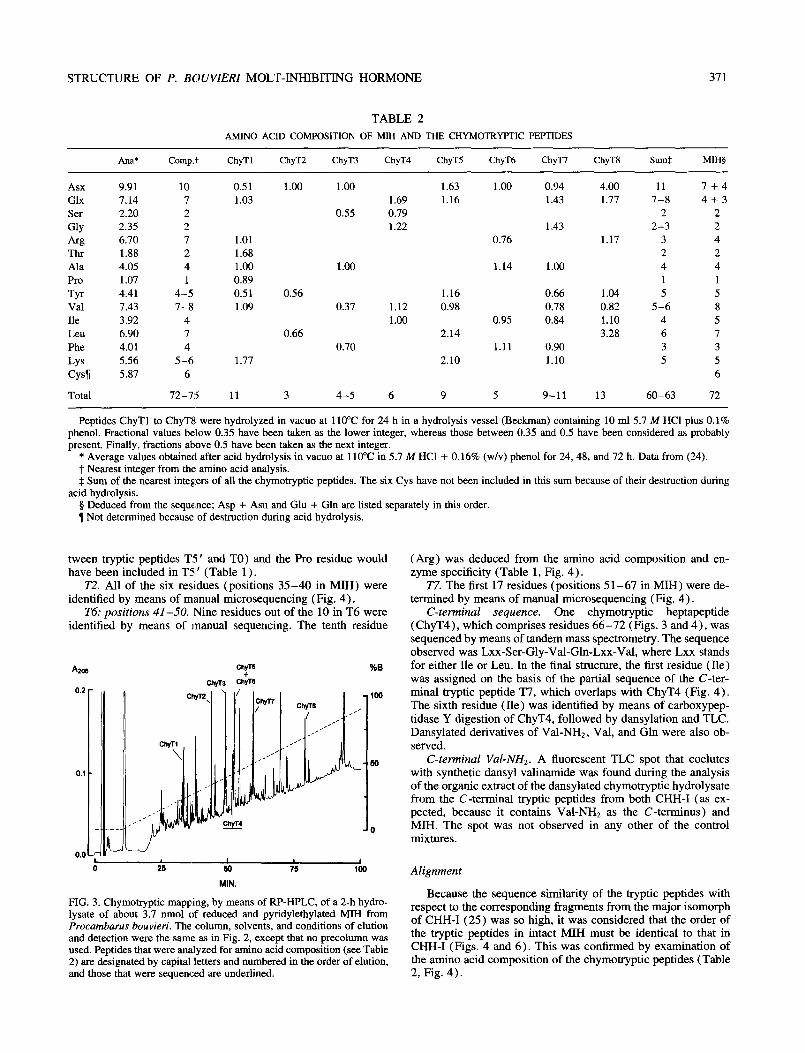
Chymotryptic peptides (Fig. 3) were generated to confirm the order of the tryptic peptides in the MIH molecule, to confirm the sequence of the putative tryptic peptide, and to complete the C- terminal sequence. Their compositional analyses are presented in Table 2.
Sequence of the Tryptic and Chymotryptic Peptides
In general, no unexpected problems were found during the manual Edman degradation. The observed partial degree of deamidation of glutamine and asparagine did not preclude the unequivocal identification of acidic and amidated residues. A small amount of the previous residue was observed in each cycle. However, manual microsequencing of chymotryptic peptide ChyTl was unsuccessful from the very first cycle.
N-terminal sequence. Six of the seven remaining residues in T4 after treatment with PGAP were determined by manual microse- quencing. The seventh residue (Lys) was assigned on the basis of compositional analysis and enzyme specificity (Table 1, Fig. 4). The first residue was obviously pGlu, because no N-terminal residue had been observed prior to the digestion with PGAP.
Tl. Manual microsequencing allowed the identification of four of the five residues that compose this peptide (positions 9-13 in MIH). The fifth residue (Arg) was assigned on the basis of the amino acid composition and enzyme specificity (Table 1, Fig. 4).
T3: positions 14-17 in MIH. The first residue (Ala) was de- termined by means of end-group analysis whereas the last one (Lys) was deduced from the amino acid composition and trypsin specificity. The third (Phe) and the fourth (Lys) residues were released by the carboxypeptidase B digestion. Finally, the second residue (Be) was assigned according to the amino acid compo- sition (Table 1, Fig. 4).
TS’: positions 18-31 in MIH. Manual degradation deter- mined the order of 13 of the 14 residues that compose this peptide. The fourteenth residue (Arg) was assigned on the basis of compositional analysis and enzyme specificity (Table 1, Fig. 4).
