conservation prioritisation of south australian marine biodiversity: effectiveness and efficiency of...
TRANSCRIPT

School of Environmental Sciences - ENV University of East Anglia - UEA
Conservation prioritisation of South Australian marine biodiversity:
Effectiveness and efficiency of the protected areas network
A dissertation presented in part-fulfilment of the degree of Master of Science in Applied Ecology and Conservation, in accordance with the regulations of the University of East Anglia. Supervision of Dr Aldina Franco (UEA) and co-supervision of Dr Bronwyn Gillanders and Dr Bertram Ostendorf (University of Adelaide – UofA).
Rebecca Borges e Silva August - 2013
School of Biological Sciences University of East Anglia Norwich Research Park Norwich NR4 7TJ
@2013 Rebecca Borges e Silva This copy of the dissertation has been supplied on condition that anyone who consults it is understood to recognize
that its copyright rests with the author and that no quotation from the dissertation, nor any information derived
there from, may be published without the author’s prior written consent. Moreover, it is supplied on the
understanding that it represents an internal University document and that neither the University nor the author are
responsible for the factual or interpretive correctness of the dissertation.

Borges, R. 2013
2
ABSTRACT
In the marine environment, protected areas represent a strategy to reduce biodiversity loss.
However, non-systematic design often leads to reserves that do not contribute to biodiversity
representativeness. This study aimed to evaluate the effectiveness and efficiency of the new
South Australian marine parks system through 1) a biodiversity coverage analysis that
compares zones with different levels of protection; and 2) an administrative units analysis that
assesses efficiency in the representation of species and habitats obtained from designs at the
state and at the regional scales. An expansion of the no-take zones to 10% of the study area was
also investigated. The outer boundaries of South Australian marine protected areas provide
good coverage of biodiversity (mean = 66.59%, se = 3.543), but only a small amount of the
distribution of species and habitats are preserved in no-take zones (mean = 13.82%, se = 2.053),
while some features are completely excluded from these higher protection level zones. The
administrative units analysis showed no considerable decrease in efficiency when priorities are
set at the regional scale, although, at this design scale, solutions present concentrated proposed
reserves near the borders of the bioregions – edge artefacts. However, for the expansion of the
no-take zones to 10% of the State waters, efficiency is considerably reduced in the regional
approach, but without enhanced edge artefacts. For the 10%-expansion scenario, comparing
and combining different solutions might be preferred to spread out new no-take zones and
consequently reduce the social conflicts that these new areas could cause.
Key-words: marine protected areas – spatial planning – gap analysis – administrative units –
edge artefacts

Borges, R. 2013
3
TABLE OF CONTENTS
ACKNOWLEDGEMENTS_________________________________________________________________________________4
1. INTRODUCTION_______________________________________________________________________________________6
1.1 Global marine biodiversity conservation 6
1.2 Marine conservation in South Australia 7
1.3 Project aims and relevance for conservation 8
2. METHODS______________________________________________________________________________________________9
2.1 Study area 9
2.2 Software 9
2.3 Source and processing of datasets 12
2.4 Effectiveness: gap analysis 13
2.5 Statistical analysis 14
2.6 Efficiency: administrative units analysis 14
2.7 Expansion of the no-take zones 15
3. RESULTS______________________________________________________________________________________________16
3.1 Effectiveness: gap analysis 16
3.2 Efficiency: administrative units analysis 21
3.3 Expansion of the no-take zones 25
4. DISCUSSION__________________________________________________________________________________________29
4.1 Gap analysis and expansion of no-take zones 29
4.2 Administrative units analysis 31
5. CONCLUSIONS_______________________________________________________________________________________34
6. CONSERVATION IMPLICATIONS _________________________________________________________________35
7. REFERENCES________________________________________________________________________________________37
I. APPENDIX A__________________________________________________________________________________________41
II. APPENDIX B_________________________________________________________________________________________42
III. APPENDIX C________________________________________________________________________________________45
IV. APPENDIX D________________________________________________________________________________________46

Borges, R. 2013
4
ACKNOWLEDGEMENTS
I thank my supervisors Dr Aldina Franco, Dr Bronwyn Gillanders and Dr Bertram
Ostendorf for the academic and technical support to deal with conservation concerns and with a
range of pieces of software that are supposed to help answer these issues, but that also raise
others ecological questions in the process.
Many thanks also to the European Commission and its Education, Audiovisual and
Culture Executive Agency (EACEA) for the Erasmus Mundus scholarship, and to the European
Master in Applied Ecology (EMAE) coordinators and secretaries Yves Caubet, Mark Hassall,
Wilhelm Windhorst, Martin Zimmer, Paulo Sousa, Mathieu Siccard, Sophie Levesque, Freddie-
Jeanne Richard and Simon Allen.
I am grateful to the South Australian Department of Environment, Water and Natural
Resources (DEWNR) for kindly providing all the spatial layers used in this study. My special
thanks to Rosemary Paxinos and Alison Wright, for explaining details of the marine parks design
process in South Australia and giving ideas for the study.
Luckily, I was surrounded by ArcGIS specialists: many thanks also to professor Andrew
Lovett and my EMAE-mate Abdullah Nagy, for extra support with the software. Many thanks
also to our full-time EMAE statistics advisor Pamela Castañeda. For all the help with maps,
directions and interactions, I am so grateful to Juliana Pille Arnold, my official research partner,
with all her experience in daytime excursions and especially nocturnal field trips. I also thank
Maria Fernanda Tirado for expert assistance in ArcGIS, Zonation, and isolation; Edgar Bracho for
the political and philosophical advice, and Samy Bracho, for important help in understanding
foreign and unknown languages.
I thank my family and friends who are not around but somehow always supported me,
even though most of them have absolutely no idea what this work is all about. Two of them,
though, were really present despite being so far away: Muito, muito obrigada Aline Andrade e
Paula Aragão pelas conversas que me salvaram tantas vezes. For the ones that are around and
understand what I’m doing, each one of you had a special part in this experience and
consequently in this work.

Borges, R. 2013
5
“And forget not that the earth delights to feel your bare feet and the winds long to play with your hair.”
Kahlil Gibran

Borges, R. 2013
6
1. INTRODUCTION
1.1 Global marine biodiversity conservation
Human societies depend on services and goods provided by ecosystems (MEA, 2003). In
the marine environment, fish populations, for instance, provide food and employment on which
livelihoods depend, besides regulatory and provisional functions that benefit the ecosystems
(Holmlund and Hammer, 1999). Fish stocks as a resource have been collapsing at ever higher
rates, causing the decline of stability, water quality and recovery potential of seas and oceans.
Biodiversity loss is impairing the capacity of marine ecosystems to provide food, maintain water
quality, and recover from perturbations (Worm et al., 2006). These declining trends are still
reversible, and marine protected areas (MPAs) have been shown, for example, to help recover
species richness, which consequently leads to increases in fisheries productivity (Worm et al.,
2006; Worm et al., 2009; Vandeperre et al., 2011).
Protected areas play a role in in situ biodiversity conservation, and their management
effectiveness serves as a valid and measurable indicator of the achievement of conservation
goals (Chape et al., 2005). However, non-systematic design has led to the selection of protected
areas that do not contribute to biodiversity representativeness. The effectiveness of the
solutions aimed to meet conservation targets should therefore be reviewed, so that political
decisions can be taken to improve the representativeness and persistence of protected areas
(Margules and Pressey, 2000).
Random MPA designs yield less benefits compared to an optimized spatial management,
even when the same total area is protected (Rassweiler et al., 2012). The complementarity
approach, which is incorporated in reserve selection algorithms for systematic planning, is
more efficient than simple selection criteria (such as buying available intact land), even when
considering the biodiversity inventory costs. Governments and organizations should, therefore,
invest in high quality surveys before establishing protected areas networks (Balmford and
Gaston, 1999).

Borges, R. 2013
7
1.2 Marine conservation in South Australia
South Australian State waters hold one of the highest levels of marine endemism in
Australia (Edyvane, 1999), with 90-95% of its known species either endemic or of restricted
range. Its marine ecosystems are considered to have national and global conservation
significance due to the fact that 75% of the red algae, 85% of the fish species and 95% of the sea
grasses are endemic to the southern regions. However, with more than 90% of South
Australians living on or near the coast, biodiversity has suffered from increased human pressure
on the marine habitats. The local government has identified pollution, bioinvasion and
environmental destruction as the main threats in the region and has pointed to considerable
habitat losses in coastal wetlands, dune systems, reefs and seagrass communities (Government
of South Australia, 2004).
In order to help safeguard marine biodiversity within its coastal waters, the South
Australian government has recently expanded the marine parks system (DEH, 2009). However,
even though it comprises about 46% of the coastal waters, the final network still leaves out
some of South Australian biodiversity (Conservation Council SA, 2013). This might be especially
relevant for the most threatened sub-regions or for the no-take zones inside the parks, since
these zones make up only 6% of the State waters. The astonishingly large size of the network
might mislead the general public’s impression that biodiversity is being effectively protected
and result in what Agardy et al. (2011) called a “dangerous illusion of protection”.
Furthermore, reserve solutions were originally designed at the regional level taking into
consideration targets that reflected conservation priorities specific to each of the eight South
Australian marine bioregions (DEH, 2009). However, local-scale designs are less efficient in
representing biodiversity when compared to a more global approach (Strange et al., 2006;
Vasquez et al., 2008; Bladt et al., 2009).

Borges, R. 2013
8
1.3 Project aims and relevance for conservation
This study aimed to evaluate effectiveness and efficiency of the South Australian marine
parks system in representing marine biodiversity through 1) a gap analysis that compares no-
take and lower-protection zones (effectiveness); and 2) an administrative units analysis that
assesses the representation of biodiversity by state- versus bioregion-scale designs (design scale
efficiency). Moreover, a third aim of this study is to propose an expansion of the no-take areas
using different design methods, through which efficiency is once more analysed.
Such a combination of problem identification and proposal of alternative solutions is a
much-needed applied approach to the establishment of protected areas, and this case study
raises methodological issues that might help better understand and therefore improve spatial
planning in various conservation contexts.

Borges, R. 2013
9
2. METHODS
2.1 Study area
The study area comprises South Australian State waters: all estuarine and marine waters
from the highest tide to 3 nm out to sea including all bays and gulfs (Interim Marine and Coastal
Regionalisation Technical Group, 2008). Coastal waters cover an area of 60,282 km2 and 5,716
km of coastline. However, only the areas of the eight South Australian marine bioregions (Day et
al., 2008) inside coastal waters were considered; offshore islands were excluded from this study
(Figure 1).
2.2 Software
Reserve selection algorithms are mathematical tools that facilitate the spatial design of
protected areas by indicating which parts of the landscape have the highest conservation values,
according to the amount and type of biodiversity that they hold. Some of the main principles
that have been recently incorporated into these tools are 1) complementarity: selected sites
should complement each other so that the biodiversity they hold together meets defined targets;
and 2) connectivity: reserve selection favours sites in the landscape that are spatially connected,
which therefore reduces the total perimeter of the network of sites and minimises edge effects.
Borges, R. 2013
10
Figure 1. Study area divided into eight marine bioregions.
Bioregions
Coorong
Eucla
Eyre
Gulf St Vincent
Murat
North Spencer Gulf
Otway
Spencer Gulf
0 40 80 12020Nautical Miles
Ü

Borges, R. 2013
11
The publicly available software Zonation produces, through iterative cell removal, a
nested sequence of highly connected landscape structures, showing core areas of biodiversity
distribution and buffer zones of removed cells around them (Moilanen, 2007). The method used
here was Core Area Zonation, which yields a performance curve that plots the areas remaining
in the landscape versus the mean distribution of biodiversity features (species and habitats)
held inside these remaining areas. Core Area Zonation was chosen because it increases the
coverage of range-restricted features that occur in low-diversity regions (Moilanen et al., 2012)
and balances the protection granted to the fauna species considered here, which have very
small distribution relative to the other features.
The other algorithm used is Marxan, along with its graphical user interface Zonae Cogito
(Segan et al., 2011). Marxan deals with the “minimum set problem” and aims to design a reserve
system that achieves biodiversity representation targets for the smallest possible cost (Ball and
Possingham, 2000). This software uses an objective function that combines the cost of the
system and a penalty for not meeting targets for the conservation features, and it might also
consider a measure of fragmentation, through the user-defined parameter BLM – Boundary
Length Modifier- and a cost threshold penalty. A penalty for not achieving targets might also be
inserted in the design process, the Conservation Feature Penalty Factor – CFPF. The lower the
final value of the object function, the better the reserve system.
∑
∑
∑
Concerning the reserve design process, Marxan and Core Area Zonation differ in that
targets for Marxan are set for each biodiversity feature, and, in this study, a minimum cost to
achieve those targets is presented in terms of total size of the reserve system. Zonation, on the
other hand, shows a gradient in the study area that reflects the removal order of cells based on

