screening and selecting arbuscular mycorrhizal fungi for inoculating micropropagated apple...
TRANSCRIPT

ORIGINAL PAPER
Screening and selecting arbuscular mycorrhizal fungi forinoculating micropropagated apple rootstocks in acid soils
Jose Renato Pereira Cavallazzi ÆOsmar Klauberg Filho Æ Sidney Luiz Sturmer ÆPaul T. Rygiewicz Æ Margarida Matos de Mendonca
Received: 8 March 2006 / Accepted: 5 September 2006 / Published online: 17 July 2007
� Springer Science+Business Media B.V. 2007
Abstract Santa Catarina state is the largest pro-
ducer of apples in Brazil. Soils in this region have
low pH and high levels of aluminum and manga-
nese, requiring high inputs of fertilizers and
amendments increasing costs of apple production.
Inoculation of arbuscular mycorrhizal fungi can
improve the establishment of micropropagated
apple plants in such adverse soil conditions. Soil
samples were collected from apple orchards in the
Cacador, Fraiburgo and Sao Joaquim regions to
develop a corn bioassay to identify mycorrhizal
communities with high infectivity. Eleven fungal
species were identified from one Cacador soil with
the highest infectivity. Glomus etunicatum SCT110,
Scutellospora pellucida SCT111, Acaulospora scro-
biculata SCT112 and Scutellospora heterogama
SCT113 were brought into single-species culture
and used in a plant growth and nutrient uptake
experiment using micropropagated apple (Malus
prunifolia), cultivated at three soil pH. Colonization
by fungal isolates significantly affected plant
height, shoot and root dry weights, and root:shoot
ratio. Soil pH also significantly affected all growth
parameters except shoot dry weight. Mycorrhizal
inoculation also significantly altered tissue concen-
trations of P, Zn, Cu, Ca, S, Na, N, K, Fe and Al.
Association with mycorrhizal fungi increased P
concentration and also decreased Al concentrations
in the shoots. Overall, G. etunicatum and S.
pellucida were the most effective isolates to
promote plant growth and nutrient uptake.
Inoculation of apple rootstock with selected fun-
gal isolates during the acclimatization stage
represents a useful strategy for producing micro-
propagated apples that can withstand acidic soil
conditions.
Keywords Acid soils � Aluminum � Brazil �Micropropagated apple � Mycorrhizae �Plant–microorganisms interaction �Nutrient content � Plant growth
J. R. P. Cavallazzi � M. M. de Mendonca (&)
Departamento de Microbiologia e Parasitologia,
Universidade Federal de Santa Catarina, Cx. P. 476,
Florianopolis, SC, 88040-900, Brazil
e-mail: [email protected]
O. K. Filho
Departamento de Solos, Universidade do Estado de Santa
Catarina, Cx. P. 281, Lages, SC, 88520-000, Brazil
S. L. Sturmer
Departamento de Ciencias Naturais, Universidade
Regional de Blumenau, Cx. P. 1507,
Blumenau, SC, 89010-971, Brazil
P. T. Rygiewicz
Western Ecology Division, National Health and
Environmental Effects Research Laboratory, U.S.
Environmental Protection Agency, Corvallis,
OR, 97333, USA
123
Plant Cell Tiss Organ Cult (2007) 90:117–129
DOI 10.1007/s11240-006-9163-6

Introduction
The State of Santa Catarina is the largest producer of
apples in Brazil, with an annual production of
750,000 tons, representing ca 53% of the country’s
production (ICEPA/SC 1999). In the 1970s, apples
ranked as the third most important agricultural
product in the country. Consequently, areas support-
ing apple production were greatly increased due to
governmental subsidies. This was especially the case
in Santa Catarina. Brazil reached self-sufficiency in
apple production in 1990, and today produces an
excess of apples, which are exported to Europe and
North America. During the period of rapid expansion
of apple orchards, demand for rootstock was higher
than production rates. As a result, reproductive
material of inferior health was used which contrib-
uted to high rates of viral diseases that caused
reduced orchard productivity leading to extensive
replanting of orchards. In Santa Catarina, most
orchards and apple nurseries are located on soils
with low pH and high aluminum levels (Basso and
Suzuki 1992). Plant growth in these soils is limited by
several factors including high concentrations of Mn
and H and low solubilities of P, Mg and Ca
(Marschner 1991). As a consequence, large inputs
of fertilizers and lime are required, making apples an
agricultural product in the state having among the
highest costs of production.
It is well known that the endomycorrhizal associ-
ation, established by arbuscular mycorrhizal fungi
(AMF) colonizing roots, promotes growth and nutri-
tion of plants by increasing uptake of minerals from
soil and protects plants against potentially phytotoxic
elements, e.g., Cd and Pb (Weissenhorn and Leyal
1995; Weissenhorn et al. 1995). This type of
mycorrhizal fungi occurs naturally in soils and
colonizes the roots of the vast majority of plant
species. Colonization by AMF in acidic soils
increases plant biomass production (O’Donnell
et al. 1992; Soedarjo and Habte 1993) and uptake
of P (Habte and Soedarjo 1996), K and Mg (Borie
and Rubio 1999). The endomycorrhizal association
also decreases uptake of toxic elements such as Cd
(El-Kherbawy et al. 1989), Mn (Arines et al. 1989)
and Al (Borie and Rubio 1999), reducing plant tissue
concentrations of these elements.
Several reports have shown that the arbuscular
endomycorrhizal association is more efficient in
promoting growth of apple plants than is applying
fertilizers only. Inoculation with AMF isolates
resulted in mean increases in plant height and
biomass of 735 and 1,161%, respectively, compared
with non-inoculated plants (Geddeda et al. 1984).
The authors also observed a negative correlation
between soil P levels and root colonization rates,
reflecting the consequences of adding fertilizers to
the soil. In a P fertilizer experiment, Miller et al.
(1985) inoculated apple plants with one of six
isolates of Glomus or an isolate of Gigaspora, and
found that only the plants growing in soils not
receiving the P fertilizer treatment were responsive
to the mycorrhizal association. Plenchette et al.
(1981) inoculated apple (Malus pumila) in sterile
soils (pH 4.7) and demonstrated that mycorrhizal
plants were significantly taller and more uniform in
height compared with non-inoculated plants. Dry
mass of the stem and leaves, and the leaf surface
area per plant of the mycorrhizal plants increased
335, 204 and 153%, respectively, compared with
non-inoculated plants.
In vitro micropropagation can be used to
produce pathogen-free and, especially, virus-free
apple plantlets while also achieving increases in
scale and speed of production (Grattapaglia and
Machado 1991). However, several problems limit
the widespread use of micropropagated plants.
Upon transfer of plantlets from in vitro to post
vitro conditions, high mortality frequently occurs
since the photosynthetic capability needs to im-
prove and root systems need to develop further
(George 1996). This conditioning of the plants to
new growth conditions leads to an extended
weaning stage that is often accompanied by high
losses of plantlets, and large inputs of fertilizers,
pesticides and other chemicals. Additionally, woody
plants can pose a particular problem in that the root
systems produced in vitro may not develop further
leading to plantlet death or severe reductions in
growth (George 1996). This characteristic might
limit commercial production of some genotypes of
apples. Sterilized substrates used during plantlet
acclimation impair establishment of the symbiotic
association, causing problems for survival and
118 Plant Cell Tiss Organ Cult (2007) 90:117–129
123

growth of seedlings after transplant (Ravolanirina
et al. 1989). Branzanti et al. (1992) observed that
AMF colonization yielded micropropagated apple
plants significantly more uniform and taller than
those plants growing in fertilized substrates.
Granger et al. (1983) observed that clones of apple
Malling 7 micropropagated and inoculated with
Glomus epigaeum produced higher biomass and had
higher P and Cu concentrations when compared
with non-mycorrhizal plants. The endomycorrhizal
association also increased tolerance of microprop-
agated apples to several stresses during rooting and
acclimation stages, reducing the time necessary to
produce seedlings (Uosukainen and Vestberg 1994).
However, the majority of studies involving micro-
propagated apples and AMF used fungal isolates
originating from areas with varying soil conditions
supporting different plant communities, paying little
attention to controlling or assessing soil conditions
such as pH.