Residues 32-34. These residues constitute a tryptic peptide (TO in Fig. 4) that could not be recovered by means of HPLC (Fig. 2). However, they are included within the chymotryptic peptide ChyTl (positions 3 l-44 in MIH, Figs. 3 and 4) that also includes tryptic peptide T2 and four residues of tryptic peptide T6, as shown by amino acid analysis (Table 3, Fig. 4). Manual microsequencing of ChyTl was unsuccesful, but by subtracting the amino acid compositions of the latter two peptides from that of ChyTl we concluded that the missing tryptic peptide must have the composition (Lys*, Pro). Because we are dealing with a tryptic peptide, a Lys residue must occupy the third position. The order of the remaining residues was deduced as Lys-Pro, accordingly to the sequence similarity between MIH and CHH and the specificity of trypsin, because if the sequence of TO were Pro-Lys-Lys-, trypsin could not have cut the Arg-Pro bond (be-
STRUCTURE OF P. BOUVIERZ MOLT-INHIBITING HORMONE 371
TABLE 2
AMINO ACID COMPOSITION OF MIH AND THE CHYMOTRYPTIC PEPTIDES
Ad Camp.” ChyTl ChyT2 ChyT3 ChyT4 ChyTS ChyT6 ChyT7 ChyTB SllmS MIH$
Asx 9.91 10 0.51 1.00 1.00 1.63 1 .oo 0.94 4.00 11 7+4 Glx 7.14 7 1.03 1.69 1.16 1.43 1.77 7-8 4+3 Ser 2.20 2 0.55 0.79 2 2 Gly 2.35 2 1.22 1.43 2-3 2 Arg 6.70 7 1.01 0.76 1.17 3 4 Thr 1.88 2 1.68 2 2 Ala 4.05 4 1.00 1 .oo 1.14 1.00 4 4 Pro 1.07 1 0.89 1 1
Tyr 4.41 4-5 0.51 0.56 1.16 0.66 1.04 5 5 Val 7.43 7-8 1.09 0.37 1.12 0.98 0.78 0.82 5-6 8 Ile 3.92 4 1.00 0.95 0.84 1.10 4 5 Leu 6.90 7 0.66 2.14 3.28 6 7 Phe 4.01 4 0.70 1.11 0.90 3 3
Lys 5.56 5-6 1.77 2.10 1.10 5 5 Cysll 5.87 6 6
Total 72-75 11 3 4-5 6 9 5 9-11 13 60-63 72
Peptides ChyTl to ChyT8 were hydrolyzed in vacua at 110°C for 24 h in a hydrolysis vessel (Beckman) containing 10 ml 5.7 M HCl plus 0.1% phenol. Fractional values below 0.35 have been taken as the lower integer, whereas those between 0.35 and 0.5 have been considered as probably present. Finally, fractions above 0.5 have been taken as the next integer.
* Average values obtained after acid hydrolysis in vacua at 110°C in 5.7 M HCl + 0.16% (w/v) phenol for 24, 48, and 72 h. Data from (24). t Nearest integer from the amino acid analysis. $ Sum of the nearest integers of all the chymotryptic peptides. The six Cys have not been included in this sum because of their destruction during
acid hydrolysis. 5 Deduced from the sequence; Asp + Asn and Glu + Gln are listed separately in this order. 1 Not determined because of destruction during acid hydrolysis.
tween tryptic peptides T5 ’ and TO) and the Pro residue would have been included in T5’ ’ (Table 1) .
72. All of the six residues (positions 35-40 in MIH) were identified by means of manual microsequencing (Fig. 4).
T6: positions 41-50. Nine residues out of the 10 in T6 were identified by means of manual sequencing. The tenth residue
FIG. 3. Chymotryptic mapping, by means of RP-HPLC, of a 2-h hydro- lysate of about 3.7 nmol of reduced and pyridylethylated MIH from Procambarus bouvieri. The column, solvents, and conditions of elution and detection were the same as in Fig. 2, except that no precolumn was used. Peptides that were analyzed for amino acid composition (see Table 2) are designated by capital letters and numbered in the order of elution, and those that were sequenced are underlined.
(Arg) was deduced from the amino acid composition and en- zyme specificity (Table 1, Fig. 4).
27. The first 17 residues (positions 51-67 in MIH) were de- termined by means of manual microsequencing (Fig. 4).
C-terminal sequence. One chymotryptic heptapeptide (ChyT4), which comprises residues 66-72 (Figs. 3 and 4), was sequenced by means of tandem mass spectrometry. The sequence observed was Lxx-Ser-Gly-Val-Gln-Lxx-Val, where Lxx stands for either Ile or Leu. In the final structure, the first residue (Ile) was assigned on the basis of the partial sequence of the C-ter- minal tryptic peptide T7, which overlaps with ChyT4 (Fig. 4). The sixth residue (Ile) was identified by means of carboxypep- tidase Y digestion of ChyT4, followed by dansylation and TLC. Dansylated derivatives of Val-NH2, Val, and Gln were also ob- served.
