communities of arbuscular mycorrhizal fungi in grassland: seasonal variability and effects of...
TRANSCRIPT

Folia Geobotanica 36." 243-263, 2001
COMMUNITIES OF ARBUSCULAR MYCORRHIZAL FUNGI IN GRASSLAND: SEASONAL VARIABILITY AND EFFECTS OF ENVIRONMENT AND HOST PLANTS
Petr ~milauer
Faculty of Biological Sciences, University of South Bohemia, Brani~ovsk~ 31, CZ-370 05 ~esk~ Bud~jovice, Czech Republic," e-mail [email protected]
Keywords: Achillea millefolium, Fertilization, Fungal morphotypes, Mowing, Poa pratensis, Plantago lanceolata, Seasonal dynamics
Abstract: The seasonal dynamics of a community of endomycorrhizal fungal morphotypes in the roots of three grassland species (Achillea millefolium, Poa angustifolia, Plantago lanceolata) was evaluated, together with the effects of experimental treatment (mowing and phosphorus application) and the host plant properties.
Strong seasonal variability was found in the ftmgal community, where clear seasonal patterns can be distinguished for several fungal morphotypes. The sampling date explained 20 to 30% of the total compositional variability for all three host species. The Plantago roots host the highest number of arbuscular mycorrhizal (AM) fungal populations. There are two co-dominant fungal morphotypes in Achillea roots (assigned to the genera Scutellospora and Glomus) and only one strongly dominant morphotype (assigned to Glomus), in the Poa roots. All three host species have a comparable pattern of richness of AM morphotypes with a single peak in the summer, possibly aligned with the flowering time of the host.
This study found only a limited effect of mowing (on the abundance ofScutellospora-like morphotypes in the Poa roots) and no effect of fertilization with phosphorus. The effect of developmental age of individual host plants on the composition of the endomycorrhizal community was found in Plantago, but not in Achillea or Poa.
INTRODUCTION
Arbuscular mycorrhizal (AM) fungi enter into endomycorrhizal symbiosis with almost 80% of the higher plant species (SMITH & READ 1997). The AM fungi were shown to provide plants with a variety of benefits beyond the acquisition of soil phosphorus, including the suppression of fungal pathogens (NEWSHAM et al. 1994), the change of attractiveness for foliar feeders (GANGE & WEST 1994) and the improvement of water supply (ALLEN 1982). Many ecological studies targeted the influence of AM symbiosis on the competitive ability of host plants (e.g. FITTER 1977, HARTNETT et al. 1993, MOORA & ZOBEL 1996), which together with the other types of interaction between the host and the AM fungi might have a profound effect upon the structure and diversity of plant communities (GRIME et al. 1987, FRANCIS & READ 1994, VAN DER HEIJDEN et al. 1998a, HARTNETT & WILSON 1999).
For a long time, the AM infection was (and still continues to be) studied as a black box, where the total infection level is used as the primary descriptor to assess the importance of mycorrhizal symbiosis for the host plants. Yet, it is now known that AM fungi form quite diverse communities not only at the level o f the whole plant community, but also within individual root systems or their parts (CLAPP et al. 1995, BEVER et al. 1996, MERRYWEATHER &

244 Smilauer P.
FITTER 1998a). From this observation alone and from the other, more direct experiments (e.g. SMITH et al. 2000), we can conclude that the individual AM fungal symbionts differ not only in the extent of their effectiveness, but also in the type of benefit they are providing (NEWSHAM et al. 1995). The variability of effects of different AM fungi on host species and their assemblages have already been demonstrated (VAN DER HEIJDEN et al. 1998b). Some of the AM fungal populations can even act as parasites of their plant hosts (JOHNSON et al. 1997). Knowledge of the ecological roles of individual fungal populations, however, cannot be obtained from comparative greenhouse experiments, performed in very un-natural conditions and with cultural plants. We need to study the response of the communities of AM fungi to the environmental factors and their spatial and temporal variability, including variability across the different host plants, using more natural conditions (see e.g. JOHNSON et al. 1997).
Because the taxonomy of AM fungi is based primarily on the morphology of their spores, which is now often supplemented with molecular biological data, the study of AM fungal communities and of their diversity is also based primarily on the retrieval of spores (GEMMA et al. 1989, JOHNSON et al. 1991, BENTIVENGA & HETRICK 1992, CHRISTIE & KILPATRICK 1992, MORTON et al. 1996, EGERTON-WARBURTON & ALLEN 2000). Yet, all the studies based on the production of spores of AM fungi suffer from a low temporal resolution caused by the spores' persistence in soil. Even more important is the high bias caused by the differential sporulation intensity of individual fungal populations (BEVER et al. 1996), scaling up to a situation where the dominant fungal symbiont can produce no or very few spores, so that the spore counts may provide a very skewed image (CLAPP et al. 1995, SANDERS et al. 1996).
Therefore, alternative approaches to studying communities of AM fungi within plant roots are needed. Among them, molecular biological methods are probably the most precise and elegant ones, and range from the direct studies of nucleic acids (CLAPP et al. 1995, SANDERS et al. 1996) through isozyme studies (TISSERANT et al. 1998) and immunofluorescence methods (WILSON et al. 1983). Nevertheless, few studies have used those methods to address real ecological questions (CLAPP et al. 1995, HELGASON et al. 1999). Three drawbacks limit the application of these molecular biological methods: (i) they are still too expensive for any medium-sized ecological project (with a hundred or more different samples); (ii) there are limitations caused by the peculiar properties of AM fungi (including genetic diversity within individual, heterokaryotic spores and fungal colonies - SANDERS et al. 1996); and (iii) these methods mostly provide information only about the presence of particular fungal genotypes, not about their abundance or the developmental stage of their colonies (but see HELGASON et al. 1999).
An alternative approach to evaluating the morphological properties of fungal colonies within plant roots was adopted by several researchers. Most of them differentiated between the morphologically very distinct "fine endophyte" and the other, "coarse" AM endophytes. Using even such a simple taxonomic distinction has often brought interesting ecological patterns into the studies (e.g. RABATIN 1979, SANDERS 8z FITTER 1992, CHRISTIE 8Z KILPATRICK 1992, RILLIG et al. 1999). A few authors have looked in more detail at the morphological variability of different AM fungi within host roots (ABBoTr 1982, BRUNDRETT & KENDRICK 1990, BLASCHKE 1991 ). THOMSON et al. (1992) addressed the effect of phosphorus application on endomycorrhizal fungi, and differentiated endomycorrhizal morphotypes within the plant roots, some to the species level. An exceptional and also isolated study in this area is that of MERRYWEATHER & FITTER (1998a and 1998b). These authors performed an extensive, quantitative study of the morphology of AM fungal colonies within the roots of Hyacinthoides

