2004 rodriguez freshw crayfish
TRANSCRIPT

1
Comparison of the impact of the freshwater decapod species Austropotamobius pallipes(indigenous) and Procambarus clarkii (non-indigenous), on the submerged vegetation of
two Mediterranean wetlands.
RODRIGUEZ, C. F.1, BECARES, E.2 & FUERTES, B.3
1 Instituto de Medio Ambiente. Universidad de León. La Serna 56, 24071 León, SPAIN.2 Dpto. Ecología. Facultad de Biología, Universidad de León. Campus de Vegazana, 24071 León, SPAIN.3 Dpto. Biología Animal, Facultad de Biología, Universidad de León. Campus de Vegazana, 24071 León,
SPAIN.
ABSTRACT
Various studies indicate that the omnivorous activity of alien freshwater decapod species can drastically reducesubmerged aquatic vegetation cover. However, there is little information regarding the impact of native crayfishspecies on vegetation cover.
This paper contributes data for two shallow water systems that have been sampled over the same period of timeand using the same methodology, one (Chozas Lake, León) inhabited by the American red swamp crayfish(Procambarus clarkii) and the other (La Ercina Lake, Asturias) with one of the largest native crayfishpopulations in the Iberian Peninsula (Austropotamobius pallipes).
The data collected show that even at high densities the native A. pallipes is capable of coexisting with submergedmacrophyte cover (90% Characeae) whereas the invasion by P. clarkii of a wetland dominated by macrophyteshas led, in a brief period of time, to the total elimination of the vegetation cover, critically altering thefunctioning of the system and the composition of the trophic chain.
Keywords: herbivory, crayfish, native species, invasive, macrophytes, lake
INTRODUCTION
Freshwater crayfish are considered keystone members of trophic chains as a result of their frequently highbiomass, their longevity and their omnivorous feeding habits (Matthews et al, 1993). It has been demonstratedthat many crayfish species are capable of modifying their environment by acting on different levels of the trophicchain (Momot et al, 1978), regulating primary production through the consumption of macrophytes (Feminella &Resh, 1989; Olsen et al, 1991; Rodríguez et al, 2003) and preying on plant eaters (Weber & Lodge, 1990;Nystrom & Pérez, 1998) such as macroinvertebrate populations (Correia, 2002) and other groups from highertrophic levels that prey on them (Rodríguez et al, 2004).
Given the fundamental role played by submerged macrophytes in the ecology of wetlands and shallow lakes(Carpenter & Lodge, 1986; see Jeppesen et al, 1997 for a review) any agent capable of reducing them to belowcertain critical thresholds is worthy of special attention, as the changes produced by the loss of vegetation coverin these systems reach all levels of the trophic chain (Carpenter & Kitchell, 1993) and have a feedback effect,stabilising the deteriorated situation (Scheffer, 1990) and favouring the development of phytoplankton (Blindow,1993; see also for a revision Scheffer, 1998).Following the invasion of alien crayfish species in different parts of the world (Gherardi & Holdich, (1999)describe the present situation of exotic crayfish in Europe, while Lodge et al (2000) analyze the situation inNorth America) the study of the impact of their presence on invaded systems (Harper et al, 1990; Olsen et al,1991; Nyström, 1999; Correia, 2002; Rodríguez et al 2003 among others) and on indigenous species (Hill andLodge, 1999; Westman & Savolainen, 2001; Gherardi et al, 2002) has captured the interest of some limnologists

2
and astacologists, while others have focused on developing habitat inventories and requirements (Neveu, 2000;Rallo & García-Arberas, 2002) of native species, which have undergone great decline since the spread of exoticones. These alien species mainly come from North America and are vectors of aphanomycosis or crayfish plaguealthough they are resistant to it (Dieguez-Uribeondo & Soderhall, 1993).
The white clawed crayfish Austropotamobius pallipes (Lereboullet), currently listed as a vulnerable species(IUCN, 1996), is one of the 4-6 (according to different authors) indigenous European species and the only onepresent in Spain and Portugal, although various taxonomic studies (Grandjean et al, 2002) indicate that in actualfact the Iberian populations consist of introduced populations of Austropotamobius italicus. As a result of thereduction in the white-clawed crayfish’s range, which has seen all over Europe and has been very pronounced inSpain, most studies have focused on determining the causes of this decline and on the study of some populations(for example Matthews & Reynolds, 1995 in Ireland; Bernardo et al, 1997 in Portugal; Troschel, 1997 inGermany; Neveu, 2000 in France; García-Arberas & Rallo, 1999 and Royo et al, 2002 in Spain); however, theeffects of the species on the rest of the trophic network has never been fully established. In the Iberian Peninsula(Spain and Portugal) and generally throughout its European original distribution area, the white-clawed crayfishcurrently inhabits the upper parts of some catchments (Gil-Sanchez & Alba Tercedor, 2002) and small systemsisolated from the main catchment basin (Rallo et al, 2001) where its presence does not seem to trigger thechanges connected with the introduction of exotic decapod species, chiefly those changes relating to thedisappearance of submerged vegetation (Matthews & Reynolds, 1995). In Spain this species is currently includedin various recovery plans (Alonso et al, 2000).
