are protected areas truly protected? the impact of road traffic on vertebrate fauna
TRANSCRIPT

ORI GIN AL PA PER
Are protected areas truly protected? The impact of roadtraffic on vertebrate fauna
Nuria Garriga • Xavier Santos • Albert Montori • Alex Richter-Boix •
Marc Franch • Gustavo A. Llorente
Received: 9 January 2012 / Accepted: 6 July 2012 / Published online: 25 July 2012� Springer Science+Business Media B.V. 2012
Abstract The extension of road networks is considered one of the major factors affecting
fauna survival. Roadkill has been documented widely and affects all taxonomic groups.
Although roadkill is associated mainly with traffic density, some life-history traits of
species and the area surrounding roads are expected to modify number of roadkills both
taxonomically and geographically. Here we studied the number of roadkills of vertebrates
in an extensive region in the northeastern Iberian Peninsula. We surveyed 820 km of 41
roads in two different seasons (spring and autumn), that differ in traffic intensity. In
addition, we covered zones with distinct climatic characteristics and levels of protection of
the surrounding habitats. Amphibians showed the highest number of roadkills whereas
reptiles, birds and mammals had similar rates. General Linear Model tests showed no
differences in roadkills by climatic region; however, differences in number of roadkills
were linked to protection status, with the highest number of casualties in highly protected
areas. Redundancy Analysis demonstrated that the number of amphibians and reptiles
killed was associated with roads in highly protected areas whereas that of mammals and
birds was linked to unprotected areas. Protected areas often receive many visitors, which in
turn may increase wildlife casualties as a result of greater traffic density. We recommend
that correction measures be taken to reduce the high number of vertebrate fauna killed
along roads that cross protected areas.
Electronic supplementary material The online version of this article (doi:10.1007/s10531-012-0332-0)contains supplementary material, which is available to authorized users.
N. Garriga (&) � X. Santos � A. Montori � M. Franch � G. A. LlorenteDep. Biologia Animal, Universitat de Barcelona, Avgda. Diagonal 643, 08028 Barcelona, Spaine-mail: [email protected]
X. SantosCentro de Investigacao em Biodiversidade e Recursos Geneticos (CIBIO), Universidade do Porto,Campus Agrario de Vairao, 4485-661 Vairao, Portugal
A. Richter-BoixDepartment of Population Biology and Conserv Biol, Uppsala Universitet, Norbyvagen 18 D,752 36 Uppsala, Sweden
123
Biodivers Conserv (2012) 21:2761–2774DOI 10.1007/s10531-012-0332-0

Keywords Roadkills � Protected area � Habitat fragmentation � Road ecology �Vertebrates
Introduction
In recent decades, anthropogenic disturbance of wildlife habitats and biota has increased
because of human population growth and human–environment interactions (Wittmeyer
et al. 2009). The extension of road networks is one of the major disturbances for fauna
(Jaarsma et al. 2006). Road construction and exploitation entails habitat destruction,
population fragmentation (Mader 1984), and consequently vertebrate mortality (Forman
and Alexander 1998; Trombulak and Frissell 2000; Lesbarreres et al. 2003; Malo et al.
2004). Demographic decline in fauna located near roads has occurred (Jones 2000;
Mumme et al. 2000; Gibbs and Shriver 2005; Row et al. 2007), even for common species
that have, a priori, large enough populations to sustain high road mortality (Forman and
Alexander 1998).
The incidence of animals killed on roads is influenced by extrinsic (e.g. habitat near
roads, climate and local weather conditions, and transportation characteristics) and
intrinsic (e.g. species life-history traits) factors (Ashley and Robinson 1996; Clevenger
et al. 2003; Puky 2005), among others. Trombulak and Frissell (2000) reported that traffic
volume is the greatest contributor to roadkills, whereas Hels and Buchwald (2001)
observed that amphibian’s vulnerability to road mortality were mainly determined by the
velocity of species, their diurnal activity patterns, as well as the diurnal traffic activity.
Despite evidence that road mortality has no geographical or taxonomical restrictions
(Taylor and Goldingay 2004), it does not affect all taxonomic groups in the same way.
Amphibians and reptiles are the most affected (Ashley and Robinson 1996; Forman and
Alexander 1998; Smith and Dodd 2003; Glista et al. 2008), and the decline of their
populations has been attributed to road mortality worldwide (Fahrig et al. 1995; Gibbs and
Shriver 2002, 2005; Marchand and Litvaitis 2004; Steen and Gibbs 2004). Most amphibian
roadkills occur during individual adult migration to breeding ponds (van Gelder 1973;
Cooke, 1995; Glista et al. 2008). Such migration is strongly related to rainfall pattern and
annual pond flooding (Salvador and Carrascal 1990; Jakob et al. 2003). With regard to
reptiles, their activity is more related to temperature oscillations (Adolph and Porter 1993)
and their use of the road surface as a thermoregulatory source (Bernardino and Dalrymple
1992; Rudolph et al. 1999). Roadkill also occurs in mammals and birds, which use roads as
dispersal corridors (Huey 1941; Getz et al. 1978) or food sources (Dhindsa et al. 1988;
Pinowski 2005).
Road traffic is also detrimental to wildlife in protected areas (Bernardino and Dalrymple
1992; Drews 1995; Clevenger et al. 2003; Ramp et al. 2006), and is a major concern of US
National Park Service managers (Ament et al. 2008). Protected areas are commonly fre-
quented by tourists and local residents, and therefore subjected to regular traffic. Local
fauna is thus endangered by roads that cross natural areas (Hartmann et al. 2011). Ber-
nardino and Dalrymple (1992) reported an example in the Everglades National Park, where
the seasonal activity of the snake community coincides with the periods of greatest tour-
ism. This finding suggests that wildlife populations in National Parks are not necessarily
more protected than those inhabiting areas outside the boundaries of these areas (Newmark
1995; Ament et al. 2008). When a particular site is protected because of its wildlife
diversity and quality, it would be reasonable to assume that road traffic will decrease
wildlife unless appropriate measures are taken to minimize this impact.