Communities of AM fungi in grassland 245
non-scripta, and then employed the study results to investigate the seasonal dynamics of the fungal populations within this host.
In this study, I was interested primarily in the comparative seasonal dynamics of AM fungi within the roots of three host species dominating an oligotrophic grassland, using the approach of intra-radical morphotypes similar to MERRVWEATHER & FITTER (1998a), and combining their results with information on the morphological descriptions of the intraradical colonies of different AM fungal taxa, as available from the International Culture Collection of VA Mycorrhizal Fungi (INVAM; http://invam.caf.wvu.edu). A comparison of the three host species is difficult to perform on the basis of AM fungal morphotypes of uncertain taxonomical status, so the presented results focus, in this respect, on a comparison of diversity and richness among the different host plant species.
Another interesting question concerns the effects of grassland management upon the composition of AM fungal communities. Several authors found a significant effect of fertilization on total mycorrhizal infection within a plant community (BENTIVENGA & HETRICK 1992, CHRISTIE & KILPATRICK 1992). There are also a few studies where this effect was studied in more detail, either looking directly at the taxonomic composition of AM fungal communities (EGERTON-WARBURTON & ALLEN 2000) or at least at its indication by the relative frequency of vesicles, arbuscules and hyphae (TITUS & LEPg 2000). Some of these papers also included a study of the effects that mowing the grassland has on the AM fungal infection (BEr~TWENGA & HETPdCK 1992, TITUS & LEPg 2000).
The host plant's nutritional status and age probably also have an important effect upon the symbiotic AM fungi, yet the knowledge of these factors is absent.
This study, therefore, targets the following four questions: (1) What is the extent of seasonal variability in the community of AM fungal morphotypes
within plant roots and how does this variability differ between the three grassland plant species?
(2) Do the host species differ in the diversity of AM fungal communities they host and what is the seasonal pattern of this diversity?
(3) Does the abandonment of mowing or the increased availability of phosphorus change the communities of AM fungi and/or their seasonal dynamics?
(4) Do the properties of plant individuals (their age, reproductive effort, presence of other fungal endophytes within the roots) influence the composition of AM fungal communities in their roots?
In this paper, I refer to the concept of arbuscular mycorrhizal (AM) fungal morphotypes using the acronym AMM.
MATERIALS AND METHODS
Research site
The research site is located near village of the Zvikov (10 krn east of (;esk6 Bud6jovice, Czech Republic, 48059 ' N, 14o36 , E, 500 m a.s.l.). The vegetation is an oligotrophic, traditionally-managed meadow on a shallow valley slope. The meadow is cut once a year, usually in the middle of June. More information can be found in ~MILAUER & ~MILAUEROVA (2000).
The plant species nomenclature follows ROTHMALER (1976).

246 Smilauer P.
Description of experiment
The seasonal dynamics of endomycorrhizal symbiosis were studied on plots under four different experimental regimes, resulting from a factorial combination of two experimental treatments: abandonment of mowing (using mown plots as control) and increased availability of phosphorus. The basic experimental plots were 1 × 1 m in size, separated by 0.5 m wide separation zones and arranged into four square blocks, each with four rows and four columns of plots. The two treatments were allocated in a split-plot arrangement, with eight adjacent sampling plots (in four rows and two columns) being mown and the other eight plots left un-mown, in each of the four blocks.
Four experimental blocks were established in 1997 and the experimental treatment was started the same year for two of them. In 1998, the treatment of the second pair of blocks started and these blocks were used for collecting plant material in 2000. The plants were collected from plots where the treatments had been in place for two seasons except for the last two collections in 1999, when the new blocks were used because the vegetation in the original two blocks had been disturbed by the constant removal of plants. The fertilization treatment was performed each autumn (September or October) and each spring (April) thereafter. At each application, 37 g of sodium phosphate (Na3PO4 . 12 H20), corresponding to 3 g of P, were distributed in 4 litres of water over each 1 m 2 plot. The grassland was mown in the first half of June.
In 1999, the dynamics of the endomycorrhizal fungal community were studied on three host plant species. Achillea millefolium is one of the most abundant herbs on the site. Poa angustifolia is the dominant grass and also Plantago lanceolata is a frequent and abundant plant at the site, and has a high level of endomycorrhizal infection throughout the whole year. In 2000, the seasonal variability of fungal community composition was studied only in the roots of Plantago and the samples were collected only from the mown plots.
The samples were taken every 3-4 weeks from mid-April to early October in 1999 and every two weeks from late March to mid-August in 2000. Plantago samples were also collected during a warm period in late February 2000.
Sample processing At each sampling time, five randomly-selected specimens of each species were collected
from each of the sampled plots. Each of the replicates was collected from a different sampling plot, but when three host species were sampled (in 1999), each chosen sampling plot was used to collect all three species. Therefore, not all sampling plots were used at each time - a subset of 20 plots was selected randomly each sampling time.
In the laboratory, roots were washed and the aboveground parts were described. For all three species, we counted the total number of leaves, number of non-damaged leaves, reproductive effort (estimated from the number of flower stalks and phenological development of their flowers) and weighed the total leaf biomass. For Plantago we additionally measured the diameter (two measurements in perpendicularly-positioned directions) and the height of its mesocorm (the accumulated nodes on the rosette base). The dimensions of the mesocorm provide a reasonable estimate of plant age. In 2000, we also classified each Plantago individual as young (without a prominent mesocorm, with primary roots dominating the root system), middle-aged or old.

Communities of AM fungi in grassland 247
The collected plant roots were stained using the modified staining procedure of PHILLIP & HAYMAN (1970), using a cold solution of KOH (applied overnight, with different timing for the three host species), and hot staining with Chlorazol Black E. Each stained sample was transferred into destaining solution and later mounted on a slide.
Morphotaxonomic units I modified the approach of MERRYWEATHER & FITTER (1998a) by developing a
classification of fungal morphotypes for each sampling time and for each host plant species, describing quantitatively their morphological features and then merging these partial classifications across the season at the end of the year 1999. The classification systems for the three host species were separate: morphological differences in colonization among the host species could have been due to the presence of different AM fungi or to the known effects of host on the morphology of fungal colonies (LACKIE et al. 1987, BRUNDRETT & KENDRICK 1990).
The properties of the more frequently occurring AMMs are summarized for the individual host species in Tables 1-3. Colour images demonstrating morphological properties of individual AMMs are available from http://regent.bf.jcu.cz/amms/index.htm. The xxGx AMMs are putatively assigned to mycorrhizal fungal species from the genus Glomus, the xxSc type to one or (probably) more species from the genus Scutellospora, and the xxA type to one or more species from the genus Acaulospora. The FE AMM refers to the group of "fine endophyte" morphotypes and its code is not preceded by a host species-specific prefix in this paper, as I think this morphotype is comparable across all three host species.
For each sample, I inspected the whole slide, recorded and described (using a scoring sheet with 18 different morphological features) each AMM found on the slide, and estimated the percentage of the root length occupied by each type and the percentage of root length with any mycorrhizal infection. These are semi-quantitative estimates based on a visual inspection of the whole root subsample on the slide. I have not used the standard grid-intersect scoring method because of the high number of samples and because of the need to devote a large amount of time to identifying and documenting the AMMs for each sample. The estimates of percentage share of root length infected by particular AMMs had to be adjusted to account for the spatial overlap of fungal colonies of different AMMs in the same root segment. In some samples (particularly those poorly stained or with intensive mycorrhizal colonization) I was not able to assign all colonies to one of the AMMs. In such case, the percentage of root length occupied by undetermined arbuscular fungi was recorded. This fraction was later distributed proportionally among the recognized AMMs, except the fine endophyte AMM, which was deemed to be always recognizable in the roots. Additionally, the staining quality of each slide was scored and this value was used together with the percentage of undetermined mycorrhizal colonies to calculate the weight of the particular observation in the statistical analyses. The observation weight was equal to 1 - (PUF/100.0), where PUF is the "percentage of unrecognized fungal types". This weight was further multiplied by 0.75 or 0.5 for somewhat or substantially understained/overstained slides. Slides that were much overstained (with infection regions having an uniformly dark blue colour) or understained (so that only vesicles appeared slightly against the root background) were excluded from the data.
To compare the diversity of recognized AMMs with the diversity of mycorrhizal spores, we used two approaches to extract spores from the field soil. First, we isolated the spores directly