Several factors influence the efficiency of the
endomycorrhizal association, e.g., genotype of the
fungal isolate (Uosukainen and Vestberg 1994),
plant host species (Branzanti et al. 1992; Guillemin
et al. 1992) and soil physical, chemical and biotic
properties (Habte and Soedarjo 1996). Therefore,
targeted and specific screening and selecting of
AMF isolates should precede large-scale inoculation
programs and need to consider the host plant
species and the specific environmental conditions
under which the inoculated plants will be grown.
The work we report here is part of a larger project
which had the objective of selecting appropriate
technologies to produce healthy propagated apple
rootstock that would be adapted to the acidic soil
conditions found in Santa Catarina. A multidisci-
plinary approach was undertaken, encompassing
microbiological- and plant-based strategies. The
specific aim of the research was to integrate the
use of AMF isolates and the techniques of micro-
propagation to produce apple rootstock that would
tolerate transplanting into acidic soil conditions. If
successful, the integrated approach could lead to
producing large amounts of plant material that
would be readily available to nurseries. Specifi-
cally, we strove to define a strategy to recover
native AMF isolates from orchard soils and assess
the inoculation of the most competitive species/
isolates on the growth of micropropagated apple
rootstocks, using sterilized substrates adjusted to
pH values likely to be found in large-scale
production operations.
Methods and materials
Sampling sites
The study was conducted in three areas producing
apple in the plateau region (900–1,500 m) in the
central part of the state of Santa Catarina, South
Brazil (Table 1). The climate is Koeppen type Cfb
(i.e., mesothermic humid with cool summers and
harsh winters), with precipitation evenly distributed
throughout the year in the form of rain (annual total:
1,400–1,800 mm). Mean annual temperature ranges
from 14 to 18�C, and relative humidity ranges from
75 to 85%. Soils are often acidic and have high
aluminum content.
In each of the two orchards in each of the three
apple producing areas, ten trees were selected from
under which four soil samples (500 g each) were
taken and combined, resulting in ten samples per
orchard. The four sub-samples were collected from
the soil surface to 20 cm depth, at 20–40 cm from the
stem with one sub-sample coming from each of the
four cardinal directions. Orchards were maintained
free of weeds and, therefore, no other plants were
found growing near each sampled apple tree. Soil was
collected exclusively from around apple feeder roots
that were detected by following a main root. Each
pooled sample was placed in a plastic bag and then
stored at 4�C before processing.
Mycorrhizal infectivity
A corn bioassay was used to estimate mycorrhizal
infectivity of the soil from each orchard (Moorman
and Reeves 1979). Soil samples from each orchard
were homogenized, mixed (1:1, vol:vol) with
washed, sterilized sand and then the mixture placed
in 0.5 l pots. One 6-day-old seedling of hybrid corn
(Agroeste—AS 532), pre-germinated in sterilized
sand was transplanted to each pot. Six pots were set
up for each orchard and sampled 40 days after
transplanting. Corn roots were gently separated from
the soil, washed and stained according to Koske and
Plant Cell Tiss Organ Cult (2007) 90:117–129 119
123
Gemma (1989). Roots were scored for percent
mycorrhizal colonization following Giovannetti and
Mosse (1980) and the value used as an estimate of
mycorrhizal infectivity in each orchard.
AMF species diversity and establishing fungal
isolates
Only soil samples from the orchard having the
greatest mycorrhizal infectivity were used to assess
species diversity associated with apples and to
establish fungal isolates in pure culture. Spores were
recovered from 50 g of soil from each of the ten
pooled samples by wet sieving (Gerdemann and
Nicolson 1963) followed by centrifugation in a 50%
sucrose solution. Spores were grouped into morpho-
types, mounted in PVLG (polyvinyl-lacto-glycerol)
and Melzer’s reagent and then identified either
according to original species description protocols
or reference cultures described on the International
Culture Collection of Arbuscular Mycorrhizal Fungi
web site (http://invam.caf.wvu.edu).
Trap cultures were set up according to Stutz and
Morton (1996), by mixing field soil (1:1, vol:vol)
with sterilized sand. The mixture was placed in 1.5 kg
pots; seeds (50–60) of Sorghum bicolor L. were sown
in each pot and then covered with 1 cm sterilized
vermiculite. After 4 months in the greenhouse, three
100 g sub-samples of each pot were collected to
isolate and identify spores as described above.
Pure cultures of AMF were obtained following
procedures of Morton et al. (1993). Fresh spores
obtained from trap cultures were separated into
morphotypes using a dissecting microscope. Spores
of each morphotype were inoculated onto the root
systems of germinated Sorghum bicolor and Pasp-
alum notatum. Host plants were grown in a sterilized
mixture of sand and a red–yellow spodosol (2:1,
vol:vol), maintained in the greenhouse and watered
with distilled water. After 4 months, watering was
stopped, and plants and the planting mix were
allowed to dry in situ for 4–10 days. Spores were
extracted from the planting mix as described above
and fungal cultures were checked for purity. Suffi-
cient inocula, derived from the single-species cul-
tures, were produced to use in the plant growth
experiment. The volumes of inocula were increased
by mixing the soil inoculum from each single culture
with sterilized substrate (sand: red–yellow spodosol,
1:1, vol:vol). Each inoculum mixture was placed in
1.5 kg pots, seeded with Sorghum and Paspalum, and
maintained in the greenhouse. Pots filled only with
sterilized substrate and seeded with both hosts as
described above served as the non-inoculated control
treatment. After 4 months, plant tops were discarded
and the soil associated with the root balls was
chopped, homogenized and stored at 4�C.
Efficiency of AMF isolates in acid soils
Efficiency of AMF isolates was studied using the
Marubakaido rootstock of apple (Malus prunifolia)
growing in acidic soil conditions. Micropropagated
plantlets were produced in the Plant Morphogenesis
Laboratory at the Universidade Federal de Santa
Catarina from apical meristems grown in modified
Murashige–Skoog medium, following the methodol-
ogy of Nunes et al. (1991).
Plants were transferred from in vitro conditions to
Styrofoam trays containing 60 ml cells filled with a
mixture of sterilized carbonized rice shells and
vermiculite (1:1, vol:vol). Styrofoam trays containing
the micropropagated plants were placed inside a
plastic tray holding ca 1 cm of water. A glass cover
with a 2-cm opening on one side was placed over
both the Styrofoam and plastic trays. Plants were
Table 1. Location and
characteristics of apple
orchards sampled
Region Location Age (year) Apple cultivar Code
Cacador EPAGRI Experimental
Station
3 Fuji and Gala CA1
18 Fuji CA2
Sao Joaquim EPAGRI Experimental
Station
17 Rome Beauty
and Fuji
SJ1
4 Fuji SJ2
Fraiburgo Fischer Company 12 Belgolden FR1
7 Fuji FR2
120 Plant Cell Tiss Organ Cult (2007) 90:117–129
123

maintained in a growth chamber (27�C) until roots
developed.
After 20 days, plants with roots were individually
transplanted into the 120 ml cells of another Styro-
foam tray that contained a sterilized mixture of
soil:sand:feedstock (1:1:1, vol:vol:vol). This mixture
had the following characteristics: pH 6.0,
P > 50 mg kg�1, 9.8% organic matter, and 7.5 and
2.8 cmolc kg�1 of Ca and Mg, respectively. During
the transplanting procedure, soil inoculum (spores,
hyphae, pieces of colonized roots) derived from one
single-species isolate of AMF was added to each cell
as a 30-cm layer between two layers of the sterilized
planting mixture. Control plants received the same
amount of substrate without AMF. Plants were grown
in greenhouse conditions for 30 days and were
irrigated only with distilled water. When plants
reached 6–10 cm height, the mycorrhizal efficiency
experiment was initiated and the plants and planting
substrate were transferred to 1.5 kg pots containing
soil with different pH.
The planting substrate was prepared by mixing a
latossolo bruno alico (EMBRAPA 1999) from the
Fraiburgo region, with a native pH of 4.0. This soil
was mixed (1:1, vol:vol) with washed, sterilized sand.
The mixture was divided into three portions: one was
kept at its native pH and the other two portions were
adjusted to either pH 5.0 or 6.0 by adding CaCO3
using incubation with neutralizing curves as de-
scribed by Melo (1985). Substrate was sterilized
(121�C, 1 h) twice with a 24-h interval between
autoclaving sessions. No fertilizer was added during
the experiment and soil chemical characteristics of
the three substrates are presented in Table 2. After
transplanting, a layer of sterilized vermiculite was
added to the substrate surface.