C-terminal Val-NH,. A fluorescent TLC spot that coelutes with synthetic dansyl valinamide was found during the analysis of the organic extract of the dansylated chymotryptic hydrolysate from the C-terminal tryptic peptides from both CHH-I (as ex- pected, because it contains Val-NH2 as the C-terminus) and MIH. The spot was not observed in any other of the control mixtures.
Alignment
Because the sequence similarity of the tryptic peptides with respect to the corresponding fragments from the major isomorph of CHH-I (25) was so high, it was considered that the order of the tryptic peptides in intact MIH must be identical to that in CHH-I (Figs. 4 and 6). This was confirmed by examination of the amino acid composition of the chymotryptic peptides (Table 2, Fig. 4).
372 AGUILAR ET AL.
,----_-___--_________ChyT7-__________________,,________c~y*~______, ,________________~~______-______,,_________~~_______,,_______~~___ pGlu-Val-Phe-Asp-Gln-Ala-Cys-Lys-Gly-Ile-Tyr-Asp-A~g-Ala-Ile-Ph~- ___ __ 7 --7 -- , __, __, __, ___ __, __, __, __, ___ 1 __L ___ ~__
AZOE
0. TZ+TS+T7 %B
,-_______--___________ChyTs-________________,~___~hy~~_,~_______ __,,__________-_______-_________T~,__-____________--______,)-___ Lys-Lys-Leu-Glu-Leu-Val-CyS-Asp-Cys-Ty~-Asn-Leu-Tyr-Arg-Lys- 7__1__7 __, __, --, _-, _-, -_, _-, -_, _-, -_, __) __, __ _I___
--______----____~-CbyT1-__-----___------__~~~~~~,~_---CbyT~----__ __T~-_,~_____-___T2__________,~_____-___________TS__-_________ Pro-Lys-Val-Ala-Thr-Thr-Cys-Arg-Glu-Asn_n-Ser-Val- ___ ___I__, --, --, --, _-, _',(__, __, --, --, __, --, --, --,
__,I_--________-__________________ ChyTS_________________________ ______,~________________________________________T~_____________ Phe-Arg-Gln-Cys-LeU-Asp-Asp-Leu-Leu-Leu-Leu-Ile-~n-Val-V~l-~p-Glu- __, ___/__, __T _-, __, _-, --, _-, --, -_, --, --, --, __, --,
__,,_____________-_____________I _________________-________---_____,
Tyr-Ile-Ser-Gly-Val-Gln-Ile-Val-NH, MIH P. boovlerf --7 --rl--71
) 4 0 30 60 90
MIN.
(_-)~-~~--(~~--)
y-- r-- z-- 7--
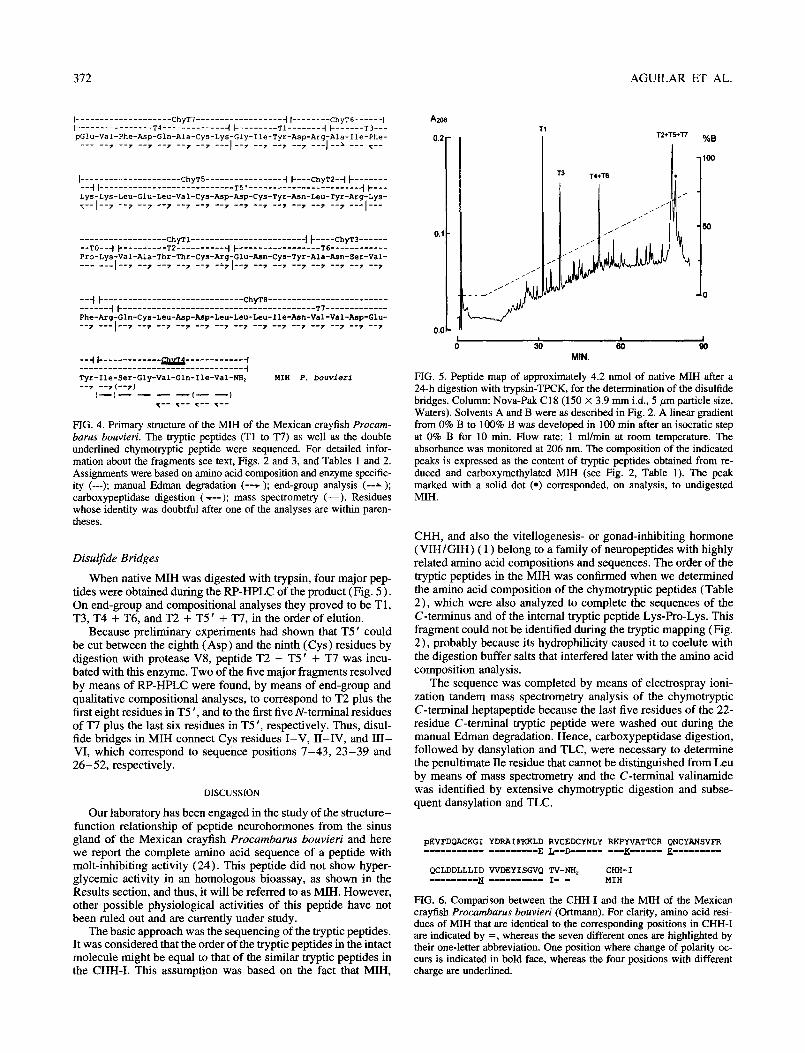
FIG. 4. Primary structure of the MIH of the Mexican crayfish Procam- bums bouvieri. The tryptic peptides (Tl to T7) as well as the double underlined chymotryptic peptide were sequenced. For detailed infor- mation about the fragments see text, Figs. 2 and 3, and Tables 1 and 2. Assignments were based on amino acid composition and enzyme specific- ity (---); manual Edman degradation (--7 ); end-group analysis (--A ); carboxypeptidase digestion ( Y--); mass spectrometry (-). Residues whose identity was doubtful after one of the analyses are within paren- theses.