2762 Biodivers Conserv (2012) 21:2761–2774
123

The following factors may simultaneously affect road mortality rates: protection status
of the area, number of species present, traffic intensity, and the climate and season sur-
veyed. Most studies addressing road mortality have designed surveys in small local
regions, comprising a few lineal kilometers (e.g. van Gelder 1973; Vijayakumar et al.
2001; Smith and Dodd 2003; Langen et al. 2007, 2009; Elzanowski et al. 2009), but over
extensive periods (Cooke 1995; Clevenger et al. 2003) or intensively surveyed (Ashley and
Robinson 1996; Clevenger et al. 2001b; Glista et al. 2008).
Nevertheless, comparison of the relevance of these factors calls for the design of
stratified surveys on a road network within an extensive region. European countries are
suitable candidates for this purpose as not only do they have dense road networks but also
many protected areas. This is the case of Catalonia (northeastern Spain), where
1,244,876 ha of protected area (39.6 % of the total extension) coexist with more than
12,000 km of a dense road network. We quantified vertebrate number of roadkills in
Catalonia and examined the most significant factors (extrinsic and intrinsic) that explain
this cause of death. Here we sought to determine some of the factors that influence roadkill
occurrence. For this purpose we addressed the following questions: (1) Are there more
roadkills in spring (breeding season) than in autumn (dispersion season)?; (2) Is there any
difference, in terms of number of roadkills, between vertebrate groups?; (3) Does clima-
tology affect the number of casualties by, for example, increasing amphibian roadkills in
the rainiest areas?; (4) Do protected areas register a higher incidence of roadkills?
Materials and methods
Study area
Catalonia has high climatic variability, with annual rainfall ranging from less than 400 to
1,000 mm, and temperatures showing wide differences throughout the region, with a mean
August temperature lower than 16 �C in some localities, while in others it exceeds 26 �C.
Following the study by Martin-Vide (1992), the study area was organized in four main
equally represented climatic regions on the basis of annual rainfall and temperature: Py-
renean, Continental Dry, Humid, and Littoral (Fig. 1a). The Pyrenean region is moun-
tainous with elevations above 1,000 meters. It is characterized by cold winters and mild
summers and high rainfall throughout the year, with the maximum in the summer and the
minimum in the winter. The Continental Dry region is characterized by cold winters and
hot summers and low rainfall, which is concentrated mainly in spring and autumn. The
Humid region has mild winters and hot summers. Although it has lower rainfall than the
Pyrenean region, the Humid region has a high rainfall pattern, which is concentrated
mainly in spring and autumn, with the maximum falling in autumn and the minimum in
winter. Finally, the Littoral region is a little drier and warmer than the Humid region, but
with small thermal fluctuation (Martin-Vide 1992). We classified roads in these climatic
regions on the basis of the predominant climate each road passes through.
Given the geographic location of the study area, fauna from several biogeographic
origins, namely Centro European, Ibero–Maghrebian and circum-Mediterranean con-
verge, thus implying high species richness (Carretero et al. 1999). Excluding fish, the
study area includes 14 amphibians, 33 reptiles, 96 mammals and 395 bird species. The
list of vertebrate species present in the 10 9 10 km UTM squares where roads are located
was obtained from the published atlases of the region (Llorente et al. 1995; Pleguezuelos
et al. 2002; Estrada et al. 2004; Palomo et al. 2007; Herrando et al. 2011). Moreover, in
Biodivers Conserv (2012) 21:2761–2774 2763
123
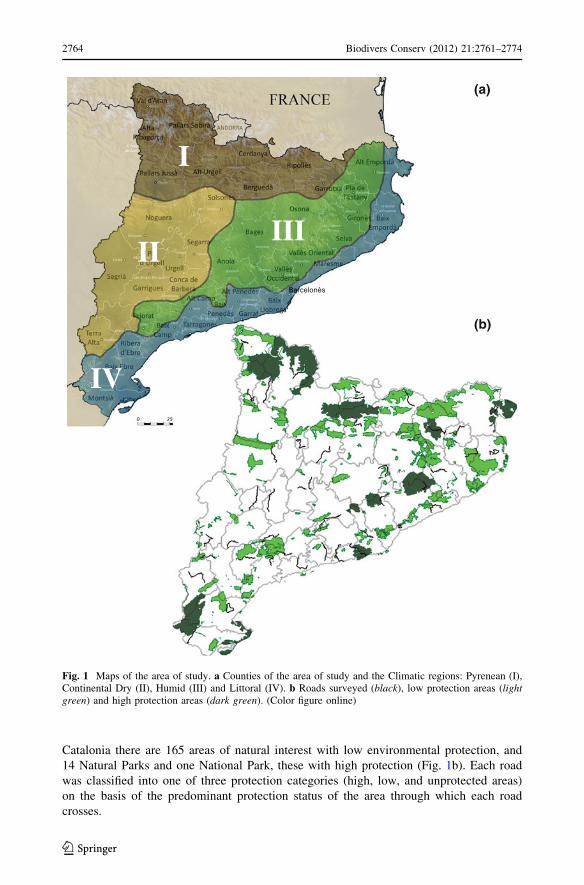
Catalonia there are 165 areas of natural interest with low environmental protection, and
14 Natural Parks and one National Park, these with high protection (Fig. 1b). Each road
was classified into one of three protection categories (high, low, and unprotected areas)
on the basis of the predominant protection status of the area through which each road
crosses.