A completely randomized design was used con-
sisting of three pH levels (4.0, 5.0 and 6.0) and five
mycorrhizal inoculation treatments (four single-spe-
cies AMF isolates and the non-inoculated control)
with eight replicates per treatment combination.
Plants were maintained under greenhouse conditions
and watered only with distilled water to field
capacity. After 70 days, plant height was measured
and shoots were separated from roots. Mycorrhizal
colonization was assessed using 0.3–0.5 g roots that
were randomly sampled, stained according to Koske
and Gemma (1989) and then scored as percent root
colonization following Giovanetti and Mosse (1980).
Shoots and the remaining root fractions were oven
dried (60�C, 3 days) and weighed to determine dry
mass. The equivalent amounts of root dry mass
removed to determine mycorrhizal colonization were
added to the mass measured for the remaining root
fractions to calculate total root weights. Total dry
mass of mycorrhizal and non-mycorrhizal plants was
used to calculate relative mycorrhizal dependency
(RMD) according to Plenchette et al. (1983):
RMD = [(dry mass myc plants – dry mass non-myc
plants)/dry mass myc plants] · 100. Mineral concen-
trations of N, P, K, Ca, Mg, S, Cu, Zn, Fe, Mn, Na, B
and Al were measured using the shoots according to
Tedesco et al. (1995).
Statistical analyses
All dependent variable data were checked for homo-
geneity of variance according to the test of Burtlett.
Mineral concentrations of N, P, K, Ca, Mg and S
were square root transformed, and percent root
colonization for the corn bioassay and apple plants
were square root + 1 transformed. Root colonization
for the corn bioassay was analyzed by one-way
ANOVA, while colonization of apple plants in the
efficiency experiment was analyzed by two-way
ANOVA. Means were separated by the least signif-
icance difference test (P = 0.05). Statistical analyses
were performed using STATGRAPHICS1 Plus,
version 2.1.
Results
Mycorrhizal inoculum potential of apple orchards
After 40 days, mycorrhizal colonization of corn
growing in soil from Cacador (CA1) was significantly
Table 2. Soil chemical properties of the native soil (pH 4.0)
and limed soil (pH 5.0 and 6.0) used in the AMF efficiency
experiment
Soil pH P K OM Al Ca Mg
mg kg�1 (%) cmolc kg�1
4.0 5.2 99 6.4 2.7 2.0 1.0
5.0 2.7 78 6.0 0.3 2.7 1.6
6.0 4.2 72 5.6 – 4.7 2.0
Plant Cell Tiss Organ Cult (2007) 90:117–129 121
123
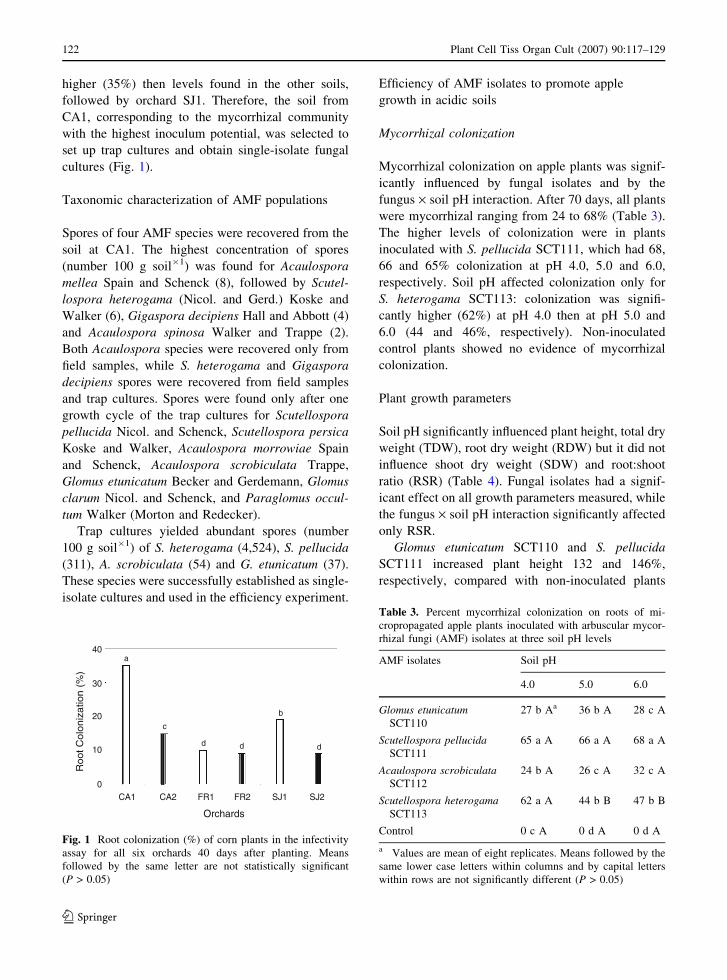
higher (35%) then levels found in the other soils,
followed by orchard SJ1. Therefore, the soil from
CA1, corresponding to the mycorrhizal community
with the highest inoculum potential, was selected to
set up trap cultures and obtain single-isolate fungal
cultures (Fig. 1).
Taxonomic characterization of AMF populations
Spores of four AMF species were recovered from the
soil at CA1. The highest concentration of spores
(number 100 g soil�1) was found for Acaulospora
mellea Spain and Schenck (8), followed by Scutel-
lospora heterogama (Nicol. and Gerd.) Koske and
Walker (6), Gigaspora decipiens Hall and Abbott (4)
and Acaulospora spinosa Walker and Trappe (2).
Both Acaulospora species were recovered only from
field samples, while S. heterogama and Gigaspora
decipiens spores were recovered from field samples
and trap cultures. Spores were found only after one
growth cycle of the trap cultures for Scutellospora
pellucida Nicol. and Schenck, Scutellospora persica
Koske and Walker, Acaulospora morrowiae Spain
and Schenck, Acaulospora scrobiculata Trappe,
Glomus etunicatum Becker and Gerdemann, Glomus
clarum Nicol. and Schenck, and Paraglomus occul-
tum Walker (Morton and Redecker).
Trap cultures yielded abundant spores (number
100 g soil�1) of S. heterogama (4,524), S. pellucida
(311), A. scrobiculata (54) and G. etunicatum (37).
These species were successfully established as single-
isolate cultures and used in the efficiency experiment.
Efficiency of AMF isolates to promote apple
growth in acidic soils
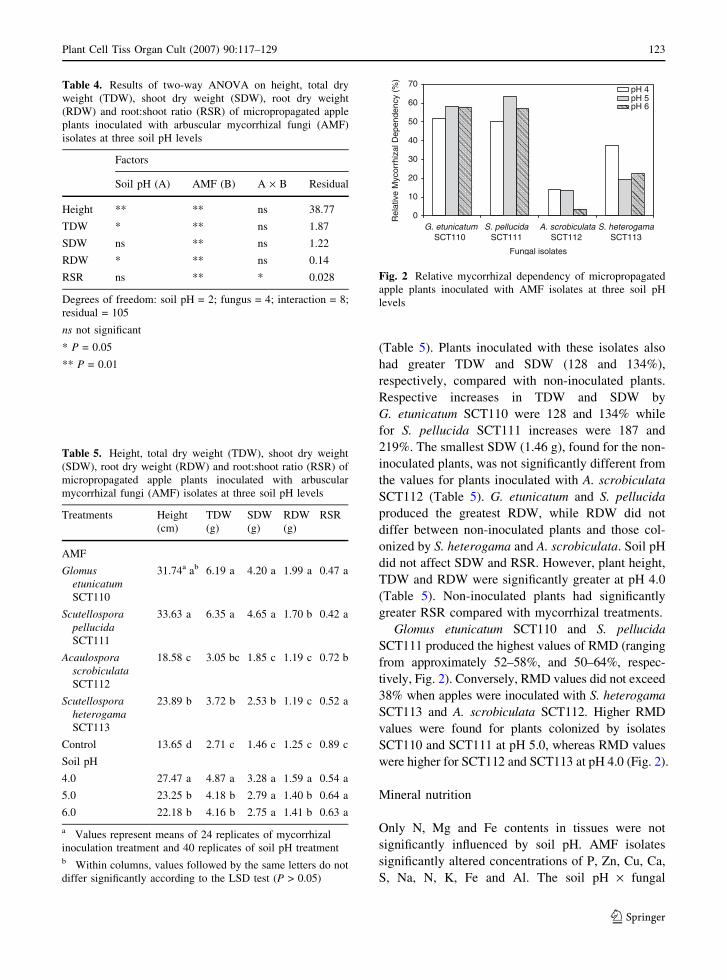
Mycorrhizal colonization
Mycorrhizal colonization on apple plants was signif-
icantly influenced by fungal isolates and by the
fungus · soil pH interaction. After 70 days, all plants
were mycorrhizal ranging from 24 to 68% (Table 3).