FIG. 5. Peptide map of approximately 4.2 mnol of native MIH after a 24-h digestion with trypsin-TPCK, for the determination of the disulfide bridges. Column: Nova-Pak Cl8 (150 X 3.9 mm i.d., 5 pm particle size, Waters). Solvents A and B were as described in Fig. 2. A linear gradient from 0% B to 100% B was developed in 100 min after an isocratic step at 0% B for 10 min. Flow rate: 1 ml/mm at room temperature. The absorbance was monitored at 206 nm. The composition of the indicated peaks is expressed as the content of tryptic peptides obtained from re- duced and carboxymethylated MIH (see Fig. 2, Table 1). The peak marked with a solid dot (0) corresponded, on analysis, to undigested MIH.
Dim&de Bridges
When native MIH was digested with trypsin, four major pep-
tides were obtained during the RP-HPLC of the product (Fig. 5). On end-group and compositional analyses they proved to be Tl, T3, T4 + T6, and T2 + T5’ + T7, in the order of elution.
Because preliminary experiments had shown that T5 ’ could be cut between the eighth (Asp) and the ninth (Cys) residues by digestion with protease V8, peptide T2 + T5 ’ + T7 was incu- bated with this enzyme. Two of the five major fragments resolved by means of RP-HPLC were found, by means of end-group and qualitative compositional analyses, to correspond to T2 plus the first eight residues in T5 ‘, and to the first five N-terminal residues of T7 plus the last six residues in T5 ‘, respectively. Thus, disul- fide bridges in MIH connect Cys residues I-V, II-IV, and III- VI, which correspond to sequence positions 7-43, 23-39 and 26-52, respectively.
CHH, and also the vitellogenesis- or gonad-inhibiting hormone (VIH/GIH) ( 1) belong to a family of neuropeptides with highly related amino acid compositions and sequences. The order of the tryptic peptides in the MM was confirmed when we determined the amino acid composition of the chymotryptic peptides (Table 2), which were also analyzed to complete the sequences of the C-terminus and of the internal tryptic peptide Lys-Pro-Lys. This fragment could not be identified during the tryptic mapping (Fig. 2)) probably because its hydrophilicity caused it to coelute with the digestion buffer salts that interfered later with the amino acid composition analysis.
DISCUSSION
Our laboratory has been engaged in the study of the structure- function relationship of peptide neurohorrnones from the sinus gland of the Mexican crayfish Procambarus bouvieri and here we report the complete amino acid sequence of a peptide with molt-inhibiting activity (24). This peptide did not show hyper- glycemic activity in an homologous bioassay, as shown in the Results section, and thus, it will be referred to as MIH. However, other possible physiological activities of this peptide have not been ruled out and are currently under study.