The red swamp crayfish Procambarus clarkii (Girard) is a generalist crayfish that was introduced into Europefor the first time in Spain in 1974 (Gutiérrez-Yurrita & Montes, 1999). Since then it has spread throughoutalmost all of Europe (Barbaresi & Gherardi, 2000). Its impact on wetlands has been described in terms ofchanges to the breeding success of other species as a result of alterations to the habitat and the destruction of theirfood supplies (Gutiérrez-Yurrita & Montes, 1999; Rodríguez et al, 2004), and in terms of direct effects on theaquatic flora and fauna (Nyström & Pérez, 1998; Angeler et al., 2001; Correia, 2002; Cronin, 2002; Rodríguez etal., 2003).
During the summer sampling carried out in the year 2000 as part of the European ECOFRAME project,information was collected from various shallow lakes and wetland systems, including two inhabited by twodifferent crayfish species: La Ercina Lake (Picos de Europa National Park) with one of the largest A. pallipespopulations in the north of the Iberian Peninsula, and Chozas Lake which suffers the effects of the introductionof P. clarkii.
This paper compares, with regard to these two systems, the data collected for the different physical and chemicalwater parameters, as well as for vegetation cover and the composition of macrophyte species in the presence ofan indigenous crayfish and an alien crayfish and the implications of this on the current state of these systems.
METHODS
During the summer of 2000 samples were taken at Chozas de Arriba Lake and La Ercina Lake to collect data forthe physical and chemical parameters recommended in the APHA-AWWA-WPCF Standard Methods for thedetermination of water quality. To determine the trophic status of the lake the Chlorophyll a (Chl.a) and TotalPhosphorus (TP) limits indicated by the OECD (1982) were taken into account. In turn, transects were madefrom the boat, along which the cover, PVI (percentage of volume occupied by vegetation) and the composition ofmacrophyte species present every ten metres were recorded. The number of transects was set in relation to theperimeter of the lake.
With the aim of comparing the densities of each of the crayfish species, a number of unbaited traps were placedin each system, the number being proportional to the size of each lake, specifically one trap for every 150 metresof shoreline, comprising an interception net that guided the animals in transit towards two fykes where they were

3
retained. The traps were placed perpendicular to the shore and remained in the lakes for one night (12 hours)before being collected the following morning. The size of the mesh of the traps established a selectivity in thecaptures of individuals with a total length of over 60 mm.
For all the specimens captured the total number of captures per trap was recorded as was the mean weight for theP. clarkii species. For the A. pallipes population, as well as the above-mentioned information, additional data onsex-ratio, weight and total length of each individual was also collected.
During September 2000 the true density of the P. clarkii at Chozas Lake was estimated using a capture-marking-recapture technique (Krebs, 1990). For the capture, a set of 20 traps baited with fresh spleen was used. The trapswere extracted every 15 minutes and the bait replaced if necessary, in daily series of three hours until dusk. Atthe end of the day the unmarked individuals captured were marked by means of a cut on the exopodites uropodsor noted down as marked, before being returned to the water.
RESULTS
Chozas Lake
Chozas Lake is a small body of shallow water (9 ha with a maximum depth of 1.8 m) located in León (north westSpain) and an argillaceous bottom, surrounded by a small area of semi-permanent wetland that is home to animportant nesting colony of lapwings (Vanellus vanellus) and other waterfowl. The lake has traditionally beenused as an irrigation reservoir which is why in the 1950s retaining walls were constructed on the south and westsides to increase the depth. Notably, the water level fluctuates significantly, both on a seasonal basis due to themarked drought in summer (a drop in the water level of almost 1 metre compared to its maximum capacity and areduction in the flooded area to about 3 ha in the summer) and from year to year (there was a severe episode ofdrought in 1993). The local residents have used the lake for various marginal, unregulated hunting activities(hunting of birds and amphibians, fishing).
Fernández-Aláez (1984) undertook the first limnological study of the system as an example of a wetland areawith abundant and diverse aquatic vegetation, a situation that remained unchanged until 1997, when the presenceof P. clarkii was detected. Prior to that year there is no record of crayfish presence. From 1994 to the present thelake has been monitored regularly and various studies (Rodriguez et al, 2003 and 2004) reflect the changes thathave taken place since the introduction of Procambarus clarkii in this system.