The higher levels of colonization were in plants
inoculated with S. pellucida SCT111, which had 68,
66 and 65% colonization at pH 4.0, 5.0 and 6.0,
respectively. Soil pH affected colonization only for
S. heterogama SCT113: colonization was signifi-
cantly higher (62%) at pH 4.0 then at pH 5.0 and
6.0 (44 and 46%, respectively). Non-inoculated
control plants showed no evidence of mycorrhizal
colonization.
Plant growth parameters
Soil pH significantly influenced plant height, total dry
weight (TDW), root dry weight (RDW) but it did not
influence shoot dry weight (SDW) and root:shoot
ratio (RSR) (Table 4). Fungal isolates had a signif-
icant effect on all growth parameters measured, while
the fungus · soil pH interaction significantly affected
only RSR.
Glomus etunicatum SCT110 and S. pellucida
SCT111 increased plant height 132 and 146%,
respectively, compared with non-inoculated plants
0CA1 CA2 FR1 FR2 SJ1 SJ2
10
20
30
40
Roo
t Col
oniz
atio
n (%
)
Orchards
a
c
d d
b
d
Fig. 1 Root colonization (%) of corn plants in the infectivity
assay for all six orchards 40 days after planting. Means
followed by the same letter are not statistically significant
(P > 0.05)
Table 3. Percent mycorrhizal colonization on roots of mi-
cropropagated apple plants inoculated with arbuscular mycor-
rhizal fungi (AMF) isolates at three soil pH levels
AMF isolates Soil pH
4.0 5.0 6.0
Glomus etunicatumSCT110
27 b Aa 36 b A 28 c A
Scutellospora pellucidaSCT111
65 a A 66 a A 68 a A
Acaulospora scrobiculataSCT112
24 b A 26 c A 32 c A
Scutellospora heterogamaSCT113
62 a A 44 b B 47 b B
Control 0 c A 0 d A 0 d A
a Values are mean of eight replicates. Means followed by the
same lower case letters within columns and by capital letters
within rows are not significantly different (P > 0.05)
122 Plant Cell Tiss Organ Cult (2007) 90:117–129
123
(Table 5). Plants inoculated with these isolates also
had greater TDW and SDW (128 and 134%),
respectively, compared with non-inoculated plants.
Respective increases in TDW and SDW by
G. etunicatum SCT110 were 128 and 134% while
for S. pellucida SCT111 increases were 187 and
219%. The smallest SDW (1.46 g), found for the non-
inoculated plants, was not significantly different from
the values for plants inoculated with A. scrobiculata
SCT112 (Table 5). G. etunicatum and S. pellucida
produced the greatest RDW, while RDW did not
differ between non-inoculated plants and those col-
onized by S. heterogama and A. scrobiculata. Soil pH
did not affect SDW and RSR. However, plant height,
TDW and RDW were significantly greater at pH 4.0
(Table 5). Non-inoculated plants had significantly
greater RSR compared with mycorrhizal treatments.
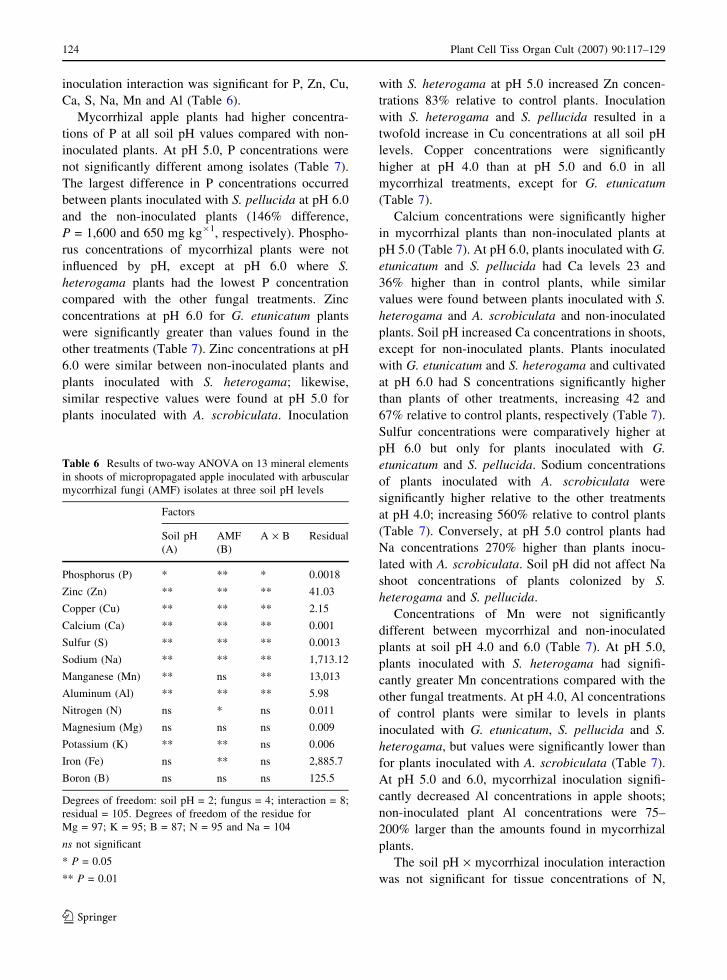
Glomus etunicatum SCT110 and S. pellucida
SCT111 produced the highest values of RMD (ranging
from approximately 52–58%, and 50–64%, respec-
tively, Fig. 2). Conversely, RMD values did not exceed
38% when apples were inoculated with S. heterogama
SCT113 and A. scrobiculata SCT112. Higher RMD
values were found for plants colonized by isolates
SCT110 and SCT111 at pH 5.0, whereas RMD values
were higher for SCT112 and SCT113 at pH 4.0 (Fig. 2).
Mineral nutrition
Only N, Mg and Fe contents in tissues were not
significantly influenced by soil pH. AMF isolates
significantly altered concentrations of P, Zn, Cu, Ca,
S, Na, N, K, Fe and Al. The soil pH · fungal
Table 4. Results of two-way ANOVA on height, total dry
weight (TDW), shoot dry weight (SDW), root dry weight
(RDW) and root:shoot ratio (RSR) of micropropagated apple
plants inoculated with arbuscular mycorrhizal fungi (AMF)
isolates at three soil pH levels
Factors
Soil pH (A) AMF (B) A · B Residual
Height ** ** ns 38.77
TDW * ** ns 1.87
SDW ns ** ns 1.22
RDW * ** ns 0.14
RSR ns ** * 0.028
Degrees of freedom: soil pH = 2; fungus = 4; interaction = 8;
residual = 105
ns not significant
* P = 0.05
** P = 0.01
Table 5. Height, total dry weight (TDW), shoot dry weight
(SDW), root dry weight (RDW) and root:shoot ratio (RSR) of
micropropagated apple plants inoculated with arbuscular
mycorrhizal fungi (AMF) isolates at three soil pH levels
Treatments Height
(cm)
TDW
(g)
SDW
(g)
RDW
(g)
RSR
AMF
GlomusetunicatumSCT110
31.74a ab 6.19 a 4.20 a 1.99 a 0.47 a
ScutellosporapellucidaSCT111
33.63 a 6.35 a 4.65 a 1.70 b 0.42 a
AcaulosporascrobiculataSCT112
18.58 c 3.05 bc 1.85 c 1.19 c 0.72 b
ScutellosporaheterogamaSCT113
23.89 b 3.72 b 2.53 b 1.19 c 0.52 a
Control 13.65 d 2.71 c 1.46 c 1.25 c 0.89 c
Soil pH
4.0 27.47 a 4.87 a 3.28 a 1.59 a 0.54 a
5.0 23.25 b 4.18 b 2.79 a 1.40 b 0.64 a
6.0 22.18 b 4.16 b 2.75 a 1.41 b 0.63 a
a Values represent means of 24 replicates of mycorrhizal
inoculation treatment and 40 replicates of soil pH treatmentb Within columns, values followed by the same letters do not
differ significantly according to the LSD test (P > 0.05)
G. etunicatumSCT110
S. pellucidaSCT111
Fungal isolates
0
10
20
30
40
50
60
70pH 4pH 5pH 6
Rel
ativ
e M
ycor
rhiz
al D
epen
denc
y (%
)
A. scrobiculataSCT112
S. heterogama SCT113
Fig. 2 Relative mycorrhizal dependency of micropropagated
apple plants inoculated with AMF isolates at three soil pH
levels
Plant Cell Tiss Organ Cult (2007) 90:117–129 123
123
inoculation interaction was significant for P, Zn, Cu,
Ca, S, Na, Mn and Al (Table 6).