po
Tab
le 1
. Mor
phol
ogic
al c
hara
cter
isti
cs o
f the
mos
t im
port
ant a
rbus
cula
r m
ycor
rhiz
al m
orph
otyp
es i
n th
e ro
ots
ofA
chil
lea
mil
lefo
liu
m (
am).
H -
hyp
hae,
V -
vesi
cles
, oo
E
P -
entr
y po
int,
G -
G
lom
us,
Sc
- S
cute
llo
spo
ra,
Ac
- A
cau
losp
ora
, F
E -
"f
ine
endo
phyt
e".
Dis
trib
utio
n in
A
uxil
iary
H
ypha
e at
ent
ry p
oint
In
trar
adic
al h
ypha
e A
rbus
cule
s V
esic
les
root
cor
tex
cell
s
am
G1
in
ner
and
oute
r ab
sent
no
t pro
min
ent,
mul
tipl
e-S
para
llel
, 3.
2-5.
1Fxm
nu
mer
ous
shap
ed H
di
amet
er,
som
etim
es
sept
ate,
wit
h pe
gs
freq
uent
, 40
-70
~tm
long
ov
al o
r le
mon
-sha
ped
am
Sc
mos
tly
in o
uter
so
met
imes
pr
omin
ent
coil
ing
of H
nea
r of
ten
wit
h pr
omin
ent
furt
her
from
EP,
ab
sent
pr
esen
t en
try
poin
t, e
xtra
radi
cal
H
proc
esse
s, 5
.5--
6.5
~tm
bu
t th
en n
umer
ous
ofte
n at
tach
ed
diam
eter
, of
ten
very
co
iled
am
Ac
inne
r an
d ou
ter
abse
nt
prom
inen
t coi
ling
of H
, but
us
uall
y no
t wel
l vis
ible
, H
the
re u
sual
ly th
icke
r th
an t
he f
eede
r H
char
acte
rist
ic H
str
uctu
res
(und
ulat
e, c
rest
-lik
e) o
ften
fo
rmed
; V o
ften
for
med
he
re
para
llel
, oft
en th
inne
r th
an n
ear
EP
(2-
3 ~u
n);
stai
n fa
intl
y
very
thin
(1-
2 ~t
m),
usua
lly
seve
ral H
ru
n to
geth
er
freq
uent
in
mat
ure
colo
nies
oft
en f
illi
ng
com
plet
ely
the
cell
s (b
rick
-lik
e ap
pear
ance
)
seem
to o
ccur
onl
y in
pa
rtic
ular
dev
elop
men
tal
stag
es o
f co
lony
(of
ten
abse
nt,
mas
sive
de
velo
pmen
t if p
rese
nt)
FE
m
ostl
y in
out
er
abse
nt
rare
ly s
een,
nea
r E
P 50
-80
pan
long
, of
ten
wit
h ir
regu
lar
shap
e
smal
l (a
roun
d 7p
ro)
vesi
cle-
like
str
uctu
res
occu
rrin
g ne
ar E
P
~c
e-- "13

Tab
le 2
. M
orph
olog
ical
cha
ract
eris
tics
of
the
mos
t im
port
ant
arbu
scul
ar m
ycor
rhiz
al m
orph
otyp
es i
n th
e ro
ots
ofP
oa
a
ng
ust
ifo
lia
(p
a).
Add
itio
nall
y,
A c
aulo
spor
a-li
ke
AM
Ms
wer
e oc
casi
onal
ly s
een
(wit
h nu
mer
ous
row
s of
bri
ck-l
ike
arbu
scul
es,
no v
esic
les
and
usua
lly
a po
or s
tain
ing
of th
e th
ick
hyph
ae).
H
- h
ypha
e, V
- v
esic
les,
EP
- en
try
poin
t, G
-
Glo
mus
, Sc
- S
cute
llos
pora
, F
E -
"fi
ne e
ndop
hyte
".
Dis
trib
utio
n in
A
uxil
iary
H
ypha
e at
ent
ry p
oint
In
trar
adic
al h
ypha
e A
rbus
cule
s V
esic
les
root
cor
tex
cell
s
paG
1 m
ostl
y in
ver
y ab
sent
no
t pro
min
ent
smoo
th,
rare
, 3-
4 ~r
n ve
ry f
requ
ent
in m
ost
thin
roo
ts w
ith
in d
iam
eter
, Par
is-
colo
nies
on
e or
few
cel
l -t
ype
infe
ctio
n la
yers
appe
ar n
umer
ousl
y at
lat
er s
tage
s,
35--
40 ~
tm lo
ng
FE
in
out
er c
orte
x ab
sent
se
em to
oc
cu
r on
ly in
la
yers
, if
pa
rtic
ular
dev
elop
men
tal
diff
eren
tiat
ed
stag
es o
f co
lony
(of
ten
abse
nt,
mas
sive
de
velo
pmen
t if p
rese
nt)
paSc
di
tto
rare
ly s
een
atta
ched
ext
rara
dica
l H
4-
6 la
in in
dia
met
er,
scat
tere
d, p
oorl
y vi
sibl
e ab
sent
of
ten
visi
ble;
pro
min
ent
outl
ine
wit
h pr
ojec
tion
s hy
phal
coi
ling
char
acte
rist
ic H
str
uctu
res
very
thin
(1-
2 In
n),
(und
ulat
e, c
rest
-lik
e) o
ften
us
uall
y se
vera
l H r
un
form
ed;
V o
ften
pre
sent
her
e to
geth
er
smal
l (ar
ound
7 ~
tm)
vesi
cle-
like
str
uctu
res
occu
rrin
g ne
ar E
P
o O 3 3 t- 5"
g~
.-i
O.
¢O
¢D

IX)
Tab
le 3
. Mor
phol
ogic
al c
hara
cter
isti
cs o
f the
mos
t im
port
ant a
rbus
cula
r m
ycor
rhiz
al m
orph
otyp
es i
n th
e ro
ots
ofP
lan
tag
o l
ance
olat
a (p
/).
H -
hyph
ae,
V -
vesi
cles
, A
- a
rbus
cule
s, E
P -
ent
ry p
oint
, G
- G
lom
us,
Sc -
Scu
tell
ospo
ra,
Ac
- A
caul
ospo
ra,
FE
- "
fine
end
ophy
te".
Dis
trib
utio
n in
A
uxil
iary
H
ypha
e at
ent
ry p
oint
In
trar
adic
al h
ypha
e A
rbus
cule
s ro
ot c
orte
x ce
lls
plG
l in
ner
and
oute
r ab
sent
pr
omin
ent,
but
H a
rran
gem
ent
mul
tipl
e-S-
shap
ed,
not
coil
ing
3-6
lam
thic
k, u
sual
ly
wit
h m
any
knob
s,
olde
r H
sep
tate
plG
2
mai
nly
inne
r ab
sent
no
t muc
h ap
pare
nt,
sim
ple
3-41
am t
hick
, so
me
arra
ngem
ent
smoo
th o
r w
ith
not-
-s
o pr
omin
ent
knob
s;
olde
r H
sep
tate
plG
3
oute
r ab
sent
pr
omin
ent
coil
ing,
al
l wit
h pr
omin
ent p
egs
sim
ilar
to p
lSc
type
or
pro
cess
es,
thic
k (5
-7 ~
tm)
prom
inen
t co
ilin
g, H
som
ewha
t th
icke
r he
re (
6-9
~tm
) p
lSc
mai
nly
in o
uter
se
en
plA
c V
in
oute
r, A
ab
sent
S-
like
arr
ange
men
t m
ainl
y in
inn
er
of H
nea
r E
P
char
acte
rist
ic H
str
uctu
res
(und
ulat
e, c
rest
-lik
e) o
ften
fo
rmed
; V
oft
en p
rese
nt h
ere
FE
m
ostl
y in
out
er
abse
nt
thic
k (4
-8 o
m),
oft
en
wit
h pr
oces
ses
or p
egs;
ne
ver
sept
ate
poor
ly (
vari
ably
) st
aini
ng,
diff
icul
t to
obse
rve
in r
oots
wit
h hi
gh A
M i
nfec
tion
very
thin
(1-
2 ~t
m),
usua
lly
seve
ral H
run
to
geth
er
typi
cal
Glo
mus
-lik
e A
, w
ith
two
base
bra
nche
s;
do n
ot f
ill w
hole
cel
l;
scat
tere
d
at m
atur
ity
fill
the
who
le
cell
; num
erou
s, i
n co
mpl
ete
row
s
very
rare
, sc
atte
red
scat
tere
d to
num
erou
s,
rela
tive
ly th
ick
trun
k
fill
ing
the
who
le c
ells
; of
ten
bric
k-li
ke
appe
aran
ce v
ery
fain
tly
stai
ning
, ra
rely
obs
erve
d
seem
to o
ccur
onl
y in
pa
rtic
ular
dev
elop
men
tal
stag
es o
f col
ony
(oft
en
abse
nt,
mas
sive
de
velo
pmen
t if p
rese
nt)
Ves
icle
s
larg
e (5
0-80
lam
long
),
lem
on-s
hape
d, w
ith
one
larg
e, e
asil
y vi
sibl
e va
cuol
e
30-5
0 ~t
m in
dia
met
er,
oval
or
alm
ost r
ound
, us
uall
y op
aque
(n
o va
cuol
e se
en)
larg
e (4
0-80
I.tm
), ov
al
or ir
regu
larl
y sh
aped
, in
tens
ivel
y st
aini
ng,
near
EP
abse
nt
rare
, fo
rmed
mor
e or
less
just
nea
r E
P
smal
l (a
roun
d 7
Ixm
) ve
sicl
e-li
ke s
truc
ture
s oc
curr
ing
near
EP
(/)c
~0
e- "O