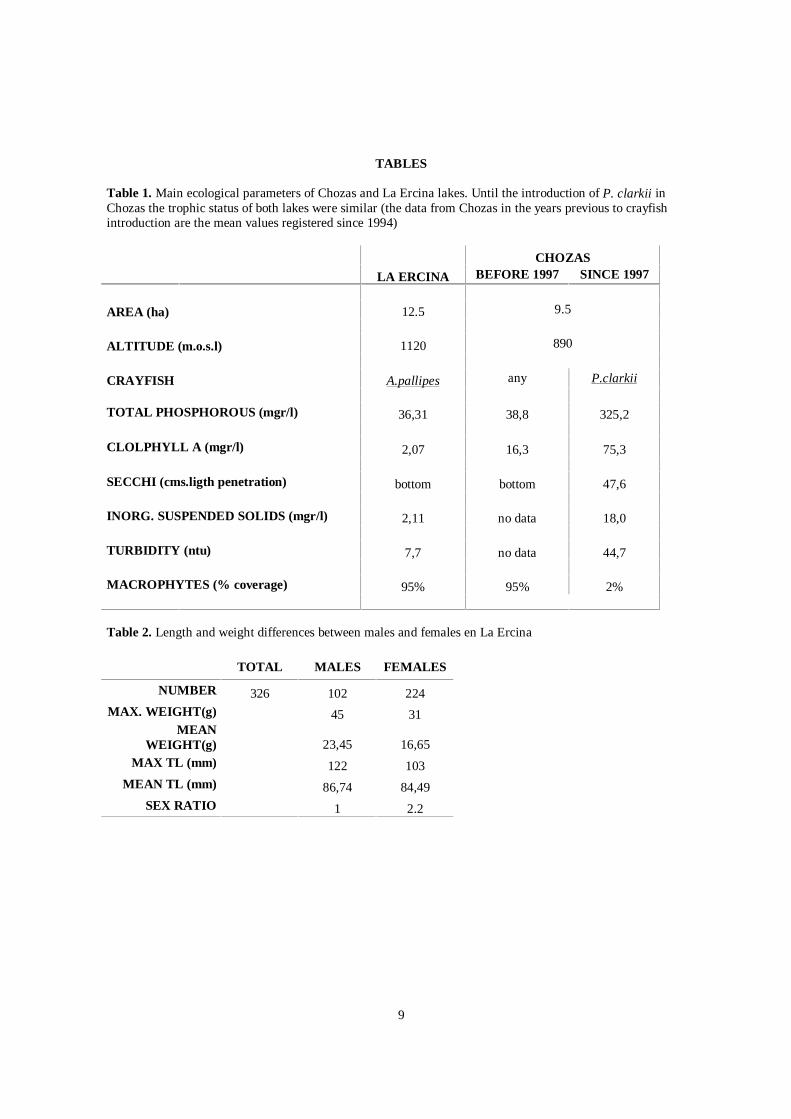
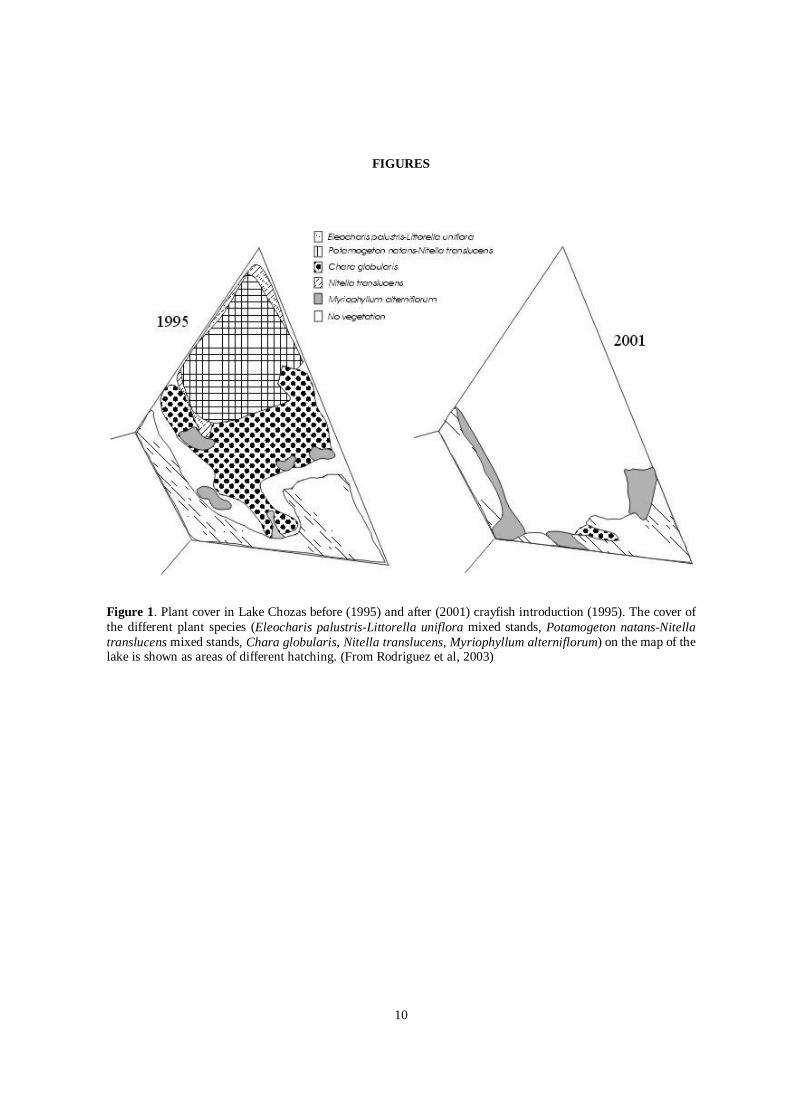
Table 1 sums up the data collected at Chozas for the period prior to the detection of the red swamp crayfish plusthe data collected during the present study with regard to the most relevant water quality parameters, Chl.a andturbidity. From 1997 the concentration of TP and Chl.a in the water column has increased and the trophic state ofthe lake has been classified as hypertrophic. Vegetation cover (see Figure 1) was less than 2% in the summer of2002, with only two of the seven macrophyte species described in 1984 present: Litorella uniflora andMyriophyllum alterniflorum occupying a small area close to the shores.
The CPUE recorded for P. clarkii at Chozas was 7.3 ind./trap, with a total of 22 individuals captured and a meanweight of 18 g. Furthermore, a skewing in the sex-ratio of 1.49:1 in favour of the males was determined. In thesummer of 2002 a capture-mark-recapture study was carried out, which made it possible to give an estimate ofthe true density of around 1 ind./m2 (Rodriguez et al, 2003)
La Ercina Lake
La Ercina Lake is located on the Asturian side of the Picos de Europa National Park at an altitude of 1108 metresabove sea level. Its maximum floodable area is around 12.5 ha, with a mean depth of 1.05 m and a maximumdepth of 1.90 m with a soft clay bottom throughout.

4
The data for the main water quality parameters, Chl.a and turbidity, are shown in table 1. Because of its TP(36.31µg/l) and Chl.a (2.07 µgr/l) levels it has been classified as oligotrophic. The vegetation cover recorded inthe summer of 2000 was close to 85%, composed exclusively of a single species, Chara sp, which on averageoccupied 41% of the volume of the lake (PVI).
According to Pardo (1942) the population of Austropotamobius pallipes (Austropotamobius italicus according toGrandjean et al (2002)) of La Ercina Lake was probably introduced in the 18th century and today forms part ofthe Natural Park’s natural ecosystem; nevertheless Fernandez-Bernaldo & García-Fernandez (1987) point outthat