Borges, R. 2013
12
their conservation value. Therefore, targets for Zonation are taken as the percentage of the area
remaining in the landscape, and the software presents how much of the distribution of
biodiversity features is retained at each stage of the cell removal process.
Zonation also has a reserve design tool based on administrative units, in which the eight
marine bioregions can be analysed together, with only a few easily-created extra input files. For
a similar approach, Marxan would require separate analyses, with individually-built input files,
a much more time-consuming approach.
The software ArcGIS Desktop 10.1 was used for the gap analysis, while the package SPSS
version 18 was used to perform the statistical tests.
2.3 Source and processing of datasets
The data used for all three analyses correspond to a sub-set of the spatial layers with the
distributions of species and habitats used by the South Australian Department of Environment,
Water and Natural Resources (DEWNR) to select the areas for the creation of the current
marine parks network at the initial stages of the selection process. Layers with the location of
the marine parks and the delimitation of the State waters and its bioregions were also provided
by DEWNR. Concerning resolution, the marine benthic habitats were mapped at a 1:100,000
scale. However, metadata in general were scarcely available.
The layers were independently formatted to meet requirements for Marxan, and then
further formatted for Zonation. Among the large dataset provided, the initial layers selected
were those with the best levels of certainty and resolution (avoiding, for example, labels such as
“likely to be present”). Other redundant layers were either merged with the primarily selected
ones or excluded from the analysis. (For further information on data limitation, refer to
Appendix A.)
Initial processing of GIS layers consisted of creating small buffers around data-points
(just so their area could be calculated – a requirement for Marxan) and spatially limiting all
features to the study area. For the Marxan analysis, the area was divided into 17,014 planning

Borges, R. 2013
13
cells with varying sizes of up to 4km2 (the ones at the edge were smaller), and the area of the
distribution of biodiversity features and protected areas inside each cell was tabulated. The final
processing of the tabulated areas and the creation of a list of features used in Marxan input files
was done using the interface Zonae Cogito. Other required input files were generated using
ArcGIS and Excel spreadsheets.
For Zonation, independent grid cells of 1km2 that fall completely inside the study area
were used and separately inserted into the software. Zonation, contrary to Marxan, requires that
all cells have the same size. Therefore, the total area for the two sets of analyses is not exactly
the same. Furthermore, the number of conservation features used as inputs for both pieces of
software was also slightly different (157 for Marxan and 151 for Zonation), because the layer
preparation procedure for Marxan is more sensitive to small-range features (For a complete list
of the biodiversity features used, refer to Appendix B). Thus, comparison between the solutions
proposed by the two packages is limited.
2.4 Effectiveness: gap analysis
A gap analysis consists of combining conservation features’ distributions (such as
species ranges) and maps of protected areas to show how well biodiversity is represented in the
existing network of reserves (Jennings, 2000; Rodrigues et al., 2004).
This part of the analysis, therefore, aims to assess the proportion of the distribution of
each biodiversity feature in the study area held by the marine protected areas. The approach
was applied to the entire South Australian coastal waters and to two individual bioregions: 1)
North Spencer Gulf, which presents high levels of endemism, especially in species that usually
occur in tropical ecosystems; and 2) Spencer Gulf, which, chosen due to high diversity and
productivity, supports important recreational and commercial fisheries (DEH, 2009).
This measure of effectiveness was done in regards to the protection level of the zones
inside the parks. Using the software ArcGIS Desktop 10.0, the area of the selected conservation

Borges, R. 2013
14
features was calculated: 1) inside the no-take zones; and 2) inside the remaining, lower-
protection zones. For the present study, the denomination “no-take zone” refers to the locally-
assigned Sanctuary Zones and Restricted Access Zones, which actually still allow “low-impact
fishing” (DEWNR, 2013). These areas could roughly be placed in the categories I to III from the
protected areas classification created by the International Union for Conservation of Nature
(IUCN). (For a more-detailed clarification of the zones designations used in this study, refer to
Appendix C).
2.5 Statistical analysis
A paired-sample t-test (t) was used to compare the total area of the distribution of
biodiversity features covered by no-takes zones with the expected distribution that would be
covered based solely on the size of these zones (random selection of MPAs). The square root of
the areas was used to perform the test. A Wilcoxon signed rank test (Z) was used to compare the
total area of the distributions of biodiversity features inside no-takes zones with the area
covered by the other lower-protection zones.
2.6 Efficiency: administrative units analysis
The aim of this analysis is to test if more biodiversity is supported in a system designed
at the bioregional or at the state scales. Using Zonation, the planning scales for reserve selection
were compared based on performance (species distribution coverage over different proportions
of the landscape being protected), for the 46%-area target, which corresponds to the extension
of the entire network.
To reduce the total network perimeter, a range of Boundary Length Penalty (BLP) values
were visually inspected and the parameter was then set to 1. Other general parameters were
kept as default. Specific parameters for the administrative units analysis followed similar
methodology to Moilanen and Arponen (2011): beta A = 0.999 and q = 0.001.
For the administrative units analysis, five scenarios were compared:

Borges, R. 2013
15
1) the state scale planning (mode 0);
2) a regional scale planning (mode 2) using South Australian eight marine bioregions:
2a) with equal weights applied to the bioregions and 2b) with weights proportional to the size
of each bioregion; and
3) a regional scale planning (mode 2) using eight randomly-created administrative units:
3a) with equal weights applied to the units and 3b) with weights proportional to the size of each
unit.
2.7 Expansion of the no-take zones
As a more practical approach to conservation management, an expansion of total size of
the no-take zones is proposed using two different pieces of software: Zonation and Marxan. In
the case of Zonation, the expansion occurs preferably inside the network, but is proposed as well
outside. The CBD-10%-area target was chosen for the expansion. The state and bioregional
planning scales were again examined in terms of efficiency for the expansion scenario, to check
how these results compare with those from the administrative units analysis.
The Marxan design, on the other hand, was set to only allow the expansion of the no-take
zones inside the system. Moreover, targets are defined per biodiversity feature. However, setting
a 10%-area target should yield a best solution that covers about 10% of South Australian coastal
waters, due to the fact that some features considered here are spread-out through the whole of
the study area. Marxan allows for easy identification of the conservation features for which the
10% could not be met inside the current marine parks system. Boundary Length Modifier - BLM
and Conservation Feature Penalty Factor – CFPF were calibrated to 1000 and 1, respectively, and
the software was set to propose 1000 solutions for the design problem. Here, reserve design is
done only at the state scale.

Borges, R. 2013
16
3. RESULTS
3.1 Effectiveness: gap analysis
The current South Australian network provides good coverage of biodiversity (mean =
66.59%, se = 3.543), but representation showed considerable variation among the different
species and habitats: some are well-represented while others have very little of their
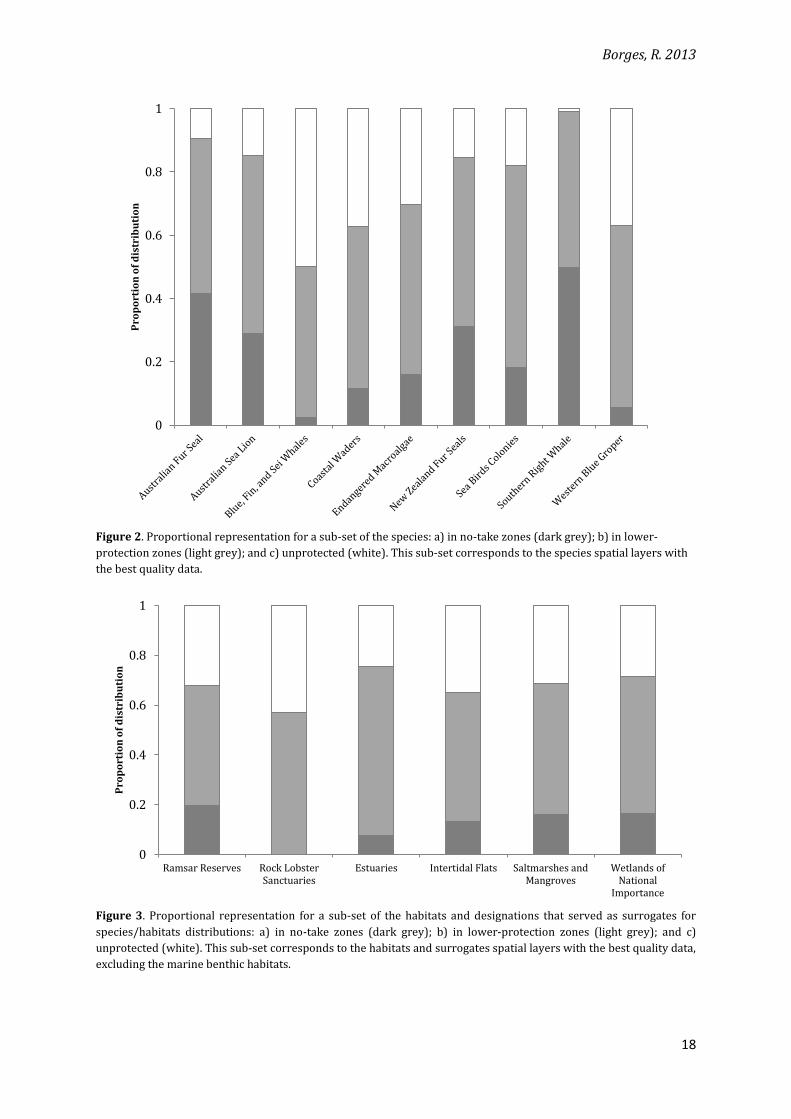
distributions protected (Figures 2 and 3). Coverage of the distribution of features in no-take
zones is significantly smaller than that inside the lower-protection-level zones (Wilcoxon signed
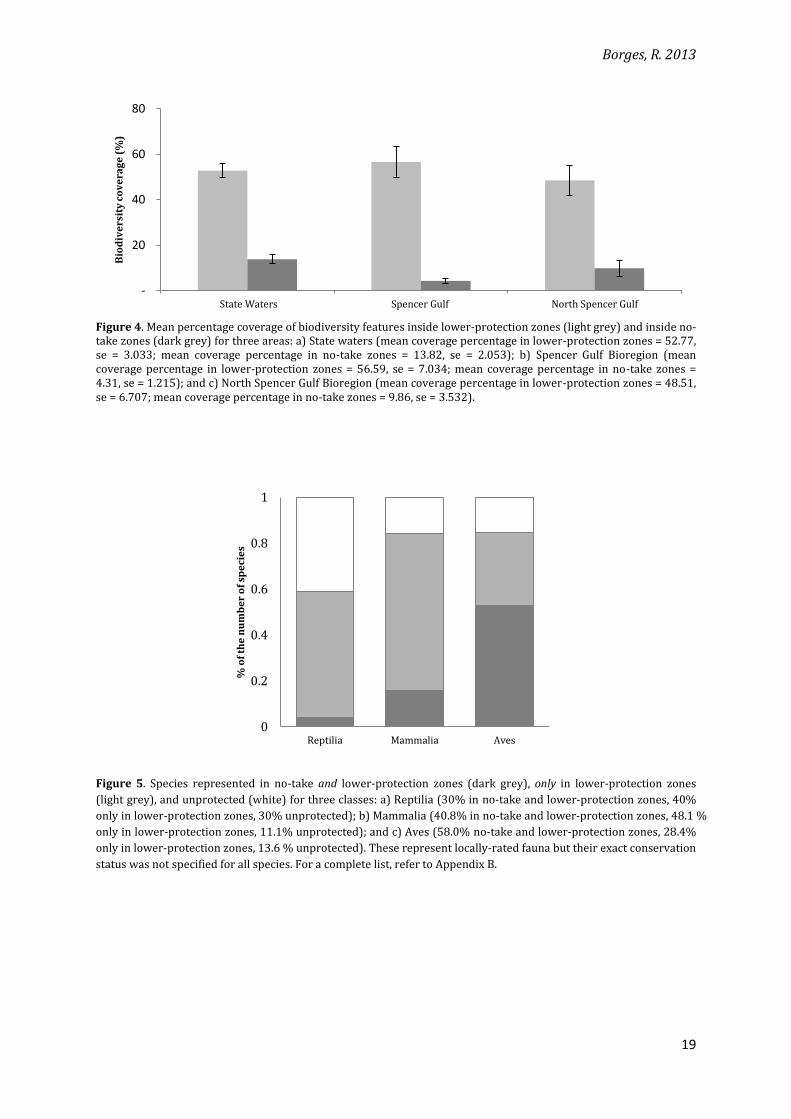
rank test Z = -5.214, df = 38, p < 0.001, N = 39) (Figure 4). Furthermore, the proportion of
species and habitats in no-take zones is not significantly different from a random design
(paired-sample t-test t = 0.869, df = 38, p = 0.390, N = 39).
The same pattern is observed for both the Spencer Gulf and the North Spencer Gulf
bioregions (Figure 4). Coverage for each feature in no-take zones is significantly smaller than
that inside the other zones (Spencer Gulf: Wilcoxon signed rank test Z = -4.045, df = 24, p <
0.001; North Spencer Gulf: Z = -3.622, df = 19, p < 0.001). Coverage for each feature in no-take
zones is not significantly different from what would be expected based solely on the size of
these zones in comparison to the networks in each of these bioregions (Spencer Gulf: t = -0.510,
df = 24, p = 0.614, N = 25; North Spencer Gulf: t = 0.991; df = 19; p = 0.334, N = 20). Moreover,
the proportion of no-take zones inside the marine parks is slightly larger than for the whole of
the State waters: 7% of the total bioregion area for Spencer Gulf and 9% for North Spencer Gulf.
(For a detailed description of the proportions of biodiversity features inside the different types
of zones, refer to Appendix D.)
In terms of number of species, the class Aves seems to be well-represented in all zones
at the State waters level. Classes Mammalia and Reptilia, however, follow the general pattern of
low-coverage in no-take zones (Figure 5).