Mycorrhizal apple plants had higher concentra-
tions of P at all soil pH values compared with non-
inoculated plants. At pH 5.0, P concentrations were
not significantly different among isolates (Table 7).
The largest difference in P concentrations occurred
between plants inoculated with S. pellucida at pH 6.0
and the non-inoculated plants (146% difference,
P = 1,600 and 650 mg kg�1, respectively). Phospho-
rus concentrations of mycorrhizal plants were not
influenced by pH, except at pH 6.0 where S.
heterogama plants had the lowest P concentration
compared with the other fungal treatments. Zinc
concentrations at pH 6.0 for G. etunicatum plants
were significantly greater than values found in the
other treatments (Table 7). Zinc concentrations at pH
6.0 were similar between non-inoculated plants and
plants inoculated with S. heterogama; likewise,
similar respective values were found at pH 5.0 for
plants inoculated with A. scrobiculata. Inoculation
with S. heterogama at pH 5.0 increased Zn concen-
trations 83% relative to control plants. Inoculation
with S. heterogama and S. pellucida resulted in a
twofold increase in Cu concentrations at all soil pH
levels. Copper concentrations were significantly
higher at pH 4.0 than at pH 5.0 and 6.0 in all
mycorrhizal treatments, except for G. etunicatum
(Table 7).
Calcium concentrations were significantly higher
in mycorrhizal plants than non-inoculated plants at
pH 5.0 (Table 7). At pH 6.0, plants inoculated with G.
etunicatum and S. pellucida had Ca levels 23 and
36% higher than in control plants, while similar
values were found between plants inoculated with S.
heterogama and A. scrobiculata and non-inoculated
plants. Soil pH increased Ca concentrations in shoots,
except for non-inoculated plants. Plants inoculated
with G. etunicatum and S. heterogama and cultivated
at pH 6.0 had S concentrations significantly higher
than plants of other treatments, increasing 42 and
67% relative to control plants, respectively (Table 7).
Sulfur concentrations were comparatively higher at
pH 6.0 but only for plants inoculated with G.
etunicatum and S. pellucida. Sodium concentrations
of plants inoculated with A. scrobiculata were
significantly higher relative to the other treatments
at pH 4.0; increasing 560% relative to control plants
(Table 7). Conversely, at pH 5.0 control plants had
Na concentrations 270% higher than plants inocu-
lated with A. scrobiculata. Soil pH did not affect Na
shoot concentrations of plants colonized by S.
heterogama and S. pellucida.
Concentrations of Mn were not significantly
different between mycorrhizal and non-inoculated
plants at soil pH 4.0 and 6.0 (Table 7). At pH 5.0,
plants inoculated with S. heterogama had signifi-
cantly greater Mn concentrations compared with the
other fungal treatments. At pH 4.0, Al concentrations
of control plants were similar to levels in plants
inoculated with G. etunicatum, S. pellucida and S.
heterogama, but values were significantly lower than
for plants inoculated with A. scrobiculata (Table 7).
At pH 5.0 and 6.0, mycorrhizal inoculation signifi-
cantly decreased Al concentrations in apple shoots;
non-inoculated plant Al concentrations were 75–
200% larger than the amounts found in mycorrhizal
plants.
The soil pH · mycorrhizal inoculation interaction
was not significant for tissue concentrations of N,
Table 6 Results of two-way ANOVA on 13 mineral elements
in shoots of micropropagated apple inoculated with arbuscular
mycorrhizal fungi (AMF) isolates at three soil pH levels
Factors
Soil pH
(A)
AMF
(B)
A · B Residual
Phosphorus (P) * ** * 0.0018
Zinc (Zn) ** ** ** 41.03
Copper (Cu) ** ** ** 2.15
Calcium (Ca) ** ** ** 0.001
Sulfur (S) ** ** ** 0.0013
Sodium (Na) ** ** ** 1,713.12
Manganese (Mn) ** ns ** 13,013
Aluminum (Al) ** ** ** 5.98
Nitrogen (N) ns * ns 0.011
Magnesium (Mg) ns ns ns 0.009
Potassium (K) ** ** ns 0.006
Iron (Fe) ns ** ns 2,885.7
Boron (B) ns ns ns 125.5
Degrees of freedom: soil pH = 2; fungus = 4; interaction = 8;
residual = 105. Degrees of freedom of the residue for
Mg = 97; K = 95; B = 87; N = 95 and Na = 104
ns not significant
* P = 0.05
** P = 0.01
124 Plant Cell Tiss Organ Cult (2007) 90:117–129
123
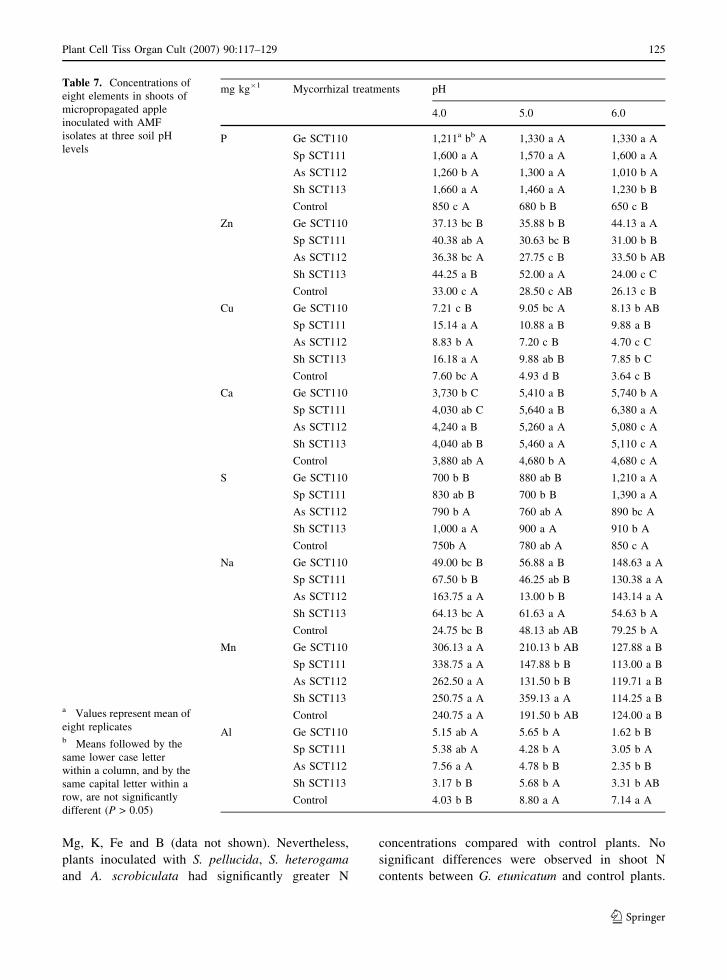
Mg, K, Fe and B (data not shown). Nevertheless,
plants inoculated with S. pellucida, S. heterogama
and A. scrobiculata had significantly greater N
concentrations compared with control plants. No
significant differences were observed in shoot N
contents between G. etunicatum and control plants.