this introduction was made later, at the beginning of the Twentieth century. On the other hand Muñoz-Goyanes (1967) remarks the high densities of A. pallipes in La Ercina, in contrast with other lakes in the Picos deEuropa National Park. The main biometric records for this population are summed up in table 2. The traps laid inthis system showed an apparently high density of A. pallipes, with a total of 326 crayfish captured and a CPUE of81.5 ind./trap. The largest individual recorded was a male with a TL of 122 mm. The heaviest individual weighed45 g and was also a male. It is worth mentioning that 38.2% of the males and 22.2% of the females had a totallength of over 90 mm and the mean total length and weight was greater for the males than for the females. Thesex-ratio observed in La Ercina during the study was 1:2.20 in favour of the females.
DISCUSSION
Although the two surveyed systems have crayfish populations, it is clear that the effect of their presence is verydifferent depending on whether it is the native white-clawed crayfish Austropotamobius pallipes that is present orthe alien species Procambarus clarkii. Furthermore, although the current trophic status of the systems showsgreat differences – La Ercina oligotrophic and Chozas hypertrophic – the condition of the latter system prior tothe invasion of the red swamp crayfish (Fernández-Alaez, 1984) makes it possible to attribute the presentdeterioration to the depredatory action of P. clarkii on the vegetation, shedding light on the different roles thatthe two species play in the systems.
The red swamp crayfish appears to be the main culprit for the destruction of the vegetation cover at Chozas(Rodríguez et al, 2003). This has given rise to a notable increase in the levels of nutrients and Chl. a in the water,which in turn, as a result of a trophic cascade effect (Carpenter & Kitchell, 1993; Blindow et al, 2000) hasbrought about an enormous loss in animal taxa which to some degree or other depended on the submergedvegetation for food, shelter or laying sites (Rodríguez et al, 2005).