Borges, R. 2013
17
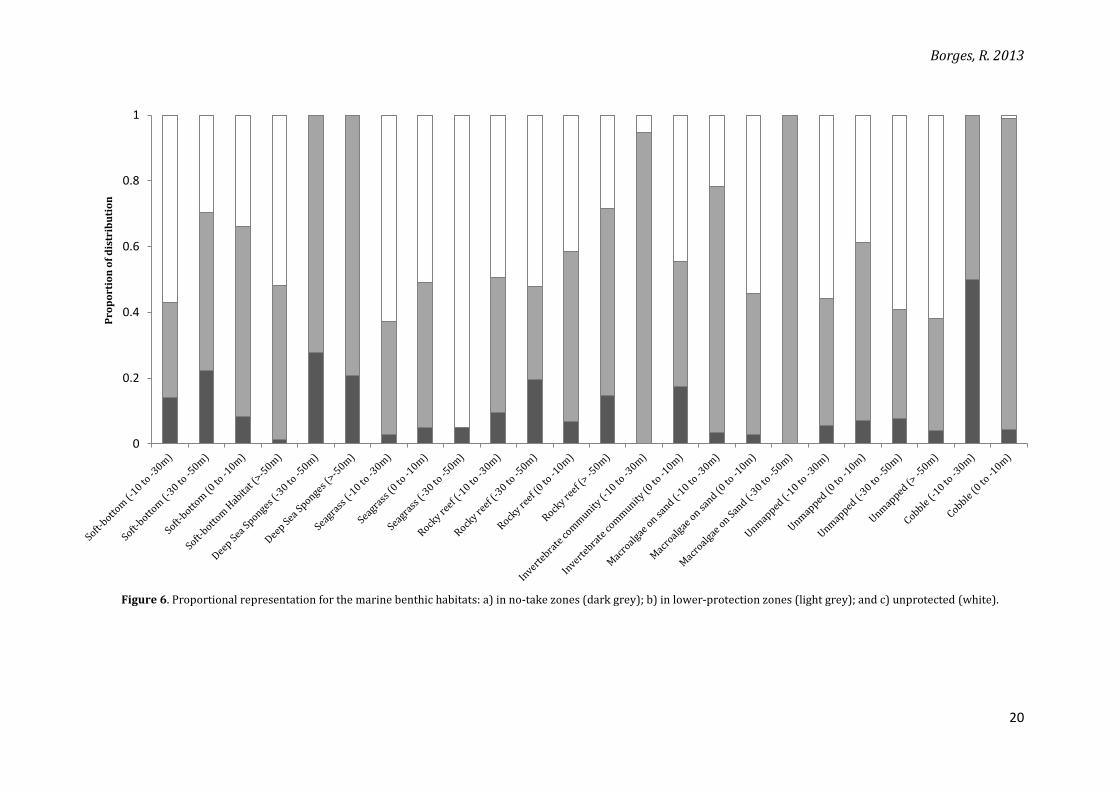
The marine benthic habitats have large proportions of their distributions inside the
parks: mostly between 20 and 50% and a large amount with above-50% representation (Figure
6). However, when only no-take zones are considered, most habitats have a maximum of 20% of
their distributions covered (only 4 out of 24 habitats have more 20% inside no-take zones). This
result is not negative per se, since no-take zones make up only 6% of the study area. However,
there is a considerable contrast between the two types of zones considered, and no-take areas
are able to represent much less biodiversity than the lower-protection ones.
Borges, R. 2013
18
Figure 2. Proportional representation for a sub-set of the species: a) in no-take zones (dark grey); b) in lower-
protection zones (light grey); and c) unprotected (white). This sub-set corresponds to the species spatial layers with
the best quality data.
Figure 3. Proportional representation for a sub-set of the habitats and designations that served as surrogates for
species/habitats distributions: a) in no-take zones (dark grey); b) in lower-protection zones (light grey); and c)
unprotected (white). This sub-set corresponds to the habitats and surrogates spatial layers with the best quality data,
excluding the marine benthic habitats.
0
0.2
0.4
0.6
0.8
1
Ramsar Reserves Rock LobsterSanctuaries
Estuaries Intertidal Flats Saltmarshes andMangroves
Wetlands ofNational
Importance
Pro
po
rtio
n o
f d
istr
ibu
tio
n
0
0.2
0.4
0.6
0.8
1
Pro
po
rtio
n o
f d
istr
ibu
tio
n
Borges, R. 2013
19
Figure 4. Mean percentage coverage of biodiversity features inside lower-protection zones (light grey) and inside no-take zones (dark grey) for three areas: a) State waters (mean coverage percentage in lower-protection zones = 52.77, se = 3.033; mean coverage percentage in no-take zones = 13.82, se = 2.053); b) Spencer Gulf Bioregion (mean coverage percentage in lower-protection zones = 56.59, se = 7.034; mean coverage percentage in no-take zones = 4.31, se = 1.215); and c) North Spencer Gulf Bioregion (mean coverage percentage in lower-protection zones = 48.51, se = 6.707; mean coverage percentage in no-take zones = 9.86, se = 3.532).
Figure 5. Species represented in no-take and lower-protection zones (dark grey), only in lower-protection zones
(light grey), and unprotected (white) for three classes: a) Reptilia (30% in no-take and lower-protection zones, 40%
only in lower-protection zones, 30% unprotected); b) Mammalia (40.8% in no-take and lower-protection zones, 48.1 %
only in lower-protection zones, 11.1% unprotected); and c) Aves (58.0% no-take and lower-protection zones, 28.4%
only in lower-protection zones, 13.6 % unprotected). These represent locally-rated fauna but their exact conservation
status was not specified for all species. For a complete list, refer to Appendix B.
-
20
40
60
80
State Waters Spencer Gulf North Spencer Gulf
Bio
div
ers
ity
co
ve
rag
e (
%)
0
0.2
0.4
0.6
0.8
1
Reptilia Mammalia Aves
% o
f th
e n
um
be
r o
f sp
eci
es
Borges, R. 2013
20
Figure 6. Proportional representation for the marine benthic habitats: a) in no-take zones (dark grey); b) in lower-protection zones (light grey); and c) unprotected (white).
0
0.2
0.4
0.6
0.8
1P
rop
ort
ion
of
dis
trib
uti
on