The sequence was completed by means of electrospray ioni- zation tandem mass spectrometry analysis of the chymotryptic C-terminal heptapeptide because the last five residues of the 22- residue C-terminal tryptic peptide were washed out during the manual Edman degradation. Hence, carboxypeptidase digestion, followed by dansylation and TLC, were necessary to determine the penultimate Ile residue that cannot be distinguished from Leu by means of mass spectrometry and the C-terminal valinamide was identified by extensive chymotryptic digestion and subse- quent dansylation and TLC.
pEVFDQACKG1 YDRAIFKKLD RVCEDCYNLY RKPYVATTCR QNCYANSVFR
The basic approach was the sequencing of the tryptic peptides. It was considered that the order of the tryptic peptides in the intact molecule might be equal to that of the similar tryptic peptides in the CHH-I. This assumption was based on the fact that MIH,
QCLDDLLLID WDEYISGVQ TV-NH, CHH-I I_=E====_~ ===*lD===,= I= _ HIH
FIG. 6. Comparison between the CHH-I and the MM of the Mexican crayfish Procumbarus bouvieri (Grtmann). For clarity, amino acid resi- dues of MM that are identical to the corresponding positions in CHH-I are indicated by =, whereas the seven different ones are highlighted by their one-letter abbreviation. One position where change of polarity oc- curs is indicated in bold face, whereas the four positions with different charge are underlined.

STRUCTURE OF P. BOUVZERZ MOLT-INHIBITING HORMONE 373
pEVFDQAC-KGIYDRAIFKKLELVCDLB3'NLYRKPKVATTCRENCYANSVFR pEVFDQAC-KGVYDRNLFKKLDRVCEDCYNLYRKPFVATTCRENCYSNWVFR DTFDHSC-KGIYDRELFRKLDRVCEDCYNVFlUPKVATECKSNCFVNKRFN RVINDECPNLIGNFtDLYKKVl~WICEDCSNIFRKTGMASLCRRNCFFNEDF'V RVINDDCPNLIGNRDLYKKVl~WICDDCANIYRSTGMASLCRKDCFFNEDFL
= = = =====z ==I== _-- ---z === == == = =
QCLDDLLLINWDEYISGVQIV-NH> Prb-MIH QCLDDLLLSNVIDEYVSNVQ~ Hoa-MIH/CHH VCVADLRHDVSRFLKMANSALS Pev-MIH-like WCVHATERSEELRDLEEWVGILGAGRD Cam-MIH WCVRATERSEDLAQLKQWVTILGAGRI Cas-MIH
== =
FIG. 7. Amino acid sequence comparison among MIH of the crayfish Procambarus bouvieri (Prb-MIH) and MIH of the lobster Homarus americanus (Hoa-MIH), the shrimp Penaeus vannamei (Pev-MIH-like), the shore crab Carcinus maenas (Cam-MIH), and the blue crab Calli- nectes sapidus (Cas-MIH). Gaps in the hrst three sequences were intro- duced to maintain the conservation of the Cys residues among all the hormones. Prb-MIH has been taken as reference for the calculation of the degree of similarity, which is based on the longest hormone in any pair compared. Totally conserved residues are indicated in bold face whereas stretches that also include conservative substitutions are under- lined with heavy lines.
It is remarkable that four out of the seven differences found in comparing the sequences of the MIH and the two CHH from Procambarus bouvieri (Fig. 6) lead to a change of the residue charge, whereas two are conservative and one involves only a change of polarity. The former four residues could be important for the interaction with the MIH receptor and, at the same time, they could hinder an effect of MIH on the CHH receptor for we know that MIH is unable to elicit a rise of the circulating glu- cose levels in Procambarus bouvieri, as shown in the Results section, despite its 90% sequence similarity with the CHH. It should be noted that the origin of six out of the seven differ- ences is, certainly, due to point mutations. This implies that the Procambarus CHH and MIH are products of different genes, as has also been demonstrated for Carcinus and Callinectes at the genetic level (33,45). These genes could be expressed dif- ferentially in Procambarus, as has been observed in brachyuran crustaceans ( 14).
When we compare the sequences of the five MIH known so far (Fig. 7), it is easy to realize that MIH is a conserved molecule. Fourteen out of 78 residues or 18% (based on the longest MIH) are identical among all the MIH. If we take into account conser- vative substitutions, the level of sequence similarity reaches 38.5% (30 out of 78 residues) and the most conserved region comprises residues 13-53. Thus, the C-terminal residues, which
are clustered in a separate molecular region, could include the majority of the residues involved in species or group specificity. In addition, the degree of sequence similarity between the MIH from any pair of organisms matches their phylogenetic distance calculated on the basis of morphological criteria. A high degree of sequence conservation has also been observed among the CHH known so far and, moreover, four residues located in the C-ter- minus have been proposed as plausible candidates for interaction with the CHH receptor and are thought to be responsible for species specificity (25).