However, and despite the fact that the results seem to indicate that the density of the native white-clawed crayfishappears to be high, and in any case much higher than the true density of the American red swamp crayfish atChozas, La Ercina Lake has dense beds of submerged macrophytes that take up 95% of the surface (Figure 2).Significantly, this community of macrophytes is mainly composed of a single species, Chara sp, which accordingto the scientific literature not only constitutes a preferred food item for the white-clawed crayfish (O´Keeffe,1986 in Matthews and Reynolds) but also for other freshwater decapod species and specifically for P. clarkii(Cronin, 1998; Angeler et al, 2001; Cirujano et al, 2004). Indeed it has been repeatedly confirmed (Vila-Escale etal, 2002; Rodríguez et al, 2003) that it is precisely the charophyte group that is the first to disappear after asystem is invaded by the red swamp crayfish, while studies carried out on A. pallipes in lentic environments(reservoirs and shallow lakes) show that the larger concentrations of this species in different systems (Matthewset al, 1993) are associated with the presence of charophytes, while they are all but absent from areas withoutvegetation.
The consumption of Chara sp by A. pallipes was studied by Matthews et al. (1993) using exclusion experiments,proving that although for all the density levels of the species that were used in the experiment a reduction in thelength and biomass of the charophytes was recorded, for none of the levels (max. density 148.5 g/m2) did thisreduction exceed 46.5% of the biomass. In contrast, the observations made at Chozas reflect a completedestruction of the charophyte cover (which was over 95% in 1994, Fernández-Alaez et al, 1999) with a density ofP. clarkii close to 22 g/m2. Inclusion experiments of this species and Myriophyllum alterniflorum, similar to the

5
previous ones, showed a destruction of 60% of the biomass in just 15 days with a red swamp crayfish density of44 g/m2 (Rodríguez et al, 2003)
The maintenance of Chara sp cover in lakes and wetlands inhabited by A. pallipes not only results in themaintenance of an appropriate habitat for their development, but also means that this charophyte vegetation canplay a fundamental role in maintaining other characteristics of these systems such as water transparency,moderate levels of nutrients in the water (Moss 1990) and the communities of organisms that are characteristic oftrophic chains in wetlands and lakes in clear water states (Scheffer, 1998; Blindow, 2000).
Because of its nutrient content (ver Demers & Reynolds, 2004), depth (Rallo & García-Arberas, 2000) and theavailability of food (Matthews et al, 1993) the whole La Ercina Lake can be considered an adequate habitat to therequirements of A. pallipes. In addition it has a ggod chance of persisting in the long term given its isolation fromthe rest of the waters in its basin, a factor which seems particularly important for maintaining the populations ofthe Iberian Peninsula (Gil-Sanchez & Alba Tercedor, 2002) safe from aphanomycosis.
Furthermore, both the CPUE obtained in this study and the total number of individuals captured in a single day ofsampling seem to indicate that this could be one of the most important lentic populations of native crayfish in theIberian Peninsula. Ideally it would be necessary to carry out the study over the entire phenological cycle of thespecies, as the high density could be favoured by movements connected with the phase prior to reproduction(sampling was carried out at the end of September).
The population structure of A. pallipes at La Ercina is similar to that found by Grandjean et al (2000) in Frenchstreams, with a sex-ratio skewed in favour of the females, and with almost 25% of the total population within thelegal minimum size (90 mm TL). Other authors, however (Matthews & Reynolds, 1995 in Ireland; Bernardo etal, 1997 in Portugal; Neveu, 2000 in France) have found that the percentage of individuals that exceed this legalsize is, in all cases, less than 9%. With regard to the sex-ratio, the above-mentioned study carried out in Irelandfound a greater number of males than females, while the Portuguese study found a balanced sex-ratio.Furthermore, Rallo & García-Arberas (2000) in a study carried out on a Spanish lake comparable to La Ercinafound a proportion of males that was double that of the females and they point out that these results are theopposite of those obtained from other studies that they carried out (García-Arberas & Rallo, 1999) on streams inthe same geographical environment.
It is also notable that the maximum recorded TL for males (122 mm) is greater that that recorded for otherpopulations (Almaça, 1989; Bernardo et al, 1997; Troschel, 1997; García-Arberas & Rallo, 1999). In accordancewith other studies on the subject, the males dominate the larger size classes.
CONCLUSIONS
While it is clear that it is necessary to prevent the spread of exotic crayfish species and in particular to protectcertain systems because of their vulnerability to biological invasion (such as wetlands and shallow lakes) it isalso apparent that these very systems, because of their trophic characteristics and isolated positions, are optimalplaces for the development of native European crayfish species, which must be protected effectively from contactwith exotic crayfish populations that are vectors of aphanomycosis.