21
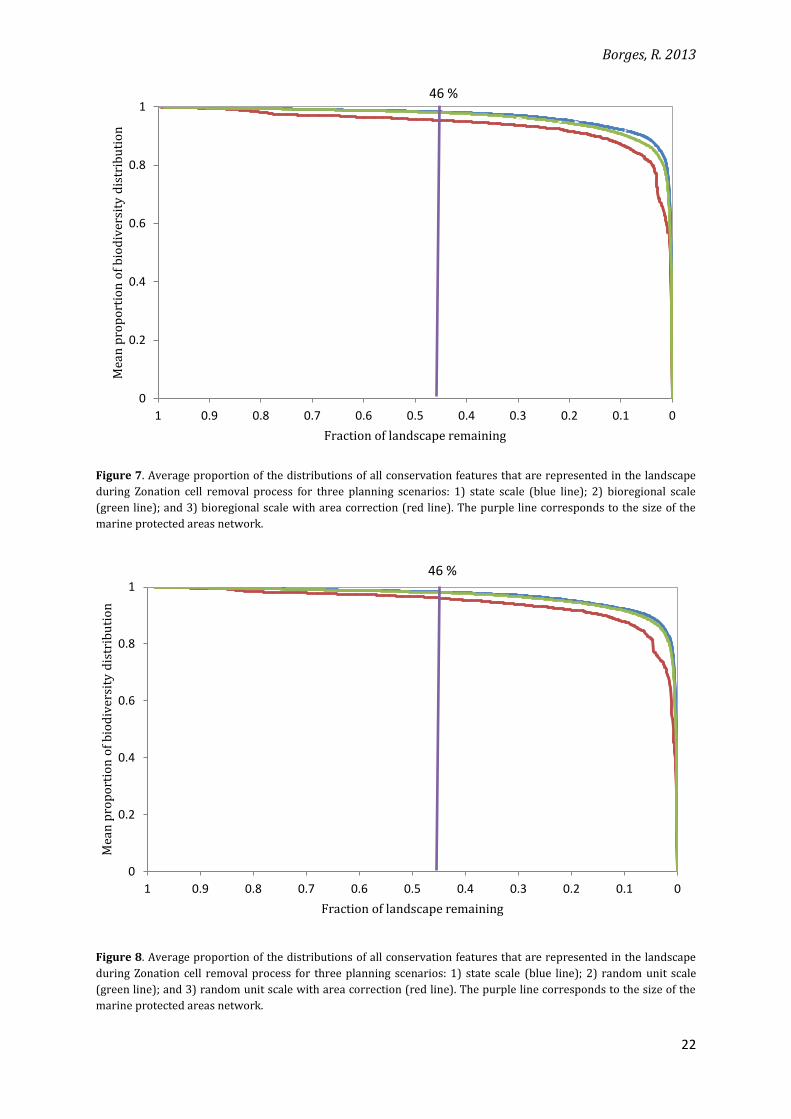
3.2 Efficiency: administrative units analysis
For most of the core-area targets (including the 46% corresponding to the size of the
network), there is no difference in efficiency of biodiversity representation between the state
and bioregional scale scenarios (Figure 7). Only for a small core-area target (< 10% of the
landscape, right-hand side of Figure 7), the area-corrected bioregional planning scale (scenarios
2b and 3b) shows a decrease in the proportion of biodiversity distribution represented in the
remaining landscape. The same pattern is observed when random units are used (Figure 8).
For all the scenarios, performance in terms of biodiversity coverage is considerably
higher than a random selection: at the 46%-area target, for instance, 98% of the mean
distribution of species and habitats is retained in the landscape, except for the corrected-area
scenarios, which yielded around 96% of biodiversity coverage. These results represent a more
than doubled efficiency when compared to a random 46%-coverage of biodiversity, from a
possible non-systematic reserve design strategy.
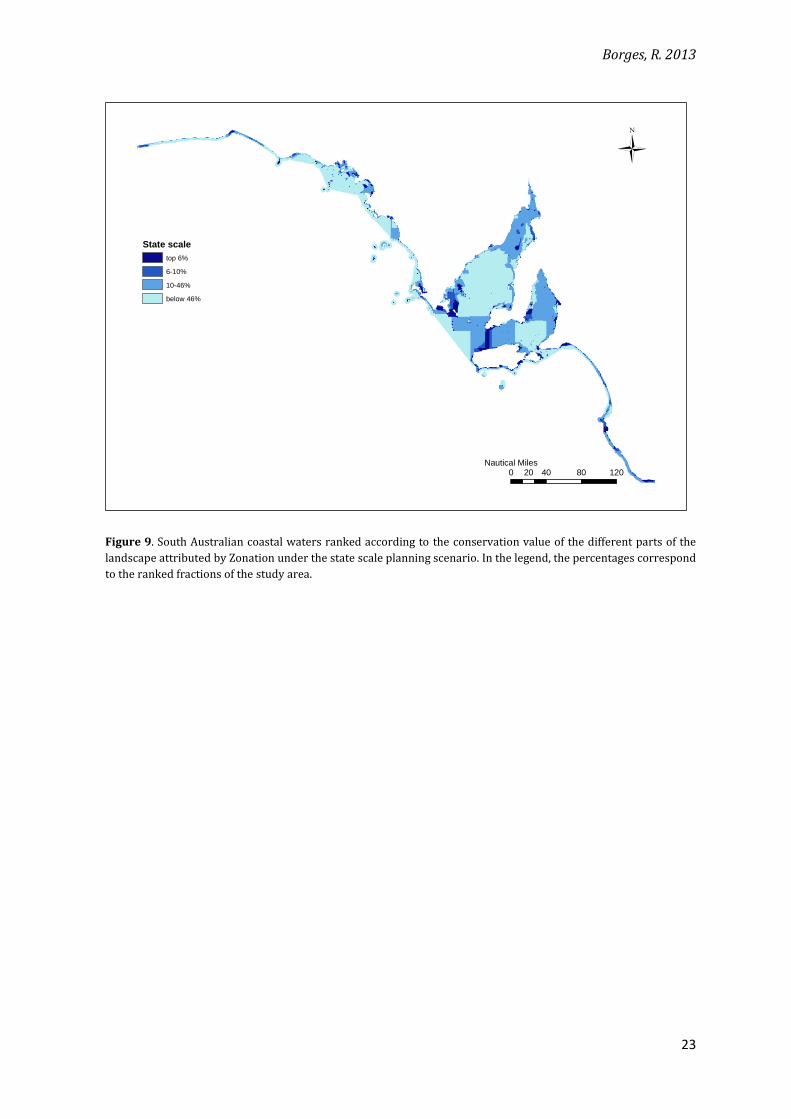
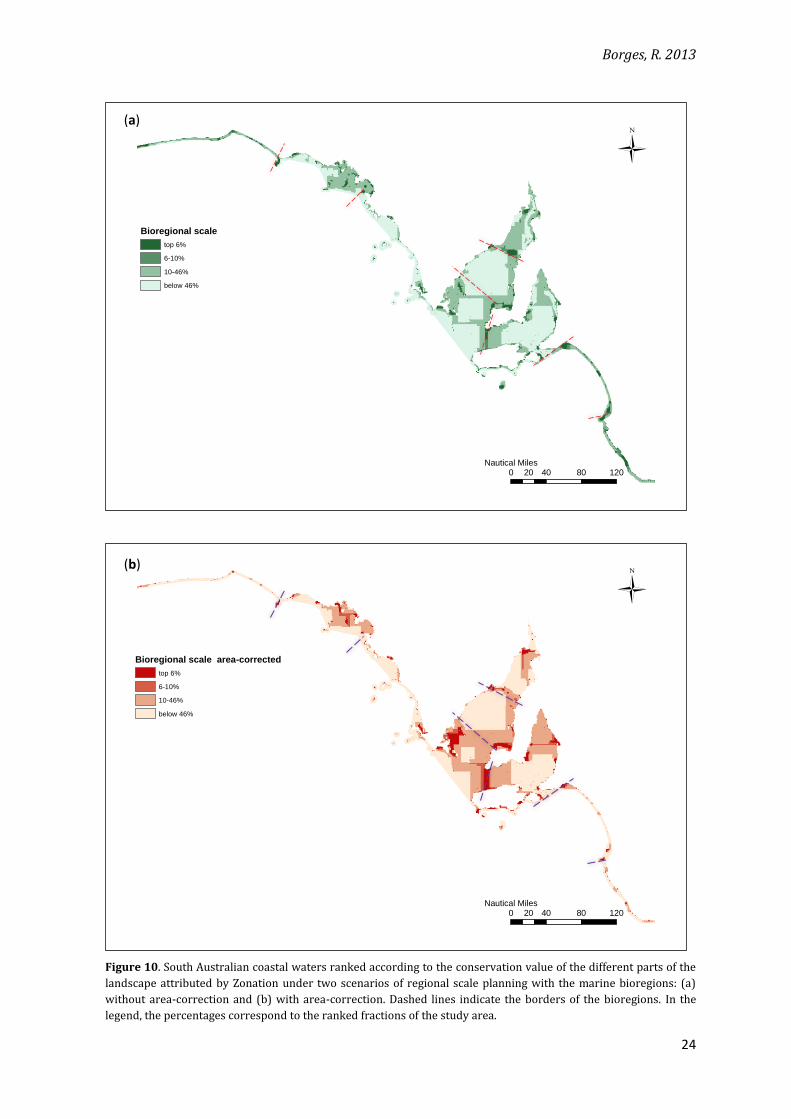
Spatial differences in the landscape ranking can be visualised for the planning scenarios
(Figures 9 and 10). A concentration of highly-ranked areas near the border between two
bioregions (what Moilanen et al. [2012] called “edge artefacts”) can be detected in the regional
planning scale for both the bioregional and the randomly-created units scenarios. It is
noteworthy that a concentration of highly ranked landscape near some of the borders can be
seen even at the state scale planning scenario, indicating areas that are of high conservation
value independently of using administrative units or not.
Another interesting observation about the area-corrected bioregional planning is that,
despite the more detectable loss in efficiency, edge artefacts are less evident in their maps than
in the ones for the regional scale without area-correction, at which design is more efficient. This
indicates that the formation of edge artefacts and loss in efficiency are not directly related.
Borges, R. 2013
22
Figure 7. Average proportion of the distributions of all conservation features that are represented in the landscape
during Zonation cell removal process for three planning scenarios: 1) state scale (blue line); 2) bioregional scale
(green line); and 3) bioregional scale with area correction (red line). The purple line corresponds to the size of the
marine protected areas network.
Figure 8. Average proportion of the distributions of all conservation features that are represented in the landscape
during Zonation cell removal process for three planning scenarios: 1) state scale (blue line); 2) random unit scale
(green line); and 3) random unit scale with area correction (red line). The purple line corresponds to the size of the
marine protected areas network.
0
0.2
0.4
0.6
0.8
1
00.10.20.30.40.50.60.70.80.91
Mea
n p
rop
ort
ion
of
bio
div
ersi
ty d
istr
ibu
tio
n
Fraction of landscape remaining
46 %
0
0.2
0.4
0.6
0.8
1
00.10.20.30.40.50.60.70.80.91
Mea
n p
rop
ort
ion
of
bio
div
ersi
ty d
istr
ibu
tio
n
Fraction of landscape remaining
46 %
Borges, R. 2013
23
Figure 9. South Australian coastal waters ranked according to the conservation value of the different parts of the
landscape attributed by Zonation under the state scale planning scenario. In the legend, the percentages correspond
to the ranked fractions of the study area.
State scale
top 6%
6-10%
10-46%
below 46%
Ü
0 40 80 12020Nautical Miles
Borges, R. 2013
24
Figure 10. South Australian coastal waters ranked according to the conservation value of the different parts of the
landscape attributed by Zonation under two scenarios of regional scale planning with the marine bioregions: (a)
without area-correction and (b) with area-correction. Dashed lines indicate the borders of the bioregions. In the
legend, the percentages correspond to the ranked fractions of the study area.
Ü
0 40 80 12020Nautical Miles
Bioregional scale
top 6%
6-10%
10-46%
below 46%
Ü
0 40 80 12020Nautical Miles
Bioregional scale area-corrected
top 6%
6-10%
10-46%
below 46%
(a)
(b)