Table 7. Concentrations of
eight elements in shoots of
micropropagated apple
inoculated with AMF
isolates at three soil pH
levels
a Values represent mean of
eight replicatesb Means followed by the
same lower case letter
within a column, and by the
same capital letter within a
row, are not significantly
different (P > 0.05)
mg kg�1 Mycorrhizal treatments pH
4.0 5.0 6.0
P Ge SCT110 1,211a bb A 1,330 a A 1,330 a A
Sp SCT111 1,600 a A 1,570 a A 1,600 a A
As SCT112 1,260 b A 1,300 a A 1,010 b A
Sh SCT113 1,660 a A 1,460 a A 1,230 b B
Control 850 c A 680 b B 650 c B
Zn Ge SCT110 37.13 bc B 35.88 b B 44.13 a A
Sp SCT111 40.38 ab A 30.63 bc B 31.00 b B
As SCT112 36.38 bc A 27.75 c B 33.50 b AB
Sh SCT113 44.25 a B 52.00 a A 24.00 c C
Control 33.00 c A 28.50 c AB 26.13 c B
Cu Ge SCT110 7.21 c B 9.05 bc A 8.13 b AB
Sp SCT111 15.14 a A 10.88 a B 9.88 a B
As SCT112 8.83 b A 7.20 c B 4.70 c C
Sh SCT113 16.18 a A 9.88 ab B 7.85 b C
Control 7.60 bc A 4.93 d B 3.64 c B
Ca Ge SCT110 3,730 b C 5,410 a B 5,740 b A
Sp SCT111 4,030 ab C 5,640 a B 6,380 a A
As SCT112 4,240 a B 5,260 a A 5,080 c A
Sh SCT113 4,040 ab B 5,460 a A 5,110 c A
Control 3,880 ab A 4,680 b A 4,680 c A
S Ge SCT110 700 b B 880 ab B 1,210 a A
Sp SCT111 830 ab B 700 b B 1,390 a A
As SCT112 790 b A 760 ab A 890 bc A
Sh SCT113 1,000 a A 900 a A 910 b A
Control 750b A 780 ab A 850 c A
Na Ge SCT110 49.00 bc B 56.88 a B 148.63 a A
Sp SCT111 67.50 b B 46.25 ab B 130.38 a A
As SCT112 163.75 a A 13.00 b B 143.14 a A
Sh SCT113 64.13 bc A 61.63 a A 54.63 b A
Control 24.75 bc B 48.13 ab AB 79.25 b A
Mn Ge SCT110 306.13 a A 210.13 b AB 127.88 a B
Sp SCT111 338.75 a A 147.88 b B 113.00 a B
As SCT112 262.50 a A 131.50 b B 119.71 a B
Sh SCT113 250.75 a A 359.13 a A 114.25 a B
Control 240.75 a A 191.50 b AB 124.00 a B
Al Ge SCT110 5.15 ab A 5.65 b A 1.62 b B
Sp SCT111 5.38 ab A 4.28 b A 3.05 b A
As SCT112 7.56 a A 4.78 b B 2.35 b B
Sh SCT113 3.17 b B 5.68 b A 3.31 b AB
Control 4.03 b B 8.80 a A 7.14 a A
Plant Cell Tiss Organ Cult (2007) 90:117–129 125
123

Concentrations of Mg and B were not affected by
mycorrhizal inoculation. Relative to non-inoculated
plants, inoculation with AMF isolates increased K
concentrations and decreased Fe concentrations in
shoots. Soil pH did not influence shoot concentrations
of N, Mg and Fe.
Discussion
The corn bioassay effectively identified orchard
soils having the highest mycorrhizal inoculum
potential. After 40 days, colonization on corn was
37% in orchard CA1 (despite the low spore count,
20 spores 100 g soil�1) compared with 10–19% in
the other orchard soils. Considering that inoculum
potential depends on fungal infectivity and propa-
gule density, AMF in CA1 were highly infective
and quickly colonized roots, a desirable selection
characteristic in screening processes (Abbott and
Robson 1981). Due to low spore counts in this soil,
mycorrhizal infectivity was caused mainly by other
fungal propagules, e.g., pieces of colonized roots
and extraradical mycelia. The low spore densities
in apple plantations were not expected as Siqueira
et al. (1989) recovered twice the number of spores
in agrosystems compared with natural systems, and
Miller et al. (1985) found spore densities in
orchards in the USA ranging from 62 to 2,150
per 100 g soil�1.
Low spore counts partially compromised identify-
ing AMF species and hindered establishing single-
species cultures. Trapping yielded seven additional
species not found sporulating in the field, indicating
that absence of sporulation should not suggest
absence of fungus (Stutz and Morton 1996). Trapping
is efficient, especially in apple orchards: 17 of 41
AMF species detected by Miller et al. (1985) were
recovered only by trap culture. In this study, the most
efficient isolates, G. etunicatum and S. pellucida,
were not detected from spores in field samples, but
found by trapping. It is possible that in the field, both
fungi allocated resources to structures enhancing
fungal efficiency such as internal and external
mycelia and arbuscules (Graham et al. 1982; Smith
and Read 1997). Sporulation can depend on the plant
species used for trapping (Bever et al. 1996);
therefore, the corn we used may have differentially
stimulated AMF species.
Increasing soil pH from 4.0 to 5.0 and 6.0 did not
influence root colonization, except for S. heterogama
where colonization declined as pH increased sug-
gesting that the isolates were adapted to abrupt
changes in pH. This plasticity may be attributable to
the long-term management of soil in the apple
orchards. In Cacador, native soils are acidic and
were limed prior to establishing orchards perhaps
promoting increased resiliency of the mycorrhizal
community to changes in soil pH as found by Mamo
and Killham (1987). According to Sieverding (1991),
sensitivity of AMF isolates to liming might be a
characteristic of the fungal species. In our study, the
highest rates of colonization was found for S.
pellucida, supporting the possibility that this fungus
invests resources preferentially to mycelial produc-
tion rather than to sporulation.
Inoculation of micropropagated apple in the post
vitro stage stimulated shoot height. After 70 days,
plants inoculated with G. etunicatum and S. pellucida
were taller than those colonized by other isolates. The
mean daily growth increment was 0.045 cm for the
first 28 days, and 0.26 cm from 29 to 42 days (an
increase of 478%). The maximal mean daily growth
rate of 0.52 cm occurred after 70 days (data not
shown). Fortuna et al. (1992) suggest that initial slow
growth rates might be related to either the: (1) lag
phase of fungal growth, reducing root colonization, or
(2) cessation of apical growth of the micropropagated
plants caused by transplanting. In this study, inocu-
lum potential was not measured for all isolates, and
only spore numbers were recorded per 100 g soil�1
(37 for G. etunicatum, 311 for S. pellucida, 4,524 for
S. heterogama and 54 for A. scrobiculata). Appar-
ently, spore numbers did not limit initiation of root
colonization. Perhaps then, the slow growth rates may
be due to transplanting the seedlings from a fertile
substrate to a low nutrient, acidic substrate.
Despite detecting 11 fungal species in CA1, 4
single-isolate cultures were obtained, and only 3 of
them were found to promote plant growth. These
results support findings of Branzanti et al. (1992) that
inoculation with AMF ensures maximum growth of
apple seedlings under low fertility, and efficiency
varies according to the isolate. Guillemin et al. (1992)
inoculated micropropagated pineapple plants with
one isolate of S. pellucida (LPA20), three isolates of
Glomus sp. (LPA21, LPA22 and LPA25) and one of
G. clarum (LPA16) and found that S. pellucida
126 Plant Cell Tiss Organ Cult (2007) 90:117–129
123

LPA20 did not promote plant growth compared with
the other isolates. Conversely, our isolate of S.
pellucida was one of the more efficacious fungi,
promoting both vegetative growth and P uptake,
supporting the hypothesis that the AMF species is not
the most appropriate taxonomic descriptor to link
physiological processes of the mycorrhizal associa-
tion with increases in plant growth (Morton 1993).
Although apple orchards receive substantial annual
inputs of fertilizers, 75% of isolates tested still
increased plant biomass contradicting the hypothesis
that long-term fertilization selects for less efficient
AMF isolates (Johnson 1993). According to Johnson,
the fungus is adapted to a rhizosphere low in root
exudates, as occurs with fertilized plants, and which
results in lower allocations to external mycelia. Even
though we did not test all species that were detected
in CA1, our results do not support the hypothesis that
fertilization selects for less mutualistic fungi.