Austropotamobius pallipes’s adaptation to the environment that it inhabits means that it can be used not only as aflagship species for the conservation of certain habitats, but that it also plays an integrated role in maintaining thewell-being of these very systems..
The introduction of non-indigenous species of crayfish as an ecological substitute to A. pallipes in waters wherethis species has disappeared must be preceded by appropriate studies of the impact of the new species on theflora and fauna of the native systems.

6
ACKNOWLEDGEMENTS
The author wish to thank Anna Homan for her contribution to the English version of this manuscript. AlvaroGonzález and Juan Coco provided scientific support. This study was funded by the European Union-fundedproject ECOFRAME.
REFERENCES
Alonso, F., Temino, C. & Dieguez-Uribeondo, J. 2000. Status of the white-clawed crayfish, Austropotamobiuspallipes (Lereboullet, 1858), in Spain: distribution and legislation. Bulletin Française de la Pêche et laPisciculture. 356: 31-53.APHA-AWWA-WPCF, 1989. Estándar Methods for the Examination of Water and Wastewater. Chapter 10. 210pp.Barbaresi, S. & Gherardi, F. 2000. The invasion of the alien crayfish Procambarus clarkii in Europe, withparticular reference to Italy. Biological invasions 2: 259-264.Bernardo, J.M., Ilheu, M. & Costa A.M. 1997. Distribution, population structure and conservation ofAustropotamobius pallipes in Portugal. Bulletin Française de la Pêche et la Pisciculture. 347: 617-624.Blindow, I., Andersson, G., Hargeby, A. and Johansson, S. 1993. Long term pattern of alternative stable states intwo shallow eutrophic lakes. Freshwater Biology 30: 159-167.Blindow, I.., Hargeby, A. and Andersson, G., 2000. Long-term waterfowl fluctuations in relation to alternativestates in two shallow lakes. F.A. Comín, J.A. Herrera & J. Ramírez (Eds.). Limnology and Aquatic Birds.Monitoring, Modelling and Management. Universidad Autónoma de Yucatán, Mérida (México).Carpenter, S.R. & Lodge, D.M. 1986. Effects of submerged macrophytes on ecosystem processes. AquaticBotany 26: 341-379).Carpenter, S.R. & Kitchell, J.F. 1993. The trophic cascade in lakes. Cambridge University Press, 386 pp.Cirujano, S., Camargo, J.A, and Gómez-Cordobés, C. 2004. Feeding preferente of the red swamp crayfishProcambarus clarkii (Girard) on living macrophytes in a Spanish wetland. Journal of Freshwater Ecology 19(2):219-226.Correia, A.M. 2002. Niche breadth and trophic diversity: feeding behaviour of the red swamp crayfish(Procambarus clarkii) towards environmental availability of aquatic macroinvertebrates in a rice field (Portugal).Acta Oecologica 23: 421-429.Cronin, G. 1998. Influence of macrophyte structure, nutritive value and chemistry on the feeding choices of ageneralist crayfish. In Jeppesen E., M. Sondergaard, M. Sondergaard, K. Christoffersen (eds.) The structuringrole of submerged macrophytes in lakes. pp 149-174. Springer, New York.Cronin, G., Lodge, D.M., Hay, M.E., Miller, M., Hill, A.M., Horvath, T., Bolser, R.C., Lindquist, N. & Wahl, M.2002. Crayfish feeding preferences for fresh water macrophytes: The influence of plant structure and chemistry.Journal of Crustacean Biology 22 (4): 708-718.Demers, A. & Reynolds, J.D. 2004. Water quality requirements of the white-clawed crayfish, Austropotamobiuspallipes, a field study. IAA 15 Abstracts.Dieguez-Uribeondo, J. & Soderhall, K. 1993. Procambarus clarkii Girard as a vector for the crayfish plaguefungus, Aphanomyces astaci Schikora. Aquaculture and Fisheries management 24 (6): 761-765.Feminella, J.W. & Resh, V.H. 1989. Submerged macrophytes and grazing crayfish: an experimental study ofherbivory in a California freshwater marsh. Holartic Ecology 12 (1): 1-8.Fernández-Aláez, M. 1984. Distribution of macrophytes and their ecological factors in lentic systems from LeónProvince (Spain). Thesis Dissertation. (in Spanish).Department of Ecology. University of León.Fernández-Aláez, M., Fernández-Aláez ,C., Rodríguez, S. and Bécares, E. 1999. Evaluation of the state ofconservation of shallow lakes in the province of León (Northwest Spain) using botanical criteria. Limnetica 17:107-117.Fernández, C. & García, E. (1987) Lagos y Lagunas de Asturias. Ayalga Ediciones, Salinas. 261 p.García-Arberas, L. & Rallo, A. 1999. Survival of natural populations of Austropotamobius pallipes in rivers inBizkaia, Basque Country (North of Iberian Peninsula). Bulletin Française de la Pêche et la Pisciculture. 356: 17-30.