Borges, R. 2013
25
3.3 Expansion of the no-take zones
3.3.1 Zonation
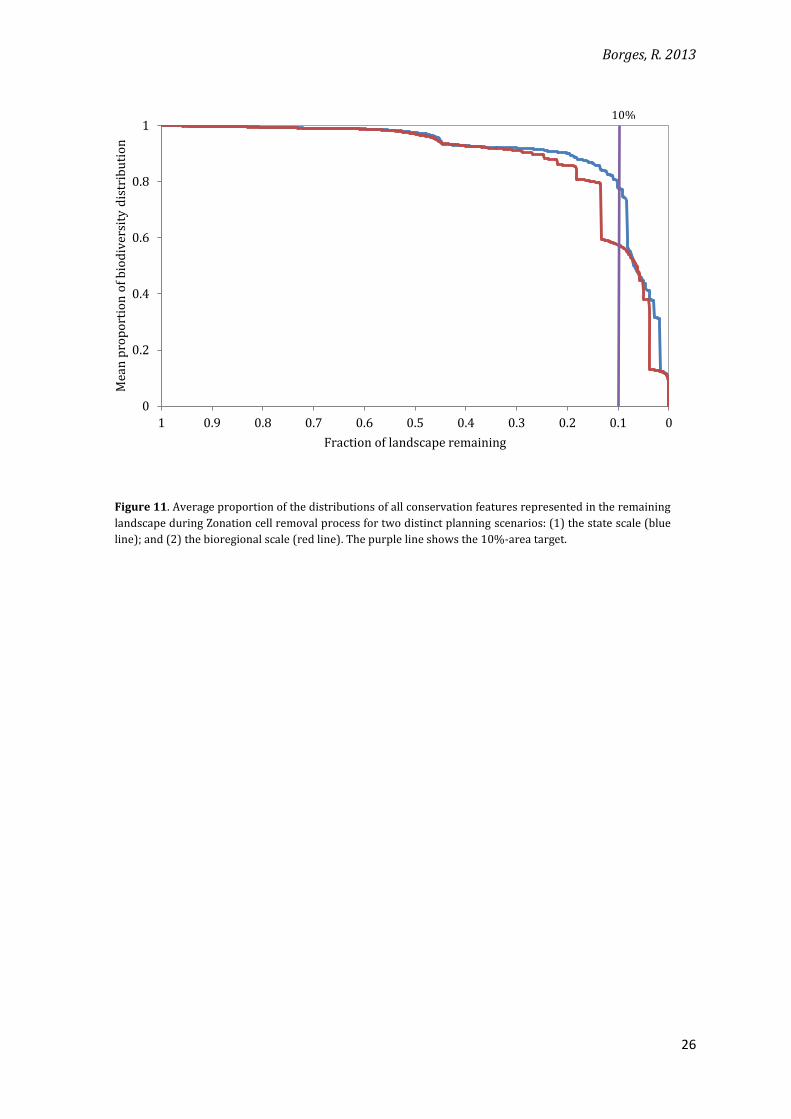
Contrary to the results from the previous section, efficiency in the expansion of the no-
take zones inside the network is considerably reduced at the bioregional scale when compared
to the state scale (from around 0.78 to 0.58 of mean proportion of biodiversity retained in the
top 10% of the landscape) (Figure 11). The loss in efficiency is not accompanied by an increase
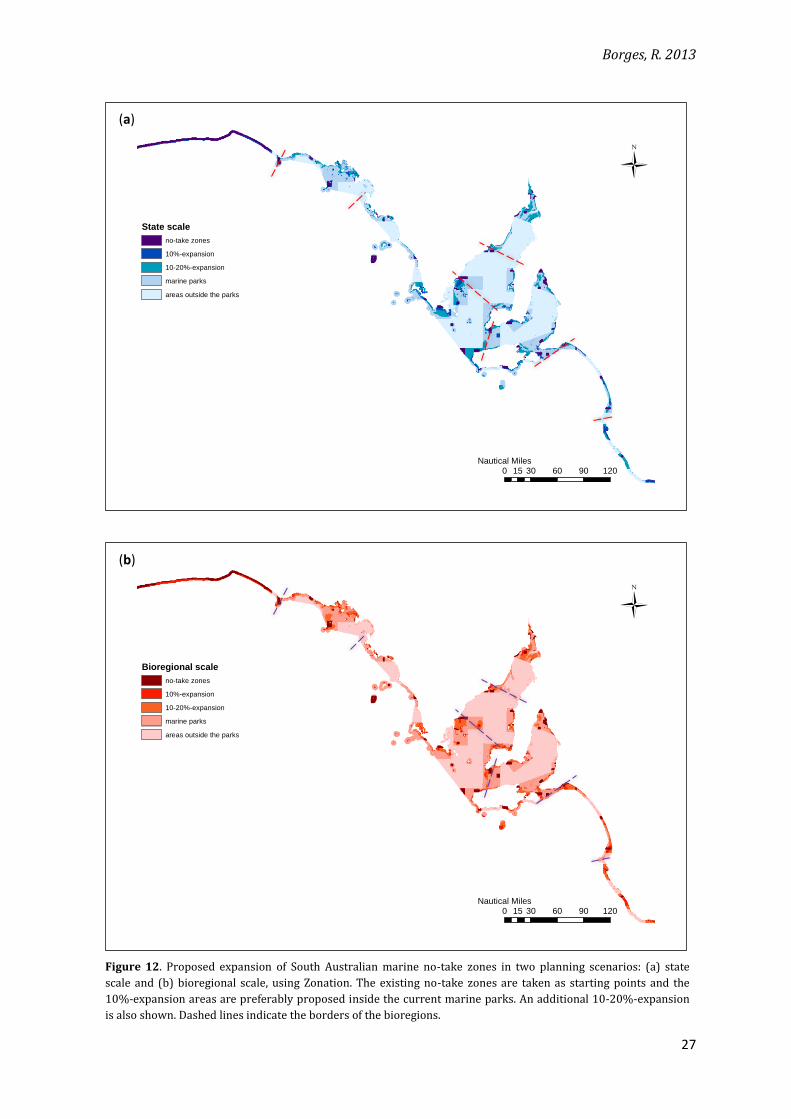
in the edge artefacts (Figure 12). This observation again reinforces that idea that edge artefacts
are not directly related to loss in efficiency.
3.3.2 Marxan
Marxan yielded solutions with a mean size of 6,183 km2 (se = 18.179), which
corresponds to about 10% of the study area. On average, 18 out of 157 conservation features
could not have their 10%-target met inside the current marine parks system.
3.3.3 Marxan versus Zonation solutions
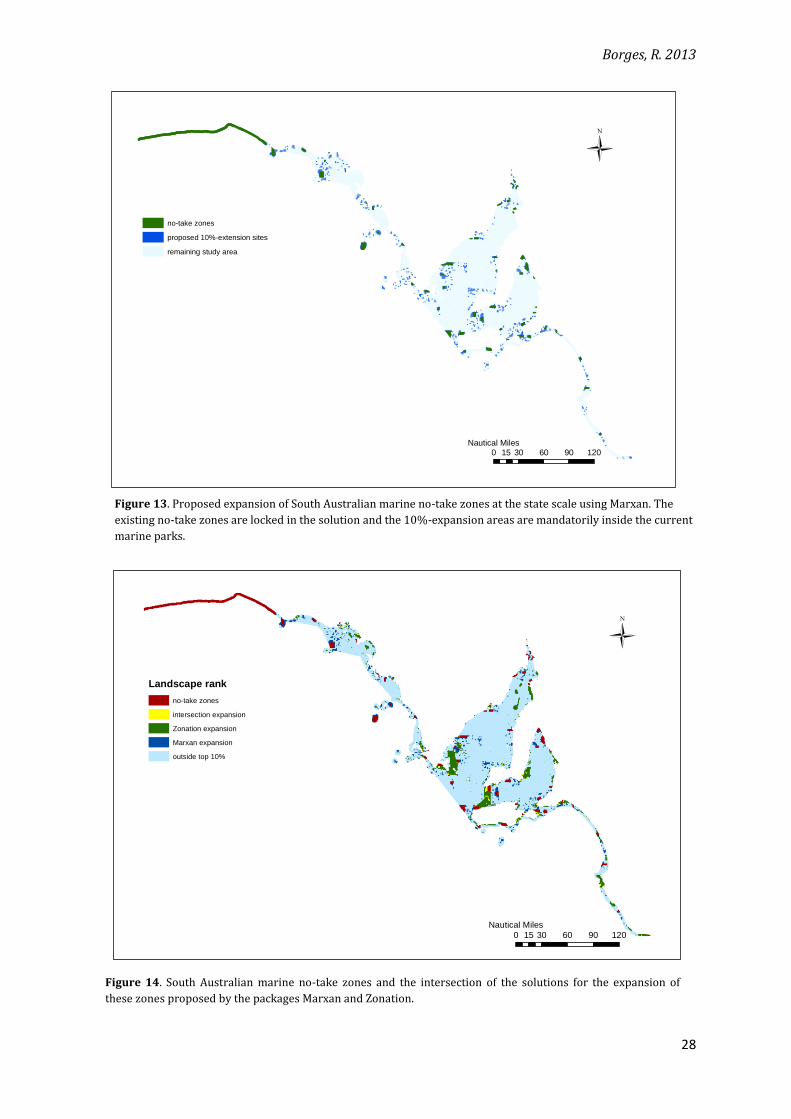
Marxan proposes less clustered sites than the 10%-area expansion proposed by
Zonation (Figure 13). Compactness could be enhanced in Marxan by increasing the boundary
length penalty. Moreover, the two approaches show little overlap in area: about 460 km2, less
than 1% of the coastal waters or 7.6% of the proposed expansion of the no-take zones (Figure
14). This spatial divergence in the solutions is probably related to the different emphasis on
connectivity in the designs using each of the packages. However, a combination of different
areas proposed by the two packages might be beneficial to meet local requirements for the
expansion of the no-take zones, since higher clustering might be preferred in some areas and
not well-accepted in others.
Borges, R. 2013
26
Figure 11. Average proportion of the distributions of all conservation features represented in the remaining
landscape during Zonation cell removal process for two distinct planning scenarios: (1) the state scale (blue
line); and (2) the bioregional scale (red line). The purple line shows the 10%-area target.
0
0.2
0.4
0.6
0.8
1
00.10.20.30.40.50.60.70.80.91
Mea
n p
rop
ort
ion
of
bio
div
ersi
ty d
istr
ibu
tio
n
Fraction of landscape remaining
10%
Borges, R. 2013
27
Figure 12. Proposed expansion of South Australian marine no-take zones in two planning scenarios: (a) state
scale and (b) bioregional scale, using Zonation. The existing no-take zones are taken as starting points and the
10%-expansion areas are preferably proposed inside the current marine parks. An additional 10-20%-expansion
is also shown. Dashed lines indicate the borders of the bioregions.
Ü
0 30 60 90 12015Nautical Miles
State scale
no-take zones
10%-expansion
10-20%-expansion
marine parks
areas outside the parks
Ü
0 30 60 90 12015Nautical Miles
Bioregional scale
no-take zones
10%-expansion
10-20%-expansion
marine parks
areas outside the parks
(a)
(b)
Borges, R. 2013
28
Figure 13. Proposed expansion of South Australian marine no-take zones at the state scale using Marxan. The
existing no-take zones are locked in the solution and the 10%-expansion areas are mandatorily inside the current
marine parks.
Figure 14. South Australian marine no-take zones and the intersection of the solutions for the expansion of
these zones proposed by the packages Marxan and Zonation.
0 30 60 90 12015Nautical Miles
Ü
no-take zones
proposed 10%-extension sites
remaining study area
ÜLandscape rank
no-take zones
intersection expansion
Zonation expansion
Marxan expansion
outside top 10%
0 30 60 90 12015
Nautical Miles

Borges, R. 2013
29
3. DISCUSSION
4. 1 Gap analysis and expansion of no-take zones
The South Australian marine parks network supports a relatively good average
proportion of the distribution of the conservation features analysed (mean = 66.59%, se =
3.543).
Other studies evaluating the present representativeness of protected areas in the
marine environment, especially comparing different protection categories, are rarely found in
the peer-reviewed literature. There may be some evaluations in the “grey literature”, such as
technical reports, but these are often difficult to access. It is difficult, therefore, to compare the
present assessment to the performance of MPAs in other regions.
Comparisons can be drawn with global compilations and a recent national evaluation of
the Australia MPAs. For example, having 46% of State waters covered by parks in South
Australia is well above the global proportion of 1.4% of protected coastal shelves (Chape et al.,
2005), and it corresponds to a representation level that is even higher than the 36% for the
MPAs in Australian commonwealth waters (not including State waters) (Barr and Possingham,
2013).
On the other hand, no-take zones are ineffective in representing many features; some
species and habitats have very little of their distributions covered or none at all. Out of the 24
marine benthic habitats analysed, for instance, 14 have less than 10% for their distribution
covered, while 2 are not included at all in no-take zones. In terms of total area reserved, South
Australian no-take zones do worse than the national MPAs network, which covers 13% of
Australian jurisdiction waters (Barr and Possingham, 2013).
Benefits to biodiversity attributed to the establishment of MPAs, such as increased
density, depend on the level of protection in marine reserves. Examples in the recent literature
either point to better results of no-take areas when compared to lower-protection categories

Borges, R. 2013
30
(e.g., Scott et al., 2012; Lester and Halpern, 2008) or solely refer to positive impacts specifically
from no-take areas (e.g., Gormley et al., 2013; Rassweiler et al., 2012).
Therefore, an increase in South Australian no-take areas would be necessary to cover
the distributions of gap or poorly-represented features. The Great Barrier Reef MPA in Australia,
for instance, now has 33% of its area included in no-take zones (Fernandes et al., 2005). The
expansion of South Australian no-take areas would reduce the protection gap between the
different zones and re-assure the commitment of the State in effectively protecting its marine
biodiversity.
But socio-political challenges to this expansion clearly exist in South Australia. Some of the
population living near the established no-take zones have already expressed dissatisfaction, and
many fear that the ban to recreational fisheries will reduce the number of tourists and affect
local economies (Novak, 2012). On the other hand, fisheries can benefit from spillover of adults
and larvae from no-take areas (Lester et al., 2009). Therefore, a geographical balance of no-take
and fishing areas could increase catches and provide benefits to local communities in the long-
run, provided these no-take zones are systematic designed.
Moreover, Lester et al. (2009) in their global synthesis concluded that reserve size does
not influence performance and even small MPAs can increase average size of organisms.
Therefore, the expansion of the higher-protection-level zones could be done through the
creation of new no-take areas far from the existing ones, so other fishing zones could benefit
from spill-over. Spreading out no-take zones across the landscape could also avoid
concentrating social conflicts and overloading certain local communities with large areas of
restricted-access zones. However, the fact that these zones are often surrounded by the others
inside large parks could be an advantage if a buffer-like expansion was chosen in some areas, as
proposed by Zonation for certain parts of the landscape. Many small no-take areas also may
have additional costs in terms of ensuring enforcement.