The sequence of the MIH-like neuropeptide from Penaeus vannamei (40) and the putative MIH from the blue crab Calli- nectes sapidus (33) were included in the comparison because they were considered good candidates for MIH, in spite of: 1) the first one having been cloned with the aid of a primer whose encoded sequence is also shared by CHH, and probably VIH, of several species, and 2) both not having been isolated nor tested by means of a bioassay. Support for this idea is that the precursors of the CHH genes cloned so far ( 11,12,45) include a CHH pre- cursor-related peptide, CPRP, which is absent in the precursors of MIH from Carcinus maenas (32)) Callinectes sapidus, the MIH-like neuropeptide from Penaeus, and GIH from Homarus americanus ( 13 ) . In addition, the signal peptides of the MIH- like precursor from Penaeus (at least 30 residues) and Callinec- tes (35 residues) resemble more closely the Carcinus MIH and Homarus GIH signal peptides (35 residues) than any of the cur- rently known CHH precursor signal peptides (Homarus CHH- A, 24 residues; Homarus CHH-B, 24 residues; Carcinus CHH, 26 residues; Orconectes CHH-A, 26 residues; Orconectes CHH- A * , 26 residues ) .
It has been found recently that D-Phe-containing isomorphs exist for CHH from Homarus americanus (39)) Procambarus clarkii (47), and Procambarus bouvieri (2). As shown in Hub- erman and Aguilar (22), the retention times in HPLC of the L- Phe- and the D-Phe-containing N-terminal tryptic peptides from CHH-I and CHH-II, respectively, are clearly distinguishable. Be- cause the elution times of the N-terminal tryptic peptides from MIH and CHH-I are identical, the Phe residue in position 3 in MIH should be the L-isomer. It will be very interesting to see whether the same posttranslational modification is present within a yet undetected MIH isomorph.
ACKNOWLEDGEMENTS
This work was made possible by grants to A. Huberman from the Consejo National de Ciencia y Tecnologfa (CONACYT) and from the United States-Mexico Foundation for Science. R. Falchetto was sup- ported by a fellowship from the European Molecular Biology Organi- zation (EMBO). The skillful assistance of Irma B. Mitre is gratefully acknowledged.
REFERENCES
Aguilar, M. B.; Quackenbush, L. S.; Hunt, D. F.; Shabanowitz, 3.; Huberman, A. Identification, purification and initial characterization of the vitellogenesis-inbibiting hormone from the Mexican crayfish Procambarus bouvieri (Ortmann). Comp. Biochem. Physiol. [B] 102:491-498; 1992. Aguilar, M. B.; Soyez, D.; Falchetto, R.; Arnott, D.; Shabanowitz, .I.; Hunt, D. F.; Hubemaan, A. Amino acid sequence of the minor isomotph of the crustacean hyperglycemic hormone (CHH-II) of the Mexican crayfish Procambarus bouvieri (Ottmann): Presence of a D-amino acid. Peptides 16:1375-1383; 1995. Allen, G. In: Work, T. S.; Burden, R. H., eds. Sequencing of proteins and peptides. Amsterdam: Elsevier/North Holland; 1981. Amott, D.; Shabanowitz, .I.; Hunt, D. F. Construction and performance of a laser desorption/ionization TOF mass spectrometer system: Apph-
5.
cations to problems in peptide, protein, and DNA structural analysis. proceedings of the 40th ASMS Conference on Mass Spectrometry and Allied Topics, Washington, DC, May 31-June 5, 1992. Amott, D.; Shabanowitz, _I.; Hunt, D. F. Mass spectrometry of pro- teins and peptides: Sensitive and accurate mass measurement and sequence analysis. Clin. Chem. 39:2005-2010; 1993. Bidlingmeyer, B. A.; Cohen, S. A.; Tarvin, T. L. Rapid analysis of amino acids using precolumn derivatization. J. Chromatogr. 336:93- 104; 1984. Chang, E. S. Endocrinology. In: Fast, A. W.; Lester, L. J., eds. Ma- rine shrimp culture: Principles and practices. Amsterdam: Elsevier Science Publishers B.V.; 1992:53-91. Chang, E. S.; Bruce, M. J.; Tamone, S. L. Regulation of crustacean molting: a multi-hormonal system. Am. Zool. 33:324-329; 1993.

374 AGUILAR ET AL.
9.
10.
11.
12.
13.
14.
15.
16.
17.
18.
19.
20.
21.
22.
23.
24.
25.
26.
27.