IV
II
I
III
(a)
(b)
Fig. 1 Maps of the area of study. a Counties of the area of study and the Climatic regions: Pyrenean (I),Continental Dry (II), Humid (III) and Littoral (IV). b Roads surveyed (black), low protection areas (lightgreen) and high protection areas (dark green). (Color figure online)
2764 Biodivers Conserv (2012) 21:2761–2774
123

Road network and sampling
Catalonia has an extension about 31,895 km2, organized in 41 counties, and a road network
of 12,068 km, with 10.1 % of highways and 89.9 % of secondary roads (Generalitat de
Catalunya 2003a). To ensure uniform sampling in the study area, roadkills were surveyed
in spring and autumn 2002 in the 41 counties, thus covering the four climatic regions and
protected and unprotected areas. In each county we randomly selected a 20-km stretch of
secondary road. These were chosen because they are uniformly distributed throughout the
study area and are mainly two-way roads, with moderate speed traffic and without a central
reservation or lateral fences. Surveys were made by observers in a car traveling at a
maximum speed of 20 km/h as slow speed is required to detect small animals such as
amphibians (Ashley and Robinson 1996; Smith and Dodd 2003; Glista et al. 2008). Even
so, there is a misdetection of carcasses, taking into account that the missing of carcasses in
by foot surveys was estimated in a 7–67 % of the total number of road victims (Hels and
Buchwald 2001). Nevertheless, as the methodology used was the same in all the roads, the
results of all the surveys are comparable. All roads were surveyed by volunteer staff on
warm sunny days (air temperature approximately between 20 and 25 �C) and during the
same days. In the Pyrenean counties, where faunal activity begins later because of low
temperatures, spring surveys were delayed by 15 days. Each road was surveyed three times
in spring (from 15 April to 20 May 2002) and three times in autumn (from 10 October to 15
November 2002). Consequently, we conducted 246 surveys covering a total of 4,920 km.
To avoid replication by counting a carcass twice, roads were surveyed fortnightly, and
animal carcasses were removed from the road once they had been registered. The
extraction of the carcasses did not bias the results by reducing the possible number of
scavengers killed, since the interval between surveys was enough to recover the dynamics
of the road casualties. Roadkills were classified into four taxonomic groups (amphibians,
reptiles, birds and mammals) and identified to species level whenever possible. The geo-
graphic location of carcasses on the road was recorded with an accuracy of 100 m.
Statistical procedures
We performed a General Linear Model (GLM) analysis to check for differences in the
number of potential species killed on each road between the four climatic regions and
levels of protection considered.
We then performed different GLMs to test for differences in the number of roadkills
(dependent variable), using the taxonomy of specimens, season, climatic region and pro-
tection status, as explanatory variables (Table 1). The model that best fitted the data also
included the interactions ‘‘taxonomy 9 season’’, ‘‘taxonomy 9 protection’’ and ‘‘season 9
protection’’. Given that traffic intensity may influence the number of roadkills, we included
IMD (Daily Mean vehicle Intensity, data from Generalitat de Catalunya 2003a) in the
GLM analysis as a covariate, nevertheless, this covariant was not informative and finally
was not included in the chosen model.
The association between number of roadkills of particular species and explanatory
variables (i.e. climatic region and protection status) was checked by Redundancy Analysis
(RDA) with the CANOCO 4.0 software (ter Braak and Smilauer 1998). RDA is a con-
strained linear ordination method in which the species data are defined as a linear com-
bination of the explanatory variables (ter Braak and Smilauer 1998). The species data
corresponded to the number of roadkills registered on each road. Species with less than 5
specimens and specimens without species attribution were excluded from the analysis.
Biodivers Conserv (2012) 21:2761–2774 2765
123
Values were squareroot-transformed to reduce the over-representation of some amphibian
species that showed high number of roadkills on single roads, and the IMD was used as a
covariate. The explanatory power of the first ordination axis and all axes together were
tested in a Monte Carlo permutation test (9,999 unrestricted permutations).
Results
The GLM analysis showed that the number of species present in the UTM squares of the
roads surveyed did not differ either by climatic region (F3,35 = 0.840, P = 0.481) or
protection status (F2,35 = 0.959, P = 0.393). In other words, the number of potential
species vulnerable to roadkill did not differ between areas.
We registered a total of 2,013 roadkills (Table 2): 853 in spring and 1,160 in autumn.
Roadkills were composed of 267 mammals (13.3 %), 253 birds (12.6 %), 245 reptiles
(12.2 %) and 1,248 amphibians (62.0 %). During the surveys we identified 11 species of
amphibians (78.6 % of all amphibian species present in the area), 17 reptile (51.5 %), 18
mammal (18.8 %) and 39 bird (9.9 %) species. The species with the highest number of
roadkills were amphibians: Salamandra salamandra with 432 roadkills, Bufo bufo with
360, and with 305. The snake Malpolon monspessulanus, with 63 specimens, was the
reptile with the highest number of casualties. Passer domesticus and Rattus norvegicuswere the bird and mammal most killed respectively, both with 52 specimens.
The whole GLM model was highly significant (F20,307 = 9.641; P \ 0.001; Table 3).
The GLM test showed differences in the number of roadkills by season (F1,307 = 8.913,
P = 0.003). The beta coefficient indicates a higher incidence in autumn than in spring
(Table 4). The number of roadkills by taxonomical group also showed significant differ-
ences (F3,307 = 45.270, P \ 0.0001), and Scheffe post hoc comparison detected differ-
ences between amphibians and the rest of the groups (P \ 0.0001). As the beta coefficients
indicated, there were more amphibians than birds and more birds than the other two
Table 1 Explanatory variables used in roadkills GLM analysis
Variable Definition Type Categories/range
n
Taxon Taxonomical group to which each roadkill belongs Nominal Amphibian 1,248
Bird 253
Mammal 267
Reptile 245
Season Season when each roadkill was found Nominal Spring 853
Autumn 1,160
Region Predominant climate of the region where each roadpasses through
Nominal Pyrenean 212
Continental dry 391
Humid 847
Littoral 563
Protectionstatus
Category of protection of thearea where the road passes through
Nominal No protection 1,140
Low protection 276
High protection 597
IMD Daily Mean vehicle Intensity Scale 126–10,466
2766 Biodivers Conserv (2012) 21:2761–2774
123
taxonomical groups. The GLM did not show differences for climatic regions, and the
interaction season 9 taxon (Table 3). Nevertheless, there was a negative significant beta
for the interaction [amphibian vs. birds 9 spring vs. autumn], which indicated higher
number of amphibian roadkills in autumn and of birds in spring (Table 4). In contrast, the
protection status of the roads sampled showed significant differences as well as its inter-
action with taxonomic group and season. The Scheffe post hoc test for the protection status
indicated differences between the number of roadkills in roads classified as high protected
and the low and non protected. The post hoc tests for the interactions showed differences in
the number of casualties of amphibians in the high protection status areas and in autumn
also in the high protected areas, than the rest of combinations (Table 3). The sign of the
betas indicated that roads with high protection status showed more amphibian and bird
roadkills than low and non protected roads. Finally, in autumn there were more roadkills in
high protected than in low protected areas (Table 4).