Plant concentrations of P, Zn and Cu generally
increase from AMF inoculation (Bolan 1991). Inoc-
ulation increased P concentrations at all pH levels,
contradicting results of Branzanti et al. (1992) where
inoculation did not alter P concentrations. Increased P
uptake can be explained by the overall low levels of P
in our soil (mg kg soil�1), 5.1 at pH 4, 2.7 at pH 5 and
4.1 at pH 6, which as suspected did not inhibit
colonization and mycorrhizal activity. Inhibition has
been observed for Acacia mangium (Habte and
Soedarjo 1996), manihot (Sieverding and Howeler
1985) and red clover (Sainz and Arines 1988); P
concentrations in mycorrhizal and non-inoculated
plants averaged 1,400 and 700 mg kg�1, respectively.
For apple plants, Plenchette et al. (1981), Geddeda
et al. (1984) and Branzanti et al. (1992) found
maximum P concentrations of 1,800, 1,900 and
1,800 mg kg�1, respectively. Our P values are below
ideal foliar concentrations (1,500–3,000 mg kg�1)
recommended by Basso et al. (1986). Only plants
inoculated with S. heterogama SCT113 attained these
recommended concentrations at all pH levels. We
analyzed leaves and stems while the recommended
concentrations are for leaves. Additionally, ideal
concentrations are expected in plants growing under
optimal relative levels of all nutrients; our substrates
were low in P and certain other nutrients.
Calcium is important for producing apples as
deficient plants reduce Ca allocation to fruits (Basso
et al. 1986). In this work, inoculation tended to
increase Ca concentrations at pH 5.0 and 6.0, but not
at pH 4.0 where inoculation did not alter amounts.
This pattern may be explained by K and Mg soil
levels, which are considered medium to high at pH
5.0 and 6.0, and sufficient to medium at pH 4.0. High
availability of K and Mg can induce Ca-deficiency in
apples (Basso et al. 1986), as what likely occurred at
pH 5.0 and 6.0 where the mycobiont promoted Ca
uptake. Despite the inoculation effect for Ca concen-
trations, we measured plant levels considered insuf-
ficient by Basso et al. (1986), i.e., below
8,000 mg kg�1.
The fungal isolates appeared to reduce Al uptake
at pH 5.0 and 6.0 where non-inoculated plants had
greater shoot Al concentrations than levels found for
plants inoculated with any of the isolates. Isolate S.
pellucida SCT111 was highly efficient at reducing Al
toxicity at all acidity levels. A protective effect
against Al was observed by Mendoza and Borie
(1998) for oat plants inoculated with an isolate of G.
etunicatum and grown in soils having high levels of
Al. Clark and Zeto (1996) found that inoculation with
Glomus isolates decreased Al concentrations and
contents of maize plants growing in two acid soils
from West Virginia. However, Borie and Rubio
(1999) studied oat varieties sensitive and tolerant to
Al and inoculated with G. etunicatum. Inoculation
increased Al concentrations in sensitive plants rela-
tive to non-inoculated plants, but not in tolerant
plants. After liming, Al concentrations in sensitive
mycorrhizal plants were half the levels measured in
non-inoculated plants. In general, these results sug-
gest that protection against high Al levels in soil
depends on the mycorrhizal community, the ability of
the host plant to tolerate this element, and on the pH
of the substrate.
Although the mycorrhizal association frequently is
considered a mutualistic symbiosis, there is a con-
tinuum of plant responses to mycorrhizal colonization
ranging from positive (mutualism) to negative (par-
asitism) (Johnson et al. 1997). We have evidence for
this continuum when apple biomass and nutrient
uptake are considered. A. scrobiculata plants exhib-
ited parasitic behavior; its effect on these processes
did not differ from responses measured for non-
inoculated plants. Alternatively, S. heterogama plants
attained greater relative shoot and total mass, but not
root mass, and Zn and Mn uptake were optimized at
pH 5.0. Finally, isolates G. etunicatum and S.
Plant Cell Tiss Organ Cult (2007) 90:117–129 127
123

pellucida behaved as mutualists. Plants inoculated by
them, at all pH levels, achieved the greatest heights,
produced greater mass than non-inoculated plants,
and attained normal shoot contents of P, Zn, Cu, Fe
and Mn. An overall analysis scheme of this sort is
paramount to identify relative abundances of efficient
and non-efficient fungi composing a mycorrhizal
community and allows for a greater understanding of
how the symbiosis works in natural systems, a
necessary requirement to utilize AMF in agriculture
(Johnson et al. 1997).
Event though the fungal isolates originated from
acidic soils, and they produced significant differ-
ences among them in attaining plant biomass and
nutrient uptake, they generally were well adapted to
the range in soil pH studied. Knowing these relative
responses is important when the goal is to produce
commercial fungal inocula for wide-scale use on
plants grown in the field, where conditions are highly
variable. The ideal inoculum should be efficient
under the diverse of conditions found among field
settings (Lovato et al. 1992). In the field, attributes of
the mycorrhizal inoculum other than colonization
efficiency are important such as reproductive capac-
ity and ability to compete with indigenous soil biota
(Lambert et al. 1980). We demonstrated that AMF
isolates stimulate plant biomass, and nutrient uptake
by micropropagated apple plants growing under a
range of acidity. Further work is needed to investi-
gate the capabilities of the same isolates directly
under field conditions. Combinations of isolates must
be evaluated striving to produce multi-isolate inoc-
ulants that are superior to single-isolate inoculants,
potentially optimizing plant performance in highly
variable field conditions.
Acknowledgments This work is part of the requirements of
the senior author for a M.S. degree in biotechnology. This
project was funded by FINEP (Financiadora de Estudos e
Projetos, Brazil). We thank CAPES (Coordenacao de
Aperfeicoamento de Pessoal de Nıvel Superior, Ministerio de
Ciencia e Tecnologia, Brazil) and CNPq (Conselho Nacional
de Desenvolvimento Cientifico e Tecnologico) for providing
scholarships to JRPC during his graduate studies and to SLS as
a Post-doctoral researcher. We are grateful to Dr Enio Pedrotii
for providing micropropagated apple seedlings, and to Dr Clori
Basso (EPAGRI, Cacador, SC) for technical support while the
soils were sampled. We thank Drs P.E. Lovato and R.G.
Linderman for valuable suggestions made to an earlier version
of the manuscript. The document was subjected to the peer and
administrative reviews of the U.S. EPA at the National Health
and Environmental Effects Research Laboratory, Western
Ecology Division, and was approved for publication.
Mention of trade names or commercial products in this paper
does not constitute endorsement or recommendation of use.
References
Abbott LK, Robson AD (1981) Infectivity and effectiveness of
vesicular–arbuscular mycorrhizal fungi: effect of inocu-
lum type. Aust J Agric Res 32:631–639
Arines J, Vilarino A, Sainz M (1989) Effect of different
inocula of vesicular–arbuscular mycorrhizal fungi on
manganese content and concentration in red clover (Tri-folium pratense L.) plants. New Phytol 112:215–219
Basso C, Suzuki A (1992) Resposta da macieira cv. Golden
Delicious a adubacao nitrogenada. Rev Bras Ciencia Solo
16:223–227
Basso C, Wilms FWW, Suzuki A (1986) Fertilidade do solo e
nutricao de macieira. In: Manual da Cultura da Macieira.