Chang, E. S.; O’Connor, J. D. Crustacea: Molting. In: Laufer, H.; Downer, R. G. H., eds. Endocrinology of selected invertebrates types. New York: Alan R. Liss; 1988:259-278. Chang, E. S.; Prestwich, G. D.; Bruce, M. J. Amino acid sequence of a peptide with both molt-inhibiting and hyperglycemic activities in the lobster, Homarus americanus. Biochem. Biophys. Res. Com- mun. 171:818-826; 1990. De Kleijn, D. P. V.; De Leeuw, E. P. H.; Van Den Berg, M. C.; Martens, G. J. M.; Van Herp, F. Cloning and expression of two mRNAs encoding structurally different crustacean hyperglycemic hormone precursors in the lobster Homarus americanus. Biochim. Biophys. Acta 1260:62-66; 1995. De Kleijn, D. P. V.; Janssen, K. P. C.; Martens, G. J. M.; Van Herp, F. Cloning and expression of two crustacean hyperglycemic hor- mones (CHH) mRNAs in the eyestalk of the crayfish Orconectes limosus. Eur. J. B&hem. 224:623-629; 1994. De Kleijn, D. P. V.; Sleutels, F. J. G. T.; Martens, G. J. M.; Van Herp, F. Cloning and expression of mRNA encoding prepro gonad inhibiting hormone (GM) in the lobster Homarus americanus. FEBS Len. 353:255-258; 1994. Dircksen, H.; Webster, S.; Keller, R. Immunocytochemical demon- stration of the neurosecretory systems containing putative moult in- hibiting hormone and hyperglycemic hormone in the eyestalk of brachyuran crustaceans. Cell Tissue Res. 251:3- 12; 1988. Femich, R. Neuroendocrine regulation of the crayfish molting gland. Diploma thesis, University of Bonn; 1987. Fingerman, M. The endocrine mechanisms of crustaceans. J. Crust. Biol. 7:1-24; 1987. Gray, W. R. A fluorescent end-group reagent for proteins and pep- tides. Biochem. J. 89:59P; 1963. Hartley, B. S. Strategy and tactics in protein chemistry. Biochem. J. 119:805-822; 1970. Hazra, A. K.; Chock, S. P. Y.; Albers, R. W. Protein determination with trinitrobenzene sulfonate: A method relatively independent of amino acid composition. Anal. Biochem. 137:437-443; 1984. Huberman, A. Hormonal control of molting in crustaceans. In: Ep- ple, A.; Scanes, C. G.; Stetson, M.-H., eds. Progress in comparative endocrinology, vol. 342 of Progress in clinical and biological re- search. Proceedings of the Eleventh International Symposium on Comparative Endocrinology, 14-20 May 1989. MQaga, Spain. New York: Wiley; 1990:205-210. Huberman, A.; Aguilar, M. B. A neurosecretory hyperglycemic hor- mone from the sinus gland of the Mexican crayfish Procambarus bouvieri (Ortmann). I. Purification and biochemical characterization of the most abundant form of the hormone. Comp. Biochem. Phys- iol. [B] 85:197-203; 1986. Huberman, A.; Aguilar, M. B. Single step purification of two hy- perglycaemic neurohormones from the sinus gland of Procambarus bouvieri. Comparative peptide mapping by means of high-perfor- mance liquid chromatography. J. Chromatogr. 443:337-349; 1988. Huberman, A.; Aguilar, M. B. A neurosecretory hyperglycemic hor- mone from the sinus gland of the Mexican crayfish Procambarus bouvieri (Ortmann). II. Structural comparison of two isoforms of the hormone. Comp. Biochem. Physiol. [B] 91:345-349; 1988. Huberman, A.; Aguilar, M. B. A neuropeptide with molt-inhibiting hormone activity from the sinus gland of the Mexican crayfish Pro- camburus bouvieri (Ortmann). Comp. Biochem. Physiol. [B] 93:299-305; 1989. Huberman, A.; Aguilar, M. B.; Brew, K.; Shabanowitz, J.; Hunt, D. F. Primary structure of the major isomorph of the crustacean hyperglycemic hormone (CHH-I) from the sinus gland of the Mex- ican crayfish Procumburus bouvieri (Ortmann) : Interspecies com- parison. Peptides 14:7-16; 1993. Hunt, D. F.; Henderson, R. A.; Shabanowitz, J.; et al. Characteri- zation of peptides bound to the class I MHC molecule IILA-22.1 by mass spectrometry. Science 255:1261-1263; 1992. Jegla, T. C.; Ruland, C.; Kegel, G.; Keller, R. The role of the Y- organ and cephalic gland in ecdysteroid production and control of
28.
29.
30.
31.
32.
33.