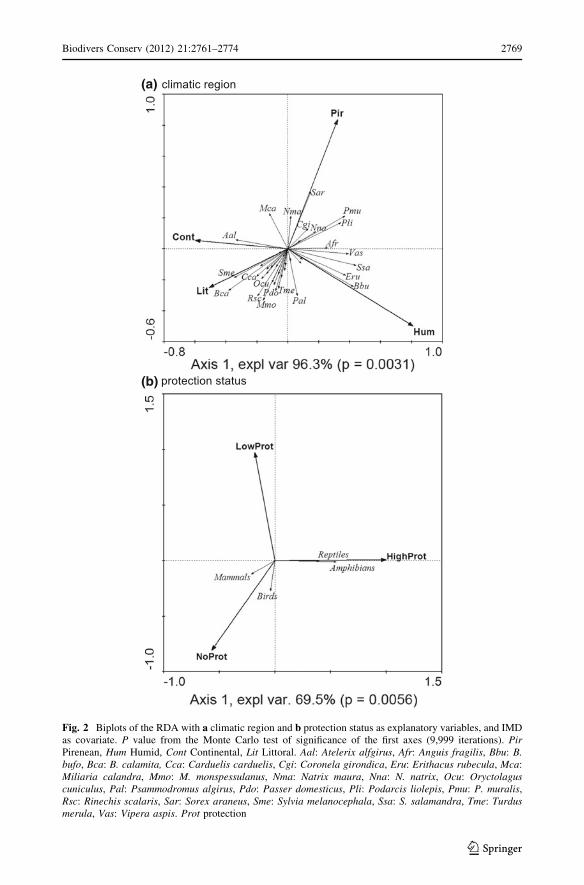
RDA showed a relation between roadkills registered and the two explanatory variables.
The test of significance of the RDA for the climatic region for all axes (F = 2.397,
P = 0.0023) and for the first axis (F = 4.708, P = 0.0056) were significant. The species
with the highest explained variance (% Expl), i.e., the amount of variation of each species
explained by climatic region, were B. bufo (24.05 %), B. calamita (21.98 %), S. salam-andra (20.72 %), Sylvia melanocephala (20.34 %) and Podarcis muralis (20.02 %)
(Fig. 2a). S. salamandra and B. bufo were more associated with Humid regions, while B.calamita and S. melanocephala were linked to Littoral areas. In contrast, P. muralis was
Table 2 Road mortality of the four taxonomic groups, both in spring and autumn
Taxonomic group Spring Autumn Total
n % n % N %
Amphibians 468 54.9 780 67.2 1,248 62.0
Birds 158 18.5 95 8.2 253 12.6
Mammals 100 11.7 167 14.4 267 13.3
Reptiles 127 14.9 118 10.2 245 12.2
Total 853 1,160 2,013
Table 3 GLM analyses of the number of roadkills for the different explanatory variables
Effect Adjusted R2 SS d.f. MS F P
Whole model 0.346 26,757.89 20 1,337.90 9.641 <0.001
Intercept 12,456.32 1 12,456.32 89.761 <0.001
Season 12,36.92 1 1,236.92 8.913 0.003
Region 243.76 3 81.25 0.586 0.625
Taxon 18,846.50 3 6,282.17 45.270 <0.001
Protection status 2,582.87 2 1,291.44 9.306 <0.001
Season 9 Taxon 1,003.91 3 334.64 2.414 0.067
Season 9 Protection status 1,455.41 2 727.71 5.244 0.006
Taxon 9 Protection status 11,145.97 6 1,857.66 13.386 <0.001
Error 42,602.93 307 138.77
In bold there are the significant effects
Biodivers Conserv (2012) 21:2761–2774 2767
123
associated with Pyrenean areas. The RDA for the protection status was also significant for
all axes (F = 2.429, P = 0.0159) and for the first one (F = 3.950, P = 0.0168).
Amphibians and reptiles were associated with high protected areas, whereas mammals and
birds were related to non-protected areas (Fig. 2b).