Communities of AM fungi in grassland 251
from the soil collected around the roots of plants from field, using the wet sieving and sucrose flotation based separation (YORK MYCORRHIZA RESEARCH GROUP 1999). Additionally, we established trap cultures (YORK MVCORRHIZA RESEARCH GROUP 1999) with the field-collected soil and Plantago roots, using Plantago lanceolata as the bait plant, and collected spores after three months of cultivation. As the observations on the collected spores are only supplementary in this study, their results are provided in the Appendix.
Data analysis
The changes in seasonal dynamics of recognized AMMs are too complex to fit into simple statistical models that use time as a quantitative explanatory variable. Therefore, I used statistical analyses where the sampling time acts as a factor. The trends can then be inferred from the graphs displaying average abundance of individual AMMs at individual sampling times (Fig. 1-6).
To compare and test changes in the whole community of AM fungi through time and due to the experimental treatments and properties of the sampled individuals, a constrained linear ordination method (redundancy analysis - RDA) was used together with a Monte Carlo permutation test (see TER BRAAK & ~M1LAUER 1998). In each test, 1999 independent randomizations were used. Because the AMMs cannot be compared across host species, I performed the tests separately for each of the three host species. The RDA used with qualitative explanatory variables (e.g. sampling time) is a modification of multivariate ANOVA, with a reduced dimensionality of the set of response variables. The estimated percentage abundance of individual AMMs was log-transformed using the log(x+l) transformation.
The fungal community diversity was measured by the number of recognized AMMs. The effects of time, of host species identity, and of experimental treatment upon the richness of AMM community were evaluated using the analysis of variance.
The effects on community composition that were found significant in the multivariate permutation tests were further tested using one-dimensional statistical models (either ANOVA models or regression models, depending on the type of the predictor) and in this case the Bonferroni correction was applied when tagging the significantly reacting response variables (i.e. the individual AMMs). The split-plot design of the experimental plots (fertilization treatments nested within the mowing treatment plots) was taken into account.
RESULTS
Seasonal dynamics of morphotypes
In the Achillea roots (Fig. 1), amSc and amG1 were the two dominant morphotypes (AMMs). The amSc AMM increased its dominance before mowing, and then was gradually replaced by the amG1 AMM. This situation reversed at the end of the season. The FE ("fine endophyte") AMM maintained a more-or-less constant abundance throughout the season.
Roots of Poa (Fig. 2) were dominated by the paG1 AMM, while the paSc population gradually decreased and finally disappeared at the end of the season. The behaviour of the FE AMM was similar to that in the Achillea roots.
The community of AMMs in the Plantago roots was the richest (Fig. 3 and Fig. 4) among the three hosts species, and also exhibited the highest equitability, with no AMM dominating.

252 S m i l a u e r P.
100 Achillea 1999
- , 8O O) t - Q
: 60 0
e " 0
~ 4o
~ 20
--0- amG1 - - ~ amSc
- I - F E .... o.... Empty
50 100
h= , cu t i E
' i
/ , i
- "0 i
150 200 250 300 [days since I J a n u a r y ]
Fig. 1. The seasonal dynamics of AM morphotypes in the roots ofAchillea millefolium in 1999. The average of the estimated percentage of root length occupied by the particular morphotype, calculated over all the experimental treatments, is plotted against the ordinal number of sampling date, relative to 1 January 1999. Mowing time is indicated by a vertical line. The averages from the first and second sampling are omitted because few reliable samples were available (the unreliability of the samples is allowed for in the statistical analyses by lower observation weights). The partial follow-up tests using ANOVA found significant difference between the sampling time in amSc, amG1 and in total infection (i.e. complement of percentage of non-infected root length).
100 Poa 1999
8O ,,¢:
t -
_o • ,., 60 0
e " 0 (I)
40 J~ ._=
paG1 - - ~ paSc
- i = FE -.c~- Empty
20
0 0
h a y c u t
.............. 0-- 0 , .
. . . . . . . i - _ ? =
~ v
50 100 150 200 250 300 [days since 1 January]
Fig. 2. The seasonal dynamics of AM morphotypes in the roots ofPoa angustifolia in 1999. The average of the estimated percentage of root length occupied by the particular morphotype, calculated over all the experimental treatments, is plotted against the ordinal number of sampling date, relative to 1 January 1999. Mowing time is indicated by a vertical line. The averages from the first and second sampling are omitted because few reliable samples were available. The partial follow-up tests using ANOVA found significant differences between the sampling times in FE, andpaG1 (and inpaSc, if ignoring Bonferroni's argument for multiple tests).
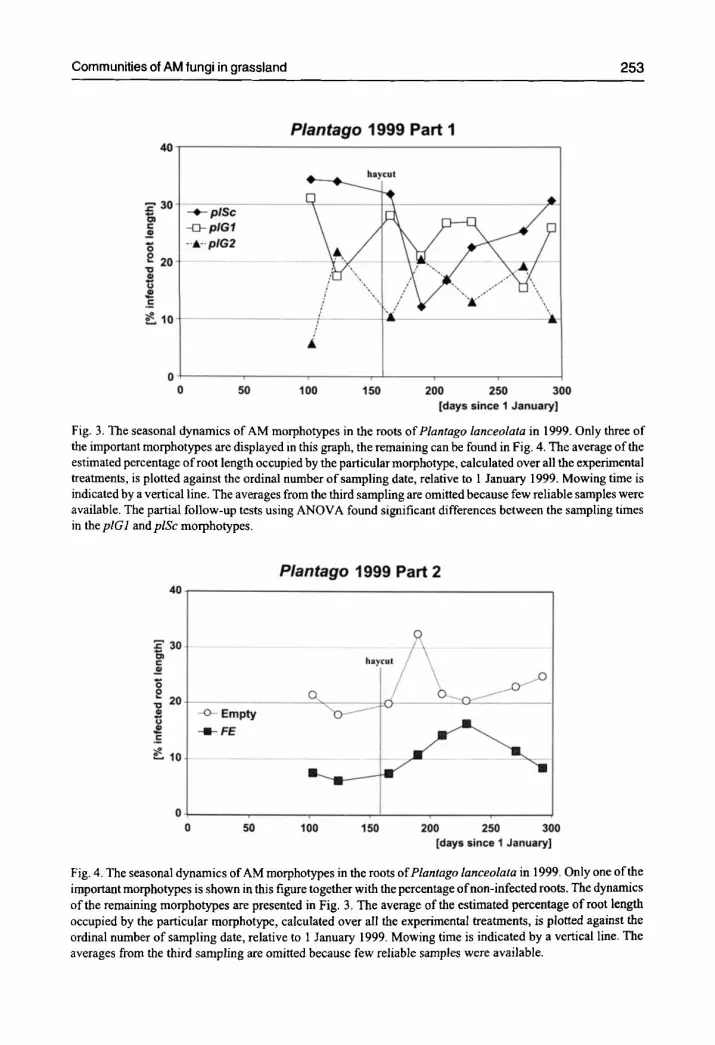
Communi t ies of AM fungi in grass land 2 5 3
40-
30- ¢m ¢- o
o 20- "O Q
Jz c
~1o
Plantago 1999 Part I
- e - p lSc - [3- p iG 1 -.A-- piG2
~ _ . ~ h a y c u t
/ ,.,. ) / ",i
/
A
tL A
0 50 100 150 200 250 300 [days since 1 January]
Fig. 3. The seasonal dynamics of AM morphotypes in the roots ofPlantago lanceolata in 1999. Only three of the important morphotypes are displayed in this graph, the remaining can be found in Fig. 4. The average of the estimated percentage of root length occupied by the particular morphotype, calculated over all the experimental treatments, is plotted against the ordinal number of sampling date, relative to 1 January 1999. Mowing time is indicated by a vertical line. The averages from the third sampling are omitted because few reliable samples were available. The partial follow-up tests using ANOVA found significant differences between the sampling times in the plG1 and plSc morphotypes.
40- Plantago 1999 Part 2
~' 30 c _o
P 20 "ID
U ~e _=
='~ 10 . . . . .
Q / \
haycut / ~\ / \ ..D
/ \ 0 ....... O O __ O.-... O ......................
~ k a a p~O
Empty
- N - F E
nL.~_...~
0 0 50 100 150 200 250 300
[days since 1 January]
Fig. 4. The seasonal dynamics of AM morphotypes in the roots ofPlantago lanceolata in 1999. Only one of the important morphotypes is shown in this figure together with the percentage of non-infected roots. The dynamics of the remaining morphotypes are presented in Fig. 3. The average of the estimated percentage of root length occupied by the particular morphotype, calculated over all the experimental treatments, is plotted against the ordinal number of sampling date, relative to 1 January 1999. Mowing time is indicated by a vertical line. The averages from the third sampling are omitted because few reliable samples were available.