34.
35.
36.
37.
38.
39.
40.
41
42.
molting in the crayfish, Orconecres limosus. J. Comp. Physiol. 152:91-95; 1983. Karas, M.; Hillenkamp, F. Laser desorption ionization of proteins with molecular masses exceeding 10000 Da. Anal. Chem. 60:2299- 2301; 1988. Kegel, G.; Reichwein, B.; Tensen, C. P.; Keller, R. Amino acid sequence of crustacean hyperglycemic hormone (CHH) from the crayfish Orconectes limosus. Emergence of a novel neuropeptide family. Peptides 12:909-913; 1991. Kegel, G.; Reichwein, B.; Weese, S.; Gaus, G.; Peter-Katalinic, J.; Keller, R. Amino acid sequence of the crustacean hyperglycemic hormone (CHH) from the shore crab, Carcinus maenus. FEBS Lett. 255:10- 14; 1989. Keller, R. Crustacean neuropeptides: structures, functions and com- parative aspects. Experientia 48:439-448; 1992. Klein, J. M.; Mangerich, S.; De Kleijn, D. P. V.; Keller, R.; Wei- demann, W. M. Molecular cloning of a crustacean putative molt inhibiting hormone (MIH) precursor. FEBS Lett. 334:139-142; 1993. Lee, K. J.; Elton, T. S.; Bej, A. K.; Watts, S. A.; Watson, R. D. Molecular cloning of a cDNA encoding putative molt-inhibiting hor- mone from the blue crab, Callinectes supidus. Biochem. Biophys. Res. Commun. 209:1126-1131; 1995. Martin, G.; Sorokine, 0.; Van Dorsselaer, A. Isolation and molecular characterization of a hyperglycemic neuropeptide from the sinus gland of the terrestrial isopod Armudillidium vulgare (Crustacea) . Eur. J. Biochem. 211:601-607; 1993. Matsubara, H.; Sasaki, R. M. High recovery of tryptophan from acid hydrolysates of proteins. B&hem. Biophys. Res. Commun. 35:175-181; 1969. Meuth, J. L.; Fox, J. L. Separation of amino acid phenylthiohydan- toin derivatives by high-pressure liquid chromatography. Anal. Biochem. 154:478-484; 1986. Quackenbush, L. S. Crustacean endocrinology: A review. Can. J. Fish. Aquat. Sci. 43:2271-2282; 1986. Soyez, D.; Le Caer, J.-P.; Noel, P. Y.; Rossier, J. Primary structure of two isoforms of the vitellogenesis inhibiting hormone from the lobster Homarus americanus. Neuropeptides 20:25-32; 199 1. Soyez, D.; Van Herp, F.; Rossier, J.; Le Caer, J.-P.; Tensen, C. P.; Lafont, R. Evidence for a conformational polymorphism of inver- tebrate neurohormones. J. Biol. Chem. 269:18295-18298; 1994. Sun, P. S. Molecular cloning and sequence analysis of a cDNA en- coding a molt-inhibiting hormone-like neuropeptide from the white shrimp Penaeus vannumei. Mol. Mar. Biol. Biotechnol. 3:1-6; 1994. Tarr, G. E. Manual Edman sequencing system. In: Shively, J. E., ed. Methods of protein microcharacterization. Clifton, NJ: The Humana Press; 1986:155-194. Tatemoto, K.; Mutt, V. Chemical determination of polypeptide hor- mones. Proc. Natl. Acad. Sci. USA 75:4115-4119; 1978.
43. Tensen, C. P.; De Kleijn, D. P. V.; Van Herp, F. Cloning and se- quence analysis of cDNA encoding two crustacean hyperglycemic hormones from the lobster Homarus americanus. Eur. J. Biochem. 200:103-106; 1991.
44. Webster, S. G. Amino acid sequence of putative moult-inhibiting hormone from the crab Carcinus maenus. Proc. R. Sot. Lond. [Biol.] 2441247-252; 1991.
45. Weidemann, W.; Gromoll, J.; Keller, R. Cloning and sequence anal- ysis of cDNA for precursor of a crustacean hyperglycemic hormone. FEBS Lett. 257:31-34; 1989.
46. Woods, K. R.; Wang, K.-T. Separation of dansyl-amino acids by polyamide chromatography. B&him. Biophys. Acta 133:369-370; 1967.
47. Yasuda, A.; Yasuda, Y.; Fujita, T.; Naya, Y. Characterization of crustacean hyperglycemic hormone from the crayfish (Procamburus clarkii): Multiplicity of molecular forms by stereoinversion and di- verse function. Gen. Comp. Endocrinol. 95:387-398; 1994.