Discussion
Composition of roadkills
The present study shows taxonomic differences in roadkills, with herpetofauna, especially
amphibians, accounting for 74.2 % of all vertebrate roadkills found on the roads studied,
both in spring and autumn. Our results are consistent with other comparative roadkill
patterns detected in surveys performed by bicycle or walking, or even by car at slow speed
(\40 km/h), which reported higher road casualties of amphibians and reptiles than other
vertebrate groups (Ashley and Robinson 1996; Smith and Dodd 2003; Dodd et al. 2004;
Glista et al. 2008). Nevertheless, Glista and DeVault (2008) also performed surveys at slow
speed (\40 km/h) and detected mammals as the most common vertebrate taxon killed. In
this case the authors stated that the seasonal sampling contributed to a bias towards
detection of larger carcasses such as raccoons or opossums to the detriment of smaller
carcasses such as birds and amphibians. As in those studies, our surveys were performed by
observers in a car travelling at low speed (\20 km/h), to facilitate the detection of small
carcasses, even though there is always a misdetection (Hels and Buchwald 2001). Puky
(2005) reported that the sampling methodology can bias the detection of vertebrate groups
killed on roads because surveys performed by vehicles travelling at high speed ([40 km/h)
Table 4 Parameter estimates and b coefficients of the significant intra-effect comparisons
Intra-effect comparison Parameterestimate
St errparam
t P b St errb
Intercept 7.962 0.840 9.474 \0.001
Spring vs autumn 2.346 0.786 -2.986 0.003 -0.161 0.054
Amph vs birds 15.850 1.361 11.645 \0.001 0.771 0.066
Rept vs birds -4.740 1.361 -3.483 \0.001 -0.231 0.066
Mamm vs birds -5.738 1.361 -4.216 \0.001 -0.279 0.066
Non vs high prot -3.242 1.081 -2.998 0.003 -0.156 0.052
Low vs high prot -2.810 1.124 -2.499 0.013 -0.122 0.049
Spring vs autumn 9 Non vs highprot
2.439 0.932 2.616 0.009 0.139 0.053
Spring vs autumn 9 Amph vsbirds
-2.869 1.127 -2.546 0.011 -0.139 0.055
Amph vs birds 9 Non vs highprot
-12.127 1.615 -7.511 \0.001 -0.487 0.065
Amph vs birds 9 Low vs highprot
-7.398 1.785 -4.144 \0.001 -0.238 0.058
Mamm vs birds 9 Non vs highprot
5.243 1.615 3.248 0.002 0.211 0.065
St err param standard error of the parameter, St err b standard error of the b, Amph amphibians, Reptreptiles, Mamm mammals, Prot protection status
2768 Biodivers Conserv (2012) 21:2761–2774
123
(b)
(a)
Fig. 2 Biplots of the RDA with a climatic region and b protection status as explanatory variables, and IMDas covariate. P value from the Monte Carlo test of significance of the first axes (9,999 iterations). PirPirenean, Hum Humid, Cont Continental, Lit Littoral. Aal: Atelerix alfgirus, Afr: Anguis fragilis, Bbu: B.bufo, Bca: B. calamita, Cca: Carduelis carduelis, Cgi: Coronela girondica, Eru: Erithacus rubecula, Mca:Miliaria calandra, Mmo: M. monspessulanus, Nma: Natrix maura, Nna: N. natrix, Ocu: Oryctolaguscuniculus, Pal: Psammodromus algirus, Pdo: Passer domesticus, Pli: Podarcis liolepis, Pmu: P. muralis,Rsc: Rinechis scalaris, Sar: Sorex araneus, Sme: Sylvia melanocephala, Ssa: S. salamandra, Tme: Turdusmerula, Vas: Vipera aspis. Prot protection
Biodivers Conserv (2012) 21:2761–2774 2769
123

show a high incidence of mammals and birds (Slater 2002; Clevenger et al. 2003; Taylor
and Goldingay 2004). This disparity of results indicates that small animals may not be
detected at higher speed, and thus bias the casualties detected to mammals and birds, even
when more amphibians or reptiles are in fact killed by traffic.
Reptile and bird roadkills were more frequent in spring, when their activity is high
because of reproduction. This result is consistent with the observation that roadkills are
usually associated with major wildlife activity during the reproductive season and with
thermoregulatory behavior in ectotherms (Bernardino and Dalrymple 1992; Rudolph et al.
1999). However, this general statement does not fit amphibians and mammals. Amphibian
activity is regulated particularly by the rainfall regime. In the study area, their main
breeding season is in spring, with a second breeding period in autumn (Richter-Boix et al.
2006), mainly in the Littoral and Humid regions, as a result of high rainfall and mild
temperatures during this season. At the time when the present study was performed, the
rainfall registered in autumn was almost two-fold that in spring (Generalitat de Catalunya
2003b). This seasonal pattern was also observed in amphibian roadkills, as we observed
almost twice the number of casualties in autumn than in spring. The association between
rainfall and amphibian roadkills is dramatic in regions with an infrequent and unpredict-
able rainfall regime, such as the Mediterranean area (Carvalho and Mira 2011). Mammals
also showed their highest number of roadkills in autumn. Several rodents, such as
Apodemus sylvaticus (the third most frequently killed mammal), breed from spring to
autumn; therefore populations reach a peak density in September (Kikkawa 1964; Torre
et al. 2002), and also increase their locomotor activity (Flowerdew 2000), which is
reflected in increased number of roadkills.
The main victims on the roads covered by our surveys were three amphibian species
with high terrestrial mobility: S. salamandra, B. bufo and B. calamita. This result supports
previous studies showing that some activity patterns, such as high migratory activity (Bar-
David et al. 2007; Schmidt et al. 2007) and terrestrial dispersion (Moore 1954; Miaud et al.
2000), make species highly vulnerable to traffic. Furthermore, the two toad species show
fidelity to breeding ponds, a behavior that makes them highly susceptible to fragmentation,
especially when there are massive migrations during the breeding season (Sinsch 1989).
The site fidelity of B. bufo is very high, with 93 % of females and 96 % of males that
survived between years returning to the same breeding ponds (Reading et al. 1991). An
example of this fidelity was registered by Cooke (1995), who recorded high number of
roadkills of B. bufo on roads near a breeding point during 21 years. However, the three
species most killed could be more detected due to a higher durability on the asphalt than
other amphibians or vertebrate species, because of their size, tough skin or unpalatability.
Therefore, further studies to determine the durability of the species on the road and
evaluate the real number of victims of the roads are needed.
Explanatory variables and number of roadkills
RDA has demonstrated that several species and groups are linked to particular climatic or
protected areas. The species with the highest amount of variation explained by the climatic
region were S. salamandra, B. bufo and B. calamita, P. muralis and S. melanocephala. This
finding indicates that species roadkills are associated with the climates for which the
species show clear preferences; for example, S. salamandra and B. bufo are more asso-
ciated with Humid areas (Llorente et al. 1995), while B. calamita and S. melanocephala are
most commonly found in Littoral areas (Dıaz et al. 1996; Gasc et al. 1997), and P. muralisin Pyrenean regions (Llorente et al. 1995).
2770 Biodivers Conserv (2012) 21:2761–2774
123

We did not detect differences in the number of species found killed per road between
the four climatic regions. This result is consistent with the lack of differences in the
number of vertebrate species present in each climatic region (see Results). In contrast, we
demonstrate that the incidence of roadkills increases as the protection level increases. It
could be proposed that this relationship is due to a greater number of species in protected
areas; however, this was not the case, as GLM analyses of species presence did not differ
between protection status areas. The most plausible explanation for this higher number of
roadkills is that wildlife populations in protected areas have higher densities and a greater
number of animals cross or use roads than in unprotected areas (Rodrigues et al. 2004). Our
results therefore agree with previous studies performed in protected areas worldwide
(Bernardino and Dalrymple 1992; Drews 1995; Kline and Swann 1998; Mazerolle 2004;
Ament et al. 2008). Paradoxically, all these studies indicate that protected areas increase
the vulnerability of wildlife to road traffic.