254 Smilauer P.
50 Plantago 2000 Part I
4o
~ 3 0 _~_ plSc e -~-plG1
-.i-. piG2
.E ...,A ."
' " t o . . . . " ~ i ",, - -
i 0 , , , ,
50 100 150 200 250 300 [days since I January]
Fig. 5. The seasonal dynamics of AM morphotypes in the roots ofPlantago lanceolata in 2000. Only three of the important morphotypes are displayed in this figure, the remaining ones can be found in Fig. 6. The average of the estimated percentage of root length occupied by the particular morphotype, calculated over all the experimental treatments, is plotted against the ordinal number of sampling date, relative to 1 January 2000. Mowing time is indicated by a vertical line. The partial follow-up tests using ANOVA found significant differences between the sampling times in the plSc morphotype.
50
4O
¢ :
0 30
o " O o u 20 = ._c
10
Plantago 2 0 0 0 Par t 2
0 '\ \ --o-.-.Emp~ -B -FE
\ (~ haycut \ ! \ - \ \ / \ \ / . , ~. , /
0 50 100 150 200 250 300 [days since 1 January]
Fig. 6. The seasonal dynamics of AM morphotypes in the roots ofPlantago lanceolata in 2000. Only one of the important morphotypes is shown in this figure together with the percentage of non-infected roots. The dynamics of the other morphotypes are presented in Fig. 5. The average of the estimated percentage of root length occupied by the particular morphotype, calculated over all the experimental treatments, is plotted against the ordinal number of sampling date, relative to 1 January 2000. Mowing time is indicated by a vertical line. The partial follow-up tests using ANOVA found significant differences between the sampling times in the FE morphotype.

Communities of AM fungi in grassland 255
Richness of A M M community '[ hxy[em
~3~ . . . . . . . . . . . . . . . . . . . . . I
0 ~ _ e _ P L . 9 9 , i -O-PL.00 il .z'J " ................ "
2 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . : i "- . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . _ --n,-AM'~ .~ . . . * - . m m-: ,~. * - . , . . , - ~,
P A ' 9 9
n s 1- •
0 o 5o Ioo .15o 2c;o ~,~ ~oo
[days since 1 J a n u a r y ]
Fig. 7. The seasonal changes of the richness (average number of recognized morphotypes) of an AM fungal community in the roots of three host plant species. The richness is plotted against the ordinal number of sampling day, relative to 1 January of the respective year. Indication of the approximate mowing time is added to the picture. The averages from the third sampling are omitted for Plantago (in year 1999) and from the first and second sampling for both Poa and Achillea, because few reliable samples were available.
Beside the four AMMs shown, an additional two occurred infrequently and only in limited parts of the season (plAc, piG3 - see description in Tab. 3). The most abundant AMMs in the Plantago roots are the plSc, plG1, andplG2 types. TheplSc type had a large abundance in the period before mowing, but then declined sharply and recovered again at the end of the season. The mowing event influences the total intensity of AM infection in Plantago roots, unlike in the other two host species (see the category Empty in Fig. 4).
The population dynamics ofAMMs of Plantago in the year 2000 are summarized in Fig. 5 and Fig. 6. The patterns in the dynamics of AMMs in this year are very similar to patterns in the previous year (displayed in Fig. 3 and Fig. 4). There was a similar trend of high abundance of theplSc type in May and early June, followed by a sharp drop of abundance in the summer time (July, August). The plG1 was the second most important player in the AMM community of Plantago, which dominated over the plSc type in early spring and increased its abundance again at the time ofplSc AMM depression in mid-summer. The fme endophyte AMM peaked in mid-summer (July, August).
Tests of the effects of sampling time upon the AMM communities composition (using the multivariate permutation test, based on RDA) reveal highly significant effects for all three host species (Tab. 4). Note that the communities differ in the total number of recognized AMMs and this fact alone influences the absolute value for the percentage of explained variability in response variables (in multivariate ordination methods, the notion of degrees of freedom extends also onto the response variables' side of the relation). Therefore, the explanatory power of the sampling time is better assessed by the F statistics, comparing the variability explained by the time with the residual variability (adjusted by the number of residual degrees of freedom). Judged from the values of these statistics, the strongest effect can be seen in 1999, for the Plantago roots, followed by Achillea and Poa.