EPAGRI, Florianopolis
Bever JD, Morton JB, Antonovics J, Schultz PA (1996) Host-
dependent sporulation and species diversity of arbuscular
mycorrhizal fungi in a mown grassland. J Ecol 84:71–82
Bolan NS (1991) A critical review on the role of mycorrhizal
fungi in the uptake of phosphorus by plants. Plant Soil
134:189–207
Borie F, Rubio R (1999) Effects of arbuscular mycorrhizae and
liming on growth and mineral acquisition of aluminum-
tolerant and aluminum-sensitive barley cultivars. J Plant
Nutr 22:121–137
Branzanti B, Gianinazzi-Pearson V, Gianinazzi S (1992)
Influence of phosphate fertilization on the growth and
nutrient status of micropropagated apple infected with
endomycorrhizal fungi during the weaning stage. Agron-
omie 12:841–845
Clark RB, Zeto SK (1996) Mineral acquisition by mycorrhizal
maize grown on acid and alkaline soil. Soil Biol Biochem
28:1495–1503
El-Kherbawy M, Angle JS, Heggo A, Chaney RL (1989) Soil
pH, rhizobia, and vesicular–arbuscular mycorrhizae
inoculation effects on growth and heavy metal uptake of
alfafa (Medicago sativa L.). Biol Fertil Soils 8:61–65
EMBRAPA (1999) Sistema Brasileiro de Classificacao de
Solos. Servicos de Producao de Informacoes, Brasilia, DF,
pp. 412
Fortuna P, Citernesi S, Morini S, Giovannetti M, Loreti F
(1992) Infectivity and effectiveness of different species of
arbuscular mycorrhizal fungi in micropropagated plants of
Mr S 2/5 plum rootstock. Agronomie 12:825–829
Geddeda YI, Trappe JM, Stebbins RL (1984) Effects of
vesicular–arbuscular mycorrhizae and phosphorus on ap-
ple seedlings. J Am Soc Hortic Sci 109:24–27
George EF (1996) Plant propagation by tissue culture—Part 2
in practice. Exegetics Limited, England, pp. 575–1361
Gerdemann JW, Nicolson TH (1963) Spores of mycorrhizal
Endogone species extracted from soil by wet sieving and
decanting. Trans Br Mycol Soc 46:235–244
Giovannetti M, Mosse B (1980) An evaluation of techniques
for measuring vesicular arbuscular mycorrhizal infection
in roots. New Phytol 84:489–500
128 Plant Cell Tiss Organ Cult (2007) 90:117–129
123

Graham JH, Linderman RG, Menge JA (1982) Development of
external hyphae by different isolates of mycorrhizal fungi
Glomus spp. in relation to root colonization and growth of
troyer citrange. New Phytol 91:183–189
Granger RL, Plenchette C, Fortin JA (1983) Effect of a
vesicular arbuscular (VA) endomycorrhizal fungus (Glo-mus epigaeum) on the growth and leaf mineral content of
two apple clones propagated in vitro. Can J Plant Sci
63:551–555
Grattapaglia D, Machado MA (1991) Micropropagacao. In:
Torres AC, Caldas LS, Boso JA (eds) Cultura de Tecidos
e Transformacao Genetica de Plantas. EMBRAPA, Bra-
silia, pp. 183–204
Guillemin JP, Gianinazzi S, Trouvelot A (1992) Screening of
arbuscular endomycorrhizal fungi for establishment of
micropropagated pineapple plants. Agronomie 12:831–836
Habte M, Soedarjo M (1996) Response of Acacia mangium to
vesicular–arbuscular mycorrhizal inoculation, soil pH,
and soil P concentration in an oxisol. Can J Bot 74:155–
161
Johnson NC (1993) Can fertilization of soil select less mutu-
alistic mycorrhizae? Ecol Appl 3:749–757
Johnson NC, Graham JH, Smith FA (1997) Functioning of
mycorrhizal associations along the mutualism–parasitism
continuum. New Phytol 135:575–585
Koske RE, Gemma JN (1989) A modified procedure for
staining roots to detect VA mycorrhizas. Mycol Res
92:486–505
Lambert DH, Cole H Jr, Baker DE (1980) Adaptation of
vesicular–arbuscular mycorrhizal to edaphic factors. New
Phytol 85:513–520
Lovato P, Guillemin JP, Gianinazzi S (1992) Application of
commercial arbuscular endomycorrhizal fungal inoculants
to the establishment of micropropagated grapevine root-
stock and pineapple plants. Agronomie 12:873–880
Mamo T, Killham KS (1987) Effect of soil liming and vesic-
ular–arbuscular mycorrhizal inoculation on the growth
and micronutrient content of the teff plant. Plant Soil
102:257–259
Marschner H (1991) Mechanisms of adaptation of plants to
acid soils. Plant Soil 134:1–20
Melo FAF (1985) Origem, natureza e componentes da acidez
do solo: criterios para calagem. In: Malavolta E (ed) Se-
minario sobre corretivos agricolas. Piracicaba, Fundacao
Cargill
Mendoza J, Borie F (1998) Effect of Glomus etunicatum on
aluminum, phosphorus, calcium and magnesium uptake of
two barley genotypes with different aluminum tolerance.
Commun Soil Sci Plant Anal 29:681–695
Miller DD, Domoto PA, Walker C (1985) Colonization and
efficacy of different endomycorrhizal fungi with apple
seedlings at two phosphorus levels. New Phytol 100:393–
402
Moorman T, Reeves FB (1979) The role of endomycorrhizae in
revegetation practices in the semi-arid west. II. A bioassay
to determine the effect of land disturbance on endomy-
corrhizal populations. Am J Bot 66:14–18
Morton JB (1993) Problems and solutions for the integration of
glomalean taxonomy, systematic biology, and the study of
endomycorrhizal phenomena. Mycorrhiza 2:97–109
Morton JB, Bentivenga SP, Wheeler WW (1993) Germplasm
in the International Collection of Arbuscular and Vesic-
ular–arbuscular Mycorrhizal Fungi (INVAM) and proce-
dures for culture development, documentation and
storage. Mycotaxon 48:491–528
Nunes JCO, Barpp A, Silva FC, Pedrotii EL (1991) Micro-
propagacao do porta-enxerto Marubakaido (Malus pruni-folia) a partir da cultura de meristemas. Rev Bras Frutic
21:191–195
O’Donnell JJ, Sylvia DM, Pitman WD, Rechcigl JE (1992)
Inoculation of Vigna parketi with mycorrhizal fungi in
acid Florida spodsol. Trop Grass 26:120–129
Plenchette C, Furlan V, Fortin JA (1981) Growth stimulation of
apple trees in unsterilized soil under field conditions with
VA mycorrhiza inoculation. Can J Bot 59:2003–2008
Plenchette C, Fortin JA, Furlan V (1983) Growth responses of
several plant species to mycorrhizae in a soil of moderate
P fertility. Plant Soil 70:199–209
Ravolanirina F, Gianinazzi S, Trouvelot A, Carre M (1989)
Production of endomycorrhizal explants of micropropa-
gated grapevine rootstocks. Agric Ecosyst Environ
29:323–327
Sainz MJ, Arines J (1988) Effects of native vesicular–arbus-
cular mycorrhizal fungi and phosphate fertilizer on red
clover growth in acid soils. J Agric Sci 111:67–73
Sieverding E (1991) Vesicular–arbuscular mycorrhiza man-
agement in tropical ecosystems. Deutsche Geselleschaft
fur Technische Zusammenarbeit, Eschborn
Sieverding E, Howeler RH (1985) Influence of species of VA
mycorrhizal fungi on cassava yield response to phospho-
rus fertilization. Plant Soil 88:213–221
Siqueira JO, Colozzi-Filho A, Oliveira E (1989) Ocorrencia de
micorrizas vesicular–arbusculares em agro e ecossistemas
do estado de Minas Gerais. Pesqui Agropecuaria Bras
24:1499–1506
Soedarjo M, Habte M (1993) Vesicular–arbuscular mycorrhizal
effectiveness in an acid soil amended with fresh organic
matter. Plant Soil 149:197–203
Smith SE, Read DJ (1997) Mycorrhizal symbiosis. Academic,
London, pp. 605
Stutz JC, Morton JB (1996) Successive pot cultures reveal high
species richness of arbuscular endomycorrhizal fungi in
arid ecosystems. Can J Bot 74:1883–1889
Tedesco MJ, Gianello C, Bissani CA, Bohnen H, Volkweiss SJ
(1995) Analise de solo, planta e outros materiais. Uni-
versidade Federal do Rio Grande do Sul, Porto Alegre
Uosukainen M, Vestberg M (1994) Effect of inoculation with
arbuscular mycorrhizas on rooting, weaning and sub-
sequent growth of micropropagated Malus (L.) Moench.
Agric Sci Finl 3:269–279
Weissenhorn I, Leyal C (1995) Root colonization of maize by a
Cd-sensitive and a Cd-tolerant Glomus mosseae and
cadmium uptake in sand culture. Plant Soil 175:233–238
Weissenhorn I, Leyal C, Berthelin J (1995) Bioavailability of
heavy metals and abundance of arbuscular mycorrhiza in
a soil polluted by atmospheric deposition from a smelter.
Biol Fertil Soils 19:22–28
Plant Cell Tiss Organ Cult (2007) 90:117–129 129
123