Conclusions
The finding that protected areas registered the highest number of roadkills is relevant for
the future management of wildlife conservation. In fact, protection status may produce a
network of roads and tourism services and consequently an increase in the number of
visitors, which in turn can degrade natural resources (Manning 2001). Roads crossing
protected areas endanger local fauna (Clevenger et al. 2003; Hartmann et al. 2011),
especially when peaks in wildlife seasonal activity coincides with tourism high seasons
(Bernardino and Dalrymple 1992). This phenomenon has been reported in one of the Parks
included in the present study. In that park, the highest number of visitors occurred in
autumn (Diputacio de Barcelona 2011), coinciding with the period of greatest number of
roadkills detected in the present study. On the other hand, the IMD did not result infor-
mative, since a single car is enough to kill a lot of animals on the asphalt. This result
indicated that, in protected areas, probably is more important animals activity patterns than
traffic intensity. Nevertheless, it would be interesting to evaluate the relation between
roadkills and season activity of species along all the year. Accordingly, the roadkills in the
protected areas should be considered by park conservation managers. Given that
amphibians is the group with the highest number of roadkills, especially in the protected
areas, correction measures should be directed to this taxonomic group and areas. Due to
their dependence on water for breeding and fidelity to breeding sites, drift fences (Langton
1989) and changes in breeding sites (Schlupp and Podloucky 1994) have been proved as
effective and can be applied in protected area (Clevenger et al. 2001a; Aresco 2003; Dodd
et al. 2004). Moreover, the enforced speed limit for cars crossing protected areas could also
be an effective measure.
Our findings demonstrate that roadkills of herpetological fauna, especially amphibian
populations in protected areas, should not be underestimated. Assessing number of road-
kills for vertebrate groups and species may shed light on their specific vulnerability to road
networks (Clevenger et al. 2003; Steen et al. 2006). Information from studies of this nature
contributes to the design of specific measures to mitigate the impact of road traffic on
wildlife.
Acknowledgments We thank the volunteers P. X. Alborna, X. Bayer, J. Camprodon, E. Cipriano, J.Galbany, D. Guixe, P. J. Jimenez, M. Martin, J. Nicolau, M. Pascual, L. Perez, L. Plaza, S. Ramos, L. Solerand F. Sort for their help with the surveys, and the two anonymous reviewers for their comments. Thanks
Biodivers Conserv (2012) 21:2761–2774 2771
123

also go to A. Sorolla for logistic support. This research was funded by the Departament de Medi Ambient,Territori i Sostenibilitat of the Generalitat de Catalunya (FBG301492). Xavier Santos was supported by apost-doctoral grant (SFRH/BPD/73176/2010) from the Fundacao para a Ciencia e a Tecnologia (FCT,Portugal).
References
Adolph S, Porter WP (1993) Temperature, activity, and lizard life histories. Am Nat 142:273–295Ament R, Clevenger AP, Yu O, Hardy A (2008) An assessment of road impacts on wildlife populations in
U.S National Parks. Environ Manage 42:480–496Aresco MJ (2003) Highway mortality of turtles and other herpetofauna at lake Jackson, Florida, USA, and
the efficacy of a temporary fence/culvert system to reduce roadkills. In: Irwin CL, Garrett P,McDermott KP (eds) Proceedings of the international conference on ecology and transportation. TheCenter for Transportation and the Environment, Raleigh, pp 433–449
Ashley EP, Robinson JT (1996) Road mortality of amphibians, reptiles and other wildlife on the long pointcauseway, Lake Erie, Ontario. Can Field Nat 110:403–412
Bar-David S, Segev O, Peleg N, Hill N, Templeton AR, Schultz CB, Blaustein L (2007) Long-distancemovements by fire salamanders (Salamandra Infraimmaculata) and implications for habitat frag-mentation. Isr J Ecol Evol 53:143–159
Bernardino FS, Dalrymple GH (1992) Seasonal activity and road mortality of the snakes of the Pa-hay-okeewetlands of Everglades National Park, USA. Biol Conserv 62:71–75
Carretero MA, Llorente GA, Montori A, Santos X, Francesch J (1999) Observed vs. potential distributions:application to the herpetofauna of a region of the Mediterranean basin. Contrib Zoogeograph EcolEastern Med Region 1:221–228
Carvalho F, Mira A (2011) Comparing annual vertebrate road kills over two time periods, 9 years apart: acase study in Mediterranean farmland. Eur J Wildl Res 57:157–174
Clevenger AP, Chruszcz B, Gunson KE (2001a) Highway mitigation fencing reduces wildlife-vehiclecollisions. Wildl Soc Bull 29:646–653
Clevenger AP, McIvor M, McIvor D, Chruszscz B, Gunson KE (2001b) Tiger Salamander, Ambystomatigrinum, movements and mortality on the Trans-Canada highway in southwestern Alberta. Can FieldNat 115:199–204
Clevenger AP, Chruszscz B, Gunson KE (2003) Spatial patterns and factors influencing small vertebratefauna road kill aggregations. Biol Conserv 109:15–26
Cooke AS (1995) Road mortality of common toads (Bufo bufo) near a breeding site, 1974–1994. Amphib-Reptil 16:87–90
Dhindsa MS, Sandhu JS, Sandhu PS, Toor HS (1988) Roadside birds in Punjab (India): relation to mortalityfrom vehicles. Environ Conserv 15:303–310
Dıaz M, Asensio B, Tellerıa JL (1996) Aves Ibericas Vol I. No paseriformes. Reyero Ediciones, MadridDiputacio de Barcelona (2011) Parc Natural del Montseny. Reserva de la Biosfera. Memoria 2010. Direccio
de Comunicacio, BarcelonaDodd CK, Barichivich WJ, Smith LL (2004) Effectiveness of a barrier wall and culverts in reducing wildlife
mortality on a heavily traveled highway in Florida. Biol Conserv 118:619–631Drews C (1995) Road kills of animals by public traffic in Mikumi National Park, Tanzania, with notes on
baboon mortality. Afr J Ecol 33:89–100Elzanowski A, Ciesiołkiewicz J, Kaczor M, Radwanska J, Urban R (2009) Amphibian road mortality in
Europe: a meta–analysis with new data from Poland. Eur J Wildl Res 55:33–43Estrada J, Pedrocchi V, Brotons L, Herrando S (eds) (2004) Atles dels ocells nidificants de Catalunya
1999–2002. Institut Catala d’Ornitologia (ICO). Lynx Edicions, BarcelonaFahrig L, Pedlar JH, Pope SE, Taylor PD, Wegner JF (1995) Effect of road traffic on amphibian density.