256 Smilauer P.
Table 4. Summary of the direct gradient analyses (RDA method) applied to the AMM community composition data separately for individual host plant species and (in case of Plantago) for individual years, using the sampling time (9 samplings in the year 1999 and 12 sampling forPlantago in the year 2000) as the explanatory variable. Percentage of variance in the community composition data explained by all the canonical axes together is given in the second column. The values in the third (F-like statistics) and fourth (Type I error estimate) column refer to the related Monte Carlo permutation test of significance for the relation between the AMM community composition and sampling time.
Host species Percentage explained F P
Achillea millefolium 33.2% 8.213 0.0005 Poa pratensis 31.5% 7.527 0.0005 Plantago lanceolata 1999 23.2% 12.378 0.0010 Plantago lanceolata 2000 21.3% 2.66 0.0005
Seasonal dynamics of AMM richness
I found significant differences in AMM community richness between host species and between sampling times (Tab. 5). The changes of richness through the season also differed between the species (as indicated by significant interaction term of the two factors). Nevertheless, the shape of the richness pattern (in Fig. 7) is more or less comparable for all three host species, culminating in the early summer (June, July), and then slowly declining towards the end of season.
Effects of mowing, phosphorus application, and host plant properties
The relation between the AMM community composition and the experimental treatments and host plant properties was tested for each explanatory variable separately, using the RDA method.
For Poa, there was a significant difference in AMM community composition between the mown and un-mown plots (P = 0.0095). The resulting ordination diagram (not shown here) suggested that when mowing was discontinued, the abundance of paSc and (to a smaller extent) ofpaG1 increased and the percentage of non-infected roots decreased. The follow-up tests (using ANOVA with effect of mowing nested within the time) indicate that this difference is significant only for the paSc AMM (P = 0.0027), which has almost double (1.9 x) the average abundance in plants from un-mown plots compared with the plants from mown plots. For Achillea, neither the experimental treatments, nor plant properties had any significant effect.
In the case ofPlantago, a nearly-significant relation between the AMM community and the volume ofmesocorm (P = 0.0720) became significant with the inclusion of data from the year 2000 (P = 0.0250). In the follow-up tests based on a linear regression model, only the increase in the percentage oftheplG2 morphotype with the increasing mesocorm volume (representing increasing age of the host plant) was found significant (P = 0.0031). Because we also performed a subjective classification of the age (young, medium-aged, or old) for each plant collected in the year 2000, the explanatory power of this descriptor was tested separately for the AMM community data from the year 2000. It was found to be the best predictor for the AMM community composition (P = 0.0225). By testing the partial effects on individual

Communities of AM fungi in grassland 257
Table 5. Summary of the results of analysis of variance using the sample richness (number of AM morphotypes) as the response variable and the identity of host plant and sampling time as factors with a full interaction.
Term d.£ Sum of squares Mean square F P
Species 2 178.09 89.05 284.7 <10 -6 Time 8 20.50 2.56 8.2 <10 -6 Species x Time 16 12.18 0.76 2.4 0.0016 Residuals 421 131.70 0.31
Table 6. Summary of the results of the general linear model using the sample richness (number of AM morphotypes) as the response variable and the host plant properties (additional to the effect of host plant species identity) as the explanatory variables.
Term d.f. Sum of squares Mean square F P
Species 2 178.09 89.05 239.5 <10 -6 Total number of leaves 1 0.10 0.10 0.3 n.s. % of undamaged leaves 1 2.23 2.23 6.1 0.014 Number of reproductive stalks 1 0.21 0.21 0.6 n.s. Residuals 442 161.84 0.37
AMMs using ANOVA, I found just one nearly significant relation with theplG2 increasing its percentage abundance with the increasing age of the host plant (P = 0.060).
In the test of effects of experimental factors (mowing and fertilization) on the richness of the AMM community, using the 1999 data, neither mowing nor phosphorus addition had a significant effect. The effects of plant characteristics (total number of leaves, percentage of undamaged leaves, number of reproductive stalks) are summarized in Tab. 6. There is a significant effect of proportion of undamaged leaves - the plants with a higher fraction of damaged leaves (mostly by herbivores or fungal pathogens) have a richer AMM community.
DISCUSSION
Seasonal variability of AMM community
The statistical analysis revealed profound and highly significant variability in the community composition of AMMs for all three host species throughout the observation period. Similar conclusions were also made by other authors studying seasonal variation of AM symbiosis (MEm~YWEATHER & FITTER 1998b, TTTUS & LEeR 2000). However, only some of the individual AMM populations show significant changes because: (i) the multivariate test is not just an "over-position" of the tests for individual AMMs, and (ii) in both the multivariate and univariate tests, time is a factor, so the individual sampling times are treated as factor levels. I f we evaluate the contents of the figures such as Fig. 5, we can see a consistency in the smooth changes over time, for example in the behaviour oftheplG1 morphotype, even though this one was not judged to be a significant pattern in ANOVA.
A very interesting pattern - observed both in the Achillea roots in 1999 and in the roots of Plantago in both years - is the low abundance of the Scutellospora-like morphotypes in the

258 Srnilauer P.
early spring, followed by its rapid increase, and then by a limited retreat in summer, possibly a consequence of the increased competition of other species (such as amG1 or plG1 in the roots of, respectively, Achillea or Plantago). This is a pattern consistent with the findings of MERRYWEATHER & FITTER (1998b) although they worked with a host plant with very different seasonality (dormant in summer). They found that a Scutellospora morphotype was replaced by either Acaulospora or Glomus morphotypes at the time of flowering.
Although Scutellospora AMMs decline after mowing in all three host species, it is likely that other factors (environmental or phenological) were responsible for this decline since half of the samples came from the unmown plots.
Patterns of species diversity and, particularly, of species richness show peaks in the summer period - especially forAchillea and Poa, and are supplemented with a peak in early spring - in Plantago. Those peaks coincide with the most intensive flowering of the host plant populations: Plantago plants flower before the mowing time and after the mowing recover quickly and renew flowering. Poa is (on our site) usually flowering at the time of hay-cut and does not create new culms after mowing. Achillea usually flowers after mowing, in July and August. The pattern of highest diversity of AM fungi aligned with the flowering time of host species is consistent with results of MERRYWEATHER & FITTER (1998b), where the high diversity of recognized AM morphotypes occurred in the spring time, when the host plant (bluebell) is flowering.
Effects of mowing, phosphorus application, and host plant properties
I found no effect of the fertilization by phosphate either upon the composition of the AMM community, or on its diversity. The effect of phosphate application on the grassland productivity was evident in the plots at the time the plants were collected (i.e. two years after the P application started), but P did not affect species composition (unpubl. results). The absence of the effect of increased availability of nutrients contrasts strongly with the other published results (BENTIVENGA & HETRICK 1992, THOMSON et al. 1992, EGERTON-WARBURTON & ALLEN 2000), and even with our results from the same research site. In a different field experiment, the fungicide-based exclusion of AM fungi was combined with the addition of P using the same application rate as in this experiment, but for a much longer time, and we found decreased total AM infection and decreased arbuscular frequency for the three plant species used in this study (see ~MILAUER & ~MILAUEROV,/~ 2000). It is also necessary to stress that some of the above-mentioned studies (namely THOMSON et al. 1992 and EGERTON-WARBURTON 8~; ALLEN 2000) worked with a much steeper gradient of nutrients availability.
Similarly, the effect of abandoned mowing was mainly seen in the increased accumulation of plant litter and in the abundance of a few species, but no species disappeared or invaded due to this change of management. It is, therefore, not surprising that mowing had little effect on AMMs, except in the AMM communities in the Achillea roots where one of the participating morphotypes increased after the mowing ceased. BENTWENGA & HETRICK (1992) made similar conclusions, namely that the effect of management practices (they were studying mowing, fertilization, and burning, in a tallgrass prairie) on the AM fungal community occurs only i fa substantial change in the host plant community is induced.
An interesting result is the relation found between the richness of AM fungal community and the extent of damage of host plant leaves. We can relate this result to the findings of