7
Gherardi, F. & Holdich, D.M. 1999. Crayfish in Europe as alien species. Crustacean Issues 11. Balkema.Rotterdam.Gherardi, F., Smietana, P. and Laurent, P. 2002. Interactions between non-indigenous and indigenous crayfishspecies. Bulletin Française de la Pêche et la Pisciculture. 367: 899-907.Gil- Sanchez, J.M. & Alba-Tercedor, J. 2002. Ecology of the native and introduced crayfishes Austropotamobiuspallipes and Procambarus clarkii in southern Spain and implications for conservation of the native species.Biological Conservation 105: 75-80Grandjean, F., Harris, J., Souty-Grosset, C. and Crandall, K.A. 2000. Systematics of the european endangeredcrayfish species Austropotamobius pallipes (Decapoda: Astacidae). Journal of Crustacean Biology 20: 522-529.Gutiérrez-Yurrita, P.J. & Montes, C. 1999. Bioenergetics and phenology of reproduction of the introduced redswamp crayfish, Procambarus clarkii, in Doñana National Park, Spain, and implications for speciesmanagement. Freshwater biology, 42,561-574Harper, D.M., Mauvati, K.M. and Muchiri, S.M. 1990. Ecology and management of Lake Naivasha, Kenya, inrelation to climatic change, alien species introductions and agricultural development. EnvironmentalConservation 17: 328-336.Hill, A.M. & Lodge, DM 1999. Replacement of resident crayfishes by an exotic crayfish: The roles ofcompetition and predation. Ecological Applications 9 (2): 678-690.IUCN, 1996. Red List of Threaded Animals. IUCN, Gland, Switzerland.Jeppesen, E., Sondergaard, M. and Christofferson, K. 1997. The structuring role of submerged macrophytes,Springer-Verlag, New York. 423 pp.Krebs, C.J. 1991 Ecological methodology. Addison Wesley, MadridLodge, D.M., Taylor, C.A., Holdich, D.M. and Skurdal, J. 2000. Nonindigenous crayfishes threaten NorthAmerican freshwater biodiversity: Lessons from Europe. Fisheries 25(8): 7-20.Matthews, M.A., Reynolds, J.D. and Keatingue, M.J. 1993 Macrophyte reduction and benthic communityalteration by the crayfish Austropotamobius pallipes (Lereboullet). Freshwater crayfish 9:289-299Matthews M.A. & J.D. Reynolds 1995. A Population Study of the white clawed crayfish Austropotamobiuspallipes (Lereboullet) in an Irish Reservoir. Proceedings of the Royal Irish Academy 95B (2): 99-109.Momot, W.T., Gowing, H. and Jones, P.D. 1978. The dynamics of crayfish and their role in ecosystems.American Midlands Naturalist 99:10-35.Moss, B. 1990. Engineering and biological approaches to the restoration from eutrophication of shallow lakes inwhich aquatic plant communities are important components, Hydrobiologia 200/201:367-77Muñoz, G. 1967. Parque Nacional de la Montaña de Covadonga. Servicio Nacional de Pesca Fluvial y Caza.Madrid.Neveu, A. 2000. Study of Austropotamobius pallipes (Crustacea, Astacidae) populations in a forest brook inNormandy. I. Demographic structures and growth: stability and variability during six years. Bulletin Française dela Pêche et la Pisciculture. 356: 71-97 2000.Neveu, A. 2000 Study of Austropotamobius pallipes (Crustacea, Astacidae) populations in a forest brook inNormandy. II. Distribution in relation to habitats structures: stability and variability during five years. BulletinFrançaise de la Pêche et la Pisciculture. 356: 99-122.Nyström, P. & Pérez, J.R. 1998. Crayfish predation on the common pond snail (Lygnea stagnalis): the effect ofhabitat complexity and snail size on foraging efficiency. Oikos 88(1):181-190.Nyström, P. !999. Ecological impact of introduced and native crayfish on freshwater communities: Europeanperspectives. In Gherardi, F. and D.M. Holdich Eds. Crayfish in Europe as alien species. A.A. Balkema.Rotterdam. 63-85.OECD (Organization for Economic Cooperation and Development) 1982. Eutrophication of waters. Monitoring,assessment and control. Final report OECD Cooperative programme on Monitoring of Inland Waters(Eutrophication Control); Environment Directorate, OECD, Paris. 154 pp.O¨keffe, C. 1986. The ecology of two populations of the freshwater crayfish Austropotamobius pallipes(Lereboullet) in Ireland. Ph. D Thesis, Department of Zoology, University of Dublin. 254 pp.Olsen, T.M., Lodge, D.M., Capelli, G.M. and Houlihan, R.J. 1991. Mechanisms of impact of an introducedcrayfish (Orconectes rusticus) on litoral congeners, snails and macrophytes. Canadian Journal of Fisheries andAquatic Science 48: 1853-1861.Rallo, A. & García-Arberas, L. 2000. Population Structure and dynamics and Habitat Conditions of the nativecrayfish Austropotamobius pallipes in a pond: a Case Study in Basque Country (Northern Iberian Peninsula).Bulletin Française de la Pêche et la Pisciculture 356: 005-016.