Borges, R. 2013
31
Depending on specific targets for the expansion, the use of Zonation or Marxan, or even
a combination of their outputs would be beneficial. If connectivity is preferred for certain areas,
Zonation is a better approach, while a Marxan-based solution might be more suitable to spread-
out expansion sites while keeping a minimised cost. The spatial solutions presented here,
however, are illustrative of these possible scenarios, and new expansion exercises should be
done with a more complete dataset, which would include social-economic variables, especially
data on recreational and commercial fisheries. It may also be useful to have additional
information on connectivity of key organisms, for example, dispersal distances and pathways.
4.2 Administrative units analysis
The visual comparison between the scenarios from planning at different scales show a
divergence in terms of areas assigned as the most valuable for conservation, and edge artefacts
can be detected in the bioregion-level scenarios. However, overall network efficiency was only
minimally reduced for a reserve design based on strong emphasis on local conservation
priorities when compared to state-scale representation (Figures 7 and 8).
These results are contrary to previous studies (Vasquez et al., 2008; Bladt et al., 2009;
Kark et al., 2009), which show that coordinated solutions at larger scales tend to increase
flexibility and therefore efficiency in site selection. Setting conservation priorities at larger
scales avoid the extra cost, for example, of covering the distribution of biodiversity features that
are locally rare but more common when other geopolitical units are considered (Strange et al.,
2006), a sort of “artificial rarity” in the solution caused by the planning scale choice.
However, considering that the marine bioregions are distinctly affected by human
pressure (DEH, 2009), it is still ecologically important to assess how each one is doing in terms
of biodiversity representation. This evaluation could then serve as a proxy to how the
establishment of protected areas is distributed and to try and avoid the concentration of areas in

Borges, R. 2013
32
regions that might be considered “cheaper” for conservation because they are not of economical
interest.
Moilanen and Arponen (2011) and Moilanen et al. (2012) used a very similar software
analysis but also found some reduction in efficiency. The latter also detected edge artefacts, and
argued that these decrease the efficiency of conservation strategies. Even though edge artefacts
were detected in the present study, these are visually less intense and therefore may not
directly cause a loss in efficiency. Edge artefacts might only reflect a re-allocation of the areas
considered by Zonation to have the highest conservation values, but not necessarily imply that
overall more areas have to be reserved so that similar proportions of biodiversity are
represented. Spatial planning procedures should ideally check for these artefacts and analyse
how this re-allocation affects the design solutions in order to decide whether these effects
should be avoided or not in the planning process.
The similarity in the results for both the bioregions and the random units indicate that,
even though bioregions were established on ecological criteria, they do not necessarily
represent areas that contain most of a certain species’ or a habitat’s distribution. This would
make these features be considered rare when present in other regions and possibly cause
differences in efficiency for both scenarios. However, it can be speculated that a higher number
of smaller units would increase the artificial-rarity phenomenon and decrease the efficiency of
the planning design.
Nevertheless, a considerable loss in efficiency was observed in the expansion of the no-
take zones inside the current system. The features’ distributions that were previously larger in
the whole of the coastal waters become medium-sized when selection is favoured inside marine
parks, which therefore allows for an enhanced artificial or scale-related rarity. The emphasis on
regional conservation priorities assigns rarity to biodiversity features in the regions that might
not have been considered rare at the state scale priority setting. As reported in the previously-
mentioned papers, this requires more area to be preserved in the bioregional scenario so that

Borges, R. 2013
33
the same mean proportion of biodiversity is overall represented, since more species and
habitats are now being recognised by the software as rare.
But again this extra area is not necessarily located in areas where MPAs are crossed by
the border between two bioregions. Edge artefacts were not increased at the bioregional scale
in the expansion scenario, when compared to the scenarios that did not consider the MPAs,
despite the considerable loss of efficiency at the 10%-area target. Even though edge artefacts
have been reported to reduce the efficiency of the spatial design process (Moilanen et al., 2012),
this loss might be attributed to extra area demanded to cover a similar proportion of
biodiversity in other areas apart from the borders of the bioregions/units.
Therefore, the different effects of the design-scale choice, whether they cause edge
artefacts, or reduce efficiency, are largely dependent on 1) spatial characteristics of biodiversity
features’ distributions; 2) the study area, whether it is continuous or segregated (as in the
expansion scenario, which favoured selection inside the current MPAs); and 3) on specific
conservation goals, since considerable differences may only be found for a restricted interval of
area-targets.

Borges, R. 2013
34
4. CONCLUSIONS
i. At the state level, South Australian marine parks effectively cover the
distributions of most of the biodiversity analysed. The Spencer Gulf and North
Spencer Gulf bioregions have similar patterns of biodiversity representation as
that for the whole of the coastal waters. However, biodiversity representation is
considered ineffective inside no-take zones, both at the state level and for those
two bioregions.
ii. The design solutions at state and bioregional scales are spatially different, but
have similar efficiency, when no pre-established parks are considered. However,
in the context of a proposed extension of no-take zones inside the current MPAs,
the bioregional-level design covers a considerably smaller proportion of
biodiversity at the 10%-area target when compared to the state-scale solution.
iii. The bioregional solutions show concentrations of high-conservation-value areas
near the borders of the bioregions (edge artefacts), but these do not seem to be
the main cause for the loss of efficiency in the planning design. The reduction in
efficiency probably depends on spatial characteristics of the distribution of the
biodiversity features included in the planning exercise, as well as specific aspects
of the study area.
iv. Marxan and Zonation yield solutions that have little spatial overlap, but similar
total area. Zonation favours the connectivity of possible extension sites, but
comparing and combining both scenarios might be preferred to spread out new
no-take zones and consequently reduce the social conflicts that these new areas
could cause.

Borges, R. 2013
35
5. CONSERVATION IMPLICATIONS
One of the aims of this study was to raise awareness and stimulate discussion on a
methodological aspect of spatial planning: the design scale. This topic had not yet been
approached for the marine environment, and specific literature for the terrestrial context is still
scarce (Vasquez et al., 2008; Bladt et al., 2009; Kark et al., 2009), with only a few studies
applying similar methodology (see Moilanen et al., 2012; Moilanen and Arponen, 2011). None of
these has considered bioregions, established on ecological criteria, as the administrative units.
Moreover, no previous work has been applied to the context of an expansion of MPAs, in which
certain parts of the study area are favoured for selection.
The inclusion of these previously-unexplored aspects of the planning process allowed
for an unprecedented discussion on the design scale, which will hopefully be carried on by
further studies, since these new aspects have important practical influence on the design of
protected areas.
As an initial step of the planning process, practitioners should examine the spatial
characteristics of the biodiversity distribution data available, bearing in mind that they might
strongly influence the final configuration and cost of the network of reserves. Secondly, spatial
characteristics of biological data might interact with the planning area and different costs for the
landscape. Certain aspects of the design solutions might depend, for example, on whether a
completely new system is being established or if an expansion of existing protected areas is
being proposed. The interaction between spatial characteristics of biodiversity distribution and
the specific planning area could then influence the solutions provided by different design scales,
even when ecological sub-units (such as the South Australian marine bioregions) are used as
planning areas. This study suggests that a larger-scale approach to spatial planning is more or at
least equally efficient and should therefore be preferred, even though loss of efficiency or edge
artefacts might actually not be verified or, when existing, these might depend on the
management targets chosen.

Borges, R. 2013
36
The use of different conservation planning packages might be advantageous to 1)
investigate how different target setting strategies influence spatial configurations, 2) assess a
variety of parameters that indicate effectiveness and efficiency of the proposed system and are
only available in either one of the packages, and 3) combine the spatial output from the different
pieces of software to reach a preferred configuration.
In the case of the South Australian marine parks, an expansion of the no-take zones to a
minimum of a 10%-area target is strongly recommended, with special emphasis on the inclusion
of gap features and the increase of the coverage of rare/endemic/threatened species and
habitats. This expansion should be conducted at the state scale and should aim also to spread
out the new no-take zones throughout the coastal waters.
However, other relevant targets and principles might strongly influence and be even
more critical for the design of the expanded no-take zones. The inclusion of fisheries
productivity as a cost metric or the decision on whether to protect highly threatened or
relatively pristine areas should also be analysed in combination with the influence of the design
scale, in order to inspect a possible trade-off between emphasis on local conservation priorities
and maximisation of efficiency in biodiversity coverage.

Borges, R. 2013
37
6. REFERENCES
Agardy T., di Sciara G.N., Christie P. (2011). Mind the gap: Addressing the shortcomings of marine
protected areas through large scale marine spatial planning. Marine Policy, 35 (2) , 226-232.
Ball, I. R. and H. P. Possingham. (2000). Marxan (v1.8.2): Marine Reserve Design Using Spatially
Explicit Annealing, a Manual.
Ball, I. R., Possingham, H. P., & Watts, M. (2009). Marxan and relatives: software for spatial
conservation prioritisation. Spatial conservation prioritisation: quantitative methods and
computational tools. Oxford University Press, Oxford, United Kingdom, 185-195.
Balmford, A., & Gaston, K.J. (1999). Why biodiversity surveys are good value. Nature, 398(6724),
204-205.
Barr, L. M., & Possingham, H. P. (2013). Are outcomes matching policy commitments in Australian
marine conservation planning?. Marine Policy, 42, 39-48.
Bladt, J., Strange, N., Abildtrup, J., Svenning, J. C., & Skov, F. (2009). Conservation efficiency of
geopolitical coordination in the EU. Journal for Nature Conservation, 17(2), 72-86.
CBD. (2006). Decisions Adopted by the Conference of the Parties to the Convention on Biological
Diversity at its Eighth Meeting (Decision VIII/15, Annex IV). Convention on Biological Diversity,
Curitiba, Brazil.
Chape, S., Harrison, J., Spalding, M., & Lysenko, I. (2005). Measuring the extent and effectiveness of
protected areas as an indicator for meeting global biodiversity targets. Philosophical Transactions
of the Royal Society B: Biological Sciences, 360(1454), 443-455.
Conservation Council SA. (2013). South Australian marine biodiversity hotspots excluded from
protection. Available at: <http://www.conservationsa.org.au/media-releases/1128-sa-marine-
biodiversity-hotspots-not-protected.html>. Access on: 29th Jan 2013.
Day, V., Paxinos, R., Emmett, J., Wright, A., & Goecker, M. (2008). The Marine Planning Framework for
South Australia: A new ecosystem-based zoning policy for marine management. Marine
Policy, 32(4), 535-543.
Department for Environment and Heritage – DEH (2009). A technical report on the outer boundaries
of South Australia’s marine parks network. Department for Environment and Heritage, South
Australia.

Borges, R. 2013
38
Department of Environment, Water and Natural Resources - DEWNR. (2013). Marine Parks - Zones
Available at: <http://www.environment.sa.gov.au/marineparks/zones>. Access on 20th June
2013.
Edyvane, K. S. (1999). Conserving marine biodiversity in South Australia–Part 2–Identification of areas
of high conservation value in South Australia. Report to Department for Environment and Heritage,
Government of South Australia, Adelaide.
Fernandes, L., Day, J. O. N., Lewis, A., Slegers, S., Kerrigan, B., Breen, D. A. N., ... & Stapleton, K. (2005).
Establishing Representative No‐Take Areas in the Great Barrier Reef: Large‐Scale
Implementation of Theory on Marine Protected Areas. Conservation Biology, 19(6), 1733-1744.
Gormley, A. M., Slooten, E., Dawson, S., Barker, R. J., Rayment, W., du Fresne, S., & Bräger, S. (2012).
First evidence that marine protected areas can work for marine mammals. Journal of Applied
Ecology, 49(2), 474-480.
Government of South Australia. Living coast strategy for South Australia. Adelaide: Natural and
Cultural Heritage, Department for Environment and Heritage; 2004.
Holmlund, C. M., & Hammer, M. (1999). Ecosystem services generated by fish populations. Ecological
Economics, 29(2), 253-268.
Interim Marine and Coastal Regionalisation Technical Group. Interim marine and coastal
regionalisation for Australia: and ecosystem-based classification for marine and coastal
environments. Version 3.3. Environment Australia, Canberra, 1998.
Jennings, M. D. (2000). Gap analysis: concepts, methods, and recent results*.Landscape
ecology, 15(1), 5-20.
Kark, S., Levin, N., Grantham, H. S., & Possingham, H. P. (2009). Between-country collaboration and
consideration of costs increase conservation planning efficiency in the Mediterranean Basin.
Proceedings of the National Academy of Sciences, 106(36), 15368-15373.
Kearney, R., Buxton, C. D., & Farebrother, G. (2012). Australia’s no-take marine protected areas:
Appropriate conservation or inappropriate management of fishing? Marine Policy, 36(5), 1064-
1071.
Kujala, H., Araújo, M. B., Thuiller, W., & Cabeza, M. (2011). Misleading results from conventional gap
analysis–Messages from the warming north. Biological Conservation, 144(10), 2450-2458.

Borges, R. 2013
39
Lester, S. E., & Halpern, B. S. (2008). Biological responses in marine no-take reserves versus partially
protected areas. Marine Ecology Progress Series, 367, 49-56.
Lester, S. E., Halpern, B. S., Grorud-Colvert, K., Lubchenco, J., Ruttenberg, B. I., Gaines, S. D., ... &
Warner, R. R. (2009). Biological effects within no-take marine reserves: a global synthesis. Marine
Ecology Progress Series, 384, 33-46.
Margules, C. R., & Pressey, R. L. (2000). Systematic conservation planning. Nature, 405(6783), 243-
253.
MEA. (2003). Millennium ecosystem assessment. Ecosystems.
Moilanen, A. (2007). Landscape zonation, benefit functions and target-based planning: unifying
reserve selection strategies. Biological Conservation, 134(4), 571-579.
Moilanen, A., Anderson, B. J., Arponen, A., Pouzols, F. M., & Thomas, C. D. (2012). Edge artefacts and
lost performance in national versus continental conservation priority areas. Diversity and
Distributions, 19(2), 171-183.
Moilanen, A., & Arponen, A. (2011). Administrative regions in conservation: balancing local priorities
with regional to global preferences in spatial planning. Biological Conservation, 144(5), 1719-
1725.
Natural Resource Management Ministerial Council (2010) Australia’s Biodiversity Conservation
Strategy 2010–2030. Australian Government, Department of Sustainability, Environment, Water,
Population and Communities, Canberra. Available at:
<http://www.environment.gov.au/biodiversity/strategy>. Access on: 31 Jan 2013.
Novak, L. (2012). Final plans for state's 19 marine parks released by SA Government. Available at:
<http://www.adelaidenow.com.au/news/south-australia/final-plans-for-states-19-marine-
parks-released-by-sa-government/story-e6frea83-1226526557652>. Access on: 8th July 2013.
Pressey, R. L., & Bottrill, M. C. (2008). Opportunism, threats, and the evolution of systematic
conservation planning. Conservation Biology, 22(5), 1340-1345.
Rassweiler, A., Costello, C., & Siegel, D. A. (2012). Marine protected areas and the value of spatially
optimized fishery management. Proceedings of the National Academy of Sciences, 109(29), 11884-
11889.