Communities of AM fungi in grassland 259
GOVERDE et al. (2000) who found that mycorrhizal symbiosis modifies palatability of host plant leaves to butterfly larvae and, consequently, substantially increases their survival and speeds-up their development. These authors found in their study that the mycorrhizal plants had a higher C/N ratio in the dry mass of leaves, unlike GANGE & WEST (1994) who actually suggested an adverse effect of AM symbiosis upon the herbivores, where the fungal symbionts represent an additional sink for carbon and thus lower the C/N ratio in the host plant biomass.
Difference in AMMs between host species This study demonstrates the persistent difference in AM fungal communities between
different plant species growing together on a grassland site. The actual morphological features ofhyphae, vesicles, and arbuscules, as well as the character of the whole fungal colony within the host roots is substantially influenced by the morphological and other properties of the host plant. The arbuscular mycorrhizae in the Poa roots can be classified mostly as Paris-type morphotypes, while those in Achillea and Plantago as Arum-type (SMITH • SMITH 1997), with the latter category featuring an extensive extracellular mycelial network within the plant roots. The fine endophyte colonies seem to have rather comparable morphological features across the three host species, but not completely so, and substantial variability within this category in individual host plant species can be seen.
The results presented in this paper indicate that the dominant colonizing species can differ between the host species (Glomus spp. morphotypes in Poa, but Scutellospora spp. and Glomus spp. swapping their lead through the season in the roots of Achillea and Plantago), but it is quite possible that even the morphotypes assigned to the Acaulospora genus can dominate roots of certain host plant species. The fact that some of the AM fungi infect the roots of several or even many plant species and that their relative importance differs between those hosts can have substantial consequences in terms of the impact of mycorrhizal fungi upon the diversity and stability of plant communities. This effect was, in fact, already demonstrated by VAN DER HEIJDEN et al. (1998a).
This study also confirms that not only can each species host more than one taxon of AM fungus, but also the same AM fungus can occur in more than one host plant species. While this conclusion cannot be made directly from the observation of AM morphotypes, it follows from the number of isolated spore-based morphotypes. Seven isolated morphotypes is not definitively a final number and will increase with continued sampling of the area (see for example the study ofBEVER et al. 1996, where more than 23 AM fungal taxa were found in an area of 75 m2), but there are approximately 90 mycorrhizal grassland plant species in this research site. This study confirms the current view that different AM species have a different affinity to particular host plant species (BEVER et al. 1996, EOM et al. 2000).
Applicability of the morphotype-based approach in the ecological studies
It is impossible to assign with certainty each observed fungal colony to one of the defined morphotypes. In the fungal communities of all three host species, but namely in the most species-rich community of AM fungi in the Plantago roots, I often found a substantial spatial overlap among colonies belonging to different morphotypes. This fact, together with an occasional imperfection of samples (overstained in the areas with a high infection intensity or partly understained), was explicitly taken into account in the statistical analyses employed in this paper. Additionally, misclassification can occur. In the case of the Plantago AMM

260 Smilauer P.
community, the most likely mistakes were between the individual Glomus-type morphotypes and also to some extent between the Scutellospora and Acaulospora morphotypes: the Acaulospora colonies sometimes lacked vesicles and when combined with an occasionally better staining ofAcaulospora hyphae, they could be incorrectly recorded as a Scutellospora type. On the other hand, the poorly staining Acaulospora hyphae can be overlooked in the areas with a high infection of the better staining types.
The endomycorrhizal symbionts can substantially influence the populations of higher plants and they are, therefore, increasingly considered in the recent ecological studies. I am convinced that the approach based on estimating the presence of recognized morphotypes is - despite its difficulty - one of the very few reasonable ways forward. Characterizing the dynamics of endomycorrhizal symbiosis using field-collected spores is just like characterizing the spatial or temporal dynamics of a community of higher plants by ignoring the plants themselves and looking only at the seed bank in the soil. The methods of molecular biology are probably the most precise and valuable approaches. Yet these methods might remain prohibitively expensive for any medium-sized ecological project for a much longer time than the ecologists can wait with their questions.
The most important future improvement of the AMM typology (except the gradual, slow enhancement based on increasing experience) is the isolation of endomycorrhizal taxa into pure cultures that can be used to study the range of morphological features within the target host plant species, in dependence on the environmental conditions. The isolation will also enable us to validate the taxonomic identity of the morphotypes using the techniques of molecular biology.
Acknowledgements: My thanks go first and foremost to my wife Marie, who contributed not only with her encouragement, but performed a large part of the collection of plants in field and their description. Another large part of the manual labour was conducted by my technician, Blanka Divigowl, and I give her my thanks here. James Merryweather provided his optimism about the work with endomycorrhizal fungi. I want to give particular thanks to Alastair Fitter and two anonymous reviewers, who provided substantial input for improving the original manuscript. The project was supported from the research grant no. 206/98/P014 of the Grant Agency of the Czech Republic and from grant MSM 123100004 of Czech Ministry of Education.
REFERENCES
ABBOTT L.K. (1982): Comparative anatomy of vesicular-arbuscular mycorrhizas formed on subterranean clover. Austral. J. Bot. 30: 485-499.
ALLEN M.F. (1982): Influence of vesicular-arbuscular mycorrhizae on water movement through Bouteloua gracilis. New Phytol. 91: 191-196.
BENTIVENGA S.P. & HETRICK B.A.D. (1992): The effect of prairie management practices on mycorrhiza} symbiosis. Mycologia 84: 522-527.
BEVER J.D., MORTON J.B., ANTONOVICS J. & SCHULTZ P.A. (1996): Host-dependent sporulation and species diversity of arbuscular mycorrhizal fungi in a mown grassland. £ Ecol. 84: 71-82.
BLASCHKE H. (1991): Distribution, mycorrhizal infection, and structure of roots ofcalcicole floral elements at treeline, Bavarian Alps, Germany. Arctic Alpine Res. 23: 444-450.
BRUNDRETT M. & KENDRICK B. (1990): The roots and mycorrhizas of herbaceous woodland plants. II. Structural aspects of morphology. New Phytol. 114: 469-479.
CHRISTIE P. & KILPATRICK D.J. (1992): Vesicular-arbuscular mycorrhiza infection in cut grassland following long-term slurry application. Soil Biol. Biochem. 24: 325-330,
CLAPP J.P., YOUNG J.P.W., MERRYWEATHER J.W. & FITTER A.H. (1995): Diversity of fungal symbionts in arbuscular mycorrhizas from a natural community. New Phytol. 130: 259-265.