8
Rallo, A. & García-Arberas, L. 2002. Diferences in abiotic water conditions between fluvial reaches and crayfishfauna in some northern rivers of the Iberian Peninsula. Aquatic Living Resources 15: 199-128.Rodríguez, C.F., Bécares, E. and Fernández-Aláez, M. 2003. Shift from clear to turbid phase in Lake Chozas(NW Spain) due to the introduction of American red swamp crayfish (Procambarus clarkii). Hydrobiologia 506-509: 421-426.Rodriguez, C.F., Bécares, E., Fernández-Alaez, M. and Fernández-Alaez, C. 2005. Loss of diversity anddegradation of wetlands as a result of introducing exotic crayfish. Biological Invasions 7: 75-85.Royo, F., Gonzalez-Cienfuegos, G. & Muzquiz, J.L. 2002. Preliminary observation of occasional mortality in thepopulation of the freshwater crayfish in the Picos de Europa (Asturias, Spain). 2002. Bulletin Française de laPêche et la Pisciculture. 367: 935-941.Scheffer, M. 1990. Multiplicity of stable states in fresh water ecosystems. Hydrobiologia 200-201:475-486.Scheffer, M. 1998. Ecology of shallow lakes. Chapman & Hall. London. 357 p.Troschel, H.J. 1997. Distribution and Ecology of Austropotamobius pallipes in Germany. Bulletin Française dela Pêche et la Pisciculture. 347: 639-647.Vila-Escalé, M., Rieradevall, M. and Prat, N. 2002. Estudio de la población de cangrejo rojo (Procambarusclarkii) sobre las comunidades vegetales sumergidas en dos torrentes del macizo de Sant Llorenç del Munt iL´Obac. Resúmenes del XI Congreso de la Asociación Española de Limnología y III Congreso Ibérico deLimnología. Cedex Publ.. MadridWeber, L.M. & Lodge, D.M. 1990. Periphytic food and predatory crayfish: relative roles in determining snaildistribution. Oecologica 82: 33-39Westman, K. & Savolainen, R. 2001. Long term study of competition between two co-occurring crayfish species,the native Astacus astacus L. and the introduced Pacifastacus leniusculus Dana, in a Finnish lake. BulletinFrançaise de la Pêche et la Pisciculture. 361: 613-627.
9
TABLES
Table 1. Main ecological parameters of Chozas and La Ercina lakes. Until the introduction of P. clarkii inChozas the trophic status of both lakes were similar (the data from Chozas in the years previous to crayfishintroduction are the mean values registered since 1994)
CHOZASLA ERCINA BEFORE 1997 SINCE 1997
AREA (ha) 12.5 9.5
ALTITUDE (m.o.s.l) 1120 890
CRAYFISH A.pallipes any P.clarkii
TOTAL PHOSPHOROUS (mgr/l) 36,31 38,8 325,2
CLOLPHYLL A (mgr/l) 2,07 16,3 75,3
SECCHI (cms.ligth penetration) bottom bottom 47,6
INORG. SUSPENDED SOLIDS (mgr/l) 2,11 no data 18,0
TURBIDITY (ntu) 7,7 no data 44,7
MACROPHYTES (% coverage) 95% 95% 2%
Table 2. Length and weight differences between males and females en La Ercina
TOTAL MALES FEMALES
NUMBER 326 102 224MAX. WEIGHT(g) 45 31
MEANWEIGHT(g) 23,45 16,65
MAX TL (mm) 122 103MEAN TL (mm) 86,74 84,49
SEX RATIO 1 2.2
10
FIGURES
Figure 1. Plant cover in Lake Chozas before (1995) and after (2001) crayfish introduction (1995). The cover ofthe different plant species (Eleocharis palustris-Littorella uniflora mixed stands, Potamogeton natans-Nitellatranslucens mixed stands, Chara globularis, Nitella translucens, Myriophyllum alterniflorum) on the map of thelake is shown as areas of different hatching. (From Rodriguez et al, 2003)

11
Figure 2. La Ercina Lake submerged macrophytes distribution. Since 1987 the coverage and species compositionhas not changed substantially. (Modified from Fernández & García, 1987)