Borges, R. 2013
40
Rodrigues, A. S., Akcakaya, H. R., Andelman, S. J., Bakarr, M. I., Boitani, L., Brooks, T. M., ... & Yan, X.
(2004). Global gap analysis: priority regions for expanding the global protected-area
network. BioScience, 54(12), 1092-1100.
Scott, R., Hodgson, D. J., Witt, M. J., Coyne, M. S., Adnyana, W., Blumenthal, J. M., ... & Godley, B. J.
(2012). Global analysis of satellite tracking data shows that adult green turtles are significantly
aggregated in Marine Protected Areas. Global Ecology and Biogeography, 21(11), 1053-1061.
Segan, D. B., Game, E. T., Watts, M. E., Stewart, R. R., & Possingham, H. P. (2011). An interoperable
decision support tool for conservation planning. Environmental Modelling & Software, 26(12),
1434-1441.
Stewart, R. R., & Possingham, H. P. (2002, August). A framework for systematic marine reserve
design in South Australia: a case study. In Inaugural World Congress on Aquatic Protected Areas,
Cairns,(unpublished).
Stewart, R. R., Noyce, T., & Possingham, H. P. (2003). Opportunity cost of ad hoc marine reserve
design decisions: an example from South Australia. Marine Ecology Progress Series, 253, 25-38.
Strange, N., Thorsen, B. J., & Bladt, J. (2006). Optimal reserve selection in a dynamic world. Biological
Conservation, 131(1), 33-41.
Vandeperre, F., Higgins, R. M., Sánchez‐Meca, J., Maynou, F., Goñi, R., Martín‐Sosa, P., ... & Santos, R. S.
(2011). Effects of no‐take area size and age of marine protected areas on fisheries yields: a meta‐
analytical approach. Fish and Fisheries, 12(4), 412-426.
Vazquez, L. B., Rodríguez, P., & Arita, H. T. (2008). Conservation planning in a subdivided world.
Biodiversity and Conservation, 17(6), 1367-1377.
Worm, B., Barbier, E. B., Beaumont, N., Duffy, J. E., Folke, C., Halpern, B. S., ... & Watson, R. (2006).
Impacts of biodiversity loss on ocean ecosystem services. Science, 314(5800), 787-790.
Worm, B., Hilborn, R., Baum, J. K., Branch, T. A., Collie, J. S., Costello, C., ... & Zeller, D. (2009).
Rebuilding global fisheries. Science, 325(5940), 578-585.

Borges, R. 2013
41
I. APPENDIX A
Potential limitations of the datasets used
The accuracy of the gap analysis results is limited by independent formatting of the spatial
layers, surrogate choice, little amount of metadata available and by the fact that interactions
with other protected areas are not considered. In the case of the rock lobster, for instance, only
point-data were available. A 1-km2 buffer was then arbitrarily established around those points,
ignoring the actual area of the sanctuaries. Therefore, sanctuary areas that might overlap with
the parks network were probably ignored, and these could include also high-protection zones.
Similarly, lobster sanctuaries, which were designated under a different law, might be considered
fully no-take zones, and the matter over conflicting regulations in intersections between areas
under different laws is unclear and beyond the scope of this study. The main message is that, in
general, features might be well represented in lower-protection zones but largely excluded from
the more strict areas. Further more, the results obtained are in accordance with a local
evaluation of the network (DEH, 2009), which validates the zones comparison in this study,
even though it did not use the exact same datasets or formatting procedures.

Borges, R. 2013
42
II. APPENDIX B
Conservation features used in the analyses
Marine habitats
Estuary
Intertidal flats
Saltmarshes and Mangroves
Wetlands of National Importance
Soft-bottom habitat ( -30m to -50m)
Soft-bottom habitat (>-50m)
Deep sea sponges (-30 to -50m)
Deep sea sponges (>-50m)
Seagrass (0 to -10m)
Seagrass (-10m to -30m)
Seagrass (-30 to -50m)
Rocky reef ( 0 to -10m)
Rocky reef (-10 to -30m)
Rocky reef (-30 to -50m)
Rocky reef (> -50m)
Invertebrate community (-10 to -30m)
Invertebrate community (0 to -10m)
Macroalgae on sand (0 to -10m)
Macroalgae on sand (-10 to -30m)
Macroalgae on Sand (-30 to -50m)*
Unmapped (0 to -10m)
Unmapped (-10 to -30m)
Unmapped (-30 to -50m)
Cobble (-10 to -30m)*
Cobble (0 to -10m)*
Relevant sites for fauna
Australian fur seal colonies
Australian sea lion colonies
Blue, fin, and sei whales
New Zealand fur seal colonies
Western blue groper survey sites
Southern right whale aggregation sites
Unspecified species
Coastal waders Endangered macroalgae Seabird colonies

Borges, R. 2013
43
Fauna
Bassiana trilineata* Caretta caretta Chelonia mydas Dermochelys coriacea Drysdalia coronoides Egernia cunninghami Lissolepis coventryi Morelia spilota Notechis scutatus Varanus rosenbergi Arctocephalus pusillus Arctocephalus tropicalis Balaenoptera acutorostrata* Balaenoptera edeni Balaenoptera musculus Balaenoptera physalus Berardius arnuxii Bettongia penicillata ogilbyi Caperea marginata Eubalaena australis Hydrurga leptonyx Hyperoodon planifrons Isoodon obesulus obesulus Kogia breviceps Kogia sima Lagostrophus fasciatus Leporillus conditor Macropus eugenii Megaptera novaeangliae Mesoplodon bowdoini Mesoplodon grayi Mirounga leonina Neophoca cinerea Petrogale lateralis pearsoni Physeter catodon Tasmacetus shepherdi Trichosurus vulpecula Acanthiza iredalei Acanthiza iredalei rosinae Actitis hypoleucos Anas rhynchotis Ardenna carneipes Ardeotis australis Arenaria interpres Biziura lobata Burhinus grallarius Calamanthus cautus Calamanthus pyrrhopygius Calidris alba Calidris melanotos Calidris subminuta Calidris tenuirostris Calyptorhynchus funereus Calyptorhynchus lathami halmaturinus Cereopsis novaehollandiae Charadrius leschenaultii Charadrius mongolus Cladorhynchus leucocephalus Corcorax melanoramphos Coturnix ypsilophora Dasyornis broadbenti Diomedea exulans Dromaius baudinianus Egretta garzetta