Communities of AM fungi in grassland 261
EGERTON-WARBURTON L.M. & ALLEN E.B. (2000): Shifts in arbuscular mycorrhizal communities along an anthropogenic nitrogen deposition gradient. Ecol. Appl. 10: 484-496.
EOM A.H., HARTNETT D.C. & WILSON G.W.T. (2000): Host plant species effects on arbuscular mycorrhizal fungal communities in tallgrass prairie. Oecologia 122: 435--444.
FITTER A.H. (1977): Influence of mycorrhizal infection on competition for phosphorus and potassium by two grasses. NewPhytol. 79:119-125.
FRANCIS R. & READ D.J. (1994): The contributions of mycorrhizal fungi to the determination of plant community structure. PI. & Soil 159:11-25.
GANGE A.C. & WEST H.M. (1994): Interactions between arbuscular mycorrhizal fungi and foliar-feeding insects in Plantago lanceolata. NewPhytol. 128: 79-87.
GEMMA J.N., KOSKE R.E. & CARREIRO M. (1989): Seasonal dynamics of selected species ofVA mycorrhizal fungi in a sand dune. Mycol. Res. 92: 317-321.
GOVERDE M., VAN DER HEIJDEN M.G.A., WIEMKEN A., SANDERS I.R. & ERHARDT A. (2000): Arbuscular mycorrhizal fungi influence life history traits of a lepidopteran herbivore. Oecologia 125: 362-369.
GRIME J.P., MACKEY J.M.L, HILLIER S.H. & READ D.J. (1987): Floristic diversity in a model system using experimental microcosms. Nature 328: 420-422.
HARTNETT D.C., HETRICK B.A.D., WILSON G.W.T. & GIBSON D.J. (1993): Mycorrhizal influence on intra- and interspecific neighbour interactions among co-occurring prairie grasses. J. EcoL 8 I: 787-795.
HARTNETT D.C. & WILSON G.W.T. (1999): Mycorrhizae influence plant community structure and diversity in tallgrass prairie. Ecology 80:1187-1195.
HELGASON T., FITTER A.H. & YOUNG J.P.W. (1999): Molecular diversity of arbuscular mycorrhizal fungi colonising Hyacinthoides non-scripta (bluebell) in a seminatural woodland. Molec. Ecol. 8: 659-666.
JOHNSON N.C., GRAHAM J.H. & SMITH F.A. (1997): Functioning of mycorrhizal associations along the mutualism-parasitism continuum. New Phytol. 135: 575-585.
JOHNSON N.C., ZAK D.R., TILMAN D. & PFLEGER F.L. (1991): Dynamics of vesicular-arbuscular mycorrhizae during old field succession. Oecologia 86: 349-358.
LACKIE S.M., GARRIOCK M.L., PETERSON R.L. & BOWLEY S.R. (1987): Influence of host plant on the morphology of the vesicular-arbuscular mycorrhizal fungus Glornus versiforme. Symbiosis 3: 147-158.
MERRYWEATHER J. & FITTER A. (1998a): The arbuscular mycorrhizal fungi ofHyacinthoides non-scripta. 1. Diversity of fungal taxa. New PhytoL 138:117-129.
MERRYWEATHER J. & FITTER A. (1998b): The arbuscular mycorrhizal fungi ofHyacinthoides non-scripta. 2. Seasonal and spatial patterns of fungal populations. New Phytol. 138:131-142.
MOORA M. & ZOBEL M. (1996): Effects ofarbuscular mycorrhiza on inter- and intraspecific competition of two grassland species. Oecologia 108: 79-84.
MORTON J.B., BENTIVENGA S.P. & BEVER J.D. (1996): Discovery, measurement, and interpretation of diversity in symbiotic endomycorrhizal fungi (Glomales, Zygomycetes). Canad. ,I.. Bot. 73, Suppl. 1: $25-$32.
NEWSHAM K.K., FITTER A.H. & WATKINSON A.R. (i 994): Root pathogenic and arbuscular mycorrhizal fimgi determine fecundity of asymptomatic plants in the field. J Ecol. 82:805-814.
NEWSHAM K.K., FITTER A.H. & WATKINSON A.R. (1995): Multi-functionality and biodiversity in arbuscular mycorrhizas. Trends EcoL Evol. 10:407-411.
PHILLIP J.M. & HAYMAN D.S. (1970): Improved procedures for clearing roots and staining parasitic and vesicular-arbuscular mycorrhizal fungi for rapid assessment of infection. Trans. Brit. MycoL Soc. 55: 15-161.
RABATIN S.C. (1979): Seasonal and edaphic variations in vesicular-arbuscular mycorrhizal infection of grasses by Glomus tenuis. New Phytol. 83: 95-102.
RILLIG M.C., FIELD C.B. & ALLEN M.F. (1999): Fungal root colonization responses in natural grasslands after long-term exposure to elevated ~itmospheric CO2. Global Change BioL 5: 577-585.
ROHtMALER W. (1976): Exkursionsflora j~r die Gebiete der DDR und der BRD. Kritischer Band. Volk und Wissen Volkseigener Verlag, Berlin.
SANDERS I.R. & FITTER A.H. (1992): The ecology and functioning of vesicular-arbuscular mycorrhizas in co-existing grassland species I. Seasonal patterns of mycorrhizal occurrence and morphology. New Phytol. 120: 517-524.

262 Smilauer P.
SANDERS I.R., CLAPP J.P. & WIEMKEN A. (1996): The genetic diversity of arbuscular mycorrhizal fungi in natural ecosystems - a key to understanding the ecology and functioning of the mycorrhizal symbiosis. New Phytol. 133: 123-134.
~MILAUER P. & ~M1LAUEROV/k M. (2000): Effect of AM symbiosis exclusion on grassland community composition. Folia Geobot. 35: 13-25.
SMITH F.A., JAKOBSEN 1. 86 SMITH S.E. (2000): Spatial differences in acquisition of soil phosphate between two arbuscular mycorrhizal fungi in symbiosis with Medicago sativa. New Phytol. 147: 357-366.
SMITH F.A. & SMITH S.E. (1997): Structural diversity in (vesicular-)arbuscular mycorrhizal symbioses. Tansley Review No. 96. New Phytol. 137: 373-388.
SMITH S.E. & READ D.J. (1997): Mycorrhizal symbioses. Ed. 2. Academic Press, London. TER BRAAK C.J.F. & ~MILAUER P. (1998): CANOCO reference manual and user's guide to CANOCO for
Windows. Software for canonical community ordination (version 4). Microcomputer Power, Ithaca. THOMSON B.D., ROBSON A.D. & ABBOTT L.K. (1992): The effect of long-term applications of
phosphorus-fertilized on populations of vesicular-arbuscular mycorrhizal fungi in pastures. Austral. J. Agric. Res. 43:1131-1143.
TISSERANT B., BRENAC V., REQUENA N., JEFFRIES P. & DODD J.C. (1998): The detection of Glomus spp. (arbuscular mycorrhizal fungi) forming mycorrhizas in three plants, at different stages of seedling development, using mycorrhiza-specific isozymes. NewPhytol. 138: 225-239.
TITUS J.H. & LEPg J.~. (2000): The response of arbuscular mycorrhizae to fertilization, mowing, and removal of dominant species in a diverse oligotrophic wet meadow. Amer. ,L Bot. 87: 392-401.
VAN DER HEIJDEN M.G.A. et al. (1998a): Mycorrhizal fungal diversity determines plant biodiversity, ecosystem variability and productivity. Nature 396: 69-72.
VAN DER HEIJDEN M.G.A. et al. (1998b): Different arbuscular mycorrhizal fungal species are potential determinants of plant community structure. Ecology 79:2082-2091.
WILSON J.M., TRINICK M.J. & PARKER C.A. (1983): The identification of vesicular-arbuscular mycorrhizal fungi using immunofluorescence. SoilBioL Biochem. 15: 439--445.
YORK MYCORRHIZA RESEARCH GROUP (1999): Techniques in Arbuscular Mycorrhiza Research. York Mycorrhiza Research Group. The University of York, York.
Received 13 September 2000, revision received 26 February 2001, accepted 20 April 2001 Encl. Appendix p. 263

Communities of AM fungi in grassland 263
APPENDIX
Among the spores collected from the experimental plots, approximately seven morphological types were found. Based on the structure of the spore wall (visualized using Melzer's reagent) and also the other spore features (presence of attached subtending hypha, presence of a germination shield), we assign these types to individual genera of AM fungi.
Working label Frequency Description Approximate diameter (~tm)
Glomus sp. - Zv01 numerous orange to red spores, assembled into raspberry-like sporocarps 70, sporocarps 220
Glomus sp. - Zv03 relatively small, dark brown spores occurring in frequent clusters, interconnected by thin hyphae 70
Glomus sp. - Zv06 frequent
Scutellospora sp. - Zv02 infrequent
Scutellospora sp. - Zv07 rare
Acaulospora sp. - Zv04
oval brown spores with attached hypha
continuous with their wall
large oval spores, gold to honey colour, with a subtending hypha attached to their sides
160-170
220-250
large elongate spores, light yellow to amber colour, sometimes a visible germination shield 350-380
quite common
large spherical, amber coloured spores with no attached hypha, but a scar is sometimes visible 230
Acaulospora sp. - Zv05 rare large spherical spores, quite translucent, with an ornamented outer wall 270