Biol Conserv 73:177–182Flowerdew JR (2000) Wood mice – small granivores/insectivores with seasonally variable patterns. In:
Halle S, Stenseth NC (eds) Activity patterns in small mammals, Ecological studies, vol 141. Springer,Berlin, pp 177–189
Forman RTT, Alexander LE (1998) Roads and their major ecological effects. Annu Rev Ecol Syst29:207–231
Gasc JP, Cabela A, Crnobrnja-Isailovic J, Dolmen D, Grossenbacher K, Haffner P, Lescure J, Martens H,Martınez Rica JP, Maurin H, Oliveira ME, Sofianidou TS, Veith M, Zuiderwijk A (1997) Atlas of
2772 Biodivers Conserv (2012) 21:2761–2774
123

amphibians and reptiles in Europe. Collection Patrimoines Naturels, 29, Societas Europaea Herpeto-logica, Museum National d’Histoire Naturelle and Service du Petrimone Naturel, Paris
Generalitat de Catalunya (2003a) Anuari estadıstic 2003, Carreteres. http://www20.gencat.cat/portal/site/ptop. Accessed 21 Dec 2011
Generalitat de Catalunya (2003b) Anuari de dades meteorologiques 2003, Dades de la Xarxa d’EstacionsMeteorologiques Automatiques. http://www.meteo.cat/servmet/index.html. Accessed 21 Dec 2011
Getz LL, Cole FR, Gates DL (1978) Interstate roadside as dispersal routes for Microtus pennsylvanicus.J Mammal 59:208–212
Gibbs JP, Shriver WG (2002) Estimating the effects of road mortality on turtle populations. Conserv Biol16:1647–1652
Gibbs JP, Shriver WG (2005) Can road mortality limit populations of pool–breeding amphibians? Wetl EcolManag 13:281–289
Glista DJ, DeVault TL (2008) Road mortality of terrestrial vertebrates in Indiana. Proc Indiana Acad Sci117:55–62
Glista DJ, DeVault TL, DeWoody JA (2008) Vertebrate road mortality predominantly impacts amphibians.Herpetol Conserv Biol 3:77–87
Hartmann PA, Hartmann MT, Martins M (2011) Snake road mortality in a protected area in the Atlanticforest of southeastern Brazil. S A J Herpetol 6:35–41
Hels T, Buchwald E (2001) The effect of road kills on amphibian population. Biol Conserv 99:331–340Herrando S, Brotons L, Estrada J, Guallar S, Anton M (2011) Atles dels ocells de Catalunya a l’hivern
2006–2009. Institut Catala d’Ornitologia (ICO). Lynx Edicions, BarcelonaHuey LM (1941) Mammlian invasion via the highway. J Mammal 22:383–385Jaarsma CF, van Langevelde F, Botma H (2006) Flattened fauna and mitigation: traffic victims related to
road, traffic, vehicle, and species characteristics. Transport Res D-Tr E 11:264–276Jakob C, Poizat G, Veith M, Seitz A, Crivelli AJ (2003) Breeding phenology and larval distribution of
amphibians in a Mediterranean pond network with unpredictable hydrology. Hydrobiologia 499:51–61Jones ME (2000) Road upgrade, road mortality and remedial measures: impacts on a population of eastern
quolls and Tasmanian devils. Wildlife Res 27:289–296Kikkawa J (1964) Movement, activity and distribution of the small rodents Clethrionomys glareolus and
Apodemus sylvaticus in woodland. J Anim Ecol 33:259–299Kline NC, Swann DE (1998) Quantifying wildlife road mortality in Saguaro National Park. In: Evink G,
Garret P, Zeigler D, Berry J (eds) Proceedings of the international conference on wildlife ecology andtransportation. Florida Department of Transportation, Tallahassee, pp 23–31
Langen TA, Machniak A, Crowe EK, Mangan C, Marker DF, Liddle N, Roden B (2007) Methodologies forsurveying herpetofauna mortality on rural highways. J Wildl Manage 71:1361–1368
Langen TA, Ogden KM, Schwarting LL (2009) Predicting hot spots of herpetofauna road mortality alonghighway networks. J Wildl Manage 73:104–114
Langton TES (ed) (1989) Amphibians and roads. ACO Polymer Products, SheffordLesbarreres D, Pagano A, Lode T (2003) Inbreeding and road effect zone in a Ranidae: the case of agile
frog, Rana dalmatina Bonaparte, 1840. C R Biol 326:S68–S72Llorente GA, Montori A, Santos X, Carretero MA (1995) Atlas dels amfibis i reptils de Catalunya i Andorra.
Editorial El Brau, FigueresMader HJ (1984) Animal habitat isolation by roads and agricultural fields. Biol Conserv 29:81–96Malo JE, Suarez F, Dıez A (2004) Can we mitigate animal–vehicle accidents using predictive models?