Borges, R. 2013
44
Egretta sacra Excalfactoria chinensis Falco peregrinus Gallinago hardwickii Haematopus fuliginosus Haematopus longirostris Haliaeetus leucogaster Halobaena caerulea Larus dominicanus Lichenostomus cratitius Limosa lapponica Limosa limosa Macronectes giganteus Macronectes halli Melanodryas cucullata Myiagra inquieta Neophema chrysostoma Neophema elegans Neophema petrophila Northiella haematogaster Numenius madagascariensis Numenius phaeopus Oxyura australis Pachycephala inornata Pandion haliaetus Petroica boodang Philomachus pugnax* Phoebetria fusca Phoebetria palpebrata Plegadis falcinellus Pluvialis fulva Podiceps cristatus Psophodes nigrogularis Psophodes nigrogularis leucogaster Stagonopleura bella Stagonopleura guttata Sterna hirundo Sternula albifrons Sternula nereis Stictonetta naevosa Stipiturus malachurus Stipiturus malachurus halmaturinus Stipiturus malachurus parimeda Strepera versicolor Thalassarche cauta Thalassarche chlororhynchos Thalassarche chrysostoma Thalassarche melanophris Thalassarche salvini Thinornis rubricollis Tringa brevipes Tringa glareola Turnix varius Xenus cinereus
Designations serving as surrogates for species
Ramsar reserves
Rock lobster sanctuaries
* Features not included in the Zonation analyses.
Borges, R. 2013
45
III. APPENDIX C
Description of the zones designations
Local denomination Description Designation in this study
General managed use No change to existing use, but managed as part of the park. All recreational activities, including fishing, are allowed.
lower-protection zone
Habitat protection Protects the sea floor. All recreational activities, including fishing, are allowed. Prawn trawling is prohibited from March 2013.
lower-protection zone
Sanctuary zones Areas of high conservation value set aside for conservation and low-impact recreation. No fishing is allowed in these zones from 1 October 2014, but diving, surfing, swimming etc. are welcome.
no-take zone
Restricted access Areas that are off limits to the public (no entry).
no-take zone
Source: DEWNR (2013).
Borges, R. 2013
46
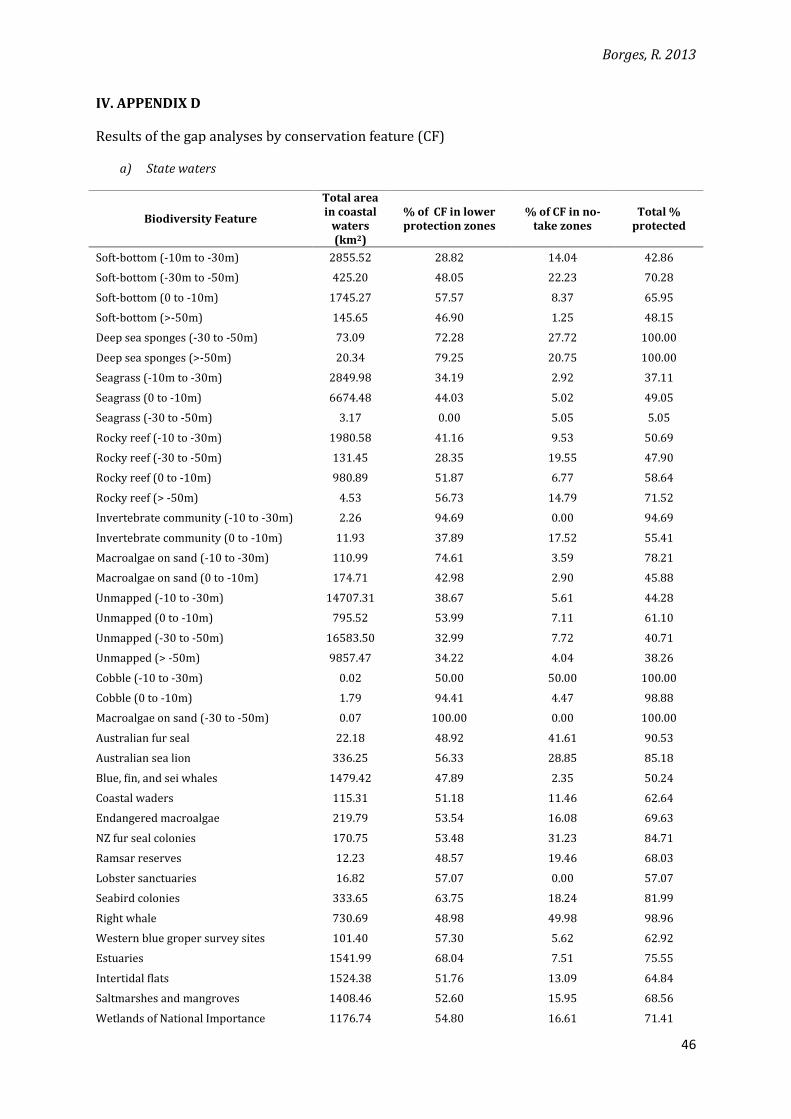
IV. APPENDIX D
Results of the gap analyses by conservation feature (CF)
a) State waters
Biodiversity Feature
Total area in coastal
waters (km2)
% of CF in lower protection zones
% of CF in no-take zones
Total % protected
Soft-bottom (-10m to -30m) 2855.52 28.82 14.04 42.86
Soft-bottom (-30m to -50m) 425.20 48.05 22.23 70.28
Soft-bottom (0 to -10m) 1745.27 57.57 8.37 65.95
Soft-bottom (>-50m) 145.65 46.90 1.25 48.15
Deep sea sponges (-30 to -50m) 73.09 72.28 27.72 100.00
Deep sea sponges (>-50m) 20.34 79.25 20.75 100.00
Seagrass (-10m to -30m) 2849.98 34.19 2.92 37.11
Seagrass (0 to -10m) 6674.48 44.03 5.02 49.05
Seagrass (-30 to -50m) 3.17 0.00 5.05 5.05
Rocky reef (-10 to -30m) 1980.58 41.16 9.53 50.69
Rocky reef (-30 to -50m) 131.45 28.35 19.55 47.90
Rocky reef (0 to -10m) 980.89 51.87 6.77 58.64
Rocky reef (> -50m) 4.53 56.73 14.79 71.52
Invertebrate community (-10 to -30m) 2.26 94.69 0.00 94.69
Invertebrate community (0 to -10m) 11.93 37.89 17.52 55.41
Macroalgae on sand (-10 to -30m) 110.99 74.61 3.59 78.21
Macroalgae on sand (0 to -10m) 174.71 42.98 2.90 45.88
Unmapped (-10 to -30m) 14707.31 38.67 5.61 44.28
Unmapped (0 to -10m) 795.52 53.99 7.11 61.10
Unmapped (-30 to -50m) 16583.50 32.99 7.72 40.71
Unmapped (> -50m) 9857.47 34.22 4.04 38.26
Cobble (-10 to -30m) 0.02 50.00 50.00 100.00
Cobble (0 to -10m) 1.79 94.41 4.47 98.88
Macroalgae on sand (-30 to -50m) 0.07 100.00 0.00 100.00
Australian fur seal 22.18 48.92 41.61 90.53
Australian sea lion 336.25 56.33 28.85 85.18
Blue, fin, and sei whales 1479.42 47.89 2.35 50.24
Coastal waders 115.31 51.18 11.46 62.64
Endangered macroalgae 219.79 53.54 16.08 69.63
NZ fur seal colonies 170.75 53.48 31.23 84.71
Ramsar reserves 12.23 48.57 19.46 68.03
Lobster sanctuaries 16.82 57.07 0.00 57.07
Seabird colonies 333.65 63.75 18.24 81.99
Right whale 730.69 48.98 49.98 98.96
Western blue groper survey sites 101.40 57.30 5.62 62.92
Estuaries 1541.99 68.04 7.51 75.55
Intertidal flats 1524.38 51.76 13.09 64.84
Saltmarshes and mangroves 1408.46 52.60 15.95 68.56
Wetlands of National Importance 1176.74 54.80 16.61 71.41
Borges, R. 2013
47
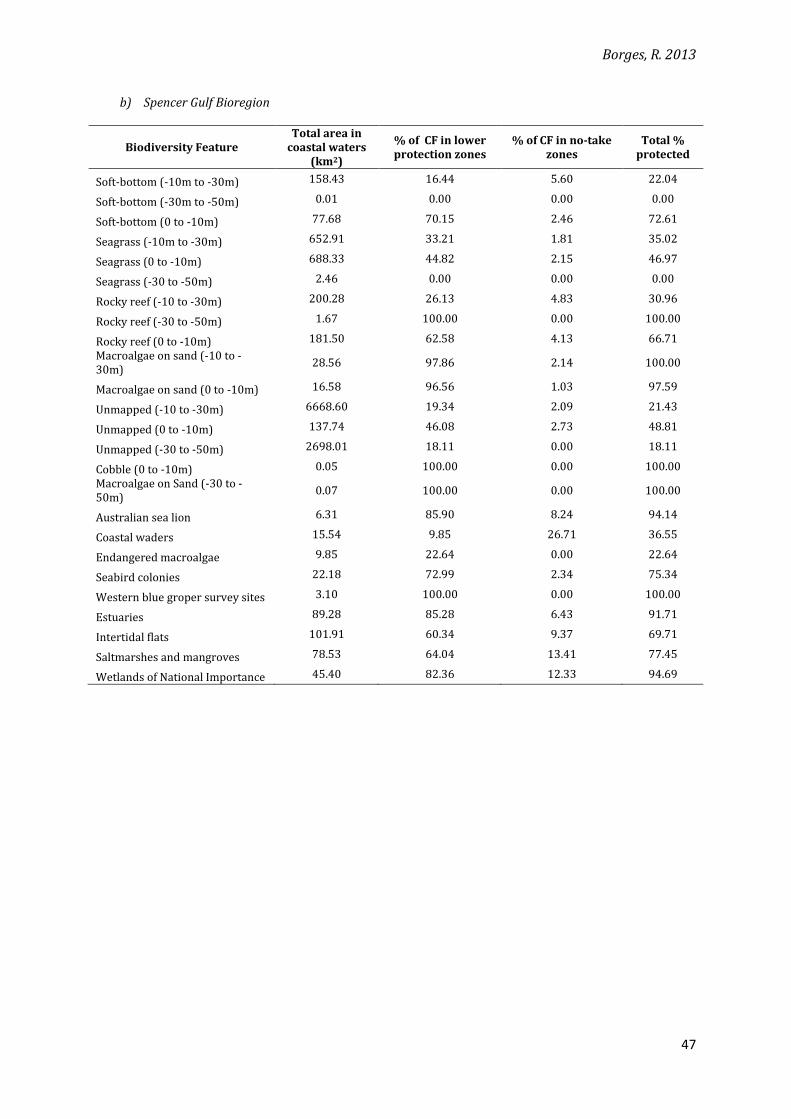
b) Spencer Gulf Bioregion
Biodiversity Feature Total area in
coastal waters (km2)
% of CF in lower protection zones
% of CF in no-take zones
Total % protected
Soft-bottom (-10m to -30m) 158.43 16.44 5.60 22.04
Soft-bottom (-30m to -50m) 0.01 0.00 0.00 0.00
Soft-bottom (0 to -10m) 77.68 70.15 2.46 72.61
Seagrass (-10m to -30m) 652.91 33.21 1.81 35.02
Seagrass (0 to -10m) 688.33 44.82 2.15 46.97
Seagrass (-30 to -50m) 2.46 0.00 0.00 0.00
Rocky reef (-10 to -30m) 200.28 26.13 4.83 30.96
Rocky reef (-30 to -50m) 1.67 100.00 0.00 100.00
Rocky reef (0 to -10m) 181.50 62.58 4.13 66.71
Macroalgae on sand (-10 to -30m)
28.56 97.86 2.14 100.00
Macroalgae on sand (0 to -10m) 16.58 96.56 1.03 97.59
Unmapped (-10 to -30m) 6668.60 19.34 2.09 21.43
Unmapped (0 to -10m) 137.74 46.08 2.73 48.81
Unmapped (-30 to -50m) 2698.01 18.11 0.00 18.11
Cobble (0 to -10m) 0.05 100.00 0.00 100.00
Macroalgae on Sand (-30 to -50m)
0.07 100.00 0.00 100.00
Australian sea lion 6.31 85.90 8.24 94.14
Coastal waders 15.54 9.85 26.71 36.55
Endangered macroalgae 9.85 22.64 0.00 22.64
Seabird colonies 22.18 72.99 2.34 75.34
Western blue groper survey sites 3.10 100.00 0.00 100.00
Estuaries 89.28 85.28 6.43 91.71
Intertidal flats 101.91 60.34 9.37 69.71
Saltmarshes and mangroves 78.53 64.04 13.41 77.45
Wetlands of National Importance 45.40 82.36 12.33 94.69
Borges, R. 2013
48
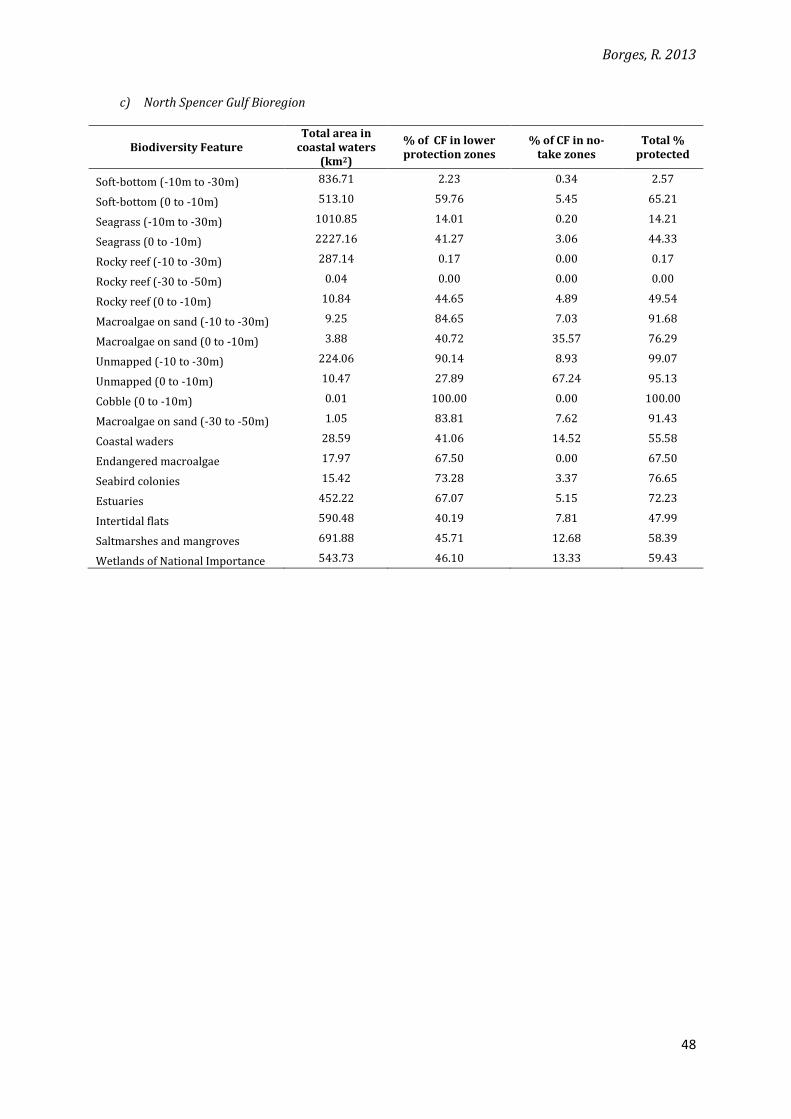
c) North Spencer Gulf Bioregion
Biodiversity Feature Total area in
coastal waters (km2)
% of CF in lower protection zones
% of CF in no-take zones
Total % protected
Soft-bottom (-10m to -30m) 836.71 2.23 0.34 2.57
Soft-bottom (0 to -10m) 513.10 59.76 5.45 65.21
Seagrass (-10m to -30m) 1010.85 14.01 0.20 14.21
Seagrass (0 to -10m) 2227.16 41.27 3.06 44.33
Rocky reef (-10 to -30m) 287.14 0.17 0.00 0.17
Rocky reef (-30 to -50m) 0.04 0.00 0.00 0.00
Rocky reef (0 to -10m) 10.84 44.65 4.89 49.54
Macroalgae on sand (-10 to -30m) 9.25 84.65 7.03 91.68
Macroalgae on sand (0 to -10m) 3.88 40.72 35.57 76.29
Unmapped (-10 to -30m) 224.06 90.14 8.93 99.07
Unmapped (0 to -10m) 10.47 27.89 67.24 95.13
Cobble (0 to -10m) 0.01 100.00 0.00 100.00
Macroalgae on sand (-30 to -50m) 1.05 83.81 7.62 91.43
Coastal waders 28.59 41.06 14.52 55.58
Endangered macroalgae 17.97 67.50 0.00 67.50
Seabird colonies 15.42 73.28 3.37 76.65
Estuaries 452.22 67.07 5.15 72.23
Intertidal flats 590.48 40.19 7.81 47.99
Saltmarshes and mangroves 691.88 45.71 12.68 58.39
Wetlands of National Importance 543.73 46.10 13.33 59.43