J Appl Ecol 41:701–710Manning R (2001) Programs that work. Visitor experience and resource protection: a framework for
managing the carrying capacity of National Parks. J Park Recreat Adm 19:93–108Marchand MN, Litvaitis JA (2004) Effects of habitat features and landscape composition on the population
structure of a common aquatic turtle in a region undergoing rapid development. Conserv Biol18:758–767
Martin-Vide J (1992) El clima Geografıa general dels Paısos Catalans. Enciclopedia Catalana, BarcelonaMazerolle MJ (2004) Amphibian road mortality in response to nightly variations in traffic intensity. Her-
petologica 60:45–53Miaud C, Sanuy D, Avrillier J (2000) Terrestrial movements of the natterjack toad Bufo calamita
(Amphibia, Anura) in a semi-arid, agricultural landscape. Amphib-Reptil 21:357–369Moore HJ (1954) Some observations on the migration of the toad (Bufo bufo). Br J Herpetol 1:194–224Mumme RL, Schoech SJ, Woolfenden GE, Fitzpatrick JW (2000) Life and death in the fast lane: demo-
graphic consequences of road mortality in the Florida Scrub-Jay. Conserv Biol 14:501–512Newmark WD (1995) Extinction of mammal populations in western North American National Parks.
Conserv Biol 9:512–526
Biodivers Conserv (2012) 21:2761–2774 2773
123

Palomo LJ, Gisbert J, Blanco JC (2007) Atlas y libro rojo de los mamıferos terrestres de Espana. DGB-SECEM-SECEMU, Madrid
Pinowski J (2005) Roadkills of vertebrates in Venezuela. Rev Bras Zool 22:191–196Pleguezuelos JM, Marquez R, Lizana M (2002) Atlas y libro rojo de los anfibios y reptiles de Espana.
Direccion General de Conservacion de la Naturaleza-Asociacion Herpetologica Espanola, MadridPuky M (2005) Amphibian road kills: a global perspective. In: Irwin CL, Garrett P, McDermott KP (eds)
Proceedings of the 2005 international conference on ecology and transportation. Center of Transpor-tation and the Environment, Raleigh, pp 325–328
Ramp D, Wilson VK, Croft DB (2006) Assessing the impact of roads in peri-urban reserves: Road-basedfatalities and road usage by wildlife in the Royal National Park, New South Wales, Australia. BiolConserv 129:348–359
Reading CJ, Loman J, Madsen T (1991) Breeding pond fidelity in common toads, Bufo bufo. J Zool225:201–211
Richter-Boix A, Llorente GA, Montori A (2006) Breeding phenology of an amphibian community in aMediterranean area. Amphib-Reptil 27:549–559
Rodrigues ASL, Andelman SJ, Bakarr MI, Boitani L, Brooks TM, Cowling RM, Fishpool LDC, da FonsecaGAB, Gaston KJ, Hoffmann M, Long JS, Marquet PA, Pilgrim JD, Pressey RL, Schipper J, SechrestW, Stuart SN, Underhill LG, Waller RW, Watts MEJ, Yan X (2004) Effectiveness of the globalprotected area network in representing species diversity. Nature 428:640–643
Row JR, Blouin-Demers G, Weatherhead PJ (2007) Demographic effects of road mortality in black rat-snakes (Elaphe obsolete). Biol Conserv 137:117–124
Rudolph C, Burgdorf S, Conner R, Schaefer R (1999) Preliminary evaluation of the impact of roads andassociated vehicular traffic on snake populations in eastern Texas. In: Evink GL, Garrett P, Zeigler D(eds) Proceedings of the third international conference on wildlife ecology and transportation. FloridaDepartment of Transportation, Tallahassee, pp 129–136
Salvador A, Carrascal LM (1990) Reproductive phenology and temporal patterns of mate access in Med-iterranean anurans. J Herpetol 24:438–441
Schlupp I, Podloucky R (1994) Changes in breeding site fidelity: a combined study of conservation andbehaviour in the common toad Bufo bufo. Biol Conserv 69:285–291
Schmidt BR, Schaub M, Steinfartz S (2007) Apparent survival of the salamander Salamandra salamandra islow because of high migratory activity. Front Zool 4:19
Sinsch U (1989) Migratory behaviour of the common toad Bufo bufo and the natterjack toad Bufo calamita.In: Langton TES (ed) Amphibians and roads, proceedings of the toad tunnel conference. ACO PolymerProducts, Bedfordshire, pp 113–125
Slater FM (2002) An assessment of wildlife road casualties—the potential discrepancy between numberscounted and numbers killed. Web Ecol 3:33–42
Smith LL, Dodd CK Jr (2003) Wildlife mortality on US highway 441 across Paynes prairie, AlachuaCounty, Florida. Florida Acad Sci 66:128–140
Steen DA, Gibbs JP (2004) Effects of roads on the structure of freshwater turtle populations. Conserv Biol18:1143–1148
Steen DA, Aresco MJ, Beilke SG, Compton BW, Condon EP, Dodd CK Jr, Forrester H, Gibbons JW,Greene JL, Johnson G, Langen TA, Oldham MJ, Oxier DN, Saumure RA, Schueler FW, Sleeman JM,Smith LL, Tucker JK, Gibbs JP (2006) Relative vulnerability of female turtles to road mortality. AnimConserv 9:269–273
Taylor BD, Goldingay RL (2004) Wildlife road kills on three major roads in north-eastern New SouthWales. Wildlife Res 31:83–91
ter Braak CJF, Smilauer P (1998) CANOCO reference manual and user’s guide to Canoco for Windows—software for canonical community ordination (version 4). Microcomputer Power, Ithaca
Torre I, Arrizabalaga A, Dıaz M (2002) Raton de campo (Apodemus sylvaticus Linnaeus, 1758). Galemys14:1–26
Trombulak SC, Frissell CA (2000) Review of ecological effects of roads on terrestrial and aquatic com-munities. Conserv Biol 14:18–30
van Gelder JJ (1973) A quantitative approach to the mortality resulting from traffic in a population of Bufobufo L. Oecologia 13:93–95
Vijayakumar SP, Vasudevan K, Ishwar NM (2001) Herpetofaunal mortality on road in the Anamalai Hills,Southern Western Ghats. Hamadryad 26:265–272
Wittmeyer G, Elsen P, Bean WT, Coleman A, Burton O, Brashares JS (2009) Accelerated human populationgrowth at protected area edges. Science 321:123–126
2774 Biodivers Conserv (2012) 21:2761–2774
123